

Abstract
Withanolides, and in particular extracts from Withania somnifera, have been used for over 3,000 years in traditional Ayurvedic and Unani Indian medical systems as well as within several other Asian countries. Traditionally, the extracts were ascribed a wide range of pharmacologic properties with corresponding medical uses, including adaptogenic, diuretic, anti-inflammatory, sedative/anxiolytic, cytotoxic, antitussive, and immunomodulatory. Since the discovery of the archetype withaferin A in 1965, approximately 900 of these naturally occurring, polyoxygenated steroidal lactones with 28-carbon ergostane skeletons have been discovered across 24 diverse structural types. Subsequently, extensive pharmacologic research has identified multiple mechanisms of action across key inflammatory pathways. In this chapter we identify and describe the major withanolides with anti-inflammatory properties, illustrate their role within essential and supportive inflammatory pathways (including NF-κB, JAK/STAT, AP-1, PPARγ, Hsp90 Nrf2, and HIF-1), and then discuss the clinical application of these withanolides in inflammation-mediated chronic diseases (including arthritis, autoimmune, cancer, neurodegenerative, and neurobehavioral). These naturally derived compounds exhibit remarkable biologic activity across these complex disease processes, while showing minimal adverse effects. As novel compounds and analogs continue to be discovered, characterized, and clinically evaluated, the interest in withanolides as a novel therapeutic only continues to grow.
Keywords: Autoimmune, Cancer, Inflammation, Neurodegenerative, NF-κB, Withaferin A, Withanolide
Introduction
Withanolides are a group of naturally occurring polyoxygenated steroidal lactones assembled on a C28 ergostane skeleton. The structural skeleton of withanolides usually varies in the nature and number of oxygenated substituents and the degree of unsaturation of the rings. Structurally diverse withanolides are typically classified based on the arrangement of the C-17 side chain into a major C-22/C-26 δ-lactone/lactol group and a minor C-23/C-26 γ-lactone/lactol group with few exceptions and about 90 % of these compounds possess ketone functionality at C-1 (Figs. 14.1, 14.2) [1–4]. Additionally, withanolides with a C-17 δ-lactone side chain, as shown in Fig. 14.1, can be further categorized into withanolides with an unmodified skeleton (e.g., withaferin A and withaperuvin B) and into those with a modified skeleton (e.g., physalin C and withametelinone) [1, 2, 4]. The occurrence of unmodified withanolides is more common in nature with approximately 580 of these naturally occurring withanolides reported in the family Solanaceae alone [1–3]. Structurally more complex withanolides with a modified skeleton both in the steroid nucleus and the side chain could possibly result from the biogenetic transformations of unmodified withanolides [3, 5].
Fig. 14.1.
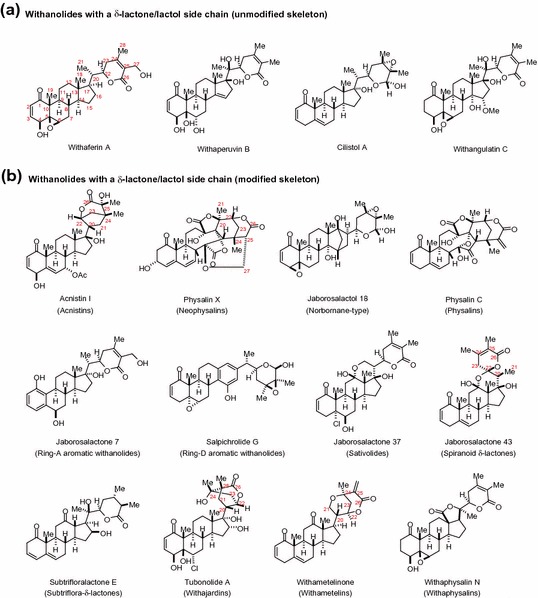
Withanolides with a δ-lactone ring
Fig. 14.2.
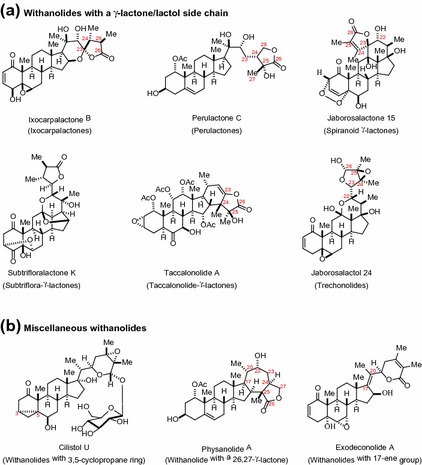
Withanolides with a γ-lactone ring and unclassified structural type
Withaferin A (WA), an archetype of this class was discovered from Withania somnifera (WS) or Ashwagandha in 1965 [6]. In the past 50 years, approximately 900 withanolides falling into 24 diverse structural types have been discovered [1, 3]. Withanolides are mainly distributed in various genera of Solanaceae, which includes Acnistus, Aureliana, Brachistus, Browallia, Datura, Deprea, Discopodium, Dunalia, Exodeconus, Hyoscyamus, Iochroma, Jaborosa, Larnax, Lycium, Mandragora, Nicandra, Physalis, Salpichroa, Saracha, Solanum, Trechonaetes, Tubocapsicum, Vassobia, Withania, and Witheringia [1–3, 7–9]. A minor population of withanolides has been isolated from other plant sources such as Dioscoreaceae, Fabaceae, Labiatae, Lamiaceae, Leguminosae, Myrtaceae, and Taccaceae [1–3] and interestingly from marine sources of Alcyoniidae family [10–12].
Structurally varied withanolides have received significant attention due to their versatile biological activities demonstrated in vitro and/or in vivo. These activities have been described as antitumor [7, 13–15], cytotoxic [16–20], apoptotic [21–23], anti-inflammatory [9, 10, 24–31], immunomodulating [32–34], antimicrobial [35–37], antistress [34], antioxidant [38], anti-neurodegenerative [39–41], radiosensitizing [42, 43], and insect antifeedant [44, 45]. Withaferin A, the most studied withanolide, possesses a wide array of the pharmacological activities described above and thus carries a great clinical potential for drug development [1, 4, 46–51]. Most notably, the antitumor and associated anti-inflammatory activities of WA and other withanolides results from targeting multiple signaling pathways simultaneously, particularly the nuclear factor kappa B (NF-κB), signal transducer and activator of transcription (STAT), and ubiquitin proteasome pathways (see Tables 14.1, 14.2) [52–57]. The potent biological activities of withanolides such as WA and tubocapsenolide A, especially the antitumor and anti-inflammatory properties have been attributed to the presence of key structural features such as an α,β-unsaturated ketone in ring A, a 5β,6β-epoxide in ring B, and a lactone side chain [1, 4, 7, 13, 30, 58–62]. Cysteine residues in the proteins are often implicated to react with these key electrophilic sites on the withanolide molecule [59, 60, 63, 64]. While other withanolides may possess α,β-unsaturated ketone and/or epoxide in some respect (e.g., paraminabeolides, capsisteroids, and chantriolides) and are bioactive, they are generally less potent than those withanolides possessing all three crucial functional groups.
Table 14.1.
Natural and semi-synthetic anti-inflammatory withanolides
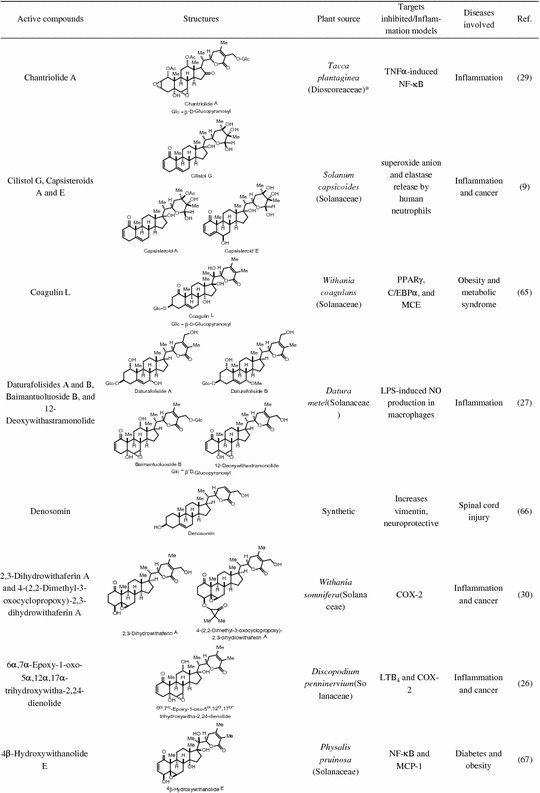
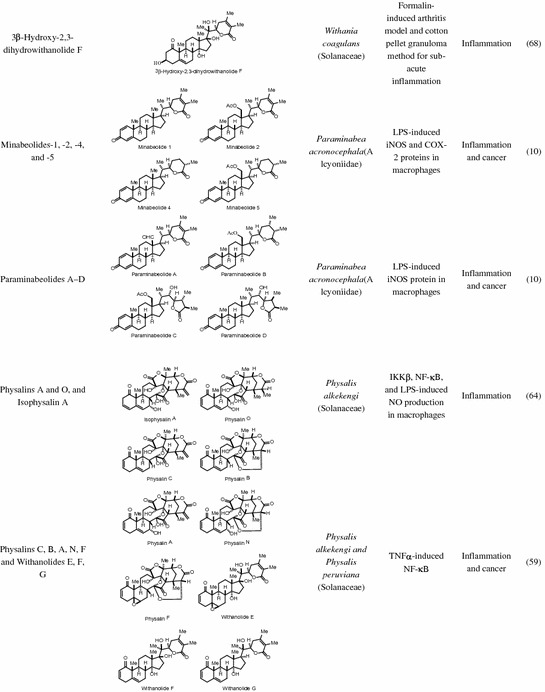

* Reference 28 had the genus Tacca assigned to family Taccaceae, which has been found in older texts but the APG II system has incorporated this genus into the family Dioscoreaceae. † Obtained from National Cancer Institute Developmental Therapeutics Program
Table 14.2.
Anti-inflammatory activity of withanolides in plant extracts
| Plant extract | Plant part | Active components | Effects | Therapeutic uses | References |
|---|---|---|---|---|---|
| Physalis peruviana (Solanaceae) | Fruits | Withanolides, polyphenols, and phytosterols | Antioxidant, anti-inflammatory, and renoprotective | Acute renal injury | [75] |
| Calyces | – | Anti-inflammatory and immunosuppressive effect on macrophages apoptopic (downregulates IL-6, TNF, and MCP-1) | Inflammation | [76] | |
| Withania somnifera (Solanaceae) | Roots | Withanolides such as withanolide A and alkaloids | Neuroprotective | Alzheimer’s disease | [77] |
| Roots | Withanolides, alkaloids, and flavanoids | Immunomodulatory, anti-inflammatory, and proapoptopic (downregulates IL-6, IL-1β, IL-8, Hsp70, and STAT-2) | Cancer | [78] | |
| Roots | – | Antioxidant, anti-inflammatory, and cytoprotective | Inflammatory bowel disease | [79] | |
| Leaves | – | Neuroprotective against glutamate neurotoxicity | Stroke and neurodegenerative disorders | [80] | |
| Withania coagulans (Solanaceae) | Fruits | – | Antioxidant, anti-inflammatory, antihyperglycemic, immunomodulatory, and renoprotective (downregulates IL-1β, IL-6, TNFα, IL-4, and IFN-γ) | Diabetes and associated renal complications | [81] |
Modulation of Inflammatory Cell Signaling Pathways by Withanolides
Inflammation is a complex immunological process by which our body fights against infection, cancer, or injury. The initial, acute stage of inflammation is mediated through the activation of immune cells, the resultant inflammatory cytokines and intracellular pathways. The initial immune mediators are CD4+ T-cells or T-helper (Th) cells and are classified as Th-1, Th-2, and Th-17. They play a crucial role in regulating the cellular and humoral immune responses through recognition of antigens presented on antigen presenting cells via the major histocompatibility complex II. Th-1 cells promote the cellular immune response (macrophages) and primarily produce interferon (IFN)-γ, tumor necrosis factor (TNF)-β, and interleukin (IL)-2, whereas Th-2 cells promote the humoral immune system (antibodies) and primarily produce IL-4 and IL-10, and Th-17 cells help recruit neutrophils early in the adaptive response, produce IL-17 cytokine, and are involved in many autoimmune diseases [82]. The alteration of normal homeostasis of any of the Th cells through aberrant recognition of self or dysregulated production of cytokines plays a major role in the formation of chronic inflammatory or autoimmune/immunomodulatory diseases. Excess cytokine production leads to the over activation of multiple downstream inflammatory pathways, including Janus kinase (JAK)/STAT, NF-κB, phosphatidylinositol-3-kinase (PI3K)/Akt, and mitogen-activated protein kinase (MAPK). A number of withanolides have demonstrated significant immunomodulatory effects, including WS extract (primarily aqueous extract), withanolide A, physalins and coagulins; however, both immuno-stimulatory and inhibitory actions have been attributed to different withanolides. WS extract (primarily withanolide A and 2,3 dihydro-3-sulphonile withanone components) is able to shift the immune response toward Th-1 polarization, activate cytotoxic natural killer cells [83, 84], and recover depleted T-cells and increase expression of Th-1 cytokines IL-2 and IFN-γ in models of stress [32, 34, 85, 86]. Conversely, coagulins isolated from Withania coagulans and primarily coagulin-H have demonstrated immunosuppressive effects similar to prednisolone, inhibiting stimulated T and B-cell lymphocyte proliferation, and Th-1 cytokine production possibly through IL-2 receptor binding [87, 88]. Similar to coagulins, physalins B, F, G, and H isolated from Physalis angulata have demonstrated immunosuppressive properties. Physalins B, F, and G showed inhibition of lipopolysaccharide (LPS)-activated macrophages along with their cytokine (TNF-α, IL-6 and IL-12) and nitric oxide (NO) production, whereas concanavalin A-induced T-cell proliferation and cytokine production in a mechanism distinct from dexamethasone [89, 90]. Physalin H has demonstrated polarization to Th-2 cells with inhibition of Th-1 cytokines IL-2 and IFN-γ, increased Th-2 cytokines IL-4 and IL-10, and induction of the heme oxygenase-1 [91].
There are strong preclinical and clinical studies that demonstrate that inflammation initially started by immune cell mediators may persist chronically, resulting in ongoing stimulation of inflammatory mediators and regulatory pathways that contribute to the pathogenesis of chronic diseases including cardiovascular, neurologic, and pulmonary diseases, as well as cancer, diabetes, and obesity [92–94]. As with acute inflammation, chronic inflammation is mediated through various signaling factors, which include proinflammatory cytokines such as TNFα, IL-1, IL-6, IL-8, IL-12, NO, adhesion molecules, and chemokines [95]. Additionally, transcription factors that regulate the expression of inflammatory mediators such as NF-κB, activator protein 1 (AP-1), peroxisome proliferator-activated receptor (PPAR)-γ, STAT3, hypoxia inducible factor-1 (HIF-1), β-catenin/Wnt, hedgehog, and nuclear factor erythroid 2-related factor 2 (Nrf2) have been linked to chronic diseases. The DNA-binding capacity of these transcription factors is modified by several signaling cascades such as JAK/STAT, MAPKs, PI3K/Akt/mechanistic target of rapamycin (mTOR), and ubiquitin proteasome system [96]. These signaling pathways have a wide range of functions and show complex crosstalk depending on the cell type and the chronic disease involved. Withanolides have emerged recently as potential therapeutics for chronic diseases due to their unique ability to modulate multiple signaling pathways. In this chapter, we will discuss several different withanolides and their seemingly broad mechanism of action in modulating key molecular pathways that connect inflammation and chronic diseases.
Withanolides Modulate the NF-κB Pathway
The NF-κB family of transcription factors plays a prominent role in the immune system and inflammation. In response to ligation by Toll-like receptors (TLRs) and IL-1 receptor family members (B-cell and T-cell receptors), NF-κB regulates the expression of several factors such as inhibitor of apoptosis protein-1 (IAP1), inducible nitric oxide synthase (iNOS), cytokines, cyclooxygenase (COX)-2, prostaglandins, growth factors, and effector enzymes [97–99]. NF-κB is activated by inflammatory cytokines, stress, free radicals, radiation, growth receptors, and TNFα leading to transcriptional regulation of several genes that are involved in proliferation, inflammation, cellular survival, apoptosis, angiogenesis, and differentiation [100–103]. The mammalian transcription NF-κB family of proteins includes RelA (p65), RelB, NF-κB2 (p52), c-Rel, and NF-κB1 (p50). In the absence of stimuli, inactive forms of NF-κB proteins are present in the cytoplasm due to their interaction with several inhibitors of kappaB (IκB) proteins. Upon exposure to stimuli, NF-κB is activated either through canonical or noncanonical pathways by regulatory IκB kinases (IKK) such as IKKα, IKKβ, and IKKγ. In the most common classical pathway, IKK phosphorylation leads to IκBα phosphorylation at serine 32 and serine 36, followed by phosphorylation, nuclear localization, DNA binding of p65–p50 complex, and transcriptional activation of NF-κB responsive genes [104]. Activation of NF-κB is essential for the survival and expression of inflammatory mediators. Hence, constitutively active NF-κB is associated with several inflammation-mediated chronic diseases such as cancer, neurological, and metabolic disorders [98].
A number of naturally occuring withanolides such as chantriolide A, physalins A, B, C, and O, viscosalactone B, WA, withanolides D, E, F and withaferin A 4,27-diacetate, a diacetyl derivative of WA have been reported to modulate the regulation of NF-κB [24, 29, 34, 52, 54, 59, 64]. Several studies have examined the beneficial effect of inhibiting transcriptional activity of NF-κB in chronic inflammatory diseases including cancer [24, 55, 56, 72]. In a study led by Aggarwal et al., various withanolides isolated from the leaf extract of WS along with their semi-synthetic acetylated derivatives were tested for their inhibitory effects on NF-κB activation induced by activators such as cigarette smoke condensate, TNF, doxorubicin, and IL-1β [54]. Withaferin A and viscosalactone B along with their 4,27-diacetyl derivative inhibited TNF-induced NF-κB activation in human myeloid leukemia KBM-5 cells, whereas physagulin D and related glycosidic withanolides were inactive. The mechanism through which the above withanolides blocked NF-κB was through the inhibitory effects of activated IκBα, with subsequent phosphorylation of IκBα and p65, followed by prevention of IκBα degradation. Blocking the degradation of IκBα in turn prevents nuclear localization of p65, and activation of NF-κB-dependent gene products such as Bcl-2-related protein A1 (Bfl-1/A1), IAP-1, c-FADD-like cFLIP, ICAM1, and COX-2 [54]. To further understand the SAR, the authors examined TNF-induced NF-κB activity after treatment of human myeloid leukemia KBM-5 cells with various withanolides isolated from the leaf extract of WS along with their synthetically modified acetylated derivatives by electrophoretic mobility shift assay (EMSA). This analysis pointed toward the importance of the α,β-unsaturated ketone in ring A as a required structure for the potent inhibition of NF-κB activity [54].
