

Highlights
-
•
Affordable, safe & potent Measles vectored HPV vaccine developed & compared.
-
•
This vaccine found immunogenic similar to classical HPV protein vaccine from Pichia.
-
•
It can be given as childhood vaccine at 9–12 months of age like Measles.
-
•
It is simple, quick and cheap to produce in high volumes for mass vaccination.
Abbreviations: HPV, human papilloma virus; CC, cervical cancer; MV, measles virus; rMV, recombinant MV; EZ, Edmonston Zagreb strain; IFN-ϒ, interferon gamma; IL-5, interleukin 5; IC50, concentration that gives a response halfway between baseline and maximal; N, nucleocapsid protein; P, phosphoprotein; M, matrix protein; F, fusion protein; H, attachment protein; NNSV, nonsegmented negative strand RNA virus
Keywords: Recombinant measles, Pichia expressed, HPV16L1, HPV18L1, Prophylactic vaccine
Abstract
Human papillomavirus (HPV) infection is the most common sexually transmitted disease worldwide. HPVs are oncogenic small double-stranded DNA viruses that are the primary causal agent of cervical cancer and other types of cancers, including in the anus, oropharynx, vagina, vulva, and penis. Prophylactic vaccination against HPV is an attractive strategy for preventing cervical cancer and some other types of cancers. However, there are few safe and effective vaccines against HPV infections. Current first-generation commercial HPV vaccines are expensive to produce and deliver.
The goal of this study was to develop an alternate potent HPV recombinant L1-based vaccines by producing HPV virus-like particles into a vaccine that is currently used worldwide. Live attenuated measles virus (MV) vaccines have a well-established safety and efficacy record, and recombinant MV (rMV) produced by reverse genetics may be useful for generating candidate HPV vaccines to meet the needs of the developing world.
We studied in non-human primate rMV-vectored HPV vaccine in parallel with a classical alum adjuvant recombinant HPV16L1 and 18L1 protein vaccine produced in Pichia pastoris. A combined prime-boost approach using both vaccines was evaluated, as well as immune interference due to pre-existing immunity against the MV.
The humoral immune response induced by the MV, Pichia-expressed vaccine, and their combination as priming and boosting approaches was found to elicit HPV16L1 and 18L1 specific total IgG and neutralizing antibody titres. Pre-existing antibodies against measles did not prevent the immune response against HPV16L1 and 18L1.
1. Introduction
Persistent infections sustained by “high-risk” genotypes of human papilloma viruses (HPVs) lead to the development of high-grade squamous intraepithelial lesions and ultimately cause cervical cancer (CC). Worldwide, approximately 70% of CCs are caused by the HPV16 or HPV18 serotypes; the remaining 30% are caused by at least 10 other high-risk HPV subtypes [1], [2], [3].
L1, the major capsid protein of HPV, capable to self-assemble into virus-like particles (VLPs), is a type-specific and highly immunogenic antigen, and can elicit high titres of neutralizing antibodies, conferring protection against HPV infection [4], [5].
Currently, prophylactic HPV vaccines based on L1 structural proteins are well established to prevent CC. The two licensed HPV vaccines, quadrivalent Gardasil from Merck (qHPV-6/11/16/18 vaccine; Kenilworth, NJ, USA) and bivalent Cervarix form GlaxoSmithKline (bHPV-16/18 vaccine; Brentford, UK), were approved in many countries and found to be safe and effective for preventing infection by vaccine-related HPV serotypes [6]. However, administration has been virtually non-existent in developing countries, mainly because of the prohibitively high cost of the vaccine and difficulties in implementing vaccination programs, which require three injections for adolescent girls, over a six-month period [6]. The recently approved nonavalent vaccine (Gardasil 9, Merck) provides protection against 5 additional oncogenic types (HPV 31, 33, 45, 52, 58) [7], [8]; however, it possesses the same difficulties as its predecessor.
Therefore, next-generation HPV vaccines are required to reduce production costs and improve immunization schedule feasibility.
This study aimed to develop a cost-effective, safe, and effective prophylactic HPV vaccine to address the needs of developing countries. We developed a recombinant HPV vaccine based on a live attenuated measles virus (MV)-vectored platform [9]. Additionally, we compared its immune response to that of the classical Pichia pastoris-produced HPV16L1 and 18L1 recombinant vaccines in non-human primates.
Live attenuated MV vaccines have a well-established safety and efficacy record and are inexpensively produced and delivered to most developing countries. MV vaccines induce life-long immunity after a single injection, and reversion to pathogenicity has never been observed [10]. In 2014, the WHO estimated that the global immunization coverage with at least the first dose of live attenuated MV vaccine was higher than 85%.
MV is an enveloped virus with a nonsegmented, negative-sense single-stranded RNA genome of 15,894 nucleotides in length. The genome contains six transcription units, which are separated by nontranscribed trinucleotide intergenic sequences and are flanked by 3′ leader and 5′ trailer sequences at the genome ends. The MeV genome encodes a total of eight proteins. The six structural proteins are the nucleocapsid protein (N), phosphoprotein (P), matrix protein (M), fusion protein (F), attachment protein (H), and the large RNA-dependent RNA polymerase protein (L). Two additional nonstructural proteins (C and V) are encoded in the P transcription unit [11].
