

Abstract
Evidence is mounting that microorganisms originating from livestock impact the air quality of the animal houses themselves and the public in the surrounding neighborhoods. The aim of this study was to develop efficient bacterial source tracking capabilities to identify sources of Escherichia coli aerosol pollution caused by pigs. Airborne E. coli were isolated from indoor air, upwind air (10 and 50 m away) and downwind air samples (10, 50, 100, 200 and 400 m away) for five swine houses using six-stage Andersen microbial samplers and Reuter-Centrifugal samplers (RCS). E. coli strains from pig fecal samples were also collected simultaneously. The enterobacterial repetitive intergenic consensus polymerize chain reaction (ERIC-PCR) and the repetitive extragenic palindromic (REP-PCR) approaches were used to study the genetic variability and to determine the strain relationships among E. coli isolated from different sites in each swine house. Results showed that 35.1% (20/57) of the bacterial DNA fingerprints from the fecal isolates matched with the corresponding strains isolated from indoor and downwind air samples (similarity ⩾90%). E. coli strains from the indoor and downwind air samples were closely related to the E. coli strains isolated from feces, while those isolated from upwind air samples (swine house C) had low similarity (61–69%). Our results suggest that some strains isolated from downwind and indoor air originated in the swine feces. Effective hygienic measures should be taken in animal farms to prevent or minimize the downwind spread of microorganism aerosol.
Keywords: Source identification, Swine house, Airborne Escherichia coli, ERIC-PCR, REP-PCR
1. Introduction
Bioaerosols contamination caused by animals maybe a threat to both animals and public health. In particular, microorganisms or their fragments can be passively or actively released into the air in response to the changes of the hygienic situation of animal houses and meteorological conditions (Jones and Harrison, 2004). Airborne microorganisms found in swine confinement units may have a direct impact on the wellbeing of the animals in different stages of growth as well as the health of human caretakers. Many airborne pathogenic microorganisms, including viruses and bacteria, can be transmitted great distances through air flow (Brown and Hovmoller, 2002; Donaldson, 1999; Lee et al., 2007; Mims and Mims, 2004; Shinn et al., 2000). Examples include Foot-and-Mouth disease virus (Donaldson et al., 1982); severe acute respiratory syndrome virus (Eubank et al., 2004; Weiss and McMichael, 2004); influenza virus (Klontz et al., 1989; Landman and Schrier, 2004); Bacillus anthracis (CDC, 2003); and Klebsiella pneumonia (Prazmo et al., 2003). These facts illustrate the need for adequate source identification of the origins and transmission of microbial pathogens. An understanding of the origin of air pollution is paramount for assessing associated health risks as well as the actions necessary to remedy the problem.
Airborne bacterial source tracking has been studied mostly by concentration differences of the total number of aerobic bacteria (Chai et al., 1999, Chai et al., 2001; Kim and Kim, 2007), and the differences of antibiotic resistances of bacteria (Chai et al., 2003; Yao et al., 2007). However, tracing the origin of airborne microorganisms is difficult using culture-based approaches. Additionally, using traditional bacterial taxonomy proves problematic when differentiating between strains that share a close genetic relationship. With the development of molecular biology, genotypic techniques were developed for studying bacterial source tracking in the field. This was initially achieved in water pollution identification (Ahmed et al., 2005; Griffith et al., 2003; Indest et al., 2005; Meays et al., 2004; van Belkum et al., 1996). Molecular methods are able to distinguish between closely related bacteria by producing DNA fingerprints that are specific for individual strains. By the comparison of these DNA fingerprints, strains from one site can be related to strains from another site. Tracing the migration of an indicator bacterium of air pollution based on comparison of their DNA fingerprints provides invaluable information. However, this technique has not yet been applied to study the origin and transmission of bioaerosols in animal houses.