Physalins A, L, G and O, and isophysalin A isolated from Physalis alkekengi were evaluated for their anti-inflammatory properties and Physalins A, O and isophysalin A showed significant inhibition of LPS-induced NO production in macrophages [64]. Based on their structural features characterized by the presence of either an α,β-unsaturated ketone in ring A (physalin O) or an α,β-unsaturated ester in lactone side chain featuring an exomethylene group (physalin A and isophysalin A), these physalins were able to conjugate with glutathione as identified by ultra-performance liquid chromatography tandem mass-spectrometry (UPLC-MS/MS) analysis. Furthermore, peptide mapping and sequencing of alkylated IKKβ using micrOTOF-MS revealed alkylation of six cysteine residues on IKKβ by physalin A indicating IKKβ as a potential target for its anti-inflammatory mechanism of action. In another related and complementary study, SARs were performed on a library of withanolides including the physalins. This study revealed the importance of the oxygenated right-side partial structure (including the lactone side chain) and the 5β,6β-epoxide, or C5–C6 olefin in the B-ring for the inhibition of NF-κB activation [59]. Withanolides and physalins with 5β,6β-epoxide inhibited NF-κB signaling through prevention of IκBα degradation and p65 nuclear localization, whereas those with C5–C6 olefin inhibited NF-κB function by blocking p65/p50 dimer binding to DNA [59].
Chantriolide A, one of the eight compounds isolated from Taccaplantaginea exhibited potent inhibition of TNFα-induced NF-κB transcriptional activity in human hepatocellular carcinoma (HepG2) cells in an NF-κB-luciferase assay [29]. In another study, withanolide D and WA from WS inhibited angiogenesis through blocking of NF-κB activity by suppressing proteasome-mediated ubiquitin degradation of IκBα in human umbilical vein endothelial cells [52]. Additionally, 4β-hydroxywithanolide E was shown to inhibit inflammatory response in adipocytes via inhibition of NF-κB transcriptional activity [67]. Inhibition of IKKβ activation by 4β-hydroxy withanolide E through suppression of IKKβ phosphorylation was mechanistically distinct from the NF-κB inhibition observed for WA, where the induction of IKKβ over-phosphorylation was shown to inhibit IKKβ activation [24]. Moreover in vivo, 4β-hydroxy withanolide E demonstrated an improvement of impaired glucose tolerance suggesting its potential role for the treatment/prevention of metabolic disorders including type 2 diabetes [67]. Overexpression of CCR7 in metastatic breast cancer cells has been associated with lymph node metastasis [69]. In breast cancer cells MDA-MB-231, tubocapsanolide A inhibited TAK1 to suppress NF-κB-mediated CCR7 expression leading to the inhibition of lymphatic invasion of breast cancer in vitro and in vivo.
In addition to the inhibition of NF-κB activation, WA and several other withanolides have been shown to directly block the expression of LPS- or TNFα-induced NF-κB-regulated inflammatory genes such as iNOS, COX-1, COX-2, and NO [10, 13, 14, 25, 26, 64, 74]. Nitric oxide is a small molecule that regulates MMPs and joints extracellular matrix, and is modulated through iNOS. COX-1 and COX-2 convert arachidonic acid to prostaglandins, which in turn cause a significant inflammatory response. COX-1 is constitutively expressed in most cell types, and is responsible for maintenance of normal physiologic function, whereas COX-2 is inducible in response to proinflammatory cytokines [26]. Nair and co-workers were the first to demonstrate the role of withanolides in inhibiting COX enzymes and provide insight into their anti-inflammatory mechanism [30]. Withaferin A, viscosalactone B, 2,3-dihydrowithaferin A, and 4-(2,2-dimethyl-3-oxocyclopropoxy)-2,3-dihydrowithaferin A were shown to inhibit COX-2 enzyme but not COX-1. Interestingly, during this study it was observed that the presence of a double bond between C-24 and C-25 in the lactone ring was essential for COX inhibitory activities and a withanolide lacking this unsaturation in the lactone ring was found to be inactive against both COX-1 and COX-2 enzymes [30].6α,7α-Epoxy-1-oxo-5α,12α,17α-trihydroxywitha-2,24-dienolide from Discopodiumpenninervium was found to inhibit COX-2 and leukotriene B4 (LTB4) but was inactive against the COX-1 enzyme [26]. Like other withanolides, withanolide sulfoxide, a sulfoxide dimer of WA was highly selective in inhibiting COX-2 compared to COX-1[74].
Daturafolisides A and B along with other known withanolides from Datura metel were shown to exhibit significant reduction in NO production in LPS-induced RAW 264.7 macrophage cells [27]. Of note, both of these compounds lack the α,β-unsaturated ketone in ring A and the 5β,6β-epoxide in ring B, however, they do possess a δ-lactone side chain. Additionally, withanolides such as paraminabeolides and minabeolides obtained from a marine source were found to inhibit LPS-induced iNOS expression in RAW 264.7 macrophages, with minabeolides also effectively inhibiting COX-2 expression [10].
Withanolide Modulation of the JAK/STAT Pathway
The JAK/STAT pathway is a key-signaling mediator of cytokines and growth factors such as platelet-derived growth factor (PDGF), epidermal growth factor (EGF), IL6, as well as oncogenic proteins [105]. Activation of STAT proteins depends on their binding to cytokines and growth factor receptors on the plasma membrane followed by tyrosine phosphorylation either directly by receptor tyrosine kinases (RTKs) or by non RTKs such as JAK or Src [106, 107]. Upon phosphorylation, cytoplasmic STAT proteins undergo dimerization via reciprocal SH2-domain/phosphotyrosine interactions followed by translocation to the nucleus for DNA binding to STAT-specific response elements leading to transcriptional activation. There are eight known STAT proteins (STATs 1A, 1B, 2, 3, 4, 5A, 5B, and 6) that play diverse biochemical roles in several important processes such as survival, proliferation, apoptosis, invasion, immune response, inflammation, and angiogenesis [105, 108, 109]. Among the eight isoforms STAT3 and STAT5 are constitutively activated in several solid tumors, including lung, bladder, breast, colon, as well as in hematological malignancies [108]. Additionally, STAT3 is also interconnected with the NF-κB pathway and plays a central role in inflammation [107].
Several chronic diseases including cancer have been shown to induce aberrant regulation of STAT3. This transcription factor promotes oncogenic processes such as invasion, metastasis, and angiogenesis as several genes involved in these mechanisms such as cyclin D1, c-Myc, vascular endothelial growth factor (VEGF), mucin 1 (MUC-1), twist family BHLH transcription factor (TWIST) are all regulated by STAT3 [110–114]. Studies have investigated the role of WA from WS in regulating STAT proteins in different cancer models including colon cancer, breast cancer, multiple myeloma, and neuroblastoma [53, 57, 115, 116]. In breast cancer, WA treatment of triple negative MDA-MB-231 and hormonally active MCF-7 cells effectively decreased the constitutive as well as the IL-6 inducible phosphorylation of JAK 2 and its downstream target STAT3 thereby inhibiting the transcriptional activity of STAT3 [115]. In renal carcinoma Caki cells, WA had a similar effect and also downregulated the expression levels of anti-apoptotic proteins that are regulated by STAT3 like Bcl-2, cyclin D1, survivin, and Bcl-xL, thereby inducing apoptosis [116]. Docking studies showed that WA not only downregulates the phosphorylation of STAT3 at the tyrosine Y705, but also prevents dimerization of STAT3 [57]. In addition to cancer cells, WA also is able to suppress the phosphorylation of STAT1/3 in murine BV2 microglial cells, leading to a reduction in LPS-induced COX-2 downregulation and PGE2 production [73]. Withacnistin, an unmodified withanolide blocked both IL-6 as well as EGF-stimulated binding of STAT3 and STAT5 to gp130 and EGF-receptor (EGFR) in MDA-MB-468 breast cancer cells. This resulted in subsequent downregulation of STAT3 tyrosine phosphorylation and decreased nuclear translocation. Further evaluation of STAT3-DNA binding and transcriptional activity after Withacnistin treatment revealed blocking of both DNA binding as well as STAT3 reporter activity. This in turn caused downregulation of STAT3 target genes Bcl-xL and MCL-1 resulting in apoptosis [71].
Modulation of the AP-1 Pathway by Withanolides
The transcription factor AP-1, which plays a key role in the inflammatory response is implicated in several diseases such as cancer, psoriasis, inflammatory bowel disease (IBD), rheumatoid arthritis (RA) and fibrosis [117]. The AP-1 complex consists of homo and hetero dimers of Jun (JunD, C-Jun, and JunB) and the Fos (FosB, C-Fos, Fra-1 and Fra-2) family of proteins [118, 119]. Cytokines, chemokines, hormones, and growth factors as well as external stress factors are known to activate AP-1 signaling. The AP-1 complex translocates to the nucleus in response to stress signaling cascades, such MAPKs and c-Jun terminal kinases [120]. This in turn leads to activation of AP-1 and regulates multiple functions such as differentiation, transformation, proliferation, and survival [121]. The crude ethanol extract of WS has been shown to inhibit the nuclear localization of both AP-1 and NF-κB in LPS-activated peripheral blood mononuclear cells (PBMC) of both normal and RA patients, as well as synovial fluid mononuclear cells (SFMC) of RA patients. This in turn led to decreased downstream transcription target genes such as MMPs, COX-2, and iNOS, all of which are known mediators of RA [122].
Withanolides Can Modulate the PPARγ Pathway
PPARγ was first discovered in adipocyte differentiation and lipid metabolism and is one of three members in this nuclear receptor family of transcription factors [123]. The other members of the PPARs in mammals are PPARα and PPARβ/δ. The PPARs activate several genes involved in inflammation, adipogenesis, lipid metabolism, glucose metabolism, cellular differentiation, development, and tumorigenesis via binding of the PPAR/retinoid X receptor heterodimer to PPAR-responsive regulatory elements [124, 125]. PPARγ plays a key role in inflammation through modulation of proinflammatory transcription factors such as NF-κB and AP-1 [113]. Treatment of 3T3-L1 adipocytes with WA resulted in phosphorylation of extracellular signal-regulated kinase (ERK), followed by decreased expressions of PPARγ leading to altered levels of Bcl2 and Bax expression, induction of apoptosis, and inhibition of adipogenesis [126]. In addition to WA, other withanolides such as plantagiolide J and I isolated from Taccaplantaginea [29] and coagulin-L isolated from Withania coagulans [65] also modulate PPARγ transcriptional activity.
Modulation of the Hsp90 Pathway by Withanolides
Heat shock proteins (Hsp) are ATP-dependent ubiquitously expressed molecular chaperones that are involved in the folding, assembly, maintenance, and transport of key regulatory proteins involved in numerous signaling pathways in the cell. Several environmental and physiological stimuli such as hypoxia, oxidative damage, inflammation, infection, and elevated temperature induce the expression of these highly conserved molecular chaperone family of proteins as a protein homeostasis and survival response [70, 114]. The Hsp90 family of proteins (Hsp90α, Hsp90β, GRP94, and TRAP1) form a large complex with other co-chaperones such as cdc37, HSP70-HSP90 organizing protein, p27, Hsp32, and Hsp70. This complex then stabilizes and maintains functional activity of proteins/kinases in many key signaling pathways, such as PI3K/Akt/mTOR, p38/MAPK, and NF-κB, all of which play critical roles in inflammation, chronic inflammatory diseases, and oncogenesis. Through inhibition of Hsp90, and therefore inhibition of its oncogenic chaperone clients, cancer cells undergo apoptosis [124, 125].
Several studies have shown that withanolides such as WA, withalongolides A and B, tubocapsenolide A, and some of their synthetically modified analoges such as withalongolide A triacetate and withalongolide B diacetateare are able to target multiple cancers such as colon, prostate, brain, breast, head and neck, skin, adrenal, and thyroid both in vitro and in vivo [17, 20, 21, 53, 63, 70, 127–137]. Withanolides such as WA and withalongolide A are known to block Hsp90 chaperone function through blocking the Hsp90/cdc37 complex, and induction of thiol-mediated oxidative stress [63, 138, 139]. The Hsp90/cdc37 complex facilitates active conformation of client kinases in particular, such as Akt, cyclin-D1, raf-1, and cdk4. Blocking this complex leads to dysfunctional or proteasome mediated degradation of these kinases within multiple oncogenic, pro-survival, and proliferative kinase cascades (p38/MAPK, PI3K/Akt/mToR, NF-κB pathways), which ultimately leads to cancer cell apoptosis [136, 138]. In addition to targeting the bulk cancer cell population, WA and withalongolide A triacetate may also target the cancer stem cell (CSC) population. These CSCs comprise a small fraction of cancer cells, and are characterized by their tumor initiating and self-renewal capacity. WA and withalongolide A triacetate block several developmental pathways such as Wnt/β-catenin, notch, and NF-κB, as well as vimentin and VEGF, all of which are important in inflammation, self-renewal, and CSC epithelial-to-mesenchymal transition [115, 140–145].
Withanolide Modulation of Nrf2 Pathway
Nuclear factor erythroid 2 related factor 2(Nrf2) is a transcription factor that regulates genes involved in redox homeostasis, inflammation, energy metabolism and cellular growth [146]. Under normal homeostatic conditions, Nrf2 is anchored in the cytoplasm as a complex with Kelch like ECH-associated protein-1 (Keap1), which facilitates ubiquitin mediated proteasome degradation of Nrf2 and decreased expression of Nrf2 target genes. However, in response to stimulation by growth factors, electrophilic stressors, and changes in redox signal, Nrf2 ubiquitination is disrupted and levels increase rapidly. Nrf2 translocates to the nucleus and upregulates expression of proteins involved in glutathione and thioredoxin-based antioxidant defense, drug metabolism and efflux, and proteins associated with heme and iron metabolism [147]. Nrf2 is engaged in crosstalk with several signaling pathways that play a critical role in the pathogenesis and progression of chronic diseases, including NF-κB, PI3K, MAPK, glycogen synthase kinase-3β, and notch [146, 147]. Molecular docking studies have shown that both WA and withanone interact with the amino acids Ala 69, Gln 75, and Phe 71 of Nrf2 [148]. In another study, WA induced reactive oxygen species (ROS) that activated JNK and stabilized Nrf2 that resulted in activation of NADPH quinone oxidoreductase and Tap73 transcriptional function leading to apoptosis of cancer cells [149]. WA was also shown to inhibit NFκB, AP-1, and Nrf2 in adriamycin-resistant human myelogenous erythroleukemic K562/Adr cells in a dose-dependent manner [72]. Moreover, compared to other tested natural products such as quercetin, only WA overcomes attenuated caspase activation and blocking of apoptosis in K562/Adr cells [72].
Modulation of the HIF-1 Pathway by Withanolides
Under normal oxygen conditions, the HIF-1 α protein is synthesized at a high rate and rendered transcriptionally inactive due to immediate hydroxylation-dependent proteasome/ubiquitin degradation by the VHL E3 ligase. However, when hypoxia is induced through impaired cellular oxygen balance, hydroxylase activity is downregulated, HIF-α protein is stabilized, and HIF-1 is activated [150]. Transcriptional activation of HIF-1 upregulates several genes that control glycolytic metabolism, angiogenesis, invasion, metastasis, and cell survival, such as VEGF, MMPs, stromal cell-derived factor-1, e-cadherin, chemokine receptor 4, EGF, and transforming growth factor beta (TGF-β) 3 [151–155]. Crosstalk between NF-κB and HIF pathways has been shown to be associated with several chronic inflammatory diseases such as cancer, RA, asthma, and chronic obstructive pulmonary diseases [156]. In solid tumors, the availability of oxygen within the tumor decreases as distance from blood vessels increases resulting in the creation of hypoxic regions [157]. This is known to be responsible in part for therapy resistance and metastatic spread [158]. Although, no study thus far directly demonstrates inhibition of HIF-1 transcriptional activation by withanolides, a few note downregulation of migration-promoting HIF-mediated genes such as VEGF, heterogeneous nuclear ribonucleoprotein K (hnRNP-K) and MMPs, which lead to restriction of angiogenesis and metastasis [159].
Withanolides for Clinical Development
As discussed above, studies show the ability of withanolides to target multiple interconnected signaling pathways such as PI3K/Akt/mTOR, JAK/STAT, AP-1, NF-κB, PPARγ, Nrf2 and MAPK. Withanolides target these pathways through multiple mechanism, such as blocking Hsp90-Cdc37 co-chaperone interaction, targeting Akt and its downstream pathways, and induction of thiol-mediated oxidative stress (summarized in Fig. 14.3). Each of these mechanisms and pathway interactions play important roles in the development of chronic inflammatory diseases. Building on the studies identifying mechanisms of action of withanolides, we will discuss the clinical importance of withanolides on inflammatory mediated diseases including chronic inflammatory/autoimmune, cancer, and neurologic.
Fig. 14.3.
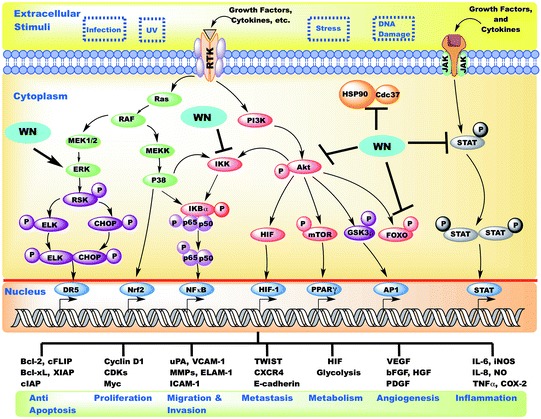
Schematic diagram representing modulation of various inflammatory pathways by withanolides
Application of Withanolides in Inflammatory/Autoimmune Diseases
Osteoarthritis and Rheumatoid Arthritis
In Ayurvedic medicine, withanolides are frequently used to treat both osteoarthritis (OA) and RA and there are several anti-inflammatory pathways affected by withanolides that contribute to chondro protection and treatment. The previously described NF-κB pathway plays a key role in arthritic inflammation, as do the downstream effectors NO via iNOS, COX-1, and COX-2 enzymes [160]. NO has been implicated in chondrocyte apoptosis in RA [161] and WS extracts have demonstrated reductions in NO in murine macrophage cell lines [122] and human chondrocytes [162]. Nonsteroidal anti-inflammatory drugs nondiscriminately inhibit COX enzymes, which with chronic use increases the risk of upper GI ulcers. Withanolides have been demonstrated to exhibit significant selective COX-2 inhibition while sparing COX-1 [13, 26, 163]. Additionally, along a parallel proinflammatory pathway with arachidonic acid, withanolides inhibited the production of LTB4 [26].
Another important mediator of arthritis is the formation of ROS leading to oxidative stress, and the protective role of Nrf2 pathway in glutathione and thioredoxin-based antioxidant defense [164]. However, the exact mechanism by which withanolides act within this pathway is still unclear as studies have demonstrated that withanolides are both ROS protective through inhibition of lipid peroxidation [13, 163] and can induce oxidative stress in rabbit chondrocytes [165, 166].
