

Abstract
The present study aimed to investigate the metabolism of Forsythoside A (FTA) by human fecal bacteria to clarify the relationship between its intestinal metabolism and its pharmacological activities. FTA was incubated with human fecal microflora in vitro to investigate its metabolic process, and highly sensitive and specific ultra-performance liquid chromatography/quadrupole time-of-flight mass spectrometry (UPLC–Q-TOF/MS) was performed using MetaboLynx™ software for metabolite analysis. Caffeic acid (CA) and hydroxytyrosol (HT) were obtained by hydrolysis of FTA, and CA was further hydrogenated to form 3,4-dihydroxybenzenepropionic acid (DCA). The anticomplementary, antimicrobial and antiendotoxin activities of FTA and its metabolites by human fecal microflora were evaluated in vitro with a hemolysis assay, the agar disc-diffusion method, the MIC value and the gel clot LAL assay, respectively. The metabolites showed higher biological activity than FTA, especially HT and DCA. Orally administered FTA may be metabolized to HT and DCA, and the pharmacological effects of FTA may be dependent on intestinal bacterial metabolism.
Chemical compounds studied in this article: Forsythoside A (PubChem CID: 45358127); Caffeic acid (PubChem CID: 689043); Hydroxytyrosol (PubChem CID: 82755); 3,4-Dihydroxybenzenepropionic acid (PubChem CID: 348154)
Keywords: Forsythoside A, Intestinal bacteria, Metabolite, Anticomplement, Antimicrobial, Antiendotoxin
Graphical abstract

1. Introduction
Fructus Forsythiae, the fruit of Forsythia suspense (Thunb.) Vahl (Oleaceae), is a well-known traditional Chinese medicine (TCM) and is widely used for the treatment of infections such as acute nephritis, pharyngitis, pyrexia erysipelas, ulcer, tonsillitis and gonorrhea [1]. Remarkably, Fructus Forsythiae was recommended for the treatment and prevention of severe acute respiratory syndrome (SARS) [2] that may be caused by the inappropriate activation of complement systems [3]. So, it is hopeful to search for anticomplementary components from Fructus Forsythiae. Its main bioactive components were identified as phenylethanoid glycosides, lignans and flavonols according to phytochemical investigations [4], [5]. Forsythoside A (FTA) is one of the main phenylethanoid glycosides from Fructus Forsythiae, which possesses strong antimicrobial, antioxidant and antiviral activities [6], [7]. The chemical structure of FTA is composed of three chemical moieties including aglycone (caffeic acid), phenylethanoid aglycone (hydroxytyrosol), and sugar moieties. Its content was approximately 8% in total, which was much higher than other ingredients, but its absolute oral bioavailability was approximately 0.5%, which was significantly lower [8]. However, FTA bioavailability demonstrated a more direct relationship with the pharmacological efficacy of forsythia extract than the representative ingredients in lignans such as phillyrin [9], [10]. According to the reports, the mechanism of FTA intestinal absorption was passive diffusion and involved paracellular transport, which is mainly governed by tight junctions [11], and FTA may be a substrate of CYP3A4, CYP2C9, CYP1A2, UGT1A6, UGT1A3, UGT1A1 and UGT1A9 for in vitro metabolism in Sprague–Dawley rat liver microsomes [12]. Moreover, most herbal medicines are orally administered, and their active components are therefore brought into contact with intestinal microflora in the alimentary tract, which are mostly composed of anaerobes [13]. Some of them are transformed by intestinal bacteria before absorption from the gastrointestinal tract. Studies on herbal component metabolism by intestinal microflora are of a great importance to understanding their biological effects [14], [15]. So far, there is no report on the metabolism of FTA by intestinal bacteria. In addition, most studies on bioactivity of FTA were focused on antibacterial and antiendotoxin activities but not anticomplementary activity. In our study, FTA was anaerobically incubated with human fecal microflora, its metabolites were identified via UPLC–Q-TOF/MS and pharmacological effects were tested.
2. Experimental
2.1. Materials and methods
2.1.1. Materials and reagents
General anaerobic medium broth (GAM broth) was purchased from Shanghai Kayon Biological Technology Co., Ltd (Shanghai, China). Sheep erythrocytes, rabbit erythrocytes and anti-sheep erythrocyte antibodies were purchased from Shanghai Fortune Biological Technological Co., Ltd (Shanghai, China). Heparin (sodium salt, 160 IU/mg) was purchased from China National Medicines Co., Ltd (Shanghai, China). Normal human serum was obtained from healthy adult donors. Guinea pig serum was obtained from healthy guinea pigs from the Laboratory Animal Research Center of Fudan University. Escherichia coli (ATCC8739), Staphylococcus aureus (ATCC25923), Klebsiella pneumonia (ATCC13883) and Candida albicans (ATCC10231) were provided by the laboratory of Dr. Mei Ge.
