

Abstract
Feline α1-acid glycoprotein (fAGP) modifies both its serum concentration and its glycan moiety during diseases. fAGP is hyposialylated in cats with feline infectious peritonitis (FIP), but not in clinically healthy cats or in cats with other diseases. This study was aimed to determine whether hyposialylated fAGP influences phagocytosis. A flow cytometric method based on ingestion of fluoresceinated bacteria and adapted to feline blood was used to assess phagocytosis of leukocytes incubated with ‘non-pathological’ fAGP (purified from sera with normal concentrations of AGP) and ‘pathological’ fAGP (purified from sera with >1.5 mg/mL hyposialylated AGP). The flow cytometric method provided repeatable results for neutrophils (coefficients of variations, CVs <15%) but not for monocytes (CVs > 20%) which had also a high individual variability. Compared with saline solution and with non-pathological fAGP, pathological fAGP significantly decreased phagocytosis in neutrophils and monocytes. This study demonstrated that hyposialylated fAGP down-regulates the phagocytic activity of feline neutrophils.
Keywords: Feline, Phagocytosis, α-1-Acid glycoprotein, Flow cytometry, Feline infectious peritonitis
1. Introduction
Feline α1-acid glycoprotein (fAGP) works as an acute phase protein in cats (Paltrinieri, 2008) and increases in the serum of cats with several inflammatory and non-inflammatory conditions (Paltrinieri et al., 2007b; Paltrinieri, 2008). The functions of AGP include the dampening of the inflammatory status to minimise tissue damages associated with the release of reactive oxygen species that characterizes the last phase of the phagocytic process of granulocytes and monocytes recruited in tissues during inflammation (Tilg et al., 1993, Atemezem et al., 2001, Hochepied et al., 2003, Miranda-Ribera et al., 2010). In addition to its serum concentration, post-translational glycosylation and phosphorylation of AGP can be modified during inflammation or diseases (Ceciliani and Pocacqua, 2007).
The glycan moiety influences several immunomodulatory activities of AGP (Hochepied et al., 2003, Ceciliani and Pocacqua, 2007). Carbohydrate moiety modifications could be important in the inflammatory response but may also be responsible for impairment of innate defences by modifying phagocyte functions. Most of the experiments focusing on the relationship between the glycan moiety of AGP and its involvement in defensive responses were performed on experimentally desialylated AGP, due to the difficulties to retrieve a sufficient number of unhealthy animals (Ceciliani et al., 2007, Miranda-Ribera et al., 2010).
In cats, hyposialylated AGP, in α(2-6)-linked and α(2-3)-linked sialic acid, has been found in feline infectious peritonitis (FIP) (Ceciliani et al., 2004). FIP is sustained by mutated viral variants of the feline coronavirus (FCoV) able to replicate into macrophages (Pedersen, 2009). The development of the disease depends on the immune response of the host (Pedersen, 1987), as suggested by the different number of lymphocytes and by the different concentration of molecules involved in inflammation or immunity found in cats with FIP compared with FCoV-infected, clinically healthy cats (Kipar et al., 2001, Berg et al., 2005, Gelain et al., 2006, Giordano and Paltrinieri, 2009). Hyposialylation seems to be a unique feature of FIP, since it does not occur in diseases other than FIP, except in a few cats infected by the feline leukemia virus (FeLV) or by the feline immunodeficiency virus (FIV) (Pocacqua et al., 2005) or in clinically healthy FCoV-infected cats, which exhibit transient increases of hypersialylated fAGP (Giordano et al., 2004, Paltrinieri et al., 2007a).
A relationship between the degree of sialylation of AGP and the clinical outcome of the infection can be hypothesised. Due to its involvement in down-regulating the inflammation, it is possible that the hyposialylation of AGP may increase the susceptibility of cats by altering the responses of phagocytes. The aim of the present study was to determine whether hyposialylated AGP purified from unhealthy cat interferes with phagocytic activity of feline granulocytes and monocytes. To fulfil this aim, we used a flow cytometric method previously employed in cats (Hoffmann-Jagielska et al., 2003, Hoffmann-Jagielska et al., 2006) to evaluate phagocytosis of fluoresceinated bacteria in whole blood.
2. Materials and methods
2.1. Animals and sample collection
Blood samples were taken from 16 clinically healthy client-owned cats under informed consent during routine wellness visits. Therefore, according to the regulations of our Institution, it was not necessary to obtain a formal authorisation by the Animal Care Committee. Blood was placed either in tubes with lithium heparin (15 U/mL), or in plain tubes and immediately transported to the laboratory to be analysed within 1 h. Specifically, anticoagulated blood was used to perform routine hematology and to assess phagocytic activity, while plain tubes were used to obtain serum by centrifugation to perform the biochemical analyses described below.
The inclusion criterion was the absence of any clinical sign and of laboratory abnormalities in a routine laboratory hematological and biochemical screening. Specifically biochemical analyses were run on serum using an automated spectrophotometer (Cobas Mira, Roche, Basel, Switzerland) using reagents provided by Real Time S.r.l (Viterbo, Italy) by measuring the following analytes: total proteins (biuret method), albumin (bromochresol method), glucose (GOD-POD method), urea (urease method), creatinine (Jaffè method), alanine aminotransferase (ALT, kinetic IFCC method), aspartate aminotransferase (AST, kinetic IFCC method), alkaline phosphatase (ALP, kinetic IFCC method), cholesterol (COD-POD-PAP method), Triglycerides (GPO-POP-PAP method), calcium (orthocresoftaleine method), phosphate (phosphomolibdate method), γ-glutamyl transferase (GGT, kinetic IFCC method), creatine kinase (CK, CK-NAC method). The serum concentration of AGP was also measured using the method described below, and only cats with results within the reference intervals were enrolled in this study. Routine hematology was performed using an automated laser hematology analyzer (Sysmex XT-2000iV, Sysmex Co., Chuo-ku, Kobe, Japan), already validated for feline blood (Lilliehöök and Tvedten, 2009) equipped with a multispecies software for veterinary use, followed by micrscopical analysis of May Grunwald Giemsa stained blood smears to perform the differential leukocyte counts.
