

Abstract
The focus of this review is to discuss findings in the last 10 years that have advanced our understanding of human NK cell responses to dengue virus. We will review recently identified interactions of activating and inhibitory receptors on NK cells with dengue virus, human NK responses to natural dengue infection and highlight possible interactions by which NK cells may shape adaptive immune responses. T cell responses to natural dengue infection will be reviewed by Laura Rivino in Chap. 10.1007/978-981-10-8727-1_17. With the advent of numerous dengue vaccine clinical trials, we will also review T and NK cell immune responses to dengue virus vaccination. As our understanding of the diverse functions of NK cell has advanced, it has become increasingly clear that human NK cell responses to viral infections are more complicated than initially recognized.
Keywords: Innate immune system, KIR peptide-MHC interactions, Inhibitory receptors on NK cells, Activationg receptors on NK cells, T-cell response to dengue vaccine
Introduction
NK cells are large granular lymphocytes that are a component of the innate immune system. NK cells kill target cells spontaneously without the requirement of priming or the expression of major histocompatibility antigens (MHC) on the target cell. NK cells lack the receptor diversity generated by DNA rearrangement found in prototype members of the adaptive immune response, T and B cells. However, similar to T cells, NK cells are able to distinguish healthy cells from virally infected cells. NK cells produce abundant antiviral cytokines, release cytotoxic granules such as granzyme B and trigger the FAS/TRAIL pathway to induce apoptosis of infected cells [35]. In order to mount a response, NK cells have to integrate signals received from many inhibitory and activating receptors to sense the environment and respond. Regulation of NK cell activity is achieved by a balance between these inhibitory and activating signals although signals received by inhibitory receptors tend to be dominant [21]. Many of the initial insights on NK cell function came from studying chronic viral infections in mice such as LCMV and MCMV [7]. The clearest demonstration that NK cells are important to humans stem from the findings that patients with NK cell deficiencies develop severe, recurrent viral infections [6, 54]. Our knowledge of select receptors on human NK cells modulated by viral infection and viral peptides that can modulate NK cell function has significantly advanced in the last decade in the context of human viral infections such as HMCV, HIV and HCV [7, 19]. Some of the findings seen in these human chronic viral infections are beginning to be observed for viral infections such as Influenza and Dengue thus opening up new areas of research that will enhance our understanding of how these innate effector cells can control acute viral infections [1, 4, 57, 58, 72]
Early and Current Observations on NK Cell Responses to Dengue Infections Using Samples from Clinical Cohorts
Human NK cells were shown to lyse dengue virus (DENV)-infected cells to a greater degree compared to uninfected cells over 30 years ago [38]. Antibodies to DENV further augmented the lysis of DENV-infected cells by NK cells. The traditional phenotype used to define human NK cells is based on the absence of CD3 (to exclude T cells) and relative expression of CD56 (neural cell adhesion molecule) and CD16 (low-affinity receptor for the Fc portion of immunoglobulin G). It is now appreciated that CD56 “bright” cells can produce cytokines abundantly but are poorly cytolytic while CD56 “dim” cells (which represent 90% of circulating NK cells) produce moderate cytokines when stimulated but are strongly cytolytic [60]. Using PBMC samples obtained from children in Thailand undergoing severe dengue infection nearly 20 years ago, Green et al. observed increased frequencies of activated NK cells (CD56+ CD69+ cells) in PBMC from patients with severe dengue disease compared to patients with milder disease suggesting that that NK cells are actively involved in the defense against dengue virus [26]. Homchampa et al. found evidence of NK cell cytotoxicity in fresh cells from children with acute dengue and the level of cytotoxicity was related to disease severity [31]. At the time, many of the activating and inhibitory receptors present on NK cells were not identified and the technologies to assess markers on individual NK cells using multiparametric flow cytometry and CyTOF were not available.
In a study published in 2008, frequencies of CD56+ CD69+ NK cells were found to be more activated in hospitalized Vietnamese infants with more severe dengue disease [15]. Recently Petitdemange et al. evaluated NK cell activation in patients from Gabon undergoing acute dengue infection [59]. They examined frequencies of subsets of NK cells and included a number of NK cell receptors NKp30, NKG2A, NKp44 and CD161 which was a significant advance from the early studies where only CD56 expression was used to identify NK cells. Compared to healthy donors, NK cells from patients who had DENV-2 infections expressed significantly less NKp30, NKG2A, and CD161, whereas expression of ILT-2 was increased at the early acute and convalescent time point. Interestingly, frequencies of NKp44+ NK cells were elevated at the early acute time point. Principal component analysis highlighted an association between NKp44 and DENV-2 infections. NKp44 has previously been shown to directly interact with the envelope protein of DENV and WNV and this subset of NK cells and its ligand in target cells is worthy of study in other cohorts [30]. While there have been only a few studies that have looked at NK cell responses in clinical cohorts, the data indicate that subsets of NK cells are involved in the immune responses to natural dengue infection thus warranting further study. NK cell responses in individuals receiving live attenuated tetravalent dengue vaccines have not been examined in any detail. Whether different subsets of NK cells respond to vaccination with live attenuated dengue vaccines versus subunit vaccines and inactivated vaccine antigens are currently unknown. Furthermore, whether the magnitude of NK cell responses is different between vaccination and natural infection has not been explored.
Activating Receptors on NK Cells
NK cells have a number of receptors that can activate NK cell reactivity (activating receptors) or dampen reactivity (inhibitory receptors). Activating receptors expressed on human NK cells include the C-type lectin receptors NKG2D and NKG2C/E, natural cytotoxicity receptors (NCR) NKp44, NKp30, NKp46, and CD16 (FC-γ-RIII), the low-affinity Fc receptor which mediates antibody dependent cytotoxicity (ADCC) (Fig. 18.1 and Table 18.1). Of the many activating receptors on NK cells and ligands on target cells, there is evidence for interaction of DENV with NKp44, the low-affinity FcR receptor CD16 and ligands for the NKG2D ligand MICB (Fig. 18.1).
Fig. 18.1.

Model of NK cell interaction with virus-infected cells. NK cells can interact with DENV infected target cells through direct and indirect mechanisms. To date, only NKp44 has been shown to directly interact with the E protein of DENV. NK cells can interact with DENV infected APCs by responding to cytokines, interacting with yet to be identified ligands on virus-infected cells and upregulating inhibitory or activating receptors on its surface. This may lead to degranulation and abundant cytokine secretion by NK cells and lysis of virus-infected cells. Alternatively, dominant interaction of inhibitory receptors on NK cells with viral proteins and peptides could dampen responses and represent an immune evasion strategy by DENV. NK cells can also be directed to DENV infected APCs through engagement of CD16 with the Fc receptor of DENV-specific Abs and target cells are lysed through ADCC
Table 18.1.