Several microorganisms have been used as indicators of fecal contamination. Among indicator bacteria, E. coli and Enterococcus spp. are most commonly used as the target organisms for bacterial source tracking in seawater (Bitton et al., 1983; Hanes and Fragala, 1967; Keswick et al., 1982; McFeters et al., 1974; Omura et al., 1982; Wait and Sobsey, 2001). E. coli are a dominant intestinal bacterial population in swine, colonizing the intestine distal to the stomach (Schierack et al., 2006). It is estimated that 1–4% of all cultivable bacteria of the colon are E. coli, and up to 1010 E. coli bacteria can be detected in 1 g of feces (Scharek et al., 2005; Selander et al., 1987). Nonpathogenic E. coli (commensal E. coli) strains are thought to maintain the physiological milieu of the gut and support digestion as well as defend against enteric pathogens. Other E. coli strains carrying virulence genes (pathogenic E. coli), can cause severe outbreaks such as neonatal and postweaning diarrhea and edema of swine (Gyles and Fairbrother, 2004; Imberechts et al., 1992). These strains are responsible for major economic losses in pig rearing (Wieler et al., 2001). In addition, E. coli is a common microorganism in the air environment, particularly in animal houses and their surroundings (Duan et al., 2006; Northcutt et al., 2004; Zucker et al., 2000). Hojovec et al. (1977) evaluated the air quality in poultry houses using E. coli as an indicator. In this experiment, E. coli was used as indicator bacterium to study the origin of bioaerosol and its transmission to the ambient air of swine houses.
In this study, the genetic similarity of E. coli strains isolated from air samples of different sites and fecal samples in five different swine houses was characterized using the ERIC-PCR and REP-PCR. From this, a deeper understanding of the mechanisms of the occurrence and spread of airborne E. coli can be obtained.
2. Materials and methods
2.1. Animal houses studied
Five swine houses were studied in Shandong, China, between January and October 2006. These houses were located outside of neighboring villages where there were no buildings or tall vegetation around the swine houses. Swine house A had a distance of more than 2000 m to the nearest village, where there were no crops planted at that time. Swine house B had a distance of more than 3000 m to the nearest village, where there were planting wheat at that time. Swine C, D and E were located to the east of the villages with distances ranging from 500 to 1000 m from the nearest village and had the same geographical characteristics. The meteorological conditions were recorded at the time of air sampling and a description of these conditions is given in Table 1 .
Table 1.
Description of swine houses studied.
| Swine house | N | Layout | Ventilation system | Age (D) | Inside |
Outside |
||||
|---|---|---|---|---|---|---|---|---|---|---|
| T (°C) | RH (%) | WS (m/s) | T (°C) | RH (%) | WS (m/s) | |||||
| A | 450 | Close | Mechanical | 90–106 | 17 | 81 | 0 | 14 | 78 | 0–0.6 |
| B | 500 | Half-close | Natural | 45–190 | 22 | 65 | 0 | 18 | 60 | 1.4–5.0 |
| C | 600 | Half-close | Natural | 70–95 | 28 | 67 | 0 | 35 | 58 | 0–1.5 |
| D | 360 | Half-close | Natural | 48–85 | 20 | 67 | 0 | 22 | 70 | 0–1.5 |
| E | 480 | Half-close | Natural | 45–80 | 22 | 65 | 0 | 24 | 70 | 0–1.5 |
Note: N=Number of animal; D=day; T=Temperature; RH=Relative Humidity; WS=Wind Speed.
