

Abstract
A switch of viral hemagglutinin receptor binding specificity from bird-type α2,3- to human-type α2,6-linked sialic acid is necessary for an avian influenza virus to become a pandemic virus. In this study, an easy-to-use strip test to detect receptor binding specificity of influenza virus was developed. A biotinylated anti-hemagglutinin antibody that bound a broad range of group 1 influenza A viruses and latex-conjugated α2,3 (blue) and α2,6 (red) sialylglycopolymers were used in an immunochromatographic strip test, with avidin and lectin immobilized on a nitrocellulose membrane at test and control lines, respectively. Accumulation of a sialylglycopolymer–virus–antibody complex at the test line was visualized by eye. The strip test could be completed in 30 min and did not require special equipment or skills, thereby avoiding some disadvantages of current methods for analyzing receptor binding specificity of influenza virus. The strip test could detect the receptor binding specificity of a wide range of influenza viruses, as well as small increases in the binding affinity of variant H5N1 viruses to α2,6 sialylglycans at viral titers >128 hemagglutination units. The strip test results were in agreement with those of ELISA virus binding assays, with correlations >0.95. In conclusion, the immunochromatographic strip test developed in this study should be useful for monitoring potential changes in the receptor binding specificity of group 1 influenza A viruses in the field.
Keywords: Immunochromatographic strip test, H5N1 avian influenza virus, Group 1 influenza A virus, Detection of receptor binding specificity, Sialylglycopolymer, Pandemic potential
Highlights
-
•
A novel immunochromatographic strip test system was developed.
-
•
The strip test was developed to detect influenza virus receptor binding specificity.
-
•
The strip test was applicable to a broad range of group 1 influenza A viruses.
-
•
The strip detected faint increases in human-type specificity of variant H5N1 viruses.
-
•
The system could be applied for easy monitoring the viral pandemic potential.
1. Introduction
Emerging infectious diseases, such as severe acute respiratory syndrome (SARS) and avian influenza, have been of increasing public concern in the past few decades. These diseases involve animal-to-human transmission of zoonotic pathogens (Pang and Guindon, 2004). In particular, an influenza pandemic would be devastating and a serious threat to human health and the global economy.
Avian influenza (AI) viruses were the origin of the influenza A viruses and also have been involved in the emergence of all past influenza pandemics (Webster et al., 1992). Therefore, surveillance of AI viruses to assess their evolution in the field is crucial for preparing for an influenza pandemic (Watanabe et al., 2012b).
Influenza viruses are classified into subtypes based on the antigenic properties of their two surface glycoproteins: hemagglutinin (HA) and neuraminidase (NA) (Webster et al., 1992). To date, 18 HA subtypes and 11 NA subtypes have been identified. Almost all possible combinations of HA and NA subtypes have been detected in influenza viruses isolated from aquatic birds, poultry and other bird species. The 18 HA subtypes are phylogenetically categorized into group 1 (H1, H2, H5, H6, H8, H9, H11, H12, H13, H16, H17 and H18) and group 2 (H3, H4, H7, H10, H14 and H15) HAs, with HAs in the same group antigenically related, although they can be distinguished by subtype-specific antibodies (Watanabe et al., 2012a).
HA is the main determinant of viral infectivity and consists of a head region and a stalk region (Imai and Kawaoka, 2012, Sriwilaijaroen and Suzuki, 2012). Influenza viruses attach to host cells by specific binding between the HA head region and sialylglycan that is expressed on the host cell surface. Influenza viruses also recognize terminal sialic acid (Sia) and galactose linkage patterns on sialylglycans (Imai and Kawaoka, 2012). Human influenza viruses preferentially bind to α2,6-linked Sia (α2,6 Sia), whereas most avian viruses preferentially bind to a sugar chain ending in α2,3-linked Sia (α2,3 Sia). This plays a key role of the interspecies barrier that prevents AI viruses from easily infecting humans. Therefore, it is believed that a switch of HA receptor specificity from α2,3 Sia to α2,6 Sia is essential for the emergence of a pandemic influenza virus (Watanabe et al., 2012b).
The highly pathogenic AI virus subtype H5N1 (H5N1 virus) that emerged in China around 1997 has become endemic in birds in some areas, including China, Viet Nam, Indonesia and Egypt (OIE, 2014). H5N1 virus can be directly transmitted from birds to humans and cause a severe respiratory disease with high morbidity (60%) (WHO, 2014). Fortunately, all human H5N1 infections have been restricted to people with close contact with infected poultry and there has been no sustained human-to-human transmission. However, repeated bird-to-human transmission may allow H5N1 viruses to acquire HA mutations that change their receptor specificity from α2,3 Sia (bird-type) to α2,6 Sia (human-type), thereby generating a pandemic virus. H5N1 virus has now diverged genetically to form 10 phylogenetically and phenotypically distinct clades (designated clades 0–9) in different geographic areas (Watanabe et al., 2013). Such complex ecology and diversification in the field increase the pandemic potential of H5N1 virus (Peiris et al., 2007). In addition, other subtype AI viruses, such as H9N2 and H7N9, have also been directly transmitted to humans (Garcia-Sastre and Schmolke, 2014).
Thus far, human-adaptive changes in AI viruses had been thought to occur during AI virus infections in humans and/or pigs. However, recent studies showed that AI virus could acquire increased human-type receptor binding affinity during viral transmission and infection in birds (Watanabe et al., 2011, Sriwilaijaroen and Suzuki, 2014). These results highlight the importance of monitoring possible changes in AI receptor binding specificity in the field to enable rapid response to the emergence of a pandemic influenza virus.