NK cell receptors and ligands
| Receptora | Ligandb | Functionc | DENV | Reference |
|---|---|---|---|---|
| CD16 | IgG | Activating | [40] | |
| CD94-NKG2C | HLA-E | Activating | [8, 36], | |
| CD94-NKG2E | HLA-E | Activating | [36] | |
| NKG2D | MICA, MICB, ULBP | Activating | GWAS [37] | [2] |
| NKp30 | Unknown | Activating | [55] | |
| NKp44 | Viral hemagglutinins | Activating | DENV protein E [30] | [11, 30] |
| NKp46 | Unknown | Activating | [56] | |
| CD161 | LLT1 | Activating and inhibitory | [46, 61] | |
| CD38 | CD31 | Activating | [20, 49, 67] | |
| KIR2DS1 | HLA-C | Activating | [52] | |
| KIR2DS2 | HLA-C | Activating | [52] | |
| KIR2DS3 | HLA-C | Activating | ||
| KIR2DS4 | HLA-Cw4 | Activating | [25] | |
| KIR2DS5 | Unknown | Activating | ||
| KIR3DS1 | HLA-Bw4? | Activating | ||
| Cytokine receptors | Type 1 IFN, IL-1,2,12,15,18 | Activating | ||
| Cytokine receptors | TGF-β | Inhibitory | ||
| KIR2DL1 | HLA-Cw4 | Inhibitory | [17, 48] | |
| KIR2DL2 | HLA-Cw3 | Inhibitory | [17, 48] | |
| KIR2DL3 | HLA-Cw3 | Inhibitory | [17, 48] | |
| KIR2DL4 | HLA-G | Activating? Inhibitory? | [23, 62] | |
| KIR2DL5A/B | Unknown | Inhibitory | ||
| KIR3DL1 | HLA-Bw4 | Inhibitory | KIR3DL1 and HLA B57 interaction [72] | [73]) |
| KIR3DL2 | HLA-B27, HLA-A3, HLA-A11 | Inhibitory | [28, 78] | |
| KIR3DL3 | HLA-Cw4 | Inhibitory | ||
| CD94-NKG2A | HLA-E | Inhibitory | [8] | |
| 2B4 | CD48 | Activating or inhibitory, coreceptor | [42, 9] | |
| LILRB1 | HLA class I | Inhibitory | Dengue and ADE [14] | [10] |
aReceptor present on the surface of NK cells
bLigand(s) bound by the NK cell receptor
cFunctional effect of receptor signaling in NK cells
In a large genome-wide association study (GWAS) performed in 2011, a single nucleotide polymorphism in MICB (ligand for NKG2D) was found to be a susceptibility locus for severe dengue disease dengue shock syndrome in over 2000 Vietnamese children [37]. In a subsequent study by the same group, MICB was also associated with the less severe form of dengue DF in both children and adults in Vietnam [77]. This genetic susceptibility of a SNP in MICB was confirmed in another study in the Thai population. In Cuban individuals (Guzman), a strong association of DF and DHF with MICA 008 and MICB 008 compared to asymptomatic individuals [24]. Interestingly soluble MICB levels were increased in the sera between pre-infection and acute illness among infants with symptomatic primary DENV infections in the Philippines and suggest an immune evasion strategy in dengue viral infection [43]. The data suggest that ligands for the activating receptor NKG2D, MICB may be involved in the immune response against DENV.
A direct interaction has been shown between the NK-activating receptor NKp44 and the DENV and WNV envelope protein [30]. Recombinant NKp44 directly bound purified DENV and WNV envelope proteins. The results suggest that NKp44 expressing NK cells are able to interact directly with the E protein on flavivirus-infected cells.
NK cells express the FcγRIIIA receptor (CD16) that binds the constant (Fc) domain of IgG antibodies. CD16 engagement is a strong activator of NK cell function and allows antigen-specific recruitment of NK responses. Very early on, NK cell-mediated lysis of DENV-2-infected Raji cells was significantly higher when these cells were pre-incubated with DENV immune serum through antibody dependent cell-mediated cytotoxicity (ADCC) [38]. More recently efforts have been made to understand in a prospective cohort whether ADCC activity in sera obtained pre-illness in a prospective cohort correlated with subsequent dengue disease severity and viremia levels. A higher level of ADCC activity measured in immune sera before secondary DENV-3 but not secondary DENV-2 infection was associated with lower subsequent viremia levels [41]. These results suggest a protective role for ADCC antibodies in sera obtained from in some children with secondary infection. In vivo, characteristics of both the Abs and NK cells in each individual are likely to impact the function of these Abs. Future studies should compare the contribution of functionally distinct Abs (those that mediate neutralization versus Abs that mediate ADCC) in the sera of individuals who subsequently develop clinical versus subclinical disease.
In order to understand soluble and membrane found factors important for activating NK cells, Lim et al. used DENV-infected monocyte-derived dendritic cells in co-culture with autologous NK cells from healthy donors [44]. The results suggest that a combination of cytokines type I IFNs, TNF-☐ secreted by DCs and cell surface receptor–ligand interactions all play some part triggering the anti-dengue response of primary human NK cells while each of these in isolation has a low to minimal response on the activation of human NK cells.
Inhibitory Receptors on NK Cells
KIRs
A major set of inhibitory receptors important for NK cell licensing is the killer cell immunoglobulin-like receptors (KIRs). The KIR receptors are expressed stochastically, leading to the generation of subpopulations of NK cells within each individual [50]. While only some KIR ligands are known, all are thought to be class I MHC molecules (Table 18.1). Epidemiological studies of immunological events, including responses to infectious diseases, autoimmune diseases, complications with pregnancy, and tumor responses, have implicated KIR/MHC interactions in protective or pathological roles [5, 34, 32, 39, 53, 63, 68].
In contrast to the T-cell receptor, which recognizes a given viral peptide-MHC complex, KIRs typically recognize subsets of MHC class I molecules with common amino acid motifs in their α1 domains ([13, 33]). However, several studies have shown that inhibitory KIRs are also sensitive to the specific peptides bound by the HLA molecules [12]. In particular, residues 7 and 8 of endogenously derived peptides bound to MHC class I can promote or abrogate binding of KIR. During viral infection, the peptide repertoire changes and a number of viral peptides are presented on MHC molecules for recognition by cytotoxic T lymphocytes. Changes in the endogenous repertoire of peptides can perturb or continue to maintain inhibition of NK cells (Fig. 18.2) depending on how strongly the viral peptide is able to bind and replace the endogenous peptide. Several viral peptides (HIV, HCV, SIV) have been shown to modulate KIR-MHC interactions thus providing a mechanism for the virus to modulate the interaction of the inhibitory KIR with its MHC ligand thus preventing the activation of subsets of NK cells [22, 47]. How changes in the peptide repertoire following viral infection alter NK cell recognition in vivo will depend on the relative amounts of viral peptides being presented, the nature of the peptides produced by infection and the frequency of NK cells expressing the relevant KIR.
Fig. 18.2.