Recombinant-MVs (rMV) produced by reverse genetics [12], [13] represent an attractive platform for generating multivalent vaccines to immunize against measles and other infectious agents or to carry therapeutic agents [14], [15], [16]. As a proof of concept, MVs have been used to express antigens derived from HIV [17], [18], hepatitis B [14], West Nile virus [19], SARS coronavirus [20], and Chikungunya virus [21], [22]. In most of these studies, rMVs expressing heterologous antigens appeared to induce long-term specific neutralizing antibodies, even in the presence of pre-existing immunity against MV [23], [24]. In a previous study, our rMV-HPV16L1 vaccine candidate was found to effectively induce specific neutralizing antibodies against HPV16L1 [9], [25] in transgenic CD46IFNAR mice susceptible to MV infection [26]. Here, we examined the development of the recombinant live attenuated MV Edmonston-Zagreb (rMVEZ) strain as a viral vector to carry heterologous genes encoding the major capsid protein L1 of HPV-type-16 and HPV-type-18 in rhesus monkeys and compared their immunogenicity in a traditional methylotrophic P. pastoris-based expression system [27].
2. Materials and methods
2.1. Cloning, rescue, characterization and production of measles vectored HPV L1 candidate vaccine viruses
Human-codon optimized genes coding for the HPV16L1 and HPV18L1 antigens were chemically synthesized at GeneArt™ (Thermo Fisher Scientific, USA). Plasmid p(+)MVEZ, delivering correctly T7-initiated and ribozyme-terminated MV antigenome, was constructed as previously described [12]. BssHII-AatII cloning of the synthetic genes as additional transcription units was in position 2 (between the P and M protein genes) of p(+)MVEZ [Fig. 1 ]. The methods used for cloning were similar to those described by Sambrook and Russell [28]. The ligated products were transformed into Escherichia coli XL10-Gold (Stratagene, USA) Ultra Competent cells. Growing colonies selected by ampicillin (100 μg/mL) resistance were screened by BssHII-AatII digestion after minipreparation of plasmid DNAs, following the Qiagen Plasmid Purification protocol. Recombinant positive clones, named p(+)MVEZHPV16L1 and p(+)MVEZHPV18L1, were used to transform XL10-Gold cells, amplified by maxipreparation following the Qiagen Plasmid Purification protocol, assayed by analytical digestion, and confirmed by primer-walking sequencing (Eurofins MWG Biotech, Germany). The rMVEZHPV16L1 and rMVEZHPV18L1 viruses were then rescued by 293T cells transfection in a five-plasmid transfection system. Single syncytia were expanded in the MRC-5 cell line to establish passage 0 (P0); the recombinant viruses were serially passaged to prepare sufficient virus stocks. The rMVEZHPVs were quantified by CCID50 microtitration in 96-well plates by Spearman and Karber formula [29].
Fig. 1.
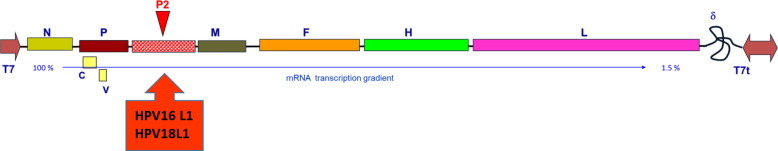
Schematic representation of rMVEZ antigenome. N: nucleocapsid protein, P: phosphoprotein, M: matrix protein, F: fusion protein, H: attachment protein, L: large RNA-dependent RNA polymerase protein, C and V represents nonstructural proteins encoded in the P transcription unit. P2 represents position in measles vector to insert HPV16L1 or HPV18L1 genes as additional transcription unit.
Media and cell lysate from Vero cells infected with either rMVEZHPV16L1 or MVEZHPV18L1 were lysed at 48 h post-infection. They were analyzed by 10% SDS–PAGE to determine the expression of HPV-specific L1 antigen by western blotting. HPV16L1 and HPV18L1 were detected with specific mouse monoclonal antibodies (Abcam, UK; Genetex, USA), by Qdot 625 Western Blot Kit (Thermo Fisher Scientific).
2.2. Cloning, expression, characterization and production of P. pastoris based HPV L1 vaccine candidates
The yeast-codon optimized HPV16 L1 and HPV18 L1 transgenes were assembled by de novo synthesis (GeneArtTM, Thermo Fisher Scientific), and cloned by BstBI/NotI (New England Biolabs, Ipswich, MA, USA) into the pPICZα expression vector (Thermo Fisher Scientific) under the control of the AOX1 promoter. The methods used for cloning were similar to those described by Sambrook and Russell [28]. The obtained pPICZ-HPV16L1 and pPICZ-HPV18L1 plasmids, propagated in E. coli and selected by Zeocin (25 μg/mL, Thermo Fisher Scientific) resistance, were linearized by SacI digestion and used to transform electrocompetent P. pastoris KM71 by electroporation (Bio-Rad GenePulser II, USA), at 1.7 kV, 25 μF, and 200 Ω. Positive clones were primary selected as resistant transformants by replica plating at different Zeocin concentrations (from 150 to 2000 μg/mL). Next, the clones were confirmed by semi-quantitative PCR and RT-PCR to assess the gene copy number integrated into the Pichia genome.
After methanol induction, the cells were lysed by acid-washed glass-beads (0.5 mm) in lysis buffer (50 mM sodium phosphate, 1 mM PMSF, 1 mM EDTA, 5% glycerol) and centrifuged to clarify the supernatant. Protein expression in the clones was confirmed by 10% SDS–PAGE under reducing conditions and by western blotting with specific mouse monoclonal antibodies (Abcam) against HPV16/18L1.