Forsythoside A was purchased from Shanghai Winherb Medical Technology Co., Ltd.; hydroxytyrosol, caffeic acid and 3, 4-dihydroxybenzenepropionic acid were purchased from J&K Chemical Ltd.; HPLC-grade acetonitrile and methanol were purchased from Merck. Ultrapure water was prepared with a Milli-Q water purification system (Millipore). All of the other chemicals and reagents were of analytical grade.
2.1.2. Instrumentation
Analyses were performed on a commercially available ACQUITY UPLC system connected online to an ACQUITY TQD triple quadrupole mass spectrometer (Waters Corp., Milford, MA, USA) with a conditioned autosampler at 4 °C. All of the data were collected using MassLynx software. The separation was performed on a Syncronis C18 column (100 × 2.1 mm i.d., 1.7 μm; Thermo, USA) with the column temperature set to 35 °C. The mobile phase consisted of (A) ultra-pure water containing 0.1% formic acid and (B) acetonitrile using a gradient elution of 5–10% B at 0–1.5 min, 10–25% B at 1.5–8 min, 25–45% B at 8–13 min, 45–100% B at 13–18 min, 100% B at 18–21 min, and 100–5% B at 21–24 min. The flow rate was 0.4 ml/min, and the injection volume was 5 μl.
The MS instrument consisted of a Waters Synapt™ QTOF/MS (Waters Corp., Milford, MA, USA). Ionization was performed in the negative electrospray (ESI) mode. Mass range was set at m/z 50–1000 Da with a 0.28 s scan time. The ESI source was operated in negative ionization mode with capillary voltage at 2.0 kV. Source and desolvation temperatures were set at 120 and 350 °C, respectively. Nitrogen was used as desolvation and cone gas with a flow rate of 700 and 50 l/h, respectively. For accurate mass measurement, the data were centroided during acquisition using an external reference (LockSpray™) comprising a 2 μg/ml solution of leucine–enkephalin (purchased from J&K Chemical Ltd.), which was infused continuously into the ESI source at a rate of 400 μl/min via a syringe pump. The data were processed using MassLynx™ 4.1 software (Waters Corp., Milford, MA, USA).
2.2. FTA metabolism in human intestinal bacteria in vitro
Fresh fecal samples were obtained from 10 healthy volunteers (five male, five female, 22 to 50 years of age), who gave informed written consent to the study protocol. The volunteers had no history of gastrointestinal disorders and had not taken antibiotics for at least three months prior to the study. The fresh fecal mixture samples were immediately homogenized with 25 times of GAM broth. The sediments were removed by filtration through three pieces of gauze. The suspension was incubated at 37 °C in an anaerobic incubator in which the air had been replaced with a gas mixture (H2 5%, CO2 10%, N2 85%). In total, 4 mg FTA was added to the human fecal suspension (10 ml), and the mixture was incubated at 37 °C in an anaerobic incubator for 24 h. The cultured mixture was removed and extracted with water saturated n-butanol. The extract was evaporated, and the residue was dissolved in methanol (0.5 ml) and analyzed with UPLC–Q-TOF/MS.
2.3. Anti-complement activity through the classical and the alternative pathways
Based on our previous described method [16], the classical and alternative complement pathway-inhibiting properties of FTA and its metabolites were investigated. The data were reported in CH50 and AP50 to indicate the concentrations that resulted in 50% inhibition of sheep erythrocytes and the concentrations that resulted in 50% rabbit erythrocyte hemolysis inhibition, respectively. Heparin sodium salt was used as the positive control.
2.4. Antimicrobial activity by the agar disc-diffusion method minimal inhibitory concentration (MIC) determination
The antimicrobial activity assay was performed on FTA and its metabolites with five test bacterial strains, and three replicates were performed for all of the samples. The positive controls were levofloxacin for bacteria and nystatin dehydrate for fungus, and the negative control was sterile distilled water. The antimicrobial effect of all of the samples was assessed in petri dishes with 20 ml nutrient agar plus 0.2 ml microorganism suspension (108 CFU/ml, O.D 0.1, λ = 590 nm). Once the agar had solidified, 200 μl of each sample and sterile distilled water (negative control) was added to wells that were 3 mm in diameter. The plates were incubated at 37 °C for 24 h, and the inhibition halos were evaluated (mm). The antimicrobial effect was determined by measuring the inhibition halos that formed around the wells [17], [18].
The MIC value was determined by the dilution method according to the National Committee for Clinical Laboratory Standards [19]. The active samples and positive controls were dissolved in dimethyl sulfoxide at a concentration of 3200 μg/ml. Twofold serial dilutions of the solution were then prepared (3200, 1600, 800, down to 6.25 μg/ml). Nutrient broth was prepared with 1% test bacteria strains (108 CFU/ml, O.D 0.1, λ = 590 nm). In total, 10 concentrations of each sample × three test bacterial strains × three replicate wells were performed in sterile round-bottom 96-well plates by comparing the sample with the non-inoculated nutrient broth. Inoculated nutrient broth (0.9 ml) and sample (0.1 ml) were placed into each well, and the plates were incubated at 37 °C for 24 h. MIC values were determined as the lowest concentration that inhibited visible bacterial growth as detected by the unaided eye.