Therapeutic Benefits of Withanolides for Osteoarthritis and Rheumatoid Arthritis In Vivo
There have been several studies indicating that withanolides have a chrondroprotective aspect. WS extract has been shown in animal models to significantly reduce the effects of collagenase on bovine Achilles tendons with even a suggestion of collagen stabilization [167]. This has also been demonstrated with inhibition of the gelatinase activity of collagenase type 2 enzyme in vitro [168]. Several collagen-induced arthritic rat models have noted significant amelioration of paw and ankle arthritis, oxidative stress, degradation of cartilage, and improvements in functional recovery and radiological score [24, 169–171]. Aqueous extracts of WS have demonstrated a significant dose-dependent reduction in adjuvant-induced hind foot pad thickness, reductions in immune complement activity [172], as well as reduced arthritic index, autoantibodies, and CRP levels with results comparable to methotrexate treatment. These rats similarly demonstrate reductions in oxidative stress through decreased lipid peroxidation, glutathione-S-transferase activity, and an increase in glutathione content and ferric-reducing ability [170]. In a monosodium urate crystal-induced rat model of gout, WS extract demonstrated a significant reduction in paw pad volume down to that of normal controls as well as analgesic and antipyretic effects without any evidence of gastric injury [173]. Despite the conflicting reports of WA increasing oxidative stress and reducing type II collagen through induction of COX-2 by microRNA-25 [166], the majority of evidence continues to support the chondroprotective and analgesic properties of withanolides in arthritis.
Clinical Activity of Withanolides for Osteoarthritis and Rheumatoid Arthritis
On the basis of the long-standing use of WS extract in the treatment of arthritis in Ayurvedic medicine and the support from animal models, there have been several human tissue and blood studies performed in recent years. WS root powder extracts were given to patients with chronic OA and mild to moderate (Grade 1–2) articular cartilage and a portion of the cartilage was explanted for analysis. Half of the cartilage samples had a significant short-term chondroprotective response as demonstrated by significant decreases in proteoglycan release [168]. Further research on the explanted cartilage also demonstrated a significant reduction in NO production as an inflammatory regulator molecule in 50 % of the patient samples [162].
In RA, WS crude ethanol extract significantly inhibited LPS-induced expression of multiple proinflammatory cytokines from PBMCs and SFMCs taken from RA and normal patients, including TNF-α, IL-1β, and IL-12 p40. The reduction in inflammatory cytokines may have resulted from suppression of LPS-activated NF-κB and AP-1, in addition to inhibition of AP-1 nuclear translocation and LPS-induced phosphorylation of IκBα [122].
Although there have been several human trials for OA and RA, they were developed using combinations of Ayurvedic drugs. Combination therapy RA-11 (that includes WS, Boswelliaserrata, Zingiberofficinale, and Curcumalonga) was given to patients in a randomized, placebo-controlled OA trial. Results demonstrated a mean reduction in pain (using visual analog scale) and in the modified WOMAC index (Western Ontario McMaster University OA Index) at 16 and 32 weeks compared to placebo without any significant in adverse events [174]. A pilot prospective study of the combination Ashwagandha and SidhMakardhwaj in RA demonstrated a significant ACR20 (20 % improvement in tender joint/swollen join counts and 20 % reduction in 3 of 5 areas, physician global assessment score, patient global assessment score, pain assessment score, patient self-assessed disability index score, and ESR) response in 56 % of patients, and a moderate response in 40 % of patients by EULAR criteria (European League Against Rheumatism) [175]. Although only part of a combination therapy and the effects of WS alone cannot be determined, these trials suggest that withanolides may have chondroprotective, anti-inflammatory, and analgesic characteristics in human joints without significant adverse events.
Applications in Systemic Lupus Erythematous and Inflammatory Bowel Disease
The pathogenesis of systemic lupus erythematous (SLE) and IBD is rooted in chronic, aberrant activation of the immune system and inflammatory pathways. SLE is a complex B-cell mediated autoimmune disease characterized by the generation of autoantibodies against nuclear antigens (antinuclear antibody) and a type III hypersensitivity reaction (antibody–antigen complexes) leading to chronic inflammation and deposition of the antibody–antigen complexes within small vessels of end organs, such as the kidneys, skin, and brain [176]. In IBD, although chronic, intermittent inflammation is a cornerstone of disease progression, the pathogenesis is more complex, involving genetic susceptibilities, perturbation of mucosal physiology/epithelial barrier, and homeostasis of intestinal innate immune system [177]. For both SLE and IBD, current therapies are based upon anti-inflammatory, immunosuppressive, or immune/biologic therapies. Ongoing therapeutic research is focused on identifying additional immune targets including modulation of the Th-1, Th-2 and Th-17 responses, inflammatory cytokines, and downstream inflammatory pathways [178, 179]. As discussed previously, withanolides including Withania coagulin, and Physalis angulata have demonstrated significant immunomodulatory effects on Th cells and inflammatory cytokines making these compounds an exciting new area of therapeutic development for inflammatory and autoimmune diseases.
Therapeutic Benefits of Withanolides for SLE and IBD In Vivo
The recent characterization of the effects of withanolides on immunomodulation has given rise to several in vivo animal models of SLE and IBD. Aqueous WS extracts were used to treat a nonautoimmune pristane-induced model of SLE in mice, which develop SLE-like symptoms, including autoantibody production, proteinuria, and nephritis. The prophylactic effects of WS extract were characterized by orally administering WS for one month prior and six months following pristane injections. WS treatment resulted in significant reductions in ROS formation within intraperitoneal macrophages, as well as reductions in inflammatory cytokines (IL-6 and TNF-α), and decreased inflammation within the kidneys, liver, and lung, however, no reduction in the humoral response autoantibodies or immune complex deposition [180, 181].
Using trinitrobenzylsulfonic acid (TNBS)-induced IBD model in rats, a rectal gel formulation from aqueous WS extract was administered from the fourth to fourteenth day, and demonstrated a dose-dependent inhibition of lipid peroxidation up to 96 %, hydrogen peroxide scavenging up to 82 %, and NO scavenging ability similar to curcumin. Histopathology demonstrated a significant decrease in colonic injury with WS treatment along with improvement in macroscopic damage when compared to untreated controls. WS-treated rats also retained their weight with improved recovery following induction of the TNBS model [79].
While the animal data for these inflammatory and autoimmune diseases remains relatively sparse, there has yet to be any major human studies using withanolides in these diseases. In the meantime, the immunomodulatory effects of withanolides continue to be characterized and hold therapeutic potential for the treatment of a number of autoimmune and immune-modulating diseases.
Application of Withanolides in Cancer Therapy
The discovery that most withanolides have cytotoxic capabilities against a wide range of cancers has initiated a large breadth of research into the cytotoxic mechanisms and potential therapeutic benefits of withanolides for cancer treatment and prevention. As previously described and depicted in Fig. 14.3, withanolides inhibit multiple aspects of inflammatory pathways. While these inflammatory pathways have complex interactions between one another, they also interact with proliferative and oncogenic pathways. Several key mechanisms of cytotoxicity from withanolides have been described and these include: (a) induction of oxidative stress, (b) inhibition of proteasome mediated ubiquitin degradation of IκBα (leading to inhibition of NF-κB and its downstream effectors AP-1 and Nrf-2), (c) inhibition of transcription factor STAT3, (d) inhibition of Hsp90 (by blocking interaction of the Cdc37 co-chaperone with Hsp90) with resultant Hsp90 client inhibition in the PI3K/Akt and MAPK pathways, (e) dysregulation of cytoskeletal and structural proteins, and (f) angiogenesis inhibition though HIF-1 [1, 137, 182, 183]. Withanolides with significant activity and identified mechanisms are depicted in Table 14.1, and their cytotoxic characteristics have been demonstrated in multiple cancer types, including breast, ovarian, colon, head and neck, renal, prostate, pancreatic, thyroid, glioblastoma, and hematologic cancers such as lymphoma, leukemia, and multiple myeloma [1, 70, 136, 182–186].
Anticancer Benefits of Withanolides in Animal Models
The groundwork done in vitro to establish the cytotoxic effect of withanolides against cancer cell lines and characterize their multiple mechanisms of therapeutic potential led to the translational evaluation of these compounds in several cancer animal models. Since cancer is a heterogeneous complex disease process, different cancers utilize different key oncogenic pathways for survival. Because different aspects of the inflammatory pathways are important in various cancer models, withanolides demonstrate cytotoxic effects on tumor growth, rate of metastatic spread, and even prevention. Looking at the STAT3 pathway in a colorectal cancer in vivo model, WS extract treatment attenuated IL-6 activation of STAT3 and demonstrated a 1.44 fold decrease in average tumor volume [53].
Following the initial characterization of a novel withanolide tubocapsenolide A to inhibit Hsp90–Hsp70 complex function in breast cancer through direct thiol oxidation [70], WA was shown to bind to the C-terminal of Hsp90, disrupt the Hsp90–Cdc37 complex, and inhibit chaperone activity in pancreatic cancer. Pancreatic cancer xenografts treated daily with withanolides demonstrated a 58 % reduction in average tumor volume compared to controls [63]. In medullary thyroid cancer, RET, a known Hsp90 client, is a key proto-oncogene that encodes for a transmembrane RTK. WA treatment of medullary thyroid xenografts not only inhibited RET phosphorylation and activation, but also inhibited phospho-Akt and phospho-ERK protein expression ex vivo. The xenograft tumors also demonstrated significant, 2-week delayed growth kinetics with 80 % of mice responding to WA treatment, as well as improved survival when compared to controls [187]. Withanolide E has also been shown to sensitize renal carcinoma cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated apoptosis through cFLIP degradation as a result of inhibited Hsp90 chaperone function. Combination treatment of mouse renal carcinoma xenografts with withanolide E and a TRAIL death receptor agonist demonstrated complete and sustained tumor responses in 55 % of treated mice compared to control or either treatment alone [188].
Cytoskeletal and structural protein inhibition by withanolides has been shown to lead to cell cycle arrest, decreased epithelial-to-mesenchymal transition (EMT), and decreased metastasis. In breast cancer, WA binds to and inhibits β-tubulin, decreasing β-tubulin protein levels in G0–G1 phase cells and severely disrupting normal spindle morphology, both of which lead to cell cycle arrest [14]. Withaferin A also induces perinuclear vimentin (a mesenchymal protein) accumulation leading to rapid vimentin depolymerization and disrupted morphology. In both murine mammary carcinoma and human xenograft breast cancer models in mice, WA and WS treatment showed a dose-dependent inhibition of tumor growth kinetics, a decrease in number of metastatic lung nodules, and an increase in ser56 phosphorylated vimentin that is indicative of disassembly [189, 190]. Subsequent studies also showed that WA treatment attenuated TNF-α and TGF-β induced EMT, increased the epithelial phenotype protein E-cadherin, and inhibited vimentin expression in addition to inhibited tumor grown kinetics and cell proliferation [23, 143]. Together, these studies characterize the ability of WA to inhibit EMT both in vitro and in vivo, which may inhibit the metastatic potential of these tumors.
Another area of research in cancer therapy is the inhibition of angiogenesis which limits the oxygen and nutrient supply to growing tumors as well as their metastatic capabilities. Withanolides demonstrate significant angiogenesis inhibition [55] and as previously described, mechanisms include blocking NF-κB activity along with inhibition of cyclin D1 expression through suppressing proteasome-mediated ubiquitin degradation of IκBα [52, 55], and the interplay between NF-κB, JAK/STAT, HIF-1 and downstream targets VEGF, hnRNP-K, and MMPs [52, 140, 159]. Using both a subcutaneous flank xenograft and lung metastasis murine model with fibrosarcoma cells, withanone and WA demonstrated significant suppression of both subcutaneous and lung metastasis tumor growth compared to controls. The ex vivo tumors then demonstrated decreased expression of hnRNP-K and downstream effectors VEGF, Erk p44/42, and MMP2 [159]. In an in vivo assay of neovascularization using subcutaneously injected Matrigel that contained VEGF and bFGF, mice treated with 3-azido withaferin A (3-azidoWA) demonstrated a marked reduction in neovascularization with both preventative and treatment dosing when compared to untreated controls [50].
Not only have withanolides demonstrated significant cytotoxicity in multiple cancer models, but they have also demonstrated the capability to prevent cancer growth and implantation [137]. The induction of the phase II enzyme quinone reductase (QR) in mouse hepatoma cells has been used as a biological screen for chemopreventive compounds, and multiple withanolides have demonstrated robust induction of QR with minimal toxicity [191]. Subsequently in rodent models, pretreatment with the WS root isolate 1-oxo-5beta, 6beta-epoxy-witha-2-enolide prevented the formation of cutaneous malignancy and were absent p53 + foci that were noted in untreated rats that had been exposed to ultraviolet B radiation and benzoyl peroxide [192]. Additionally, WS root extract inhibited benzo(a)pyrene-induced forestomach papillomagenesis by 60 % and 7,12-dimethylbenzanthracene-induced skin papillomagenesis by 45 % [193]. It has been proposed that one mechanism of cutaneous chemoprevention is through inhibition of carcinogen-induced upregulation of acetyl-CoA carboxylase by suppressing AP-1 activation [194]. In a mouse mammary tumor virus-neu (MMTV-neu) transgenic model that forms spontaneous tumors, preventative WA treatment led to a significant 50 % reduction in average mammary tumor weight, a 95 % reduction in average area of invasive carcinoma, and a 73 % reduction in incidence of lung metastasis [195]. Another important antitumor effect of these withanolides is that they can also target breast cancer stem cells. Treatment of breast cancer cells with WA resulted in decreased ability to form mammospheres as well as significantly decreased aldehyde dehydrogenase activity within the mammary tumor cells [196]. WS root extract has also been shown to reduce the formation of spontaneous estrogen receptor negative mammary tumors in the MMTV-neu mice when mice were fed a diet containing the extract [197].
Overall these in vivo studies also noted that withanolides were well tolerated without significant reductions in animal weight, necrosis, or fibrosis when compared to placebo [23, 143, 159, 189, 190, 192, 193, 195]. The promising results of withanolides observed thus far in cancer models have led to ongoing research by many centers to both identify new withanolides and evaluate existing withanolides in multiple cancer models. In the last few years, withanolides have also been shown to demonstrate tumor cytotoxicity using animal xenografts in gliomas [198], B-cell lymphomas [199], ovarian cancer [200, 201], prostate cancer [202, 203], soft tissue sarcoma [204], cervical carcinoma [205], melanoma, and mesothelioma [206]. These exciting results all point toward the incredible potential of these natural products for future clinical treatment regimens.
Clinical Applications of Withanolides for Cancer
Despite having improvements in our understanding of the multiple mechanisms of cytotoxicity with withanolide treatment across an expanding range of cancers, there has yet to be a significant human clinical placebo-controlled trial brought to completion and published that evaluates the efficacy of withanolides for the treatment of cancer. Withanolides are generally regarded as safe and have been used in human clinical trials for inflammatory and neurologic diseases, and have been evaluated for the treatment of fatigue for breast cancer patients undergoing chemotherapy [207]. Additional in vivo research on withanolides is ongoing and has overall been quite promising to identify withanolides as an important cancer therapy, however, further supporting research is needed to initiate the human clinical trials required to obtain a better understanding of the clinical treatment effects of withanolides on cancer. To date, there is no Food and Drug Administration (FDA)-approved good manufacturing process (GMP) facility currently purifying and producing purified withanolide compounds for use in clinical trials, although several facilities in other countries produce capsules of withanolide plant products and extracts that are not regulated by the FDA.
Application of Withanolides in Neurologic Diseases
Neurodegenerative Diseases
Neurodegenerative diseases are characterized by the progressive dysfunction and loss of neurons in the central nervous system (CNS) and are a major cause of dementia, cognitive and motor dysfunction. As the pathophysiology of neurodegenerative diseases such as Alzheimer’s disease (AD), Parkinson’s disease (PD), and Huntington’s disease (HD) are better understood, the major role of the immune system and neuroinflammation has become readily apparent [208]. Although the blood–brain barrier maintains some degree of separation of the CNS from the systemic immune system to aid in the immune privileged state of the CNS (its relative inability of nonself antigens to illicit an immune response), the role of the innate and adaptive immune responses has become a major focus of both study and intervention [209]. In addition to the formation and deposition of amyloid β plaques in AD, activation of microglia (macrophage-like CNS cell) and astrocytes, increased complement components, cytokines, and TLR pathways, and alterations in peripheral Th cell responses have also been described [210, 211]. In PD, mitochondrial dysfunction, oxidative stress, and altered protein handling with Lewy body deposition as well as inflammation through microglial activation, increased IL-1β, IL-6, TNF-α, and TLRs, formation of antibodies to neuronal antigens, and Th cell modulation are important. Finally in HD, mutant Huntingtin (HTT) expression in neurons and glia leading to microglia proliferation/activation, increased complement 3, 9 and neuroinflammation are keys to the pathogenesis of the disease [208, 209].
As previously discussed, withanolides inhibit multiple inflammatory pathways, and within the CNS astrocytes, Withaferin A has been shown to attenuate LPS-induced NF-κB, TNF-α, COX-2 and iNOS [212]. In addition to neuroinflammatory modulation, several withanolides including WS extract, withanosides (particularly withanoside IV and its active metabolite sominone), the synthetic withanolidedenosomin, withanolide A and coagulin Q have all demonstrated important effects on stimulating neurite outgrowth and regeneration [66, 210, 211]. In AD, additional key findings demonstrated that withanolide A significantly downregulates beta-site amyloid precursor protein (APP) cleaving enzyme 1 (BACE1; known as β-secretase, enzyme involved in production of Aβ) while it upregulates a disintegrin and metalloproteinase domain-containing protein 10 (ADAM10; α-secretase, non-amyloidogenic enzymatic processing of APP). Additionally, withanolide A increases expression of insulin-degrading enzyme, which is important in the proteolytic degradation of Aβ [213]. Multiple withanolides (WS, WA, and bracteosins) have also demonstrated significant acetylcholinesterase (AChE) or butyrylcholinesterase (BChE) inhibition [214, 215] with computational docking analysis supporting withanolide A high-affinity binding for active sites on AChE [216]. These results, combined with the known antioxidant and anti-inflammatory effects of withanolides have propelled their research forward in neurodegenerative disorders to further elucidate their mechanism of action.
In Vivo Activities of Withanolides in the Treatment of Neurodegenerative Diseases
In a systematic review and meta-analysis, Durg et al. identified and analyzed 28 studies evaluating the use of WS in neurobehavioral disorders (including AD, PD, HD and anxiety/stress) induced by brain oxidation in rodent models [217]. Overall, WS treatment had a protective effect on brain oxidative stress that corrected abnormal activity levels of super oxide dismutase (SOD), catalase, glutathione peroxidase and glutathione, lipid peroxidation, and levels of nitrite and AChE [217]. In several types of AD mouse models, including Aβ-[25–35] induced memory deficit and 5XFAD mice (5 FAD mutations carried on APP and PS1 transgenes), withanolides (withanolide A, withanoside IV and its main active metabolite, sominone, and WS extract) significantly ameliorated the impairment of spatial memory and behavioral deficits [39, 40, 210, 217], while ex vivo brains showed increased axonal and dendritic protein markers back to control levels [39, 211], increased axonal densities [40], and reduced Aβ levels and plaque depositions through upregulation of low-density lipoprotein receptor-related protein within the liver [218].
