

Abstract
Enveloped viruses, which include many medically important viruses such as human immunodeficiency virus, influenza virus and hepatitis C virus, are intracellular parasites that acquire lipid envelopes from their host cells. Success of replication is intimately linked to their ability to hijack host cell mechanisms, particularly those related to membrane dynamics and lipid metabolism. Despite recent progress, our knowledge of lipid mediated virus–host interactions remains highly incomplete. In addition, diverse experimental systems are used to study different stages of virus replication thus complicating comparisons. This review aims to present a unifying view of the widely diverse strategies used by enveloped viruses at distinct stages of their replication cycles.
Keywords: Lipidomics, Enveloped virus, Membrane, Receptor, Glycolipid, Lipid–protein interactions
1. Introduction
In recent years, the contribution of lipids to virus replication has gained increasing attention. Lipids can be envisioned as ‘multidimensional’ entities. From structurally diverse single lipid molecules (Fig. 1A), they form larger molecular aggregates as bilayers in organelle membranes. Lipids are heterogeneously distributed throughout the cell and show predominant accumulation within certain subcellular locations. Within biological membranes, they further exhibit lateral heterogeneity through the formation of specialized microdomains and by establishing asymmetry across bilayers. The overall structural and topological diversity of lipids and their aggregates results in a wide range on specific protein–lipid interactions and electrostatic considerations, which translate to potentially numerous functional roles in the replication of enveloped viruses (Table 1 ).
Fig. 1.
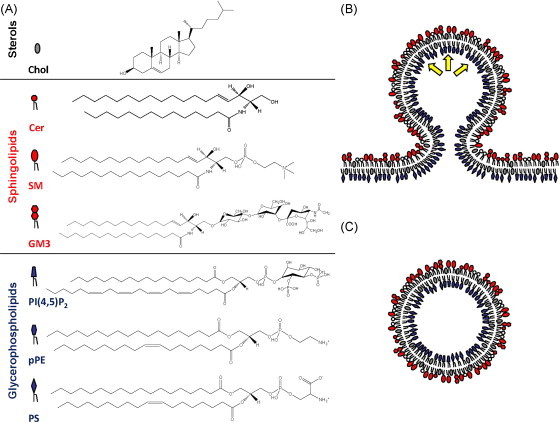
(A) Overview of lipid classes discussed in this review: cholesterol (Chol), ceramide (Cer), sphingomyelin (SM), ganglioside GM3 (GM3), phosphatidylinositol-4,5-bis-phosphate (PI(4,5)P2), plasmalogen phosphatidylethanolamine (pPE), phosphatidylserine (PS) and cardiolipin (CL); (B) lipid-bilayer arrangement during virus budding: lipid species are coloured according to their head group as described in (A). Lipids are asymmetrically distributed in the plasma membrane and virus budding occurs at sites enriched in cholesterol, sphingolipids and phosphoinositides. Yellow arrows indicate the targeting of proteins to the plasma membrane through electrostatic interaction with phosphoinositides. This results in a pushing-force finally leading to virus budding. (C) Lipid distribution in the virus envelope: enrichment of sphingolipids and cholesterol in the lipid envelope bestows fluidity and stability to the virus. Such and other lipids have also an important role in virus entry.
Table 1.
Summary of examples of virus–lipid interactions discussed in this review.
| Stage | Lipid class | Mode of action | Virus examples |
|---|---|---|---|
| Docking/fusion | Phosphatidylserines | PS addition causes a “glycosylation-specific enhancement” of virus entry | HIV (Coil and Miller, 2005b, Coil and Miller, 2005a) |
| Annexins are hijacked as co-receptors by PS expressed in the virus envelope | Influenza virus (Huang et al., 1996), HBV (Hertogs et al., 1993), HCMV (Raynor et al., 1999), RSVa (Malhotra et al., 2003), HIV (Callahan et al., 2003, Ma et al., 2004) | ||
| Molecular mimicry of apoptotic bodies facilitates macropinocytosis/virus entry | VACV (Mercer and Helenius, 2008) | ||
| Sterols/Sterol Esters | Association with apolipoproteins facilitates virus docking and subsequent entry steps | HCV (Prince et al., 1996, Agnello et al., 1999, Molina et al., 2007) | |
| Virus surface glycoproteins and receptors are clustered in “lipid rafts” | HCV (Kapadia et al., 2007, Burlone and Budkowska, 2009), HIV (Sorice et al., 2000, Sorice et al., 2001, Nguyen and Taub, 2002, Popik et al., 2002, Bhattacharya et al., 2004, Nguyen et al., 2005), RSVa (Fleming et al., 2006), influenza virus (Takeda et al., 2003) | ||
| Sphingolipids | Glycan–glycan interactions between host and virus stabilize virus attachment | Influenza virus (Huang, 1976, Leskawa et al., 1986, Kasson and Pande, 2008) | |
| GalCer, GM3 and Gb3 serve as auxiliary receptors for virus entry | HIV (Fantini et al., 1993, Seddiki et al., 1996, Hammache et al., 1999, Hug et al., 2000, Magerus-Chatinet et al., 2007) | ||
| GSLs aggregate viral receptors which have low endogenous cellular levels | EBOV, Marburg virus, VACV, HSV (reviewed in Teissier and Pecheur, 2007) | ||
| Gangliosides serve as main receptors for virus entry | SeV (Markwell et al., 1981, Hansson et al., 1984, Markwell et al., 1984, Muthing and Unland, 1994, Kaneda et al., 1997), NDV (Ferreira et al., 2004a, Ferreira et al., 2004b, Villar and Barroso, 2006) | ||
| Replication/translation | Phosphatidylserines | PS enriched in endosomal membranes is required by NSP1 for capping of viral mRNAs | SFV (Ahola et al., 1999) |
| Sterols/sterol esters | Nef accessory protein enhances cholesterol biosynthesis and impairs ABCA1 dependent cholesterol efflux | HIV (Mujawar et al., 2006) | |
| Sphingolipids | Enhanced levels of cholesterol are required to support formation of “lipid-raft” structures | RSVa (Money et al., 2009) | |
| Increased activity of SphK1 promotes accumulation of transcriptional activator IE1 | HCMV (Machesky et al., 2008) | ||
| Fatty acids | Genome replication is stimulated by saturated or monounsaturated fatty acids. FAS is highly enriched in the supernatant of infected cells | HCV (Su et al., 2002) | |
| Assembly/budding | Phosphoinositides | Gag binds specifically to PI(4,5)P2 causing a myristic switch in MA which increases membrane partitioning of Gag | HIV (Zhou et al., 1994, Ono et al., 2004, Freed, 2006) |
| Sterols | It is also possible that PI(4,5)P2 interactions with cellular proteins contribute to membrane curvature during virus assembly and budding | HIV (Chan et al., 2008) | |
| Surface exposed polybasic domains of similar matrix proteins also use phosphoinositides or other anionic phospholipids for “lipid-raft” targeting/induction of membrane curvature | Influenza virus (Ruigrok et al., 2000, Thaa et al., 2009), EBOV (Timmins et al., 2004), VSV (Gaudier et al., 2002), RSVa (Money et al., 2009) | ||
| Virus assembly occurs on lipid droplets | HCV (Miyanari et al., 2007) | ||
| Sphingolipids | Nef contains cholesterol binding sequence in its C terminus which allows transport to and association with lipid rafts | HIV (Zheng et al., 2003) | |
| Cholesterol depletion enhances virus budding | Influenza virus (Barman and Nayak, 2007) | ||
| Cholesterol depletion inhibits virus production | HIV (Ono and Freed, 2001, Pickl et al., 2001) | ||
| “Lipid-raft” dependent viruses do not co-localize on the plasma membrane | HIV, Influenza virus, EBOV (Khurana et al., 2007, Leung et al., 2008) | ||
| Fatty acids | Several viral proteins have myristoyl and palmitoyl covalent modifications for “lipid-raft” targeting | HIV, Influenza virus, EBOV, Marburg virus, measles virus, NDV, RSVa (reviewed in Hruby and Franke, 1993, Maurer-Stroh and Eisenhaber, 2004) | |
| Extracellular | Sterols/sterol esters | Cholesterol depletion of virus envelopes leads to virus inactivation due to a loss of protein core and genome integrity | HIV and SIV (Graham et al., 2003), influenza virus (Barman and Nayak, 2007, Ambrose et al., 2008) |
| Lipid molecular geometry | Inverted-cone shaped lipids like lyso PC inhibits virus envelope fusion with target membrane while cone shaped lipids like pPE, DAG and ceramide may facilitate membrane dynamics | HIV (Martin and Ruysschaert, 1995, Gunther-Ausborn and Stegmann, 1997, Brugger et al., 2006, Chan et al., 2008), TBEV (Stiasny and Heinz, 2004), influenza virus (Gunther-Ausborn et al., 1995, Chernomordik et al., 1998) | |
We will emphasize general principles of lipid involvement by using examples of well studied viruses as a foundation. Additionally, we will highlight unique cases of lipid function and address controversial concepts in virus budding. In doing so, we hope to stimulate new research efforts to elucidate new lipid functions in enveloped virus replication.
1.1. Virus entry
Enveloped viruses gain entry into their host cells via a two step process. First, the enveloped virus will engage with specific receptors on the cell surface through its fusion proteins in a process known as docking. Next, the interaction of the fusion protein with the receptor will cause a conformational change to the fusion protein which results in apposition and mixing of the viral envelope and cell membrane. This forms a membrane pore that facilitates delivery of virus content into the host cell. The involvement of lipids has been described separately in both processes and at times without a clear distinction between docking and fusion.
1.2. Glycosphingolipids as viral receptors
Cell associated glycosphingolipids (GSLs) have been widely investigated for their role as viral receptors. GSLs are ubiquitous constituents of mammalian plasma membranes, with diverse cellular roles including cell recognition, growth control, differentiation and tumorigenesis (Tettamanti, 2004). Structurally, GSLs contain a lipid group linked to a carbohydrate group via a glycosidic bond. The lipid group is typically made up of a ceramide molecule containing a sphingoid base that is derivatized on the 2-amino group with a fatty acyl chain. The attached carbohydrate group can take the form of uncharged sugars such as glucose, galactose and N-acetylglucoseamine (GlcNAc) or may contain ionized functional groups such as phosphate, sulfate or charged carbohydrate residues such as sialic acid (N-acetylneuraminic acid) in gangliosides. The high density of epitope rich GSLs on mammalian plasma membrane suggests that this class of lipids contribute to enveloped virus entry.Although it is known that the binding of cell surface glycans by influenza hemagglutinin controls viral attachment and infection of host cells, the precise involvement of GSLs have been difficult to resolve. Despite reports that influenza virus hemagglutinin (HA) can directly bind to GSLs (Gambaryan et al., 2004, Kogure et al., 2006, Hidari et al., 2007), there exists contradicting evidence that GSLs are non-essential for influenza virus infection (Ablan et al., 2001, Chu and Whittaker, 2004, Matrosovich et al., 2006). The latter findings are supported by the finding that preferred receptors for influenza virus hemagglutinin are N-linked rather than O-linked glycoproteins (Chu and Whittaker, 2004); GSLs are generally O-linked. Computer simulation of crystallographic data on hemagglutinin–ligand interactions suggests that influenza virus associated glycans contact the host glycans within 1–2 residues of the ligand binding site to stabilize virus binding (Kasson and Pande, 2008). While the virus associated glycans may be presented by glycoproteins, it is also possible that GSLs may be involved since early works showed enrichment of gangliosides in the influenza virus envelope (Huang, 1976, Leskawa et al., 1986). Therefore, GSLs may function as attachment stabilizers rather than receptors per se in influenza virus entry mechanism.Some enveloped viruses are also able to harness GSLs as auxiliary receptors when their natural protein receptors are not presented. While it is well established that HIV infects CD4 expressing immune cells, there are clinical observations that HIV can also infect CD4 negative cells such as neuronal cells (Harouse et al., 1989) and colonic epithelial cells (Fantini et al., 1993). In these cells types, HIV appears to use GSLs as substitute receptors to gain entry. A binding site for galactosyl ceramide has been mapped to the HIV Env gp41 residues 650–685 thereby providing a structural basis for GSLs in HIV entry (Alfsen and Bomsel, 2002, Yu et al., 2008). It is speculated that CD4 independent entry of HIV is mediated by the action of either a combination of CXCR4 and GalCer or solely by Gb3 in absence of CXCR4 from the host cell surface (Viard et al., 2004). Additionally, HIV transcytosis from dendritic cells to T-cells is orchestrated by GalCer (Magerus-Chatinet et al., 2007) and it has been reported that downregulation of GalCer through interferon-γ is part of a protective immune response (Yahi et al., 1994).