2.5. Antiendotoxin effect of FTA and its metabolites by the gel clot limulus amoebocyte lysate (LAL) assay
The LAL assay was performed in duplicate using commercial LAL reagent containing clotting enzyme (Zhanjiang A & C Biological Ltd., Guangdong, China). The assay was performed in pyrogen-free test tubes, to which 0.1 ml of test sample serial dilutions (1000, 500, down to 7.8 μg/ml) and 0.1 ml LAL reagent were added. The reaction solution was mixed thoroughly and placed immediately in a noncirculating water bath at 37 °C ± 1 °C for 60 ± 2 min. At the end of the incubation period, the test tubes were examined by 180° inversion for the presence of a stable solid clot. If a gel had formed and remained intact in the bottom of the reaction tube after inversion, the test was positive. The test was considered negative if a gel formed but broke or collapsed when inverted. Endotoxin standard (E. coli strain O111:B4, 0.125 EU/ml) and pyrogen-free LAL reagent water, both provided by the manufacturer, were used as controls [20].
3. Results
3.1. Metabolic processing of FTA by human intestinal bacteria in vitro
FTA was anaerobically incubated with a human fecal bacterial mixture, and the metabolic samples incubated at 6, 12 and 24 h were extracted with water-saturated n-butanol and analyzed by UPLC–Q-TOF/MS as demonstrated in Fig. 1 . In total, three peaks were observed compared with the blank sample that only contained GAM and the standard FTA solution, and their time courses were determined as demonstrated in Fig. 2 . FTA was converted to M1 and M3 at the initial time of 6 h, and M1 was increased with the incubated temporal variation. M3 degraded after incubation for 6 h, and M2 was converted at the same time. The content change between M2 and M3 was exactly opposite, indicating a mutual transformation between these two metabolites.
Fig. 1.

TIC chromatograms of FTA incubated with human intestinal bacteria.
Fig. 2.
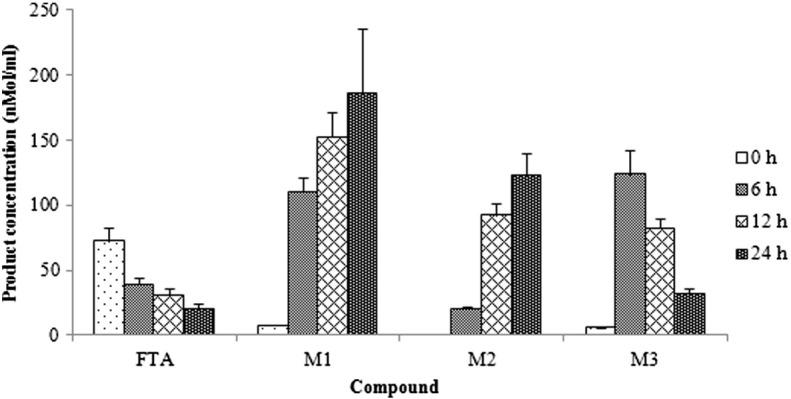
Time courses of FTA production over 24 h after incubation with human intestinal bacteria. The data are presented as the mean ± SD.
3.2. Identification of FTA metabolites
FTA metabolites M1–M3 were analyzed by UPLC–MS, which is demonstrated in Table 1 . The negative electrospray mass spectrum of FTA revealed a [M–H]− ion at m/z 623. The full mass of M3, which was detected at 2.83 min, gave a [M–H]− ion at m/z 179, indicating the loss of glucopyranose, rhamnose and C8H10O3 from FTA to form caffeic acid (CA). The molecular ion of M1 [M–H]− was 153, indicating the loss of glucopyranose, rhamnose and CA from FTA to form hydroxytyrosol (HT). The [M–H]− ion of M2 was at m/z 181, indicating the hydrogenation reaction of CA to form 3,4-dihydroxybenzenepropionic acid (DCA). To further confirm the chemical structure, three metabolites were identified by comparing the retention times and UV spectra with reference substances based on MS data. Those data are summarized in Table 1, deglycosylated and hydrogenated metabolites were present with human fecal incubation.
Table 1.
Retention time (RT), ion mass peaks and peak area of FTA and its metabolites.
| No. | RT (min) | [M –H]− m/z | Metabolite name | Formula | Area (%) |
|---|---|---|---|---|---|
| Parent Compound | 5.58 | 623.1990 | FTA | C29H36O15 | 2.75 |
| M1 | 1.80 | 153.0545 | HT | C8H10O3 | 0.93 |
| M2 | 2.73 | 181.0509 | DCA | C9H10O4 | 0.16 |
| M3 | 2.83 | 179.0335 | CA | C9H8O4 | 0.47 |
3.3. FTA metabolism with human intestinal bacteria
Based on the results above, FTA metabolism by human intestinal bacteria was proposed to function as follows (Fig. 3 ). FTA was first desugared to form CA and large amounts of HT, and CA was further hydrogenated to form DCA.