Several assay techniques have been developed to analyze receptor binding affinity of influenza virus (Smith and Cummings, 2014, Stevens et al., 2006, Watanabe et al., 2011, Yamada et al., 2006). The solid-phase virus binding assay is a quantitative system that can detect small changes in receptor binding affinity of influenza virus (Watanabe et al., 2011, Yamada et al., 2006). Glycan microarrays enable multiplex analysis using a panel of glycans with different topologies (Smith and Cummings, 2014, Stevens et al., 2006). A new assay system has been developed combining a virus binding assay and real-time RT-PCR (Takahashi et al., 2013). However, these very sensitive assay systems require a high level of technical expertise, expensive reagents and specialized equipment.
These limitations indicate the need for a system that is easier to use for detecting changes in influenza virus binding specificity. Immunochromatographic assays based on specific antigen–antibody reactions are very useful diagnostic tools and do not require specialized equipment or complicated handling procedures (Gopinath et al., 2014, Sakurai and Shibasaki, 2012). Despite their relatively moderate sensitivity, immunochromatographic assays have been used in various rapid diagnostic kits to detect virus infections for clinical diagnosis and surveillance, since they are fairly simple and rapid. In this study, we developed a new easy-to-use immunochoromatographic strip test to detect the emergence of AI viruses with increased human-type receptor specificity and confirmed the applicability of this test using AI viruses isolated in several different geographic areas.
2. Materials and methods
2.1. Virus isolation and preparation
Avian and human influenza viruses were grown in 10-day-old embryonated chicken eggs and MDCK cells, respectively. The allantoic fluids and culture supernatants were then harvested, pre-cleared by centrifugation at 3000 rpm for 20 min, passed through 0.45 µm filters, and stored as seed viruses at −80 °C. Viral titers were assayed as hemagglutination units (HAU) by hemagglutination assays as described below. All experiments with live H5N1 viruses were performed in Biosafety Level 3 (BSL3) conditions at Osaka University (Japan), Tottori University (Japan), National Institute of Hygiene and Epidemiology (Viet Nam) and Airlangga University (Indonesia).
2.2. Generation of viruses by reverse genetics
Recombinant viruses were generated with a plasmid-based reverse genetics system in the genetic background of influenza virus A/duck/Egypt/D1Br/2007 (EG/D1), which is one of the parental Egyptian H5N1 strains, as previously described (Watanabe et al., 2011). Mutant HA genes were generated by PCR-based site-directed mutagenesis. Recombinant EG/D1 virus and an EG/D1 variant carrying an HA mutation were denoted here as rEG/D1 and rEG/D1mutation, respectively. Recombinant viruses were propagated by single passage in eggs. The HA genes of the virus stocks were sequenced to detect the possible emergence of revertants during amplification. All studies with recombinant DNAs were conducted at Osaka University under the applicable laws and approved by the Biological Safety Committee of the Research Institute for Microbial Diseases, Osaka University (approval number 3439).
2.3. Hemagglutination titration
Stocks of avian and human influenza viruses were serially diluted with phosphate-buffered saline (PBS) and mixed with 0.5% turkey red blood cells (Nippon Biotest, Japan) and 0.75% guinea pig red blood cells (Nippon Biotest, Japan), respectively. Hemagglutination by avian and human influenza viruses was observed after incubation at room temperature for 30 min and 1 h, respectively, to determine their HAU titers.
2.4. Virus binding assay
Receptor binding specificity was analyzed by a solid-phase direct binding assay (Watanabe et al., 2011) with sialylglycopolymers (Totani et al., 2003; Suzuki et al., 2009) containing Sia linked to galactose through Siaα2,3-LacNAcβ-pAP or Siaα2,6LacNAcβ-pAP. A 1 µg/ml solution of each sialylglycopolymer (non-conjugated with colored latex) was prepared in PBS, and 100 µl of this solution was added to each well of 96-well microtiter plates (Polystyrene Universal-Bind Microplates, Corning, USA). The plates were then irradiated with 254 nm ultraviolet light for 10 min, and each well was washed three times with 250 µl PBS. Each well was blocked with 200 µl SuperBlock Blocking Buffer (Thermo Scientific, USA) at room temperature for 1 h. After washing with ice-cold PBS containing 0.1% Tween 20 (PBST), a serial dilution of 256 HAU of an influenza virus in PBST was added to the wells, and the plates were incubated at 4 °C for 12 h. After five washes with ice-cold PBST, monoclonal antibody against influenza virus nucleoprotein, which is a structural protein of the viral particle, was added to each well, and the plates were incubated at 4 °C for 2 h. The wells were then washed five times with ice-cold PBST and incubated with peroxidase-conjugated goat anti-immunoglobulin (Histofine Simple Stain MAX-PO, Nichirei, Japan) at 4 °C for 2 h. After washing five times with ice-cold PBST, 100 µl SureBlue Reserve Peroxidase Substrate (KPL, USA) was added to each well and, after incubation at room temperature for 10 min, absorbance at 630 nm (developed blue color) was measured.
2.5. Assembly of strip test
The test system in this study included an expansion solution, an antibody solution, and two test strips. The expansion solution was a mixture of surfactant agents: 0.2% Triton X-405, 1% Tween 20, and 50 μg DANA (N-acetyl-2.3-didehydro-2-deoxyneuraminic acid)/ml in Tris–HCl buffer (pH 7.5). These surfactant concentrations were sufficient to inactivate influenza virus infectivity and sialidase activity. The antibody solution contained 0.12 mg biotinylated anti-HA antibody C179/ml. C179 is a monoclonal antibody against the antigenically conserved HA stem region (Okuno et al., 1993, Sakabe et al., 2010), and has been reported to bind to a broad range of group 1 influenza HAs (H1, H2, H5, H6 and H9) (Pan et al., 2014, Smirnov Iu et al., 1999). A preliminary study showed that C179 reacted with all of the group 1 influenza HAs in this study, while monoclonal antibodies against the antigenically variable HA head region only reacted with some of these HAs (Table S1). C179 binding to the HA stem region has been reported to not interfere with the interaction between sialylglycans and the HA head region (Okuno et al., 1993), which enabled its use in the strip test to detect sialylglycan–HA complexes.