Heparinized blood from the first 10 cats enrolled in the study was also used to optimize the flow cytometric method described below (cats 1–5) and to evaluate the precision of this method (cats 6–10). As regards the leukocyte populations of interest for this study, differential cell counts of these cats were normal (4.52 ± 2.02 neutrophils × 103/μL; 2.86 ± 1.08 lymphocytes × 103/μL and 0.20 ± 0.07 monocytes × 103/μL). Then, additional six heparinized samples (cats 11–16) were used to assess the phagocytic responses in the presence of fAGP. Blood was placed in tubes with lithium heparin, transported to the laboratory and analysed within 1 h. Differential cell counts from blood of these cats were normal (5.27 ± 3.10 neutrophils × 103/μL; 3.02 ± 1.57 lymphocytes × 103/μL and 0.23 ± 0.11 monocytes × 103/μL).
2.2. Evaluation of phagocytic activity
Phagocytic activity was assayed in heparinised whole blood using a commercial kit (Phagotest, Glycotope Biotechnology) based on the incubation of blood with fluoresceinated (fluorescein isothiocyanate, FITC-labelled) Escherichia coli, followed by flow cytometric quantification of phagocytosis. Aliquots of 100 μL blood were prepared to assess in duplicate either non-activated phagocytosis (basal phagocytic activity of resting cells incubated on ice) or activated phagocytosis (phagocytic activity of cells incubated for 10 min in a water bath at 37 °C followed by rapid blocking of phagocytosis by immersion on ice). Before incubation, each aliquot was mixed with different amounts (see below) of FITC-labelled E. coli. After incubation, a quenching solution (100 μL) was added to inactivate the fluorescence of non-phagocytosed E. coli. Cell suspensions were washed twice with phosphate buffered saline (PBS) at 4 °C, centrifuged (5 min, 250g, 4 °C) and mixed with the lysing solution provided in the kit. After 5 min, cells were washed again and 200 μL DNA staining solution was added, followed by incubation on ice for 10 min.
Phagocytosis was assessed at 488 nm wavelength using a flow cytometer (FACSCalibur, Becton Dickinson) equipped with the CellQuest software (Becton Dickinson) by analyzing a volume of blood containing 10,000 cells in the gate of neutrophils and monocytes were gated on dot plots of side scatter (SSC) and forward scatter (FSC) analysis (Supplementary Fig. 1). These gates were designed during a previous study (Paltrinieri et al., 2012) based on cell distribution assessed using monoclonal antibodies against each leukocyte population. To exclude artifacts due to the residual fluorescence of aggregates of E. coli the intensity of positivity of DNA staining was assessed on histograms of the gated population to differentiate aploid bacteria from diploid nucleated cells. Diploid cells were back-gated to evaluate phagocytosis: two histograms (neutrophils and monocytes) were generated and data were expressed as percentage of phagocytosing cells and as median channel fluorescence (MCF) emitted by the cells. The MCF is proportional to the amount of ingested bacteria per cell. Data were recorded on a logarithmic scale and values of non-activated samples were subtracted from those of activated samples to determine the percentage of phagocytic cells and the MCF of each sample.
Different concentrations of E. coli were tested in preliminary assays. In the first test, 20 μL E. coli suspension (1 × 109 E. coli/mL) were added to each aliquot of blood. Further tests were performed using 10 μL, 5 μL and 2.5 μL E. coli suspension. Intra-assay precision was assessed on duplicate measurement of five samples analysed using the optimised method by calculating the coefficient of variation (CV = mean/standard deviation × 100).
2.3. Purification of fAGP from pooled sera and evaluation of the pattern of sialylation
Feline AGP was purified from two pools of feline sera received at our institution for routine diagnostic investigation and stored at −20 °C for a maximum of 6 months. The concentration of AGP was assessed at admission using a single radial immunodiffusion (SRID) kit (Feline a1AG Plate, Tridelta) (Paltrinieri et al., 2007a). Sera were pooled based on their AGP concentration. Forty-six samples from cats with a clinical suspicion of FIP and serum AGP concentrations >1.5 mg/mL (total volume 20.9 mL) were included in the ‘pathological pool’. Fifty-nine sera (total volume 29.9 mL) from clinically healthy cats with serum AGP concentrations <0.5 mg/mL were used to form the ‘non-pathological pool’, since this value is within the reference interval of our laboratory (<0.56 mg/mL). The concentrations of AGP assessed by SRID were 0.42 mg/mL in the non-pathological pool and 3.84 mg/mL in the pathological pool.
Feline AGP was purified from both the pools as described previously (Ceciliani et al., 2004), with the addition of a third reverse phase high performance liquid chromatography (RP-HPLC) purification step to remove contaminants such as lipopolysaccharide (LPS), which might interfere with in vitro experiments with cells. This additional step completely remove LPS from the purified protein, as demonstrated using the LAL test in a previous study (Ceciliani et al., 2007). Protein peaks recorded at each chromatographic step are reported in the Supplementary Fig. 2).