Master and working cell banks were prepared in yeast peptone dextrose media, whereas routine production batches were produced in chemically defined synthetic media by batch fermentation in 3 substages. The cells were induced by methanol for protein expression. The batch was harvested, pelleted, lysed by a high-pressure homogenizer at 28 kpsi, and clarified. The clarified lysate was loaded onto cation exchange resin to purify the L1 antigens and sterile-filtered in a 0.2 μm filter. Final concentrates were stored at 2–8 °C. To assess VLP formation, the purified HPV16L1/18L1 proteins were negatively stained with 1% uranyl acetate and examined under an electron microscope.
2.3. Final formulations of vaccine candidates
2.3.1. Recombinant measles vectored HPV
rMVEZHPV (16L1 and 18L1) virus stocks stored at −80 °C were thawed at 37 °C. Stocks were mixed at ⩾105 CCID50/serotype/mL to obtain test bivalent measles HPV vaccines and injected within 1 h.
2.3.2. Pichia expressing HPV
The purified recombinant HPV16L1 and 18L1 proteins were prepared in aluminum phosphate adjuvant as 0.225 mg/mL in a final PBS aqueous formulation. The final concentration of antigens was ∼20 μg/mL for HPV18L1 and ∼40 μg/mL for HPV16L1. The vaccine was adsorbed overnight, filled at a quantity of 1 mL/vial, and stored at 2–8 °C.
2.4. Immunization schedule, sampling scheme, and groups
Before the experiment, all non-human primates were subjected to thorough physical and veterinary examination and clinical biochemistry parameters were examined. Healthy rhesus monkeys were immunized with 1 mL vaccine formulation per animal, equivalent to one human dose (Table 1 ).
Table 1.
Experimental study design. 4 animals per test group and 3 animals with PBS were primed on day (D) 0, then boosted on D21, D180, and D342 except Pichia expressed HPV which was boosted until D180, either by intramuscular (I/M) or sub-cutaneous (S/C) administration as shown below.
| Group no. | Test groups of non-human primates | No. of animals | Immunization schedule (route) & scheme |
|---|---|---|---|
| G1 | Pichia expressed HPV | 4 | D0 (I/M),D21 (I/M),D180(I/M) |
| G2 | Measles vectored HPV (rMV-HPV) | 4 | D0 (S/C),D21 (S/C),D180(S/C),D342 (S/C) |
| G3 | Priming with Pichia and boosters with rMV-HPV | 4 | D0 (I/M) – Pichia Expressed HPV |
 | |||
| G4 | Measles vaccine primed | 4 |  |
| D342 (S/C)- rMV-HPV | |||
| G5 | PBS (negative control) | 3 | D0 (I/M), D21 (I/M),D180(I/M),D342 (I/M) |
Moreover, to assess the immune response elicited by the rMVEZHPV vaccine formulation in the case of pre-existing immunity against the MV, G4 was included, in which animals already immunized with 3 vaccine doses of attenuated measles vaccine were further boosted with 1 dose of rMVEZHPV. We injected only 2 boosters into Pichia expressing HPV, whereas 3 were given to the other groups (Table 1). Bleeding was done at day 0, 21, 33, 46, 126, 186, 193, 342, 372, 433, 492 and 586 for sera collection and subsequent analysis.
2.5. Evaluation of humoral immune response
2.5.1. Anti-HPV ELISA
The 96-well ELISA plates were precoated overnight with 50 ng/well of purified HPV16 or 18L1 proteins; the next day, the plates were washed to remove unbound proteins. The plates were blocked in 2% skim milk PBST for 1 h and washed with PBST. Sera samples were diluted at 1:80 for 16L1 and 1:40 for 18L1 in PBST, maintained for 1 h at 37 °C, and washed. Anti-simian IgG labeled with horseradish peroxidase was added and incubated for 1 h at 37 °C and washed. The TMB substrate was added and incubated for 30 min at 25 °C in the dark. The plates were read at 405 nm in a Spectramax Microplate reader (Molecular Devices, USA). One known positive sera sample was used as a standard and diluted serially for 4 parameters logistic regression curve fitting. All tested samples titrated were evaluated to determine the absolute antibody titres for HPV16/18L1 by extrapolation from the curve. A 4-fold or higher than negative control (PBS) titre increase was defined as seroconversion.
2.5.2. HPV pseudovirus-based neutralization assay
To measure specific anti-HPV L1 neutralizing antibodies in vitro, a pseudovirus-based neutralization assay (PBNA) was conducted [30] using secreted alkaline phosphatase (SEAP) as a reporter. Monkey sera were assayed either as pooled and individual samples. All testing, reagents, and materials such as the 293T cell line and reporter gene plasmids (p16sheLL, p18sheLL, pYSEAP), as well as the methods for the pseudovirus (PsV) production [31] and neutralization, were obtained from John Schiller (Laboratory of Cellular Oncology, NCI, NIH, Bethesda, MD, USA). PsV16SEAP and PsV18SEAP were produced by Lipofectamine 2000 transfection of 293TT cells, harvested, and purified by Optiprep gradient ultracentrifugation (Sigma, USA). SEAP was detected in the 72-h cell culture supernatant, using the Great ESCAPE SEAP Chemiluminescence Detection kit (Clontech, USA) according to the manufacturer instructions, using a Victor3V Multilabel Plate Reader (Perkin Elmer, USA). The neutralization titre was defined as the reciprocal of the highest dilution of serum that reduced SEAP activity by at least 50% compared to the reactivity in the control wells that received PsV but no antibody. Sera with at least 50% neutralizing activity were scored as neutralizing, and the data were statistically analyzed using GraphPad Prism 6.0 software to determine the IC50 (half maximal inhibitory concentration) values as described previously [30].