Each test strip consisted of four components assembled on a plastic backing: a sample pad, a sialylglycopolymer pad, a nitrocellulose (NC) membrane and an absorption pad ( Fig. 1A). The three pads were as follows: the sample pads were Rayon Nonwoven Fabric (Advantec), each 4 mm wide, 2 cm long and 0.24 mm thick; the sialylglycopolymer pads were glass fiber (Millipore Corporation), each 4 mm wide, 8 mm long and 0.43 mm thick; and the absorption pads were filter paper (Advantec), each 4 mm wide, 3.4 cm long and 1.35 mm thick. The NC membranes (Millipore Corporation) were 4 mm wide and 3 cm long, and the plastic backings were adhesive backing cards (Lohmann Technologies Corp.), each 4 mm wide and 7.6 cm long. A sialylglycopolymer pad was prepared by spraying it with either blue latex-conjugated α2,3 sialylglycopolymer or red latex-conjugated α2,6 sialylglycopolymer (denoted as an α2,3 Sia strip and an α2,6 Sia strip, respectively) and drying at room temperature. Our previous study showed that the latex-conjugated sialylglycopolymers are remarkably stable without any aggregation even in boiling water (Totani et al., 2003). An 0.4 μl sample of a 4 mg avidin/ml solution and a 0.4 μl sample of a 1 mg lectin/ml solution were dispensed on each NC membrane to form test and control lines, respectively, and the membrane was air-dried. The distance between test and control lines was about 7 mm.
Fig. 1.
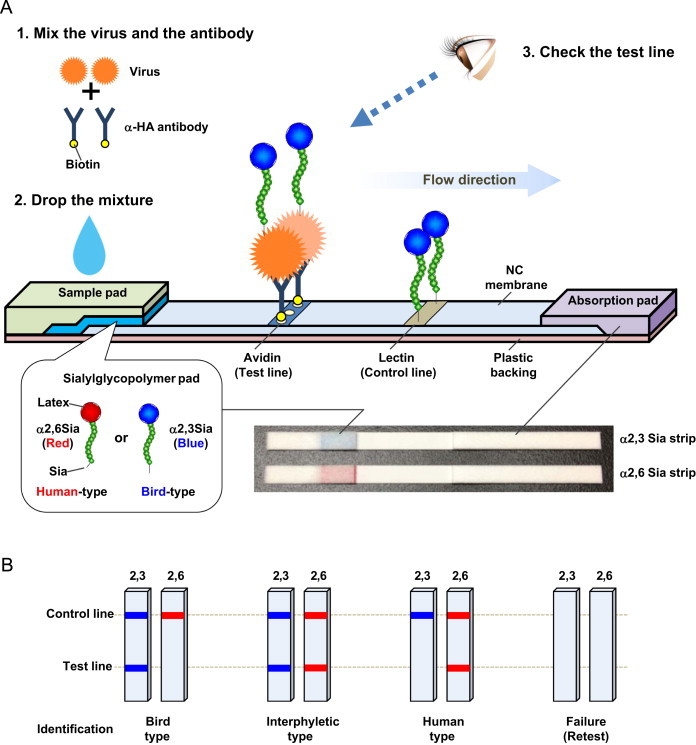
Schematic of the immunochromatographic test strip system. (A) Schematic of the test strip. A virus sample and biotinylated anti-HA antibody C179 were mixed and incubated for 15 min. The mixture was then applied on the sample pad. During migration though the sialylglycopolymer pad, virus–C179 antibody complexes formed a colored sialylglycopolymer–virus–biotinylated antibody complex, which was captured by avidin at the test line. Accumulation of the trimetric complex produced a visible test line with the color of the bound sialylglycopolymer (α2,3 Sia, blue; α2,6 Sia, red). Excess complexes and free sialylglycopolymers were captured by lectin at the control line. The strip test was completed 15 min after the virus–antibody complex was applied on the sample pad. A photograph of the strips is also shown. (B) Schematic illustration of the viral receptor binding specificity patterns on the test strip. Based on the patterns of visible bands on the α2,3 Sia and α2,6 Sia strips, viral receptor binding specificity was identified as indicated.
To assemble a test strip, an NC membrane was pasted onto the center of the plastic backing (using acrylic adhesive) and covered with the sialylglycopolymer pad at one end and the absorption pad at the other end. A sample pad was pasted onto the plastic backing at the top of the sialylglycopolymer pad. The NC membranes with attached pads were then cut into 4 mm wide test strips and stored desiccated at 4 °C.
2.6. Test strip analysis
Virus samples (30 µl) were mixed with 90 µl of expansion solution and 30 µl of antibody solution, and then incubated for 15 min to form virus–C179 complexes (Fig. 1A). Each mixture was then pipetted onto a test strip sample pad or, alternatively, a sample pad was immersed in the virus mixture. This allowed the mixture to migrate along the strip by capillary action. During migration through the sialylglycopolymer pad, the virus–C179 complex was able to form a complex with α2,3 Sia or α2,6 Sia depending on the virus HA receptor binding affinity. Principle of the subsequent reaction is mainly based on two types of interaction: avidin–biotin interaction and lectin–glycan interaction. As the virus–C179–sialylglycopolymer complex migrated along the test strip, it was captured at the test line by avidin–biotinylated C179 interaction. Accumulation of the trimeric complex resulted in visual blue and red test lines on the α2,3 Sia strips and α2,6 Sia strips, respectively. If a viral HA did not bind either sialyglycopolymer, there was no visible test line because the captured virus–C179 dimeric complex was colorless. Excess complexes and free sialylglycopolymers were captured at the control line by lectin–glycan interactions, resulting in visible blue and red control lines on the α2,3 Sia strips and α2,6 Sia strips, respectively.