Fig. 3.

The possible metabolic pathway of FTA incubated with human intestinal bacteria.
3.4. Classical and alternative anticomplementary pathway activity
The effect of FTA and its metabolites on guinea pig complement activation through the classical pathway was examined in 1:40 diluted guinea pig serum, and heparin sodium was used as the positive control. The percent of activation that 1:40 diluted guinea serum occurred in the classical pathway was 98.34 ± 5.23% in the complement control group. The percentage of activation that 1:10 diluted normal human serum occurred in the alternative pathway was 97.26 ± 7.85% in the complement control group. As Table 2 demonstrates, FTA demonstrated no anticomplementary activity for either the classical pathway or the alternative pathway. HT, CA and DCA all exhibited complement modulation properties in a dose-dependent manner. The CH50 were 0.096 ± 0.007 mg/ml, 0.321 ± 0.011 mg/ml and 0.923 ± 0.037 mg/ml for HT, DCA and CA, respectively. In the AP50 assay, the concentrations that resulted in 50% hemolysis inhibition of rabbit erythrocytes were 0.121 ± 0.012 mg/ml for HT, 0.427 ± 0.023 mg/ml for DCA and 1.038 ± 0.131 mg/ml for CA, respectively. HT demonstrated the most significant activity comparable to the positive inhibitor (heparin sodium, CH50 = 0.068 ± 0.002 mg/ml; AP50 = 0.086 ± 0.003 mg/ml).
Table 2.
Anticomplementary activity of FTA and its metabolites by human intestinal bacteria.
| Sample | CH50 (mg/ml)a | AP50 (mg/ml)a |
|---|---|---|
| FTA | NEb | NE |
| HT | 0.096 ± 0.007 | 0.121 ± 0.012 |
| DCA | 0.321 ± 0.011 | 0.427 ± 0.023− |
| CA | 0.923 ± 0.037 | 1.038 ± 0.131 |
| Heparin sodium | 0.068 ± 0.002 | 0.086 ± 0.003 |
The data are expressed as the mean ± SD of independent experiments performed in triplicate.
NE denotes that this compound has no inhibitory effect at the maximal concentration tested.
3.5. Determination of the antimicrobial activity of FTA and its metabolites
The primary antimicrobial screening was performed using the agar well diffusion method. The antimicrobial activity of FTA and its metabolites was compared with levofloxacin and nystatin dihydrate as standard antibiotics. As demonstrated in Table 3 , none of the tested samples exhibited any activity against C. albicans and K. pneumonia, but some of them possessed inhibitory activity against E. coli and S. aureus. HT demonstrated inhibitory effects against the Gram-positive bacteria S. aureus and the Gram-negative bacteria E. coli, while FTA and DCA were limited to S. aureus. The MIC values of FTA, HT and DCA against the bacteria are summarized in Table 4 . HT displayed the highest antimicrobial activity overall followed by FTA and DCA.
Table 3.
Antimicrobial activity of FTA and its metabolites by human intestinal bacteria.
| Samples | Zone of inhibition (cm) |
|||
|---|---|---|---|---|
| E. coli | S. aureus | K. pneumonia | C. albicans | |
| FTA | NEa | 0.8 | NE | NE |
| CA | NE | NE | NE | NE |
| HT | 1.2 | 1.6 | NE − | NE |
| DCA | NE | 1.1 | NE | NE |
| Levofloxacin | 2.8 | 2.3 | 2.1 | NE |
| Nystatin Dihydrate | NE | NE | NE | 2.2 |
NE denotes that this compound has no inhibitory effect at the maximal concentration tested.
Table 4.
MIC of FTA and its metabolites by human intestinal bacteria.
| Samples |
MIC (μg/ml) |
|
|---|---|---|
| E. coli | S. aureus | |
| FTA | NEa | 50 |
| HT | 50 | 25 |
| DCA | NE | 50 |
| Levofloxacin | 0.1 | 1.5 |
NE denotes that this compound has no inhibitory effect at the maximal concentration tested.
3.6. Antiendotoxin effect of FTA and its metabolites
The antiendotoxin effect was determined by the Gel clot LAL assay in vitro. As the results presented in Table 5 , FTA and CA were invalid even at the maximal concentration. However, HT and DCA were still effective when diluted four and three times, respectively. HT possessed the highest antiendotoxin effect among the four compounds.
Table 5.