2.2. Airborne Escherichia coli
Six-stage Andersen samplers (Andersen, 1958) were used to sample the indoor air for the collection of E. coli in the swine houses. Samplers were placed near the middle of the stable about 1 m above the ground. The Reuter-Centrifugal samplers (RCS) were used to collect airborne E. coli from the outdoor air at the different sites upwind (10 and 50 m away from the swine houses) and downwind (10, 50, 100, 200 and 400 m away from the swine houses). The airflow rates for the Andersen sampler and the RCS were 28.3 and 40 L min−1, respectively. The Andersen sampler was equipped with MacConkey agar No. 3 (Oxoid) plates and operated for 1–8 min and the RCS was equipped with MacConkey agar strips and also operated for 1–8 min. At each sampling site, five air samples were collected continuously. The exposed agar plates and strips were incubated at 37 °C for 48 h. All colonies that appeared on the plates and strips were screened based on their Gram reactions using the KOH assay. Gram-negative colonies were subcultured on MacConkey Agar and species identifications were made using conventional biochemical tests: the ability to ferment lactose and glucose (Kligler-Iron test), to produce hydrogen sulfide, and when in doubt, use of the API 20 E system (BioMérieux, Marcy-I’Etoile, France).
2.3. Escherichia coli in feces
Ten to 15 fresh fecal samples of healthy pigs were aseptically collected from each feedlot randomly from piles on the ground and air samples were collected simultaneously. Then, the fecal samples were immediately transported to the laboratory in ice-cooled containers and analyzed within 24 h of collection. Approximately 1 g of feces was transferred to a sterilized glass homogenizer containing 9 mL of 0.9% sodium chloride. Samples were diluted and spread onto the surface of eosin methylene blue (EMB) agar plates (Tianhe, Hangzhou, China) which were incubated at 37 °C for 18–20 h. Colonies with a metallic sheen on EMB agar were picked and streaked onto MacConkey agar No. 3 (Oxoid). After overnight incubation at 37 °C, one or two typical pink colonies were selected from each MacConkey agar plate and the isolate identifications were performed as described in Section 2.2.
2.4. DNA extraction
All E. coli isolates were grown in 5 mL of Luria-Bertani (LB) broth (Oxoid) with moderate shaking for 18 h at 37 °C. Then, 1.5 mL LB broth was removed and centrifuged at 10,000g for 2 min. The cell pellet was resuspended in 100 μL of sterile double distilled water and held in boiling water for 10 min. Bacteria were then rapidly cooled by placing on ice for 5 min. This mixture was centrifuged at 12,000g for 2 min and the supernatant removed and stored at −20 °C. These samples were used as the DNA templates for PCR analysis (Cheng et al., 2006).
2.5. ERIC-PCR and REP-PCR
The PCR reaction mixture (25 μL) contained 1.5 U Taq DNA polymerase (TaKaRa), 300 ng μL−1 of each primer, 0.875 mM of each dNTP, 1× reaction buffer and 1.5 mM MgCl2 (TaKaRa). Amplification was performed in a thermocycler (Eppendorf, Germany) as follows: an initial denaturation (94 °C, 5 min) followed by 35 cycles of denaturation (94 °C, 1 min), annealing (1 min at 42 °C), and extension (65 °C, 8 min) with a single final extension (65 °C, 16 min). The reaction products were stored at 4 °C until they were electrophoresed on a 1.2–1.5% (w/v) agarose gel containing 1× TAE and 0.5 μg mL−1 ethidium bromide along with the DL2000 DNA markers (TaKaRa). The primers ERIC1, ERIC2, REP1R and REP2I were synthesized by TaKaRa (Dalian, China) and the sequences (5′–3′) are as follows (Versalovic et al., 1991, Versalovic et al., 1994):
ERIC 1: ATGTAAGCTCCTGGGGATTCAC; ERIC 2: AAGTAAGTGACTGGGGTGAGCG;
REP1R: IIIICGICGICATCIGGC; REP2I: ICGICTTATCIGGCCTAC.