Parkinson’s disease is characterized by an age-related neurodegenerative progression that leads to resting tremor, rigidity, and akinesia though loss of dopaminergic nigro-striatal neurons, particularly within the substantia nigra [219]. The rodent models of Parkinsonism are created through brain injection of 6-Hydroxydopamine [220] or systemic administration of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) [221] or the combination of manganese ethylene-bis-dithiocarbamate (maneb) and N,N′-dimethyl-4,4′-bipyridinium dichloride (paraquat) [222]. They cause specific degeneration and toxicity to catecholaminergic and dopaminergic neurons though oxidative stress and formation of hydrogen peroxide, hydroxyl, and superoxide radicals [220–222]. Using these models, treatment with WS extract resulted in significant improvements in locomotor control (including rotation, locomotion, muscular coordination and rearing), amelioration of oxidative parameters (including LPO, glutathione and associated enzymes, SOD and catalase), correction of ex vivo rodent brain alterations to catecholamine, dopamine, and dopamine metabolites (DOPAC/HVA) in ex vivo rat brains [222–226]. Some of these effects may be due to mediation of the otherwise proapototic state through reductions of Bax and Bcl-2 expression and neuroinflammatory astrocyte activation [227].
Huntington’s disease has autosomal dominant transmission of the mutated HTT gene that leads to neuron destruction within the basal ganglia, leading to dementia and the characteristic involuntary writhing movements characteristic of Huntington’s chorea. Mutant HTT aggregates have been implicated in oxidative stress through the formation of free radicals, which creates imbalance and plays a role in subsequent neuroinflammation [228]. Huntington’s disease-like neurobehavioral and biochemical changes are induced in rodent models using neurotoxin 3-Nitropropionic acid. Treatment with WS root extract demonstrated significant dose-dependent attenuation of AChE levels, significantly improved oxidative stress markers, and improvement in general locomotor, performance and behavioral changes tested by Rotarod and Morris water [229, 230].
Although the involvement of the immune response and neuroinflammation plays a significant role in the pathogenesis of neurodegenerative diseases, the specific connections between these inflammatory pathways and the effects of withanolide treatment have not been well defined. The role of WS on reducing the oxidative stress within neurons has been demonstrated in multiple animal models of different disease processes, as well as the neuro-regenerative role of several withanolides on axonal and dendritic outgrowths and functional locomotor improvements. Connecting these effects to neuroinflammation and immunomodulation identifies an important area of future research.
Clinical Activity of Withanolides in Neurodegenerative Diseases
There is an overall paucity of research on the effects of withanolides in patients with neurodegenerative diseases. WS was a component of one study that evaluated the efficacy of a traditional Ayurveda treatment in clinically diagnosed PD patients. Treatment with a combination of eliminative cleansing and a concoction of cow’s milk with powdered Mucunapruriens, Hyoscyamusreticulatus seeds, WS and Sidacordifolia roots (analyzed to contain 200 mg L-DOPA per dose) showed significant improvements in tremor/bradykinesia, stiffness and cramp-like pain resulting in improved activities of daily living, however, no changes in other symptoms like dysphonia, dysarthria, wasting, cogwheel rigidity, shuffling gait, or other locomotor symptoms [231]. However, conclusions regarding the role of WS cannot be reached due to the mixed nature of the L-DOPA containing herbal treatment. However, the growing in vitro and animal data continue to support the use of withanolides in neurodegenerative diseases, and illustrates the potential for its use in a more controlled clinical trial.
Withanolides in Neurobehavioral/Psychiatric Diseases
There is a broad range of neurological diseases in which inflammation and particularly cytokines and microglial activation have been shown to play a pathophysiological role. These include chronic stress [232], anxiety [233], depression [234], schizophrenia [235, 236], bipolar disorder [237, 238], and obsessive–compulsive disorder (OCD) [239]. Although the study of how the immune system and inflammation contribute to these diseases is relatively recent, major inflammatory pathways and cytokines have been identified. One proposed mechanism is through the stress response, whether through external stimuli or psychological imbalances, which leads to activation of the hypothalamic-pituitary-adrenal (HPA) axis and short-term elevations in glucocorticoids (mainly cortisol) and a subsequent anti-inflammatory response. However, prolonged stress and sustained HPA activation causes cortisol resistance and initiation of pro-inflammatory pathways, such Th cell release of IL-1α and β, IL-2, IL-6, IL-10, TNF-α cytokines, activation of NF-κB pathway, and microglial activation [232–238]. In addition to the known anti-inflammatory effects of withanolides previously discussed, they have also been described as adaptogens that assist with balance and regulation of the body’s physiologic response to stressors [240]. The adaptogen mechanism of effect has been proposed to occur through modulation of the HPA axis, inducing stress-activated c-Jun N-terminal protein kinase (JNK1), inhibiting iNOS expression, and modulation of Hsp70 chaperone function [241]. Additionally, the inhibitory neurotransmitter gamma-aminobutyric acid (GABA), which plays an important inhibitory role in neurological disorders, has more recently been shown to have key anti-inflammatory interactions within the immune system through suppression of T-cells and macrophages, and inhibition of NF-κB on lymphocytes [242]. The GABA-mimetic activity of WS root extract has been shown for several decades [243] with a recent study identifying a 27 times greater affinity of WS for the highly sensitive GABAρ1 receptors compared to GABAA receptors, though WA and withanolide A were not the active GABA-mimetics [244]. These GABA-mimetic and adaptogenic effects likely play a significant role in the long-standing history of using Ashwagandha for the treatment of anxiety and neurobehavioral disorders, though the vast majority of animal and human studies have been performed in the last several decades.
In Vivo Activity of Withanolides for Neurobehavioral/Psychiatric Diseases
Most animal studies evaluating neurologic disorders have focused on the effects of WS extract using models of either chronic stress or withdrawal to induce anxiety and depression. Using a chronic, unpredictable, mild footshock to create a chronic stress model in rats, treatment with aqueous ethanol WS root extract significantly attenuated chronic, stress-induced abnormalities with improvements in biochemical imbalances (elevated blood glucose and corticosteroid levels), reduced number and severity of gastric ulcers, improved behavioral depression and sexual responses, improved cognitive memory function, and rescued immunosuppression in macrophage activity and immunologic pedal edema [245]. In depression and anxiety rodent models using chronic stress, isolation, sleep deprivation, WS extract treatment demonstrated significant antidepressant effects, and potentiated conventional antidepressants (tricyclic antidepressant imipramine and selective serotonin-reuptake inhibitor fluoxetine) measured by reduced immobility time in the forced open swim test (“behavioral despair/learned helplessness”), and an anxiolytic effect similar to benzodiazepines (lorazepam and diazepam) in the elevated plus maze, the social interaction test, and the feeding latency test [246–250].
In an OCD rodent model of marble-burying behavior, intraperitoneal injections of both methanolic and aqueous WS root extracts 30 min prior to evaluation resulted in significant dose-dependent reductions in marble-burying behavior similar to standard fluoxetine alone, and a synergistic effect in combination with fluoxetine [251]. These in vivo studies demonstrate a consistent amelioration of stress, anxiety, depressant, and OCD behaviors with corresponding corrections in biochemical abnormalities, however, the modulation of inflammatory pathways in these diseases requires further characterization.
Clinical Activities of Withanolides in Neurobehavioral/Psychiatric Diseases
Recent clinical trials using WS extract as a neurological treatment have evaluated effects on psychomotor function in patients with anxiety, bipolar disorder, and schizophrenia and provide clinical support for centuries of WS use in Ayurvedic medicine. There have been several randomized controlled trials that demonstrate significant improvement in anxiety and stress relief [252–257]. Studying ICD-10 anxiety disorders (generalized anxiety disorder, mixed anxiety and depression, panic disorder, and adjustment disorder with anxiety) assessed by Hamilton Anxiety Scale, patients were treated with 500 mg WS extract or placebo twice daily with subsequent clinically guided dose titrations. At 6 weeks, the WS extract treatment response of 88 % was significantly improved compared to the 50 % placebo response with no significant difference in overall adverse outcomes or with abrupt cessation of WS treatment [253]. Subsequent randomized controlled studies have corroborated their results and demonstrated either significantly decreased anxiety or improved stress relief using multiple anxiety scales (Hamilton Anxiety Scale, Beck Anxiety Inventory, Perceived Stress Scale), a range of WS dosing primarily between 125 and 600 mg/day for extract (one study 12,000 mg/day whole dried root powder) over the course of 60–84 days [252, 254–257]. None of the studies reported significant adverse events with WS treatment, with all described as mild or comparable to the placebo groups. However, significant limitations existed in each of the studies that included high rates of patient withdrawal from study [253], low sample size leading to bias risk and under powering, inconsistent dosing or lack of withanolide standardization, lack of comparison to standard of care anti-anxiolytics, and possible methodological flaws [252–257]. The limitations impair making definitive conclusions regarding the effectiveness of WS in anxiety disorders, but the body of study identifies WS as a relatively safe therapeutic and supports the role for conducting additional clinical studies.
In both bipolar disease and schizophrenia, cognitive impairments are consistently associated with poorer functional outcomes. As discussed previously, WS improves memory and cognitive function when treating neurodegenerative rodent models, and in healthy adults significantly improves psychomotor function as demonstrated by simple reaction times, choice discrimination, digit symbol substitution, digit vigilance, and card sorting tests compared to placebo [258]. In well maintained bipolar disease patients, a randomized controlled, trial of WS in adjunct to maintenance bipolar treatment resulted specific improvement in several cognitive function tests including the digit span backward, Flanker neutral response time, and social cognition response rating from the Penn Emotional Acuity Test, though no global cognitive improvement [259]. In schizophrenia, there is currently an ongoing randomized controlled clinical trial in the United States evaluating the effect of WS extract on symptom severity and associated stress that also aims to characterize alterations in inflammatory cytokines [260]. The results from this study will help provide valuable insight into the effect of WS on neuroinflammation and schizophrenia symptoms.
Conclusions
Withanolides are an incredibly bio-diverse group of naturally occurring steroidal lactones. These plant-derived compounds have ongoing use within Ayurvedic medical practices and are important plant-derived medicinal compounds for a variety of human diseases and conditions. With WA as the archetype of this novel class drugs, there are now approximately 900 withanolides identified from a natural source or synthetically modified. Significant strides have been made in recent years to advance our understanding of the biochemical, immunomodulatory, and anti-inflammatory mechanisms of these compounds and plant extracts. Added to this is emerging in vivo and clinical evidence of safe, efficacious treatment effects across multiple disease processes ranging from autoimmune/inflammatory disorders, cancer, neurodegenerative diseases, and neurobehavioral/psychiatric diseases. Ongoing research on withanolides continues to identify new compounds and analogs, both plant derived and synthetically altered, that are more potent or specific for use in particular diseases. Given their impressive biologic activity in a number of challenging and complex disease processes and their unique ability to synergize with many standard drug treatments, these naturally derived drug compounds undoubtedly will have an important role clinically in well-designed combination strategies once disease-specific mechanisms of action and synergy are further validated and characterized. As such, they remain a very hot area of research as a novel group of medicinal therapeutics.
Abbreviations
- 5XFAD
5 FAD mutations carried on APP and PS1 transgenes
- AChE
Acetylcholinesterase
- AD
Alzheimer’s disease
- ADAM10
Adisintegrin and metalloproteinase domain-containing protein 10, or α-secretase
- AP-1
Activator protein 1
- APP
Amyloid precursor protein
- BACE
APP cleaving enzyme 1 or β-secretase
- BChE
Butyrylcholinesterase
- Bfl-1/A1
Bcl-2-related protein A1
- C/EBPα
CCAAT/enhancer-binding proteinα
- CCR7
Chemokine (C–C motif) receptor 7
- cFLIP
C-FADD-like IL-1β-converting enzyme–inhibitory proteins
- CNS
Central nervous system
- COX
Cyclooxygenase
- CSC
Cancer stem cell
- EGF
Epidermal growth factor
- EGFR
EGF receptor
- ERK
Extracellular signal-regulated kinase
- FDA
Food and Drug Administration
- GABA
Gamma-aminobutyric acid
- GMP
Good manufacturing process
- HD
Huntington’s disease
- HIF-1
Hypoxia inducible factor-1
- HMGB1
High mobility group box 1
- hnRNP-K
Heterogeneous nuclear ribonucleoprotein
- KHPA
Hypothalamic–pituitary–adrenal
- Hsp
Heat shock protein
- HTT
Mutant Huntingtin
- IAP1
Inhibitor of apoptosis protein-1
- IBD
Inflammatory bowel disease
- ICAM
Intercellular adhesion molecule
- IFN
Interferon
- IKK
I kappa B kinase
- IL
Interleukin
- iNOS
Inducible nitric oxide synthase
- JAK
Janus kinase
- JNK1
c-Jun N-terminal protein kinase
- Keap1
Kelch like ECH-associated protein-1
- LPS
Lipopolysaccharide
- LTB4
Leukotriene B4
- MAPK
Mitogen-activated protein kinase
- MCE
Mitotic clonal expansion,
- MCP-1
Monocyte chemoattractant protein-1
- MMPs
Matrix metalloproteinases
- MMTV-neu
Mouse mammary tumor virus-neu
- MPTP
1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine
- MS
Mass spectrometry
- mTOR
Mechanistic target of rapamycin
- MUC-1
Mucin 1
- NF-κB
Nuclear factor kappa B
- NO
Nitric oxide
- Nrf2
Nuclear factor erythroid 2-related factor 2
- OCD
Obsessive-compulsive disorder
- PBMC
Peripheral blood mononuclear cells
- PD
Parkinson’s disease
- PDGF
Platelet-derived growth factor
- PGE2
Prostaglandin E2
- PI3K
Phosphatidylinositol-3-kinase
- PPARs
Peroxisome proliferator-activated receptors
- QR
Quinone reductase
- RA
Rheumatoid arthritis
- ROS
Reactive oxygen species
- RTKs
Receptor tyrosine kinases
- SAR
Structural–activity relationship
- SFMC
Synovial fluid mononuclear cells
- SOD
Super oxide dismutase
- STAT
Signal transducer and activator of transcription
- TAK1
Transforming growth factor-β-activating kinase
- TGF-β
Transforming growth factor beta
- Th
T-helper
- TLRs
Toll-like receptors
- TNBS
Trinitrobenzyl sulfonic acid
- TNF
Tumor necrosis factor
- TRAIL
Tumor necrosis factor-related apoptosis-inducing ligand
- TWIST
Twist family BHLH transcription factor
- UPLC
Ultra-performance liquid chromatography
- VEGF
Vascular endothelial growth factor
- WA
Withaferin A
- WS
Withania somnifera
Footnotes
These authors contributed equally
Contributor Information
Subash Chandra Gupta, Email: subhashchandra@bhu.ac.in.
Sahdeo Prasad, Email: sprasad@mdanderson.org.
