

Abstract
Infections may cause mortality in old age due to damaged immune responses. As zinc is required as a catalyst, structural (zinc fingers) and regulatory ion, it is involved in many biological functions, including immune responses. Low zinc ion bioavailability and impaired cell-mediated immunity are common in ageing and may be restored by physiological supplementation with zinc for 1–2 months, impacting upon morbidity and survival. This article reviews the role of zinc in immune efficacy during ageing, and also describes the main biochemical pathways involved in the role of zinc in resistance to infections in ageing in order to better understand the possible causes of immunosenescence.
Keywords: Zinc, Infections, Immunosenescence
1. Introduction
Infections disease is one of the major causes of mortality in elderly populations because of decreases in cell-mediated immune responses (Pawelec and Solana, 1997). Zinc may be crucial in this respect, because of its requirement as a catalytic component of more than 300 enzymes, as a structural constituent of many proteins, and as a regulatory ion for the stability of proteins and in preventing free radical formation (Mills, 1989). This has led to recognition of zinc as a leading element in assuring correct functioning of various tissues, organs and systems, including immune responses (Wellinghausen et al., 1997 Mocchegiani et al., 1998a). In the role of catalyst, zinc polarises the substrate activating H2O molecules, which then act as a nucleophile. Zinc ions act as a template to bring together the substrate and the nucleophile, thereby accelerating the conversion of substrate to product (Vallee and Falchuk, 1993). In a regulatory role, zinc determines the balance between gene expression of metalloproteinases (MMPs) and tissue inhibitors of matrix metalloproteinases (TIMPs) (Nagase, 1996) as well as all phases of the cell cycle by means of cyclin/CDK complex gene expression (Kilbey et al., 1999). In particular, zinc is most important in the G0/G1 phase. High doses of zinc (900 μM) arrest the cell cycle (Paramanantham et al., 1996) whereas, low doses (150 μM) inhibit apoptosis in ageing in the G0/G1 phase (Provinciali et al., 1998). This last point is crucial for ageing and infections because their immune decrement is likely due to low zinc ions bioavailability with subsequent immune cell-death (Shankar and Prasad, 1998).
However, the main task of zinc is in a structural role because of the involvement of zinc finger, DNA domains of many proteins, peptides, enzymes, hormones, transcriptional factors and growth factors, including cytokines, relevant to the maintenance of body homeostatic mechanisms affecting, as such, gene transcription (Coleman, 1992, Berg and Shi, 1996, Rink and Gabriel, in press). Zinc also regulates gene expression by affecting mRNA stability (Taylor and Blackshear, 1995) and by maintaining the extracellular matrix (Vallee and Falchuk, 1993). Moreover, zinc binds enzymes, proteins and peptides with different affinity (kd) ranging from 10−2 to 10−14 mol/l (Mocchegiani et al., 1998a), including a thymic hormone called thymulin (kd = 5.0 ± 2 × l0−7 mol/l) (Dardenne et al., 1982). The compounds are inactive or with low biological activity when zinc-binding does not occur (Mills, 1989). Zinc also plays a critical role in the structure, function, stabilisation and fluidity of biomembranes because of zinc binding to sulphydryl groups forming mercaptides (Bettger and O'Dell, 1993). It is of interest, that zinc is also required to maintain optimal functional and enzymatic activity of inducible nitric-oxide synthase (iNOS) with consequent nitric oxide (NO) production, in turn, relevant in immune responses and cytokine activity (Bogdan et al., 2000). In fact, zinc is bound to two cysteine residues: part of crystal structures of the heme domain of iNOS maintaining integrity of the cofactor (tetrahydrobiopterin) binding site of iNOS (Li et al., 1999). Because: i) NO is involved in metallothioneins (MTs) mRNA expression in the liver and hence protection against oxidative stress during inflammation (Arizonol et al., 1995) and ii) NO prevents cell-death activating a nuclear enzyme, poly (ADP-ribose) polymerase (PARP), which in turn requires two zinc finger motifs (Trucco et al., 1996), the structural task of zinc in NO production is crucial. In addition, NO is involved in zinc release from MTs to antioxidant enzymes (Maret et al., 1999).
Pro-inflammatory cytokines (IL-1 and IL-6) affect MT mRNA (Cousins and Leinart, 1988). IL-1, IL-6, MTs, NO are increased during ageing and infections, whereas, zinc and immune efficacy are decreased (Mocchegiani et al., 1998a, Capanni et al., 1998) with consequent immune cell-death by activating PARP (Szabo and Dowson, 1998, Pieper et al., 1999). Since infections are common event in elderly (Pawelec and Solana, 1997), subsequent mortality may occur because of the poor efficacy of conventional therapies in elderly populations (Mocchegiani et al., 1999b). Therefore, increasing zinc bioavailability by supplementing zinc may be of benefit for immunoresistance to infections in ageing involving a new mechanism of homeostasis among pro-inflammatory cytokines, MTs, NO and PARP (as all these factors are zinc-dependent and implicated in immune efficiency (Mocchegiani et al., 2000b)).
This review considers the efficacy of zinc supply in immune responses during ageing and accelerated ageing (e.g., Down's syndrome), as well as the main biochemical pathways of zinc, including MTs-NO-PARP homeostasis, as crucial to immunoresistance to infections in ageing. An emphasis on cytokines in ageing is described because this is altered in ageing (Rink et al., 1998) and is involved in many biochemical pathways of zinc immunobiology (Wellinghausen et al., 1997, Rink et al., 1998).
2. Zinc, infections and ageing: the role of cytokines
Many data support the view that the impact of zinc on immunocompetence is greater in cell-mediated immunity than humoral (Wellinghausen et al., 1997). In particular, chemotaxis by neutrophils and monocytes, thymic endocrine activity, antigen presentation by MHC class II molecules, natural killer (NK) activity, cytokine production and the Th1/Th2 balance are the immune functions most affected by zinc (Prasad, 1998), as well as Th3 cells (Mocchegiani et al., 2000b). In this context, cytokine production is peculiar, because of their different synthesis by Th1 or Th2 or Th3 cells. According to their function in cell-mediated immunity (Th1), in antibody mediated immunity (Th2) and in suppression (Th3), Th1 cells produce IL-2, IFN-gamma and TNF-alpha, Th2 cells produce IL-4, IL-5, IL-6, IL-10 and IL-13 (Romagnani, 1997), whereas, Th3 cells produce TGF-beta (Cobbold and Waldman, 1998). Zinc may more directly affect Th1 cells, rather than Th2 cells, by means of MHC-complexes, or by thymic hormones or by immunomodulating hormones (melatonin, thyroid hormones, IGF-I or growth hormone) (Mocchegiani et al., 1998a, Prasad, 1998), whereas zinc affects Th3 cells by zinc finger DNA domains, such as Egr-1 (Yoo et al., 1996). Although a specific role of Th3 has not been reported in zinc deficiency, an increment of TGF-beta is observed in infections (Fitzpatrick and Bielefeldt-Ohmann, 1999) as well as in ageing related to inflammation (McCaffrey, 2000), and an imbalance of Th1/Th2 activity towards Th2 is usual in ageing and infections both characterised by zinc deficiency (Prasad, 1998). Therefore a Th1/Th2 imbalance may also have to include consideration of the Th3 cells as well, as recently suggested in infections (Cobbold and Waldman, 1998). In vivo and in vitro experiments in old (Frasca et al., 1997) and infected mice (Salem et al., 1999) show that the imbalance of Th1/Th2 cytokines and TGF-beta production by Th3 cells is under the control of IL-12 produced by macrophages and resulting in decreased resistance to infections. In fact, IL-12 synthesis, which is required for Th1 development, is suppressed by increased IL-4 and TGF-beta in infected mice (Salem et al., 1999, Fresno et al., 1997) and Th1 cells (Rink et al., 1998) and IL-12 (Fagiolo et al., 1993, Lio et al., 1998) are decreased in ageing. On the other hand, activated TGF-beta has an important effect on pathogen entry, replication, persistence and latency (Fitzpatrick and Bielefeldt-Ohmann, 1999). Because of zinc affecting IL-12 production (Driessen et al., 1994), the balance of Th1/Th2 cells with also Th3 for the immunoresistance to infections may be controlled by zinc turnover, the alteration of which may cause decreased resistance to infections in old age with consequent high mortality. Supplementing zinc may be of benefit to restore the Th1/Th2 balance in ageing and infections, as documented in elderly people (Cakman et al., 1997) and in nematode-infected mice (Shi et al., 1997). Although no data on the effects of zinc supplementation on TGF-beta are available, indirect evidence showing resistance to listeriosis in mice by an increment of IL-12, TNF-alpha and IFN-gamma by Th1 and by a decrement of TGF-beta after anti-CD1 mAb treatment (Szalay et al., 1999), strongly support the possible involvement of zinc turnover in immunoresistance, because all these cytokines are directly or indirectly under the control of zinc turnover (Fabris et al., 1997).