2.6. ERIC-PCR and REP-PCR fingerprints analyses
ERIC and REP-PCR fingerprints of amplified DNA fragments obtained from the agarose gel electrophoreses were recorded. The observed bands in the gels were evaluated based on the presence (coded 1) or absence (coded 0) of polymorphic fragments for the ERIC primers. Cluster analysis was performed with NTSYS-pc (Version 2.10), a numerical taxonomy and multivariate analysis software package (Rohlf, 2000), based on Dice's similarity coefficient (S D) with a 1% position tolerance and the unweighted pair group method using arithmetic averages (UPGMA). In addition, each isolate was considered as an operational taxonomic unit (OTU). In order to reduce the number of OTUs in the dendrogram, to facilitate interpretation, isolates of ⩾90% similarity were treated as a single isolate (Borges et al., 2003).
3. Results
The number of isolates recovered and the number examined from each site (including fecal samples) at each swine house was presented in Table 2 . Table 2 showed that a total of 60 fecal samples were examined, and 57 E. coli strains were detected in this experiment. At the same time, 41 E. coli strains from indoor air and 19 E. coli strains from downwind air were detected. Only 3 E. coli strains from upwind air were detected at swine house C (Table 2).
Table 2.
Number of isolates recovered and the number examined from each site at each swine house.
| Site | Swine house |
||||
|---|---|---|---|---|---|
| A (R/E) | B (R/E) | C (R/E) | D (R/E) | E (R/E) | |
| Fecal | 10/10 | 10/10 | 14/15 | 12/15 | 11/10 |
| Upwind 50 m | 0/5 | 0/5 | 1/5 | 0/5 | 0/5 |
| Upwind 10 m | 0/5 | 0/5 | 2/5 | 0/5 | 0/5 |
| Indoor air | 5/5 | 8/5 | 12/5 | 9/5 | 7/5 |
| Downwind 10 m | 0/5 | 2/5 | 4/5 | 2/5 | 3/5 |
| Downwind 50 m | 0/5 | 2/5 | 3/5 | 0/5 | 1/5 |
| Downwind 100 m | 0/5 | 1/5 | 1/5 | 0/5 | 0/5 |
| Downwind 200 m | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 |
| Downwind 400 m | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 |
Note: R/E=The number of isolates recovered/the number examined.
The electrophoretic profiles of the ERIC-PCR products were determined for the E. coli strains isolated from indoor air, outdoor air and fecal samples. The data matrices based on the DNA fragments and the dendrogram using the NTSYS-pc software were constructed, grouping all E. coli isolates obtained in this study into different clusters or branches based on the ERIC-PCR pattern similarity. For best comparision and resolution, the ERIC DNA fragments from the isolates obtained from the same animal house were amplified in one PCR reaction and loaded on the same gel.
According to the UPGMA dendrogram (Fig. 1, Fig. 2, Fig. 3, Fig. 4, Fig. 5 ), each swine house contained fecal-obtained E. coli strains that shared 100% similarity with the corresponding air-obtained E. coli strains. Additional air sampled strains obtained either upwind, downwind, or indoor had the similarity ranged from 100% to 58% (swine house A); to 43% (swine house B); to 61% (swine house C); to 52% (swine house D); to 50% (swine house E) with the corresponding strains isolated from fecal samples. The number of E. coli strains isolated from indoor air which shared 100% similarity with fecal strains was different in 5 swine houses (Table 3 ).
Fig. 1.
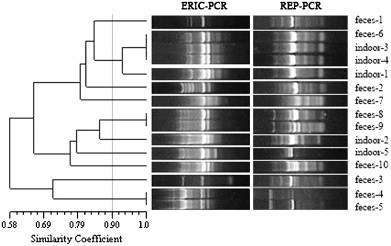
ERIC-PCR and REP-PCR fingerprints of 15 E. coli strains in swine house A. The dendrogram was constructed using UPGMA with a 1% band tolerance (feces-1 means the first E. coli strain isolated from feces; indoor-2 means the second E. coli strain isolated from indoor air; upwind50m-1 means the first E. coli strain isolated from upwind at 50 m; downwind100m-1 means the first E. coli strain isolated from downwind at 100 m).
Fig. 2.