2.6. Anti-measles antibody neutralization assay
Sera samples twofold diluted in MEM - 2% FBS were analyzed in a 96-well microtitre plate. Live attenuated MVEZ virus equivalent to 100 CCID50 per 50 μL per well was added, except to the cell control wells. Vero cells were seeded at 20,000 ± 5000 cells per well per 100 μL in cell growth media (MEM - 10% FBS) and incubated in a CO2 incubator for 7–9 days. Cells were then observed for measles-specific cytopathic effects (CPE) under an inverted microscope and recorded. The reciprocal of the last dilution of the neutralized sera sample as determined by the absence of CPE was considered as the end-point titre.
2.7. Statistical analysis
Statistical significance of the difference between data sets was determined using a two-tailed Student t-test and one-way non-parametric analysis of variance using GraphPad Prism 6.0 software for Windows.
2.8. Ethical statement
Animal experiments were performed in accordance with the national animal ethics guidelines of the Government of India after approval by the Institutional Animal Ethics Committees of Cadila Healthcare, Ltd. (Ahmedabad, India).
3. Results
3.1. Cloning, rescue, and characterization of measles vectored HPV L1 vaccine candidate viruses
The selected recombinant plasmids, respectively named p(+)MV2EZ-HPV16L1 [25] and p(+)MV2EZ-HPV18L1 (Fig. 2 ), verified by analytical digestion and primer walking sequencing (MWG Biotech, Germany), were used to transfect 293T cells to rescue the corresponding attenuated rMVEZHPV16L1 and rMVEZHPV18L1 viruses (supplementary Fig. 1). The rescued rMVEZHPV16L1 and 18L1 viruses, amplified using MRC-5 cells, grew as efficiently as the parental virus, reaching a titre of 106 or higher. Moreover, rMVEZHPVs expressed high and stable levels of the HPVL1 antigen(s) during subsequent passages. Fig. 3 shows western blot characterization of the selected rMVEZHPV18L1 virus.
Fig. 2.
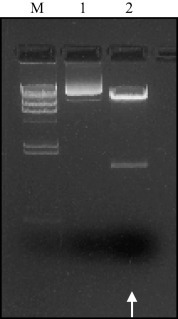
Positive p(+)MVEZHPV18L1 clone selection by restriction analysis. M: size marker, Lambda/HindIII; 1: p(+)MVEZHPV18L1; 2: p(+)MVEZHPV18L1 BssHII-AatII digested. White arrow: HPV18L1 transgene.
Fig. 3.
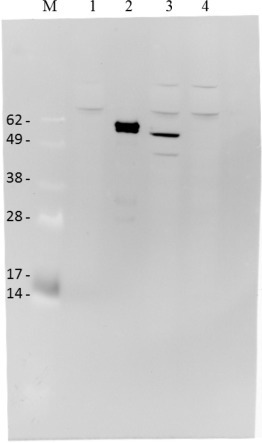
rMVEZHPV18L1 clone characterisation by western blot analysis. M: size marker, SeeBlue Plus2 standard; 1. Negative control, MVEZ; 2. Positive control, PsV18; 3. rMVEZHPV18L1 (∼58 kDa); 4. Negative control, rMVEZ-malaria. Used 1:4000 diluted specific anti-18L1 monoclonal antibody, Abcam.
3.2. Cloning, expression and selection of recombinant P. pastoris clones expressing HPV L1 vaccine candidates
The best-expressing HPV16L1 and 18L1 clones were selected based on SDS–PAGE and western blotting profiles. We confirmed them also by electron microscopy for VLP formation and they ranged in size from 30 to 80 nm and were irregular in shape (supplementary Fig. 2A and B).
3.3. Vaccination of non-human primates with vaccine candidates
The primates were vaccinated with both HPV vaccines as alone and different combinations in various test groups and PBS in the negative control (Table 1). The measles containing vaccines were immunized by the subcutaneous route (S/C) and Pichia expressing protein vaccines were immunized intramuscularly (I/M).
3.4. Humoral immune response as induced by rMVEZHPVL1 vaccines in susceptible primates and comparison with P. pastoris expressing HPV vaccine
3.4.1. Total IgG response measured by anti-HPV16L1/18L1 antibody ELISA
The HPV16L1 and HPV18L1 antibody titres (Fig. 4 A and B) were 4-fold or higher than the PBS negative control titres as early as D21 only after 1 dose in all vaccine groups. These antibody titres were slightly increased by the 2nd and 3rd boosters in both groups. At various intervals between immunization and until the end of experimentation, anti HPV16L1/18L1 titres were always ⩾4-fold higher than those in the negative control. Interestingly, when one dose of MV vectored HPV vaccine was given in measles vaccine-primed animals on D342, the anti-HPV 16 and 18L1 antibodies appeared quickly on D372 and later. This result confirms that pre-existing immunity to measles does not interfere with replication of the MV vectored HPV vaccine. Additionally, priming with Pichia and boosting with measles HPV resulted in a more than 4-fold increase in titres, demonstrating that classical and next-generation measles HPV can be combined for immunization.
Fig. 4.