Bharat B. Aggarwal, Email: bbaggarwal@gmail.com
Mark S. Cohen, Phone: (734) 615-4741, Email: cohenmar@med.umich.edu
References
- 1.Chen L-X, He H, Qiu F. Natural withanolides: an overview. Nat Prod Rep. 2011;28(4):705–740. doi: 10.1039/c0np00045k. [DOI] [PubMed] [Google Scholar]
- 2.Misico Rosana I., Nicotra Viviana E., Oberti Juan C., Barboza Gloria, Gil Roberto R., Burton Gerardo. Progress in the Chemistry of Organic Natural Products Vol. 94. Vienna: Springer Vienna; 2011. Withanolides and Related Steroids; pp. 127–229. [DOI] [PubMed] [Google Scholar]
- 3.Zhang H, Cao C-M, Gallagher RJ, Timmermann BN. Antiproliferative withanolides from several solanaceous species. Nat Prod Res. 2014;28(22):1941–1951. doi: 10.1080/14786419.2014.919286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Glotter E. Withanolides and related ergostane-type steroids. Nat Prod Rep. 1991;8(4):415–440. doi: 10.1039/np9910800415. [DOI] [PubMed] [Google Scholar]
- 5.Misico RI, Song LL, Veleiro AS, Cirigliano AM, Tettamanzi MC, Burton G, et al. Induction of quinone reductase by withanolides. J Nat Prod. 2002;65(5):677–680. doi: 10.1021/np0106337. [DOI] [PubMed] [Google Scholar]
- 6.Lavie D, Glotter E, Shvo Y (1965) Constituents of Withania somnifera Dun. Part IV. The structure of withaferin A. J Chem Soc (1): 7517–7531
- 7.Zhang H, Samadi AK, Cohen MS, Timmermann BN. Antiproliferative withanolides from the Solanaceae: a structure–activity study. Pure Appl Chem. 2012;84(6):1353–1367. doi: 10.1351/PAC-CON-11-10-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cao C-M, Wu X, Kindscher K, Xu L, Timmermann BN. Withanolides and Sucrose Esters from Physalis neomexicana. J Nat Prod. 2015;78(10):2488–2493. doi: 10.1021/acs.jnatprod.5b00698. [DOI] [PubMed] [Google Scholar]
- 9.Chen B-W, Chen Y-Y, Lin Y-C, Huang C-Y, Uvarani C, Hwang T-L, et al. Capsisteroids A–F, withanolides from the leaves of Solanum capsicoides. RSC Adv. 2015;5(108):88841–88847. doi: 10.1039/C5RA12014D. [DOI] [Google Scholar]
- 10.Chao C-H, Chou K-J, Wen Z-H, Wang G-H, Wu Y-C, Dai C-F, et al. Paraminabeolides A-F, cytotoxic and anti-inflammatory marine withanolides from the soft coral Paraminabea acronocephala. J Nat Prod. 2011;74(5):1132–1141. doi: 10.1021/np2000705. [DOI] [PubMed] [Google Scholar]
- 11.Ksebati MB, Schmitz FJ. Minabeolides: a group of withanolides from a soft coral, Minabea sp. J Organ Chem. 1988;53(17):3926–3929. doi: 10.1021/jo00252a008. [DOI] [Google Scholar]
- 12.Huang C-Y, Liaw C-C, Chen B-W, Chen P-C, Su J-H, Sung P-J, et al. Withanolide-based steroids from the cultured soft coral Sinularia brassica. J Nat Prod. 2013;76(10):1902–1908. doi: 10.1021/np400454q. [DOI] [PubMed] [Google Scholar]
- 13.Jayaprakasam B, Zhang Y, Seeram NP, Nair MG. Growth inhibition of human tumor cell lines by withanolides from Withania somnifera leaves. Life Sci. 2003;74(1):125–132. doi: 10.1016/j.lfs.2003.07.007. [DOI] [PubMed] [Google Scholar]
- 14.Antony ML, Lee J, Hahm E-R, Kim S-H, Marcus AI, Kumari V, et al. Growth arrest by the antitumor steroidal lactone withaferin A in human breast cancer cells is associated with down-regulation and covalent binding at cysteine 303 of β-tubulin. J Biol Chem. 2014;289(3):1852–1865. doi: 10.1074/jbc.M113.496844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shohat B, Joshua H. Effect of withaferin a on Ehrlich ascites tumor cells. II. Target tumor cell destruction in vivo by immune activation. Int J Cancer. 1971;8(3):487–496. doi: 10.1002/ijc.2910080317. [DOI] [PubMed] [Google Scholar]
- 16.Zhang H, Bazzill J, Gallagher RJ, Subramanian C, Grogan PT, Day VW, et al. Antiproliferative Withanolides from Datura wrightii. J Nat Prod. 2012;76(3):445–449. doi: 10.1021/np300766p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhang H, Samadi AK, Gallagher RJ, Araya JJ, Tong X, Day VW, et al. Cytotoxic withanolide constituents of Physalis longifolia. J Nat Prod. 2011;74(12):2532–2544. doi: 10.1021/np200635r. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Minguzzi S, Barata LES, Shin YG, Jonas PF, Chai H-B, Park EJ, et al. Cytotoxic withanolides from Acnistus arborescens. Phytochemistry. 2002;59(6):635–641. doi: 10.1016/S0031-9422(02)00022-5. [DOI] [PubMed] [Google Scholar]
- 19.He Q-P, Ma L, Luo J-Y, He F-Y, Lou L-G, Hu L-H. Cytotoxic withanolides from Physalis angulata L. Chem Biodivers. 2007;4(3):443–449. doi: 10.1002/cbdv.200790036. [DOI] [PubMed] [Google Scholar]
- 20.Grogan PT, Sleder KD, Samadi AK, Zhang H, Timmermann BN, Cohen MS. Cytotoxicity of withaferin A in glioblastomas involves induction of an oxidative stress-mediated heat shock response while altering Akt/mTOR and MAPK signaling pathways. Invest New Drugs. 2013;31(3):545–557. doi: 10.1007/s10637-012-9888-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Samadi AK, Tong X, Mukerji R, Zhang H, Timmermann BN, Cohen MS. Withaferin A, a cytotoxic steroid from Vassobia breviflora, induces apoptosis in human head and neck squamous cell carcinoma. J Nat Prod. 2010;73(9):1476–1481. doi: 10.1021/np100112p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Choi JK, Murillo G, Su B-N, Pezzuto JM, Kinghorn AD, Mehta RG. Ixocarpalactone A isolated from the Mexican tomatillo shows potent antiproliferative and apoptotic activity in colon cancer cells. FEBS J. 2006;273(24):5714–5723. doi: 10.1111/j.1742-4658.2006.05560.x. [DOI] [PubMed] [Google Scholar]
- 23.Stan SD, Hahm E-R, Warin R, Singh SV. Withaferin A causes FOXO3a- and Bim-dependent apoptosis and inhibits growth of human breast cancer cells in vivo. Cancer Res. 2008;68(18):7661–7669. doi: 10.1158/0008-5472.CAN-08-1510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kaileh M, Vanden Berghe W, Heyerick A, Horion J, Piette J, Libert C, et al. Withaferin A strongly elicits IκB Kinase β hyperphosphorylation concomitant with potent inhibition of its kinase activity. J Biol Chem. 2007;282(7):4253–4264. doi: 10.1074/jbc.M606728200. [DOI] [PubMed] [Google Scholar]
- 25.Oh JH, Lee T-J, Park J-W, Kwon TK. Withaferin A inhibits iNOS expression and nitric oxide production by Akt inactivation and down-regulating LPS-induced activity of NF-κB in RAW 264.7 cells. Eur J Pharmacol. 2008;599(1–3):11–17. doi: 10.1016/j.ejphar.2008.09.017. [DOI] [PubMed] [Google Scholar]
- 26.Wube AA, Wenzig E-M, Gibbons S, Asres K, Bauer R, Bucar F. Constituents of the stem bark of Discopodium penninervium and their LTB4 and COX-1 and -2 inhibitory activities. Phytochemistry. 2008;69(4):982–987. doi: 10.1016/j.phytochem.2007.11.001. [DOI] [PubMed] [Google Scholar]
- 27.Yang B-Y, Guo R, Li T, Wu J-J, Zhang J, Liu Y, et al. New anti-inflammatory withanolides from the leaves of Datura metel L. Steroids. 2014;87:26–34. doi: 10.1016/j.steroids.2014.05.003. [DOI] [PubMed] [Google Scholar]
- 28.Maldonado E, Amador S, Martínez M, Pérez-Castorena AL. Virginols A–C, three new withanolides from Physalis virginiana. Steroids. 2010;75(4–5):346–349. doi: 10.1016/j.steroids.2010.01.014. [DOI] [PubMed] [Google Scholar]
- 29.Quang TH, Ngan NTT, Minh CV, Kiem PV, Yen PH, Tai BH, et al. Plantagiolides I and J, two new withanolide glucosides from Tacca plantaginea with nuclear factor-kappaB inhibitory and peroxisome proliferator-activated receptor transactivational activities. Chem Pharm Bull. 2012;60(12):1494–1501. doi: 10.1248/cpb.c12-00550. [DOI] [PubMed] [Google Scholar]
- 30.Jayaprakasam B, Nair MG. Cyclooxygenase-2 enzyme inhibitory withanolides from Withania somnifera leaves. Tetrahedron. 2003;59(6):841–849. doi: 10.1016/S0040-4020(02)01601-0. [DOI] [Google Scholar]
- 31.Qiu L, Zhao F, Jiang Z-H, Chen L-X, Zhao Q, Liu H-X, et al. Steroids and flavonoids from Physalis alkekengi var. franchetii and their inhibitory effects on nitric oxide production. J Nat Prod. 2008;71(4):642–646. doi: 10.1021/np700713r. [DOI] [PubMed] [Google Scholar]
- 32.Malik F, Singh J, Khajuria A, Suri KA, Satti NK, Singh S, et al. A standardized root extract of Withania somnifera and its major constituent withanolide-A elicit humoral and cell-mediated immune responses by up regulation of Th1-dominant polarization in BALB/c mice. Life Sci. 2007;80(16):1525–1538. doi: 10.1016/j.lfs.2007.01.029. [DOI] [PubMed] [Google Scholar]
- 33.Habtemariam S. Cytotoxicity and immunosuppressive activity of withanolides from Discopodium penninervium. Planta Med. 1997;63(01):15–17. doi: 10.1055/s-2006-957594. [DOI] [PubMed] [Google Scholar]
- 34.Kour K, Pandey A, Suri KA, Satti NK, Gupta KK, Bani S. Restoration of stress-induced altered T cell function and corresponding cytokines patterns by Withanolide A. Int Immunopharmacol. 2009;9(10):1137–1144. doi: 10.1016/j.intimp.2009.05.011. [DOI] [PubMed] [Google Scholar]
- 35.Abou-Douh AM. New withanolides and other constituents from the fruit of Withania somnifera. Arch Pharm. 2002;335(6):267–276. doi: 10.1002/1521-4184(200208)335:6<267::AID-ARDP267>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- 36.Bravo BJA, Sauvain M, Gimenez TA, Balanza E, Serani L, Laprévote O, et al. Trypanocidal withanolides and withanolide glycosides from Dunalia brachyacantha. J Nat Prod. 2001;64(6):720–725. doi: 10.1021/np000527p. [DOI] [PubMed] [Google Scholar]
- 37.Choudhary MI, Yousuf S, Samreen AS, Atta UR. New leishmanicidal physalins from Physalis minima. Nat Prod Res. 2007;21(10):877–883. doi: 10.1080/14786410701315147. [DOI] [PubMed] [Google Scholar]
- 38.Bhattacharya SK, Satyan KS, Ghosal S. Antioxidant activity of glycowithanolides from Withania somnifera. Indian J Exp Biol. 1997;35(3):236–239. [PubMed] [Google Scholar]
- 39.Kuboyama T, Tohda C, Komatsu K. Withanoside IV and its active metabolite, sominone, attenuate Aβ(25–35)-induced neurodegeneration. Eur J Neurosci. 2006;23(6):1417–1426. doi: 10.1111/j.1460-9568.2006.04664.x. [DOI] [PubMed] [Google Scholar]
- 40.Joyashiki E, Matsuya Y, Tohda C. Sominone Improves Memory Impairments and Increases axonal density in Alzheimer’s Disease Model Mice, 5XFAD. Int J Neurosci. 2011;121(4):181–190. doi: 10.3109/00207454.2010.541571. [DOI] [PubMed] [Google Scholar]
- 41.Baitharu I, Jain V, Deep SN, Shroff S, Sahu JK, Naik PK, et al. Withanolide A prevents neurodegeneration by modulating hippocampal glutathione biosynthesis during hypoxia. PLoS ONE. 2014;9(10):e105311. doi: 10.1371/journal.pone.0105311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Devi PU. Withaferin A: a new radiosensitizer from the Indian medicinal plant Withania somnifera. Int J Radiat Biol. 1996;69(2):193–197. doi: 10.1080/095530096146020. [DOI] [PubMed] [Google Scholar]
- 43.Devi PU, Kamath R. Radiosensitizing effect of withaferin A combined with hyperthermia on mouse fibrosarcoma and melanoma. J Radiat Res. 2003;44(1):1–6. doi: 10.1269/jrr.44.1. [DOI] [PubMed] [Google Scholar]
- 44.Budhiraja RD, Krishan P, Sudhir S. Biological activity of withanolides. J Sci Ind Res. 2000;59(11):904–911. [Google Scholar]
- 45.Mareggiani G, Picollo MI, Zerba E, Burton G, Tettamanzi MC, Benedetti-Doctorovich MOV, et al. Antifeedant Activity of Withanolides from Salpichroa origanifolia on Musca domestica. J Nat Prod. 2000;63(8):1113–1116. doi: 10.1021/np0001068. [DOI] [PubMed] [Google Scholar]
- 46.Mandal C, Dutta A, Mallick A, Chandra S, Misra L, Sangwan R, et al. Withaferin A induces apoptosis by activating p38 mitogen-activated protein kinase signaling cascade in leukemic cells of lymphoid and myeloid origin through mitochondrial death cascade. Apoptosis. 2008;13(12):1450–1464. doi: 10.1007/s10495-008-0271-0. [DOI] [PubMed] [Google Scholar]
- 47.Khan ZA, Ghosh AR. Possible nitric oxide modulation in protective effects of withaferin A against stress induced neurobehavioural changes. J Med Plants Res. 2010;4(6):490–495. [Google Scholar]
- 48.Machin RP, Veleiro AS, Nicotra VE, Oberti JC, Padrón JM. Antiproliferative activity of withanolides against human breast cancer cell lines. J Nat Prod. 2010;73(5):966–968. doi: 10.1021/np9006734. [DOI] [PubMed] [Google Scholar]
- 49.Koduru S, Kumar R, Srinivasan S, Evers MB, Damodaran C. Notch-1 inhibition by Withaferin-A: a therapeutic target against colon carcinogenesis. Mol Cancer Ther. 2010;9(1):202–210. doi: 10.1158/1535-7163.MCT-09-0771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Rah B, Amin H, Yousuf K, Khan S, Jamwal G, Mukherjee D, et al. A novel MMP-2 inhibitor 3-azidowithaferin A (3-azidoWA) abrogates cancer cell invasion and angiogenesis by modulating extracellular par-4. PLoS ONE. 2012;7(9):e44039. doi: 10.1371/journal.pone.0044039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lee W, Kim TH, Ku S-K, K-j Min, Lee H-S, Kwon TK, et al. Barrier protective effects of withaferin A in HMGB1-induced inflammatory responses in both cellular and animal models. Toxicol Appl Pharmacol. 2012;262(1):91–98. doi: 10.1016/j.taap.2012.04.025. [DOI] [PubMed] [Google Scholar]
- 52.Bargagna-Mohan P, Ravindranath PP, Mohan R. Small molecule anti-angiogenic probes of the ubiquitin proteasome pathway: Potential application to choroidal neovascularization. Invest Ophthalmol Vis Sci. 2006;47(9):4138–4145. doi: 10.1167/iovs.05-1452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Choi BY, Kim B-W. Withaferin-A inhibits colon cancer cell growth by blocking STAT3 transcriptional activity. J Cancer Prevent. 2015;20(3):185–192. doi: 10.15430/JCP.2015.20.3.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ichikawa H, Takada Y, Shishodia S, Jayaprakasam B, Nair MG, Aggarwal BB. Withanolides potentiate apoptosis, inhibit invasion, and abolish osteoclastogenesis through suppression of nuclear factor-κB (NF-κB) activation and NF-κB-regulated gene expression. Mol Cancer Ther. 2006;5(6):1434–1445. doi: 10.1158/1535-7163.MCT-06-0096. [DOI] [PubMed] [Google Scholar]
- 55.Mohan R, Hammers H, Bargagna-Mohan P, Zhan X, Herbstritt C, Ruiz A, et al. Withaferin A is a potent inhibitor of angiogenesis. Angiogenesis. 2004;7(2):115–122. doi: 10.1007/s10456-004-1026-3. [DOI] [PubMed] [Google Scholar]
- 56.Ndlovu MN, Van Lint C, Van Wesemael K, Callebert P, Chalbos D, Haegeman G, et al. Hyperactivated NF-κB and AP-1 transcription factors promote highly accessible chromatin and constitutive transcription across the interleukin-6 gene promoter in metastatic breast cancer cells. Mol Cell Biol. 2009;29(20):5488–5504. doi: 10.1128/MCB.01657-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Yco LP, Mocz G, Opoku-Ansah J, Bachmann AS. Withaferin A Inhibits STAT3 and induces tumor cell death in neuroblastoma and multiple myeloma. Biochem Insights. 2014;7:1–13. doi: 10.4137/BCI.S18863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ishiguro M, Kajikawa A, Haruyama T, Morisaki M, Ikekawa N. Synthetic studies of withanolide I synthesis of AB ring moiety of withaferin A. Tetrahedron Lett. 1974;15(15):1421–1424. doi: 10.1016/S0040-4039(01)82508-9. [DOI] [Google Scholar]
- 59.Ozawa M, Morita M, Hirai G, Tamura S, Kawai M, Tsuchiya A, et al. Contribution of cage-shaped structure of physalins to their mode of action in inhibition of NF-κB activation. ACS Med Chem Lett. 2013;4(8):730–735. doi: 10.1021/ml400144e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Wijeratne EMK, Xu Y-M, Scherz-Shouval R, Marron MT, Rocha DD, Liu MX, et al. Structure–activity relationships for withanolides as inducers of the cellular heat-shock response. J Med Chem. 2014;57(7):2851–2863. doi: 10.1021/jm401279n. [DOI] [PubMed] [Google Scholar]
- 61.Damu AG, Kuo P-C, Su C-R, Kuo T-H, Chen T-H, Bastow KF, et al. Isolation, structures, and structure-cytotoxic activity relationships of withanolides and physalins from Physalis angulata. J Nat Prod. 2007;70(7):1146–1152. doi: 10.1021/np0701374. [DOI] [PubMed] [Google Scholar]
- 62.Wang H-C, Tsai Y-L, Wu Y-C, Chang F-R, Liu M-H, Chen W-Y, et al. Withanolides-induced breast cancer cell death is correlated with their ability to inhibit heat protein 90. PLoS ONE. 2012;7(5):e37764. doi: 10.1371/journal.pone.0037764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Yu Y, Hamza A, Zhang T, Gu M, Zou P, Newman B, et al. Withaferin A targets heat shock protein 90 in pancreatic cancer cells. Biochem Pharmacol. 2010;79(4):542–551. doi: 10.1016/j.bcp.2009.09.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ji L, Yuan Y, Luo L, Chen Z, Ma X, Ma Z, et al. Physalins with anti-inflammatory activity are present in Physalis alkekengi var. franchetii and can function as Michael reaction acceptors. Steroids. 2012;77(5):441–447. doi: 10.1016/j.steroids.2011.11.016. [DOI] [PubMed] [Google Scholar]
- 65.Beg M, Chauhan P, Varshney S, Shankar K, Rajan S, Saini D, et al. A withanolide coagulin-L inhibits adipogenesis modulating Wnt/β-catenin pathway and cell cycle in mitotic clonal expansion. Phytomedicine. 2014;21(4):406–414. doi: 10.1016/j.phymed.2013.10.009. [DOI] [PubMed] [Google Scholar]