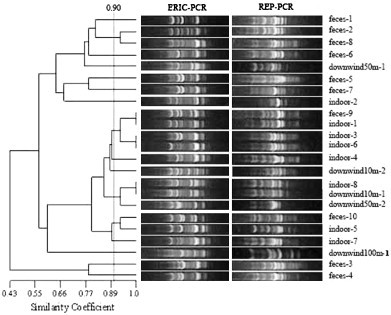
ERIC-PCR and REP-PCR fingerprints of 23 E. coli strains in swine house B. The dendrogram was constructed using UPGMA with a 1% band tolerance (feces-1 means the first E. coli strain isolated from feces; indoor-2 means the second E. coli strain isolated from indoor air; upwind50m-1 means the first E. coli strain isolated from upwind at 50 m; downwind100m-1 means the first E. coli strain isolated from downwind at 100 m).
Fig. 3.
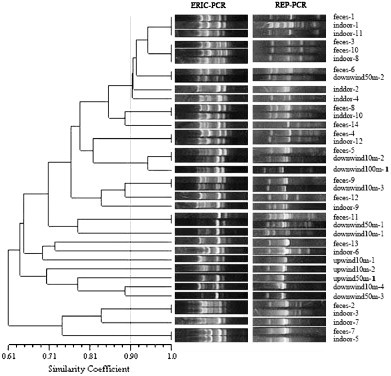
ERIC-PCR and REP-PCR fingerprints of 37 E. coli strains in swine house C. The dendrogram was constructed using UPGMA with a 1% band tolerance (feces-1 means the first E. coli strain isolated from feces; indoor-2 means the second E. coli strain isolated from indoor air; upwind50m-1 means the first E. coli strain isolated from upwind at 50 m; downwind100m-1 means the first E. coli strain isolated from downwind at 100 m).
Fig. 4.
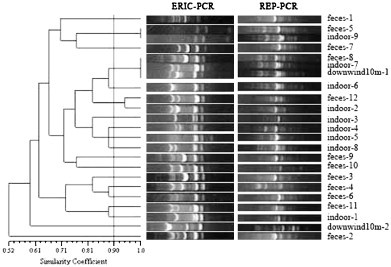
ERIC-PCR and REP-PCR fingerprints of 23 E. coli strains in swine house D. The dendrogram was constructed using UPGMA with a 1% band tolerance (feces-1 means the first E. coli strain isolated from feces; indoor-2 means the second E. coli strain isolated from indoor air; upwind50m-1 means the first E. coli strain isolated from upwind at 50 m; downwind100m-1 means the first E. coli strain isolated from downwind at 100 m).
Fig. 5.

ERIC-PCR and REP-PCR fingerprints of 22 E. coli strains in swine house E. The dendrogram was constructed using UPGMA with a 1% band tolerance (feces-1 means the first E. coli strain isolated from feces; indoor-2 means the second E. coli strain isolated from indoor air; upwind50m-1 means the first E. coli strain isolated from upwind at 50 m; downwind100m-1 means the first E. coli strain isolated from downwind at 100 m).
Table 3.
Similarity among E. coli strains isolated from indoor air and fecal samples or outdoor air samples based on the ERIC-PCR and REP-PCR methods in 5 swine houses.
| Stable | The number of strains |
||||||||
|---|---|---|---|---|---|---|---|---|---|
| Indoor air with fecal samples |
Indoor air or fecal samples with downwind samples |
Indoor air with upwind samples |
|||||||
| 100% | 90–100% | <90% | 100% | 90–100% | <90% | 100% | 90–100% | <90% | |
| A | 2/5 | 1/5 | 2/5 | – | – | – | – | – | – |
| B | 1/8 | 3/8 | 4/8 | 1/5 | 1/5 | 3/5 | – | – | – |
| C | 7/12 | 0/12 | 5/12 | 4/8 | 1/8 | 3/8 | 0 | 0 | 3/3 |
| D | 2/9 | 1/9 | 6/9 | 1/2 | 0 | 1/2 | – | – | – |
| E | 2/8 | 2/8 | 4/8 | 1/4 | 1/4 | 2/4 | – | – | – |
Note: “–”=E. coli strains were not isolated in these sites.