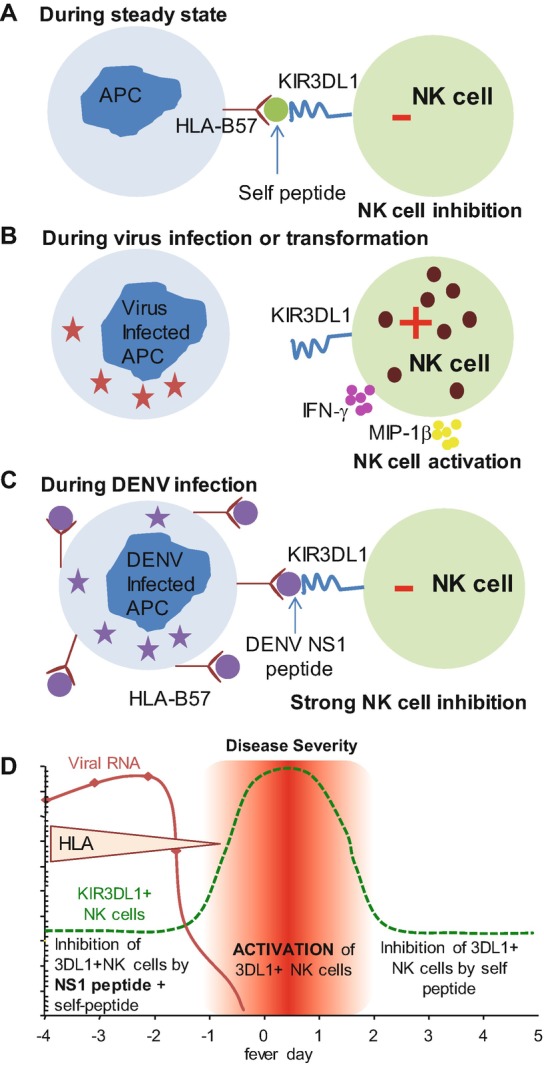
A model of NK-cell inhibition during DENV infection. Specific amino acid changes in the peptides presented by major histocompatibility complex (MHC) class I can impact KIR recognition of peptide/MHC complexes. We hypothesize that the DENV NS1 peptide generated during infection presented by HLA B57 alters the affinity of the inhibitory KIR3DL1 receptor expressed on NK cells for its putative ligand, resulting in the inhibition of NK cells early during viral infection while MHC class I is upregulated. Following the clearance of viremia, HLA-B57 levels return to normal which decreases the inhibitory signal received by KIR3DL1+ NK cells. This release of inhibition allows KIR3DL1+ NK cells to be activated at fever day 0, coincident with the activation of CD8+ T cells
KIR Peptide-MHC Interactions During Dengue Infection
During a study of HLA B57 restricted T cells in acute dengue infection, Townsley et al. found that a T cell tetramer for a CD8+ T cell epitope in the NS1 protein of DENV bound CD56dim NK cells [72]. Using depletion studies and KIR-transfected cell lines, the HLA B57 restricted dengue NS1 tetramer was found to bind the inhibitory receptor KIR3DL1 on NK cells clearly demonstrating an interaction with an inhibitory KIR. Using clinical samples from a long-standing cohort in Thailand the authors further found late activation of NK cells in HLA B57+ individuals. The peak of NK cell activation occurred at the critical time point in people with severe dengue disease which supported the hypothesis that the HLA B57 restricted NS1 peptide can modulate the function of KIR3DL1+ NK cells. The data strongly suggest that subpopulations of NK cells may contribute to disease pathogenesis and enabled the authors to speculate how the NS1 peptide may modulate function during acute dengue infection. At steady state (Fig. 18.1a), the interaction of an HLA B57 presenting endogenous peptides on an antigen presenting cell (APC) with KIR3DL1 maintains an inhibitory signal in NK cells thus preventing lysis of healthy target APCs. During many viral infections, HLA molecules are downregulated which allows NK cells to get activated and lyse virus-infected APCs (Fig. 18.1b). However it is well known that in DENV infection HLA class I molecules are upregulated [79]. We hypothesize that presentation of the NS1 peptide on HLA B57 during the viremic phase in infected individuals maintains the inhibitory interaction with KIR3DL1 (since the tetramer has been shown to bind KIR3DL1) thus delaying the activation of this important subset of NK cells (Fig. 18.1c, d). As viremia decreases, Class I expression decreases and NK cells are able to sense the change in Class I expression, triggering their activation during the critical phase of illness (around fever day 0) (Fig. 18.1d). Our clinical data support our hypothesis but we are yet to show functional alteration of NK cell responses by the NS1 peptide.
We are currently using NK cell sensitive TAP-impaired and TAP-blocked target cells (721.221) transfected with HLA alleles in functional assays. Since self-peptide is able to maintain inhibition, the use of TAP blocked and TAP impaired cell lines HLA transfected cell lines are essential to evaluate whether viral peptides added exogenously such as the DENV NS1 peptide can modulate the function of KIR3DL1+ NK cells. We anticipate that additional KIR/HLA interactions occur which are likely to be modulated by viral peptides during acute dengue.
Petitdemange et al. performed the genotypic analysis of KIR in the combination with their cognate HLA-class I ligands in 73 CHIKV and 55 DENV2 adult cases, compared with 54 healthy individuals [57]. Petitdemange et al. found no evidence of a role for KIR genotypes in patients infected with DENV-2. In contrast, Beltrame et al. detected an association between certain KIR genes and their cognate HLA ligands in the context of infection with DENV-3 in Southern Brazil [3, 57]. Differences in population origin and the infecting DENV serotype may explain these disparate results. Petitdemange et al. in a subsequent paper assessed NK cell activation by multiparametric flow cytometry in patients with DENV-2, CHIKV or in a few subjects with co-infections [59]. They found different signatures of NK cell responses between the two infections. NK cells in CHIKV infection were activated early and expressed a terminal differentiation pattern with prolonged persistence of NKG2C + CD57+ cells which the authors speculate may contribute to the chronic arthralgia seen in CHIKV infection. DENV-2 infections were associated with an increase in KIR2DL1+ NK cells which recognize HLA-C2. Together, these studies suggest that KIR-MHC interactions are likely to be important during acute dengue infection.
LILRB
LILRB is an inhibitory receptor present on monocytes, dendritic cells, and NK cells. It interacts with a wide range of MHC Class I molecules and maintains a negative feedback loop to prevent autoimmunity [70]. THE UL-18 protein of HCMV has been shown to bind with higher affinity to LILRB and protect against NK cell recognition in the context of HCMV infection. Recently using dengue-specific antibodies at neutralizing and sub-neutralizing concentrations, mechanisms of antibody-dependent enhancement (ADE) were further characterized in a resistant (THP-1R) and susceptible (THP-1S) subclone of THP-1 cells [14]. In this context, DENV was shown to interact with the inhibitory receptor LILRB. This inhibitory interaction blocked FcγRII signaling and dampened the expression of IFN stimulated genes and enhances DENV replication.
HLA-E is a non-classical MHC molecule that interacts with both activating (CD94C, D, and E) and inhibitory receptors (CD94A). Interaction of HLA-E with the inhibitory receptor NKG2A is of higher affinity compared to the interaction with most known HLA-E/peptide complexes than those transmitting activating signals. A related flavivirus, Japanese Encephalitis virus has been shown to upregulate HLA-E but no work has been published yet for DENV [69].
The Role of NK Cells in Modulating Adaptive Immune Responses
NK cells have also been implicated in shaping the adaptive response to viral infections in a number of ways including promoting maturation or elimination of DCs, perforin-dependent elimination of CD8+ T cells, and cytokine production [64]. Waggoner et al. used the model of lymphocytic choriomeningitis virus (LCMV), to show that NK cells can regulate CD4 T-cell-mediated support for the antiviral CD8 T cells [74, 76]. NK cells have also been shown to be important for long-term CD4+ T cell memory and subsequent antibody responses [18]. The data suggest that NK cells continue to participate in immune modulation well after initial infection when NK cells are traditionally thought to be active. In a previous study of CD8+ T cells by Townsley et al. frequencies of the HLA-B57-restricted epitope, were assessed over the course of acute DENV infection [71]. Given the highly conserved nature of this epitope, we predicted that PBMC from donors with secondary dengue infection would have significantly higher frequencies of B57-NS126–34 CD8+ T cells compared to PBMC from donors with primary dengue infection. While we detected tetramer-positive T cells in all subjects tested, the frequencies in subjects with secondary infections were not higher than in subjects with primary infections, with one exception. We speculated that an unidentified factor may dampen activation of CD8+ T cells directed at this epitope but had not yet identified that the NS1 peptide presented on HLA-B57 could bind KIR3DL1 an inhibitory receptor on NK cells. Our new findings suggest that NK cells could shape CD8+ T cell responses but given the lack of an authentic animal model that mimics human dengue infection it will be challenging to provide definite proof that inhibitory NK cells can modulate adaptive responses at the epitope level in vivo.