Antiendotoxin effect of FTA and its metabolites by human bacteria.
| Samples | Tubes no. |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1/2 | 1/4 | 1/8 | 1/16 | 1/32 | 1/64 | 1/128 | Positive | Negative | |
| FTA | + + | + + | + + | + + | + + | + + | + + | + + | + + | − − |
| CA | + + | + + | + + | + + | + + | + + | + + | + + | + + | − − |
| HT | − − | − − | − − | − − | + + | + + | + + | + + | + + | − − |
| DCA | − − | − − | − − | + + | + + | + + | + + | + + | + + | − − |
4. Discussion
FTA, one of the main, most abundant active ingredients in Fructus Forsythiae, had low bioavailability. There were many physiological factors such as bile, intestinal microflora, and the gastrointestinal tract pH, which would be related to drug bioavailability [21]. According to the present study, after oral administration, FTA was first metabolized into CA and HT in anaerobic conditions, and then CA was converted to DCA by human fecal bacteria. The incubation process as well as the activity tests are under 37 °C or at low temperatures due to FTA instability at high temperature. Fortunately, no metabolites in our study was found (Figure not given) when FTA was incubated with blank that only contained GAM. Accordingly, the intestinal bacterial metabolism played an important role in the generation of three metabolites. FTA was proved to deglycosylated to CA and HT under the existence of glycosidase in the intestinal microflora. The same metabolic behavior was observed in naringin [22] and anthocyanin metabolism [23]. Hydrogenation also played an important role in CA metabolism in our study. These results were different from the Parkar's report [24], which demonstrated that CA may be metabolized into 3-hydroxyphenylpropionic acid and DCA mainly when incubated for 24 h with human fecal bacteria. This difference may due to different substrate concentrations or the presence of HT during the metabolic process.
So far, there is no report on the anticomplementary effect of FTA. The anticomplementary activities of phenylethanoid glycosides from Monochasma savatieri have previously been investigated [25]. In the present study, HT, as a main human intestinal bacteria-derived metabolite of FTA, demonstrated considerable anticomplementary activity compared with the positive control drug heparin sodium for the first time. HT is present in olive oil mainly as secoiridoid derivative, and it is the most active polyphenol from olive tree leaves, olive pulp and olive oil [26]. Interestingly, previous reports indicated that extracts of olive (Olea europaea L., Oleaceae) leaves demonstrated anticomplementary activity [27]. Accordingly, HT was indispensable in the anticomplementary effect of olive leaves. Conversely, the metabolite CA had a moderate effect to inhibit the classical and alterative pathways of the complement system, which is consistent with the previous report on the anticomplementary effect study of compounds from Gnaphalium affine D. Don [28].
In our study, FTA demonstrated inhibition against the Gram-positive bacteria S. aureus but no effect against the Gram-negative bacteria E. coli and K. pneumonia as well as C. albicans. The results indicated that S. aureus was the most susceptible to FTA among the four strains tested. HT possessed the highest antibacterial activity against the Gram-positive bacteria S. aureus and the Gram-negative bacteria E. coli. This result was consistent with what has been reported by Papadopoulou [29]. Other phenolic compounds have also demonstrated good activity against S. aureus such as epigallocatechin-3-O-gallate [30] and oleuropein [31]. DCA, a major metabolite, had the same antibacterial effect against S. aureus as FTA, which supports that phenylpropionic acid was more active against Gram-positive bacteria [32]. Characterization of the interaction between these antimicrobial compounds and their target sites also need to be further studied.
LAL assays are the most sensitive assays that are currently known for endotoxin [33]. Three types of LAL endotoxin testing methods (the gel-clot method, the chromogenic method, and the turbidimetric method) have been approved by the United States Pharmacopeia for the evaluation of end-product injectable drugs, medical devices, and raw materials [34]. The gel-clot method is thought of as the most accurate and sensitive procedure for determining endotoxin content because fewer false-positive and false-negative results are observed when that method is used [35]. Our study reveals that among FTA and its intestinal bacterial metabolites, HT and DCA had an antagonistic effect against endotoxin. DCA, a possible metabolite of CA containing the o-dihydroxyl structure, expressed higher antiendotoxin, anticomplementary and antibacterial effects than CA. Previous results suggested that the olefinic double bond in CA is an important factor for enhancing antioxidant activity [36]. Thus, it is likely that the 2,3-double bond of CA is critical for the pharmacodynamic expression of DCA.
DCA was reportedly found in human plasma as a metabolite of CA after dietary supplementation and may have been formed by intestinal bacteria [37]. Wang et al. [38] also demonstrated that HT and CA were identified as two major metabolites of echinacoside (a typical phenylethanoid glycoside) in rat urine and feces samples. What's more, anticomplementary and antiendotoxin properties of HT and DCA were determined for the first time in our study. Therefore, both DCA and HT were considered to be possible and effective metabolites in the body after oral administration of phenylethanoid glycosides such as FTA. The present result indicated that the biological activities were increased when FTA was metabolized to HT and DCA by human intestinal bacteria, which were coincident with the previous report that the intestinal flora played an important role in the metabolism of orally administered or bile-excreted compounds [39]. Herbal components should be transformed to bioactive compounds by intestinal bacteria and to perform the pharmacological action of herbal medicines [40], [41]. This conclusion might help explain that FTA demonstrated a positive correlation between dose and biological activity in Forsythia suspense [42].