The results of this assay were observed by eye 15 min after deposition of the virus mixture onto the strips. The presence of double blue lines on the α2,3 Sia strip and double red lines on the α2,6 Sia strip indicated virus binding affinity to α2,3 Sia and α2,6 Sia, respectively. Only a blue control line on the α2,3 Sia strip and only a red control line on the α2,6 Sia strip indicated no binding affinity to α2,3 Sia and α2,6 Sia respectively, or that the amount of virus was below the limit of detection of this assay. If a control line did not appear within 15 min, the test was considered invalid. Thus, the complete assay took only 30 min.
2.7. Identification of viral receptor binding specificity by the strip test
In past influenza pandemics, mutant viruses with dual tropism for both α2,3 Sia and α2,6 Sia have emerged in the immediate early phase of an outbreak (Matrosovich et al. 2000). These prototype viruses subsequently switched their receptor binding specificity from bird-type to human-type sialyglycans to become established pandemic viruses. Therefore, based on the patterns of the test lines on the α2,3 Sia and α2,6 Sia strips (Fig. 1B), viral receptor binding specificity was identified as follows: (i) only a blue test line indicated typical bird-type receptor specificity; (ii) both blue and red test lines indicated increased human-type receptor affinity with residual bird-type receptor affinity, denoting an interphyletic mutant virus; and (iii) only a red test line alone indicated a switch of viral receptor affinity from bird-type to human-type receptors, denoting either a novel virus with a high pandemic potential or a pandemic virus.
2.8. Densitometric analysis
Band intensities on test strips were quantified by ImageJ software, and the test line intensity was calculated by subtracting the background intensity from the test line intensity.
3. Results and discussion
For this study, the receptor binding specificity of several virus strains that were used as reference viruses were confirmed by virus binding assays. The reference viruses were as follows: A/Suita/117/2011 (Suita/117), a human H1N1 strain; A/Japan/434/2003 (Japan/434), a human H3N2 strain; EG/D1 and recombinant rEG/D1 virus, bird H5N1 strains; and rEG/D1Q192H and rEG/D128Δ/I151T, mutant rEG/D1 viruses with enhanced human-type receptor specificity. Previous studies reported that HA Q192H and 128Δ/I151T mutations increased viral binding affinity for α2,6 Sia (Watanabe et al., 2011). Therefore, rEG/D1Q192H and rEG/D1128Δ/I151T were generated by introducing the Q192H and 128Δ/I151T mutations into EG/D1 HA to produce representative interphyletic viruses for this study. The virus binding assays showed that Suita/117 and Japan/434 had binding specificity for α2,6 Sia ( Fig. 2A and B). In contrast, EG/D1 and rEG/D1 had binding specificity for α2,3 Sia (Fig. 2C and D). Interphyletic viruses rEG/D1Q192H and rEG/D1128Δ/I151T had significantly increased binding to α2,6 Sia, with binding to both α2,3 Sia and α2,6 Sia (Fig. 2E and F). The binding curves of these viruses were typical Sia binding specificity profiles for human viruses, bird viruses, and interphyletic viruses, respectively (Boltz et al., 2010, Watanabe et al., 2011, Yamada et al., 2006).
Fig. 2.
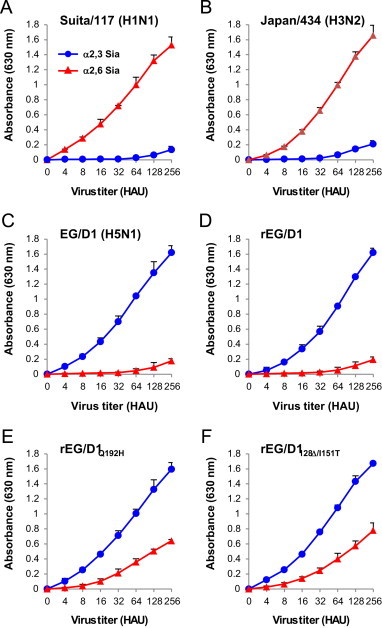
Receptor binding specificity of the reference viruses in this study. Virus binding assays of the reference viruses to sialylglycopolymers containing either α2,3 Sia (blue circle) or α2,6 Sia (red triangle). (A) Human H1N1 strain. (B) Human H3N2 strain. (C and D) Bird H5N1 strains. (E and F) H5N1 mutant strains with increased α2,6 Sia binding affinity and residual α2,3 Sia binding. Each data point is the mean±SD of three independent experiments. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
We evaluated the sensitivity and specificity of the strip test by investigating the reactivity of 8–512 HAU of the reference viruses to α2,3 Sia and α2,6 Sia. Suita/117 gave clear red test lines on α2,6 Sia strips for virus titers of 32–512 HAU ( Fig. 3A). However, even at the highest titer, 512 HAU, Suita/117 produced no visible blue test line on the α2,3 Sia strip, indicating human-type receptor specificity. Japan/434, which is classified as a group 2 influenza A virus, produced no visible test lines on either α2,6 Sia or α2,3 Sia strips (Fig. 3B). These results showed that the visible test lines produced on the test strips were the result of specific C179 binding to group 1 influenza HAs.
