

Abstract
Paternal investment is predicted to be a facultative calculation based on expected fitness returns and modulated by a host of social predictors including paternity uncertainty. However, the direct role of paternity confidence on the patterns of paternal investment is relatively unknown, in part due to a lack of research in populations with high levels of paternity uncertainty. Additionally, much of the work on paternity certainty uses cues of paternity confidence rather than direct assessments from fathers. We examine the effect of paternity assertions on the multiple measures of paternal investment in Himba pastoralists. Despite a high degree of paternity uncertainty, Himba have strong norms associated with social fatherhood, with men expected to invest equally in biological and non-biological offspring. Our behavioural data show patterns that largely conform to these norms. For domains of investment that are highly visible to the community, such as brideprice payments, we find no evidence of investment biased by paternity confidence. However, more private investment decisions do show some evidence of sex-specific titration. We discuss these results in light of broader considerations about paternal care and the mating–parenting trade-off.
Keywords: paternity certainty, paternal investment, life-history theory
1. Introduction
Paternal investment is a calculated risk. Care from fathers can significantly impact the fitness prospects of offspring, but where paternity is uncertain, such care can result in the misallocation of time, energy and resources. As a result, paternity certainty is thought to be one of the most critical factors determining the level of investment that males provide [1,2]. Where they do provide care, males are expected either to titrate their investment based on paternity confidence, or to balance sunk costs in others' offspring with either somatic or reproductive gains [3]. They are also expected to care more when the risks of non-paternity are outweighed by the potential benefit of their care on offspring fitness and where there are few substitutes who can provide care in their place [4,5].
Despite widespread acceptance of the importance of paternity, establishing strong causal evidence linking paternity certainty to paternal care is challenging. Factors such as male condition, expected returns on investment and future reproductive value can cloud the predicted relationship between paternity and paternal care [6]. As a result, evidence connecting them is decidedly mixed [7]. For example, in some bird species where males engage in paternal care and extra-pair paternity is common, more attractive males are less likely to provide care, investing instead in mating effort [8]. In other cases, paternity certainty appears to modulate certain forms of care, but not others [9,10]. Nevertheless, some studies, mainly of birds, provide strong support for the role of paternity certainty in paternal care. In particular, experimental manipulations and within-species comparisons provide the most robust evidence [11–13].
In humans, the picture is more complicated. Fathers can take on both a social (pater) and a biological (genitor) role, and the two are not always synchronized. Furthermore, human parental investment is long-lasting and multifaceted, and is embedded within broader sets of cultural norms, multi-layered social networks, and institutional structures and systems. Given this complexity, paternity is clearly just one among many factors in the decision of whether and how much care fathers will provide. Here, we review the evidence that cues, perceptions and assertions of paternity certainty impact paternal care and investment in humans, and provide a unique case study of both norms and behaviour in a setting where low paternity certainty is concurrent with considerable paternal obligations.
(a). Human studies linking paternity and paternal care
There is a substantial literature linking cues of paternity to the patterns of investment in humans. Many studies rely on facial resemblance between father and offspring as a cue of paternity, correlated with one or more measures of paternal care [14–20]. Others relate perceived facial resemblance to ratings of father-offspring closeness and parenting quality [21–23]. These studies are supported by the evidence of cognitive mechanisms to detect relatedness in assessments of facial morphology [24,25] and social biases that promote paternal confidence [26,27]. Olfactory cues have also been studied, with similar effects [14,28]. There are, however, some limitations to using physical resemblance as a cue of paternity. It is possible that a reversed causal link from care to resemblance is driving the correlation [29], or that resemblance is triggering care due to assortative preferences, rather than because it is a reliable cue of paternity [30,31]. Another limitation is that almost all of these studies rely on self-reported measures of investment, and given generalized expectations that men should engage in paternal care, this method could result in biases in reporting [29].
In addition to the methodological limitations described above, current studies have almost exclusively been conducted in western societies, where the rate of extra-pair paternity is very low [30]. In one of the only studies using actual paternity assertions, Anderson et al. [31] found that low paternity confidence resulted in reduced reported involvement in children's schooling and time spent with children in groups. Pregnancies associated with low paternity confidence were also associated with the greater risk of divorce. However, low paternity confidence was reported for only 49 of 3066 pregnancies (1.46%), and in another 3% of pregnancies, men refused to answer questions about paternity. Teasing out the effects of paternity uncertainty on paternal investments and paternal care may be more fruitful in societies where extra-pair paternity is more common. This would allow for larger sample sizes of men with low paternity confidence, and where extra-marital sex is less stigmatized, greater ability to collect reliable data on paternity confidence.
(b). Paternity confidence and paternal care decisions
While there is a general expectation that men will titrate paternal care based on paternity confidence, there are several instances when it might be beneficial for men to treat biological and non-biological children equally. First, when offspring bring in substantial benefits to the household, fathers may be incentivized to treat children equally, or to bias investment towards those who are harder working, better contributors or members of a favoured gender. For example, among the Kpelle, men rely on the production of their children for domestic and agricultural labour, and may insist on social fatherhood of a child even when their paternity is suspect, in order to capitalize on the economic value children bring to subsistence activities [32]. As they age, men may also rely on their children to support them, another advantage that can come regardless of paternity [33]. Treating a non-biological child well could engender greater reciprocity if it were viewed as less obligatory. Fathers can also reap economic benefits from daughters' marriages, where brideprice is paid. This could cause a sex bias in investment, as competition between sons (biological and non) may be greater than competition between daughters [34,35].