- 66.Teshigawara K, Kuboyama T, Shigyo M, Nagata A, Sugimoto K, Matsuya Y, et al. A novel compound, denosomin, ameliorates spinal cord injury via axonal growth associated with astrocyte-secreted vimentin. Br J Pharmacol. 2013;168(4):903–919. doi: 10.1111/j.1476-5381.2012.02211.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Takimoto T, Kanbayashi Y, Toyoda T, Adachi Y, Furuta C, Suzuki K, et al. 4β-Hydroxywithanolide E isolated from Physalis pruinosa calyx decreases inflammatory responses by inhibiting the NF-κB signaling in diabetic mouse adipose tissue. Int J Obes. 2014;38(11):1432–1439. doi: 10.1038/ijo.2014.33. [DOI] [PubMed] [Google Scholar]
- 68.Budhiraja RD, Sudhir S, Garg KN. Antiinflammatory activity of 3 β-hydroxy-2,3-dihydro-withanolide F. Planta Med. 1984;50(02):134–136. doi: 10.1055/s-2007-969651. [DOI] [PubMed] [Google Scholar]
- 69.Pan M-R, Chang H-C, Wu Y-C, Huang C-C, Hung W-C. Tubocapsanolide A inhibits transforming growth factor-β-activating kinase 1 to suppress NF-κB-induced CCR7. J Biol Chem. 2009;284(5):2746–2754. doi: 10.1074/jbc.M806223200. [DOI] [PubMed] [Google Scholar]
- 70.Chen W-Y, Chang F-R, Huang Z-Y, Chen J-H, Wu Y-C, Wu C-C. Tubocapsenolide A, a novel withanolide, inhibits proliferation and induces apoptosis in MDA-MB-231 cells by thiol oxidation of heat shock proteins. J Biol Chem. 2008;283(25):17184–17193. doi: 10.1074/jbc.M709447200. [DOI] [PubMed] [Google Scholar]
- 71.Zhang X, Blaskovich MA, Forinash KD, Sebti SM. Withacnistin inhibits recruitment of STAT3 and STAT5 to growth factor and cytokine receptors and induces regression of breast tumours. Br J Cancer. 2014;111(5):894–902. doi: 10.1038/bjc.2014.349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Suttana W, Mankhetkorn S, Poompimon W, Palagani A, Zhokhov S, Gerlo S, et al. Differential chemosensitization of P-glycoprotein overexpressing K562/Adr cells by withaferin A and Siamois polyphenols. Mol Cancer. 2010;9(1):1–22. doi: 10.1186/1476-4598-9-99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Min K-J, Choi K, Kwon TK. Withaferin A down-regulates lipopolysaccharide-induced cyclooxygenase-2 expression and PGE2 production through the inhibition of STAT1/3 activation in microglial cells. Int Immunopharmacol. 2011;11(8):1137–1142. doi: 10.1016/j.intimp.2011.02.029. [DOI] [PubMed] [Google Scholar]
- 74.Mulabagal V, Subbaraju GV, Rao CV, Sivaramakrishna C, DeWitt DL, Holmes D, et al. Withanolide sulfoxide from Aswagandha roots inhibits nuclear transcription factor-kappa-B, cyclooxygenase and tumor cell proliferation. Phytother Res. 2009;23(7):987–992. doi: 10.1002/ptr.2736. [DOI] [PubMed] [Google Scholar]
- 75.Ahmed Lamiaa Ali. Renoprotective Effect of Egyptian Cape Gooseberry Fruit (Physalis peruvianaL.) against Acute Renal Injury in Rats. The Scientific World Journal. 2014;2014:1–7. doi: 10.1155/2014/273870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Martínez W, Ospina LF, Granados D, Delgado G. In vitro studies on the relationship between the anti-inflammatory activity of Physalis peruviana extracts and the phagocytic process. Immunopharmacol Immunotoxicol. 2010;32(1):63–73. doi: 10.3109/08923970903143957. [DOI] [PubMed] [Google Scholar]
- 77.Kurapati Kesava Rao Venkata, Atluri Venkata Subba Rao, Samikkannu Thangavel, Nair Madhavan P. N. Ashwagandha (Withania somnifera) Reverses β-Amyloid1-42 Induced Toxicity in Human Neuronal Cells: Implications in HIV-Associated Neurocognitive Disorders (HAND) PLoS ONE. 2013;8(10):e77624. doi: 10.1371/journal.pone.0077624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Aalinkeel R, Hu Z, Nair BB, Sykes DE, Reynolds JL, Mahajan SD, et al. Genomic analysis highlights the role of the JAK-STAT signaling in the anti-proliferative effects of dietary flavonoid—’Ashwagandha’ in prostate cancer cells. Evid Based Complement Altern Med. 2010;7(2):177–187. doi: 10.1093/ecam/nem184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Pawar P, Gilda S, Sharma S, Jagtap S, Paradkar A, Mahadik K, et al. Rectal gel application of Withania somnifera root extract expounds anti-inflammatory and muco-restorative activity in TNBS-induced Inflammatory Bowel Disease. BMC Complement Altern Med. 2011;11(1):1–9. doi: 10.1186/1472-6882-11-34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Kataria H, Wadhwa R, Kaul SC, Kaur G. Water extract from the leaves of Withania somnifera protect RA differentiated C6 and IMR-32 cells against glutamate-induced excitotoxicity. PLoS ONE. 2012;7(5):e37080. doi: 10.1371/journal.pone.0037080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Ojha Shreesh, Alkaabi Juma, Amir Naheed, Sheikh Azimullah, Agil Ahmad, Fahim Mohamed Abdelmonem, Adem Abdu. Withania coagulansFruit Extract Reduces Oxidative Stress and Inflammation in Kidneys of Streptozotocin-Induced Diabetic Rats. Oxidative Medicine and Cellular Longevity. 2014;2014:1–9. doi: 10.1155/2014/201436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Chaplin DD. Overview of the immune response. J Allergy Clin Immunol. 2010;125(2 Suppl 2):S3–S23. doi: 10.1016/j.jaci.2009.12.980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Mikolai J, Erlandsen A, Murison A, Brown KA, Gregory WL, Raman-Caplan P, et al. In vivo effects of Ashwagandha (Withania somnifera) extract on the activation of lymphocytes. J Altern Complement Med. 2009;15(4):423–430. doi: 10.1089/acm.2008.0215. [DOI] [PubMed] [Google Scholar]
- 84.Davis L, Kuttan G. Effect of Withania somnifera on cell mediated immune responses in mice. J Exp Clin Cancer Res. 2002;21(4):585–590. [PubMed] [Google Scholar]
- 85.Khan B, Ahmad SF, Bani S, Kaul A, Suri KA, Satti NK, et al. Augmentation and proliferation of T lymphocytes and Th-1 cytokines by Withania somnifera in stressed mice. Int Immunopharmacol. 2006;6(9):1394–1403. doi: 10.1016/j.intimp.2006.04.001. [DOI] [PubMed] [Google Scholar]
- 86.Bani S, Gautam M, Sheikh FA, Khan B, Satti NK, Suri KA, et al. Selective Th1 up-regulating activity of Withania somnifera aqueous extract in an experimental system using flow cytometry. J Ethnopharmacol. 2006;107(1):107–115. doi: 10.1016/j.jep.2006.02.016. [DOI] [PubMed] [Google Scholar]
- 87.Mesaik MA, Zaheer Ul H, Murad S, Ismail Z, Abdullah NR, Gill HK, et al. Biological and molecular docking studies on coagulin-H: Human IL-2 novel natural inhibitor. Mol Immunol. 2006;43(11):1855–1863. doi: 10.1016/j.molimm.2005.10.020. [DOI] [PubMed] [Google Scholar]
- 88.Huang C-F, Ma L, Sun L-J, Ali M, Arfan M, Liu J-W, et al. Immunosuppressive withanolides from withania coagulans. Chem Biodivers. 2009;6(9):1415–1426. doi: 10.1002/cbdv.200800211. [DOI] [PubMed] [Google Scholar]
- 89.Soares MB, Bellintani MC, Ribeiro IM, Tomassini TC, dos Santos RR. Inhibition of macrophage activation and lipopolysaccaride-induced death by seco-steroids purified from Physalis angulata L. Eur J Pharmacol. 2003;459(1):107–112. doi: 10.1016/S0014-2999(02)02829-7. [DOI] [PubMed] [Google Scholar]
- 90.Soares MBP, Brustolim D, Santos LA, Bellintani MC, Paiva FP, Ribeiro YM, et al. Physalins B, F and G, seco-steroids purified from Physalis angulata L., inhibit lymphocyte function and allogeneic transplant rejection. Int Immunopharmacol. 2006;6(3):408–414. doi: 10.1016/j.intimp.2005.09.007. [DOI] [PubMed] [Google Scholar]
- 91.Yu Y, Sun L, Ma L, Li J, Hu L, Liu J. Investigation of the immunosuppressive activity of Physalin H on T lymphocytes. Int Immunopharmacol. 2010;10(3):290–297. doi: 10.1016/j.intimp.2009.11.013. [DOI] [PubMed] [Google Scholar]
- 92.Aggarwal BB, Vijayalekshmi RV, Sung B. Targeting inflammatory pathways for prevention and therapy of cancer: short-term friend, long-term foe. Clin Cancer Res. 2009;15(2):425–430. doi: 10.1158/1078-0432.CCR-08-0149. [DOI] [PubMed] [Google Scholar]
- 93.Costa G, Francisco V, Lopes MC, Cruz MT, Batista MT. intracellular signaling pathways modulated by phenolic compounds: application for new anti-inflammatory drugs discovery. Curr Med Chem. 2012;19(18):2876–2900. doi: 10.2174/092986712800672049. [DOI] [PubMed] [Google Scholar]
- 94.Libby P. Inflammatory mechanisms: the molecular basis of inflammation and disease. Nutr Rev. 2007;65(12 Pt 2):S140–S146. doi: 10.1301/nr.2007.dec.S140-S146. [DOI] [PubMed] [Google Scholar]
- 95.Santangelo C, Vari R, Scazzocchio B, Di Benedetto R, Filesi C, Masella R. Polyphenols, intracellular signalling and inflammation. Ann Ist Super Sanita. 2007;43(4):394–405. [PubMed] [Google Scholar]
- 96.O’Neill LA. Targeting signal transduction as a strategy to treat inflammatory diseases. Nat Rev Drug Discov. 2006;5(7):549–563. doi: 10.1038/nrd2070. [DOI] [PubMed] [Google Scholar]
- 97.Ghosh DM, George BS, Bhatia A, Sidhu OP. Characterization of Withania somnifera leaf transcriptome and expression analysis of pathogenesis-related genes during salicylic acid signaling. PLoS ONE. 2014;9(4):e94803. doi: 10.1371/journal.pone.0094803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Li Q, Verma IM. NF-kappaB regulation in the immune system. Nat Rev Immunol. 2002;2(10):725–734. doi: 10.1038/nri910. [DOI] [PubMed] [Google Scholar]
- 99.Bonizzi G, Karin M. The two NF-kappaB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 2004;25(6):280–288. doi: 10.1016/j.it.2004.03.008. [DOI] [PubMed] [Google Scholar]
- 100.Brasier AR. The NF-kappaB regulatory network. Cardiovasc Toxicol. 2006;6(2):111–130. doi: 10.1385/CT:6:2:111. [DOI] [PubMed] [Google Scholar]
- 101.Gilmore TD. Introduction to NF-kappaB: players, pathways, perspectives. Oncogene. 2006;25(51):6680–6684. doi: 10.1038/sj.onc.1209954. [DOI] [PubMed] [Google Scholar]
- 102.Perkins ND. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat Rev Mol Cell Biol. 2007;8(1):49–62. doi: 10.1038/nrm2083. [DOI] [PubMed] [Google Scholar]
- 103.Tian B, Brasier AR. Identification of a nuclear factor kappa B-dependent gene network. Recent Prog Horm Res. 2003;58:95–130. doi: 10.1210/rp.58.1.95. [DOI] [PubMed] [Google Scholar]
- 104.Karin M. NF-kappaB as a critical link between inflammation and cancer. Cold Spring Harb Perspect Biol. 2009;1(5):a000141. doi: 10.1101/cshperspect.a000141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Darnell JE., Jr STATs and gene regulation. Science. 1997;277(5332):1630–1635. doi: 10.1126/science.277.5332.1630. [DOI] [PubMed] [Google Scholar]
- 106.Darnell JE., Jr Transcription factors as targets for cancer therapy. Nat Rev Cancer. 2002;2(10):740–749. doi: 10.1038/nrc906. [DOI] [PubMed] [Google Scholar]
- 107.Yu H, Pardoll D, Jove R. STATs in cancer inflammation and immunity: a leading role for STAT3. Nat Rev Cancer. 2009;9(11):798–809. doi: 10.1038/nrc2734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Yu H, Jove R. The STATs of cancer—new molecular targets come of age. Nat Rev Cancer. 2004;4(2):97–105. doi: 10.1038/nrc1275. [DOI] [PubMed] [Google Scholar]
- 109.Takeda K, Noguchi K, Shi W, Tanaka T, Matsumoto M, Yoshida N, et al. Targeted disruption of the mouse Stat3 gene leads to early embryonic lethality. Proc Natl Acad Sci USA. 1997;94(8):3801–3804. doi: 10.1073/pnas.94.8.3801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Cheng GZ, Zhang WZ, Sun M, Wang Q, Coppola D, Mansour M, et al. Twist is transcriptionally induced by activation of STAT3 and mediates STAT3 oncogenic function. J Biol Chem. 2008;283(21):14665–14673. doi: 10.1074/jbc.M707429200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Niu G, Wright KL, Huang M, Song L, Haura E, Turkson J, et al. Constitutive Stat3 activity up-regulates VEGF expression and tumor angiogenesis. Oncogene. 2002;21(13):2000–2008. doi: 10.1038/sj.onc.1205260. [DOI] [PubMed] [Google Scholar]
- 112.Ahmad R., Rajabi H., Kosugi M., Joshi M. D., Alam M., Vasir B., Kawano T., Kharbanda S., Kufe D. MUC1-C Oncoprotein Promotes STAT3 Activation in an Autoinductive Regulatory Loop. Science Signaling. 2011;4(160):ra9–ra9. doi: 10.1126/scisignal.2001426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Kiuchi N, Nakajima K, Ichiba M, Fukada T, Narimatsu M, Mizuno K, et al. STAT3 is required for the gp130-mediated full activation of the c-myc gene. J Exp Med. 1999;189(1):63–73. doi: 10.1084/jem.189.1.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Masuda M, Suzui M, Yasumatu R, Nakashima T, Kuratomi Y, Azuma K, et al. Constitutive activation of signal transducers and activators of transcription 3 correlates with cyclin D1 overexpression and may provide a novel prognostic marker in head and neck squamous cell carcinoma. Cancer Res. 2002;62(12):3351–3355. [PubMed] [Google Scholar]
- 115.Lee J, Hahm ER, Singh SV. Withaferin A inhibits activation of signal transducer and activator of transcription 3 in human breast cancer cells. Carcinogenesis. 2010;31(11):1991–1998. doi: 10.1093/carcin/bgq175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Um HJ, Min K-J, Kim DE, Kwon TK. Withaferin A inhibits JAK/STAT3 signaling and induces apoptosis of human renal carcinoma Caki cells. Biochem Biophys Res Commun. 2012;427(1):24–29. doi: 10.1016/j.bbrc.2012.08.133. [DOI] [PubMed] [Google Scholar]
- 117.Schonthaler HB, Guinea-Viniegra J, Wagner EF. Targeting inflammation by modulating the Jun/AP-1 pathway. Ann Rheum Dis. 2011;70(Suppl 1):i109–i112. doi: 10.1136/ard.2010.140533. [DOI] [PubMed] [Google Scholar]
- 118.Zenz R, Wagner EF. Jun signalling in the epidermis: From developmental defects to psoriasis and skin tumors. Int J Biochem Cell Biol. 2006;38(7):1043–1049. doi: 10.1016/j.biocel.2005.11.011. [DOI] [PubMed] [Google Scholar]
- 119.Hess J, Angel P, Schorpp-Kistner M. AP-1 subunits: quarrel and harmony among siblings. J Cell Sci. 2004;117(Pt 25):5965–5973. doi: 10.1242/jcs.01589. [DOI] [PubMed] [Google Scholar]
- 120.Wagner EF, Nebreda AR. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat Rev Cancer. 2009;9(8):537–549. doi: 10.1038/nrc2694. [DOI] [PubMed] [Google Scholar]
- 121.Eferl R, Wagner EF. AP-1: a double-edged sword in tumorigenesis. Nat Rev Cancer. 2003;3(11):859–868. doi: 10.1038/nrc1209. [DOI] [PubMed] [Google Scholar]
- 122.Singh D, Aggarwal A, Maurya R, Naik S. Withania somnifera inhibits NF-kappaB and AP-1 transcription factors in human peripheral blood and synovial fluid mononuclear cells. Phytother Res. 2007;21(10):905–913. doi: 10.1002/ptr.2180. [DOI] [PubMed] [Google Scholar]
- 123.Pratt WB, Toft DO. Regulation of signaling protein function and trafficking by the hsp90/hsp70-based chaperone machinery. Exp Biol Med (Maywood) 2003;228(2):111–133. doi: 10.1177/153537020322800201. [DOI] [PubMed] [Google Scholar]
- 124.Sawai A, Chandarlapaty S, Greulich H, Gonen M, Ye Q, Arteaga CL, et al. Inhibition of Hsp90 down-regulates mutant epidermal growth factor receptor (EGFR) expression and sensitizes EGFR mutant tumors to paclitaxel. Cancer Res. 2008;68(2):589–596. doi: 10.1158/0008-5472.CAN-07-1570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Stebbins CE, Russo AA, Schneider C, Rosen N, Hartl FU, Pavletich NP. Crystal structure of an Hsp90-geldanamycin complex: targeting of a protein chaperone by an antitumor agent. Cell. 1997;89(2):239–250. doi: 10.1016/S0092-8674(00)80203-2. [DOI] [PubMed] [Google Scholar]
- 126.Park HJ, Rayalam S, Della-Fera MA, Ambati S, Yang JY, Baile CA. Withaferin A induces apoptosis and inhibits adipogenesis in 3T3-L1 adipocytes. Biofactors. 2008;33(2):137–148. doi: 10.1002/biof.5520330206. [DOI] [PubMed] [Google Scholar]
- 127.Subramanian C, Zhang H, Gallagher R, Hammer G, Timmermann B, Cohen M. Withanolides are potent novel targeted therapeutic agents against adrenocortical carcinomas. World J Surg. 2014;38(6):1343–1352. doi: 10.1007/s00268-014-2532-0. [DOI] [PubMed] [Google Scholar]
- 128.Samadi AK, Bazzill J, Zhang X, Gallagher R, Zhang H, Gollapudi R, et al. Novel withanolides target medullary thyroid cancer through inhibition of both RET phosphorylation and the mammalian target of rapamycin pathway. Surgery. 2012;152(6):1238–1247. doi: 10.1016/j.surg.2012.08.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Yadav VR, Prasad S, Sung B, Kannappan R, Aggarwal BB. Targeting inflammatory pathways by triterpenoids for prevention and treatment of cancer. Toxins. 2010;2:2428–2466. doi: 10.3390/toxins2102428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Zhang X, Mukerji R, Samadi AK, Cohen MS. Down-regulation of estrogen receptor-alpha and Rearranged during Transfection tyrosine kinase is associated with Withaferin A-induced apoptosis in MCF-7 breast cancer cells. BMC Complement Altern Med. 2011;11:84. doi: 10.1186/1472-6882-11-84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Zhang X, Samadi AK, Roby KF, Timmermann B, Cohen MS. Inhibition of cell growth and induction of apoptosis in ovarian carcinoma cell lines CaOV3 and SKOV3 by natural withanolide Withaferin A. Gynecol Oncol. 2012;124(3):606–612. doi: 10.1016/j.ygyno.2011.11.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Zhang X, Timmermann B, Samadi AK, Cohen MS. Withaferin a induces proteasome-dependent degradation of breast cancer susceptibility gene 1 and heat shock factor 1 proteins in breast cancer cells. ISRN Biochem. 2012;2012:707586. doi: 10.5402/2012/707586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Samadi AK, Cohen SM, Mukerji R, Chaguturu V, Zhang X, Timmermann BN, et al. Natural withanolide withaferin A induces apoptosis in uveal melanoma cells by suppression of Akt and c-MET activation. Tumor Biol. 2012;33(4):1179–1189. doi: 10.1007/s13277-012-0363-x. [DOI] [PubMed] [Google Scholar]