3. Therapeutic use of zinc in infections during ageing
The relevant role played by zinc deficiency in immune decrements during infections and ageing is well documented during the last decade (Wellinghausen et al., 1997 Prasad, 1998, Mocchegiani et al., 1998a). However, there are dispersions of data due to the great variability in habitat conditions, in health status and individual dietary attitudes (Fabris and Mocchegiani, 1995). This leads to considering zinc deficiency related more to the single individual's particular status rather than to the general effects of alterations of zinc-dependent factors with subsequent possible low zinc ion bioavailability for immune responses. Indeed, some investigators have reported normal zinc content both in the circulation and in lymphocytes during ageing (Prasad et al., 1993). Therefore, supplementing zinc is not recommended by them. This is misleading because zinc is bound to many enzymes and proteins (Mills, 1989). Since atomic absorption spectrophotometry (AAS) assesses bound and unbound zinc, the real zinc ion bioavailability is not evident when using this technique (Mocchegiani et al., 1998a). Although ecto-5 nucleotidase may be useful to examine zinc deficiency (Prasad et al., 1993), specific methodological procedures to detect free zinc ions in plasma and tissues are not available, because zinc binds to more than 300 enzymes. We have developed a methodological procedure for testing active zinc-bound thymulin (ZnFTS) (AT) and total thymulin (TT) (active zinc-bound ZnFTS + inactive zinc-unbound FTS) after in vitro zinc addition to plasma samples. The ratio between TT/AT is the non-saturable fraction of thymulin (FTS) by zinc ions. Because of the existence of a strict inverse correlation between plasma zinc levels and the ratio itself, the latter is a good marker to detect zinc ion bioavailability and, consequently, the real zinc deficiency. The following cut-off points were established: (i) TT/AT > 2 (log−2) = marked zinc deficiency; (ii) TT/AT < 2 (log−2) = mild zinc deficiency; (iii) TT/AT = 1 (log−2): normal zinc values (Fabris et al., 1984). This methodological procedure has led us to identify low zinc ion bioavailability in ageing and in various pathologies despite plasma zinc levels in the normal range (Table 1 ). This also led us to seek the possible biological causes of low zinc ion bioavailability in ageing. Zinc-binding proteins, such as the metallothioneins (MT), are involved due to their higher binding affinity (kd = l0−13 M) for zinc compared to thymulin (10−7 M) (see Mocchegiani et al., 1998a). However, this does not exclude other zinc-binding proteins with higher kd, such as α − 2 macroglobulin (10−10 M), from being increased in ageing and inflammation (Cakman et al., 1996). MT bind preferentially to zinc rather than copper in ageing (Hamer, 1986) and more zinc ions bound to MT in old liver than young (E. Mocchegiani, unpublished results). This last recent finding by our laboratory strongly suggest MTs as sequesters of zinc in constant stress, such as in ageing, inducing low zinc ion bioavailability for normal immune responses (Mocchegiani et al., 1998a). As such, MT may move from having a protection in young-adults to a dangerous one for immune responses in the aged (Mocchegiani et al., 1997, Mocchegiani et al., 1998a). On the other hand, MT mRNA is increased in ageing (Mocchegiani et al., 1997) associated with impaired immune functions (NK activity) also during the circadian cycle (Mocchegiani et al., 2000a). Therefore, high zinc-bound MT is deleterious for immune efficiency in ageing. Because of no significant, further increments of already high liver MT levels by zinc in old mice (Mocchegiani et al., in press), zinc supplementation can be recommended in ageing in order to keep the immune system more efficient by means of major zinc ion bioavailability. Subsequent limited appearance of infections or relapses in elderly population may occur. Indeed low zinc bioavailability is a risk factor for immune deficiency and subsequent infection relapses in elderly population (Mocchegiani et al., 1999b). On the other hand, high MT levels are present in lymphocytes of old people (Yurkow and Makhijani, 1998) suggesting also the measure of zinc content in lymphocytes may be misleading in testing zinc ion bioavailability. Following these considerations physiological supplementation of zinc (RDA) (USDA, 1976) was carried out in ageing and in syndromes of accelerated ageing (Down's syndrome) for short periods, even knowing the possible harmful effect of zinc toxicity in immune responses (Chandra, 1984). However, there is no general consensus for supplementing zinc in Down's syndrome (DS) and in physiological ageing because contradictory data exist in real zinc deficiency. Indeed, some authors have reported normal or slightly reduced zinc content in lymphocytes of Down's syndrome subjects (Lockitch et al., 1989) and in the elderly population (Bodgen et al., 1990) with modest effects on immune efficiency. This may also be misleading, because, despite plasma zinc levels in the normal range for age in DS and ageing (Fabris et al., 1984), the TT/AT ratio is >2 (log−2) (Table 1) suggesting the presence of low zinc ion bioavailability in both conditions. In agreement with others (Stabile et al., 1991, Lesourd, 1997), physiological supplementing zinc (lozenge of 12 μg Zn++/day) for one or two months in elderly populations and in DS restores immune responses, and decreases oxidative damage with no modifications to copper levels (Fabris et al., 1993). Reductions (more than 50%) of infectious episodes in DS and elderly subjects occur after zinc treatment with maintenance of good health and better cognitive performance also after 4 months and 1 yr of follow-up (Fabris et al., 1993, Johnson and Porter, 1997, Fortes et al., 1998, Mocchegiani et al., 1999b). Similarly, physiological zinc treatment (18 μg Zn++/day) in drinking water in presenescent mice (from the age of 12 months) restores immune responses with reduction of mortality (35%) by infections of mycoplasma pulmonis or coronavirus murini with consequent significant prolonged survival as compared to normal mice (Mocchegiani et al., 1998b, Mocchegiani et al., 2000c). Moreover, supplementing zinc restores the Th1/Th2 balance in an elderly population (Cakman et al., 1997). It is also of interest to note that supplementing zinc is of benefit in restoring immune efficiency and subsequent significant reductions of infections relapses also in young-adult people affected by various pathological conditions (Table 2 ). Therefore, zinc is crucial to maintain normal immune efficiency against infections over the entire life span. As a consequence, the lack of zinc ion bioavailability is pivotal to better understand the possible causes of immunosenescence.