The varying combinations of inhibitory and activating receptors on NK cells and the number of unknown ligands make it difficult to assess changes in absolute frequencies of subsets of NK cells between subjects with mild or severe dengue illness. Furthermore, for meaningful comparisons to be made samples must be collected at multiple points and compared in subjects with mild and severe dengue disease rather than compare responses in all subjects with dengue disease and responses in healthy subjects. Since the hallmark of dengue hemorrhagic fever DHF (severe disease) is plasma leakage, if NK cells are hypothesized to contribute to DHF, then changes in the frequency or function of subsets of NK cells during or prior to defervescence the critical phase of illness must be demonstrated. In vitro assays will need to be performed in order to understand which receptors and ligands interact with virus-infected cells and are important for cytokine production and cytolytic activity in subsets of NK cells following DENV infection as little information exists to date. Basic biological studies of NK cells and multiple studies using clinical samples in chronic viral infections suggest that subsets of NK cells are an important contributor to the overall immune response. We have a partial understanding of how NK cells can regulate dengue immune responses but these findings point to multiple avenues whereby NK cells may control DENV infection and shape adaptive immune responses.
T-Cell Responses to Dengue Vaccination
T cell immune responses to natural dengue infection will be reviewed by Rivino et al. in Chap. 10.1007/978-981-10-8727-1_15. With the advent of a number of dengue tetravalent vaccine trials, T cell responses are being evaluated in vaccinated individuals [66]. These studies indicate that CD4 and CD8 T cells are directed against a number of epitopes across the entire DENV genome. In support of smaller studies performed in individuals receiving monovalent DENV vaccines, the data indicate that non-structural proteins are dominant targets of T cell responses following tetravalent vaccination as well [51]. There are significant differences in the composition of dengue antigens included in each vaccine which are likely to impact the immune response elicited [66]. Prior flavivirus immunity in many volunteers who received dengue vaccines in endemic countries will also influence the magnitude and quality of T cell immune responses. Here, published work to characterize CD4+ and CD8+ T-cell responses to dengue virus antigens and peptides will be reviewed.
The Sanofi Pasteur vaccine is a chimeric Yellow fever virus that expresses the structural components (prM and E) of each of the four serotypes of dengue [65, 66]. In three independent phase 1 clinical trials conducted in Australia and in the USA, Dengue 1–4 and Yellow Fever 17D (YF 17D)-specific CD4 and CD8 cellular responses induced by tetravalent chimeric dengue vaccines (CYD) were analyzed in flavivirus-naive or immune individuals [27]. Significant YF 17D NS3- specific CD8 responses and dengue serotype-specific T helper responses were detected. These responses were dominated by serotype 4 in naive individuals, but a booster vaccination broadened serotype-specific responses. A similar, broader response was seen after primary tetravalent immunization in subjects with pre-existing dengue 1 or 2 immunity caused by prior monovalent live- attenuated dengue vaccination. There was an absence of a Th2 response, and a more dominant response to IFN-☐ compared to TNF-α, for both CD4 and CD8 responses.
CD4 and CD8 T cell responses were assessed before and 28 days after a first and third injection of Sanofi Pasteur live attenuated CYD-TDV and 1 year after the third injection in 80 adolescents and adults enrolled in a phase II trial in Singapore [29]. CD4/IFN-γ and TNF-α responses specific to dengue NS3 were detected before vaccination. Vaccination induced YF-17D-NS3-specific CD8 IFN-γ responses. Using a multiplex analysis of peripheral blood mononuclear cells (PBMC) supernatants after restimulation with each the CYD vaccine viruses, higher levels of IFN-γ were detected compared with TNF-α, and low-level IL-13 levels.
The Takeda vaccine is a live-attenuated tetravalent dengue vaccine (TDV) candidate that consists of an attenuated DENV-2 strain (TDV-2), and 3 chimeric viruses containing the premembrane (prM) and envelope (E) genes of DENV-1, -3, and -4 expressed in the context of the TDV-2 genome (TDV-1, TDV-3, TDV-4, respectively). CD8+ T-cell responses in flavivirus-naive human volunteers vaccinated with 2 doses of the Takeda vaccine 90 days apart via two routes (subcutaneous or intradermal) were assessed using overlapping peptide pools against the NSl, NS3 and NS5 proteins [16]. The TDV vaccine-elicited CD8+ T cell TNF-☐ and IFN-☐ responses against the nonstructural NS l, NS3, and NS5 proteins of DENV-2. Responses were highest on day 90 after the first dose and were still detectable on 180 days after the second dose. In addition, CD8+ T cells were multifunctional, producing 2 cytokines simultaneously, and cross-reactive to NS proteins of the other 3 DENV serotypes.
For the NIH vaccine, attenuation was achieved by deleting a portion of the 31 untranslated region (UTR) of the dengue genome. Longitudinal specimens were collected in flavivirus-naive volunteers and tested for T cell responses against inactivated DENV antigen preparations made in Vero cells following low dose vaccination with a live attenuated DENV-1 candidate vaccine (DEN1L130) from the NIH. CD4+ T cells were found to secrete IFN-γ, TNF-α and IL-2, 3 weeks following exposure to DENV-1 and were detected upto 6 weeks. T cells produced 2 cytokines simultaneously [45]. There was little cross-reactivity in T cell responses.
Weiskopf et al analyzed CD8+ T cell responses in PBMC from healthy naïve volunteers in Vermont who received either monovalent or tetravalent live attenuated DENV vaccine from the NIH [75]. Using peptide pools (9- and 10-mer peptides that were predicted for their binding affinity to 27 major histocompatibility complex (MHC) class I molecules to monovalent and tetravalent DENV vaccines from the NIH. They found broad responses (IFN-☐ and TNF-☐ responses) to both structural and non-structural proteins were elicited in response to a monovalent vaccine (up to 47 months post-vaccination) while responses to a tetravalent vaccine (up to 12 months post-vaccination) were directed against the non-structural proteins.
Immune responses in vaccinated individuals are continuing to be monitored. How durable T cell responses to any of the attenuated vaccines in clinical trials is of interest as waning immunity to dengue is proposed to increase the risk of severe illness.
Questions
Are distinct subsets of NK cells elevated in natural DENV infection and do they differ between mild and severe disease, primary or secondary dengue infection?
Can viral peptides modulate NK cell function by interacting with inhibitory and activating KIRs?
Can distinct subsets of NK cells be induced following vaccination and how do they differ from natural infection?
Do preexisting dengue antibodies modulate NK cell responses?
Are there new ligands on target cells that are important for flavivirus infections?
Discussion of Chapter 18 at Dengue and Zika: Control and Antiviral Treatment Strategies
This discussion was held at the 2nd Advanced Study Week on Emerging Viral Diseases at Praia do Tofo, Mozambique.Transcribed by Hilgenfeld R and Vasudevan SG (Eds); approved by Dr. Anuja Mathew.
Siew Pheng Lim: Do you see sequence homology of your NS1 peptide with the self derived peptide and also within the sequence of the protein for Dengue?
Anuja Mathew: Definitely, there is some homology between the NS1 peptide and self peptide. There is no strong motif for peptide binding to KIRs, even for self peptides although positions 7 and 8 of bound peptides have been shown to bind KIRs. Our dengue virus peptide does match reasonably. Since self peptide can mediate inhibition, we still have to proove that the dengue peptide can modulate function in vivo.
Katja Fink: Is this a dominant peptide? Do all patients display this dominant NS1 peptide?