- 134.Cohen Stephanie M., Mukerji Ridhwi, Timmermann Barbara N., Samadi Abbas K., Cohen Mark S. A novel combination of withaferin A and sorafenib shows synergistic efficacy against both papillary and anaplastic thyroid cancers. The American Journal of Surgery. 2012;204(6):895–901. doi: 10.1016/j.amjsurg.2012.07.027. [DOI] [PubMed] [Google Scholar]
- 135.Motiwala Hashim F., Bazzill Joseph, Samadi Abbas, Zhang Huaping, Timmermann Barbara N., Cohen Mark S., Aubé Jeffrey. Synthesis and Cytotoxicity of Semisynthetic Withalongolide A Analogues. ACS Medicinal Chemistry Letters. 2013;4(11):1069–1073. doi: 10.1021/ml400267q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Grogan PT, Sarkaria JN, Timmermann BN, Cohen MS. Oxidative cytotoxic agent withaferin A resensitizes temozolomide-resistant glioblastomas via MGMT depletion and induces apoptosis through Akt/mTOR pathway inhibitory modulation. Invest New Drugs. 2014;32(4):604–617. doi: 10.1007/s10637-014-0084-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Vyas AR, Singh SV. Molecular targets and mechanisms of cancer prevention and treatment by withaferin A, a naturally occurring steroidal lactone. AAPS J. 2014;16(1):1–10. doi: 10.1208/s12248-013-9531-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Grover A, Shandilya A, Agrawal V, Pratik P, Bhasme D, Bisaria VS, et al. Hsp90/Cdc37 chaperone/co-chaperone complex, a novel junction anticancer target elucidated by the mode of action of herbal drug withaferin A. BMC Bioinform. 2011;12(Suppl 1):S30. doi: 10.1186/1471-2105-12-S1-S30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Gambhir L, Checker R, Sharma D, Thoh M, Patil A, Degani M, et al. Thiol dependent NF-kappaB suppression and inhibition of T-cell mediated adaptive immune responses by a naturally occurring steroidal lactone withaferin A. Toxicol Appl Pharmacol. 2015;289(2):297–312. doi: 10.1016/j.taap.2015.09.014. [DOI] [PubMed] [Google Scholar]
- 140.Saha S, Islam MK, Shilpi JA, Hasan S. Inhibition of VEGF: a novel mechanism to control angiogenesis by Withania somnifera’s key metabolite withaferin A. In Silico Pharmacol. 2013;1:11. doi: 10.1186/2193-9616-1-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.White Peter T., Subramanian Chitra, Zhu Qing, Zhang Huaping, Zhao Huiping, Gallagher Robert, Timmermann Barbara N., Blagg Brian S.J., Cohen Mark S. Novel HSP90 inhibitors effectively target functions of thyroid cancer stem cell preventing migration and invasion. Surgery. 2016;159(1):142–151. doi: 10.1016/j.surg.2015.07.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Lee DH, Lim I-H, Sung E-G, Kim J-Y, Song I-H, Park YK, et al. Withaferin A inhibits matrix metalloproteinase-9 activity by suppressing the Akt signaling pathway. Oncol Rep. 2013;30(2):933–938. doi: 10.3892/or.2013.2487. [DOI] [PubMed] [Google Scholar]
- 143.Lee J, Hahm ER, Marcus AI, Singh SV. Withaferin A inhibits experimental epithelial-mesenchymal transition in MCF-10A cells and suppresses vimentin protein level in vivo in breast tumors. Mol Carcinog. 2015;54(6):417–429. doi: 10.1002/mc.22110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Lee J, Sehrawat A, Singh SV. Withaferin A causes activation of Notch2 and Notch4 in human breast cancer cells. Breast Cancer Res Treat. 2012;136(1):45–56. doi: 10.1007/s10549-012-2239-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Amin Hina, Nayak Debasis, ur Rasool Reyaz, Chakraborty Souneek, Kumar Anmol, Yousuf Khalid, Sharma Parduman Raj, Ahmed Zabeer, Sharma Neelam, Magotra Asmita, Mukherjee Debaraj, Kumar Lekha Dinesh, Goswami Anindya. Par-4 dependent modulation of cellular β-catenin by medicinal plant natural product derivative 3-azido Withaferin A. Molecular Carcinogenesis. 2015;55(5):864–881. doi: 10.1002/mc.22328. [DOI] [PubMed] [Google Scholar]
- 146.O’Connell MA, Hayes JD. The Keap1/Nrf2 pathway in health and disease: from the bench to the clinic. Biochem Soc Trans. 2015;43(4):687–689. doi: 10.1042/BST20150069. [DOI] [PubMed] [Google Scholar]
- 147.Hayes JD, Chowdhry S, Dinkova-Kostova AT, Sutherland C. Dual regulation of transcription factor Nrf2 by Keap1 and by the combined actions of beta-TrCP and GSK-3. Biochem Soc Trans. 2015;43(4):611–620. doi: 10.1042/BST20150011. [DOI] [PubMed] [Google Scholar]
- 148.Vaishnavi K, Saxena N, Shah N, Singh R, Manjunath K, Uthayakumar M, et al. Differential activities of the two closely related withanolides, Withaferin A and Withanone: bioinformatics and experimental evidences. PLoS ONE. 2012;7(9):e44419. doi: 10.1371/journal.pone.0044419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Kostecka A, Sznarkowska A, Meller K, Acedo P, Shi Y, Mohammad Sakil HA, et al. JNK-NQO1 axis drives TAp73-mediated tumor suppression upon oxidative and proteasomal stress. Cell Death Dis. 2014;5:e1484. doi: 10.1038/cddis.2014.408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Scholz CC, Taylor CT. Targeting the HIF pathway in inflammation and immunity. Curr Opin Pharmacol. 2013;13(4):646–653. doi: 10.1016/j.coph.2013.04.009. [DOI] [PubMed] [Google Scholar]
- 151.Arya M, Ahmed H, Silhi N, Williamson M, Patel HR. Clinical importance and therapeutic implications of the pivotal CXCL12-CXCR4 (chemokine ligand-receptor) interaction in cancer cell migration. Tumour Biol. 2007;28(3):123–131. doi: 10.1159/000102979. [DOI] [PubMed] [Google Scholar]
- 152.Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100(1):57–70. doi: 10.1016/S0092-8674(00)81683-9. [DOI] [PubMed] [Google Scholar]
- 153.Conway EM, Collen D, Carmeliet P. Molecular mechanisms of blood vessel growth. Cardiovasc Res. 2001;49(3):507–521. doi: 10.1016/S0008-6363(00)00281-9. [DOI] [PubMed] [Google Scholar]
- 154.Laderoute KR, Calaoagan JM, Gustafson-Brown C, Knapp AM, Li GC, Mendonca HL, et al. The response of c-jun/AP-1 to chronic hypoxia is hypoxia-inducible factor 1 alpha dependent. Mol Cell Biol. 2002;22(8):2515–2523. doi: 10.1128/MCB.22.8.2515-2523.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Masoud GN, Li W. HIF-1 pathway: role, regulation and intervention for cancer therapy. Acta Pharm Sin B. 2015;5(5):378–389. doi: 10.1016/j.apsb.2015.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.D’Ignazio L, Bandarra D, Rocha S (2016) NF-kappaB and HIF crosstalk in immune responses. FEBS J 283(3):413–424 [DOI] [PMC free article] [PubMed]
- 157.Semenza GL. Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene. 2010;29(5):625–634. doi: 10.1038/onc.2009.441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Vaupel P. Prognostic potential of the pre-therapeutic tumor oxygenation status. Adv Exp Med Biol. 2009;645:241–246. doi: 10.1007/978-0-387-85998-9_36. [DOI] [PubMed] [Google Scholar]
- 159.Gao R, Shah N, Lee JS, Katiyar SP, Li L, Oh E, et al. Withanone-rich combination of Ashwagandha withanolides restricts metastasis and angiogenesis through hnRNP-K. Mol Cancer Ther. 2014;13(12):2930–2940. doi: 10.1158/1535-7163.MCT-14-0324. [DOI] [PubMed] [Google Scholar]
- 160.Hitchon CA, El-Gabalawy HS. Oxidation in rheumatoid arthritis. Arthritis Res Ther. 2004;6(6):265–278. doi: 10.1186/ar1447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.van’t Hof RJ, Hocking L, Wright PK, Ralston SH. Nitric oxide is a mediator of apoptosis in the rheumatoid joint. Rheumatology (Oxford) 2000;39(9):1004–1008. doi: 10.1093/rheumatology/39.9.1004. [DOI] [PubMed] [Google Scholar]
- 162.Sumantran VN, Chandwaskar R, Joshi AK, Boddul S, Patwardhan B, Chopra A, et al. The relationship between chondroprotective and antiinflammatory effects of Withania somnifera root and glucosamine sulphate on human osteoarthritic cartilage in vitro. Phytother Res. 2008;22(10):1342–1348. doi: 10.1002/ptr.2498. [DOI] [PubMed] [Google Scholar]
- 163.Subbaraju GV, Vanisree M, Rao CV, Sivaramakrishna C, Sridhar P, Jayaprakasam B, et al. Ashwagandhanolide, a bioactive dimeric thiowithanolide isolated from the roots of Withania somnifera. J Nat Prod. 2006;69(12):1790–1792. doi: 10.1021/np060147p. [DOI] [PubMed] [Google Scholar]
- 164.Wruck CJ, Fragoulis A, Gurzynski A, Brandenburg LO, Kan YW, Chan K, et al. Role of oxidative stress in rheumatoid arthritis: insights from the Nrf2-knockout mice. Ann Rheum Dis. 2011;70(5):844–850. doi: 10.1136/ard.2010.132720. [DOI] [PubMed] [Google Scholar]
- 165.Yu SM, Kim SJ. Production of reactive oxygen species by withaferin A causes loss of type collagen expression and COX-2 expression through the PI3K/Akt, p38, and JNK pathways in rabbit articular chondrocytes. Exp Cell Res. 2013;319(18):2822–2834. doi: 10.1016/j.yexcr.2013.08.026. [DOI] [PubMed] [Google Scholar]
- 166.Kim JH, Kim SJ. Overexpression of microRNA-25 by withaferin A induces cyclooxygenase-2 expression in rabbit articular chondrocytes. J Pharmacol Sci. 2014;125(1):83–90. doi: 10.1254/jphs.13232FP. [DOI] [PubMed] [Google Scholar]
- 167.Ganesan K, Sehgal PK, Mandal AB, Sayeed S. Protective effect of Withania somnifera and Cardiospermum halicacabum extracts against collagenolytic degradation of collagen. Appl Biochem Biotechnol. 2011;165(3–4):1075–1091. doi: 10.1007/s12010-011-9326-8. [DOI] [PubMed] [Google Scholar]
- 168.Sumantran VN, Kulkarni A, Boddul S, Chinchwade T, Koppikar SJ, Harsulkar A, et al. Chondroprotective potential of root extracts of Withania somnifera in osteoarthritis. J Biosci. 2007;32(2):299–307. doi: 10.1007/s12038-007-0030-3. [DOI] [PubMed] [Google Scholar]
- 169.Rasool M, Varalakshmi P. Protective effect of Withania somnifera root powder in relation to lipid peroxidation, antioxidant status, glycoproteins and bone collagen on adjuvant-induced arthritis in rats. Fundam Clin Pharmacol. 2007;21(2):157–164. doi: 10.1111/j.1472-8206.2006.00461.x. [DOI] [PubMed] [Google Scholar]
- 170.Khan MA, Subramaneyaan M, Arora VK, Banerjee BD, Ahmed RS. Effect of Withania somnifera (Ashwagandha) root extract on amelioration of oxidative stress and autoantibodies production in collagen-induced arthritic rats. J Complement Integr Med. 2015;12(2):117–125. doi: 10.1515/jcim-2014-0075. [DOI] [PubMed] [Google Scholar]
- 171.Gupta A, Singh S. Evaluation of anti-inflammatory effect of Withania somnifera root on collagen-induced arthritis in rats. Pharm Biol. 2014;52(3):308–320. doi: 10.3109/13880209.2013.835325. [DOI] [PubMed] [Google Scholar]
- 172.Rasool M, Varalakshmi P. Immunomodulatory role of Withania somnifera root powder on experimental induced inflammation: an in vivo and in vitro study. Vascul Pharmacol. 2006;44(6):406–410. doi: 10.1016/j.vph.2006.01.015. [DOI] [PubMed] [Google Scholar]
- 173.Rasool M, Varalakshmi P. Suppressive effect of Withania somnifera root powder on experimental gouty arthritis: an in vivo and in vitro study. Chem Biol Interact. 2006;164(3):174–180. doi: 10.1016/j.cbi.2006.09.011. [DOI] [PubMed] [Google Scholar]
- 174.Chopra A, Lavin P, Patwardhan B, Chitre D. A 32-week randomized, placebo-controlled clinical evaluation of RA-11, an Ayurvedic drug, on osteoarthritis of the knees. J Clin Rheumatol. 2004;10(5):236–245. doi: 10.1097/01.rhu.0000138087.47382.6d. [DOI] [PubMed] [Google Scholar]
- 175.Kumar G, Srivastava A, Sharma SK, Rao TD, Gupta YK. Efficacy & safety evaluation of Ayurvedic treatment (Ashwagandha powder & Sidh Makardhwaj) in rheumatoid arthritis patients: a pilot prospective study. Indian J Med Res. 2015;141(1):100–106. doi: 10.4103/0971-5916.154476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Gottschalk TA, Tsantikos E, Hibbs ML. Pathogenic inflammation and its therapeutic targeting in systemic lupus erythematosus. Front Immunol. 2015;6:550. doi: 10.3389/fimmu.2015.00550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Khor B, Gardet A, Xavier RJ. Genetics and pathogenesis of inflammatory bowel disease. Nature. 2011;474(7351):307–317. doi: 10.1038/nature10209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Yildirim-Toruner Cagri, Diamond Betty. Current and novel therapeutics in the treatment of systemic lupus erythematosus. Journal of Allergy and Clinical Immunology. 2011;127(2):303–312. doi: 10.1016/j.jaci.2010.12.1087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Triantafillidis JK, Merikas E, Georgopoulos F. Current and emerging drugs for the treatment of inflammatory bowel disease. Drug Des Dev Ther. 2011;5:185–210. doi: 10.2147/DDDT.S11290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Minhas U, Minz R, Bhatnagar A. Prophylactic effect of Withania somnifera on inflammation in a non-autoimmune prone murine model of lupus. Drug Discov Ther. 2011;5(4):195–201. doi: 10.5582/ddt.2011.v5.4.195. [DOI] [PubMed] [Google Scholar]
- 181.Minhas U, Minz R, Das P, Bhatnagar A. Therapeutic effect of Withania somnifera on pristane-induced model of SLE. Inflammopharmacology. 2012;20(4):195–205. doi: 10.1007/s10787-011-0102-8. [DOI] [PubMed] [Google Scholar]
- 182.Samadi AK. Potential anticancer properties and mechanisms of action of withanolides. Enzymes. 2015;37:73–94. doi: 10.1016/bs.enz.2015.05.002. [DOI] [PubMed] [Google Scholar]
- 183.Dar NJ, Hamid A, Ahmad M. Pharmacologic overview of Withania somnifera, the Indian Ginseng. Cell Mol Life Sci. 2015;72(23):4445–4460. doi: 10.1007/s00018-015-2012-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Widodo N, Priyandoko D, Shah N, Wadhwa R, Kaul SC. Selective killing of cancer cells by Ashwagandha leaf extract and its component withanone involves ROS signaling. PLoS ONE. 2010;5(10):e13536. doi: 10.1371/journal.pone.0013536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Nishikawa Y, Okuzaki D, Fukushima K, Mukai S, Ohno S, Ozaki Y, et al. Withaferin A induces cell death selectively in androgen-independent prostate cancer cells but not in normal fibroblast cells. PLoS ONE. 2015;10(7):e0134137. doi: 10.1371/journal.pone.0134137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Park JW, Min KJ, Kim DE, Kwon TK. Withaferin A induces apoptosis through the generation of thiol oxidation in human head and neck cancer cells. Int J Mol Med. 2015;35(1):247–252. doi: 10.3892/ijmm.2014.1983. [DOI] [PubMed] [Google Scholar]
- 187.Samadi AK, Mukerji R, Shah A, Timmermann BN, Cohen MS. A novel RET inhibitor with potent efficacy against medullary thyroid cancer in vivo. Surgery. 2010;148(6):1228–1236. doi: 10.1016/j.surg.2010.09.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Henrich CJ, Brooks AD, Erickson KL, Thomas CL, Bokesch HR, Tewary P, et al. Withanolide E sensitizes renal carcinoma cells to TRAIL-induced apoptosis by increasing cFLIP degradation. Cell Death Dis. 2015;6:e1666. doi: 10.1038/cddis.2015.38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Thaiparambil JT, Bender L, Ganesh T, Kline E, Patel P, Liu Y, et al. Withaferin A inhibits breast cancer invasion and metastasis at sub-cytotoxic doses by inducing vimentin disassembly and serine 56 phosphorylation. Int J Cancer. 2011;129(11):2744–2755. doi: 10.1002/ijc.25938. [DOI] [PubMed] [Google Scholar]
- 190.Yang Z, Garcia A, Xu S, Powell DR, Vertino PM, Singh S, et al. Withania somnifera root extract inhibits mammary cancer metastasis and epithelial to mesenchymal transition. PLoS ONE. 2013;8(9):e75069. doi: 10.1371/journal.pone.0075069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Su B-N, Gu J-Q, Kang Y-H, Park E-J, Pezzuto JM, Kinghorn AD. Induction of the phase II enzyme, quinone reductase, by withanolides and norwithanolides from solanaceous species. Mini Rev Org Chem. 2004;1(1):115–123. doi: 10.2174/1570193043488926. [DOI] [Google Scholar]
- 192.Mathur S, Kaur P, Sharma M, Katyal A, Singh B, Tiwari M, et al. The treatment of skin carcinoma, induced by UV B radiation, using 1-oxo-5beta, 6beta-epoxy-witha-2-enolide, isolated from the roots of Withania somnifera, in a rat model. Phytomedicine. 2004;11(5):452–460. doi: 10.1016/j.phymed.2003.05.004. [DOI] [PubMed] [Google Scholar]
- 193.Padmavathi B, Rath PC, Rao AR, Singh RP. Roots of Withania somnifera inhibit forestomach and skin carcinogenesis in mice. Evid Based Complement Altern Med. 2005;2(1):99–105. doi: 10.1093/ecam/neh064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Li Wenjuan, Zhang Chunjing, Du Hongyan, Huang Vincent, Sun Brandi, Harris John P., Richardson Quitin, Shen Xinggui, Jin Rong, Li Guohong, Kevil Christopher G., Gu Xin, Shi Runhua, Zhao Yunfeng. Withaferin A suppresses the up-regulation of acetyl-coA carboxylase 1 and skin tumor formation in a skin carcinogenesis mouse model. Molecular Carcinogenesis. 2015;55(11):1739–1746. doi: 10.1002/mc.22423. [DOI] [PubMed] [Google Scholar]
- 195.Hahm ER, Lee J, Kim SH, Sehrawat A, Arlotti JA, Shiva SS, et al. Metabolic alterations in mammary cancer prevention by withaferin A in a clinically relevant mouse model. J Natl Cancer Inst. 2013;105(15):1111–1122. doi: 10.1093/jnci/djt153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Kim SH, Singh SV. Mammary cancer chemoprevention by withaferin A is accompanied by in vivo suppression of self-renewal of cancer stem cells. Cancer Prev Res (Phila) 2014;7(7):738–747. doi: 10.1158/1940-6207.CAPR-13-0445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Khazal KF, Hill DL, Grubbs CJ. Effect of Withania somnifera root extract on spontaneous estrogen receptor-negative mammary cancer in MMTV/Neu mice. Anticancer Res. 2014;34(11):6327–6332. [PMC free article] [PubMed] [Google Scholar]