Some members of the paramyxovirus family depend solely on host cell associated GSLs as receptors (Villar and Barroso, 2006). Newcastle disease virus (NDV) binds to a broad range of GSLs with sialic acid either attached to terminal sugars (GM3, GD1a and GT1b) or to internal sugars (GM2, GM1 and GD1b) (Ferreira et al., 2004b, Villar and Barroso, 2006). It has been shown that modification of the GSL presentation on host cell surfaces by sialidase treatment clearly impaired NDV entry (Anastasia et al., 2008). Moreover, gangliosides have been shown to cause conformational changes in the NDV surface protein known as HN, thus mediating virus fusion (Ferreira et al., 2004a). Importantly, one should note that other paramyxoviruses exhibit different ganglioside specificities as well. For example, Sendai virus interact with gangliosides having a terminally attached sialic acid such as GD1a, GT1b, and GQ1b, and also recognize neolacto-series gangliosides (Markwell et al., 1981, Markwell et al., 1984, Hansson et al., 1984, Muthing and Unland, 1994, Kaneda et al., 1997).
1.3. Phosphatidylserine and its binding proteins are implicated in enveloped virus entry
Besides GSLs, phosphatidylserine (PS) has also been implicated in enhancing enveloped virus entry. When target cells bearing functional enveloped virus receptors are pre-treated with PS liposomes, cell surface PS level is increased and the specific enveloped virus infectivity is enhanced by up to 20-fold (Coil and Miller, 2005a). This effect is limited to enveloped viruses and does not occur through increases in virus receptor levels or virus binding, thereby indicating that virus fusion in enhanced. In some cases, cell specific glycosylation of viral receptors near the active virus binding site result in a block to functional receptor–virus interactions. However, with pre-treatment of PS liposomes, such blocks are relieved and the cells become permissible to retrovirus infection. This effect is referred to as “glycosylation-specific enhancement” by PS (Coil and Miller, 2005b).
Virus associated PS also appear to enhance enveloped virus entry. Lipid asymmetry exists in all resting mammalian cells. PS and phosphatidylethanolamine (PE) are lipids oriented towards the cytoplasm, while phosphatidylcholine (PC) and sphingomyelin (SM) are oriented towards the extracellular space. Virus infection of host cells can activate apoptosis which is accompanied by the externalization of PS to the outer leaflet of the plasma membrane (Balasubramanian and Schroit, 2003). This has been documented during HIV infection of macrophage cells. HIV virions produced from these cells are decorated with PS on the outer surface of their envelope (Callahan et al., 2003). In this system, incubation of PS-specific binding protein Annexin V or PS with purified HIV particles reduces its ability to infect macrophages but not T-cells. Later studies identified the macrophage PS-binding protein annexin II as the cellular cofactor supporting macrophage HIV-1 infection (Ma et al., 2004).
Numerous facets of virus fusion may be enhanced by a large increase in plasma membrane or virus coat PS concentration, including lipid packing, alterations in bilayer curvature, changes in membrane fluidity and locally induced changes in the bilayer phase. However, the involvement of annexin II highlights a specific structural mechanism by which PS enhances virus fusion through interaction with the annexin protein family. After all, a number of enveloped viruses including influenza virus (Huang et al., 1996), Hepatitis B virus (Hertogs et al., 1993), human cytomegalovirus (Raynor et al., 1999) and respiratory synticial virus (Malhotra et al., 2003) have been shown to use different annexins as co-receptors for entry. The hijacking of annexin as viral co-receptors may therefore represent a way for viruses to overcome host cell apoptosis and maximize infectivity of virus particles that are enriched with PS molecules on the outer virus coat.
Lastly, another interesting example of PS involvement during enveloped virus entry comes from a recent study of vaccinia virus (Mercer and Helenius, 2008). As with other viruses, vaccinia virus infection leads to apoptosis and production of virus particles that present PS on the outer layer of their envelope. This virus appears to use externalized PS in macropinocytosis and apoptotic mimicry to infect host cells and these effects can be inhibited by incubating the virus with annexin (Mercer and Helenius, 2008). Such a mechanism explains why vaccinia virus is able to infect many different cell types, because PS-mediated macropinocytosis of apoptotic material is common to most cells (Henson et al., 2001, Albert, 2004). In addition, the virus may avoidimmune detection by mimicking apoptotic bodies since clearance of apoptotic debris suppresses the activation of innate immune responses (Albert, 2004). However, the application of this mechanism to other viruses may be limited because it appears to work in favor of vaccinia virus and other enveloped pathogens of large sizes that are too big for regular endocytic mechanisms but are similar in size to apoptotic bodies (Henson et al., 2001).
1.4. Cholesterol transport pathway is a facilitator of virus entry
The entry mechanism of hepatitis C virus offers another interesting example of molecular mimicry. It has been shown that infectious HCV particles in patient sera commonly associate with apolipoproteins and are of low density, i.e. very low density lipoprotein (VLDL)-HCV (Thomssen et al., 1992, Agnello et al., 1999). This is because high density HCV particles are degraded in the host cells, whereas only VLDL-HCV particles are successfully released from infected cells (Gastaminza et al., 2006, Gastaminza et al., 2008). The VLDL-HCV association is important in engaging host cell lipoprotein receptors (e.g. human scavenger receptor B1 or LDL receptor) through an apolipoprotein mediated mechanism (Agnello et al., 1999, Andre et al., 2002, Molina et al., 2007). This, in turn, facilitates the CD81 dependent transport of HCV to tight junction proteins claudin-1 and occludin (Brazzoli et al., 2008) which finally leads to the uptake of HCV particles into host cells by endocytosis. In this way, HCV actually mimics the molecular identity of lipoprotein to hijack lipoprotein transport mechanism for its entry into its host. Furthermore, human scavenger receptor B1 has been additionally suggested to be a crucial mediator of HCV entry by regulating CD81 clustering into cholesterol-enriched plasma membrane microdomains (Kapadia et al., 2007, Burlone and Budkowska, 2009).
1.5. The importance of lipid microenvironment on virus protein receptors
Both envelope and host membrane lipids may also play a role in facilitating fusion by stereospecific interaction with the viral fusion machinery partners. A recurring theme for such interactions lies in lipid-raft dependent clustering of receptors. Clustering of sufficient HA proteins into lipid rafts in the virus envelope is required to ensure efficient virus fusion of the next host (Takeda et al., 2003). Other enveloped viruses that display similar mechanisms of raft dependent clustering of fusion glycoproteins include retrovirus Env (Bhattacharya et al., 2004) and RSV F protein (Fleming et al., 2006). The issue of lipid-raft dependent assembly and budding will be dealt with in later sections of this review.
In cells producing functional retroviral receptors, a direct association between the HIV receptor CD4 (Sorice et al., 2000, Popik et al., 2002), co-receptors CCR5 (Nguyen and Taub, 2002, Nguyen et al., 2005) or CXCR4 (Sorice et al., 2001) with lipid rafts has been observed. While CD4 and CCR5 reside in different plasma membrane microenvironments of resting cells, binding of the third variable region V3-containing gp120 core to CD4 and CCR5 induces association between these receptors to form the HIV entry complex. An important requirement in this reaction is the presence of plasma membrane cholesterol since its removal abolishes gp120 core-induced association between CD4 and CCR5. GSLs also play a significant role here. Several studies identified GalCer (Fantini et al., 1993, Seddiki et al., 1996, Magerus-Chatinet et al., 2007), GM3 and Gb3 (Hammache et al., 1999) as major interaction partners during HIV entry (Hug et al., 2000).During the CD4 dependent infection pathway, GM3 and Gb3 come into play only after binding to co-receptors CXCR4/CCR5 (Nehete et al., 2002). On the one hand, when GSLs are expressed in excess in certain cell lines, they become a barrier to prevent HIV receptor clustering, thus blocking subsequent events necessary for the formation of a functional fusion pore and infection (Rawat et al., 2004). On the other hand, when cells express low endogenous levels of viral receptors, cell surface GSLs are known to be involved in promoting receptor aggregation which is required for the formation of the HIV entry complex (Rawat et al., 2006). Numerous other viruses, including Ebola and Marburg viruses, vaccinia virus and herpes simplex virus, has been reported to use similar strategies for co-receptor clustering (reviewed in Teissier and Pecheur, 2007).
1.6. Intracellular expression and replication of viral genomes
The next step in productive infection lies in the expression and replication of viral genomes. It will be clear from numerous studies presented below that successful enveloped virus replication entails extensive manipulation of host cell membranes and lipid metabolism.
1.7. Intracellular membrane structures are modified for virus replication
Positive strand (+) RNA viruses are well known to replicate in the cytoplasm of the host cell in association with cytoplasmic membranes. Since (+) RNA genomes have the same polarity as cellular mRNA, translation of the genome is initiated by the host cell translation machinery into one or multiple viral polyproteins, which are co- and post-translationally processed by viral and host cell proteases. The accumulation of viral-RNA-dependent RNA polymerase (RdRp), various accessory non-structural proteins, viral-RNA and host cell factors induces the deformation of various cellular organelles to create replication complexes (RCs).
For a majority of (+) viruses, including dengue virus (DENV) and severe acute respiratory syndrome (SARS) virus, the endoplasmic reticulum (ER) appears to be the common origin for membrane modification (Miller and Krijnse-Locker, 2008). In the case of HCV, it appears that ER derived membranes are rearranged around lipid droplets (Miyanari et al., 2007). Exceptions include the Semliki Forest Virus and Flock house virus (FHV) which appear to use endosomes/lysosomes and mitochondria membranes respectively to derive their RCs.
There are numerous unanswered questions with regards to the biogenesis and functions of virus-induced membrane alterations. A number of viral and cellular protein factors involved in the formation of RCs have been described. Virtually nothing is known however about specific membrane types and lipid requirements which help regulating the process. As mentioned above, lipids are heterogeneously distributed in different cellular organelles. Thus the association of different (+) RNA viruses with specific cellular organelles would allow speculation that membrane lipid composition may influence virus replication. For instance, SFV non-structural protein (NSP1), the capping enzyme of viral mRNAs, requires association with anionic membrane phospholipids, particularly PS which are enriched in endosomal membranes, for activation (Ahola et al., 1999). Similarly, it was shown that FHV protein A, the essential viral protein necessary for RC assembly in FHV, associates efficiently with anionic phospholipids, and in particular the mitochondrial-specific anionic phospholipid cardiolipin thereby providing a structural basis for mitochondria targeting (Stapleford et al., 2009). The stimulation of HCV genome replication by exogenously added saturated or monounsaturated fatty acids (Kapadia and Chisari, 2005) may also be linked to the enhanced formation for lipid droplet formation seen in HCV infected cells (Miyanari et al., 2007).