After each step of the purification protocol, protein concentrations were quantified by spectrophotometric measurement at 280 nm and fAGP concentrations were determined by SRID. The presence of fAGP in the purified protein fractions was assessed by Western blot analysis using an anti-fAGP polyclonal antibody raised in sheep kindly provided by Professor David Eckersall (University of Glasgow, UK) and an alkaline phosphatase-conjugated goat anti-sheep secondary antibody. Western blots were developed using the Amplified AP Immun-Blot Kit (Bio-Rad).
AGP sialylation was characterised in a further Western blotting experiment using the biotin-conjugated lectins Sambucus nigra agglutinin (SNAI, 2 μg/mL; Vector Laboratories) and Maackia amurensis agglutinin (MAA, 5 μg/mL; Vector Laboratories) specific for sialic acid α(2-6)-linked and α(2-3)-linked to galactose, respectively.
Glycan analysis of fAGP-lectins binding was performed following a peroxidase reaction using the Vectastain ABC Kit (Vector Laboratories) as described previously (Ceciliani et al., 2004) and developed using the HRP Immun-Blot Assay Kit (Bio-Rad Laboratories). Pathological and non-pathological AGP were analysed in the same blot. Densitometric analysis was then performed using the Imagemaster 1D software (Amersham Biosciences) that analysed digital images of the membranes acquired with a scanner in order to display profile plots of fAGP peaks and to calculate the area under each peak.
2.4. Effects of fAGP on the phagocytic activity of feline leucocytes
The effect of fAGP on phagocytosis was assessed on blood from six clinically healthy cats with AGP concentration ranging from 0.24 to 0.31 mg/mL. Samples were divided in six aliquots (200 μL each) that were added with 30 μL of the following solutions: (1) saline solution, to be placed on ice after incubation (non-activated negative control); (2) saline solution to activate phagocytosis as described above (activated negative control: baseline phagocytosis); (3) albumin, to evaluate the effect on phagocytosis of a non-specific protein (i.e. non-AGP); (4) ‘non-pathological fAGP’, prepared as described above; (5) AGP from clinically healthy cattle (Ceciliani et al., 2007): bovine AGP (bAGP) served to test the effect on phagocytosis of a AGP from a species other than cats; and (6) ‘pathological fAGP’, prepared as described above. The final protein concentration in wells 3–6 was 1.5 mg/mL. The 6 aliquots of blood added with saline solution or with the proteins listed above, were used to assess the phagocytic activity in duplicate. To this aim each aliquot was used to fill two wells of a microtitre plate (100 μL each). The plate was then incubated for 30 min at 37 °C with 5% CO2. After incubation, blood from each well was used to assess phagocytosis using the optimised protocol described above.
2.5. Statistical analysis
Statistical analyses were performed using an Excel spreadsheet with the Analyse-it set of macroinstructions (Analyse-it Software, version 2.21). Non parametric analyses were used since a Kolgomorov Smirnov normality test demonstrated that data did not have a normal distribution. Results recorded after incubation in the different experimental conditions were compared to each other with the Friedmann test. Results obtained with each protein were then compared with those obtained with saline solution with a Wilcoxon signed rank paired t test. The same test was used to compare the results obtained with non pathological feline AGP and those obtained with pathological feline AGP or bovine AGP. Values were considered to be significant when P < 0.05.
3. Results
3.1. Optimisation of the protocol to evaluate phagocytosis of feline leukocytes
The application of the manufacturer’s protocol demonstrated that the signal exceeded the maximum visible fluorescence (Supplementary Fig. 3). Conversely, fluorescence histograms were completely visible using 5 μL or 2.5 μL E. coli suspension containing respectively 2.5 × 108 and 1.25 × 108 E. coli/mL. The former dilution (5 μL) was finally selected, since an excess of bacteria is needed to not underestimate phagocytic activity. At this dilution, the FSC vs. SSC plot showed that most of the phagocytic cells were neutrophils and monocytes, although a residual non-specific signal was found on lymphocytes. Histograms of gated neutrophils and monocytes demonstrated a moderate background signal in non-activated cells (Fig. 1 a and c; Table 1 ) and a strong positive signal in activated neutrophils. Positivity was less intense for activated monocytes, which also showed a high individual variability (Fig. 1c and d; Table 1). Intra-assay imprecision of the modified method was high in non-activated samples (Supplementary Table 1) and low (CVs < 15%) in activated samples, although individual cases had CVs > 20% (especially for monocytes).
Fig. 1.
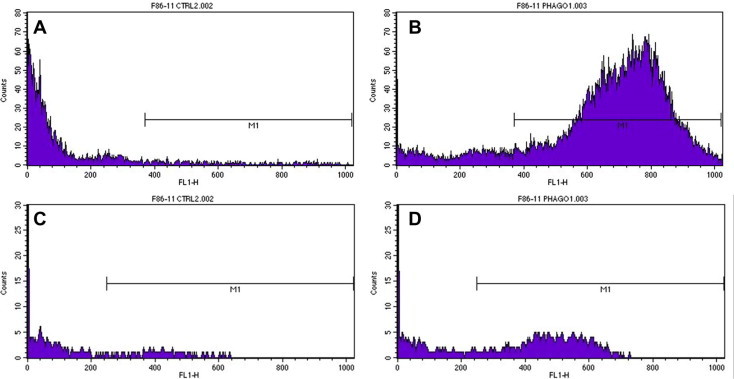
Examples of histograms obtained in non-activated neutrophils (A), activated neutrophils (B), non-activated monocytes (C) and activated monocytes (D). The gate designed in the non-activated population to identify the percentage of non-specific phagocytosis is indicated with ‘M1’. The comparisons of histograms A and C with those in B and D, respectively demonstrates the increased fluorescence obtained after stimulation compared with non-activated cells (i.e. cells kept on ice to block phagocytosis), detectable as a peak in the M1 area, that is not present in non activated cells.