Anuja Mathew: So all of the HLA-B57 people that we have tested do. But we have only tested a few. B57 is unfortunately an uncommon allele in Thailand, so our access to samples were limited. If it was on HLA-A11 we had hundreds of samples to test.
Laura Rivino: I just have a question on the first part: when you tested the two tetramers, the A11 and the B57, you tested them in a secondary Dengue-2 patient. So when you tested the different Dengue-2 peptide did you also test the other serotypes?
Anuja Mathew: We have done studies, where we had all three tetramers, three variants together, and then single variants of each of the peptides. Specifically for secondary infection we would test with whatever they had currently. But for this particular donor, I know we have tested the individual variants.
Laura Rivino: Do you see a very different response?
Anuja Mathew: No, not a striking difference. You would expect that maybe you would see one, but no. In some instances, and I do not know if it is something unique about the serotype, but for Dengue-1, we see a dominant binding of Dengue-1 variant peptide tetramer. There may be something about Dengue-1 that we still have to figure out.
Laura Rivino: I want to make a comment on B57. So in ARDS, this is published very well. HLA Bs seem to be better at presenting, but I think they are just better at inducing T-cell responses in general.
Anuja Mathew: I think that is true. We have had that experience, too. I think all of the B57 donors we tested are responders. But in general, the frequencies are not as high as we expected to see given how conserved the B57 epitope is. You know, frequencies are similar to what we see in A11+ donors. It is fairly low.
Laura Rivino: And have you tested other HLAs, apart from the A11-NS3 peptide?
Anuja Mathew: We have tested A2, A11, B7, B57.
Laura Rivino: And you find A11 is partcularly low?
Anuja Mathew: We detect low responses to many HLA restricted epitopes. Every now and then, you’ll get one or two donors that have a high frequency of TET+ T cells. We have mutiple time points during acute infection, and that is the strength of our studies. It is easy to miss peak frequencies when you use only one or two time points. We have about five time points in each individual during acute infection. That is our strength of our studies, I think.
George Gao: Regarding your NS1 peptide which might be the B57. If they are binding to the KIR, is that peptide detected?
Anuja Mathew: We really hope, it is. We show binding of NK cells to B57 NS1 tetramers. We show binding of the B57 NS1 tetramers with KIR3DL1 transfectants. But the question really is: can it alter NK cell functional responses? We have been struggling to find proof for that. This is a challenge in the field including those working more actively on HIV viral peptides.
Subhash Vasudevan: What does it mean for severe dengue? Because you are saying that the NK cell response is during the defervesence period and in secondary dengue at least that is the period where you can get severe disease. So how does it fit the observed T-cell response in that period?
Anuja Mathew: What is new about this study is the unexpected NK cell regulation for a CD8+ T cell peptide. We were looking at frequencies of B57 NS1 CD8 T cells to determine whether frequencies of these T cells correlate with severe disease. Here we find an interaction with an inhibitory receptor on NK cells, which is something to consider if you do not see what you expect.
Subhash Vasudevan: So are the cytokines downstream of any importance?
Anuja Mathew: That is what we hope. Our expectation is that during the viremic phase this peptide is able to inibit NK cells. When the viremia ends, there is no longer presentation of the NS1 peptide, and that leads to NK-cell inhibition being released. So now suddenly the NK cells get activated and release the cytokines which may contribute to the cytokine storm seen during end of viremia and the critical phase of illness. That is our thinking, but that is still to proven.
Contributor Information
Rolf Hilgenfeld, Email: hilgenfeld@biochem.uni-luebeck.de.
Subhash G. Vasudevan, Phone: +65656565166718, FAX: +6565+65-6221-2529, Email: subhash.vasudevan@duke-nus.edu.sg
Anuja Mathew, Email: mathewa@uri.edu.
References
- 1.Aranda-Romo S, Garcia-Sepulveda CA, Comas-Garcia A, Lovato-Salas F, Salgado-Bustamante M, Gomez-Gomez A, Noyola DE. Killer-cell immunoglobulin-like receptors (KIR) in severe a (H1N1) 2009 influenza infections. Immunogenetics. 2012;64:653–662. doi: 10.1007/s00251-012-0623-3. [DOI] [PubMed] [Google Scholar]
- 2.Bauer S, Groh V, Wu J, Steinle A, Phillips JH, Lanier LL, Spies T. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science. 1999;285:727–729. doi: 10.1126/science.285.5428.727. [DOI] [PubMed] [Google Scholar]
- 3.Beltrame LM, Sell AM, Moliterno RA, Clementino SL, Cardozo DM, Dalalio MM, Fonzar UJ, Visentainer JE. Influence of KIR genes and their HLA ligands in susceptibility to dengue in a population from southern Brazil. Tissue Antigens. 2013;82:397–404. doi: 10.1111/tan.12256. [DOI] [PubMed] [Google Scholar]
- 4.Beltran D, Lopez-Verges S. NK cells during dengue disease and their recognition of dengue virus-infected cells. Front Immunol. 2014;5:1–6. doi: 10.3389/fimmu.2014.00192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bettencourt A, Silva AM, Carvalho C, Leal B, Santos E, Costa PP, Silva BM. The role of KIR2DS1 in multiple sclerosis: KIR in Portuguese MS patients. J Neuroimmunol. 2014;269(1–2):52–55. doi: 10.1016/j.jneuroim.2014.01.009. [DOI] [PubMed] [Google Scholar]
- 6.Biron CA, Byron KS, Sullivan JL. Severe herpesvirus infections in an adolescent without natural killer cells. N Engl J Med. 1989;320:1731–1735. doi: 10.1056/NEJM198906293202605. [DOI] [PubMed] [Google Scholar]
- 7.Blish CA. Natural killer cell diversity in viral infection: why and how much? Pathog Immun. 2016;1:165–192. doi: 10.20411/pai.v1i1.142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Braud VM, Allan DS, O’callaghan CA, Soderstrom K, D’andrea A, Ogg GS, Lazetic S, Young NT, Bell JI, Phillips JH, Lanier LL, Mcmichael AJ. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature. 1998;391:795–799. doi: 10.1038/35869. [DOI] [PubMed] [Google Scholar]
- 9.Brown MH, Boles K, Van Der Merwe PA, Kumar V, Mathew PA, Barclay AN. 2B4, the natural killer and T cell immunoglobulin superfamily surface protein, is a ligand for CD48. J Exp Med. 1998;188:2083–2090. doi: 10.1084/jem.188.11.2083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Brown D, Trowsdale J, Allen R. The Lilr family: modulators of innate and adaptive immune pathways in health and disease. Tissue Antigens. 2004;64:215–225. doi: 10.1111/j.0001-2815.2004.00290.x. [DOI] [PubMed] [Google Scholar]