1.8. Virus infection impacts host lipid metabolism
Modification of host lipid metabolism by HIV infection has been very well investigated. In fact, an increased risk of arthrosclerosis and coronary artery disease due to dyslipidemia is a recognized clinical problem in HIV-infected patients (Escaut et al., 2003, El-Sadr et al., 2005) as is the appearance of antibodies against GM2 (Wu et al., 1999), GM3 (Griggi et al., 1994), and PS (Silvestris et al., 1996). At the cellular level, it was found that changes in cellular morphology including the appearance of lipid bodies in macrophages, led to the formation of foam cells common in arthrosclerosis (Mujawar et al., 2006). Gene expression analysis of both HIV-infected cells indicated that numerous genes involved in cholesterol metabolism and trafficking are altered by infection (Giri et al., 2006). Interestingly, it was found that expressing the HIV Nef accessory protein alone is able to simultaneously increase cholesterol biosynthesis and impair ATP-binding cassette transporter A1 (ABCA1)-dependent cholesterol efflux from human macrophages (Mujawar et al., 2006). Furthermore, Nef contains a cholesterol binding sequence in its C terminus which leads to re-distribution of cholesterol and progeny viruses to promote infectious HIV particle formation (Zheng et al., 2003).
Other members of the retrovirus family, including MLV (Beer et al., 2003) and SIV (Mansfield et al., 2007), also appear to increase cholesterol loading in infected cells either through enhanced biosynthesis or uptake from extracellular or dietary sources. More recently, it was reported that various genes involved in cholesterol biosynthesis were also up-regulated in paramyxovirus RSV infected cell (Money et al., 2009). A common feature appears to be an association with lipid rafts during assembly and budding of these viruses. Curiously, influenza virus, which also buds from lipid rafts does not show similar manipulation of the cholesterol biosynthetic pathway. Instead, only cases of acute influenza infection with febrile convulsion would result in the enhancement of APOL1 and ABCD2 which are associated with cholesterol levels and fatty acid metabolism respectively (Kawada et al., 2006).
However, the most severe case of virus-induced lipogenesis occurs during HCV infection. Pathologically, this results in liver steatosis which is the process of abnormal retention of lipids in the liver. Gene expression analysis of liver derived cRNA from HCV infected chimpanzees reveal many changes in genes associated with lipid metabolism (Su et al., 2002). Some genes such as peroxisome proliferator activated receptor α (PPAR-α), flotillin 2 and hepatic lipase C are repressed while UDP-glucose ceramide glucosyltransferase (UGCG), serum response element binding protein (SREBP), lipase A and fatty acid synthase (FAS) are expressed at higher levels. More recently, proteomic analysis of HCV infected cell culture supernatant revealed highly enriched fractions of FAS and apolipoprotein E (Yang et al., 2008). These metabolic alteration would likely support the formation of lipid droplets that are for HCV budding.
Interesting studies have also been carried out on human cytomegalovirus (HCMV) infected cells. HCMV appears to increase metabolic flux of citrate through the tricarboxylic acid cycle and its efflux to the fatty acid biosynthesis pathway as malonyl-CoA (Munger et al., 2008). This mirrors a similar need for lipogenesis as in HCV infections. On a separate note, HCMV infection also appears to stimulate sphingolipid biosynthesis in infected host cells during early stage of infection (Machesky et al., 2008). Specifically, HCMV causes an accumulation of sphingosine kinase 1 (SphK1) and increased activity of SphK1, resulting in the increase in dihydrosphingosine-1-phosphate. More importantly, SphK1 activity promotes the accumulation of the HCMV transcriptional transactivator IE1. In addition, there is also enhancement of de novo sphingolipid synthesis that increases dihydrosphingosine and ceramide levels in the infected host cell although the consequences of these effects are still unknown.
1.9. Virus assembly
Viral assembly and budding represents the last stage of the journey through the host cell. At this stage, newly synthesized virus genomes and its protein products are targeted to membrane assembly sites where final budding also takes place. Depending on the virus, these sites can occur at various intracellular organelles such as nuclear envelopes (e.g. herpes simplex virus), ER (e.g. dengue virus), Golgi (e.g. cytomegalovirus) or lipid droplets (e.g. hepatitis C virus) as well as distinct microdomains in the plasma membrane (e.g. human immunodeficiency virus, influenza A virus and respiratory syncitial virus).
1.10. Role of plasma membrane microdomains during virus budding
A widely discussed concept stems from the idea that enveloped viruses are targeted to and bud from specialized plasma membrane microdomains (Fig. 1B). The synergistic and partially lipid-driven packaging of cholesterol, sphingolipids and saturated glycerophospholipids into membrane microdomains accounts for the unique biophysical characteristics of this liquid-ordered (lo) state (Simons and Vaz, 2004, Hanzal-Bayer and Hancock, 2007). Certain proteins with a high affinity for lipid rafts are also selectively incorporated into lipid rafts. The molecular signal for lipid-raft affinity of peripheral and transmembrane proteins is based on covalent modifications with myristoyl and palmitoyl residues (Brown, 2006). Most importantly, it is well known that viral proteins are myristoylated and/or palmitoylated (Resh, 1999, Maurer-Stroh and Eisenhaber, 2004). For instance retrovirus Gag and Nef proteins are typically myristoylated (Hizi et al., 1989, Shoji et al., 1989, Yu and Felsted, 1992, Ono and Freed, 1999, Campbell et al., 2001) while Env glycoproteins are palmitoylated (Resh, 1999). Palmitoylation has also been observed in HA and M2 proteins of influenza virus (Melkonian et al., 1999, Tatulian and Tamm, 2000, Thaa et al., 2009). Such observations are in line with studies showing that “lipid rafts” are hijacked during HIV (Ono and Freed, 2001, Pickl et al., 2001, Nguyen et al., 2003) and influenza virus (Scheiffele et al., 1999, Takeda et al., 2003, Calistri et al., 2009) assembly and budding. Other examples of lipid-raft dependent viruses include Ebola and Marburg viruses (Bavari et al., 2002), measles virus (Manie et al., 2000, Vincent et al., 2000), Newcastle disease virus (Dolganiuc et al., 2003, Laliberte et al., 2006) and respiratory syncytial virus (Marty et al., 2004, Money et al., 2009).
The interpretation of some studies pertaining to raft-associated enveloped virus budding is inherently controversial. One common method of assigning lipid-raft association is through co-flotation of viral proteins with lipid-raft membranes and marker proteins in density gradients after cold detergent treatments (Briggs et al., 2003). This procedure is based on the idea that pre-existing lipid raft form insoluble detergent resistant membranes (DRM) when treated with low concentrations of a non-ionic detergent. However it is clear that the use of different detergents can alter membrane phase separation and lipid-raft composition (Pike et al., 2005). Hence, neither presence nor absence of proteins in DRM fraction is sufficient to assign lipid-raft association (Hanzal-Bayer and Hancock, 2007). A second common method is to disrupt lipid raft by cholesterol depletion from the plasma membrane using β-cyclodextrins (Briggs et al., 2003). However, cyclodextrin treatment may result in additional cholesterol-independent effects on membrane protein mobility that can negatively impact assembly and budding (Shvartsman et al., 2003). Nonetheless, there is equally compelling evidence for lipid-raft association from observations that (1) viral proteins are co-localized with raft markers by immunofluorescence microscopy (Briggs et al., 2003) and (2) purified virus particles are enriched in raft lipids including cholesterol, sphingomyelin and glycosphingolipids relative to bulk cellular lipid composition (Aloia et al., 1993, van Genderen et al., 1994, Scheiffele et al., 1999, Brugger et al., 2006, Chan et al., 2008).
The heterogeneity of cellular lipid rafts and its biological implications for enveloped virus assembly and budding is highly intriguing. For example, the depletion of cholesterol from the plasma membrane enhances influenza virus production (Barman and Nayak, 2007) as opposed to HIV production which is significantly inhibited (Ono and Freed, 2001, Pickl et al., 2001). Additionally, when influenza HA and HIV gag proteins are expressed in the same cells, they do not co-localize with each other on the plasma membrane (Khurana et al., 2007). Similarly, the Ebola virus glycoprotein (GP) and HIV envelope protein (Env) also do not co-localize with each other on the plasma membrane and that pseudotyped virus like particles (VLP) exclusively carried either only GP or Env despite their expression in the same producer cell (Leung et al., 2008). Putting these findings into a common context, it is clear that these enveloped viruses (and likely others) use different types of plasma membrane lipid rafts for virus production and that these microdomains are distinct with regard to their lipid and protein composition.
1.11. Phosphoinositides target virus proteins to the assembly site
While lipid rafts provide an attractive description of how viral membranes are formed, the precise molecular interaction between viral protein and the membrane is less evident. An early clue to this issue was provided by the observation that MA domain of retroviral Gag directs the polypeptide precursor to the plasma membrane via a bipartite motif consisting of an N-terminal myristic acid that is covalently attached (Bryant and Ratner, 1990) and a highly basic cluster of amino acid residues (Yuan et al., 1993). However, myristoylation modification alone cannot be solely responsible for Gag transport to the plasma membrane since not all myristoylated proteins are associated with the plasma membrane (Resh, 1999). Moreover, non-myristoylated Gag from retroviruses like Rous sarcoma virus (RSV) and EIAV, are still targeted to the plasma membrane for assembly and budding (Conte and Matthews, 1998). Hence, the additional signal for plasma membrane binding must reside within the basic cluster of positively charged amino acid residues.
The cytosolic surface of the plasma membrane carries an appreciable negative charge due to the large proportion of negatively charged phospholipids (Fig. 1B). The most abundant of these phospholipids is the monovalent acidic phosphatidylserine (PS), which represents 10–20% of plasma membrane lipids (Vance and Steenbergen, 2005). In addition to PS, other negatively charged phospholipids appear to be enriched in the inner leaflet of the plasma membrane including phosphatidic acid (PA), phosphatidylinositol (PI) and their polyphosphorylated derivatives (collectively termed as phosphoinositides) such as phosphatidylinositol-4,5-bisphosphate (PI(4,5)P2) and phosphatidylinositol-3,4,5-triphosphate (PI(3,4,5)P3). While PI(4,5)P2 and PI(3,4,5)P3 represent only a minor fraction of the plasma membrane lipids, they play multiple roles in diverse cellular functions (Rusten and Stenmark, 2006). Therefore, their concentration is tightly regulated enzymatically by a significant number of kinases and phosphatases (Downes et al., 2005) and spatially by reversible phosphoinositide binding proteins such as MARCKS (McLaughlin and Murray, 2005).
The complementary charge state of Gag MA and inner plasma membrane surface intuitively suggests an electrostatic mechanism by which Gag interacts with the membrane. This early hypothesis was supported by liposome pull-down assays showing that Gag can be enriched by liposomes containing PS (Zhou et al., 1994). A link between phosphoinositides and retrovirus assembly was provided by the observation that inositol polyphosphate molecules are able to promote cell free assembly of virus like particles from recombinant HIV-1 Gag molecules (Campbell et al., 2001). In cell based systems, the depletion of plasma membrane PI(4,5)P2 inhibits HIV assembly and leads to accumulation of Gag at late endosomes and multi vesicular bodies (Ono et al., 2004). The work of Saad et al. (2006) provided a structural framework for Gag and PI(4,5)P2 binding during HIV-1 assembly. Essentially, Gag binds specifically to PI(4,5)P2 through the hydrophobic cleft in Gag MA domain and the inositol headgroup and 2′ fatty acid chain of the lipid molecule. This interaction causes the MA myristic acid moiety to flip out from a sequestered to an exposed conformation thereby increasing Gag membrane partitioning.