Table 1.
TT/AT ratio and plasma zinc levels in the elderly population and in various conditionsa
| Condition | Age (range years) | TT/AT (range log−2) | Plasma zinc levels (range μg/dl) | r |
|---|---|---|---|---|
| Elderly population | 60–83 | 2–3 | 80–95 | −0.77 (P < 0.01) |
| Alzheimer's Dementia | 70–75 | 2–3 | 80–90 | −0.67 (P < 0.05) |
| AIDS | 20–35 | 5–6 | 77–95 | −0.82 (P < 0.01) |
| Down's syndrome | 8–25 | 4–5 | 85–120 | −0.85 (P < 0.01) |
| Cystic fibrosis | 2–13 | 4–5 | 65–89 | −0.87 (P < 0.01) |
| Diabetes juvenile type I | 20–37 | 3–4 | 90–120 | −0.80 (P < 0.01) |
| Head traumatic injury | 18–30 | 3–4 | 90–110 | −0.78 (P < 0.01) |
| Premature infants | 1–2 (months) | 3–4 | 100–120 | −0.77 (P < 0.01) |
| Allergy (Asthma and atropy) | 3–15 | 1–2 | 110–130 | −0.45 (not significant) |
| Solid tumours (lung, colon, cervix, prostate) | 35–55 | 4–5 | 85–95 | −0.82 (P < 0.01) |
| Leukemia (ALL) | 2–14 | 4–5 | 80–120 | −0.68 (P < 0.05) |
| Chronic renal failure | 37–46 | 3–4 | 80–100 | −0.70 (P < 0.05) |
| Iperprolactinemia | 30–42 | 2–3 | 88–97 | −0.73 (P < 0.05) |
| Crohn's Disease | 40–55 | 2–3 | 85–95 | −0.75 (P < 0.05) |
| Dwarfism (children) | 5–15 | 2–3 | 87–100 | −0.78 (P < 0.01) |
| Dwarfism (adult) | 27–45 | 2–3 | 80–95 | −0.66 (P < 0.05) |
Normal levels of plasma zinc (range) for age (Immunology Ctr. INRCA, Ancona, Italy): 1–20 yr=120–150 μg/dl; 21–40 yr=95–120 μg/dl; 41–60 yr=80–95 μg/dl; 61–80 yr=80–87 μg/dl; >80 yr=70–80 μg/dl. Coefficient of inverse correlation (r) between TT/AT and plasma zinc levels for single condition was calculated also adding data of respective age-matched healthy controls or young-adult healthy people for elderly people (Fabris and Mocchegiani, 1995). Range TT/AT=1–1.5 (log−2) in healthy controls. (Fabris et al., 1984). Normal zinc ion bioavailability exists in allergy.
Table 2.
Benefit of physiological zinc (RDA) as resistance to infections in humansa
| Time of treatment | Condition | % of infections reduction |
|---|---|---|
| 2 months | Down's syndrome (Bronchitis, Gastrointestinal infections) | 50 |
| 2 months | Malnutrition (Gastrointestinal infections) | 35 |
| 2 months | Acrodermatitis enteropathica (Skin lesions, Gastrointestinal infections) | 38 |
| 1 month | Congenital herpes (herpes simplex II) | 50 |
| 2 months | Leprosy | 40 |
| 2 months | Malaria | 35 |
| 2 months | Chronic diarrhea (Salmonella) | 30 |
| 1 month | Acute lower respiratory infections (Bronchitis) | 35 |
| 2 months | Burns (Skin lesions, Bronchopneumonia) | 30 |
| 1 month | AIDS (Candida albicans, Pneumocistis carinii, Toxoplasma gondii, Cytomegalovirus, Criptococcus, Salmonella, Mycobacterium tuberculosis) | 55 |
| 1–3 months | Elderly population (Chronic Bronchitis Obstructive, Bronchopneumonia) | 30 |
| 1 year | Sickle cell disease (Bronchitis and Urinary tract infections) | 50 |
| 3 weeks | Pregnancy (Bacterial Vaginosis) | 53 |
| 1 month | Preschool children (Malaria) | 55 |
| 2 months | Common cold (cough, fever, respiratory tract secretions) | 50 |
Infections or symptoms related to the condition are in brackets (for specific references, see Fabris and Mocchegiani 1995, Shankar and Prasad 1998).
4. Zinc interaction with other micronutrients and zinc toxicity
The beneficial effect of physiological zinc supplementation on the immune system must be, however, related to the competition phenomena between zinc and several cations such as cadmium, lead, calcium, iron, manganese and copper. Beneficial effects of zinc on ameliorating toxicity of cadmium and lead, accentuation of zinc deficiency by administration of calcium and phytate, and production of hypocupremia by excessive zinc intake in humans and animal models, are some of the examples of competition phenomena (Hill, 1976). Such competition occurs, because these ions have similar valence shell electronic structure and, therefore, can be antagonists to each other biologically. Copper, zinc and cadmium with similar orbitals, configurations and coordination numbers have been shown to interact with each other (Hurley et al., 1983). Such interactions occur between zinc and iron (Fe++) at the level of cysteine-histidine ligands for the formation of iron or zinc fingers proteins (Prasad, 1993). In the course of an excess of iron, a preferential binding of iron over zinc with the metal free-protein occurs. The binding of iron results, however, in the loss of nucleic acid-binding activity (Klausner et al., 1993). An excess of zinc as well as zinc deficiency impairs DNA-protein interactions of zinc finger domains with their cognate DNA target sites (Thiesen and Bach, 1991). As a result, the production of some transcriptional factors (SP1, TFIIIA) relevant for cell proliferation and differentiation is impaired (Thiesen and Bach, 1991). The same impairment of zinc finger DNA domains occurs in excess or deficiency of copper (Thiesen and Bach, 1991). This reinforces the notion of the relevance of interactions between zinc and copper and, more in general, also with other metals in the efficiency of immunity (Chandra, 1984). Thus, a limited range of bioavailability exists for each metal: it may be narrow for some and broad for others. Indeed, the beneficial effect of zinc is strictly dependent on the dose and the length of the treatment. Zinc accumulation or imbalance of zinc-to-copper ratios may occur despite low doses of zinc (Fabris and Mocchegiani, 1995). Thus, harmful side effects on the cardiovascular system and brain may appear with increases of both low density lipoprotein and cholesterol (Fosmire 1990), and neural cell-death (Kim et al., 1999). Therefore, caution in zinc supplementation is warranted for successful treatment and to avoid undesirable and harmful side effects. Zinc supplementation must not exceed 2–3 times the RDA/day, for short periods (1–2 months) and in periodical cycles. Such a treatment does not interfere with copper absorption (Fabris and Mocchegiani, 1995). Zinc in picolinate form is the best supplement (Wapnir et al., 1983).