Second, men's investment in their children may serve as a signal to current and future mates and allies of their generosity, their sociability or their provisioning ability [36–38]. In the animal literature, Kokko et al. [39] describe a similar concept, where males compete for females by demonstrating their prowess as good providers and there is a substantial literature more broadly positing that paternal care can serve as mating effort [40,41]. Furthermore, violating social norms related to parental care could come at a social or reputational cost; it may be more advantageous for men to provide care than to suffer a cost for shirking their duties. Such costs may include reduced reproductive success through the loss of a marital partner, or being seen as a less desirable partner by future mates or allies.
Third, the accrual of children, regardless of biological paternity, can in and of itself be an important form of social capital [32,33,42]. Among Yoruba, an older man who is impotent would secretly allow another man to father his children, whom he would provide for even after his death, all the while retaining his role as their father [43]. In other cases, children, and the relationships they sow throughout their lives, can extend the networks and prestige of a man [44]. Where children have this kind of social value, men may be incentivized to treat them equally, regardless of paternity.
Considering these factors together, we propose that the extent to which paternity certainty is associated with paternal care should be variable and domain specific. We examine both patterns of investment and social norms surrounding investment among Himba pastoralists living in northern Namibia. This population has a high rate of concurrency, which is largely normalized [45,46], and an extra-pair paternity rate of 48% [47]. This results in a large sample of cases of suspected non-paternity and relative ease in collecting paternity assertions. Fathers have been shown to be quite accurate in their paternity assertions, correctly assessing paternity 73% of the time [47]. In order to understand context-specific paternal care, we assess the link between paternity confidence and paternal care in four domains: child nutrition, livestock transfers, brideprice payments and rates of fosterage. Each domain has a specific set of risks and benefits associated with it, leading to a deeper understanding of the impact of paternity on paternal care decisions. In particular, the domains vary in terms of their social visibility, which we predict will be particularly important to men's investment decisions. As the visibility of paternal care increases, its potential as a signal to mates or allies rises, as do the risks of reputational harm that would come from shirking investment expectations. This should lead to less titration in more public domains. For greater contextualization, we match our behavioural outcomes to a series of vignettes addressing men's perceptions of how paternity will affect paternal care, as well as their perceptions of how common norm violations are, and what the potential punishments are when violations occur.
2. Data and methods
(a). Study population
This study was part of a longitudinal project on paternity, parenting, and maternal and child health in Himba pastoralists. Household size ranges is 5–25 individuals, typically including three or more generations. Households are patrilocal, typically with a male head-of-household, his wives and children, and other relatives. While polygyny is common, female autonomy is high, and women may freely choose to divorce their husbands. All marriages are arranged through kin and brideprice is paid, although a large proportion of second marriages are love marriages with kin approval. Compared to many pastoralists in sub-Saharan Africa, Himba brideprice is fairly small (typically 1–2 cows and 1–2 small stock) and is almost always paid entirely by a single individual. Himba men typically give their children small gifts of livestock during adolescence, while the bulk of inheritance, in the form of cattle wealth, is passed matrilaterally, although shifts towards patrilineal wealth transfer may be under way [48]. Fosterage in this population is very common, and related to the needs and restrictions of the focal household as well as the households who care for fostered children [49]. Informal (non-marital) partnerships are common and not heavily stigmatized [50]. This is probably due to a myriad of factors, including a female-biased sex ratio of 0.714 [51], ecological conditions that necessitate spousal separation, and a cultural history of double descent and strong matrifocal ties. However, husbands are always recognized as the social father of children born to their wives. Children that are the result of affairs are referred to as omoka or omokati, reflecting the knowledge that the biological father is someone other than their social father. As with other pastoralists [52], Himba men provide little and infrequent direct care, but do participate in indirect care, including provisioning, investing in education and paying brideprice for their sons. However, relative to other pastoralists, almost all of whom practice patrilineal inheritance, resource transfers from fathers to children are minimal.
(b). Data collection
This study uses data from a 10-year project on the marital and family dynamics of Himba living in northwest Namibia. The measures of investment we use were therefore chosen in part because they were measures that were included in that larger study. However, the four investment outcomes (anthropometrics, brideprice, livestock gifts and fosterage decisions) relate to a broad spectrum of decisions about paternal care. This is relevant, because fathers have been shown to be important outside the traditional parenting arenas of provisioning and direct care [5,53]. In addition, some forms of care are more public than others, and the measures we chose represent both private and public investments, allowing us to investigate the role of social status and reputational impacts on investment decisions. The multiplex data used in this study mean that slightly different samples of men and children were included for each of the five analyses (norms study and four realms of investment). Electronic supplementary material, table S1 provides demographic information on each sub-sample.
(i). Determining paternity confidence
Paternity confidence was determined by asking men whether each of the children born to their wives was fathered by them (not omoka) or by someone else (omoka). To maximize sample size, where paternal assertions were unavailable, maternal assertions were used and where these were not available and the child was at least 16 years of age, self-assertions of omoka status were used. Here we rely on paternity assertions rather than genetic paternity, as it is the beliefs of men about paternity that are most relevant to investment. However, as these data are from the same set of households used in the genetic paternity study, which showed high accuracy in detecting nonpaternity events, we are confident that there is a strong link between paternity assertions and genetic paternity. Four domains of paternal care were measured, matched to the four vignettes presented in the norms study: child nutritional status, livestock gifts, brideprice (sons only) and fosterage rates. For all study participants, age was calculated using the local year name system [54].