An interesting feature of this myristic switch model is that it reconciles an apparent conflict with the lipid-raft model (Freed, 2006). As mentioned above, lipid rafts are concentrated in saturated glycerophospholipids. Because the predominant form of PI(4,5)P2 in cells contains a saturated 1′ fatty acid side (C18:0) chain and a poly-unsaturated 2′ side chain (C20:4), PI(4,5)P2 may equilibrate between raft and non-raft domains in the membrane. During Gag binding, the 2′ side chain is extruded from the lipid bilayer and packed into MA, with myristic acid taking its place in the bilayer. Hence, the PI(4,5)P2–Gag complex is now anchored by the fully saturated 1′ fatty acid and myristic acid and will exhibit increased affinity for lipid rafts.
Importantly, the function of PI(4,5)P2 dependent membrane targeting is not confined to HIV but other retroviruses as well (Murray et al., 2005, Chan et al., 2008). In addition, other families of enveloped viruses such as influenza virus (Ruigrok et al., 2000, Thaa et al., 2009), Ebola virus (Timmins et al., 2004), VSV (Gaudier et al., 2002) and RSV (Money et al., 2009) which expresses similar matrix proteins with surface exposed polybasic domains, may also use phosphoinositides or other anionic phospholipids in a similar mechanism for plasma membrane targeting.
Besides serving as a targeting signal for viral structural proteins, further roles for PI(4,5)P2 or other phosphoinositides isoforms remain a distinct possibility. PI(4,5)P2 is intimately involved in the inward and outward bending of plasma membrane. During endocytosis, BAR domain proteins bind to PI(4,5)P2 rich membranes to form inward invaginations (Itoh et al., 2005, Hess et al., 2007). Conversely, during the formation of filopodia, MIM and IRSp53, proteins which contain BAR-like domains, can lead to the formation of outward bending of PI(4,5)P2 rich membranes (Mattila et al., 2007). If PI(4,5)P2 lipids are highly enriched in viral envelopes, as is the case with retroviruses (Chan et al., 2008), it is possible that the PI(4,5)P2 interaction with cellular protein may contribute to the induction of membrane curvature during virus assembly and budding.
2. Extracellular virus
2.1. Lipid-bilayer functions as a protective shell for extracellular virus
Between leaving one host cell and infecting the next, an enveloped virus is exposed to an extremely hostile environment. The extracellular environment is full of degradative enzymes released from dead or leaky cells or deliberately secreted by immune cells as defense against infections. In addition, physical damage such as shearing by mechanical forces and chemical modification by ultraviolet light and extreme pH conditions are also possible. Thus, the most obvious and simplistic view of the lipid envelope is to provide a protective shell for the fragile nucleic acid an extracellular enveloped virus in order to ensure successful transmission. High levels of lipid ordering and membrane rigidity has been reported with HIV (Aloia et al., 1988, Aloia et al., 1993) and a range of other retroviruses as well, including equine infectious anemia virus (EIAV), bovine leukemia virus (BLV), murine leukemia virus (MLV) and avian myelolastosis virus (Slosberg and Montelaro, 1982). However, extracellular protection is perhaps most important for enveloped viruses that rely on air borne transmission such as the flu virus. More recent work showed that the flu virus envelope increases its equilibrium of lipid ordered to disordered phases at lower than physiological temperatures, which would lead to the increased survival of the virus at environmental temperatures (Polozov et al., 2008).
While enveloped viruses derive their lipid envelope from host membranes, they acquire a unique lipid composition that distinguishes them from the bulk lipid composition of host membranes (Fig. 1C). The lipid inventory of purified enveloped viruses including HIV (Aloia et al., 1988, Aloia et al., 1993, Brugger et al., 2006, Chan et al., 2008), HSV (van Genderen et al., 1994), RSV (Quigley et al., 1971, Quigley et al., 1972), VSV (McSharry and Wagner, 1971, van Meer and Simons, 1982) and Influenza Virus (Scheiffele et al., 1999, Blom et al., 2001) have been analyzed biochemically. The striking result from these analyses is that viral envelopes typically feature a high cholesterol/sphingomyelin to phospholipids ratio and also a propensity for highly saturated phospholipid lipid inventory, consistent with lipid microdomains. The structural basis for membrane rigidity and protection is thus three folds: (1) The sphingolipids interact laterally through van der Waals interactions and extensive hydrogen bonding between the sphingosine backbones and between the sugar head groups (Ramstedt and Slotte, 2006). (2) The majority of sphingolipids and phospholipids have saturated, and therefore unlinked, acyl chains that allow tighter packing of laterally associated lipids and a higher gel–liquid phase transition temperature (Ramstedt and Slotte, 2002). (3) The hydroxyl group of cholesterol hydrogen bonds with the ceramide group of sphingolipids, while its planar sterol ring interacts with the saturated acyl chain (Xu and London, 2000, Ramstedt and Slotte, 2006). As a proof of concept, it has been shown that the removal of virion-associated cholesterol using β-cyclodextrin permeabilized the viral membrane of HIV, SIV and influenza virus and the enveloped viruses became inactivated due to a loss in their protein core and genome integrity (Graham et al., 2003, Barman and Nayak, 2007, Ambrose et al., 2008).
2.2. Lipids that mediate virus budding and entry
An equally intuitive role of the virus envelope is to provide the medium by which the nascent retrovirus can form a protrusion at the assembly site before finally budding from the site through fission (Fig. 1B). This envelope composition must subsequently allow efficient fusion of the extracellular virus with the membrane of a new host cell. For both events to occur, both cell and virus associated membrane bilayers must generate membrane curvature through a coordinated sequence of events involving membrane proteins and lipids to provide the physical forces required to disrupt the original membrane bilayers (McMahon and Gallop, 2005, Zimmerberg and Kozlov, 2006). In many enveloped viruses, fission occurs through interaction of the late domain of their matrix protein with the endosomal sorting complex required for transport (ESCRT) machinery, which is a collection of approximately 20 host proteins that mediate cargo protein sorting into the endosomal pathway under normal cellular function (Calistri et al., 2009). During fusion, a part of this energy is provided by the interaction of the viral fusion machinery with its partner receptor, e.g. HIV Env with CD4, Influenza hemagglutinin with cell sialic acid containing cell surface receptors and Dengue E-glycoprotein with DC-SIGN. A further aspect of dynamic membrane energy is provided by the presence of particular lipids that are capable of producing curvature spontaneously or through interaction with protein partners as discussed above with phosphoinositides during budding.
The underlying basis of lipids producing spontaneous curvature depends very much on its individual molecular geometry. Lipids that have conical and inverted-conical shapes promote negative and positive spontaneous curvature respectively. Conical lipids are characterized by a relatively small head group compared to its fatty acyl portion and are represented by PA, cholesterol, ceramide, diacylglycerol (DG) and phosphatidylethanolamine (PE). Inverted-conical lipids are characterized by a large head group compared to its fatty acyl portion and are represented mainly by lysolipids, i.e. lipids with a single fatty acyl chain. On the other hand, PS and phosphatidylcholine (PC) have relatively low spontaneous curvature because their large head group compensates for the double fatty acyl chains, giving these lipids a cylindrical shape. In theory, membrane in a state of dynamic flux should exhibit local accumulations of curvature forming lipids and a concomitant decrease in cylindrical lipids in membrane regions with a highly bent structure. This would lead to an energetically favorable state that allows membrane rupture and mixing needed to complete fission or fusion activities.
One would expect lipids to play a major role in directing membrane activity in viruses during budding and entry. Earlier studies showed that virus envelopes of VSV (Fong et al., 1976, Fong and Brown, 1978), Rous sarcoma virus (Quigley et al., 1971), Newcastle disease virus (Quigley et al., 1971), Sendai virus (Barnes et al., 1987) and Influenza virus (Rothman et al., 1976, Bukrinskaya et al., 1987, Zhang et al., 2000, Blom et al., 2001) were enriched in overall PE content. Most importantly, recent lipidomics analysis of HIV particles found strong enrichment of plasmalogen PE (pPE) molecules (Brugger et al., 2006, Chan et al., 2008), a lipid class with strong fusogenic activity (Glaser and Gross, 1994), compared to total cellular levels. The loss of carbonyl oxygen and the presence of the vinyl ether double bond give pPE different physical properties when compared to their diacyl PE counterparts. For instance, pPE tends to form non-bilayer Hex II structures at or below 30 °C, while diacyl analogues form these structures at much higher temperatures (Glaser and Gross, 1994, Lohner, 1996). Energetically, this would mean easier formation of the hemi-fusion stalk-like state so that budding and entry become less challenging for enveloped viruses. Consistent with this idea, it has been shown in both class I (Gunther-Ausborn et al., 1995, Martin and Ruysschaert, 1995, Gunther-Ausborn and Stegmann, 1997, Chernomordik et al., 1998) and class II (Stiasny and Heinz, 2004) viruses, the presence of inverted-cone shaped lipids in the target membrane severely inhibit the fusion mechanism while cone shaped lipids such as PE enhance it.
3. Conclusion
Many of the enveloped viruses presented here are the cause of serious infectious diseases around the world. We have summarized reported evidence that these enveloped viruses are able to manipulate host cell lipid metabolism and membrane dynamics to support a wide range of replication strategies. A deeper understanding of these mechanisms may therefore contribute to the development of new therapeutics for the treatment of such diseases.
Acknowledgments
Research in the laboratory of M.R.W. is supported in part by the Singapore National Research Foundation under CRP Award No. 2007-04, the Biomedical Research Council of Singapore (R-183-000-211-305) and the National Medical Research Council (R-183-000-224-213) and the SystemsX.ch project LipidX. R.B.C. is a recipient of a scholarship from the National University of Singapore (NUS). L.T. is a recipient of a scholarship from the NUS Graduate School for Integrative Sciences and Engineering (NGS).