5. Biochemical pathways of zinc affecting immunoresistance to infections in ageing
The mechanism/s by which zinc may affect the immune system are multifaceted due to zinc's widespread action on different enzymes, peptides, transcriptional factors and cytokines involved in various physiological steps of immune development and reactivity. The main biochemical pathways of zinc with possible relevance to immunoresistance to infections in ageing are summarised in Table 3 . Zinc directly influences immune cells by means of multiple enzymes at the level of replication and transcription. These include DNA and RNA polymerases, thymidine-kinase, terminal deoxyribonucleotidyl transferase, and ornithine decarboxylase. All these enzymes are zinc-dependent and involved in mitosis (Chester, 1989). Other zinc-dependent enzymes, such as nucleoside phosphorylase, ecto-5 nucleotidase and protein kinase C (PKC) are directly involved in zinc affects on the immune system because of their presence on lymphocyte membranes (Prasad, 1993). Thus, zinc affects membrane stability by competing with thiols and influencing peroxidative damage (Bettger and O'Dell, 1993). In addition, zinc protects against oxidative stress by pro-inflammatory cytokines with transcriptional factor (NF-kB and AP-1) activation (Hennig et al., 1996). This occurs, because transcriptional factors NF-kB and AP-1 as well as many others involved in cell proliferation or in preventing apoptosis (MTF-1, RING, TFIIIA, EGR-1, EGR-2, BTE, WT-1, TRAF-2, ZEB, KS-1) contain zinc finger-like domains that are influenced by changes in the intracellular pool of zinc (Shankar and Prasad, 1998, Rink and Gabriel, in press). Moreover, zinc plays a role in T-lymphocyte activation by means of tyrosine kinase p56lck, an essential protein in the early steps of activation. Zinc stimulates autophosphorylation of tyrosine residues by p56lck and subsequent phosphorylation of the antigen receptor complex involving CD45 (Pernelle et al., 1991). Reduction of p56lck activity, such as in ageing (Shankar and Prasad, 1998), could reduce the ability of chemokine receptors on T-cells to bind the CC or CXC families of chemokines. This would result in a reduction in the migration of naive T-cells into inflammatory sites and subsequent formation of primed memory T-cells which seem reduced in aged mice (Borghesi and Nicoletti, 1995). By contrast, normal zinc turnover induces p56lck to inhibit viral penetration by phosphorylation of membrane proteins via PKC (Pasternak, 1986). A relevant pathway of zinc in affecting the immune system is related to the discovery of apoptosis prevention by zinc. Apoptosis is a mechanism involved in decreased immune responses during ageing and infections (Monti et al., 1992, Shankar and Prasad, 1998), and low zinc bioavailability is a usual event in both conditions (Fabris and Mocchegiani, 1995). Although, apoptosis prevention is dependent on zinc dose (Fraker and Telford, 1997), the mechanisms by which zinc prevents apoptosis apply at multiple levels. Inhibition of Ca++/Mg++ DNA endonuclease activity, activation of nuclear factors NF-kB and AP-1, inhibition of caspase-3, increase of the Bcl-2/Bax ratio, and blockade of the ligand binding glucocorticoid receptors are the most studied. Zinc prevents apoptosis in ageing, especially by endonuclease enzymes activation (Fraker and Telford, 1997), or by the blockade of the ligand binding glucocorticoid receptors (Provinciali et al., 1998). Another direct pathway is related to the discovery of zinc requirement in the biological activity of a thymic hormone, called thymulin (ZnFTS), which is essential in T-cell maturation and differentiation, in cytokine production, in natural killer cell activity and, consequently, in the efficiency of the entire immune system in host defence (Dardenne et al., 1982, Mocchegiani et al., 1998a). As a consequence, a correct balance of Th1/Th2 cytokines occurs, as demonstrated in the elderly (Cakman et al., 1997, Rink et al., 1998). Such a paradigm may also include Th3 cells because of their control of IL-12 (Fresno et al., 1997), which are in turn decreased in ageing and affected by zinc (Driessen et al. 1994) and thymulin (Hadden, 1994).
Table 3.
Main biochemical pathways of zinc relevant to immunoresistance to infections in ageinga
| Direct effects |
|---|
| DNA-RNA polymerases activation |
| Thymidine–Kinase activation |
| Terminal deoxyribonucleotidyl transferase activation |
| Ornithine decarboxylase activation |
| Ecto-5 nucleotidase activation |
| Protein Kinase-C activation |
| Membrane stability (competing with thiols) |
| Transcriptional factor activation (NF-kB, AP-1) |
| Apoptosis prevention |
| Thymulin activation (ZnFTS) |
| Balance of Th1/Th2/Th3 paradigm (with subsequent cytokine production) |
| MHC class II restricted activation |
| p56lck autophosphorylation |
| Indirect effects |
| Endocrine cell activation (pineal gland, thyroid gland, adenohypophysis, β-cells of pancreas) |
| Hormone receptor superfamily activation (melatonin, growth hormone, Nerve Growth Factor, insulin, IGF-I, thyroid hormones) |
| Metallothioneins-Nitric-oxide-PARP homeostasis |
See for specific references: Shankar and Prasad 1998, Rink and Gabriel, in press, Mocchegiani et al., 2000b, Fraker and Telford, 1997.
Among the indirect pathways, the action of immunomodulating hormones [melatonin, growth hormone, thyroid hormones (T3 and T4) and insulin-like growth factor-1 (IGF-1)] has been recently proposed, because all these hormones can also affect immunity by means of the zinc pool (Mocchegiani et al., 1998a, Mocchegiani et al., 1998b). These are, in turn, required to activate nuclear hormone receptors (Fabris et al., 1997). An indirect pathway of zinc as an antioxidant (Bray and Bettger, 1990), the homeostasis among zinc-bound MT, NO and PARP has been recently proposed, because it is related to the presence of massive and constant stress in ageing (Mocchegiani et al., 2000b). Because of strict links between MT and NO and between NO and PARP, in turn, controlled by the pro-inflammatory cytokines (IL-1 and IL-6) and zinc turnover (see Mocchegiani et al., 2000b), an alteration in one step of this homeostasis may finally divert PARP towards cell-death rather than DNA-repair. Indeed, zinc-bound MT are increased in ageing and acquire a dangerous role for immunity (Mocchegiani et al., 1997). NO is also augmented in ageing (Capanni et al., 1998) with the final result of no protective role by NO against oxidative damage (McBride et al., 1999) and no prevention of apoptosis induced by PARP (Pieper et al., 1999). In contrast, moderate stress in normal zinc bioavailability triggers PARP for DNA repair, as in young-adults (Dantzer et al., 1998). Therefore, altered zinc turnover might be the “primus movens” of decreased immunoresistance to infections in ageing by means of this altered biochemical homeostasis, via pro-inflammatory cytokines (Mocchegiani et al., 2000b). Supplementation of zinc in ageing may correct the defect. Indeed, the increase of MT mRNA caused by zinc (Cousins and Lee-Ambrose, 1992) does not affect the already high liver MT levels (Mocchegiani et al., in press). This suggests that MT are saturated by pre-existing zinc ions in ageing. The increased MT mRNA acts as a reserve in MT production for a prompt response against continuous oxidative damage (Mocchegiani et al., 2000c, Mocchegiani et al., in press) normally affecting NO and PARP activity, as it occurs in young-adult age. In addition, MT regain their original role of protection in releasing zinc to superoxide dismutase (SOD) (Maret et al., 1999), which is decreased in ageing with subsequent cellular oxidative damage (Mocchegiani et al., 1998a). Analagously, the same phenomenon of regained protection occurs for increased alpha-2 macroglobulin in cancer as a result of zinc supplementation (Mocchegiani et al., 1999a). Therefore, supply zinc in ageing is of benefit either to regain MT role of protection or to induce more zinc ion bioavailable in immune efficiency and in NO normal activity, as recently suggested by Sprietsma (1999).