5. Conclusion
Our results demonstrated that the presence of intestinal bacteria played an important role in FTA metabolism after oral administration. The metabolic process is involved in hydrolytic reactions and the generation of non-polar low-molecular-weight byproducts, and FTA was accompanied by active metabolites. The results indicated that the pharmacological effects of FTA may be dependent on the metabolism of FTA by intestinal bacteria, which also clarified the low FTA bioavailability. However, further studies should be performed to elucidate anticomplementary and antimicrobial activity in vivo as well as the mechanism of action related to pharmacological activity.
Acknowledgments
This work was financially supported by the National Project in Significant Creation of New Drugs during the Eleventh Five-Year Plan Period (2009ZX09502-013). The authors are grateful to the laboratory of Dr. Mei Ge for providing all of the bacterial strains and technical support of the antimicrobial tests and Dr. Lei Feng from the Instrumental Analysis Center of Shanghai Jiao Tong University for the sample analysis.
References
- 1.Jiao J., Gai Q.Y., Fu Y.J., Zu Y.G., Luo M., Zhao C.J. Microwave-assisted ionic liquids treatment followed by hydro-distillation for the efficient isolation of essential oil from Fructus forsythiae seed. Sep Purif Technol. 2013;107:228–237. [Google Scholar]
- 2.Lau T.F., Leung P.C., Wong E.L., Fong C., Cheng K.F., Zhang S.C. Using herbal medicine as a means of prevention experience during the SARS crisis. Am J Chin Med. 2005;33:345–356. doi: 10.1142/S0192415X05002965. [DOI] [PubMed] [Google Scholar]
- 3.Chen D.F. Isolation and characterization of anti-complementary agents from medicinal plants. Planta Med. 2011;77:S9. [Google Scholar]
- 4.Guo H., Liu A.H., Li L., Guo D.A. Simultaneous determination of 12 major constituents in Forsythia suspensa by high performance liquid chromatography-DAD method. J Pharm Biomed. 2007;43:1000–1006. doi: 10.1016/j.jpba.2006.09.033. [DOI] [PubMed] [Google Scholar]
- 5.Xia Y., Yang B., Wang Q., Liang J., Wei Y., Yu H. Quantitative analysis and chromatographic fingerprinting for the quality evaluation of Forsythia suspensa extract by HPLC coupled with photodiode array detector. J Sep Sci. 2009;32:1–13. doi: 10.1002/jssc.200900488. [DOI] [PubMed] [Google Scholar]
- 6.Ma Y.Y., Zhang Z.W., Li H.W., Sun J.H., Xu C.Y., Wu G.J. Effects of forsythoside A on the expression of IFN-α and Mx1. Sci Agric Sin. 2010;43:3237–3243. [Google Scholar]
- 7.Jiao J., Fu Y.J., Zu Y.G., Luo M., Wang W., Zhang L. Enzyme-assisted microwave hydro-distillation essential oil from Fructus forsythia, chemical constituents, and its antimicrobial and antioxidant activities. Food Chem. 2012;134:235–243. [Google Scholar]
- 8.Wang G.N., Pan R.L., Liao Y.H., Chen Y., Tang J.T., Chang Q. An LC–MS/MS method for determination of forsythiaside in rat plasma and application to a pharmacokinetic study. J Chromatogr B. 2010;878:102–106. doi: 10.1016/j.jchromb.2009.11.029. [DOI] [PubMed] [Google Scholar]
- 9.Zhou W., Qin K.M., Shan J.J., Ju W.Z., Liu S.J., Cai B.C. Improvement of intestinal absorption of forsythoside A in weeping forsythia extract by various absorption enhancers based on tight junctions. Phytomedicine. 2012;20:47–58. doi: 10.1016/j.phymed.2012.09.014. [DOI] [PubMed] [Google Scholar]
- 10.Qu H., Zhang Y., Wang Y., Li B., Sun W. Antioxidant and antibacterial activity of two compounds (forsythiaside and forsythin) isolated from Forsythia suspense. J Pharm Pharmacol. 2008;60:261–267. doi: 10.1211/jpp.60.2.0016. [DOI] [PubMed] [Google Scholar]