- 11.Cantoni C, Bottino C, Vitale M, Pessino A, Augugliaro R, Malaspina A, Parolini S, Moretta L, Moretta A, Biassoni R. NKP44, a triggering receptor involved in tumor cell lysis by activated human natural killer cells, is a novel member of the immunoglobulin superfamily. J Exp Med. 1999;189:787–796. doi: 10.1084/jem.189.5.787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cassidy SA, Cheent KS, Khakoo SI. Effects of peptide on NK cell-mediated Mhc I recognition. Front Immunol. 2014;5:133. doi: 10.3389/fimmu.2014.00133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cassidy S, Mukherjee S, Myint TM, Mbiribindi B, North H, Traherne J, Mulder A, Claas FH, Purbhoo MA, Das J, Khakoo SI. Peptide selectivity discriminates NK cells from KIR2DL2- and KIR2DL3-positive individuals. Eur J Immunol. 2015;45:492–500. doi: 10.1002/eji.201444613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chan KR, Ong EZ, Tan HC, Zhang SL, Zhang Q, Tang KF, Kaliaperumal N, Lim AP, Hibberd ML, Chan SH, Connolly JE, Krishnan MN, Lok SM, Hanson BJ, Lin CN, Ooi EE. Leukocyte immunoglobulin-like receptor B1 is critical for antibody-dependent dengue. Proc Natl Acad Sci U S A. 2014;111(7):2722. doi: 10.1073/pnas.1317454111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chau TN, Quyen NT, Thuy TT, Tuan NM, Hoang DM, Dung NT, Lien Le B, Quy NT, Hieu NT, Hieu LT, Hien TT, Hung NT, Farrar J, Simmons CP. Dengue in vietnamese infants--results of infection-enhancement assays correlate with age-related disease epidemiology, and cellular immune responses correlate with disease severity. J Infect Dis. 2008;198:516–524. doi: 10.1086/590117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chu H, George SL, Stinchcomb DT, Osorio JE, Partidos CD. CD8+ T-cell responses in Flavivirus-Naive individuals following immunization with a live-attenuated tetravalent dengue vaccine candidate. J Infect Dis. 2015;212:1618–1628. doi: 10.1093/infdis/jiv258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Colonna M, Borsellino G, Falco M, Ferrara GB, Strominger JL. HLA-C is the inhibitory ligand that determines dominant resistance to Lysis by NK1- and NK2-specific natural killer cells. Proc Natl Acad Sci U S A. 1993;90:12000–12004. doi: 10.1073/pnas.90.24.12000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cook KD, Waggoner SN, Whitmire JK. NK cells and their ability to modulate T cells during virus infections. Crit Rev Immunol. 2014;34:359–388. doi: 10.1615/CritRevImmunol.2014010604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Das J, Khakoo SI. NK cells: tuned by peptide? Immunol Rev. 2015;267:214–227. doi: 10.1111/imr.12315. [DOI] [PubMed] [Google Scholar]
- 20.Deaglio S, Zubiaur M, Gregorini A, Bottarel F, Ausiello CM, Dianzani U, Sancho J, Malavasi F. Human CD38 and CD16 are functionally dependent and physically associated in natural killer cells. Blood. 2002;99:2490–2498. doi: 10.1182/blood.V99.7.2490. [DOI] [PubMed] [Google Scholar]
- 21.Della Chiesa M, Marcenaro E, Sivori S, Carlomagno S, Pesce S, Moretta A. Human NK cell response to pathogens. Semin Immunol. 2014;26:152–160. doi: 10.1016/j.smim.2014.02.001. [DOI] [PubMed] [Google Scholar]
- 22.Fadda L, Korner C, Kumar S, Van Teijlingen NH, Piechocka-Trocha A, Carrington M, Altfeld M. Hla-CW*0102-restricted HIV-1 P24 epitope variants can modulate the binding of the inhibitory KIR2DL2 receptor and primary NK cell function. PLoS Pathog. 2012;8:e1002805. doi: 10.1371/journal.ppat.1002805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Faure M, Long EO. KIR2DL4 (CD158D), an NK cell-activating receptor with inhibitory potential. J Immunol. 2002;168:6208–6214. doi: 10.4049/jimmunol.168.12.6208. [DOI] [PubMed] [Google Scholar]
- 24.Garcia G, Del Puerto F, Perez AB, Sierra B, Aguirre E, Kikuchi M, Sanchez L, Hirayama K, Guzman MG. Association of MICA and MICB alleles with symptomatic dengue infection. Hum Immunol. 2011;72:904–907. doi: 10.1016/j.humimm.2011.06.010. [DOI] [PubMed] [Google Scholar]
- 25.Graef T, Moesta AK, Norman PJ, Abi-Rached L, Vago L, Older Aguilar AM, Gleimer M, Hammond JA, Guethlein LA, Bushnell DA, Robinson PJ, Parham P. KIR2DS4 is a product of gene conversion with KIR3DL2 that introduced specificity for HLA-A*11 while diminishing avidity for HLA-C. J Exp Med. 2009;206:2557–2572. doi: 10.1084/jem.20091010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Green S, Pichyangkul S, Vaughn DW, Kalayanarooj S, Nimmannitya S, Nisalak A, Kurane I, Rothman AL, Ennis FA. Early CDd69 expression on peripheral blood lymphocytes from children with dengue hemorrhagic fever. J Infect Dis. 1999;180:1429–1435. doi: 10.1086/315072. [DOI] [PubMed] [Google Scholar]
- 27.Guy B, Nougarede N, Begue S, Sanchez V, Souag N, Carre M, Chambonneau L, Morrisson DN, Shaw D, Qiao M, Dumas R, Lang J, Forrat R. Cell-mediated immunity induced by chimeric tetravalent dengue vaccine in naive or flavivirus-primed subjects. Vaccine. 2008;26:5712–5721. doi: 10.1016/j.vaccine.2008.08.019. [DOI] [PubMed] [Google Scholar]
- 28.Hansasuta P, Dong T, Thananchai H, Weekes M, Willberg C, Aldemir H, Rowland-Jones S, Braud VM. Recognition of HLA-A3 and HLA-A11 by KIR3DL2 is peptide-specific. Eur J Immunol. 2004;34:1673–1679. doi: 10.1002/eji.200425089. [DOI] [PubMed] [Google Scholar]
- 29.Harenberg A, Begue S, Mamessier A, Gimenez-Fourage S, Ching Seah C, Wei Liang A, Li Ng J, Yun Toh X, Archuleta S, Wilder-Smith A, Shek LP, Wartel-Tram A, Bouckenooghe A, Lang J, Crevat D, Caillet C, Guy B. Persistence of Th1/Tc1 responses one year after tetravalent dengue vaccination in adults and adolescents in Singapore. Hum Vaccin Immunother. 2013;9:2317. doi: 10.4161/hv.25562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hershkovitz O, Rosental B, Rosenberg LA, Navarro-Sanchez ME, Jivov S, Zilka A, Gershoni-Yahalom O, Brient-Litzler E, Bedouelle H, Ho JW, Campbell KS, Rager-Zisman B, Despres P, Porgador A. NKp44 receptor mediates interaction of the envelope glycoproteins from the West Nile and dengue viruses with NK cells. J Immunol. 2009;183:2610–2621. doi: 10.4049/jimmunol.0802806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Homchampa P, Sarasombath S, Suvatte V, Vongskul M. Natural killer cells in dengue hemorrhagic fever/dengue shock syndrome. Asian Pac J Allergy Immunol. 1988;6:95–102. [PubMed] [Google Scholar]
- 32.Ito M, Okuno T, Fujii T, Mutoh K, Oguro K, Shiraishi H, Shirasaka Y, Mikawa H. ACTH therapy in infantile spasms: relationship between dose of ACTH and initial effect or long-term prognosis. Pediatr Neurol. 1990;6(4):240. doi: 10.1016/0887-8994(90)90114-G. [DOI] [PubMed] [Google Scholar]