6. Conclusions and future prospects
Zinc is required as a catalytic, structural and regulatory ion for enzymes, proteins and transcription factors. Therefore, it is a key trace element for many body homeostatic mechanisms, including immune responses. Low zinc ion bioavailability results in limited immunoresistance to infections in ageing. Physiological supplementation of zinc for 1–2 months restores immune responses, thereby, reducing the incidence of infections and, consequently, prolonging survival (Ames, 1998). Its use may be of relevance in poor developing countries thanks to the low cost of zinc, where infection incidence is high (Sazawal et al., 1995, Black, 1998).
Among the biochemical pathways of zinc which affect the immune system, zinc-bound MT, NO and PARP homeostasis, via pro-inflammatory cytokines, is also involved because pro-inflammatory cytokines, MT and NO are all increased in ageing. Since NO activates PARP, which has a double role (DNA-repair or cell-death depending on the level of oxidative stress which can be massive in ageing (Capanni et al., 1998)), PARP activity may be addressed towards cell-death rather than DNA-repair (Pieper et al., 1999) (Fig. 1 ). Zinc supplementation in ageing might nonetheless restore the DNA repair function of PARP. This last point is under investigation. Moreover, normal PARP and NO activities are implicated in healthy longevity (Muiras et al., 1998, DeBenedictis et al., 1998 Wingrove and O'Farrell, 1999). Because of increased MT in the liver and atrophic thymus of old mice (Mocchegiani et al., 1998a), their normal production may also be involved in facilitating healthy longevity. Polymorphisms within the human genes encoding NO synthases, PARP and MT exist (Martin et al., 1999, West et al., 1990). The data are, however, fragmentary limited to each factor alone. Genetic studies must be performed in the context of more genes (Schachter et al., 1993). Therefore, the genetic study of this homeostasis in its totality might provide insight into possible biological and genetic markers of immunosenescence. These genetic studies are currently in progress in our laboratory.
Fig. 1.
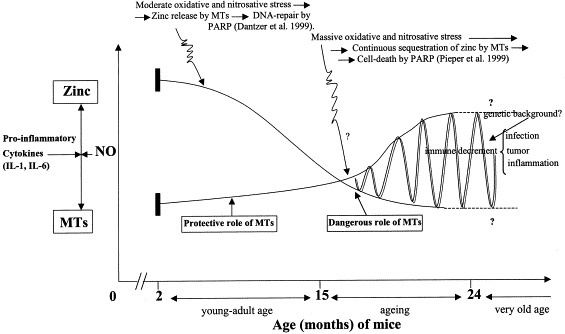
Possible different role of MTs during the whole life. Protective in young-adult age and dangerous in old age with final results of DNA-repair or cell-death by PARP, respectively. This is strictly dependent by oxidative and nitrosative stress status which is moderate in young age, massive in old age (Mocchegiani et al., 2000b). Hatched line, very old age; ?, currently under investigation in our laboratory.
Acknowledgements
This paper was supported by INRCA, Italian Health Ministry (R.F. 99/107 to E. Mocchegiani) and the European Community (ImAginE project: Coordinator Prof. G. Pawelec).
References
- Ames B.N. Micronutrients prevent cancer and delay aging. Toxicol. Lett. 1998;102–103:5–18. doi: 10.1016/s0378-4274(98)00269-0. [DOI] [PubMed] [Google Scholar]
- Arizonol K., Kagawa S., Hamada H., Ariyoshi T. Nitric oxide mediated metalliothonein induction by lipopolysaccharide. Res. Commun. Mol. Pathol. Pharmacol. 1995;90:49–58. [PubMed] [Google Scholar]
- Berg J.M., Shi Y. The galvanization of biology: a growing appreciation for the roles of zinc. Science. 1996;271:1081–1085. doi: 10.1126/science.271.5252.1081. [DOI] [PubMed] [Google Scholar]
- Bettger W.J., O'Dell B.L. Physiological role of zinc in the plasma membrane of mammalian cells. J. Nutr. Biochem. 1993;4:194–207. [Google Scholar]
- Black R.E. Therapeutic and preventive effects of zinc on serious childhood infectious diseases in developing countries. Am. J. Clin. Nutr. 1998;68(Suppl):476S–479S. doi: 10.1093/ajcn/68.2.476S. [DOI] [PubMed] [Google Scholar]
- Bodgen J.D., Oleske J.M., Lavenhar M.A., Munves E.M., Kemp F.W., Bruening K.S. Effects of one year of supplementation with zinc and other micronutrients on cellular immunity in the elderly. Am. J. Clin. Nutr. 1990;9:214–225. doi: 10.1080/07315724.1990.10720372. [DOI] [PubMed] [Google Scholar]
- Bogdan C., Rollinghoff M., Diefenbach A. Reactive oxygen and reactive nitrogen intermediates in innate and specific immunity. Curr. Opin. Immunol. 2000;12:64–76. doi: 10.1016/s0952-7915(99)00052-7. [DOI] [PubMed] [Google Scholar]
- Borghesi C., Nicoletti C. In vivo and in vitro study of the primary and secondary antibody response to a bacterial antigen in aged mice. Int. J. Exp. Pathol. 1995;76:419–424. [PMC free article] [PubMed] [Google Scholar]
- Bray T.M., Bettger W.J. The physiological role of zinc as antioxidant. Free Radic. Biol. Med. 1990;8:281–291. doi: 10.1016/0891-5849(90)90076-u. [DOI] [PubMed] [Google Scholar]
- Cakman I., Rohwer J., Schuz R.M., Kirchner H., Rink L. Dysregulation of Th1 and Th2 and subpopulation in elderly. Mech. Ageing Develop. 1996;87:197–209. doi: 10.1016/0047-6374(96)01708-3. [DOI] [PubMed] [Google Scholar]
- Cakman I., Kirchner H., Rink L. Zinc supplementation reconstitutes the production of interferon-alpha by leukocytes from elderly persons. J. Interferon Cytokine Res. 1997;17:469–472. doi: 10.1089/jir.1997.17.469. [DOI] [PubMed] [Google Scholar]
- Capanni C., Squarzoni S., Petrini S., Villanova M., Muscari C., Maraldi N.A., Guarnieri C., Caldarera C.M. Increase neuronal nitric-oxide synthase in rat skeletal muscle during ageing. Biochem. Biophys. Res. Comm. 1998;245:216–219. doi: 10.1006/bbrc.1998.8404. [DOI] [PubMed] [Google Scholar]
- Chandra R.K. Excessive intake of zinc impairs immune response. J. Am. Med. Ass. 1984;252:1443–1446. [PubMed] [Google Scholar]
- Chester J.K. Biochemistry of zinc in cell division and tissue growth. In: Mills C., editor. Zinc in human biology. Springer; London: 1989. pp. 109–118. [Google Scholar]
- Coleman J.E. Zinc proteins, enzymes, storage proteins, transcription factor and replication proteins. Annu. Rev. Biochem. 1992;61:897–946. doi: 10.1146/annurev.bi.61.070192.004341. [DOI] [PubMed] [Google Scholar]
- Cobbold S., Waldman H. Infectious tollerance. Curr. Opin. Immunol. 1998;10:518–524. doi: 10.1016/s0952-7915(98)80217-3. [DOI] [PubMed] [Google Scholar]
- Cousins R.J., Lee-Ambrose L.M. Nuclear zinc uptake and interaction and metallothioneins gene expression are influenced by dietary zinc in rats. J. Nutr. 1992;122:56–64. doi: 10.1093/jn/122.1.56. [DOI] [PubMed] [Google Scholar]
- Cousins R.J., Leinart A.S. Tissue specific regulation of zinc metabolism and metallothionein genes by interleukin-1. FASEB J. 1988;2:2844–2850. doi: 10.1096/fasebj.2.13.2458983. [DOI] [PubMed] [Google Scholar]
- Dantzer F., Nashever H.P., Vanesch J.L., de Murcia G., Menisser de Murcia J. Functional association of poly (ADP-ribose) polymerase alpha-primase complex: a link between DNA strand break detection and DNA replication. Nucleic Acids Res. 1998;26:1891–1898. doi: 10.1093/nar/26.8.1891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dardenne M., Pleau J.M., Nabama B., Lefancier P., Denien M., Choay J., Bach J.F. Contribution of zinc and other metals to the biological activity of the serum thymic factor. Proc. Natl. Acad. Sci. 1982;79:5370–5373. doi: 10.1073/pnas.79.17.5370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeBenedictis G., Carotenuto L., Carrieri G., De Luca M., Falcone E., Rose G., Cavalcanti S., Corsonello F., Feraco E., Baggio G., Bertolini S., Mari D., Mattace R., Yashin A.I., Bonafè M., Franceschi C. Gene/longevity association studies at four ausosomal loci (REN, THO, PARP, SOD2) Eur. J. Hum. Genet. 1998;6:534–541. doi: 10.1038/sj.ejhg.5200222. [DOI] [PubMed] [Google Scholar]