- 11.Zhou W., Di L.Q., Wang J., Shan J.J., Liu S.J., Ju W.Z. Intestinal absorption of forsythoside A in in situ single-pass intestinal perfusion and in vitro Caco-2 cell models. Acta Pharmacol Sin. 2012;33:1069–1079. doi: 10.1038/aps.2012.58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhou W., Di L.Q., Shan J.J., Bi X.L., Chen L.T., Wang L.C. In vitro metabolism in Sprague–Dawley rat liver microsomes of forsythoside A in different compositions of Shuang-Huang-Lian. Fitoterapia. 2011;82:1222–1230. doi: 10.1016/j.fitote.2011.08.009. [DOI] [PubMed] [Google Scholar]
- 13.Akao T., Hayashi T., Kobashi K. Intestinal bacterial hydrolysis is indispensable to absorption of 18 beta-glycyrrhetic acid after oral administration of glycyrrhizin in rats. J Pharm Pharmacol. 1994;46:135–137. doi: 10.1111/j.2042-7158.1994.tb03756.x. [DOI] [PubMed] [Google Scholar]
- 14.Kim D.H., Jung E.A., Sohng I.S., Han J.A., Kim T.H., Han M.J. Intestinal bacterial metabolism of flavonoids and its relation to some biological activities. Arch Pharm Res. 1998;21:17–23. doi: 10.1007/BF03216747. [DOI] [PubMed] [Google Scholar]
- 15.Trinh H.T., Jang S.Y., Han M.J., Kawk H.Y., Beak N.I., Kim D.H. Metabolism of wogonoside by human fecal microflora and its anti-pruritic effect. Biomol Ther. 2009;17:211–216. [Google Scholar]
- 16.Xing S.H., Wang M.Y., Peng Y., Chen D.F., Li X.B. Simulated gastrointestinal tract metabolism and pharmacological activities of water extract of Scutellaria baicalensis roots. J Ethnopharmacol. 2014;152:183–189. doi: 10.1016/j.jep.2013.12.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Boussaada O., Chriaa J., Nabli R., Ammar S., Saidana D., Mahjoub M.A. Antimicrobial and antioxidant activities of methanol extracts of Evax pygmaea (Asteraceae) growing wild in Tunisia. World J Microbiol Biotechnol. 2008;24:1289–1296. [Google Scholar]
- 18.Murray P.R., Baron E.J., Pfaller M.A., Tenover F.C., Yolken R.H. American Society for Microbiology; Washington DC, USA: 1995. Manual of clinical microbiology; pp. 1327–1341. [Google Scholar]
- 19.National Committee for Clinical Laboratory Standards . National Committee for Clinical Laboratory Standards; Villanova, PA, USA: 1985. Approved standard document M7A. [Google Scholar]
- 20.Joiner T.J., Kraus P.F., Kupiec T.C. Comparison of endotoxin testing methods for pharmaceutical products. Int J Pharm Compd. 2002;6:408–409. [PubMed] [Google Scholar]
- 21.Hurst S., Loi C.M., Brodfuehrer J., El-Kattan A. Impact of physiological, physicochemical and biopharmaceutical factors in absorption and metabolism mechanisms on the drug oral bioavailability of rats and humans. Expert Opin Drug Metab Toxicol. 2007;3:469–489. doi: 10.1517/17425225.3.4.469. [DOI] [PubMed] [Google Scholar]
- 22.Jang I.S., Kang K.H., Sung C.K., Kim D.H. Metabolism of saikosaponin c and naringin by human intestinal bacteria. Arch Pharm Res. 1997;20:420–424. doi: 10.1007/BF02973933. [DOI] [PubMed] [Google Scholar]
- 23.Keppler K., Humpf H.U. Metabolism of anthocyanins and their phenolic degradation products by the intestinal microflora. Bioorg Med Chem. 2005;13:5195–5205. doi: 10.1016/j.bmc.2005.05.003. [DOI] [PubMed] [Google Scholar]
- 24.Parkar S.G., Trower T.M., Stevenson D.E. Fecal microbial metabolism of polyphenols and its effects on human gut microbiota. Anaerobe. 2013;23:12–19. doi: 10.1016/j.anaerobe.2013.07.009. [DOI] [PubMed] [Google Scholar]
- 25.Li M., Shi M.F., Liu Y.L., Xu Q.M., Yang S.L. Phenylethanoid glycosides from Monochasma savatieri and their anticomplement activity through the classical pathway. Planta Med. 2012;78:1381–1386. doi: 10.1055/s-0032-1314982. [DOI] [PubMed] [Google Scholar]
- 26.Granados-Principal S., Quiles J.L., Ramirez-Tortosa C.L., Sanchez-Rovira P., Ramirez-Tortosa M.C. Hydroxytyrosol: from laboratory investigations to future clinical trials. Nutr Rev. 2010;68:191–206. doi: 10.1111/j.1753-4887.2010.00278.x. [DOI] [PubMed] [Google Scholar]
- 27.Pieroni A., Heimler D., Pieters L., van Poel B., Vlietinck A.J. In vitro anti-complementary activity of flavonoids from olive (Olea europaea L.) leaves. Pharmazie. 1996;51:765–768. [PubMed] [Google Scholar]