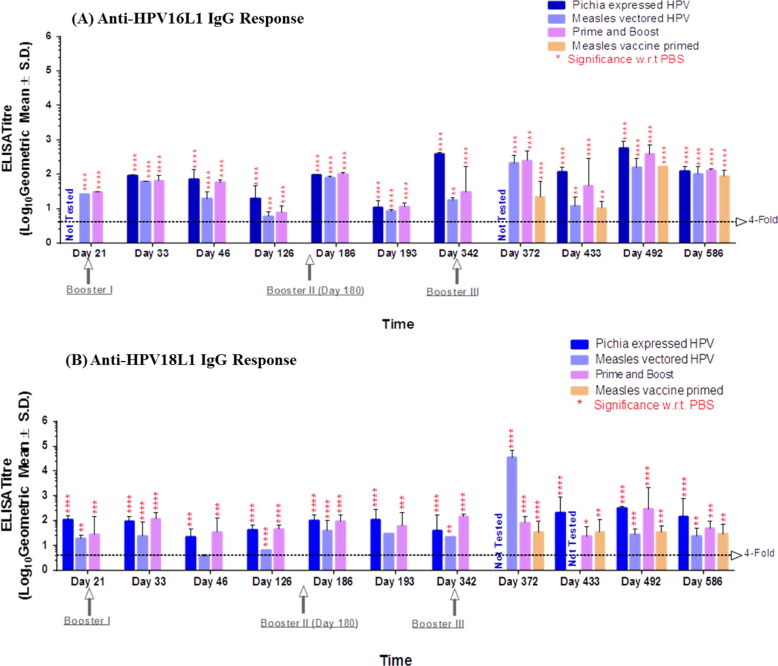
Anti-HPV16L1 and anti-HPV18L1 antibody response. (A). Anti-HPV16L1 total IgG titres measured by ELISA; (B). Anti-HPV18L1 total IgG titres measured by ELISA. These figures represents anti-HPV 16 L1 and anti-HPV18 L1 IgG antibody titres elicited in monkey groups immunized with our test vaccines at different time intervals. Statistical significance of the difference in titre between groups was determined by one-way non-parametric analysis of variance. P < 0.001 was considered statistically significant. Significance was calculated with respect to PBS negative control group.
Notably, our next-generation measles vectored HPV vaccine showed antibody titres that were comparable at different intervals until the end of the study and were similar to those of the classical Pichia expressed HPV vaccine (Fig. 4A and B).
3.4.2. Neutralizing antibodies against HPV L1 measured by PBNA
We demonstrated that the measles vectored and classical Pichia expressing vaccines induced remarkable levels of HPV-specific neutralizing antibodies for both HPV16L1 and 18L1 (Fig. 5 A and B). Anti-HPV16L1 antibody titres appeared as early as D21 after 1 dose in the vaccinated groups, whereas anti-HPV18L1 titres were detected on D33. The titres were boosted by the 2nd and 3rd doses and were maintained at high levels until the end of the experiment. Antibody titres against HPV18L1 were consistently relatively lower than those against HPV16L1.
Fig. 5.
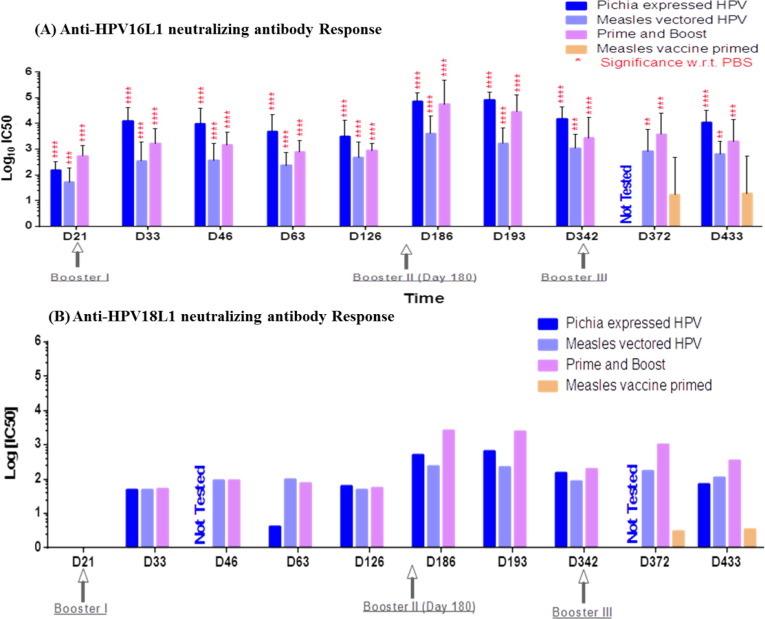
Anti-HPV16L1 and Anti-HPV18L1 neutralizing antibody determination. (A). Anti-HPV16L1 neutralizing antibodies measured by 16PBNA; (B). Anti-HPV18L1 neutralizing antibodies measured by 18PBNA. These figures represents anti-HPV 16 L1 and anti-HPV18 L1 neutralizing antibody titres elicited in monkey groups immunized with our test vaccines at different time intervals. Statistical significance of the difference in titre between groups was determined by one-way non-parametric analysis of variance. P < 0.001 was considered statistically significant. Significance was calculated with respect to PBS negative control group, Note- sera were tested as pool for anti HPV18 L1 neutralization therefore no statistical significance calculated.
3.5. Anti-measles neutralizing antibody response of measles vectored HPV vaccine
The anti-measles neutralizing antibody response induced by the measles vectored vaccine was comparable to that of commercial measles vaccine immunization (Fig. 6 ). Thus, the titre induced against measles was not affected by incorporation of the HPV transgene. This demonstrates that the measles vector can be used for both measles immunization and vectored antigens.