Fig. 3.
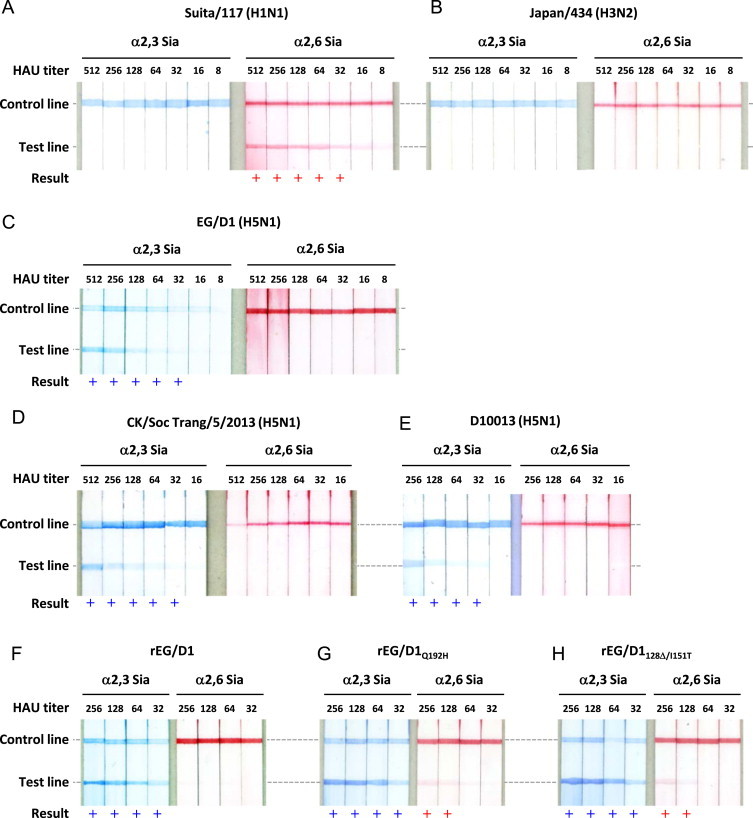
Sensitivity and specificity of the test strips. Test strip patterns as a function of viral HAU titers. (A) Human H1N1 strain. (B) Human H3N2 strain. (C–F) Bird H5N1 strains. (G and H) H5N1 mutant strains with increased α2,6 Sia binding affinity and residual binding to α2,3 Sia. + Indicates a positive result.
In contrast, EG/D1 gave clear blue test lines on α2,3 Sia test strips when the virus titer was 32–512 HAU (Fig. 3C). However, even the highest EG/D1 titer, 512 HAU, produced no visible red test line on α2,6 Sia strips, showing bird-type receptor specificity. To further assess the receptor specificity of H5N1 viruses that were circulating in geographical areas other than Egypt, we tested A/chicken/Soc Trang/5/2013 and A/tree sparrow/Indonesia/D10013/2010, which were typical bird-type H5N1 viruses isolated in Viet Nam and Indonesia, respectively. These two viruses produced blue test lines only on α2,3 Sia strips, with the same sensitivity (Fig. 3D and E). These results showed that the test strips consistently detected the receptor binding specificity of group 1 influenza viruses.
The reactivity of the test strips to the interphyletic viruses with titers of 32–256 HAU was then evaluated. Both rEG/D1Q192H and rEG/D1128Δ/I151T produced clear blue test lines on α2,3 Sia strips at all virus titers tested (Fig. 3G and H). In addition, 128–256 HAU of these viruses produced visible red test lines on α2,6 Sia strips, in contrast to the results with the parental rEG/D1 virus (Fig. 3F). These results showed that the test strips were able to detect human-type receptor binding by interphyletic viruses at higher titers (>128 HAU). Based on these results, we used virus isolates with titers of 128–512 HAU for further studies.
Band intensities on the test strips were quantified by densitometry and plotted as a function of virus titer. The results showe d that the intensity profiles ( Fig. 4A–F) were similar to the virus binding assay results (Fig. 2A–F). Scatter plots showed that an increase in band intensity was strongly correlated with binding affinity on both α2,3 Sia (R=0.966, Fig. 4G) and α2,6 Sia strips (R=0.957, Fig. 4H), indicating that the results obtained with the test strips and by binding assays were highly correlated. Therefore, the test strips produced a semi-quantitative measure of viral receptor binding affinity.
Fig. 4.
Quantification of band intensities at the strip test line. (A–F) Intensities of the bands shown in Fig. 3 were quantified by ImageJ software. The intensities of the bands in the α2,3 Sia strips (blue circle) and α2,6 Sia strips (red triangle) were expressed relative to the maximum values for each virus, and plotted as a function of virus HAU titer. (A) Human H1N1 strain. (B) Human H3N2 strain. (C and D) Bird H5N1 strains. (E and F) H5N1 mutant strains with increased human-type receptor affinity. For the human H3N2 strain (B), the intensity values were calculated relative to the maximum values for the human H1N1 strain (A) that was analyzed at the same time. Each data point is the mean±SD of three independent experiments. (G and H) Scatter plots of virus binding affinity (x-axis) versus test line band intensity (y-axis) on α2,3 Sia strips (G) and α2,6 Sia strips (H). Virus binding affinity was measured as absorbance (630 nm) in virus binding assays. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
A long-term storage test was then performed to evaluate the stability and reproducibility of the strip test. Investigations using the same batch of test strips showed that the test strips were stable for at least 12 months at 4 °C without any changes in sensitivity or specificity (Table S2).