Table 1.
Results obtained in five clinically healthy cats using the optimised flow cytometric method.
| Non-activated | Activated | Difference | |
|---|---|---|---|
| Mean ± SD (median) Min–Max | Mean ± SD (median) Min–Max | Mean ± SD (median) Min–Max | |
| % PMN | 0.8 ± 0.2 (0.9) 0.5–1.0 | 75.6 ± 9.3 (70.7) 67.2–89.9 | 74.7 ± 9.2 (70.2) 66.3–88.9 |
| MCF PMN | 1.1 ± 0.1 (1.0) 1.0–1.2 | 614.6 ± 79.3 (578.5) 545–709 | 613.6 ± 79.3 (577.5) 544–708 |
| % Mono | 10.6 ± 8.1 (7.8) 2.5–20.8 | 39.1 ± 15.8 (37.3) 17.5–55.3 | 28.5 ± 9.9 (34.5) 13.1–36.3 |
| MCF Mono | 14.5 ± 19.8 (8.0) 1.0–48.5 | 147.3 ± 116.8 (103.5) 47.0–344.5 | 132.8 ± 113.1 (102.5) 46.0–330.5 |
SD, standard deviation; Min, minimum; Max, maximum;% PMN, percentage of phagocytic neutrophils; MCF PMN, median channel fluorescence intensity of neutrophils;% Mono, percentage of phagocytic monocytes; MCF PMN, median channel fluorescence intensity of monocytes.
3.2. Purification of AGP
After purification, 0.66 mg non-pathological fAGP and 2.20 mg pathological fAGP were recovered. Coomassie blue staining confirmed that AGP was purified to homogeneity and Western blot analysis confirmed that the purified protein was AGP in both samples (Supplementary Fig. 5). The densitometric analysis of the glycosylation patterns of non-pathological and pathological fAGP (Fig. 2 ) revealed that, compared with non-pathological AGP, the sialic acid α(2-6)-linked content of pathological fAGP (lectin SNAI) decreased to 63% and the sialic acid α(2-3)-linked to galactose (lectin MAA) decreased to 42%. These results confirmed that pathological fAGP was hyposialylated.
Fig. 2.

Western blot analysis of non-pathological (NP) and pathological (P) fAGP stained with the lectins Sambucus nigra agglutinin (SNAI) and Maackia amurensis agglutinin (MAA) that are specific for sialic acid α2-6)-linked and α2-3)-linked to galactose, respectively. In Western blots for both lectins (A), each lane was loaded with 2 μg purified protein. Non-pathological protein reacts strongly with both lectins when compared with the pathological protein. The signal recorded with the two lectins in the pathological pool is weak. For each lectin, the reactivity with SNAI and MAAI has been quantified by a densitometry and the quantification of the area under the peaks (B) confirms that for both lectins, the value corresponding to pathological fAGP is lower than that of non pathological fAGP.
3.3. Effect of AGP on phagocytosis of feline phagocytes
Results recorded in controls and in the presence of fAGP or other proteins are summarised in Fig. 3 . Pathological fAGP decreased in all the four flow cytometric parameters either compared with saline solution or compared with non-pathological fAGP. A significant decrease of phagocytosing monocytes compared with saline solution was induced by bAGP. No other significant differences were found, but non-pathological fAGP had a strong individual variability, due to a single cat that had extremely low values, and monocytes had a wide individual variability for almost all the parameters. The inhibitory activity on neutrophils was AGP-specific, since the same concentration of other proteins was ineffective.
Fig. 3.
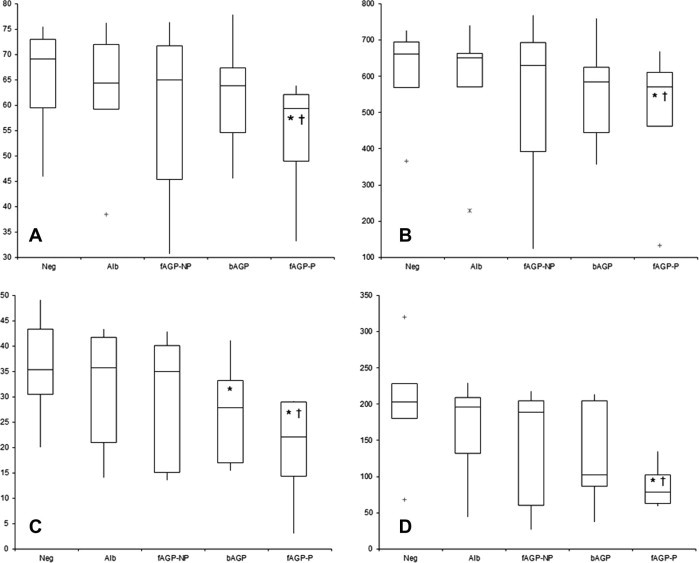
Results obtained after incubation of whole blood with saline solution (Neg), bovine albumin (Alb), non-pathological feline AGP (fAGP-NP), bovine AGP (bAGP) and pathological feline AGP (fAGP-P). The four graphs display the percentage of phagocytosing neutrophils (A), the median channel fluorescence (MCF) of neutrophils (B), the percentage of phagocytosing monocytes (C) and the MCF of monocytes (D). The boxes indicates the I–III interquartile range (IQR), the horizontal line indicates the median values, whiskers extend to further observation within the I quartile minus 1.5 ∗ IQR or to further observation within the III quartile plus 1.5 ∗ IQR. Asterisks indicate values that are significantly different from the negative control (saline solution) and the symbols † indicate values that are significantly different from non pathological fAGP: as regards neutrophils, both the percentage of the phagocytic cells and the MCF were significantly lower after incubation with pathological fAGP compared with controls and with non pathological fAGP. Conversely, as regards monocytes, the percentage of the phagocytic cells and the MCF were significantly lower after incubation with pathological fAGP compared with controls and with non-pathological fAGP. The incubation with bovine AGP induced a significant decrease of the percentage of the phagocytic cells only compared with controls but not with non-pathological fAGP.