- 198.Kataria Hardeep, Kumar Sushil, Chaudhary Harshita, Kaur Gurcharan. Withania somnifera Suppresses Tumor Growth of Intracranial Allograft of Glioma Cells. Molecular Neurobiology. 2015;53(6):4143–4158. doi: 10.1007/s12035-015-9320-1. [DOI] [PubMed] [Google Scholar]
- 199.McKenna MK, Gachuki BW, Alhakeem SS, Oben KN, Rangnekar VM, Gupta RC, et al. Anti-cancer activity of withaferin A in B-cell lymphoma. Cancer Biol Ther. 2015;16(7):1088–1098. doi: 10.1080/15384047.2015.1046651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Kakar SS, Ratajczak MZ, Powell KS, Moghadamfalahi M, Miller DM, Batra SK, et al. Withaferin a alone and in combination with cisplatin suppresses growth and metastasis of ovarian cancer by targeting putative cancer stem cells. PLoS ONE. 2014;9(9):e107596. doi: 10.1371/journal.pone.0107596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Fong MY, Jin S, Rane M, Singh RK, Gupta R, Kakar SS. Withaferin A synergizes the therapeutic effect of doxorubicin through ROS-mediated autophagy in ovarian cancer. PLoS ONE. 2012;7(7):e42265. doi: 10.1371/journal.pone.0042265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Yang H, Shi G, Dou QP. The tumor proteasome is a primary target for the natural anticancer compound Withaferin A isolated from “Indian winter cherry”. Mol Pharmacol. 2007;71(2):426–437. doi: 10.1124/mol.106.030015. [DOI] [PubMed] [Google Scholar]
- 203.Srinivasan S, Ranga RS, Burikhanov R, Han SS, Chendil D. Par-4-dependent apoptosis by the dietary compound withaferin A in prostate cancer cells. Cancer Res. 2007;67(1):246–253. doi: 10.1158/0008-5472.CAN-06-2430. [DOI] [PubMed] [Google Scholar]
- 204.Lahat G, Zhu QS, Huang KL, Wang S, Bolshakov S, Liu J, et al. Vimentin is a novel anti-cancer therapeutic target; insights from in vitro and in vivo mice xenograft studies. PLoS ONE. 2010;5(4):e10105. doi: 10.1371/journal.pone.0010105. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 205.Munagala R, Kausar H, Munjal C, Gupta RC. Withaferin A induces p53-dependent apoptosis by repression of HPV oncogenes and upregulation of tumor suppressor proteins in human cervical cancer cells. Carcinogenesis. 2011;32(11):1697–1705. doi: 10.1093/carcin/bgr192. [DOI] [PubMed] [Google Scholar]
- 206.Yang H, Wang Y, Cheryan VT, Wu W, Cui CQ, Polin LA, et al. Withaferin A inhibits the proteasome activity in mesothelioma in vitro and in vivo. PLoS ONE. 2012;7(8):e41214. doi: 10.1371/journal.pone.0041214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Biswal BM, Sulaiman SA, Ismail HC, Zakaria H, Musa KI. Effect of Withania somnifera (Ashwagandha) on the development of chemotherapy-induced fatigue and quality of life in breast cancer patients. Integr Cancer Ther. 2013;12(4):312–322. doi: 10.1177/1534735412464551. [DOI] [PubMed] [Google Scholar]
- 208.Amor S, Peferoen LA, Vogel DY, Breur M, van der Valk P, Baker D, et al. Inflammation in neurodegenerative diseases–an update. Immunology. 2014;142(2):151–166. doi: 10.1111/imm.12233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Amor S, Woodroofe MN. Innate and adaptive immune responses in neurodegeneration and repair. Immunology. 2014;141(3):287–291. doi: 10.1111/imm.12134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Zhao J, Nakamura N, Hattori M, Kuboyama T, Tohda C, Komatsu K. Withanolide derivatives from the roots of Withania somnifera and their neurite outgrowth activities. Chem Pharm Bull (Tokyo) 2002;50(6):760–765. doi: 10.1248/cpb.50.760. [DOI] [PubMed] [Google Scholar]
- 211.Kuboyama T, Tohda C, Komatsu K. Neuritic regeneration and synaptic reconstruction induced by withanolide A. Br J Pharmacol. 2005;144(7):961–971. doi: 10.1038/sj.bjp.0706122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Martorana F, Guidotti G, Brambilla L, Rossi D. Withaferin A inhibits nuclear factor-kappaB-dependent pro-inflammatory and stress response pathways in the astrocytes. Neural Plast. 2015;2015:381964. doi: 10.1155/2015/381964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Patil SP, Maki S, Khedkar SA, Rigby AC, Chan C. Withanolide A and asiatic acid modulate multiple targets associated with amyloid-beta precursor protein processing and amyloid-beta protein clearance. J Nat Prod. 2010;73(7):1196–1202. doi: 10.1021/np900633j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Choudhary MI, Yousuf S, Nawaz SA, Ahmed S, Atta ur R. Cholinesterase inhibiting withanolides from Withania somnifera. Chem Pharm Bull (Tokyo) 2004;52(11):1358–1361. doi: 10.1248/cpb.52.1358. [DOI] [PubMed] [Google Scholar]
- 215.Riaz N, Malik A, Nawaz SA, Muhammad P, Choudhary MI. Cholinesterase-inhibiting withanolides from Ajuga bracteosa. Chem Biodivers. 2004;1(9):1289–1295. doi: 10.1002/cbdv.200490091. [DOI] [PubMed] [Google Scholar]
- 216.Grover A, Shandilya A, Agrawal V, Bisaria VS, Sundar D. Computational evidence to inhibition of human acetyl cholinesterase by withanolide a for Alzheimer treatment. J Biomol Struct Dyn. 2012;29(4):651–662. doi: 10.1080/07391102.2012.10507408. [DOI] [PubMed] [Google Scholar]
- 217.Durg S, Dhadde SB, Vandal R, Shivakumar BS, Charan CS. Withania somnifera (Ashwagandha) in neurobehavioural disorders induced by brain oxidative stress in rodents: a systematic review and meta-analysis. J Pharm Pharmacol. 2015;67(7):879–899. doi: 10.1111/jphp.12398. [DOI] [PubMed] [Google Scholar]
- 218.Sehgal N, Gupta A, Valli RK, Joshi SD, Mills JT, Hamel E, et al. Withania somnifera reverses Alzheimer’s disease pathology by enhancing low-density lipoprotein receptor-related protein in liver. Proc Natl Acad Sci. 2012;109(9):3510–3515. doi: 10.1073/pnas.1112209109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Galvan A, Wichmann T. Pathophysiology of parkinsonism. Clin Neurophysiol. 2008;119(7):1459–1474. doi: 10.1016/j.clinph.2008.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Przedborski S, Levivier M, Jiang H, Ferreira M, Jackson-Lewis V, Donaldson D, et al. Dose-dependent lesions of the dopaminergic nigrostriatal pathway induced by intrastriatal injection of 6-hydroxydopamine. Neuroscience. 1995;67(3):631–647. doi: 10.1016/0306-4522(95)00066-R. [DOI] [PubMed] [Google Scholar]
- 221.Przedborski S, Vila M. The 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model: a tool to explore the pathogenesis of Parkinson’s disease. Ann N Y Acad Sci. 2003;991:189–198. doi: 10.1111/j.1749-6632.2003.tb07476.x. [DOI] [PubMed] [Google Scholar]
- 222.Prakash J, Yadav SK, Chouhan S, Singh SP. Neuroprotective role of Withania somnifera root extract in maneb-paraquat induced mouse model of parkinsonism. Neurochem Res. 2013;38(5):972–980. doi: 10.1007/s11064-013-1005-4. [DOI] [PubMed] [Google Scholar]
- 223.RajaSankar S, Manivasagam T, Sankar V, Prakash S, Muthusamy R, Krishnamurti A, et al. Withania somnifera root extract improves catecholamines and physiological abnormalities seen in a Parkinson’s disease model mouse. J Ethnopharmacol. 2009;125(3):369–373. doi: 10.1016/j.jep.2009.08.003. [DOI] [PubMed] [Google Scholar]
- 224.Ahmad M, Saleem S, Ahmad AS, Ansari MA, Yousuf S, Hoda MN, et al. Neuroprotective effects of Withania somnifera on 6-hydroxydopamine induced Parkinsonism in rats. Hum Exp Toxicol. 2005;24(3):137–147. doi: 10.1191/0960327105ht509oa. [DOI] [PubMed] [Google Scholar]
- 225.RajaSankar S, Manivasagam T, Surendran S. Ashwagandha leaf extract: a potential agent in treating oxidative damage and physiological abnormalities seen in a mouse model of Parkinson’s disease. Neurosci Lett. 2009;454(1):11–15. doi: 10.1016/j.neulet.2009.02.044. [DOI] [PubMed] [Google Scholar]
- 226.Sankar SR, Manivasagam T, Krishnamurti A, Ramanathan M. The neuroprotective effect of Withania somnifera root extract in MPTP-intoxicated mice: an analysis of behavioral and biochemical variables. Cell Mol Biol Lett. 2007;12(4):473–481. doi: 10.2478/s11658-007-0015-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Prakash J, Chouhan S, Yadav SK, Westfall S, Rai SN, Singh SP. Withania somnifera alleviates parkinsonian phenotypes by inhibiting apoptotic pathways in dopaminergic neurons. Neurochem Res. 2014;39(12):2527–2536. doi: 10.1007/s11064-014-1443-7. [DOI] [PubMed] [Google Scholar]
- 228.Crotti A, Glass CK. The choreography of neuroinflammation in Huntington’s disease. Trends Immunol. 2015;36(6):364–373. doi: 10.1016/j.it.2015.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Kumar P, Kumar A. Effects of root extract of Withania somnifera in 3-nitropropionic acid-induced cognitive dysfunction and oxidative damage in rats. Int J Health Res. 2008;1(3):139–149. [Google Scholar]
- 230.Kumar P, Kumar A. Possible neuroprotective effect of Withania somnifera root extract against 3-nitropropionic acid-induced behavioral, biochemical, and mitochondrial dysfunction in an animal model of Huntington’s disease. J Med Food. 2009;12(3):591–600. doi: 10.1089/jmf.2008.0028. [DOI] [PubMed] [Google Scholar]
- 231.Nagashayana N, Sankarankutty P, Nampoothiri MR, Mohan PK, Mohanakumar KP. Association of L-DOPA with recovery following Ayurveda medication in Parkinson’s disease. J Neurol Sci. 2000;176(2):124–127. doi: 10.1016/S0022-510X(00)00329-4. [DOI] [PubMed] [Google Scholar]
- 232.Tian R, Hou G, Li D, Yuan TF. A possible change process of inflammatory cytokines in the prolonged chronic stress and its ultimate implications for health. Sci World J. 2014;2014:780616. doi: 10.1155/2014/780616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Furtado M, Katzman MA. Neuroinflammatory pathways in anxiety, posttraumatic stress, and obsessive compulsive disorders. Psychiatry Res. 2015;229(1–2):37–48. doi: 10.1016/j.psychres.2015.05.036. [DOI] [PubMed] [Google Scholar]
- 234.Raison CL, Capuron L, Miller AH. Cytokines sing the blues: inflammation and the pathogenesis of depression. Trends Immunol. 2006;27(1):24–31. doi: 10.1016/j.it.2005.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Tohmi M, Tsuda N, Watanabe Y, Kakita A, Nawa H. Perinatal inflammatory cytokine challenge results in distinct neurobehavioral alterations in rats: implication in psychiatric disorders of developmental origin. Neurosci Res. 2004;50(1):67–75. doi: 10.1016/j.neures.2004.05.010. [DOI] [PubMed] [Google Scholar]
- 236.Potvin S, Stip E, Sepehry AA, Gendron A, Bah R, Kouassi E. Inflammatory cytokine alterations in schizophrenia: a systematic quantitative review. Biol Psychiatry. 2008;63(8):801–808. doi: 10.1016/j.biopsych.2007.09.024. [DOI] [PubMed] [Google Scholar]
- 237.Goldstein BI, Kemp DE, Soczynska JK, McIntyre RS. Inflammation and the phenomenology, pathophysiology, comorbidity, and treatment of bipolar disorder: a systematic review of the literature. J Clin Psychiatry. 2009;70(8):1078–1090. doi: 10.4088/JCP.08r04505. [DOI] [PubMed] [Google Scholar]
- 238.Berk M, Kapczinski F, Andreazza AC, Dean OM, Giorlando F, Maes M, et al. Pathways underlying neuroprogression in bipolar disorder: focus on inflammation, oxidative stress and neurotrophic factors. Neurosci Biobehav Rev. 2011;35(3):804–817. doi: 10.1016/j.neubiorev.2010.10.001. [DOI] [PubMed] [Google Scholar]
- 239.Gray SM, Bloch MH. Systematic review of proinflammatory cytokines in obsessive–compulsive disorder. Curr Psychiatry Rep. 2012;14(3):220–228. doi: 10.1007/s11920-012-0272-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Rege NN, Thatte UM, Dahanukar SA. Adaptogenic properties of six rasayana herbs used in Ayurvedic medicine. Phytother Res. 1999;13(4):275–291. doi: 10.1002/(SICI)1099-1573(199906)13:4<275::AID-PTR510>3.0.CO;2-S. [DOI] [PubMed] [Google Scholar]
- 241.Panossian A, Wikman G. Evidence-based efficacy of adaptogens in fatigue, and molecular mechanisms related to their stress-protective activity. Curr Clin Pharmacol. 2009;4(3):198–219. doi: 10.2174/157488409789375311. [DOI] [PubMed] [Google Scholar]
- 242.Prud’homme GJ, Glinka Y, Wang Q. Immunological GABAergic interactions and therapeutic applications in autoimmune diseases. Autoimmun Rev. 2015;14(11):1048–1056. doi: 10.1016/j.autrev.2015.07.011. [DOI] [PubMed] [Google Scholar]
- 243.Mehta AK, Binkley P, Gandhi SS, Ticku MK. Pharmacological effects of Withania somnifera root extract on GABAA receptor complex. Indian J Med Res. 1991;94:312–315. [PubMed] [Google Scholar]
- 244.Candelario M, Cuellar E, Reyes-Ruiz JM, Darabedian N, Feimeng Z, Miledi R, et al. Direct evidence for GABAergic activity of Withania somnifera on mammalian ionotropic GABAA and GABArho receptors. J Ethnopharmacol. 2015;171:264–272. doi: 10.1016/j.jep.2015.05.058. [DOI] [PubMed] [Google Scholar]
- 245.Bhattacharya SK, Muruganandam AV. Adaptogenic activity of Withania somnifera: an experimental study using a rat model of chronic stress. Pharmacol Biochem Behav. 2003;75(3):547–555. doi: 10.1016/S0091-3057(03)00110-2. [DOI] [PubMed] [Google Scholar]
- 246.Gupta GL, Rana AC. Protective effect of Withania somnifera dunal root extract against protracted social isolation induced behavior in rats. Indian J Physiol Pharmacol. 2007;51(4):345–353. [PubMed] [Google Scholar]
- 247.Gupta GL, Rana AC. Effect of Withania somnifera Dunal in ethanol-induced anxiolysis and withdrawal anxiety in rats. Indian J Exp Biol. 2008;46(6):470–475. [PubMed] [Google Scholar]
- 248.Bhattacharya SK, Bhattacharya A, Sairam K, Ghosal S. Anxiolytic-antidepressant activity of Withania somnifera glycowithanolides: an experimental study. Phytomedicine. 2000;7(6):463–469. doi: 10.1016/S0944-7113(00)80030-6. [DOI] [PubMed] [Google Scholar]
- 249.Kumar A, Kalonia H. Protective effect of Withania somnifera Dunal on the behavioral and biochemical alterations in sleep-disturbed mice (Grid over water suspended method) Indian J Exp Biol. 2007;45(6):524–528. [PubMed] [Google Scholar]
- 250.Shah PC, Trivedi NA, Bhatt JD, Hemavathi KG. Effect of Withania somnifera on forced swimming test induced immobility in mice and its interaction with various drugs. Indian J Physiol Pharmacol. 2006;50(4):409–415. [PubMed] [Google Scholar]
- 251.Kaurav BP, Wanjari MM, Chandekar A, Chauhan NS, Upmanyu N. Influence of Withania somnifera on obsessive compulsive disorder in mice. Asian Pac J Trop Med. 2012;5(5):380–384. doi: 10.1016/S1995-7645(12)60063-7. [DOI] [PubMed] [Google Scholar]
- 252.Cooley K, Szczurko O, Perri D, Mills EJ, Bernhardt B, Zhou Q, et al. Naturopathic care for anxiety: a randomized controlled trial ISRCTN78958974. PLoS ONE. 2009;4(8):e6628. doi: 10.1371/journal.pone.0006628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Andrade C, Aswath A, Chaturvedi SK, Srinivasa M, Raguram R. A double-blind, placebo-controlled evaluation of the anxiolytic efficacy of an ethanolic extract of withania somnifera. Indian J Psychiatry. 2000;42(3):295–301. [PMC free article] [PubMed] [Google Scholar]
- 254.Chandrasekhar K, Kapoor J, Anishetty S. A prospective, randomized double-blind, placebo-controlled study of safety and efficacy of a high-concentration full-spectrum extract of ashwagandha root in reducing stress and anxiety in adults. Indian J Psychol Med. 2012;34(3):255–262. doi: 10.4103/0253-7176.106022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Auddy B, Hazra J, Mitra A, Abedon B, Ghosal S. A standardized Withania somnifera extract significantly reduces stress-related parameters in chronically stressed humans: a double-blind, randomized, placebo-controlled study. J Am Neutraceut Assoc. 2008;11:50–56. [Google Scholar]
- 256.Khyati S, Anup T. A randomized double blind placebo controlled study of ashwagandha on generalized anxiety disorder. Int Ayurvedic Med J. 2013;1:1–7. [Google Scholar]
- 257.Pratte MA, Nanavati KB, Young V, Morley CP. An alternative treatment for anxiety: a systematic review of human trial results reported for the Ayurvedic herb ashwagandha (Withania somnifera) J Altern Complement Med. 2014;20(12):901–908. doi: 10.1089/acm.2014.0177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Pingali U, Pilli R, Fatima N. Effect of standardized aqueous extract of Withania somnifera on tests of cognitive and psychomotor performance in healthy human participants. Pharmacogn Res. 2014;6(1):12–18. doi: 10.4103/0974-8490.122912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Chengappa KN, Bowie CR, Schlicht PJ, Fleet D, Brar JS, Jindal R. Randomized placebo-controlled adjunctive study of an extract of withania somnifera for cognitive dysfunction in bipolar disorder. J Clin Psychiatry. 2013;74(11):1076–1083. doi: 10.4088/JCP.13m08413. [DOI] [PubMed] [Google Scholar]
- 260.Chengappa KN (2014) Withania Somnifera: an immunomodulator and anti-inflammatory agent for schizophrenia. In: ClinicalTrials.gov. National Library of Medicine (US), Bethesda (MD). [cited 2015 Dec 10]. https://clinicaltrials.gov/show/NCT01793935. p NLM identifier: NCT01793935