To verify the potential applicability of the test strip, the HA receptor specificity of 63 influenza virus strains that had been isolated in the several geographical areas were examined. The results showed that 4 strains that were classified as Egyptian clade 2.2.1 sublineage A/B viruses had dual binding affinity to both α2,3 Sia and α2,6 Sia, indicating that they were interphyletic viruses ( Fig. 5 and Table 1). This was in agreement with the previous report that sublineage A/B viruses have generally acquired an increased human-type receptor binding affinity (Watanabe et al., 2011). In contrast, all the H5N1 strains that were isolated in Viet Nam and Indonesia showed typical bird-type receptor specificity. Several variant-type strains with increased α2,6 Sia binding affinity have been reported in some Asian countries (Auewarakul et al., 2007, Boltz et al., 2010). However, acquisition of human-type receptor affinity by Asian H5N1 viruses seemed to be strain-specific, which may explain why the Asian H5N1 viruses tested in this study did not show dual receptor specificity. Further evaluation of the test strips using larger numbers of Asian H5N1 strains may detect interphyletic viruses in these areas.
Fig. 5.
Representative photographs of application of the strip test to viruses isolated in other areas. (A) H5N1 strains isolated in Egypt. (B) H5N1 strains isolated in Viet Nam. (C) H5N1 strains isolated in Indonesia. + Indicates a positive result.
Table 1.
Summary of strip test results in this study.
| Lineage | Subtype | Country | Strain | Clade (sublineage) | HAU | α2,3 Sia strip | α2,6 Sia strip | Identification |
|---|---|---|---|---|---|---|---|---|
| Group 1 | H5N1 HPAIV | Egypt | A/duck/Egypt/D1Br/2007 | 2.2.1 | 256 | + | − | Bird |
| A/duck/Egypt/D3Li12/2007 | 2.2.1 | 256 | + | − | Bird | |||
| A/chicken/Egypt/C1Tr13/2007 | 2.2.1 | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD1-5/2008 | 2.2.1 | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD5-3/2008 | 2.2.1 | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD8-14/2008 | 2.2.1 | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD5-3/2008 | 2.2.1 | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD12-3/2008 | 2.2.1 (A) | 256 | + | + | Dual | |||
| A/Egypt/N04822/2009 | 2.2.1 (A) | 256 | + | + | Dual | |||
| A/goose/Egypt/0929-NLQP/2009 | 2.2.1 (B) | 256 | + | + | Dual | |||
| A/Egypt/N2039/2009 | 2.2.1 (B) | 256 | + | + | Dual | |||
| A/chicken/Egypt/RIMD25-1/2009 | 2.2.1 (C) | 256 | + | – | Bird | |||
| A/chicken/Egypt/RIMD26-3/2009 | 2.2.1 (C) | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD13-1/2008 | 2.2.1 (C) | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD27-2/2009 | 2.2.1 (C) | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD28-1/2009 | 2.2.1 (C) | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD29-3/2009 | 2.2.1 (C) | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD4-3/2008 | 2.2.1 (D) | 256 | + | − | Bird | |||
| A/chicken/Egypt/RIMD11/2008 | 2.2.1 (D) | 256 | + | − | Bird | |||
| Viet Nam | A/Vietnam/HN3028/2003 | 1 | 256 | + | − | Bird | ||
| A/Vietnam/HN3040/2003 | 1 | 256 | + | − | Bird | |||
| A/Vietnam/HN3062/2004 | 1 | 128 | + | − | Bird | |||
| A/Vietnam/30259/ 2004 | 1 | 128 | − | − | un | |||
| A/chicken/Soc Trang/1/2012 | 1 | 128 | + | − | Bird | |||
| A/duck/Soc Trang/8/2012 | 1 | 512 | + | − | Bird | |||
| A/chicken/Soc Trang/3/2013 | 1 | 512 | + | − | Bird | |||
| A/chicken/Soc Trang/4/2013 | 1 | 512 | + | − | Bird | |||
| A/chicken/Soc Trang/5/2013 | 1 | 512 | + | − | Bird | |||
| A/Vietnam/CM32/2011 | 2.3.2.1 | 256 | + | − | Bird | |||
| A/muscovy duck/Vietnam/LBM14/2011 | 2.3.2.1 (VN-A) | 128 | + | − | Bird | |||
| A/muscovy duck/Vietnam/LBM227/2011 | 2.3.2.1 (VN-C) | 128 | + | − | Bird | |||
| A/Vietnam/30850/ 2005 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN31203/2007 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN31244/2007 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN31312/2007 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN31323/2007 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN31388/2007 | 2.3.4 | 256 | + | − | Bird | |||
| A/duck/Vietnam/G12/2008 | 2.3.4 | 128 | + | − | Bird | |||
| A/Vietnam/HN31394/2008 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN31412/2008 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN31413/2008 | 2.3.4 | 256 | − | − | un | |||
| A/Vietnam/HN31432/2008 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN31461/2008 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN31673/2009 | 2.3.4 | 256 | + | − | Bird | |||
| A/Vietnam/HN 36250/2010 | 2.3.4 | 256 | + | − | Bird | |||
| Indonesia | A/Indonesia/5/2005 | 2.1.3 | 256 | + | − | Bird | ||
| A/tree sparrow/Indonesia/D10013/2010 | 2.1.3 | 512 | + | − | Bird | |||
| A/chicken/Indonesia/D12069/2012 | un | 256 | + | − | Bird | |||
| A/chicken/Indonesia/D12021/2012 | un | 128 | + | − | Bird | |||
| A/chicken/Indonesia/D12067/2012 | un | 256 | + | − | Bird | |||
| A/chicken/Indonesia/D12069/2012 | un | 256 | + | − | Bird | |||
| A/chicken/Indonesia/D12096/2012 | un | 256 | + | − | Bird | |||
| Thailand | A/Thailand/Kan353/2004 | 1 | 256 | + | − | Bird | ||
| China | A/Shanghai/1/2006 | 2.3.4 | 256 | + | − | Bird | ||
| Japan | A/crow/Kyoto/53/2004 | 2.5 | 256 | + | − | Bird | ||
| H5N1 LPAIV | Japan | A/mallard/Hokkaido/24/09 | 256 | + | − | Bird | ||
| H5N3 | Hong Kong | A/duck/Hong Kong/313/4/78 | 256 | + | − | Bird | ||
| Seasonal H1N1 | Japan | A/Narita/1/2009 | 256 | − | + | Human | ||
| USA | A/New Caledonia/20/99 | 256 | − | + | Human | |||
| H1N1pdm | Japan | A/Suita/112/2011 | 256 | − | + | Human | ||
| Japan | A/Suita/117/2011 | 256 | − | + | Human | |||
| Group 2 | H3N2 | USA | A/New York/55/2004 | 256 | − | − | un | |
| Japan | A/Japan/434/2003 | 256 | − | − | un | |||
Abbreviations: HPAIV, highly pathogenic avian influenza virus; LPAIV, low pathogenic avian influenza virus; +, positive result; −, negative result; un, unknown; Bird, bird-type receptor specificity; Dual, dual receptor affinity; Human, human-type receptor specificity.