- Driessen C., Hirv K., Rink L., Kirchner H. Induction of cytokines by zinc ions in human peripheral blood mononuclear cells and separated monocytes. Lymphokine Cytokine Res. 1994;13:15–20. [PubMed] [Google Scholar]
- Fabris N., Mocchegiani E. Zinc, human diseases and aging. Aging Clin. Exp. Res. 1995;7:77–93. doi: 10.1007/BF03324297. [DOI] [PubMed] [Google Scholar]
- Fabris N., Mocchegiani E., Albertini G. Psychoendocrine-immune interactions in Down's syndrome: role of zinc. In: Castells S., Wisniewski K.E., editors. Growth Hormone Treatment in Down's syndrome. Wiley; New York: 1993. pp. 203–218. [Google Scholar]
- Fabris N., Mocchegiani E., Amadio L., Zannotti M., Licastro F., Franceschi C. Thymic hormone deficiency in normal aging and Down's syndrome: is there a primary failure of the thymus? Lancet. 1984;1:983–986. doi: 10.1016/s0140-6736(84)92325-0. [DOI] [PubMed] [Google Scholar]
- Fabris N., Mocchegiani E., Provinciali M. Plasticity of neuroendocrine-immune interaction in aging. Exp. Gerontol. 1997;32:415–429. doi: 10.1016/s0531-5565(96)00166-0. [DOI] [PubMed] [Google Scholar]
- Fagiolo U., Cossarizza A., Sala E., Fanales-Belasio E., Ortolani C., Cozzi D., Monti D., Franceschi C., Paganelli R. Increased cytokines production in mononuclear cells of healthy and elderly people. Eur. J. Immunol. 1993;23:2375–2378. doi: 10.1002/eji.1830230950. [DOI] [PubMed] [Google Scholar]
- Fitzpatrick D.R., Bielefeldt-Ohmann H. Transforming growth factor beta in infectious disease: always there for the host and the pathogen. Trends Microbiol. 1999;7:232–236. doi: 10.1016/s0966-842x(99)01498-5. [DOI] [PubMed] [Google Scholar]
- Fortes C., Forastiere F., Agabiti N., Fano V., Pacifici R., Virgili F., Piras G., Guidi L., Bartoloni C., Tricceri A., Zuccaro P., Ebrahim S., Petrucci G.A. The effect of zinc and vitamin A supplementation on immune response in an older population. J. Am. Geniatr. Soc. 1998;46:19–26. doi: 10.1111/j.1532-5415.1998.tb01008.x. [DOI] [PubMed] [Google Scholar]
- Fosmire C.J. Zinc toxicity. Am. J. Clin. Nutr. 1990;51:225–227. doi: 10.1093/ajcn/51.2.225. [DOI] [PubMed] [Google Scholar]
- Fraker P.J., Telford W.G. A reappraisal of the role of zinc in life and death decision of cells. Proc. Soc. Exp. Biol. Med. 1997;215:229–236. doi: 10.3181/00379727-215-44132. [DOI] [PubMed] [Google Scholar]
- Frasca D., Pucci S., Goso G., Barattini O., Barile S., Pioli C., Doria G. Regulation of cytokine production in aging: use of recombinant cytokines to up regulate mitogen-stimulated spleen cells. Mech. Ageing Develop. 1997;93:157–169. doi: 10.1016/s0047-6374(96)01825-8. [DOI] [PubMed] [Google Scholar]
- Fresno M., Kopf M., Rivas L. Cytokines and infectious diseases. Immunology Today. 1997;18:56–58. doi: 10.1016/s0167-5699(96)30069-8. [DOI] [PubMed] [Google Scholar]
- Hadden J.W. T-cell adjuvants. Int. J. Immunopharmacol. 1994;16:703–710. doi: 10.1016/0192-0561(94)90090-6. [DOI] [PubMed] [Google Scholar]
- Hamer D.H. Metallothionein review. Ann. Rev. Biochem. 1986;55:913–951. doi: 10.1146/annurev.bi.55.070186.004405. [DOI] [PubMed] [Google Scholar]
- Hill C.H. Mineral interrelationships. In: Prasad A.S., editor. II. Academic Press; New York: 1976. pp. 281–300. (Trace Elements in Human Health and Diseases). [Google Scholar]
- Hennig B., Toborek M., McClain C.J. Antiatherogenic properties of zinc implications in endothelial cell metabolism. Nutrition. 1996;12:711–717. doi: 10.1016/s0899-9007(96)00125-6. [DOI] [PubMed] [Google Scholar]
- Hurley, L.S., Keen, C.L., Lonnerdal, B., 1983. Aspects of trace element interactions during development. Fed. Proc. 1735–1739. [PubMed]
- Johnson M.A., Porter K.H. Micronutrient supplementation and infection in institutionalized elders. Nutr. Rev. 1997;55:400–404. doi: 10.1111/j.1753-4887.1997.tb01582.x. [DOI] [PubMed] [Google Scholar]
- Kilbey A., Stephens V., Bartholomew C. Loss of cell cycle control by deregulation of cyclin-dependent Kinase 2 activity in Evi-1 transformed fibrolast. Cell Growth Differ. 1999;10:601–610. [PubMed] [Google Scholar]
- Kim Y.H., Kim E.Y., Gwag B.J., Sohn S., Koh J.Y. Zinc-induced cortical neuronal death. Neuroscience. 1999;89:175–182. doi: 10.1016/s0306-4522(98)00313-3. [DOI] [PubMed] [Google Scholar]
- Klausner R.D., Rouault T.A., Harford J.B. Regulation the fate of mRNA: the control of cellular iron metabolism. Cell. 1993;72:19–28. doi: 10.1016/0092-8674(93)90046-s. [DOI] [PubMed] [Google Scholar]
- Lesourd B.M. Nutrition and immunity in the elderly: modification of immune responses with nutritional treatments. Am. J. Clin. Nutr. 1997;66(Suppl):478S–484S. doi: 10.1093/ajcn/66.2.478S. [DOI] [PubMed] [Google Scholar]
- Li H., Raman C.S., Glaser C.B., Blasko E., Young T.A., Parkinson J.F., Whitlow M., Poulos T.L. Crystal structures of zinc-free and bound heme domain of human inducible nitric-oxide synthase. Implications for dimer stability and comparision with endothelial nitric-oxide synthase. J. Biol. Chem. 1999;274:21276–21284. doi: 10.1074/jbc.274.30.21276. [DOI] [PubMed] [Google Scholar]
- Lio D., D'Anna C., Gervasi F., Scola L., Potestio M., Di Lorenzo G., Listi F., Colombo A., Candore G., Caruso G. Interleukin-12 release by mitogen-stimulated mononuclear cells in the elderly. Mech. Ageing Develop. 1998;102:211–219. doi: 10.1016/s0047-6374(98)00016-5. [DOI] [PubMed] [Google Scholar]
- Lockitch G., Puterman M., Godolphin W., Sheps S., Tingle A.J., Quigley G. Infection and immunity in Down syndrome: a trial of long-term low oral doses of zinc. J. Pediatr. 1989;114:781–787. doi: 10.1016/s0022-3476(89)80136-2. [DOI] [PubMed] [Google Scholar]
- Maret W., Jacob C., Vallee B.L., Fisher E.H. Inhibitory sites in enzymes: zinc removal and reactivation by thionein. Proc. Natl. Acad. Sci. 1999;96:1936–1940. doi: 10.1073/pnas.96.5.1936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin J., Calzada J.E., Nieto A. Inducible nitric oxide synthase (NOS2) gene polymorphism and parasitic diseases. Lancet. 1999;353:72–75. doi: 10.1016/S0140-6736(05)74833-9. [DOI] [PubMed] [Google Scholar]
- McBride A.G., Borutaite V., Brown G.C. Superoxide dismutase and hydrogen peroxide cause rapid nitric oxide breakdown, peroxynitrite production and subsequent cell death. Biochem. Biophys. Acta. 1999;1454:275–288. doi: 10.1016/s0925-4439(99)00046-0. [DOI] [PubMed] [Google Scholar]
- McCaffrey T.A. TGF-betas and TGF-beta receptors in atherosclerosis. Cytokine Growth Factor Rev. 2000;11:103–114. doi: 10.1016/s1359-6101(99)00034-9. [DOI] [PubMed] [Google Scholar]
- Mills, C.F. (Ed.), 1989. Zinc in Human Biology. Springer, London.