3.1. The similarity among E. coli strains isolated from indoor air and fecal samples
As shown in the UPGMA dendrograms, 33.3% (14/42) of E. coli strains (Table 3) isolated from indoor air samples shared 100% similarity with corresponding strains isolated from fecal samples in the five swine houses. In swine house A, strains indoor-3 and -4 shared 100% similarity with feces-6, while the same was observed for indoor-1 and feces-9 (swine house B); indoor-1, -11 and feces-1 (swine house C); indoor-8 and feces-3, -10 (swine house D); indoor-9 and feces-5 (swine house D); indoor-3 and feces-5 (swine house E); (Fig. 1, Fig. 2, Fig. 3, Fig. 4, Fig. 5). In addition, 23.8% (10/42) of E. coli strains isolated from indoor air samples had ⩾90% but <100% similarity with the fecal strains. In swine house A, indoor-1 and feces-6 shared 93% similarity; in swine house B, indoor-3, -6 and feces-9 shared 92% similarity as did indoor-5 and feces-10; in swine house C, over 90% similarity existed among indoor-2, -4 and feces -1, -3, -6, -10; in swine house D indoor-2 and feces-12 shared 93% similarity; in swine house E, indoor-4 and feces-5 had 94% similarity (Fig. 1, Fig. 2, Fig. 3, Fig. 4, Fig. 5).
Of the strains isolated from indoor air samples, 50.0% (21/42) had<90% similarity with strains isolated from fecal samples in the five swine houses. For instance, in swine house A indoor-5 and feces-8, -9 shared 79% similarity; in swine house B indoor-8 and feces-9 had 79% similarity. All isolates with<90% similarity were treated as independent strains.
3.2. The similarity among E. coli strains isolated from downwind air samples and indoor air or fecal samples
A total of 36.8% (7/19) E. coli strains (Table 3) isolated from downwind air samples at different sampling sites had 100% similarity with corresponding strains isolated from indoor air or fecal samples in the five swine houses. In swine house B downwind10m-1 and indoor-8 had 100% similarity as did downwind10m-2 and feces-5, downwind50m-2 and feces-6, downwind10m-3 and feces-9 (swine house C); downwind10m-1, feces-8 and indoor-7 (swine house D); downwind10m-3 and feces-6 (swine house E) (Fig. 2, Fig. 3, Fig. 4, Fig. 5). In addition, 15.8% (3/19) E. coli strains isolated from downwind air samples at different sampling sites had ⩾90% but <100% similarity with corresponding strains isolated from indoor air or fecal samples. For example, in swine house B, downwind50m-2 and indoor-8 shared 92% similarity; in swine house C downwind100m-1 and feces-5 shared 94% similarity; in swine house E downwind10m-1, indoor-3 and feces-5 had 92% similarity (Fig. 2, Fig. 3, Fig. 5). Isolates with ⩾90% similarity were treated as the same strain (Borges et al., 2003).
In addition, 47.4% (9/19) E. coli strains (Table 3) isolated from downwind air samples had <90% similarity with corresponding strains isolated from indoor air or fecal samples in the swine houses B through E. For example, in swine house B, downwind50m-1 and feces-1 had low similarity (77%); in swine house C, downwind10m-1 and feces-11 shared only 76% similarity, and downwind10m-4, downwind50m-3 and feces-1 shared only 64% similarity; in swine house D, downwind10m-2 and feces-1 had only 59% similarity; in swine house E, downwind10m-2 with feces-5 shared only 84% similarity. All the isolates with <90% similarity were treated as different isolates.