4. Discussion
This study demonstrated that hyposialylated fAGP purified from unhealthy cats can interfere with phagocytosis. The phagocytic activity of feline neutrophils and monocytes was determined by a flow cytometric protocol that copes with the need to work with low volumes of blood, as frequently occurs in cats, and provides the opportunity to work with whole blood, which paints a more reliable picture of cell–cell interactions that occur in blood. The main limitation of tests performed in whole blood is that other substance in plasma (cytokines, pro-inflammatory molecules, or even endogenous AGP) may interfere with cell responses. The study design adopted in this study, however, minimized the possible influence of endogenous substances since on one side all the cats were clinically healthy and have a normal concentration of AGP, and on the other side an aliquot of whole blood was incubated with saline solution. Therefore, the only difference between this aliquot and activated aliquots was the amount (and, as regards fAGP, the type of sialylation) of activator(s). Therefore, it is very likely that the different responses recorded after incubation with AGP or albumin actually depend on the presence of activators rather than on endogenous molecules.
This or similar methods were already been employed in cats, (Hoffmann-Jagielska et al., 2003, Hoffmann-Jagielska et al., 2006, Hanel et al., 2003, Chen et al., 2006, Jaso-Friedmann et al., 2008, Craig et al., 2009, Webb et al., 2009, Keegan and Webb, 2010). However, the methods cited above were not specifically customed for feline cells. A preliminary step of optimisation to feline blood was therefore carried out, followed by the evaluation of imprecision. This approach allowed us to improve the quality of the signal by decreasing the number of E. coli added to cell suspensions, likely due to a decreased non specific binding of bacteria to cell surfaces. The residual positivity on lymphocytes, possibly due to non-specific adherence of bacteria or to the low phagocytic activity of lymphocytes (Tizard, 2009, Parra et al., 2012) did not influence the interpretation of neutrophil and monocyte, since the phagocytic activity of these cells was evaluated on gated cell populations that do not contain lymphocytes (Paltrinieri et al., 2012).
Using the optimized method, intra-assay imprecision was higher in non-activated cells, as expected for datasets with low mean values (Westgard, 2003), but acceptable in activated cells, with CVs comparable to those reported previously (Hoffmann-Jagielska et al., 2003). Neutrophils had a higher phagocytic activity than monocytes, since phagocytosis of monocytes must be activated by cytokines (Netea et al., 2008, Tizard, 2009). Results of monocytes were also biased by the high imprecision and individual variability.
In the second part of the experiment, we determined the phagocytosis-modulatory activity of fAGP by incubating feline blood with AGP purified from healthy and unhealthy animals. This study was not focused to assess the phagocytic responses of cats with FIP. Nevertheless, AGP purified from sick cats was strongly hyposialylated for both α(2-3)-linked and α(2-6)-linked sialic acid, with a magnitude similar to that found in cats with FIP (Ceciliani et al., 2004). This finding, coupled with the high AGP concentration in sera from sick animals, which is also consistent with FIP (Duthie et al., 1997), suggests that, although the clinical diagnosis of FIP was not confirmed post-mortem, it is very likely that most of the pathological sera came from FIP affected cats.
The last step of purification (RP-HPLC) that removes contaminating LPS reduced the purification yield, as already reported (Ceciliani et al., 2007). By consequence, the amount of purified AGP was sufficient only for a limited number of replicas. In spite of this limitation, the pathological and strongly hyposialylated fAGP down-regulated phagocytosis of neutrophils and, to a lesser extent, of monocytes, when added at concentrations similar to those found in FIP. The high variability of monocytes depends on the intrinsic variability of their responses, potentially amplified by the low number of cats tested, rather than on a true effect of AGP. The possible influence of subclinical conditions was also unlikely since the cats were clinically and clinico-pathologically healthy.
In humans and cattle, AGP modulates monocyte and neutrophil functions (Vasson et al., 1994, Lecchi et al., 2008, Rinaldi et al., 2008, Miranda-Ribera et al., 2010). AGP interacts with target cells through its glycan moiety, and the experimental desialylation inhibits immunomodulatory activities such as the apoptosis protecting function (Ceciliani et al., 2007) and the degranulatory inhibitory activity (Miranda-Ribera et al., 2010).