(ii). Assessing norms of paternal investment
A convenience sample of Himba men (n = 16) answered a series of counterfactual vignettes, designed to illuminate the social norms surrounding biased paternal investment in this community. Each vignette described a scenario where a father biased investment against an omoka child. The vignettes addressed four domains of paternal care: (i) providing meat to a child after a ceremony, (ii) the amount of a livestock gift, (iii) the amount paid in brideprice for a son's marriage and (iv) the decision to foster out a child to the wife's family. After each vignette, men were presented with a set of comprehension questions and were then asked: (i) whether such a bias in investment was OK or not OK, (ii) whether that kind of bias was common and (iii) if it occurred what, if any, punishment would occur.
(iii). Child nutrition
To assess the influence of paternity confidence on child growth, anthropometrics collected between 2010 and 2018 were used. A digital SECA scale and fixed stadiometer were used for all measurements. To estimate the effects of paternity confidence on anthropometric variables, height, weight and BMI were predicted independently by sex, with age splines using all anthropometric data collected for individuals under 20 years of age (n > 600, to leverage the full longitudinal dataset). Using these fitted models, age-independent standardized residuals were calculated separately for males and females. Individual measurements were excluded if standardized residuals were greater than or less than three standard deviations from the mean, indicating measurement error. Finally, these residuals were then predicted using linear models from paternity assertions, sex and sex by paternity interactions. Varying intercepts by individual child were included to correct for repeated measures. Any children who were known to be fostered were excluded from the analysis, as this is known to have additional negative impacts on child health [49,55].
(iv). Livestock gifts
Himba fathers were asked for each of their children, the type and quantity of livestock gifts given. When fathers weren't interviewed, cattle gifts were collected from self-reports to increase sample size. Fostered children were excluded from this analysis, since fosterage may reduce the probability of receiving gifts from fathers. Livestock gifts were converted to tropical livestock units (TLU) using standard values [56], and hurdle-gamma models were used to predict TLU, with sex, standardized age (via a spline) and paternity predicting both binary and continuous outcome components of the model. To account for non-independence among fathers with multiple children, varying intercepts by father and slopes by sex and paternity were included in the final model.
(v). Brideprice
Amount of brideprice paid in TLU as reported through marriage histories was estimated with hurdle-gamma models. Varying intercepts were used to correct for repeated marital records by individual men and fixed effects for standardized age at marriage and paternity. An additional dummy variable was used to indicate whether the marriage in question was the first marriage, as brideprice is more standardized for first marriages and expectations that social fathers will pay are greater. All predictors were included in both the binary and continuous outcome components of the model.
(vi). Fosterage decisions
Data were collected on fosterage status for children and adults. Adults were asked if they were fostered and their relationship to their foster parent. Children's fosterage status was ascertained by interviews with foster parents and biological parents. Binomial regressions were used to assess the probability of fosterage, using sex, paternity and age. Generational effects of fosterage were expected across adults in the sample, and age effects in children were expected to be non-linear, so standardized age was fitted as a spline in all models. Children (less than 20 years of age) and adults (greater than or equal to 20 years of age) were also analysed individually (see electronic supplementary material, table S11).
(vii). Additional details on statistical models
Model comparisons were used to assess the impact of predictors on model fit. Initial models included only intercepts and relevant predictors, which were built upon by adding predictors for paternity certainty, sex and an interaction parameter. Models were fit to RStan in R v. 3.3.2 [57] using the brms package [58] with regularizing priors. Full models for each analysis and posterior summary statistics are in the electronic supplementary material. For each outcome, we sampled from the posterior for males and females, both omoka and non-omoka, and calculated the per cent difference in these posterior distributions (percentage of the posterior difference > 0), which indicate the size and direction of difference in posterior outcome. Where relevant we also calculate the posterior probabilities of a positive or negative effect for predictors (Pr[b > or < 0]). See electronic supplementary material for detailed model descriptions and posterior predictions.
3. Results
(a). Social norms about paternal investment
For all four domains of investment, there was strong consensus that preferentially investing in biological children was unacceptable (figure 1). However, respondents indicated that norm violations were relatively common. Punishments varied by investment domain, with verbal sanctions and reputational harm most commonly reported. Respondents also noted that more severe repercussions were possible, including removal of the child from the family (to be raised by maternal kin), and divorce.
Figure 1.

Counterfactual vignette responses.
(b). Paternity confidence and child anthropometrics
Overall, the effects of paternity confidence on nutritional status were modest, but exhibit a sex-specific pattern, with daughters more negatively affected than sons (electronic supplementary material, tables S2–S4). This occurs despite the general finding that sons are more nutritionally stressed than daughters, a result similar to previous analyses from this population, which have shown boys to be more likely to suffer from stunting, wasting and thinness [49,55], a common pattern in sub-Saharan Africa [59].
Results from a model comparison show nearly identical WAIC values across the three models, indicating that the addition of predictors does not improve out of sample deviance. Inclusion of paternity has little effect on models predicting height, but paternity has a modest negative effect on both weight and BMI residuals (Pr([b < 0) = 86.1% and 94.6%, respectively). Calculating posterior predictions from models including both paternity and sex indicates that girls, but not boys, have lower predicted weight and BMI when omoka (96.7% and 97.9% difference in posterior greater than 0, respectively, for girls; figure 2).
Figure 2.

Posterior predictions for anthropometrics: density plots illustrate the predicted difference in posteriors of omoka status by sex. Curves that fall further from zero demonstrate a larger effect of omoka status for that sex. Posteriors are derived from models that include sex, paternity and an interaction effect. Posterior mean and 95% prediction intervals shown. (Online version in colour.)