3.3. The similarity among E. coli strains isolated from upwind air samples and indoor air or fecal samples
Airborne E. coli strains were isolated from upwind air samples only for swine house C and all three of these strains (upwind10m-1, 2 and upwind50m-1) had low similarity (61–69%) with corresponding strains isolated from indoor air or fecal samples (Fig. 3, Table 3).
4. Discussion
An effective method for bacterial source tracking is one that can correctly identify the origin of environmental isolates; has the greatest accuracy, and is quick and cost effective. There are several techniques in use for this purpose as mentioned in the historical background. The use of PCR-based molecular techniques, which depend on DNA-sequence polymorphisms, has become wide spread. PCR reactions are easy to execute, relatively inexpensive, and highly reliable.
ERIC also known as intergenic repeat units is another family of prokaryotic repetitive sequences. In E. coli there are 30–50 copies of ERIC sequences, and ERIC units tend to occur singly, including a potential stem-loop structure, and are located in non-coding regions (Hulton et al., 1991; Martin et al., 1992; Sharples and Lloyd, 1990). REP sequences are present from 500 to 1000 base pairs in length and are found in clusters in which successive copies are found in alternate orientations (Gilson et al., 1984; Martin et al., 1992; Stern et al., 1984). And REP-PCR is a widely used method for DNA typing analysis and has been shown to successfully differentiate strains of Bartonella (Rodriguez-Barradas et al., 1995), Citrobacter diversus (Woods et al., 1992), Enterobacter aerogenes (Georghiou et al., 1995), Burkholderiaa cepacia (Hamill et al., 1995), Salmonella enterica (Johson and Calbots, 2000) and E. coli (Dombek et al., 2000). So, the ERIC-PCR and REP-PCR techniques have been devised for characterization of bacteria and are widely employed to distinguish species, strains and serotypes (Chansiripornchai et al., 2001; de Moura et al., 2001; Lupsiki and Weinstock, 1992; van Belkum, 1994; Versalovic et al., 1994). McLellan et al. (2003) found that ERIC-PCR and REP-PCR yielded comparable results using E. coli as the target organism. In this experiment, our results showed that the discriminatory power yielded by the techniques of ERIC-PCR and REP-PCR was very well to differentiate between strains isolated from different sites in each swine house.
In the present study, we analyzed 120 E. coli isolates originating from five swine houses. As illustrated in Fig. 1, Fig. 2, Fig. 3, Fig. 4, Fig. 5, the grouping of REP-PCR fingerprints was highly similar with those obtained from ERIC-PCR. This showed that the two approaches yield an analogous taxonomic resolution, suggesting that the two approaches can be fully applied for the study of the transmission of bioaerosols from animal houses to their environments. From the results, we can see that 48.8% E. coli strains isolated from indoor air samples had ⩾90% similarity with corresponding strains isolated from fecal samples in each swine house. That is, the fecal strains could be aerosolized and spread to the indoor air. In contrast, however, 51.2% of E. coli isolated from indoor air samples had a far genetic distance (<90% similarity) from the fecal samples. This observation might result from the following reasons: (i) the strains isolated from the indoor air originated from other sources (e.g. feed, water, etc.); (ii) these strains are present in the fecal samples, but were not isolated from the fecal samples and could not be compared.
Similarly, 52.6% (10/19) of strains isolated from downwind air samples had a ⩾90% similarity with strains isolated from indoor air and/or fecal samples, indicating that the strains in the indoor air could spread to the outdoor air, especially the downwind air, via air exchange. Some strains isolated from downwind (47.4%) however had a <90% similarity with strains corresponding from the indoor air or fecal samples in swine houses B–C, while no E. coli strains were isolated from downwind of house A. These results indicated that some strains isolated from downwind sites did not originate from the swine stables. It is obvious that outdoor air can be easily affected by anthropogenic activities (Lighthart, 1984) and environmental factors.