The finding that hyposialylated fAGP purified from unhealthy cats can down-regulate phagocytosis is remarkable and gives some insight in the relationship between the sialylation degree of AGP and the modulation of its functions during diseases where post-translational modifications of the proteins occur. The results of this study do not allow us to formulate any hypothesis on the mechanism(s) responsible for this finding. Hyposialylation has been found to be specific in FIP, but not in clinically healthy, FCoV infected cats (Ceciliani et al., 2004, Paltrinieri et al., 2008). Although previous report, performed with methods different from those reported in this study, suggested that phagocytosis is not suppressed in cats wit FIP (Knotek et al., 2000), the hyposialylation of fAGP, which we found to down-regulate phagocytosis, may influence the clinical outcome of FCoV infection by inducing a failure of cellular responses to the virus. Macrophages and neutrophils are involved in FIP immunopathology, being the most abundant, while hardly effective, populations in pyogranulomatous lesions (Kipar et al., 1998, Paltrinieri et al., 1998). It would be interesting to investigate whether other cell-mediated functions are hampered by desialylated fAGP. Among these, the potential of AGP of modulate apoptosis through variations in the glycan moiety (Williams et al., 1997, De Vries et al., 2004, Kagaya et al., 2005, Ceciliani et al., 2007) could be interesting, since lymphocyte apoptosis occurs in cats with FIP (Takano et al., 2007), but not in FCoV-infected, clinically healthy cats (Paltrinieri et al., 2003).
In conclusion, using a modified flow cytometric method to evaluate phagocytosis in feline whole blood, hyposialylated fAGP down-regulated neutrophil phagocytosis, thus supporting the hypothesis that post-translational modifications of AGP contributes to the pathogenesis of diseases involving cell-mediated responses, such as FIP (Ceciliani et al., 2004). Further experiments on the phagocytic activity in FIP affected cats are envisaged to confirm this hypothesis.
Conflict of interest
None of the authors of this paper has a financial or personal relationship with other people or organisations that could inappropriately influence or bias the content of the paper.
Acknowledgements
This study has been funded with the grant PUR2008 from the University of Milan and by the European Social Fund (Fondo Sociale Europeo, Regione Lombardia), through the grant “Dote Ricerca”. The Authors thank Debora Groppetti and Dr. Stefano Comazzi for providing samples and for assistance with flow cytometric analysis. We thank Ms Irene Ongaro for her support in the purification of AGP and Dr Alessio Scarafoni for his help with densitometric analysis. We acknowledge the very important gift of the anti-fAGP antibody from Professor David Eckersall, University of Glasgow, UK.
Footnotes
Supplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/j.rvsc.2013.04.026.
Appendix A. Supplementary data
Supplementary Fig. 1.
Supplementary Fig. 2.

Supplementary Fig. 3.
Supplementary Fig. 4.
Intra-assay imprecision of flow cytometric parameters expressing phagocytosis.
References
- Atemezem A., Mbemba E., Vassy R., Slimani H., Saffar L., Gattegno L. Human α1-acid glycoprotein binds to CCR5 expressed on the plasma membrane of human primary macrophages. Biochemical Journal. 2001;356:121–128. doi: 10.1042/0264-6021:3560121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berg A.L., Ekman K., Belák S., Berg M. Cellular composition and interferon-gamma expression of the local inflammatory response in feline infectious peritonitis (FIP) Veterinary Microbiology. 2005;111:15–23. doi: 10.1016/j.vetmic.2005.07.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceciliani F., Pocacqua V. The acute phase protein alpha1-acid glycoprotein: a model for altered glycosylation during diseases. Current Protein and Peptide Science. 2007;8:91–108. doi: 10.2174/138920307779941497. [DOI] [PubMed] [Google Scholar]
- Ceciliani F., Grossi C., Giordano A., Pocacqua V., Paltrinieri S. Decreased sialylation of the acute phase protein α1-acid glycoprotein in feline infectious peritonitis (FIP) Veterinary Immunology and Immunopathology. 2004;99:229–236. doi: 10.1016/j.vetimm.2004.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceciliani F., Pocacqua V., Miranda-Ribera A., Bronzo V., Lecchi C., Sartorelli P. Alpha(1)-acid glycoprotein modulates apoptosis in bovine monocytes. Veterinary Immunology and Immunopathology. 2007;116:145–152. doi: 10.1016/j.vetimm.2007.01.006. [DOI] [PubMed] [Google Scholar]
- Chen Y.J., Kyles A.E., Gregory C.R. In vitro evaluation of the effect of a novel immunosuppressive agent, FTY720, on the function of feline neutrophils. American Journal of Veterinary Research. 2006;67:588–592. doi: 10.2460/ajvr.67.4.588. [DOI] [PubMed] [Google Scholar]
- Craig B.W., McCord K.W., Twedt D.C. Assessment of oxidative stress in leukocytes and granulocyte function following oral administration of a silibinin-phosphatidylcholine complex in cats. American Journal of Veterinary Research. 2009;70:57–62. doi: 10.2460/ajvr.70.1.57. [DOI] [PubMed] [Google Scholar]