References
- Ablan S., Rawat S.S., Blumenthal R., Puri A. Entry of influenza virus into a glycosphingolipid-deficient mouse skin fibroblast cell line. Arch. Virol. 2001;146:2227–2238. doi: 10.1007/s007050170033. [DOI] [PubMed] [Google Scholar]
- Agnello V., Abel G., Elfahal M., Knight G.B., Zhang Q.X. Hepatitis C virus and other flaviviridae viruses enter cells via low density lipoprotein receptor. Proc. Natl. Acad. Sci. U.S.A. 1999;96:12766–12771. doi: 10.1073/pnas.96.22.12766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahola T., Lampio A., Auvinen P., Kaariainen L. Semliki Forest virus mRNA capping enzyme requires association with anionic membrane phospholipids for activity. EMBO J. 1999;18:3164–3172. doi: 10.1093/emboj/18.11.3164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albert M.L. Death-defying immunity: do apoptotic cells influence antigen processing and presentation? Nat. Rev. Immunol. 2004;4:223–231. doi: 10.1038/nri11308. [DOI] [PubMed] [Google Scholar]
- Alfsen A., Bomsel M. HIV-1 gp41 envelope residues 650–685 exposed on native virus act as a lectin to bind epithelial cell galactosyl ceramide. J. Biol. Chem. 2002;277:25649–25659. doi: 10.1074/jbc.M200554200. [DOI] [PubMed] [Google Scholar]
- Aloia R.C., Jensen F.C., Curtain C.C., Mobley P.W., Gordon L.M. Lipid composition and fluidity of the human immunodeficiency virus. Proc. Natl. Acad. Sci. U.S.A. 1988;85:900–904. doi: 10.1073/pnas.85.3.900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aloia R.C., Tian H., Jensen F.C. Lipid composition and fluidity of the human immunodeficiency virus envelope and host cell plasma membranes. Proc. Natl. Acad. Sci. U.S.A. 1993;90:5181–5185. doi: 10.1073/pnas.90.11.5181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ambrose Z., Compton L., Piatak M., Jr., Lu D., Alvord W.G., Lubomirski M.S., Hildreth J.E., Lifson J.D., Miller C.J., KewalRamani V.N. Incomplete protection against simian immunodeficiency virus vaginal transmission in rhesus macaques by a topical antiviral agent revealed by repeat challenges. J. Virol. 2008;82:6591–6599. doi: 10.1128/JVI.02730-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anastasia L., Holguera J., Bianchi A., D’Avila F., Papini N., Tringali C., Monti E., Villar E., Venerando B., Munoz-Barroso I., Tettamanti G. Over-expression of mammalian sialidase NEU3 reduces Newcastle disease virus entry and propagation in COS7 cells. Biochim. Biophys. Acta. 2008;1780:504–512. doi: 10.1016/j.bbagen.2007.11.011. [DOI] [PubMed] [Google Scholar]
- Andre P., Komurian-Pradel F., Deforges S., Perret M., Berland J.L., Sodoyer M., Pol S., Brechot C., Paranhos-Baccala G., Lotteau V. Characterization of low- and very-low-density hepatitis C virus RNA-containing particles. J. Virol. 2002;76:6919–6928. doi: 10.1128/JVI.76.14.6919-6928.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balasubramanian K., Schroit A.J. Aminophospholipid asymmetry: a matter of life and death. Annu. Rev. Physiol. 2003;65:701–734. doi: 10.1146/annurev.physiol.65.092101.142459. [DOI] [PubMed] [Google Scholar]
- Barman S., Nayak D.P. Lipid raft disruption by cholesterol depletion enhances influenza A virus budding from MDCK cells. J. Virol. 2007;81:12169–12178. doi: 10.1128/JVI.00835-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes J.A., Pehowich D.J., Allen T.M. Characterization of the phospholipid and fatty acid composition of Sendai virus. J. Lipid Res. 1987;28:130–137. [PubMed] [Google Scholar]
- Bavari S., Bosio C.M., Wiegand E., Ruthel G., Will A.B., Geisbert T.W., Hevey M., Schmaljohn C., Schmaljohn A., Aman M.J. Lipid raft microdomains: a gateway for compartmentalized trafficking of Ebola and Marburg viruses. J. Exp. Med. 2002;195:593–602. doi: 10.1084/jem.20011500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beer C., Buhr P., Hahn H., Laubner D., Wirth M. Gene expression analysis of murine cells producing amphotropic mouse leukaemia virus at a cultivation temperature of 32 and 37 degrees C. J. Gen. Virol. 2003;84:1677–1686. doi: 10.1099/vir.0.18871-0. [DOI] [PubMed] [Google Scholar]
- Bhattacharya J., Peters P.J., Clapham P.R. Human immunodeficiency virus type 1 envelope glycoproteins that lack cytoplasmic domain cysteines: impact on association with membrane lipid rafts and incorporation onto budding virus particles. J. Virol. 2004;78:5500–5506. doi: 10.1128/JVI.78.10.5500-5506.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blom T.S., Koivusalo M., Kuismanen E., Kostiainen R., Somerharju P., Ikonen E. Mass spectrometric analysis reveals an increase in plasma membrane polyunsaturated phospholipid species upon cellular cholesterol loading. Biochemistry. 2001;40:14635–14644. doi: 10.1021/bi0156714. [DOI] [PubMed] [Google Scholar]
- Brazzoli M., Bianchi A., Filippini S., Weiner A., Zhu Q., Pizza M., Crotta S. CD81 is a central regulator of cellular events required for hepatitis C virus infection of human hepatocytes. J. Virol. 2008;82:8316–8329. doi: 10.1128/JVI.00665-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Briggs J.A., Wilk T., Fuller S.D. Do lipid rafts mediate virus assembly and pseudotyping? J. Gen. Virol. 2003;84:757–768. doi: 10.1099/vir.0.18779-0. [DOI] [PubMed] [Google Scholar]
- Brown D.A. Lipid rafts, detergent-resistant membranes, and raft targeting signals. Physiology (Bethesda) 2006;21:430–439. doi: 10.1152/physiol.00032.2006. [DOI] [PubMed] [Google Scholar]
- Brugger B., Glass B., Haberkant P., Leibrecht I., Wieland F.T., Krausslich H.G. The HIV lipidome: a raft with an unusual composition. Proc. Natl. Acad. Sci. U.S.A. 2006;103:2641–2646. doi: 10.1073/pnas.0511136103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bryant M., Ratner L. Myristoylation-dependent replication and assembly of human immunodeficiency virus 1. Proc. Natl. Acad. Sci. U.S.A. 1990;87:523–527. doi: 10.1073/pnas.87.2.523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bukrinskaya A.G., Molotkovsky J.G., Vodovozova E.L., Manevich Y.M., Bergelson L.D. The molecular organization of the influenza virus surface. Studies using photoreactive and fluorescent labeled phospholipid probes. Biochim. Biophys. Acta. 1987;897:285–292. doi: 10.1016/0005-2736(87)90424-x. [DOI] [PubMed] [Google Scholar]
- Burlone M.E., Budkowska A. Hepatitis C virus cell entry: role of lipoproteins and cellular receptors. J. Gen. Virol. 2009;90:1055–1070. doi: 10.1099/vir.0.008300-0. [DOI] [PubMed] [Google Scholar]
- Calistri A., Salata C., Parolin C., Palu G. Role of multivesicular bodies and their components in the egress of enveloped RNA viruses. Rev. Med. Virol. 2009;19:31–45. doi: 10.1002/rmv.588. [DOI] [PubMed] [Google Scholar]
- Callahan M.K., Popernack P.M., Tsutsui S., Truong L., Schlegel R.A., Henderson A.J. Phosphatidylserine on HIV envelope is a cofactor for infection of monocytic cells. J. Immunol. 2003;170:4840–4845. doi: 10.4049/jimmunol.170.9.4840. [DOI] [PubMed] [Google Scholar]
- Campbell S., Fisher R.J., Towler E.M., Fox S., Issaq H.J., Wolfe T., Phillips L.R., Rein A. Modulation of HIV-like particle assembly in vitro by inositol phosphates. Proc. Natl. Acad. Sci. U.S.A. 2001;98:10875–10879. doi: 10.1073/pnas.191224698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan R., Uchil P.D., Jin J., Shui G., Ott D.E., Mothes W., Wenk M.R. Retroviruses human immunodeficiency virus and murine leukemia virus are enriched in phosphoinositides. J. Virol. 2008;82:11228–11238. doi: 10.1128/JVI.00981-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chernomordik L.V., Frolov V.A., Leikina E., Bronk P., Zimmerberg J. The pathway of membrane fusion catalyzed by influenza hemagglutinin: restriction of lipids, hemifusion, and lipidic fusion pore formation. J. Cell Biol. 1998;140:1369–1382. doi: 10.1083/jcb.140.6.1369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu V.C., Whittaker G.R. Influenza virus entry and infection require host cell N-linked glycoprotein. Proc. Natl. Acad. Sci. U.S.A. 2004;101:18153–18158. doi: 10.1073/pnas.0405172102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coil D.A., Miller A.D. Enhancement of enveloped virus entry by phosphatidylserine. J. Virol. 2005;79:11496–11500. doi: 10.1128/JVI.79.17.11496-11500.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coil D.A., Miller A.D. Phosphatidylserine treatment relieves the block to retrovirus infection of cells expressing glycosylated virus receptors. Retrovirology. 2005;2:49. doi: 10.1186/1742-4690-2-49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conte M.R., Matthews S. Retroviral matrix proteins: a structural perspective. Virology. 1998;246:191–198. doi: 10.1006/viro.1998.9206. [DOI] [PubMed] [Google Scholar]
- Dolganiuc V., McGinnes L., Luna E.J., Morrison T.G. Role of the cytoplasmic domain of the Newcastle disease virus fusion protein in association with lipid rafts. J. Virol. 2003;77:12968–12979. doi: 10.1128/JVI.77.24.12968-12979.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Downes C.P., Gray A., Lucocq J.M. Probing phosphoinositide functions in signaling and membrane trafficking. Trends Cell Biol. 2005;15:259–268. doi: 10.1016/j.tcb.2005.03.008. [DOI] [PubMed] [Google Scholar]
- El-Sadr W.M., Mullin C.M., Carr A., Gibert C., Rappoport C., Visnegarwala F., Grunfeld C., Raghavan S.S. Effects of HIV disease on lipid, glucose and insulin levels: results from a large antiretroviral-naive cohort. HIV Med. 2005;6:114–121. doi: 10.1111/j.1468-1293.2005.00273.x. [DOI] [PubMed] [Google Scholar]
- Escaut L., Monsuez J.J., Chironi G., Merad M., Teicher E., Smadja D., Simon A., Vittecoq D. Coronary artery disease in HIV infected patients. Intensive Care Med. 2003;29:969–973. doi: 10.1007/s00134-003-1740-0. [DOI] [PubMed] [Google Scholar]
- Fantini J., Cook D.G., Nathanson N., Spitalnik S.L., Gonzalez-Scarano F. Infection of colonic epithelial cell lines by type 1 human immunodeficiency virus is associated with cell surface expression of galactosylceramide, a potential alternative gp120 receptor. Proc. Natl. Acad. Sci. U.S.A. 1993;90:2700–2704. doi: 10.1073/pnas.90.7.2700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferreira L., Villar E., Munoz-Barroso I. Conformational changes of Newcastle disease virus envelope glycoproteins triggered by gangliosides. Eur. J. Biochem. 2004;271:581–588. doi: 10.1111/j.1432-1033.2003.03960.x. [DOI] [PubMed] [Google Scholar]
- Ferreira L., Villar E., Munoz-Barroso I. Gangliosides and N-glycoproteins function as Newcastle disease virus receptors. Int. J. Biochem. Cell Biol. 2004;36:2344–2356. doi: 10.1016/j.biocel.2004.05.011. [DOI] [PubMed] [Google Scholar]