(c). Paternity confidence and livestock gifts
Models to predict the TLU of livestock gifts indicate little support for the prediction that omoka children receive fewer gifts (electronic supplementary material, table S5; figure 3). While boys tend to receive more livestock gifts from fathers generally, there is little difference in the predicted gifts within sex between omoka and non-omoka boys and girls, after correcting for individual father ID (49.3% and 59.6% difference in posterior greater than 0, respectively), or in models predicting reported gifts from adult men and women (56.2% and 66.7% difference in posterior greater than 0, respectively). Father's wealth was also included in the model, as this was expected to positively increase the size of gifts, but the inclusion of wealth did not increase model fit nor did it impact the probability of omoka children receiving livestock (see electronic supplementary material, tables S6 and S7).
Figure 3.

Posterior predictions for cattle gifts and brideprice: posterior predictions for livestock gifts separated by children and adults, and posterior predictions from the hurdle model for the father's payment of son's brideprice. Density plots illustrate the predicted difference in posteriors between omoka and non-omoka by sex for gifts, and in males only for brideprice. Posterior mean and 95% prediction intervals shown. (Online version in colour.)
(d). Paternity confidence and brideprice
Model results and comparisons show little evidence that paternity confidence biases the payment of brideprice by fathers. Inclusion of paternity assertions does not improve model fit, nor does the predictor impact posterior predictions (figure 3). Other relevant variables including standardized age at marriage and whether the marriage in question was a first marriage did predict increased brideprice payments (see electronic supplementary material, table S9).
(e). Paternity confidence and fosterage
Model predictions indicate a strong effect of paternity confidence on the probability of a child being fostered. In the model including only paternity confidence, posterior distributions of the predictor illustrate that, regardless of sex, omoka offspring are more likely to be fostered (Pr(b > 0) = 96.6%; see electronic supplementary material, table S10). However, in the best-fit model, where sex and a sex-by-paternity interaction are included, the effect of paternity confidence disappears. The full model illustrates that, for sons, but not daughters, low paternity confidence increases the probability of fosterage (99.9% and 34.6% difference in posterior less than 0, respectively; figure 4). Results were similar when adults and children were run in separate models, although prediction intervals are wider and overlap zero (see electronic supplementary material, table S11).
Figure 4.
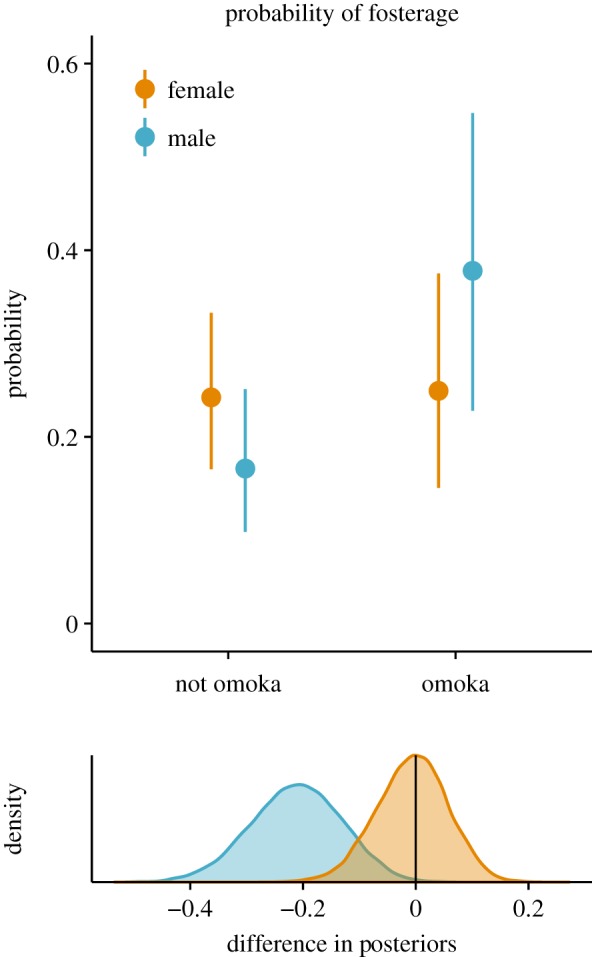
Posterior predictions for fosterage: posterior predictions for probability of being fostered. Density plots illustrate the predicted difference in posteriors of omoka – non-omoka by sex. Posteriors derived from models that include sex, paternity and an interaction effect. Posterior mean and 95% prediction intervals shown. (Online version in colour.)
4. Discussion
Despite expectations that men should bias investment against offspring of suspect paternity, our results demonstrate little inclination towards favouritism. Where we do see effects, they are sex- and domain-specific. This reflects the general finding in our vignette study that the titration of investment based on paternity is unacceptable in this population (figure 1), despite a very high risk of non-paternity. Data collected on social norms indicate that there can be social costs to biasing care, through both verbal and material sanctions, as well as reputational harm. Following this, we assess the patterns of paternal care and their relation to paternity confidence among Himba in the context of broader decisions that men are making about maintaining their households, their social networks and their romantic partnerships.
(a). Biased investment and the economic value of children
Where we see paternity confidence associated with investment outcomes in these data, the patterns are sex-specific. Daughters' nutritional status is negatively impacted when paternity confidence is low, and omoka sons are more likely to be fostered out. To understand these patterns, it is helpful to contextualize men's investment decisions within the framework of the household economy. Himba have a childhood sexual division of labour where girls tend to work harder and provide labour that is less substitutable. Additionally, sons and daughters have differential value to the family. Brideprice makes sons costly and daughters a source of revenue [35]. Among Himba, child marriage is common, meaning daughters can bring this revenue in from as young as 2–3 years of age. Coupled with their work in the household, daughters have value to their fathers from very early on. The value provided by Himba sons occurs much later in life, when they have the size and strength to contribute to herding and digging wells, and can contribute to the ritual maintenance of the herd and the compound.