Fig. 6.
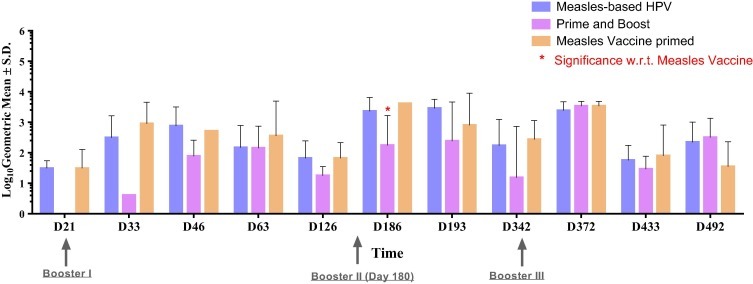
Anti-measles neutralizing antibody response. Figure represents anti-measles neutralizing antibody titres elicited in monkey groups immunized with our test vaccines at different time intervals. Statistical significance of the difference in titre between groups was determined by one-way non-parametric analysis of variance. P < 0.001 was considered statistically significant. Significance was calculated with respect to measles alone group.
4. Discussion
The main goal of our study was to establish and evaluate a next-generation measles vectored rMVEZ-HPV vaccine.
The rMVEZHPV vaccine was found to be comparable to the classical Pichia expressed HPV vaccine as shown by immune profiling in terms of generating HPV 16L1/18L1-specific total IgG, neutralizing antibodies, and related cellular immune responses in non-human primates.
The measles vectored HPV vaccine showed titre levels 4-fold or higher compared to those in the negative control for anti-HPV16L1/18L1 IgG, and neutralizing antibodies levels too comparable to the classical Pichia expressed HPV vaccine group.
The cost effectiveness of the rMVEZ-HPV vaccine is better than the vaccines produced by the market leaders Gardasil and Cervarix. Vaccination costs can vary by country; in developed countries, Gardasil costs approximately $120 per dose, and Cervarix, $100 [32]. Commercially, HPV vaccines are offered at $4.5. In developed countries, measles vaccines cost approximately $10 and are offered to UNICEF at a price of $0.24. Both commercial HPV vaccines require a complete three-dose vaccination regimen to confer protective immunity. Therefore, these vaccines cannot fulfill the needs of HPV vaccination in developing countries or low-resource areas. The process of vaccine production is complex and involves numerous steps (seed and batch fermentation, lysis, clarification, chromatography, ultracentrifugation and/or ultrafiltration, sterile filtration, adjuvant adsorption, and filling) when using yeast or other classical recombinant expression systems such as baculovirus. In contrast, live measles vaccines can be produced quickly and simply using MRC-5 cells for virus propagation in roller bottles followed by harvesting of the supernatant, clarification, blending, final filling, and lyophilization. The yield of live virus measles HPV was quite high at approximately 400,000 doses or more per batch of 100 roller bottles. The yield of yeast-based HPV vaccine production is lower at approximately 50,000 doses from a 20 L batch.
Therefore, the cost of producing our live attenuated measles vectored HPV vaccine is much lower than that of yeast or recombinant protein vaccines.
Moreover, the anti-HPV18L1 titres were always lower than anti-HPV16L1 in this study. However, this is common for HPV vaccines [33], [34]. Thus, the titres obtained by our vaccine also support the strong-weak antigen paradigm of HPVL1 antigens.
To generate a balanced and desirable level of immune response, we tested two different antigen delivery systems either alone or in combination using a heterologous prime and boost approach. We found comparable results in terms of total IgG and neutralization antibody titres in the different vaccine groups. Therefore, these vaccines can be combined for use in the field with already commercially available VLP vaccines. Furthermore, to verify the effect of pre-existing immunity against measles on the anti-HPV immune response by the recombinant vaccine candidate, we administered boosters of the MV-HPV vaccine in MV-primed animals. We observed significant increases in titres post-boost with HPV16/18L1. Thus, individuals with preimmune titres of measles can also be vaccinated for HPV.
There is also an extensive safety record of live viral vaccines including measles which proves that nonsegmented negative strand RNA virus (NNSV) genetic exchange is not of safety concern. There could be a theoretical risk posed by NNSV genetic exchange, which already exists in nature and was not created by the use of live vaccines. Indeed, the vaccine strains have restricted infectivity and replication capacity to participate in the dual infections with wild virus necessary for genetic exchange [35].
5. Conclusion
The data obtained from our immunogenicity studies are encouraging. Our findings provide a foundation for the development of a novel, cost-effective, prophylactic HPV vaccine. The rMVEZ-HPV vaccine induced high titres of HPV16 and HPV18 neutralizing antibodies in non-human primates, with no reduction in either neutralizing antibody titres for measles or production yields compared to the parent strain. Clinical trials of this approach are now planned.
Conflicts of interest
The authors have no actual or potential conflict of interest including any financial, personal, or other relationships with or amongst the organization for the submitted work.
Submission declaration
The work submitted has not been published previously (except in the form of an abstract or as part of a published lecture or academic thesis or as an electronic preprint) and it is not under consideration for publication elsewhere. This publication is approved by all authors.
Role of funding agency
The work to develop the HPV new candidate vaccine was provided by Cadila Healthcare, Ltd., Ahmedabad, India and Etna Biotech, Ltd., Catania, Italy.