- Fleming E.H., Kolokoltsov A.A., Davey R.A., Nichols J.E., Roberts N.J., Jr. Respiratory syncytial virus F envelope protein associates with lipid rafts without a requirement for other virus proteins. J. Virol. 2006;80:12160–12170. doi: 10.1128/JVI.00643-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fong B.S., Brown J.C. Asymmetric distribution of phosphatidylethanolamine fatty acyl chains in the membrane of vesicular stomatitis virus. Biochim. Biophys. Acta. 1978;510:230–241. doi: 10.1016/0005-2736(78)90023-8. [DOI] [PubMed] [Google Scholar]
- Fong B.S., Hunt R.C., Brown J.C. Asymmetric distribution of phosphatidylethanolamine in the membrane of vesicular stomatitis virus. J. Virol. 1976;20:658–663. doi: 10.1128/jvi.20.3.658-663.1976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freed E.O. HIV-1 Gag: flipped out for PI(4,5)P(2) Proc. Natl. Acad. Sci. U.S.A. 2006;103:11101–11102. doi: 10.1073/pnas.0604715103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gambaryan A.S., Tuzikov A.B., Pazynina G.V., Webster R.G., Matrosovich M.N., Bovin N.V. H5N1 chicken influenza viruses display a high binding affinity for Neu5Acalpha2-3Galbeta1-4(6-HSO3)GlcNAc-containing receptors. Virology. 2004;326:310–316. doi: 10.1016/j.virol.2004.06.002. [DOI] [PubMed] [Google Scholar]
- Gastaminza P., Cheng G., Wieland S., Zhong J., Liao W., Chisari F.V. Cellular determinants of hepatitis C virus assembly, maturation, degradation, and secretion. J. Virol. 2008;82:2120–2129. doi: 10.1128/JVI.02053-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gastaminza P., Kapadia S.B., Chisari F.V. Differential biophysical properties of infectious intracellular and secreted hepatitis C virus particles. J. Virol. 2006;80:11074–11081. doi: 10.1128/JVI.01150-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaudier M., Gaudin Y., Knossow M. Crystal structure of vesicular stomatitis virus matrix protein. EMBO J. 2002;21:2886–2892. doi: 10.1093/emboj/cdf284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giri M.S., Nebozhyn M., Showe L., Montaner L.J. Microarray data on gene modulation by HIV-1 in immune cells: 2000–2006. J. Leukoc. Biol. 2006;80:1031–1043. doi: 10.1189/jlb.0306157. [DOI] [PubMed] [Google Scholar]
- Glaser P.E., Gross R.W. Plasmenylethanolamine facilitates rapid membrane fusion: a stopped-flow kinetic investigation correlating the propensity of a major plasma membrane constituent to adopt an HII phase with its ability to promote membrane fusion. Biochemistry. 1994;33:5805–5812. doi: 10.1021/bi00185a019. [DOI] [PubMed] [Google Scholar]
- Graham D.R., Chertova E., Hilburn J.M., Arthur L.O., Hildreth J.E. Cholesterol depletion of human immunodeficiency virus type 1 and simian immunodeficiency virus with beta-cyclodextrin inactivates and permeabilizes the virions: evidence for virion-associated lipid rafts. J. Virol. 2003;77:8237–8248. doi: 10.1128/JVI.77.15.8237-8248.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griggi T., Bauer R., Garofalo T., Kukel S., Lenti L., Massetti A.P., Muller C., Sorice M., Pontieri G.M. Autoantibodies against ganglioside GM3 represent a portion of anti-lymphocyte antibodies in AIDS patients. Scand. J. Immunol. 1994;40:77–82. doi: 10.1111/j.1365-3083.1994.tb03436.x. [DOI] [PubMed] [Google Scholar]
- Gunther-Ausborn S., Praetor A., Stegmann T. Inhibition of influenza-induced membrane fusion by lysophosphatidylcholine. J. Biol. Chem. 1995;270:29279–29285. doi: 10.1074/jbc.270.49.29279. [DOI] [PubMed] [Google Scholar]
- Gunther-Ausborn S., Stegmann T. How lysophosphatidylcholine inhibits cell–cell fusion mediated by the envelope glycoprotein of human immunodeficiency virus. Virology. 1997;235:201–208. doi: 10.1006/viro.1997.8699. [DOI] [PubMed] [Google Scholar]
- Hammache D., Yahi N., Maresca M., Pieroni G., Fantini J. Human erythrocyte glycosphingolipids as alternative cofactors for human immunodeficiency virus type 1 (HIV-1) entry: evidence for CD4-induced interactions between HIV-1 gp120 and reconstituted membrane microdomains of glycosphingolipids (Gb3 and GM3) J. Virol. 1999;73:5244–5248. doi: 10.1128/jvi.73.6.5244-5248.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansson G.C., Karlsson K.A., Larson G., Stromberg N., Thurin J., Orvell C., Norrby E. A novel approach to the study of glycolipid receptors for viruses. Binding of Sendai virus to thin-layer chromatograms. FEBS Lett. 1984;170:15–18. doi: 10.1016/0014-5793(84)81359-9. [DOI] [PubMed] [Google Scholar]
- Hanzal-Bayer M.F., Hancock J.F. Lipid rafts and membrane traffic. FEBS Lett. 2007;581:2098–2104. doi: 10.1016/j.febslet.2007.03.019. [DOI] [PubMed] [Google Scholar]
- Harouse J.M., Kunsch C., Hartle H.T., Laughlin M.A., Hoxie J.A., Wigdahl B., Gonzalez-Scarano F. CD4-independent infection of human neural cells by human immunodeficiency virus type 1. J. Virol. 1989;63:2527–2533. doi: 10.1128/jvi.63.6.2527-2533.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henson P.M., Bratton D.L., Fadok V.A. The phosphatidylserine receptor: a crucial molecular switch? Nat. Rev. Mol. Cell. Biol. 2001;2:627–633. doi: 10.1038/35085094. [DOI] [PubMed] [Google Scholar]
- Hertogs K., Leenders W.P., Depla E., De Bruin W.C., Meheus L., Raymackers J., Moshage H., Yap S.H. Endonexin II, present on human liver plasma membranes, is a specific binding protein of small hepatitis B virus (HBV) envelope protein. Virology. 1993;197:549–557. doi: 10.1006/viro.1993.1628. [DOI] [PubMed] [Google Scholar]
- Hess S.T., Gould T.J., Gudheti M.V., Maas S.A., Mills K.D., Zimmerberg J. Dynamic clustered distribution of hemagglutinin resolved at 40 nm in living cell membranes discriminates between raft theories. Proc. Natl. Acad. Sci. U.S.A. 2007;104:17370–17375. doi: 10.1073/pnas.0708066104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hidari K.I., Shimada S., Suzuki Y., Suzuki T. Binding kinetics of influenza viruses to sialic acid-containing carbohydrates. Glycoconj. J. 2007;24:583–590. doi: 10.1007/s10719-007-9055-y. [DOI] [PubMed] [Google Scholar]
- Hizi A., Henderson L.E., Copeland T.D., Sowder R.C., Krutzsch H.C., Oroszlan S. Analysis of gag proteins from mouse mammary tumor virus. J. Virol. 1989;63:2543–2549. doi: 10.1128/jvi.63.6.2543-2549.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hruby D.E., Franke C.A. Viral acylproteins: greasing the wheels of assembly. Trends Microbiol. 1993;1:20–25. doi: 10.1016/0966-842x(93)90020-r. [DOI] [PubMed] [Google Scholar]
- Huang R.T. Sphingolipids of influenza viruses. Biochim. Biophys. Acta. 1976;424:90–97. doi: 10.1016/0005-2760(76)90053-9. [DOI] [PubMed] [Google Scholar]
- Huang R.T., Lichtenberg B., Rick O. Involvement of annexin V in the entry of influenza viruses and role of phospholipids in infection. FEBS Lett. 1996;392:59–62. doi: 10.1016/0014-5793(96)00783-1. [DOI] [PubMed] [Google Scholar]
- Hug P., Lin H.M., Korte T., Xiao X., Dimitrov D.S., Wang J.M., Puri A., Blumenthal R. Glycosphingolipids promote entry of a broad range of human immunodeficiency virus type 1 isolates into cell lines expressing CD4, CXCR4, and/or CCR5. J. Virol. 2000;74:6377–6385. doi: 10.1128/jvi.74.14.6377-6385.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Itoh T., Erdmann K.S., Roux A., Habermann B., Werner H., De Camilli P. Dynamin and the actin cytoskeleton cooperatively regulate plasma membrane invagination by BAR and F-BAR proteins. Dev. Cell. 2005;9:791–804. doi: 10.1016/j.devcel.2005.11.005. [DOI] [PubMed] [Google Scholar]
- Kaneda M., Takeuchi K., Inoue K., Umeda M. Localization of the phosphatidylserine-binding site of glyceraldehyde-3-phosphate dehydrogenase responsible for membrane fusion. J. Biochem. 1997;122:1233–1240. doi: 10.1093/oxfordjournals.jbchem.a021886. [DOI] [PubMed] [Google Scholar]
- Kapadia S.B., Barth H., Baumert T., McKeating J.A., Chisari F.V. Initiation of hepatitis C virus infection is dependent on cholesterol and cooperativity between CD81 and scavenger receptor B type I. J. Virol. 2007;81:374–383. doi: 10.1128/JVI.01134-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapadia S.B., Chisari F.V. Hepatitis C virus RNA replication is regulated by host geranylgeranylation and fatty acids. Proc. Natl. Acad. Sci. U.S.A. 2005;102:2561–2566. doi: 10.1073/pnas.0409834102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasson P.M., Pande V.S. Structural basis for influence of viral glycans on ligand binding by influenza hemagglutinin. Biophys. J. 2008;95:L48–50. doi: 10.1529/biophysj.108.141507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawada J., Kimura H., Kamachi Y., Nishikawa K., Taniguchi M., Nagaoka K., Kurahashi H., Kojima S., Morishima T. Analysis of gene-expression profiles by oligonucleotide microarray in children with influenza. J. Gen. Virol. 2006;87:1677–1683. doi: 10.1099/vir.0.81670-0. [DOI] [PubMed] [Google Scholar]
- Khurana S., Krementsov D.N., de Parseval A., Elder J.H., Foti M., Thali M. Human immunodeficiency virus type 1 and influenza virus exit via different membrane microdomains. J. Virol. 2007;81:12630–12640. doi: 10.1128/JVI.01255-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kogure T., Suzuki T., Takahashi T., Miyamoto D., Hidari K.I., Guo C.T., Ito T., Kawaoka Y., Suzuki Y. Human trachea primary epithelial cells express both sialyl(alpha2-3)Gal receptor for human parainfluenza virus type 1 and avian influenza viruses, and sialyl(alpha2-6)Gal receptor for human influenza viruses. Glycoconj. J. 2006;23:101–106. doi: 10.1007/s10719-006-5442-z. [DOI] [PubMed] [Google Scholar]
- Laliberte J.P., McGinnes L.W., Peeples M.E., Morrison T.G. Integrity of membrane lipid rafts is necessary for the ordered assembly and release of infectious Newcastle disease virus particles. J. Virol. 2006;80:10652–10662. doi: 10.1128/JVI.01183-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leskawa K.C., Hogan E.L., Dasgupta S., Chien J.L., Erwin R.E., Brostoff S.W. Lipid content of swine influenza and other vaccines. Lipids. 1986;21:342–346. doi: 10.1007/BF02535698. [DOI] [PubMed] [Google Scholar]
- Leung K., Kim J.O., Ganesh L., Kabat J., Schwartz O., Nabel G.J. HIV-1 assembly: viral glycoproteins segregate quantally to lipid rafts that associate individually with HIV-1 capsids and virions. Cell Host Microbe. 2008;3:285–292. doi: 10.1016/j.chom.2008.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lohner K. Is the high propensity of ethanolamine plasmalogens to form non-lamellar lipid structures manifested in the properties of biomembranes? Chem. Phys. Lipids. 1996;81:167–184. doi: 10.1016/0009-3084(96)02580-7. [DOI] [PubMed] [Google Scholar]
- Ma G., Greenwell-Wild T., Lei K., Jin W., Swisher J., Hardegen N., Wild C.T., Wahl S.M. Secretory leukocyte protease inhibitor binds to annexin II, a cofactor for macrophage HIV-1 infection. J. Exp. Med. 2004;200:1337–1346. doi: 10.1084/jem.20041115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Machesky N.J., Zhang G., Raghavan B., Zimmerman P., Kelly S.L., Merrill A.H., Jr., Waldman W.J., Van Brocklyn J.R., Trgovcich J. Human cytomegalovirus regulates bioactive sphingolipids. J. Biol. Chem. 2008;283:26148–26160. doi: 10.1074/jbc.M710181200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magerus-Chatinet A., Yu H., Garcia S., Ducloux E., Terris B., Bomsel M. Galactosyl ceramide expressed on dendritic cells can mediate HIV-1 transfer from monocyte derived dendritic cells to autologous T cells. Virology. 2007;362:67–74. doi: 10.1016/j.virol.2006.11.035. [DOI] [PubMed] [Google Scholar]