The greater potential economic value of daughters' work may both help and harm them when it comes to paternal investment. When paternity confidence is low, girls may be coerced into working harder, or may choose to provide more labour, to compensate for being omoka. One of our informants explains one case of this type: ‘At first a man was treating the child nice, but then as she grew he realized the child wasn't his. He started treating her badly, but he didn't send her away. Now he likes her again because she is hard-working and she helps him so much.’ Sons' labour, on the other hand, is more invariable and not as energy-intensive until they reach adolescence, which means there are fewer opportunities for them to work harder as children, and less chance that extra work will impact their nutritional status. Previous work in this population illustrates a similar effect in the context of fosterage, which is also associated with increased demands, this time in order to provide labour for one's foster parent. Akin to these data, that study showed that fosterage status particularly impacts girls’ nutritional outcomes [55].
Relatedly, our results indicate that omoka boys, but not omoka girls, are more likely to be fostered out. Here, the economic value of daughters makes them valuable to their father's household, while sons' labour is less critical. By preferentially fostering out omoka boys, Himba men minimize the costs of misallocated investment while still reaping economic value from omoka girls. In addition, there are social benefits to be had by strengthening ties with kin via fosterage, further helping fathers to recoup any losses from investment provided to omoka children.
(b). Domain visibility and the social costs of investment bias
Unlike growth outcomes and fosterage decisions, paternal investments in livestock transfers, show no evidence of biased investment. Livestock transfers are highly visible to the community and are culturally resonant forms of wealth in pastoralist populations [60,61]. As such, biasing investment in these domains may be paradoxically costly to Himba men if they trigger sanctions or reputational harm. In several cases, men reported that if a man refused to pay brideprice for his son, another relative would step in, but with significant consequences for the father. One man reported, ‘The brother of the husband said to him, “If you don't pay the brideprice I will pay it, but from now on that son will be mine. Everything he needs I will provide for him.” Then his family will feel shame and people will think badly of the husband.’ The man saves himself the payment of a few stock, but could create ill will or later retributions, whose costs outweigh those initial savings.
In addition to the elevated costs of biased investment in visible domains, the importance of social fatherhood in Himba culture may further explain why we don't see predicted investment biases resulting from paternity uncertainty. The pronatalist view in Himba culture means that men reap reputational rewards by having many children, regardless of their paternity. One man states, ‘My wives here have boyfriends and have kids with them, and I care for those kids. When I die those children will wear the necklace showing that their father died … It is good for people to remember you.’ Another remarks, ‘They call me the father of [name redacted]. They call me with the name of my children.’
Finally, being generous towards children who are omoka can also benefit men's reproductive and political networks. Generosity has been reported as one of the most important traits that Himba women look for in a partner [45]. Providing for omoka children, particularly in more public arenas, can alert others to a man's prosocial tendencies. As one Himba man explains when asked why he provides so much for children he knows to be omoka: ‘The women are happy with a man who is generous. When the men are together for a meeting, the generous men are regarded as special. The chief is chosen based on his generosity. Even other men, like me, I am listened to more because I am generous.’ Other studies have shown similar links between paternal care and relationship maintenance in industrialized populations [16,62]. In this population, where a high level of paternity uncertainty is coupled with paternal care, benefits to men's reputation and relationships via provisioning are likely to be substantial.
(c). Limitations and future directions
This study is one of very few to map paternity assertions onto measures of investment, and the only one to do so in a population where paternity uncertainty is high. Contrary to predictions derived from the animal literature, titration of investment based on paternity certainty is relatively modest. By highlighting the dual social/biological nature of fatherhood, we illustrate the importance of social context to men's parenting decisions. In particular, social fatherhood introduces a set of expectations about care that can increase the costs of norm violation, regardless of suspected paternity. Likewise, benefits can accrue across domains, which outweigh the costs of what would otherwise be viewed as ‘misallocated' investment.
Further study bridging behavioural patterns of paternal care with an understanding of local norms and context would help to address some additional questions raised by these data. For example, despite strong continuity in Himba perceptions that men should not shirk their duties towards omoka children, their perceptions of which types of bias will be most prominent do not map onto our behavioural data very well. For example, livestock norm violations are reported to be common, but our data show no difference in giving to omoka and non-omoka children. This could be due to a more nuanced pattern than our data could reveal (e.g. goats may be easier to distribute unevenly than cattle) or could be the result of self-reports reiterating cultural norms instead of reflecting actual assessments [63]. Another possibility is that cultural change is causing norms about parenting and paternal roles to shift. For example, a recent study from the same population showed that a majority of men preferred patrilineal inheritance to the current system where cattle are transferred matrilaterally [48]. The same study also showed generational differences in views of women's autonomy, which is relevant in that it could lead to restrictions on concurrent partnerships, and more attention to paternity and stigmatization of omoka children. Follow-up studies could test between these possibilities.
The literature on paternity and investment could also be strengthened by additional work in a larger range of populations that capitalize on different types of paternal care. In particular, pastoralists like the Himba provide little direct care to offspring, and their level of paternal investment is generally low relative to populations using other subsistence strategies. Finally, while we measured norms for each domain of investment, we didn't measure norms related to sex-specific investment, which was revealed in analyses of the fosterage and anthropometric data.