Acknowledgements
We acknowledge John Schiller, Laboratory of Cellular Oncology, NCI, NIH, for supporting us throughout the project. Our sincere thanks to the entire Zydus Team. Special Thanks to the team members of VTC-Virology.
Footnotes
Supplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/j.vaccine.2016.07.051.
Appendix A. Supplementary material
References
- 1.De Flora S., La Maestra S. Epidemiology of cancers of infectious origin and prevention strategies. J Prev Med Hyg. 2015;56(1):15–20. [PMC free article] [PubMed] [Google Scholar]
- 2.De Sanjose S., Quint W.G., Alemany L., Geraets D.T., Klaustermeier J.E., Lloveras B. Human papillomavirus genotype attribution in invasive cervical cancer: a retrospective cross-sectional worldwide study. Lancet Oncol. 2010;11(11):1048–1056. doi: 10.1016/S1470-2045(10)70230-8. [DOI] [PubMed] [Google Scholar]
- 3.Santos-López G., Márquez-Domínguez L., Reyes-Leyva J., Vallejo-Ruiz V. General aspects of structure, classification and replication of human papillomavirus. Rev Med Inst Mex Seguro. 2015;53(2):166–171. [PubMed] [Google Scholar]
- 4.Kirnbauer R., Booy F., Cheng N., Lowy D.R., Schiller J.T. Papillomavirus L1 major capsid protein self-assembles into virus-like particles that are highly immunogenic. Proc Natl Acad Sci USA. 1992;89(24):12180–12184. doi: 10.1073/pnas.89.24.12180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Roden R.B., Greenstone H.L., Kirnbauer R., Booy F.P., Jessie J., Lowy D.R. In vitro generation and type-specific neutralization of a human papillomavirus type 16 virion pseudotype. J Virol. 1996;70(9):5875–5883. doi: 10.1128/jvi.70.9.5875-5883.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Einstein M.H., Takacs P., Chatterjee A., Sperling R.S., Chakhtoura N., Blatter M.M. Comparison of long-term immunogenicity and safety of human papillomavirus (HPV)-16/18 AS04-adjuvanted vaccine and HPV-6/11/16/18 vaccine in healthy women aged 18–45 years: end-of-study analysis of a Phase III randomized trial. Hum Vaccin Immunother. 2014;10(12):3435–3445. doi: 10.4161/hv.36121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Printz C. FDA approves Gardasil 9 for more types of HPV. Cancer. 2015;121(8):1156–1157. doi: 10.1002/cncr.29374. [DOI] [PubMed] [Google Scholar]
- 8.Vesikari T., Brodszki N., van Damme P., Diez-Domingo J., Icardi G., Petersen L.K. A randomized, double-blind, Phase III study of the immunogenicity and safety of a 9-valent human papillomavirus L1 virus-like particle vaccine (V503) versus Gardasil® in 9–15-year-old girls. Pediatr Infect Dis J. 2015;34(9):992–998. doi: 10.1097/INF.0000000000000773. [DOI] [PubMed] [Google Scholar]
- 9.Schiller J.T., Müller M. Next generation prophylactic human papillomavirus vaccines. Lancet Oncol. 2015;16:217–225. doi: 10.1016/S1470-2045(14)71179-9. [DOI] [PubMed] [Google Scholar]
- 10.Griffin D. Measles virus. In: Knipe D., Howley P., editors. 4th ed. vol. 2. Lippincott Raven Publishers; Philadelphia: 2001. pp. 1401–1441. (Field’s virology). [Google Scholar]
- 11.Beaty S.M., Lee B. Constraints on the genetic and antigenic variability of measles virus. Viruses. 2016;8(4):109–129. doi: 10.3390/v8040109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Radecke F., Spielhofer P., Schneider H., Kaelin K., Huber M., Dötsch C. Rescue of measles viruses from cloned DNA. EMBO J. 1995;14:5773–5784. doi: 10.1002/j.1460-2075.1995.tb00266.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Billeter M.A., Naim H.Y., Udem S.A. Reverse genetics of measles virus and resulting multivalent recombinant vaccines: applications of recombinant measles viruses. Curr Top Microbiol Immunol. 2009;329:129–162. doi: 10.1007/978-3-540-70523-9_7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Singh M., Cattaneo R., Billeter M.A. A recombinant measles virus expressing hepatitis B virus surface antigen induces humoral immune responses in genetically modified mice. J Virol. 1999;73(6):4823–4828. doi: 10.1128/jvi.73.6.4823-4828.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Tangy F., Naim H.Y. Live attenuated measles vaccine as a potential multivalent pediatric vaccination vector. Viral Immunol. 2005;18(2):317–326. doi: 10.1089/vim.2005.18.317. [DOI] [PubMed] [Google Scholar]
- 16.Singh M., Billeter M.A. A recombinant measles virus expressing biologically active human interleukin-12. J Gen Virol. 1999;80:101–106. doi: 10.1099/0022-1317-80-1-101. [DOI] [PubMed] [Google Scholar]
- 17.Liniger M., Zuniga A., Morin T.N., Combardiere B., Marty R., Wiegand M. Recombinant measles viruses expressing single or multiple antigens of human immunodeficiency virus (HIV-1) induce cellular and humoral immune responses. Vaccine. 2009;27(26):3299–3305. doi: 10.1016/j.vaccine.2009.01.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Stebbings R., Li B., Lorin C., Koutsoukos M., Février M., Mee E.T. Immunogenicity of a recombinant measles HIV-1 subtype C vaccine. Vaccine. 2013;31(51):6079–6086. doi: 10.1016/j.vaccine.2013.09.072. [DOI] [PubMed] [Google Scholar]