- 33.Jamil KM, Khakoo SI. KIR/HLA interactions and pathogen immunity. J Biomed Biotechnol. 2011;2011:298348. doi: 10.1155/2011/298348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Jiang Y, Chen O, Cui C, Zhao B, Han X, Zhang Z, Liu J, Xu J, Hu Q, Liao C, Shang H. KIR3DS1/L1 and HLA-BW4-80I are associated with HIV disease progression among HIV typical Progressors and long-term Nonprogressors. BMC Infect Dis. 2013;13:405. doi: 10.1186/1471-2334-13-405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Jost S, Altfeld M. Control of human viral infections by natural killer cells. Annu Rev Immunol. 2013;31:163–194. doi: 10.1146/annurev-immunol-032712-100001. [DOI] [PubMed] [Google Scholar]
- 36.Kaiser BK, Barahmand-Pour F, Paulsene W, Medley S, Geraghty DE, Strong RK. Interactions between NKG2x immunoreceptors and HLA-E ligands display overlapping affinities and thermodynamics. J Immunol. 2005;174:2878–2884. doi: 10.4049/jimmunol.174.5.2878. [DOI] [PubMed] [Google Scholar]
- 37.Khor CC, Chau TN, Pang J, Davila S, Long HT, Ong RT, Dunstan SJ, Wills B, Farrar J, Van Tram T, Gan TT, Binh NT, Tri Le T, Lien Le B, Tuan NM, Tham NT, Lanh MN, Nguyet NM, Hieu NT, Van NVCN, Thuy TT, Tan DE, Sakuntabhai A, Teo YY, Hibberd ML, Simmons CP. Genome-wide association study identifies susceptibility loci for dengue shock syndrome at MICB and PLCE1. Nat Genet. 2011;43:1139–1141. doi: 10.1038/ng.960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kurane I, Hebblewaite D, Brandt WE, Ennis FA. Lysis of dengue virus-infected cells by natural cell-mediated cytotoxicity and antibody-dependent cell-mediated cytotoxicity. J Virol. 1984;52:223–230. doi: 10.1128/jvi.52.1.223-230.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kusnierczyk P. Killer cell immunoglobulin-like receptor gene associations with autoimmune and allergic diseases, recurrent spontaneous abortion, and neoplasms. Front Immunol. 2013;4:8. doi: 10.3389/fimmu.2013.00008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lanier LL, Ruitenberg JJ, Phillips JH. Functional and biochemical analysis of CD16 antigen on natural killer cells and granulocytes. J Immunol. 1988;141:3478–3485. [PubMed] [Google Scholar]
- 41.Laoprasopwattana K, Libraty DH, Endy TP, Nisalak A, Chunsuttiwat S, Vaughn DW, Ennis FA, Rothman AL, Green S. Antibody dependent cellular cytotoxicity in pre-secondary dengue virus serotype 3 (DV3) but not in DV2 infection plasma samples inversely correlated with viremia levels. J Infect Dis. 2007;195:1108–1116. doi: 10.1086/512860. [DOI] [PubMed] [Google Scholar]
- 42.Latchman Y, Mckay PF, Reiser H. Identification of the 2B4 molecule as a counter-receptor for CD48. J Immunol. 1998;161:5809–5812. [PubMed] [Google Scholar]
- 43.Libraty DH, Zhang L, Obcena A, Brion JD, Capeding RZ. Circulating levels of soluble MICB in infants with symptomatic primary dengue virus infections. PLoS One. 2014;9:e98509. doi: 10.1371/journal.pone.0098509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lim DS, Yawata N, Selva KJ, Li N, Tsai CY, Yeong LH, Liong KH, Ooi EE, Chong MK, Ng ML, Leo YS, Yawata M, Wong SB. The combination of Type I IFN, TNF-alpha, and cell surface receptor engagement with dendritic cells enables NK cells to overcome immune evasion by dengue virus. J Immunol. 2014;193:5065–5075. doi: 10.4049/jimmunol.1302240. [DOI] [PubMed] [Google Scholar]
- 45.Lindow JC, Borochoff-Porte N, Durbin AP, Whitehead SS, Fimlaid KA, Bunn JY, Kirkpatrick BD. Primary vaccination with low dose live dengue 1 virus generates a proinflammatory, multifunctional T cell response in humans. PLoS Negl Trop Dis. 2012;6:e1742. doi: 10.1371/journal.pntd.0001742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ljutic B, Carlyle JR, Filipp D, Nakagawa R, Julius M, Zuniga-Pflucker JC. Functional requirements for signaling through the stimulatory and inhibitory mouse NKR-P1 (CD161) NK cell receptors. J Immunol. 2005;174:4789–4796. doi: 10.4049/jimmunol.174.8.4789. [DOI] [PubMed] [Google Scholar]
- 47.Lunemann S, Martrus G, Holzemer A, Chapel A, Ziegler M, Korner C, Garcia Beltran W, Carrington M, Wedemeyer H, Altfeld M. Sequence variations in HCV core-derived epitopes Alter binding of KIR2DL3 to HLA-C *03:04 and modulate NK cell function. J Hepatol. 2016;65:252–258. doi: 10.1016/j.jhep.2016.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Maenaka K, Juji T, Stuart DI, Jones EY. Crystal structure of the human P58 killer cell inhibitory receptor (KIR2DL3) specific for HLA-CW3-related MHC Class I. Structure. 1999;7:391–398. doi: 10.1016/S0969-2126(99)80052-5. [DOI] [PubMed] [Google Scholar]
- 49.Mallone R, Funaro A, Zubiaur M, Baj G, Ausiello CM, Tacchetti C, Sancho J, Grossi C, Malavasi F. Signaling through CD38 induces NK cell activation. Int Immunol. 2001;13:397–409. doi: 10.1093/intimm/13.4.397. [DOI] [PubMed] [Google Scholar]
- 50.Manser AR, Weinhold S, Uhrberg M. Human KIR repertoires: shaped by genetic diversity and evolution. Immunol Rev. 2015;267:178–196. doi: 10.1111/imr.12316. [DOI] [PubMed] [Google Scholar]
- 51.Mathew A, Townsley E, Ennis FA. Elucidating the role of T cells in protection against and pathogenesis of dengue virus infections. Future Microbiol. 2014;9:411–425. doi: 10.2217/fmb.13.171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Moretta A, Sivori S, Vitale M, Pende D, Morelli L, Augugliaro R, Bottino C, Moretta L. Existence of both inhibitory (p58) and activatory (p50) receptors for HLA-C molecules in human natural killer cells. J Exp Med. 1995;182:875–884. doi: 10.1084/jem.182.3.875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Ozturk OG, Sahin G, Karacor ED, Kucukgoz U. Evaluation of KIR genes in recurrent miscarriage. J Assist Reprod Genet. 2012;29:933–938. doi: 10.1007/s10815-012-9811-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Parolini S, Bottino C, Falco M, Augugliaro R, Giliani S, Franceschini R, Ochs HD, Wolf H, Bonnefoy JY, Biassoni R, Moretta L, Notarangelo LD, Moretta A. X-linked lymphoproliferative disease. 2B4 molecules displaying inhibitory rather than activating function are responsible for the inability of natural killer cells to kill Epstein-Barr virus-infected cells. J Exp Med. 2000;192:337–346. doi: 10.1084/jem.192.3.337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Pende D, Parolini S, Pessino A, Sivori S, Augugliaro R, Morelli L, Marcenaro E, Accame L, Malaspina A, Biassoni R, Bottino C, Moretta L, Moretta A. Identification and molecular characterization of NKP30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J Exp Med. 1999;190:1505–1516. doi: 10.1084/jem.190.10.1505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Pessino A, Sivori S, Bottino C, Malaspina A, Morelli L, Moretta L, Biassoni R, Moretta A. Molecular cloning of NKP46: a novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicity. J Exp Med. 1998;188:953–960. doi: 10.1084/jem.188.5.953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Petitdemange C, Wauquier N, Jacquet JM, Theodorou I, Leroy E, Vieillard V. Association of HLA Class-I and inhibitory KIR genotypes in gabonese patients infected by chikungunya or dengue Type-2 viruses. PLoS One. 2014;9:e108798. doi: 10.1371/journal.pone.0108798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Petitdemange C, Wauquier N, Rey J, Hervier B, Leroy E, Vieillard V. Control of acute dengue virus infection by natural killer cells. Front Immunol. 