- Malhotra R., Ward M., Bright H., Priest R., Foster M.R., Hurle M., Blair E., Bird M. Isolation and characterisation of potential respiratory syncytial virus receptor(s) on epithelial cells. Microbes Infect. 2003;5:123–133. doi: 10.1016/s1286-4579(02)00079-5. [DOI] [PubMed] [Google Scholar]
- Manie S.N., de Breyne S., Vincent S., Gerlier D. Measles virus structural components are enriched into lipid raft microdomains: a potential cellular location for virus assembly. J. Virol. 2000;74:305–311. doi: 10.1128/jvi.74.1.305-311.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mansfield K.G., Carville A., Wachtman L., Goldin B.R., Yearley J., Li W., Woods M., Gualtieri L., Shannon R., Wanke C. A diet high in saturated fat and cholesterol accelerates simian immunodeficiency virus disease progression. J. Infect. Dis. 2007;196:1202–1210. doi: 10.1086/521680. [DOI] [PubMed] [Google Scholar]
- Markwell M.A., Fredman P., Svennerholm L. Receptor ganglioside content of three hosts for Sendai virus. MDBK, HeLa, and MDCK cells. Biochim. Biophys. Acta. 1984;775:7–16. doi: 10.1016/0005-2736(84)90228-1. [DOI] [PubMed] [Google Scholar]
- Markwell M.A., Svennerholm L., Paulson J.C. Specific gangliosides function as host cell receptors for Sendai virus. Proc. Natl. Acad. Sci. U.S.A. 1981;78:5406–5410. doi: 10.1073/pnas.78.9.5406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin I., Ruysschaert J.M. Lysophosphatidylcholine inhibits vesicles fusion induced by the NH2-terminal extremity of SIV/HIV fusogenic proteins. Biochim. Biophys. Acta. 1995;1240:95–100. doi: 10.1016/0005-2736(95)00171-4. [DOI] [PubMed] [Google Scholar]
- Marty A., Meanger J., Mills J., Shields B., Ghildyal R. Association of matrix protein of respiratory syncytial virus with the host cell membrane of infected cells. Arch. Virol. 2004;149:199–210. doi: 10.1007/s00705-003-0183-9. [DOI] [PubMed] [Google Scholar]
- Matrosovich M., Suzuki T., Hirabayashi Y., Garten W., Webster R.G., Klenk H.D. Gangliosides are not essential for influenza virus infection. Glycoconj. J. 2006;23:107–113. doi: 10.1007/s10719-006-5443-y. [DOI] [PubMed] [Google Scholar]
- Mattila P.K., Pykalainen A., Saarikangas J., Paavilainen V.O., Vihinen H., Jokitalo E., Lappalainen P. Missing-in-metastasis and IRSp53 deform PI(4,5)P2-rich membranes by an inverse BAR domain-like mechanism. J. Cell Biol. 2007;176:953–964. doi: 10.1083/jcb.200609176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maurer-Stroh S., Eisenhaber F. Myristoylation of viral and bacterial proteins. Trends Microbiol. 2004;12:178–185. doi: 10.1016/j.tim.2004.02.006. [DOI] [PubMed] [Google Scholar]
- McLaughlin S., Murray D. Plasma membrane phosphoinositide organization by protein electrostatics. Nature. 2005;438:605–611. doi: 10.1038/nature04398. [DOI] [PubMed] [Google Scholar]
- McMahon H.T., Gallop J.L. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature. 2005;438:590–596. doi: 10.1038/nature04396. [DOI] [PubMed] [Google Scholar]
- McSharry J.J., Wagner R.R. Lipid composition of purified vesicular stomatitis viruses. J. Virol. 1971;7:59–70. doi: 10.1128/jvi.7.1.59-70.1971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melkonian K.A., Ostermeyer A.G., Chen J.Z., Roth M.G., Brown D.A. Role of lipid modifications in targeting proteins to detergent-resistant membrane rafts. Many raft proteins are acylated, while few are prenylated. J. Biol. Chem. 1999;274:3910–3917. doi: 10.1074/jbc.274.6.3910. [DOI] [PubMed] [Google Scholar]
- Mercer J., Helenius A. Vaccinia virus uses macropinocytosis and apoptotic mimicry to enter host cells. Science. 2008;320:531–535. doi: 10.1126/science.1155164. [DOI] [PubMed] [Google Scholar]
- Miller S., Krijnse-Locker J. Modification of intracellular membrane structures for virus replication. Nat. Rev. Microbiol. 2008;6:363–374. doi: 10.1038/nrmicro1890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyanari Y., Atsuzawa K., Usuda N., Watashi K., Hishiki T., Zayas M., Bartenschlager R., Wakita T., Hijikata M., Shimotohno K. The lipid droplet is an important organelle for hepatitis C virus production. Nat. Cell Biol. 2007;9:1089–1097. doi: 10.1038/ncb1631. [DOI] [PubMed] [Google Scholar]
- Molina S., Castet V., Fournier-Wirth C., Pichard-Garcia L., Avner R., Harats D., Roitelman J., Barbaras R., Graber P., Ghersa P., Smolarsky M., Funaro A., Malavasi F., Larrey D., Coste J., Fabre J.M., Sa-Cunha A., Maurel P. The low-density lipoprotein receptor plays a role in the infection of primary human hepatocytes by hepatitis C virus. J. Hepatol. 2007;46:411–419. doi: 10.1016/j.jhep.2006.09.024. [DOI] [PubMed] [Google Scholar]
- Money V.A., McPhee H.K., Mosely J.A., Sanderson J.M., Yeo R.P. Surface features of a Mononegavirales matrix protein indicate sites of membrane interaction. Proc. Natl. Acad. Sci. U.S.A. 2009;106:4441–4446. doi: 10.1073/pnas.0805740106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mujawar Z., Rose H., Morrow M.P., Pushkarsky T., Dubrovsky L., Mukhamedova N., Fu Y., Dart A., Orenstein J.M., Bobryshev Y.V., Bukrinsky M., Sviridov D. Human immunodeficiency virus impairs reverse cholesterol transport from macrophages. PLoS Biol. 2006;4:e365. doi: 10.1371/journal.pbio.0040365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munger J., Bennett B.D., Parikh A., Feng X.J., McArdle J., Rabitz H.A., Shenk T., Rabinowitz J.D. Systems-level metabolic flux profiling identifies fatty acid synthesis as a target for antiviral therapy. Nat. Biotechnol. 2008;26:1179–1186. doi: 10.1038/nbt.1500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murray P.S., Li Z., Wang J., Tang C.L., Honig B., Murray D. Retroviral matrix domains share electrostatic homology: models for membrane binding function throughout the viral life cycle. Structure. 2005;13:1521–1531. doi: 10.1016/j.str.2005.07.010. [DOI] [PubMed] [Google Scholar]
- Muthing J., Unland F. A comparative assessment of TLC overlay technique and microwell adsorption assay in the examination of influenza A and Sendai virus specificities towards oligosaccharides and sialic acid linkages of gangliosides. Glycoconj. J. 1994;11:486–492. doi: 10.1007/BF00731285. [DOI] [PubMed] [Google Scholar]
- Nehete P.N., Vela E.M., Hossain M.M., Sarkar A.K., Yahi N., Fantini J., Sastry K.J. A post-CD4-binding step involving interaction of the V3 region of viral gp120 with host cell surface glycosphingolipids is common to entry and infection by diverse HIV-1 strains. Antiviral Res. 2002;56:233–251. doi: 10.1016/s0166-3542(02)00130-4. [DOI] [PubMed] [Google Scholar]
- Nguyen D.G., Booth A., Gould S.J., Hildreth J.E. Evidence that HIV budding in primary macrophages occurs through the exosome release pathway. J. Biol. Chem. 2003;278:52347–52354. doi: 10.1074/jbc.M309009200. [DOI] [PubMed] [Google Scholar]
- Nguyen D.H., Giri B., Collins G., Taub D.D. Dynamic reorganization of chemokine receptors, cholesterol, lipid rafts, and adhesion molecules to sites of CD4 engagement. Exp. Cell Res. 2005;304:559–569. doi: 10.1016/j.yexcr.2004.11.022. [DOI] [PubMed] [Google Scholar]
- Nguyen D.H., Taub D. CXCR4 function requires membrane cholesterol: implications for HIV infection. J. Immunol. 2002;168:4121–4126. doi: 10.4049/jimmunol.168.8.4121. [DOI] [PubMed] [Google Scholar]
- Ono A., Ablan S.D., Lockett S.J., Nagashima K., Freed E.O. Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc. Natl. Acad. Sci. U.S.A. 2004;101:14889–14894. doi: 10.1073/pnas.0405596101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ono A., Freed E.O. Binding of human immunodeficiency virus type 1 Gag to membrane: role of the matrix amino terminus. J. Virol. 1999;73:4136–4144. doi: 10.1128/jvi.73.5.4136-4144.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ono A., Freed E.O. Plasma membrane rafts play a critical role in HIV-1 assembly and release. Proc. Natl. Acad. Sci. U.S.A. 2001;98:13925–13930. doi: 10.1073/pnas.241320298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pickl W.F., Pimentel-Muinos F.X., Seed B. Lipid rafts and pseudotyping. J. Virol. 2001;75:7175–7183. doi: 10.1128/JVI.75.15.7175-7183.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pike L.J., Han X., Gross R.W. Epidermal growth factor receptors are localized to lipid rafts that contain a balance of inner and outer leaflet lipids: a shotgun lipidomics study. J. Biol. Chem. 2005;280:26796–26804. doi: 10.1074/jbc.M503805200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polozov I.V., Bezrukov L., Gawrisch K., Zimmerberg J. Progressive ordering with decreasing temperature of the phospholipids of influenza virus. Nat. Chem. Biol. 2008;4:248–255. doi: 10.1038/nchembio.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popik W., Alce T.M., Au W.C. Human immunodeficiency virus type 1 uses lipid raft-colocalized CD4 and chemokine receptors for productive entry into CD4(+) T cells. J. Virol. 2002;76:4709–4722. doi: 10.1128/JVI.76.10.4709-4722.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prince A.M., Huima-Byron T., Parker T.S., Levine D.M. Visualization of hepatitis C virions and putative defective interfering particles isolated from low-density lipoproteins. J. Viral Hepat. 1996;3:11–17. doi: 10.1111/j.1365-2893.1996.tb00075.x. [DOI] [PubMed] [Google Scholar]
- Quigley J.P., Rifkin D.B., Reich E. Phospholipid composition of Rous sarcoma virus, host cell membranes and other enveloped RNA viruses. Virology. 1971;46:106–116. doi: 10.1016/0042-6822(71)90010-9. [DOI] [PubMed] [Google Scholar]
- Quigley J.P., Rifkin D.B., Reich E. Lipid studies of Rous sarcoma virus and host cell membranes. Virology. 1972;50:550–557. doi: 10.1016/0042-6822(72)90406-0. [DOI] [PubMed] [Google Scholar]
- Ramstedt B., Slotte J.P. Membrane properties of sphingomyelins. FEBS Lett. 2002;531:33–37. doi: 10.1016/s0014-5793(02)03406-3. [DOI] [PubMed] [Google Scholar]
- Ramstedt B., Slotte J.P. Sphingolipids and the formation of sterol-enriched ordered membrane domains. Biochim. Biophys. Acta. 2006;1758:1945–1956. doi: 10.1016/j.bbamem.2006.05.020. [DOI] [PubMed] [Google Scholar]
- Rawat S.S., Gallo S.A., Eaton J., Martin T.D., Ablan S., KewalRamani V.N., Wang J.M., Blumenthal R., Puri A. Elevated expression of GM3 in receptor-bearing targets confers resistance to human immunodeficiency virus type 1 fusion. J. Virol. 2004;78:7360–7368. doi: 10.1128/JVI.78.14.7360-7368.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rawat S.S., Viard M., Gallo S.A., Blumenthal R., Puri A. Sphingolipids, cholesterol, and HIV-1: a paradigm in viral fusion. Glycoconj. J. 2006;23:189–197. doi: 10.1007/s10719-006-7924-4. [DOI] [PubMed] [Google Scholar]