- Mocchegiani E., Ciavattini A., Santarelli L., Tibaldi A., Muzzioli M., Bonazzi P., Giacconi R., Fabris N., Garzetti G.G. Role of zinc and α-2 macroglobulin on thymic endocrine activity and on peripheral immune efficiency (Natural Killer activity and IL-2) in cervical carcinoma. Br. J. Cancer. 1999;79:244–250. doi: 10.1038/sj.bjc.6690040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mocchegiani E., Muzzioli M., Cipriano C., Giacconi R. Zinc, T-cell pathways, ageing: role of metallothioneins. Mech. Ageing Develop. 1998;106:183–204. doi: 10.1016/s0047-6374(98)00115-8. [DOI] [PubMed] [Google Scholar]
- Mocchegiani E., Muzzioli M., Cipriano C., Giacconi R. Metallothioneins and extrathymic functions (liver Natural Killer activity) during the circadian cycle in young and old mice. In: Centeno J.A., Collery P., Vernet G., Finkelman R.B., Gibb H., Etienne J.C., editors. Vol. 6. John Libbey Eurotext; Paris: 2000. pp. 150–154. (Metal Ions in Biology and Medicine). [Google Scholar]
- Mocchegiani E., Muzzioli M., Giacconi R. Zinc and immunoresistence to infection in aging: new biological tools. Trends Pharmacological Science. 2000;21:205–208. doi: 10.1016/s0165-6147(00)01476-0. [DOI] [PubMed] [Google Scholar]
- Mocchegiani E., Muzzioli M., Giacconi R. Zinc, metallothioneins, immune responses, survival, ageing. Biogerontology. 2000;1:133–200. doi: 10.1023/a:1010095930854. [DOI] [PubMed] [Google Scholar]
- Mocchegiani, E., Muzzioli, M., Giacconi, R., Cipriano, C., in press. The zinc and metallothioneins homeostasis in thyroid thymus axis efficiency as new biological clock of ageing. J. Clin. Invest.
- Mocchegiani E., Muzzioli M., Gaetti R., Veccia S., Viticchi C., Scalise G. Contribution of zinc to reduce CD4+ risk factor for infection relapse in aging: parallelism with HIV. Int. J. Immunopharmacol. 1999;21:271–281. doi: 10.1016/s0192-0561(99)00009-0. [DOI] [PubMed] [Google Scholar]
- Mocchegiani E., Santarelli L., Tibaldi A., Muzzioli M., Bulian D., Cipriano C., Olivieri F., Fabris N. Presence of links between zinc and melatonin during the circadian cycle in old mice: effects on thymic endocrine activity and on the survival. J. Neuroimmunol. 1998;86:111–122. doi: 10.1016/s0165-5728(97)00253-1. [DOI] [PubMed] [Google Scholar]
- Mocchegiani E., Verbanac D., Santarelli L., Tibaldi A., Muzzioli M., Radosevic-Stasic B., Milin C. Zinc and metallothioneins on cellular immune effectiveness during liver regeneration in young and old mice. Life Science. 1997;61:1125–1145. doi: 10.1016/s0024-3205(97)00646-2. [DOI] [PubMed] [Google Scholar]
- Monti D., Troiano L., Propea F., Grassili E., Cossarizza A., Barozzi D., Pelloni M.C., Tamassia M.G., Bellomo G., Franceschi C. Apoptosis-programmed cell death: a role in the aging process? Am. J. Clin. Nutr. 1992;55:1208–1214. doi: 10.1093/ajcn/55.6.1208S. [DOI] [PubMed] [Google Scholar]
- Muiras M.L., Muller M., Schachter F., Burkle A. Increased poly (ADP-ribose) polymerase activity in lymphoblastoid cell lines from centenarians. J. Mol. Med. 1998;76:346–354. doi: 10.1007/s001090050226. [DOI] [PubMed] [Google Scholar]
- Nagase H. Matrix metalloproteinases. In: Hopper N.M., editor. Zinc, Metalloproteases in Health and Disease. Taylor & Francis; London: 1996. pp. 153–204. [Google Scholar]
- Paramanantham R., Bay B.H., Sit K.H. Flow cytometric evaluation of the DNA profile and cell cycle of zinc supplemented human Chang liver cells. Acta Paediatr. Jpn. 1996;38:334–338. doi: 10.1111/j.1442-200x.1996.tb03501.x. [DOI] [PubMed] [Google Scholar]
- Pasternak C.A. A novel form of host defence: membrane protection by Ca++ and Zn++ Biosc. Rep. 1986;261:81–91. doi: 10.1007/BF01121871. [DOI] [PubMed] [Google Scholar]
- Pawelec G., Solana R. Immunosenescence. Immunology Today. 1997;18:514–516. doi: 10.1016/s0167-5699(97)01145-6. [DOI] [PubMed] [Google Scholar]
- Pernelle J.J., Creuzet C., Loeb J., Gacon G. Phosphorilation of the lymphoid cell kinase p56 lck is stimulated by micromolar concentrations of Zn2+ FEBS Lett. 1991;281:278–282. doi: 10.1016/0014-5793(91)80411-u. [DOI] [PubMed] [Google Scholar]
- Pieper A.A., Verma A., Zhang J., Snyder S.H. Poly (ADP-ribose) polymerase, nitric-oxide and cell-death. Trends Pharmacol. Sci. 1999;19:287–298. doi: 10.1016/s0165-6147(99)01292-4. [DOI] [PubMed] [Google Scholar]
- Prasad A.S. Biochemistry of zinc. Plenum Press; New York: 1993. [Google Scholar]
- Prasad A.S. Zinc and immunity. Mol. Cell Biochem. 1998;188:63–69. [PubMed] [Google Scholar]
- Prasad A.S., Fitzgerald J.T., Hess J.W., Kaplan J., Pelen F., Dardenne M. Zinc deficiency in elderly patients. Nutrition. 1993;9:218–224. [PubMed] [Google Scholar]
- Provinciali M., Di Stefano G., Stronati S. Flow cytometric analysis of CD3/TCR complex, zinc, and glucocorticoid-mediated regulation of apoptosis and cell cycle distribution in thymocytes from old mice. Cytometry. 1998;32:1–8. [PubMed] [Google Scholar]
- Rink L., Cakman I., Kirchner H. Altered cytokines in the elderly. Mech. Ageing Develop. 1988;102:199–209. doi: 10.1016/s0047-6374(97)00153-x. [DOI] [PubMed] [Google Scholar]
- Rink, L., Gabriel, P., 2000. Zinc and the immune system. Br. J. Nutr. [DOI] [PubMed]