- De Vries B., Walter S.J., Wolfs T.G.A.M., Hochepied T., Räbinä J., Heeringa P., Parkkinen J., Libert C., Buurman W.A. Exogenous alpha-1-acid glycoprotein protects against renal ischemia-reperfusion injury by inhibition of inflammation and apoptosis. Transplantation. 2004;78:1116–1124. doi: 10.1097/01.tp.0000138096.14126.ca. [DOI] [PubMed] [Google Scholar]
- Duthie S., Eckersall P.D., Addie D.P., Lawrence C.E., Jarrett O. Value of α1-acid glycoprotein in the diagnosis of feline infectious peritonitis. Veterinary Record. 1997;141:299–303. doi: 10.1136/vr.141.12.299. [DOI] [PubMed] [Google Scholar]
- Gelain M.E., Meli M., Paltrinieri S. Whole blood cytokine profiles in cats infected by feline coronavirus and healthy non-FCoV infected specific pathogen free cats. Journal of Feline Medicine and Surgery. 2006;8:389–399. doi: 10.1016/j.jfms.2006.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giordano A., Paltrinieri S. Interferon-γ in the serum and effusions of cats with feline coronavirus infection. The Veterinary Journal. 2009;180:396–398. doi: 10.1016/j.tvjl.2008.02.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giordano A., Spagnolo V., Colombo A., Paltrinieri S. Changes in some acute phase proteins and in imunnoglobulin concentrations in cats with FIP or exposed to FCoV. The Veterinary Journal. 2004;167:38–44. doi: 10.1016/S1090-0233(03)00055-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanel R.M., Crawford P.C., Hernandez J., Benson N.A., Levy J.K. Neutrophil function and plasma opsonic capacity in colostrum-fed and colostrum-deprived neonatal kittens. American Journal of Veterinary Research. 2003;64:538–543. doi: 10.2460/ajvr.2003.64.538. [DOI] [PubMed] [Google Scholar]
- Hochepied T., Berger F.G., Baumann H., Libert C. Alpha(1)-acid glycoprotein: an acute phase protein with inflammatory and immunomodulating properties. Cytokine and Growth Factor Reviews. 2003;14:25–34. doi: 10.1016/s1359-6101(02)00054-0. [DOI] [PubMed] [Google Scholar]
- Hoffmann-Jagielska M., Winnicka A., Jagielski D., Lechowski R. Influence of dexamethasone on some cellular aspects of the immune system in cats. Veterinary Research Communications. 2003;27:643–652. doi: 10.1023/a:1027376530463. [DOI] [PubMed] [Google Scholar]
- Hoffmann-Jagielska M., Winnicka A., Jagielski D., Micuń J., Zmudzka M., Lechowski R. Influence of naturally acquired feline leukemia virus (FeLV) infection on the phagocytic and respiratory burst activity of neutrophils and monocytes of peripheral blood. Polish Journal of Veterinary Sciences. 2006;8:93–97. [PubMed] [Google Scholar]
- Jaso-Friedmann L., Leary J.H., 3rd, Praveen K., Waldron M., Hoenig M. The effects of obesity and fatty acids on the feline immune system. Veterinary Immunology and Immunopathology. 2008;122:146–152. doi: 10.1016/j.vetimm.2007.10.015. [DOI] [PubMed] [Google Scholar]
- Kagaya N., Kamiyoshi A., Tagawa Y., Akamatsu S., Isoda K., Kawase M., Yagi K. Suppression of cell death in primary rat hepatocytes by alpha1-acid glycoprotein. Journal of Biosciences and Bioenginering. 2005;99:81–83. doi: 10.1263/jbb.99.81. [DOI] [PubMed] [Google Scholar]
- Keegan R.F., Webb C.B. Oxidative stress and neutrophil function in cats with chronic renal failure. Journal of Veterinary Internal Medicine. 2010;24:514–519. doi: 10.1111/j.1939-1676.2010.0498.x. [DOI] [PubMed] [Google Scholar]
- Kipar A., Bellmann S., Kremendhal J., Khoeler K., Reinacher M. Cellular composition, coronavirus antigen expression and production of specific antibodies in lesions in feline infectious peritonitis. Veterinary Immunology and Immunopathology. 1998;65:243–257. doi: 10.1016/S0165-2427(98)00158-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kipar A., Kohler K., Leukert W., Reinacher M. A comparison of lymphatic tissues from cats with spontaneous feline infectious peritonitis (FIP), cats with FIP virus infection but no FIP, and cats with no infection. Journal of Comparative Pathology. 2001;125:182–191. doi: 10.1053/jcpa.2001.0501. [DOI] [PubMed] [Google Scholar]
- Knotek Z., Toman M., Faldyna M. Clinical and immunological characteristics of cats affected by feline infectious peritonitis. Acta Veterinaria Brno. 2000;69:51–60. [Google Scholar]
- Lecchi C., Ceciliani F., Bernasconi S., Franciosi F., Bronzo V., Sartorelli P. Bovine alpha-1 acid glycoprotein can reduce the chemotaxis of bovine monocytes and modulate CD18 expression. Veterinary Research. 2008;39:50. doi: 10.1051/vetres:2008027. [DOI] [PubMed] [Google Scholar]
- Lilliehöök I., Tvedten H. Validation of the Sysmex XT-2000iV hematology system for dogs, cats, and horses. I. Erythrocytes, platelets, and totalleukocyte counts. Veterinary Clinical Pathology. 2009;38:163–174. doi: 10.1111/j.1939-165X.2009.00125.x. [DOI] [PubMed] [Google Scholar]
- Miranda-Ribera A., Lecchi C., Bronzo V., Scaccabarozzi L., Sartorelli P., Franciosi F., Ceciliani F. Down-regulatory effect of alpha 1-acid glycoprotein on bovine neutrophil degranulation. Comparative Immunology Microbiology and Infectious Diseases. 2010;33:291–306. doi: 10.1016/j.cimid.2008.10.009. [DOI] [PubMed] [Google Scholar]
- Netea M.G., Lewis E.C., Azam T., Joosten L.A., Jaekal J., Bae S.Y., Dinarello C.A., Kim S.H. Interleukin-32 induces the differentiation of monocytes into macrophage-like cells. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:3515–3520. doi: 10.1073/pnas.0712381105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paltrinieri S. The feline acute phase reaction. The Veterinary Journal. 2008;177:26–35. doi: 10.1016/j.tvjl.2007.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paltrinieri S., Cammarata Parodi M., Cammarata G., Mambretti M. Type IV hypersensitivity in the pathogenesis of FIPV-induced lesions. Journal of Veterinary Medicine B. 1998;45:151–159. doi: 10.1111/j.1439-0450.1998.tb00778.x. [DOI] [PubMed] [Google Scholar]