- Raynor C.M., Wright J.F., Waisman D.M., Pryzdial E.L. Annexin II enhances cytomegalovirus binding and fusion to phospholipid membranes. Biochemistry. 1999;38:5089–5095. doi: 10.1021/bi982095b. [DOI] [PubMed] [Google Scholar]
- Resh M.D. Fatty acylation of proteins: new insights into membrane targeting of myristoylated and palmitoylated proteins. Biochim. Biophys. Acta. 1999;1451:1–16. doi: 10.1016/s0167-4889(99)00075-0. [DOI] [PubMed] [Google Scholar]
- Rothman J.E., Tsai D.K., Dawidowicz E.A., Lenard J. Transbilayer phospholipid asymmetry and its maintenance in the membrane of influenza virus. Biochemistry. 1976;15:2361–2370. doi: 10.1021/bi00656a018. [DOI] [PubMed] [Google Scholar]
- Ruigrok R.W., Barge A., Durrer P., Brunner J., Ma K., Whittaker G.R. Membrane interaction of influenza virus M1 protein. Virology. 2000;267:289–298. doi: 10.1006/viro.1999.0134. [DOI] [PubMed] [Google Scholar]
- Rusten T.E., Stenmark H. Analyzing phosphoinositides and their interacting proteins. Nat. Methods. 2006;3:251–258. doi: 10.1038/nmeth867. [DOI] [PubMed] [Google Scholar]
- Saad J.S., Miller J., Tai J., Kim A., Ghanam R.H., Summers M.F. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc. Natl. Acad. Sci. U.S.A. 2006;103:11364–11369. doi: 10.1073/pnas.0602818103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheiffele P., Rietveld A., Wilk T., Simons K. Influenza viruses select ordered lipid domains during budding from the plasma membrane. J. Biol. Chem. 1999;274:2038–2044. doi: 10.1074/jbc.274.4.2038. [DOI] [PubMed] [Google Scholar]
- Seddiki N., Ben Younes-Chennoufi A., Benjouad A., Saffar L., Baumann N., Gluckman J.C., Gattegno L. Membrane glycolipids and human immunodeficiency virus infection of primary macrophages. AIDS Res. Hum. Retroviruses. 1996;12:695–703. doi: 10.1089/aid.1996.12.695. [DOI] [PubMed] [Google Scholar]
- Shoji S., Tashiro A., Furuishi K., Takenaka O., Kida Y., Horiuchi S., Funakoshi T., Kubota Y. Antibodies to an NH2-terminal myristoyl glycine moiety can detect NH2-terminal myristoylated proteins in the retrovirus-infected cells. Biochem. Biophys. Res. Commun. 1989;162:724–732. doi: 10.1016/0006-291x(89)92370-x. [DOI] [PubMed] [Google Scholar]
- Shvartsman D.E., Kotler M., Tall R.D., Roth M.G., Henis Y.I. Differently anchored influenza hemagglutinin mutants display distinct interaction dynamics with mutual rafts. J. Cell Biol. 2003;163:879–888. doi: 10.1083/jcb.200308142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silvestris F., Frassanito M.A., Cafforio P., Potenza D., Di Loreto M., Tucci M., Grizzuti M.A., Nico B., Dammacco F. Antiphosphatidylserine antibodies in human immunodeficiency virus-1 patients with evidence of T-cell apoptosis and mediate antibody-dependent cellular cytotoxicity. Blood. 1996;87:5185–5195. [PubMed] [Google Scholar]
- Simons K., Vaz W.L. Model systems, lipid rafts, and cell membranes. Annu. Rev. Biophys. Biomol. Struct. 2004;33:269–295. doi: 10.1146/annurev.biophys.32.110601.141803. [DOI] [PubMed] [Google Scholar]
- Slosberg B.N., Montelaro R.C. A comparison of the mobilities and thermal transitions of retrovirus lipid envelopes and host cell plasma membranes by electron spin resonance spectroscopy. Biochim. Biophys. Acta. 1982;689:393–402. doi: 10.1016/0005-2736(82)90274-7. [DOI] [PubMed] [Google Scholar]
- Sorice M., Garofalo T., Misasi R., Longo A., Mattei V., Sale P., Dolo V., Gradini R., Pavan A. Evidence for cell surface association between CXCR4 and ganglioside GM3 after gp120 binding in SupT1 lymphoblastoid cells. FEBS Lett. 2001;506:55–60. doi: 10.1016/s0014-5793(01)02830-7. [DOI] [PubMed] [Google Scholar]
- Sorice M., Garofalo T., Misasi R., Longo A., Mikulak J., Dolo V., Pontieri G.M., Pavan A. Association between GM3 and CD4-Ick complex in human peripheral blood lymphocytes. Glycoconj. J. 2000;17:247–252. doi: 10.1023/a:1026501609699. [DOI] [PubMed] [Google Scholar]
- Stapleford K.A., Rapaport D., Miller D.J. Mitochondrion-enriched anionic phospholipids facilitate flock house virus RNA polymerase membrane association. J. Virol. 2009;83:4498–4507. doi: 10.1128/JVI.00040-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiasny K., Heinz F.X. Effect of membrane curvature-modifying lipids on membrane fusion by tick-borne encephalitis virus. J. Virol. 2004;78:8536–8542. doi: 10.1128/JVI.78.16.8536-8542.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su A.I., Pezacki J.P., Wodicka L., Brideau A.D., Supekova L., Thimme R., Wieland S., Bukh J., Purcell R.H., Schultz P.G., Chisari F.V. Genomic analysis of the host response to hepatitis C virus infection. Proc. Natl. Acad. Sci. U.S.A. 2002;99:15669–15674. doi: 10.1073/pnas.202608199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takeda M., Leser G.P., Russell C.J., Lamb R.A. Influenza virus hemagglutinin concentrates in lipid raft microdomains for efficient viral fusion. Proc. Natl. Acad. Sci. U.S.A. 2003;100:14610–14617. doi: 10.1073/pnas.2235620100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tatulian S.A., Tamm L.K. Secondary structure, orientation, oligomerization, and lipid interactions of the transmembrane domain of influenza hemagglutinin. Biochemistry. 2000;39:496–507. doi: 10.1021/bi991594p. [DOI] [PubMed] [Google Scholar]
- Teissier E., Pecheur E.I. Lipids as modulators of membrane fusion mediated by viral fusion proteins. Eur. Biophys. J. 2007;36:887–899. doi: 10.1007/s00249-007-0201-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tettamanti G. Ganglioside/glycosphingolipid turnover: new concepts. Glycoconj. J. 2004;20:301–317. doi: 10.1023/B:GLYC.0000033627.02765.cc. [DOI] [PubMed] [Google Scholar]
- Thaa B., Herrmann A., Veit M. The polybasic region is not essential for membrane binding of the matrix protein M1 of influenza virus. Virology. 2009;383:150–155. doi: 10.1016/j.virol.2008.10.001. [DOI] [PubMed] [Google Scholar]
- Thomssen R., Bonk S., Propfe C., Heermann K.H., Kochel H.G., Uy A. Association of hepatitis C virus in human sera with beta-lipoprotein. Med. Microbiol. Immunol. 1992;181:293–300. doi: 10.1007/BF00198849. [DOI] [PubMed] [Google Scholar]
- Timmins J., Ruigrok R.W., Weissenhorn W. Structural studies on the Ebola virus matrix protein VP40 indicate that matrix proteins of enveloped RNA viruses are analogues but not homologues. FEMS Microbiol. Lett. 2004;233:179–186. doi: 10.1111/j.1574-6968.2004.tb09480.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Genderen I.L., Brandimarti R., Torrisi M.R., Campadelli G., van Meer G. The phospholipid composition of extracellular herpes simplex virions differs from that of host cell nuclei. Virology. 1994;200:831–836. doi: 10.1006/viro.1994.1252. [DOI] [PubMed] [Google Scholar]
- van Meer G., Simons K. Viruses budding from either the apical or the basolateral plasma membrane domain of MDCK cells have unique phospholipid compositions. EMBO J. 1982;1:847–852. doi: 10.1002/j.1460-2075.1982.tb01258.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vance J.E., Steenbergen R. Metabolism and functions of phosphatidylserine. Prog. Lipid Res. 2005;44:207–234. doi: 10.1016/j.plipres.2005.05.001. [DOI] [PubMed] [Google Scholar]
- Viard M., Parolini I., Rawat S.S., Fecchi K., Sargiacomo M., Puri A., Blumenthal R. The role of glycosphingolipids in HIV signaling, entry and pathogenesis. Glycoconj. J. 2004;20:213–222. doi: 10.1023/B:GLYC.0000024253.48791.d9. [DOI] [PubMed] [Google Scholar]
- Villar E., Barroso I.M. Role of sialic acid-containing molecules in paramyxovirus entry into the host cell: a minireview. Glycoconj. J. 2006;23:5–17. doi: 10.1007/s10719-006-5433-0. [DOI] [PubMed] [Google Scholar]
- Vincent S., Gerlier D., Manie S.N. Measles virus assembly within membrane rafts. J. Virol. 2000;74:9911–9915. doi: 10.1128/jvi.74.21.9911-9915.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu X., Okada N., Momota H., Irie R.F., Okada H. Complement-mediated anti-HIV-1 effect induced by human IgM monoclonal antibody against ganglioside GM2. J. Immunol. 1999;162:533–539. [PubMed] [Google Scholar]
- Xu X., London E. The effect of sterol structure on membrane lipid domains reveals how cholesterol can induce lipid domain formation. Biochemistry. 2000;39:843–849. doi: 10.1021/bi992543v. [DOI] [PubMed] [Google Scholar]
- Yahi N., Spitalnik S.L., Stefano K.A., De Micco P., Gonzalez-Scarano F., Fantini J. Interferon-gamma decreases cell surface expression of galactosyl ceramide, the receptor for HIV-1 GP120 on human colonic epithelial cells. Virology. 1994;204:550–557. doi: 10.1006/viro.1994.1568. [DOI] [PubMed] [Google Scholar]
- Yang W., Hood B.L., Chadwick S.L., Liu S., Watkins S.C., Luo G., Conrads T.P., Wang T. Fatty acid synthase is up-regulated during hepatitis C virus infection and regulates hepatitis C virus entry and production. Hepatology. 2008;48:1396–1403. doi: 10.1002/hep.22508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu G., Felsted R.L. Effect of myristoylation on p27 nef subcellular distribution and suppression of HIV-LTR transcription. Virology. 1992;187:46–55. doi: 10.1016/0042-6822(92)90293-x. [DOI] [PubMed] [Google Scholar]
- Yu H., Alfsen A., Tudor D., Bomsel M. The binding of HIV-1 gp41 membrane proximal domain to its mucosal receptor, galactosyl ceramide, is structure-dependent. Cell Calcium. 2008;43:73–82. doi: 10.1016/j.ceca.2007.04.011. [DOI] [PubMed] [Google Scholar]
- Yuan X., Yu X., Lee T.H., Essex M. Mutations in the N-terminal region of human immunodeficiency virus type 1 matrix protein block intracellular transport of the Gag precursor. J. Virol. 1993;67:6387–6394. doi: 10.1128/jvi.67.11.6387-6394.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J., Pekosz A., Lamb R.A. Influenza virus assembly and lipid raft microdomains: a role for the cytoplasmic tails of the spike glycoproteins. J. Virol. 2000;74:4634–4644. doi: 10.1128/jvi.74.10.4634-4644.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng Y.H., Plemenitas A., Fielding C.J., Peterlin B.M. Nef increases the synthesis of and transports cholesterol to lipid rafts and HIV-1 progeny virions. Proc. Natl. Acad. Sci. U.S.A. 2003;100:8460–8465. doi: 10.1073/pnas.1437453100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou W., Parent L.J., Wills J.W., Resh M.D. Identification of a membrane-binding domain within the amino-terminal region of human immunodeficiency virus type 1 Gag protein which interacts with acidic phospholipids. J. Virol. 1994;68:2556–2569. doi: 10.1128/jvi.68.4.2556-2569.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmerberg J., Kozlov M.M. How proteins produce cellular membrane curvature. Nat. Rev. Mol. Cell. Biol. 2006;7:9–19. doi: 10.1038/nrm1784. [DOI] [PubMed] [Google Scholar]