2014;5:1–5. doi: 10.3389/fimmu.2014.00209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Petitdemange C, Wauquier N, Devilliers H, Yssel H, Mombo I, Caron M, Nkoghe D, Debre P, Leroy E, Vieillard V. Longitudinal analysis of natural killer cells in dengue virus-infected patients in comparison to chikungunya and chikungunya/dengue virus-infected patients. PLoS Negl Trop Dis. 2016;10:e0004499. doi: 10.1371/journal.pntd.0004499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Poli A, Michel T, Theresine M, Andres E, Hentges F, Zimmer J. CD56bright natural killer (NK) cells: an important NK cell subset. Immunology. 2009;126:458–465. doi: 10.1111/j.1365-2567.2008.03027.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Pozo D, Vales-Gomez M, Mavaddat N, Williamson SC, Chisholm SE, Reyburn H. CD161 (human NKR-P1A) signaling in NK cells involves the activation of acid sphingomyelinase. J Immunol. 2006;176:2397–2406. doi: 10.4049/jimmunol.176.4.2397. [DOI] [PubMed] [Google Scholar]
- 62.Rajagopalan S, Long EO. KIR2DL4 (CD158D): an activation receptor for HLA-G. Front Immunol. 2012;3:258. doi: 10.3389/fimmu.2012.00258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ramirez-De Los Santos S, Sanchez-Hernandez PE, Munoz-Valle JF, Palafox-Sanchez CA, Rosales-Rivera LY, Garcia-Iglesias T, Daneri-Navarro A, Ramirez-Duenas MG. Associations of killer cell immunoglobulin- like receptor genes with rheumatoid arthritis. Dis Markers. 2012;33:201–206. doi: 10.1155/2012/486320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Raulet DH. Interplay of natural killer cells and their receptors with the adaptive immune response. Nat Immunol. 2004;5:996–1002. doi: 10.1038/ni1114. [DOI] [PubMed] [Google Scholar]
- 65.Rothman AL. Immunity to dengue virus: a tale of original antigenic sin and tropical cytokine storms. Nat Rev Immunol. 2011;11:532–543. doi: 10.1038/nri3014. [DOI] [PubMed] [Google Scholar]
- 66.Rothman AL, Currier JR, Friberg HL, Mathew A. Analysis of cell-mediated immune responses in support of dengue vaccine development efforts. Vaccine. 2015;33:7083–7090. doi: 10.1016/j.vaccine.2015.09.104. [DOI] [PubMed] [Google Scholar]
- 67.Sconocchia G, Titus JA, Mazzoni A, Visintin A, Pericle F, Hicks SW, Malavasi F, Segal DM. CD38 triggers cytotoxic responses in activated human natural killer cells. Blood. 1999;94:3864–3871. [PubMed] [Google Scholar]
- 68.Seymour LA, Nourse JP, Crooks P, Wockner L, Bird R, Tran H, Gandhi MK. The presence of KIR2DS5 confers protection against adult immune thrombocytopenia. Tissue Antigens. 2014;83:154–160. doi: 10.1111/tan.12295. [DOI] [PubMed] [Google Scholar]
- 69.Shwetank, Date OS, Carbone E, Manjunath R. Inhibition of ERK and proliferation in NK cell lines by soluble HLA-E released from Japanese encephalitis virus infected cells. Immunol Lett. 2014;162:94–100. doi: 10.1016/j.imlet.2014.07.010. [DOI] [PubMed] [Google Scholar]
- 70.Thomas R, Matthias T, Witte T. Leukocyte immunoglobulin-like receptors as new players in autoimmunity. Clin Rev Allergy Immunol. 2010;38:159–162. doi: 10.1007/s12016-009-8148-8. [DOI] [PubMed] [Google Scholar]
- 71.Townsley E, Woda M, Thomas SJ, Kalayanarooj S, Gibbons RV, Nisalak A, Srikiatkhachorn A, Green S, Stephens HA, Rothman AL, Mathew A. Distinct activation phenotype of a highly conserved novel HLA-B57-restricted epitope during dengue virus infection. Immunology. 2014;141:27–38. doi: 10.1111/imm.12161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Townsley E, O’connor G, Cosgrove C, Woda M, Co M, Thomas SJ, Kalayanarooj S, Yoon IK, Nisalak A, Srikiatkhachorn A, Green S, Stephens HA, Gostick E, Price DA, Carrington M, Alter G, Mcvicar DW, Rothman AL, Mathew A. Interaction of a dengue virus NS1-derived peptide with the inhibitory receptor KIR3DL1 on natural killer cells. Clin Exp Immunol. 2016;183:419–430. doi: 10.1111/cei.12722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Vivian JP, Duncan RC, Berry R, O’connor GM, Reid HH, Beddoe T, Gras S, Saunders PM, Olshina MA, Widjaja JM, Harpur CM, Lin J, Maloveste SM, Price DA, Lafont BA, Mcvicar DW, Clements CS, Brooks AG, Rossjohn J. Killer cell immunoglobulin-like receptor 3DL1-mediated recognition of human leukocyte antigen B. Nature. 2011;479:401–405. doi: 10.1038/nature10517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Waggoner SN, Cornberg M, Selin LK, Welsh RM. Natural killer cells act as rheostats modulating antiviral T cells. Nature. 2012;481:394–398. doi: 10.1038/nature10624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Weiskopf D, Angelo MA, Bangs DJ, Sidney J, Paul S, Peters B, De Silva AD, Lindow JC, Diehl SA, Whitehead S, Durbin A, Kirkpatrick B, Sette A. The human CD8+ T cell responses induced by a live attenuated tetravalent dengue vaccine are directed against highly conserved epitopes. J Virol. 2015;89:120–128. doi: 10.1128/JVI.02129-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Welsh RM, Waggoner SN. NK cells controlling virus-specific T cells: rheostats for acute vs. persistent infections. Virology. 2013;435:37–45. doi: 10.1016/j.virol.2012.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Whitehorn J, Chau TN, Nguyet NM, Kien DT, Quyen NT, Trung DT, Pang J, Wills B, Van Vinh Chau N, Farrar J, Hibberd ML, Khor CC, Simmons CP. Genetic variants of MICB and PLCE1 and associations with non-severe dengue. PLoS One. 2013;8:e59067. doi: 10.1371/journal.pone.0059067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wong-Baeza I, Ridley A, Shaw J, Hatano H, Rysnik O, Mchugh K, Piper C, Brackenbridge S, Fernandes R, Chan A, Bowness P, Kollnberger S. KIR3DL2 binds to HLA-B27 dimers and free H chains more strongly than other HLA class I and promotes the expansion of T cells in ankylosing spondylitis. J Immunol. 2013;190:3216–3224. doi: 10.4049/jimmunol.1202926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Yossef R, Rosental B, Appel MY, Hershkovitz O, Porgador A. Upregulation of MHC class I expression following dengue virus infection: the mechanism at the promoter level. Expert Rev Anti-Infect Ther. 2012;10:285–287. doi: 10.1586/eri.12.7. [DOI] [PubMed] [Google Scholar]