- 19.Brandler S., Tangy F. Vaccines in development against West Nile virus. Viruses. 2013;5(10):2384–2409. doi: 10.3390/v5102384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Escriou N., Callendret B., Lorin V., Combredet C., Marianneau P., Février M. Protection from SARS coronavirus conferred by live measles vaccine expressing the spike glycoprotein. Virology. 2014;453:32–41. doi: 10.1016/j.virol.2014.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ramsauer K., Schwameis M., Firbas C., Müllner M., Putnak R.J., Thomas S.J. Immunogenicity, safety, and tolerability of a recombinant measles-virus-based chikungunya vaccine: a randomised, double-blind, placebo-controlled, active-comparator, first-in-man trial. Lancet Infect Dis. 2015;15(5):519–527. doi: 10.1016/S1473-3099(15)70043-5. [DOI] [PubMed] [Google Scholar]
- 22.Brandler S., Ruffié C., Combredet C., Brault J.B., Najburg V., Prevost M.C. A recombinant measles vaccine expressing chikungunya virus-like particles is strongly immunogenic and protects mice from lethal challenge with chikungunya virus. Vaccine. 2013;31(36):3718–3725. doi: 10.1016/j.vaccine.2013.05.086. [DOI] [PubMed] [Google Scholar]
- 23.Lorin C1, Mollet L., Delebecque F., Combredet C., Hurtrel B. A single injection of recombinant measles virus vaccines expressing human immunodeficiency virus (HIV) type 1 clade B envelope glycoproteins induces neutralizing antibodies and cellular immune responses to HIV. J Virol. 2004;78(1):146–157. doi: 10.1128/JVI.78.1.146-157.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Brandler S1, Lucas-Hourani M., Moris A., Frenkiel M.P., Combredet C., Février M. Pediatric measles vaccine expressing a dengue antigen induces durable serotype-specific neutralizing antibodies to dengue virus. PLoS Negl Trop Dis. 2007;1(3):e96. doi: 10.1371/journal.pntd.0000096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cantarella G1, Liniger M., Zuniga A., Schiller J.T., Billeter M., Glueck R. Recombinant measles virus-HPV vaccine candidates for prevention of cervical carcinoma. Vaccine. 2009;27(26):3385–3390. doi: 10.1016/j.vaccine.2009.01.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mrkic B1, Pavlovic J., Rülicke T., Volpe P., Buchholz C.J., Hourcade D. Measles virus spread and pathogenesis in genetically modified mice. J Virol. 1998;72:7420–7427. doi: 10.1128/jvi.72.9.7420-7427.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bazan SB1, de Alencar Muniz Chaves A., Aires K.A., Cianciarullo A.M., Garcea R.L., Ho P.L. Expression and characterization of HPV-16 L1 capsid protein in Pichia pastoris. Arch Virol. 2009;154(10):1–16. doi: 10.1007/s00705-009-0484-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sambrook J.F., Russell D.W. Molecular Cloning: A Laboratory Manual. 3rd ed. vol. 1. Cold Spring Harbor Laboratory Press; 2001. Directional cloning. 17(1.84) [Google Scholar]
- 29.Wulff N.H., Tzatzaris M., Young P.J. Monte Carlo simulation of the Spearman-Kaerber TCID50. J Clin Bioinformat. 2012;2(5):1–5. doi: 10.1186/2043-9113-2-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pastrana D.V., Buck C.B., Pang Y.Y., Thompson C.D., Castle P.E., FitzGerald P.C. Reactivity of human sera in a sensitive, high-throughput pseudovirus-based papillomavirus neutralization assay for HPV16 and HPV18. Virology. 2004;321(2):205–216. doi: 10.1016/j.virol.2003.12.027. [DOI] [PubMed] [Google Scholar]
- 31.Buck C.B., Thompson C.D., Pang Y.Y., Lowy D.R., Schiller J.T. Maturation of papillomavirus capsids. J Virol. 2005;79(5):2839–2846. doi: 10.1128/JVI.79.5.2839-2846.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Monie A., Hung C.F., Roden R., Wu T.C. Cervarix™: a vaccine for the prevention of HPV 16, 18-associated cervical cancer. Biologics. 2008;2(1):107–113. [PMC free article] [PubMed] [Google Scholar]
- 33.Day P.M., Pang Y.Y., Kines R.C., Thompson C.D., Lowy D.R., Schiller J.T. A human papillomavirus (HPV) in vitro neutralization assay that recapitulates the in vitro process of infection provides a sensitive measure of HPV L2 infection-inhibiting antibodies. Clin Vaccine Immunol. 2012;19(7):1075–1082. doi: 10.1128/CVI.00139-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kemp T.J., García-Piñeres A., Falk R.T., Poncelet S., Dessy F., Giannini S.L. Evaluation of systemic and mucosal anti-HPV16 and anti-HPV18 antibody responses from vaccinated women. Vaccine. 2008;26(30):3608–3616. doi: 10.1016/j.vaccine.2008.04.074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Peter L.C., Alexander B., Brian R.M. What are the risks—hypothetical and observed—of recombination involving live vaccines and vaccine vectors based on nonsegmented negative-strain RNA viruses? J Virol. 2008;82(19):9805–9806. doi: 10.1128/JVI.01336-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.