- Romagnani S. The Th1/Th2 paradigm. Immunology Today. 1997;18:263–266. doi: 10.1016/s0167-5699(97)80019-9. [DOI] [PubMed] [Google Scholar]
- Salem M.L., Matsuzaki G., Madkour G.A., Nomoto K. Beta-estradiol induced decrease in IL-12 and TNF-alpha expression suppress macrophages functions in the course of Listeria monocytogens infection in mice. Int. J. Immunopharmacol. 1999;21:481–497. doi: 10.1016/s0192-0561(99)00027-2. [DOI] [PubMed] [Google Scholar]
- Sazawal S., Black R.E., Bhan M.K., Bhandari N., Sinha A., Jalla S. Zinc supplementation in young children with acute diarrhea in India. N. Engl. J. Med. 1995;333:839–844. doi: 10.1056/NEJM199509283331304. [DOI] [PubMed] [Google Scholar]
- Schachter F., Cohen D., Kirkwood T. Prospects for the genetics of human longevity. Hum. Genet. 1993;91:519–526. doi: 10.1007/BF00205074. [DOI] [PubMed] [Google Scholar]
- Shankar A.H., Prasad A.S. Zinc and immune function: the biological basis of altered resistance to infections. Am. J. Clin. Nutr. 1998;68(Suppl):447S–463S. doi: 10.1093/ajcn/68.2.447S. [DOI] [PubMed] [Google Scholar]
- Shi H.N., Koski K.G., Stevenson M.M., Scott M.E. Zinc deficiency and energy restriction modify immune responses in mice during both primary and challenge infection with Heligmosomoides polygyrus (Nematoda) Parasite Immunol. 1997;19:363–373. doi: 10.1046/j.1365-3024.1997.d01-223.x. [DOI] [PubMed] [Google Scholar]
- Sprietsma J.E. Modern diets and diseases: NO-zinc balance. Under Th1, zinc and nitrogen monoxide (NO) collectively protect against viruses, AIDS, autoimmunity, diabetes, allergies, asthma, infectious diseases, atherosclerosis and cancer. Med. Hypothesis. 1999;53:6–16. doi: 10.1054/mehy.1999.0867. [DOI] [PubMed] [Google Scholar]
- Stabile A., Pesaresi M.A., Stabile A.M., Pastore M., Sopo S.M., Ricci R., Celestini E., Segni G. Immunodeficiency and plasma zinc levels in children with Down's syndrome: a long term follow-up of oral zinc supplementation. Clin. Immunol. Immunopathol. 1991;58:207–216. doi: 10.1016/0090-1229(91)90137-y. [DOI] [PubMed] [Google Scholar]
- Szabo C., Dowson V.L. Role of poly (ADP-ribose) synthetase in inflammation and ischemia-reperfusion. Trends Pharmacol. Sci. 1998;19:287–298. doi: 10.1016/s0165-6147(98)01193-6. [DOI] [PubMed] [Google Scholar]
- Szalay G., Ladel C.H., Blum C., Brossay L., Kronenberg M., Kaufmann S.H. Cutting edge: anti-CD1 monoclonal antibody treatment reverses the production patterns of TGF-beta2 and Th1 cytokines and ameliorates listeriosis in mice. J. Immunol. 1999;162:6955–6958. [PubMed] [Google Scholar]
- Taylor G.A., Blackshear P.J. Zinc inhibits turnover of labile mRNA in intact cells. J. Cell Physiol. 1995;162:378–387. doi: 10.1002/jcp.1041620310. [DOI] [PubMed] [Google Scholar]
- Thiesen H.J., Bach C. Transition metals modulate DNA-protein interaction of SP1 zinc finger domain with its cognate traget site. Biochem. Biophys. Res. Comm. 1991;176:551–557. doi: 10.1016/s0006-291x(05)80219-0. [DOI] [PubMed] [Google Scholar]
- Trucco C., Flatter E., Fribourg S., de Murcia G., Menisser-de Murcia J. Mutations in the amino terminal domain of the human poly (ADP-ribose) polymerase that affect catalytic activity but not its DNA binding capacity. FEBS Lett. 1996;399:313–316. doi: 10.1016/s0014-5793(96)01347-6. [DOI] [PubMed] [Google Scholar]
- US Recommended Daily Allowance (USDA) for Dietary Intakes of Minerals, 1976. Federal Register 41, 46172–46176.
- Vallee B.L., Falchuk K.H. The biochemical basis of zinc physiology. Phys. Res. 1993;73:79–118. doi: 10.1152/physrev.1993.73.1.79. [DOI] [PubMed] [Google Scholar]
- Wapnir R.A., Khani D.E., Bayne M.A., Lifshitz F. Absorption of zinc by the rat ileum: effects of histidine and other low-molecular-weight ligands. J. Nutr. 1983;133:1346–1354. doi: 10.1093/jn/113.7.1346. [DOI] [PubMed] [Google Scholar]
- West A.K., Stallings R., Hildebrand C.E., Chiu R., Karin M., Richards R.I. Human metallothionein genes: structure of the functional locus at 16q3. Genomics. 1990;8:513–518. doi: 10.1016/0888-7543(90)90038-v. [DOI] [PubMed] [Google Scholar]
- Wellinghausen N., Kirchner H., Rink L. The immunobiology of zinc. Immunology Today. 1997;18:519–521. doi: 10.1016/s0167-5699(97)01146-8. [DOI] [PubMed] [Google Scholar]
- Wingrove J.A., O'Farrell P.H. Nitric oxide contributes to behavioral, cellular and developmental responses to low oxygen in Drosophila. Cell. 1999;98:105–114. doi: 10.1016/S0092-8674(00)80610-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoo Y.D., Chiou C.J., Choi K.S., Yi Y., Michelson S., Kim S., Hayward G.S., Kim S.J. The IE2 regulatory protein of human cytomegalovirus induces expression of the human transforming growth factor beta 1 gene through an Erg-1 binding site. J. Virol. 1996;70:7062–7070. doi: 10.1128/jvi.70.10.7062-7070.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yurkow E.J., Makhijani P.R. Flow cytometric determination of metallothionein levels in human peripheral blood lymphocytes: utility in enviromental exposure assessment. J. Toxicol. Environmental Health. 1998;157:445–457. doi: 10.1080/009841098158737. [DOI] [PubMed] [Google Scholar]