5. Conclusion
Within evolutionary biology, there is a strong prediction that males will titrate care based on the certainty of paternity. However, in both the human and non-human literature, evidence for such titration is inconsistent. Complicating the matter in humans is the fact that most studies linking paternity and paternal care rely on proxy measures of paternity like physical resemblance. In addition, a western sampling bias means that most studies are from populations where extra-pair paternity is rare. Here we present evidence from a population with a high rate of extra-pair paternity, and thus where decisions about paternal care are highly likely to be affected by paternity confidence, and yet we find very little evidence of titration. Instead, men appear to be making strategic decisions about whether to bias care. These decisions balance the potential loss of resources diverted to non-biological offspring with the reputational benefits of providing care. Biases also reflect complex household dynamics that affects sons and daughters differently. These data point to the need for studies of paternal care in a wider range of societies and call for more nuanced studies that consider the role of paternity alongside other drivers of men's decisions.
Supplementary Material
Acknowledgements
We would like to thank the community of Omuhonga for their continued support and to the individuals who participated in the study. John Jakurama, Kenanbangu Ngombe and Cancy Louis acted as research assistants and translators in Namibia. Members of the Behavior, Evolution and Culture (BEC) group at UCLA provided valuable feedback to this manuscript.
Ethics
Human subject permissions were issued by the UCLA Office of the Human Research Protection Program.
Data accessibility
Data and code associated with this manuscript can be accessed at https://osf.io/bjkn8.
Authors' contributions
Authors contributed equally to this manuscript.
Competing interests
We declare we have no competing interests.
Funding
This work was funded by the National Science Foundation (BCS-1534682).
References
- 1.Queller DC. 1997. Why do females care more than males? Proc. R. Soc. Lond B 264, 1555–1557. ( 10.1098/rspb.1997.0216) [DOI] [Google Scholar]
- 2.Trivers RL. 1972. Parental investment and sexual selection. In Sexual selection & the descent of man (ed. Campbell B.), pp. 136–179. New York, NY: Aldine de Gruyter. [Google Scholar]
- 3.Geary D. 2000. Evolution and proximate expression of human paternal investment. Psychol. Bull. 126, 55–77. ( 10.1037/0033-2909.126.1.55) [DOI] [PubMed] [Google Scholar]
- 4.Jones NGB, Marlowe FW, Hawkes K, O'Connell JF, Marlowe FW, Hawkes K, O'Connell JF. 2000. Paternal investment and hunter-gatherer divorce rates. In Adaptation and human behavior (eds Cronk L, Chagnon N, Irons W), pp. 49–68. New York, NY: Walter de Gruyter. [Google Scholar]
- 5.Scelza BA. 2010. Fathers' presence speeds the social and reproductive careers of sons. Curr. Anthropol. 51, 295–303. ( 10.1086/651051) [DOI] [Google Scholar]
- 6.Kempenaers B, Sheldon BC. 1997. Studying paternity and paternal care: pitfalls and problems. Anim. Behav. 53, 423–427. ( 10.1006/anbe.1996.0377) [DOI] [Google Scholar]
- 7.Sheldon BC. 2002. Relating paternity to paternal care. Phil. Trans. R. Soc. Lond. B 357, 341–350. ( 10.1098/rstb.2001.0931) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Møller AP, Thornhill R. 1998. Male parental care, differential parental investment by females and sexual selection. Anim. Behav. 55, 1507–1515. ( 10.1006/anbe.1998.0731) [DOI] [PubMed] [Google Scholar]
- 9.Møller AP, Birkhead TR. 1993. Certainty of paternity covaries with paternal care in birds. Behav. Ecol. Sociobiol. 33, 261–268. ( 10.1007/BF02027123) [DOI] [Google Scholar]
- 10.Schwagmeyer PL, Clair RCS, Moodie JD, Lamey TC, Schnell GD, Moodie MN. 1999. Species differences in male parental care in birds: a reexamination of correlates with paternity. Auk 116, 487–503. ( 10.2307/4089381) [DOI] [Google Scholar]
- 11.Dixon A, Ross D, O'Malley SLC, Burke T. 1994. Paternal investment inversely related to degree of extra-pair paternity in the reed bunting. Nature 371, 698–700. ( 10.1038/371698a0) [DOI] [Google Scholar]
- 12.Neff BD. 2003. Decisions about parental care in response to perceived paternity. Nature 422, 716–719. ( 10.1038/nature01528) [DOI] [PubMed] [Google Scholar]
- 13.Sheldon BC, Ellegren H. 1998. Paternal effort related to experimentally manipulated paternity of male collared flycatchers. Proc. R. Soc. Lond. B 265, 1737–1742. ( 10.1098/rspb.1998.0496) [DOI] [Google Scholar]
- 14.Alvergne A, Faurie C, Raymond M. 2009. Father–offspring resemblance predicts paternal investment in humans. Anim. Behav. 78, 61–69. ( 10.1016/j.anbehav.2009.03.019) [DOI] [Google Scholar]