- Paltrinieri S., Gelain M.E., Ceciliani F., Ribera A.M., Battilani M. Association between fecal shedding of feline coronavirus (FCoV) and serum α1-acid glycoprotein (AGP) sialylation. Journal of Feline Medicine and Surgery. 2008;10:514–518. doi: 10.1016/j.jfms.2008.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paltrinieri S., Giordano A., Tranquillo V., Guazzetti S. Critical assessment of the diagnostic value of feline α1-acid glycoprotein for feline infectious peritonitis using likelihood ratios approach. Journal of Veterinary Diagnostic Investigation. 2007;19:266–272. doi: 10.1177/104063870701900306. [DOI] [PubMed] [Google Scholar]
- Paltrinieri S., Metzger C., Battilani M., Pocacqua V., Gelain M.E., Giordano A. Serum α1-acid glycoprotein (AGP) concentration in non-symptomatic cats with feline coronavirus (FCoV) infection. Journal of Feline Medicine and Surgery. 2007;9:271–277. doi: 10.1016/j.jfms.2007.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paltrinieri S., Ponti W., Comazzi S., Giordano A., Poli G. Shifts in circulating lymphocyte subsets in cats with feline infectious peritonitis (FIP): pathogenic role and diagnostic relevance. Veterinary Immunology and Immunopathology. 2003;96:141–148. doi: 10.1016/S0165-2427(03)00156-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paltrinieri S., Marchini I., Gelain M.E. Flow cytometric detection of alpha-1-acid glycoprotein on feline circulating leucocytes. Australian Veterinary Journal. 2012;90:291–296. doi: 10.1111/j.1751-0813.2012.00948.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parra D., Rieger A.M., Li J., Zhang Y., Randall L.M., Hunter C.A., Barreda D.R., Sunyer J.O. Pivotal advance: peritoneal cavity B-1 B cells have phagocytic and microbicidal capacities and present phagocytosed antigen to CD4+ T cells. Journal of Leukocyte Biology. 2012;91:525–536. doi: 10.1189/jlb.0711372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pedersen N.C. Virologic and immunologic aspects of feline infectious peritonitis virus infection. Advances in Experimental Medicine and Biology. 1987;218:529–550. doi: 10.1007/978-1-4684-1280-2_69. [DOI] [PubMed] [Google Scholar]
- Pedersen N.C. A review of feline infectious peritonitis virus infection: 1963–2008. Journal of Feline Medicine and Surgery. 2009;11:225–258. doi: 10.1016/j.jfms.2008.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pocacqua V., Provasi E., Paltrinieri S., Gelain E., Comunian C., Ceciliani F. Glycan moiety modifications of feline alpha1-acid glycoprotein in retrovirus (FIV, FeLV) affected cats. Veterinary Immunology and Immunopathology. 2005;107:17–26. doi: 10.1016/j.vetimm.2005.02.021. [DOI] [PubMed] [Google Scholar]
- Rinaldi M., Ceciliani F., Lecchi C., Moroni P., Bannerman D.D. Differential effects of alpha1-acid glycoprotein on bovine neutrophil respiratory burst activity and IL-8 production. Veterinary Immunology and Immunopathology. 2008;126:199–210. doi: 10.1016/j.vetimm.2008.07.001. [DOI] [PubMed] [Google Scholar]
- Takano T., Hohdatsu T., Hashida Y., Kaneko Y., Tanabe M., Koyama H. A possible involvement of TNF-alpha in apoptosis induction in peripheral blood lymphocytes of cats with feline infectious peritonitis. Veterinary Microbiology. 2007;31(119):121–131. doi: 10.1016/j.vetmic.2006.08.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tilg H., Vannier E., Vachino G., Dinarello C., Mier J.J.W. Antiinflammatory properties of hepatic acute phase proteins: preferential induction of interleukin 1 (IL-1) receptor antagonist over IL-1 beta synthesis by human peripheral blood mononuclear cells. Journal of Experimental Medicine. 1993;178:1629–2636. doi: 10.1084/jem.178.5.1629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tizard I.R. seventh ed. W.B. Saunders Company; St. Louis, Missouri: 2009. Veterinary Immunology – An Introduction. pp.496. [Google Scholar]
- Vasson M.P., Roch-Arveiller M., Couderc R., Baguet J.C., Raichvarg D. Effects of α1-acid glycoprotein on human polymorphonuclear neutrophils: influence of glycan microheterogeneity. Clinica Chimica Acta. 1994;224(65):71. doi: 10.1016/0009-8981(94)90121-x. [DOI] [PubMed] [Google Scholar]
- Webb C.B., McCord K.W., Twedt D.C. Assessment of oxidative stress in leukocytes and granulocyte function following oral administration of a silibinin-phosphatidylcholine complex in cats. American Journal of Veterinary Research. 2009;70:57–62. doi: 10.2460/ajvr.70.1.57. [DOI] [PubMed] [Google Scholar]
- Westgard J.O. second ed. Westgard QC; Madison, WI: 2003. Basic Method Validation. pp. 292. [Google Scholar]
- Williams J.P., Weiser M.R., Pechet T.T., Kobzik L., Moore F.D., Jr., Hechtman H.B. Alpha 1-acid glycoprotein reduces local and remote injuries after intestinal ischemia in the rat. American Journal of Physiology. 1997;273:G1031–G1035. doi: 10.1152/ajpgi.1997.273.5.G1031. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Intra-assay imprecision of flow cytometric parameters expressing phagocytosis.