Clade 2.2.1 sublineage A-D viruses have been defined by Watanabe et al., 2012.
In this study, two H5N1 virus strains produced no visible test lines on either α2,3 Sia or α2,6 Sia test strips (Table 1), even though the titers of these viruses were >128 HAU. Epidemiological studies have reported a few exceptional virus strains that recognized distinct sialylglycan topologies (Xiong et al., 2013). Thus, these two viruses may have such an exceptional receptor binding specificity. However, these viruses may not have bound the C179 antibody due to amino acid change(s) at the antigen epitope in the HA stem region. Further genetic analyses and characterization of these two viruses are needed to elucidate the reason(s) why these viruses did not produce bands on the test strips.
4. Conclusions
The results described above demonstrated the utility of the test strip system developed in this study to analyze the receptor specificity of influenza viruses. Compared with conventional methods to analyze receptor binding affinity of influenza virus, the test strip system was simple and easy to use: only a drop of virus and the test strips were required, without any additional methods or equipment. The test strips were shown to be applicable to a broad range of group 1 influenza A vruses and allowed semi-quantitative detection of their receptor binding affinities. These results indicate that the test strips could be applied as a screening device to monitor possible evolutionary changes in AI receptor binding specificity in the field.
The sensitivity of the test strips was not adequate for rapid diagnosis of clinical samples (e.g., nasopharyngeal swabs). However, traditional methods for determining viral genome sequences, pathogenicity and drug resistance using isolated viruses are still routinely used in public health and academic laboratories worldwide (Kumar and Henrickson, 2012). Since isolated virus samples contain significantly more virus than clinical samples, application of the strip test to influenza virus analysis should enable us to more easily and comprehensively monitor AI evolution and pandemic potential in the field. The strip test system described here could then be applied for rapid diagnosis in the field when its sensitivity is improved.
Acknowledgments
The authors would like to thank Dr. Tadahiro Sasaki and Dr. Kazuo Takahashi for assistance with this project; Dr. Masato Tashiro and Dr. Takato Odagiri for valuable discussions. This work was supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology, Japan (JSPS KAKENHI Grant numbers 23791134 and 23406017) and by Japan Initiative for Global Research Network on Infectious Diseases (J-GRID).
Footnotes
Supplementary data associated with this article can be found in the online version at doi:10.1016/j.bios.2014.10.036.
Contributor Information
Yohei Watanabe, Email: nabe@biken.osaka-u.ac.jp.
Yasuo Suzuki, Email: suzukiy@isc.chubu.ac.jp.
Appendix A. Supplementary materials
Supplementary material
Supplementary material
References
- Auewarakul P., Suptawiwat O., Kongchanagul A., Sangma C., Suzuki Y., Ungchusak K., Louisirirotchanakul S., Lerdsamran H., Pooruk P., Thitithanyanont A., Pittayawonganon C., Guo C.T., Hiramatsu H., Jampangern W., Chunsutthiwat S., Puthavathana P. J. Virol. 2007;81(18):9950–9955. doi: 10.1128/JVI.00468-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boltz D.A., Douangngeun B., Phommachanh P., Sinthasak S., Mondry R., Obert C., Seiler P., Keating R., Suzuki Y., Hiramatsu H., Govorkova E.A., Webster R.G. J. Gen. Virol. 2010;91(Pt 4):949–959. doi: 10.1099/vir.0.017459-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Sastre A., Schmolke M. Lancet. 2014;383(9918):676–677. doi: 10.1016/S0140-6736(14)60163-X. [DOI] [PubMed] [Google Scholar]
- Gopinath S.C., Tang T.H., Chen Y., Citartan M., Tominaga J., Lakshmipriya T. Biosens. Bioelectron. 2014;61C:357–369. doi: 10.1016/j.bios.2014.05.024. [DOI] [PubMed] [Google Scholar]
- Imai M., Kawaoka Y. Curr. Opin. Virol. 2012;2(2):160–167. doi: 10.1016/j.coviro.2012.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar S., Henrickson K.J. Clin. Microbiol. Rev. 2012;25(2):344–361. doi: 10.1128/CMR.05016-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matrosovich M., Tuzikov A., Bovin N., Gambaryan A., Klimov A., Castrucci M.R., Donatelli I., Kawaoka Y. J. Virol. 2000;74(18):8502–8512. doi: 10.1128/jvi.74.18.8502-8512.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- OIE, 2014. 〈http://www.oie.int/animal-health-in-the-world/update-on-avian-influenza/〉.