- 15.Alvergne A, Faurie C, Raymond M. 2010. Are parents' perceptions of offspring facial resemblance consistent with actual resemblance? Effects on parental investment. Evol. Hum. Behav. 31, 7–15. ( 10.1016/j.evolhumbehav.2009.09.002) [DOI] [Google Scholar]
- 16.Apicella CL, Marlowe FW. 2004. Perceived mate fidelity and paternal resemblance predict men's investment in children. Evol. Hum. Behav. 25, 371–378. ( 10.1016/j.evolhumbehav.2004.06.003) [DOI] [Google Scholar]
- 17.Billingsley J, Antfolk J, Santtila P, Lieberman D. 2018. Cues to paternity: do partner fidelity and offspring resemblance predict daughter-directed sexual aversions? Evol. Hum Behav. 39, 290–299. ( 10.1016/j.evolhumbehav.2018.02.001) [DOI] [Google Scholar]
- 18.Heijkoop M, Dubas JS, van Aken MAG. 2009. Parent–child resemblance and kin investment: Physical resemblance or personality similarity? Eur. J. Dev. Psychol. 6, 64–69. ( 10.1080/17405620802642306) [DOI] [Google Scholar]
- 19.Prokop P, Obertová Z, Fedor P. 2010. Paternity cues and mating opportunities: what makes fathers good? Acta Ethol. 13, 101–107. ( 10.1007/s10211-010-0079-0) [DOI] [Google Scholar]
- 20.Tracey MR, Polachek SW. 2018. If looks could heal: Child health and paternal investment. J. Health Econ. 57, 179–190. ( 10.1016/j.jhealeco.2017.11.007) [DOI] [PubMed] [Google Scholar]
- 21.Yu Q, Zhang Q, Xiong Q, Jin S, Zou H, Guo Y. 2019. The more similar, the more warmth: the effect of parent–child perceived facial resemblance on parenting behavior. Pers. Individ. Differ. 138, 358–362. ( 10.1016/j.paid.2018.10.027) [DOI] [Google Scholar]
- 22.Yu Q, Zhang Q, Jin S, Chen J, Shi Y, Li Y. 2017. The effect of perceived facial resemblance on parent-child relationship. Pers. Individ. Differ. 116, 359–365. ( 10.1016/j.paid.2017.05.016) [DOI] [Google Scholar]
- 23.Chang L, Lu HJ, Lee LNY, Li T, Sui SS. 2010. Patrilocal residence and father–child resemblance beliefs in relation to paternal investment. Parenting 10, 274–285. ( 10.1080/15295192.2010.492039) [DOI] [Google Scholar]
- 24.Alvergne A, Perreau F, Mazur A, Mueller U, Raymond M. 2014. Identification of visual paternity cues in humans. Biol. Lett. 10, 20140063 ( 10.1098/rsbl.2014.0063) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Platek SM, Kemp SM. 2009. Is family special to the brain? An event-related fMRI study of familiar, familial, and self-face recognition. Neuropsychologia 47, 849–858. ( 10.1016/j.neuropsychologia.2008.12.027) [DOI] [PubMed] [Google Scholar]
- 26.McLain DK, Setters D, Moulton MP, Pratt AE. 2000. Ascription of resemblance of newborns by parents and nonrelatives. Evol. Hum. Behav. 21, 11–23. ( 10.1016/S1090-5138(99)00029-X) [DOI] [Google Scholar]
- 27.Regalski JM, Gaulin SJC. 1993. Whom are Mexican infants said to resemble? Monitoring and fostering paternal confidence in the Yucatan. Ethol. Sociobiol. 14, 97–113. ( 10.1016/0162-3095(93)90010-F) [DOI] [Google Scholar]
- 28.Dubas JS, Heijkoop M, van Aken MAG. 2009. A preliminary investigation of parent–progeny olfactory recognition and parental investment. Hum. Nat. 20, 80–92. ( 10.1007/s12110-009-9058-4) [DOI] [Google Scholar]
- 29.van de Mortel T. 2008. Faking it: social desirability response bias in self-report research. Aust. J. Adv. Nurs. 25, 40–48. [Google Scholar]
- 30.Anderson KG. 2006. How well does paternity confidence match actual paternity? evidence from worldwide nonpaternity rates. Curr. Anthropol. 47, 513–520. ( 10.1086/504167) [DOI] [Google Scholar]
- 31.Anderson KG, Kaplan H, Lancaster JB. 2007. Confidence of paternity, divorce, and investment in children by Albuquerque men. Evol. Hum. Behav. 28, 1–10. ( 10.1016/j.evolhumbehav.2006.06.004) [DOI] [Google Scholar]
- 32.Bledsoe C. 1980. The manipulation of Kpelle social fatherhood. Ethnology 19, 29–45. ( 10.2307/3773318) [DOI] [Google Scholar]
- 33.Isiugo-Abanihe UC. 1994. Reproductive motivation and family-size preferences among Nigerian men. Stud. Fam. Plann. 25, 149–161. ( 10.2307/2137941) [DOI] [PubMed] [Google Scholar]
- 34.Mace R. 1996. Biased parental investment and reproductive success in Gabbra pastoralists. Behav. Ecol. Sociobiol. 38, 75–81. ( 10.1007/s002650050219) [DOI] [PubMed] [Google Scholar]
- 35.Mulder MB. 1998. Brothers and sisters. Hum. Nat. 9, 119–161. ( 10.1007/s12110-998-1001-6) [DOI] [PubMed] [Google Scholar]
- 36.Bird RB, Ready E, Power EA. 2018. The social significance of subtle signals. Nat. Hum. Behav. 2, 452–457. ( 10.1038/s41562-018-0298-3) [DOI] [PubMed] [Google Scholar]
- 37.Barker JL, Power EA, Heap S, Puurtinen M, Sosis R. 2019. Content, cost, and context: a framework for understanding human signaling systems. Evol. Anthropol. 28, 86–99. ( 10.1002/evan.21768) [DOI] [PubMed] [Google Scholar]
- 38.Hamilton WD. 1964. The genetical evolution of social behaviour. II. J. Theor. Biol. 7, 17–52. ( 10.1016/0022-5193(64)90039-6) [DOI] [PubMed] [Google Scholar]