- 28.Li J., Huang D., Chen W., Xi Z., Chen C., Huang G. Two new phenolic glycosides from Gnaphalium affine D. Don and their anti-complementary activity. Molecules. 2013;18:7751–7760. doi: 10.3390/molecules18077751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Papadopoulou C., Soulti K., Roussis I.G. Potential antimicrobial activity of red and white wine phenolic extracts against strains of Staphylococcus aureus, Escherichia coli and Candida albicans. Food Technol Biotechnol. 2005;43:41–46. [Google Scholar]
- 30.Taguri T., Tanaka T., Kouno I. Antibacterial spectrum of plant polyphenols and extracts depending upon hydroxyphenyl structure. Biol Pharm Bull. 2006;29:2226–2235. doi: 10.1248/bpb.29.2226. [DOI] [PubMed] [Google Scholar]
- 31.Bisignano G., Tomaino A., Lo Cascio R., Crisafi G., Uccella N., Saija A. On the in-vitro antimicrobial activity of oleuropein and hydroxytyrosol. J Pharm Pharmacol. 1999;51:971–974. doi: 10.1211/0022357991773258. [DOI] [PubMed] [Google Scholar]
- 32.Cueva C., Moreno-Arribas M.V., Martín-Alvarez P.J., Bills G., Vicente M.F., Basilio A. Antimicrobial activity of phenolic acids against commensal, probiotic and pathogenic bacteria. Res Microbiol. 2010;161:372–382. doi: 10.1016/j.resmic.2010.04.006. [DOI] [PubMed] [Google Scholar]
- 33.Firca J.R., Rudbach R.J. Liss; New York: 1982. Endotoxins and their detection with the Lymulus Amebocyte Lysate Test; pp. 121–128. [Google Scholar]
- 34.Hausmann M.J., Yulzari R., Lewis E., Saisky Y., Douvdevani A. Gel clot LAL assay in the initial management of peritoneal dialysis patients with peritonitis: a retrospective study. Nephrol Dial Transplant. 2000;15:680–683. doi: 10.1093/ndt/15.5.680. [DOI] [PubMed] [Google Scholar]
- 35.Morris H.C., Monaco L.A., Steele A., Wainwright N. Setting a standard: the limulus amebocyte lysate assay and the assessment of microbial contamination on spacecraft surfaces. Astrobiology. 2010;10:845–852. doi: 10.1089/ast.2009.0446. [DOI] [PubMed] [Google Scholar]
- 36.Moon J.H., Terao J. Antioxidant activity of caffeic acid and dihydrocaffeic acid in lard and human low-density lipoprotein. J Agric Food Chem. 1998;46:5062–5065. [Google Scholar]
- 37.Goldstein D.S., Stull R., Markey S.P., Marks E.S., Keiser H.R. Dihydrocaffeic acid: a common contaminant in the liquid chromatographic–electrochemical measurement of plasma catecholamines in man. J Chromatogr. 1984;311:148–153. doi: 10.1016/s0378-4347(00)84701-5. [DOI] [PubMed] [Google Scholar]
- 38.Wang Y., Hao H., Wang G., Tu P., Jiang Y., Liang Y. An approach to identifying sequential metabolites of a typical phenylethanoid glycoside, echinacoside, based on liquid chromatography–ion trap-time of flight mass spectrometry analysis. Talanta. 2009;80:572–580. doi: 10.1016/j.talanta.2009.07.027. [DOI] [PubMed] [Google Scholar]
- 39.Zuo F., Zhou Z.M., Yan M.Z., Liu L.M., Xiong Y.L., Zhang Q. Metabolism of constituents in Huangqin-Tang, a prescription in Traditional Chinese Medicine, by human intestinal flora. Biol Pharm Bull. 2002;25:558–563. doi: 10.1248/bpb.25.558. [DOI] [PubMed] [Google Scholar]
- 40.Lee D.S., Kim Y.S., Ko C.N., Cho K.H., Bae H.S., Lee K.S. Fecal metabolic activities of herbal components to bioactive compounds. Arch Pharm Res. 2002;25:165–169. doi: 10.1007/BF02976558. [DOI] [PubMed] [Google Scholar]
- 41.Sousa T., Paterson R., Moore V., Carlsson A., Abrahamsson B., Basit A.W. The gastrointestinal microbiota as a site for the biotransformation of drugs. Int J Pharm. 2008;363:1–25. doi: 10.1016/j.ijpharm.2008.07.009. [DOI] [PubMed] [Google Scholar]
- 42.Zhou W., Zhu X.X., Yin A.L., Cai B.C., Wang H.D., Di L.Q. Effect of various absorption enhancers based on tight junctions on the intestinal absorption of forsythoside A in Shuang-Huang-Lian, application to its antivirus activity. Pharmacogn Mag. 2014;10:9–17. doi: 10.4103/0973-1296.126651. [DOI] [PMC free article] [PubMed] [Google Scholar]