- Okuno Y., Isegawa Y., Sasao F., Ueda S. J. Virol. 1993;67(5):2552–2558. doi: 10.1128/jvi.67.5.2552-2558.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan Y., Sasaki T., Kubota-Koketsu R., Inoue Y., Yasugi M., Yamashita A., Ramadhany R., Arai Y., Du A., Boonsathorn N., Ibrahim M.S., Daidoji T., Nakaya T., Ono K.I., Okuno Y., Ikuta K., Watanabe Y. Biochem. Biophys. Res. Commun. 2014;450(1):42–48. doi: 10.1016/j.bbrc.2014.05.060. [DOI] [PubMed] [Google Scholar]
- Pang T., Guindon G.E. EMBO Rep. 2004;(5 Spec No):S11–S16. doi: 10.1038/sj.embor.7400226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peiris J.S., de Jong M.D., Guan Y. Clin. Microbiol. Rev. 2007;20(2):243–267. doi: 10.1128/CMR.00037-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakabe S., Iwatsuki-Horimoto K., Horimoto T., Nidom C.A., Le M., Takano R., Kubota-Koketsu R., Okuno Y., Ozawa M., Kawaoka Y. Antivir. Res. 2010;88(3):249–255. doi: 10.1016/j.antiviral.2010.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakurai A., Shibasaki F. Viruses. 2012;4(8):1235–1257. doi: 10.3390/v4081235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smirnov Iu A., Lipatov A.S., Okuno I., Gitel’man A.K. Vopr. Virusol. 1999;44(3):111–115. [PubMed] [Google Scholar]
- Smith D.F., Cummings R.D. Curr. Opin. Virol. 2014;7C:79–87. doi: 10.1016/j.coviro.2014.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sriwilaijaroen N., Suzuki Y. Proc. Jpn. Acad. Ser. B. Phys. Biol. Sci. 2012;88(6):226–249. doi: 10.2183/pjab.88.226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sriwilaijaroen N., Suzuki Y. Methods Mol. Biol. 2014;1200:447–480. doi: 10.1007/978-1-4939-1292-6_38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevens J., Blixt O., Paulson J.C., Wilson I.A. Nat. Rev. Microbiol. 2006;4(11):857–864. doi: 10.1038/nrmicro1530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki, Y., Asai, A., Suzuki, T., Hidari, K., Murata, T., Usui, T., Takeda, S., Yamada, K., Noguchi, T., 2009. United States Patent Application Publication, Pub No. US2009/0181362A1
- Takahashi T., Kawakami T., Mizuno T., Minami A., Uchida Y., Saito T., Matsui S., Ogata M., Usui T., Sriwilaijaroen N., Hiramatsu H., Suzuki Y., Suzuki T. PLoS One. 2013;8(10):e78125. doi: 10.1371/journal.pone.0078125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Totani K., Kubota T., Kuroda T., Murata T., Hidari K.I., Suzuki T., Suzuki Y., Kobayashi K., Ashida H., Yamamoto K., Usui T. Glycobiology. 2003;13(5):315–326. doi: 10.1093/glycob/cwg032. [DOI] [PubMed] [Google Scholar]
- Watanabe Y., Ibrahim M.S., Ellakany H.F., Kawashita N., Daidoji T., Takagi T., Yasunaga T., Nakaya T., Ikuta K. J. Gen. Virol. 2012;93(Pt 10):2215–2226. doi: 10.1099/vir.0.044032-0. [DOI] [PubMed] [Google Scholar]
- Watanabe Y., Ibrahim M.S., Suzuki Y., Ikuta K. Trends Microbiol. 2012;20(1):11–20. doi: 10.1016/j.tim.2011.10.003. [DOI] [PubMed] [Google Scholar]
- Watanabe Y., Ibrahim M.S., Ellakany H.F., Kawashita N., Mizuike R., Hiramatsu H., Sriwilaijaroen N., Takagi T., Suzuki Y., Ikuta K. PLoS Pathog. 2011;7(5):e1002068. doi: 10.1371/journal.ppat.1002068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe Y., Ibrahim M.S., Ikuta K. EMBO Rep. 2013;14(2):117–122. doi: 10.1038/embor.2012.212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster R.G., Bean W.J., Gorman O.T., Chambers T.M., Kawaoka Y. Microbiol. Rev. 1992;56(1):152–179. doi: 10.1128/mr.56.1.152-179.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO, 2014. 〈http://www.who.int/influenza/human_animal_interface/avian_influenza/en/〉.
- Xiong X., Tuzikov A., Coombs P.J., Martin S.R., Walker P.A., Gamblin S.J., Bovin N., Skehel J.J. Virus Res. 2013;178(1):12–14. doi: 10.1016/j.virusres.2013.08.007. [DOI] [PubMed] [Google Scholar]
- Yamada S., Suzuki Y., Suzuki T., Le M.Q., Nidom C.A., Sakai-Tagawa Y., Muramoto Y., Ito M., Kiso M., Horimoto T., Shinya K., Sawada T., Kiso M., Usui T., Murata T., Lin Y., Hay A., Haire L.F., Stevens D.J., Russell R.J., Gamblin S.J., Skehel J.J., Kawaoka Y. Nature. 2006;444(7117):378–382. doi: 10.1038/nature05264. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material
Supplementary material