- 39.Kokko H, Jennions MD. 2008. Parental investment, sexual selection and sex ratios. J. Evol. Biol. 21, 919–948. ( 10.1111/j.1420-9101.2008.01540.x) [DOI] [PubMed] [Google Scholar]
- 40.Marlowe F. 1999. Male care and mating effort among Hadza foragers. Behav. Ecol. Sociobiol. 46, 57–64. ( 10.1007/s002650050592) [DOI] [Google Scholar]
- 41.Stiver KA, Alonzo SH. 2009. Parental and mating effort: is there necessarily a trade-off? Ethology 115, 1101–1126. ( 10.1111/j.1439-0310.2009.01707.x) [DOI] [Google Scholar]
- 42.Coleman JS. 1988. Social capital in the creation of human capital. AJS 94, S95–S120. ( 10.1086/228943) [DOI] [Google Scholar]
- 43.Caldwell JC, Orubuloye IO, Caldwell P. 1991. The destabilization of the traditional Yoruba sexual system. Popul. Dev. Rev. 17, 229–262. ( 10.2307/1973730) [DOI] [Google Scholar]
- 44.Schoen R, Kim YJ, Nathanson CA, Fields J, Astone NM. 1997. Why do Americans want children? Popul. Dev. Rev. 23, 333–358. ( 10.2307/2137548) [DOI] [Google Scholar]
- 45.Scelza BA, Prall SP. 2018. Partner preferences in the context of concurrency: what Himba want in formal and informal partners. Evol. Hum. Behav. 39, 212–219. ( 10.1016/j.evolhumbehav.2017.12.005) [DOI] [Google Scholar]
- 46.Wolputte SV. 2016. Sex in troubled times: moral panic, polyamory and freedom in north-west Namibia. Anthropol. Southern Africa 39, 31–45. ( 10.1080/23323256.2016.1147967) [DOI] [Google Scholar]
- 47.Scelza BA, Prall SP, Swinford N, Gopalan S, Atkinson E, McElreath R, Sheehama J, Henn BM. 2020. High rate of extra-pair paternity in a human population demonstrates diversity in human reproductive strategies. Sci. Adv. 6, eaay6195 ( 10.1126/sciadv.aay6195) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Scelza BA, Prall SP, Levine NE. 2019. The disequilibrium of double descent: changing inheritance norms among Himba pastoralists. Phil. Trans. R. Soc. B 374, 20180072 ( 10.1098/rstb.2018.0072) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Scelza BA, Silk JB. 2014. Fosterage as a system of dispersed cooperative breeding. Hum. Nat. 25, 448–464. ( 10.1007/s12110-014-9211-6) [DOI] [PubMed] [Google Scholar]
- 50.Scelza BA. 2011. Female choice and extra-pair paternity in a traditional human population. Biol. Lett. 7, 889–891. ( 10.1098/rsbl.2011.0478) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Scelza BA, et al. 2020. Patterns of paternal investment predict cross-cultural variation in jealous response. Nat. Hum. Behav. 4, 20–26. ( 10.1038/s41562-019-0654-y) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Marlowe F. 2000. Paternal investment and the human mating system. Behav. Processes 51, 45–61. ( 10.1016/S0376-6357(00)00118-2) [DOI] [PubMed] [Google Scholar]
- 53.Gray P, Anderson KG. 2010. Fatherhood: evolution and human paternal behavior. Cambridge, MA: Harvard University Press. [Google Scholar]
- 54.Scelza BA. 2011. Female mobility and postmarital kin access in a patrilocal society. Hum. Nat. 22, 377–393. ( 10.1007/s12110-011-9125-5) [DOI] [PubMed] [Google Scholar]
- 55.Prall SP, Scelza BA. 2017. Child fosterage and sex-biased nutritional outcomes among Namibian pastoralists. Am. J. Hum. Biol. 29, e23058 ( 10.1002/ajhb.23058) [DOI] [PubMed] [Google Scholar]
- 56.Bollig M. 2010. Risk management in a hazardous environment: a comparative study of two pastoral societies. Berlin, Germany: Springer Science & Business Media. [Google Scholar]
- 57.Stan Development Team. 2019. RStan: the R interface to Stan. See http://mc-stan.org/.
- 58.Bürkner P-C. 2017. brms: an R package for bayesian multilevel models using Stan. J. Stat. Softw. 80, 1–28. ( 10.18637/jss.v080.i01) [DOI] [Google Scholar]
- 59.Wamani H, Åstrøm AN, Peterson S, Tumwine JK, Tylleskär T. 2007. Boys are more stunted than girls in Sub-Saharan Africa: a meta-analysis of 16 demographic and health surveys. BMC Pediatr. 7, 17 ( 10.1186/1471-2431-7-17) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mulder MB, Fazzio I, Irons W, McElreath RL, Bowles S, Bell A, Hertz T, Hazzah L. 2010. Pastoralism and wealth inequality: revisiting an old question. Curr. Anthropol. 51, 35–48. ( 10.1086/648561) [DOI] [Google Scholar]
- 61.Hutchinson S. 1992. The cattle of money and the cattle of girls among the Nuer, 1930–83. Am. Ethnol. 19, 294–316. ( 10.1525/ae.1992.19.2.02a00060) [DOI] [Google Scholar]
- 62.Anderson KG, Kaplan H, Lancaster J. 1999. Paternal care by genetic fathers and stepfathers I: reports from Albuquerque Men. Evol. Hum. Behav. 20, 405–431. ( 10.1016/S1090-5138(99)00023-9) [DOI] [Google Scholar]
- 63.Du J, Mace R. 2018. Parental investment in Tibetan populations does not reflect stated cultural norms. Behav. Ecol. 29, 106–116. ( 10.1093/beheco/arx134) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data and code associated with this manuscript can be accessed at https://osf.io/bjkn8.