Airborne E. coli was not collected in upwind air samples from four of the swine houses (A, B, D and E) or⩾200 m downwind of all five swine houses. This was likely due to the following reasons: (i) the concentrations of airborne E. coli were very low in these sampling sites; (ii) environmental factors such as relative humidity, temperature, ultraviolet (UV) radiation, oxygen content, specific ions, various pollutants and air-associated factors influenced the ability of microorganisms to survive in the air (Hermann et al., 2007); (iii) the aerosolization of microorganisms and sampling stress lead to a loss of culture viability (Marthi et al., 1990; Stewart et al., 1995).
Based on these findings, it can be concluded that the use of ERIC-PCR and REP-PCR are beneficial, and these approaches have shown to be useful for assessing the genetic diversity of isolates from different geographic locations (Mehta et al., 2002). Moreover, the consumables and labor costs for ERIC-PCR and REP-PCR analyses are significantly lower than those for other genotyping methods (Olive and Bean, 1999).
Taken together, by using the accurate ERIC-PCR and REP-PCR technologies, E. coli in feces was found aerosolized and spread to outdoor air, especially to the downwind air of the swine houses via air exchange. Airborne microbes released from swine farms may pose health risks to exposed animals and human beings, especially farm workers (Donham et al., 1986). However, further studies are needed to determine if these microbial contaminants actually cause measurable human or animal health effects.
Acknowledgment
This study was supported by the Grant no. 30571381 “Study on spread model of microorganism aerosol from animal stable” from the National Natural Science Foundation of China (NSFC).
References
- Ahmed W., Neller R., Katouli M. Host species-specific metabolic fingerprint database for Enterococci and Escherichia coli and its application to identify sources of fecal contamination in surface waters. Appl. Environ. Microbiol. 2005;71:4461–4468. doi: 10.1128/AEM.71.8.4461-4468.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersen A. New sampler for the collection, sizing, and enumeration of viable airborne particles. J. Bacterial. 1958;76:471–484. doi: 10.1128/jb.76.5.471-484.1958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bitton G., Farrah S.R., Ruskin R.H., Butner J., Chou Y.J. Survival of pathogenic and indicator organisms in groundwater. Groundwater. 1983;21:405–410. [Google Scholar]
- Borges L.G.d.A., Vechia V.n.D., Corçã G. Characterisation and genetic diversity via REP-PCR of Escherichia coli isolates from polluted waters in southern Brazil. FEMS Microbiol. Ecol. 2003;45:173–180. doi: 10.1016/S0168-6496(03)00147-8. [DOI] [PubMed] [Google Scholar]
- Brown J.K.M., Hovmoller M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science. 2002;297:537–541. doi: 10.1126/science.1072678. [DOI] [PubMed] [Google Scholar]
- Centers for Disease Control and Prevention (CDC) Follow-up of deaths among US postal service workers potentially exposed to Bacillus anthracis—district of Columbia, 2001–2002. MMWR Weekly. 2003;52:937–938. [PubMed] [Google Scholar]
- Chai T.J., Chai J.Q., Mueller W. Airborne microorganisms of a calf stable and spreading to its environment. Chin. J. Prev. Vet. Med. 1999;21:311–313. [Google Scholar]
- Chai T.J., Zhang J.X., Hou Y.P., Zhao Y.L. Spread of microorganism aerosol in animal house to its environments. Anim. Husb. Vet. Med. 2001;33:10–12. (in Chinese) [Google Scholar]
- Chai T.J., Zhao Y.L., Liu W.B., Liu H. The resistance against antibiotics of bacteria from a poultry house and their spreading to surroundings of the house. Chin. J. Prev. Vet. Med. 2003;25:209–214. [Google Scholar]
- Chansiripornchai N., Ramasoota P., Sasipreyajan J., Svenson S.B. Differentiation of avian Escherichia coli (APEC) isolates by random amplified polymorphic DNA (RAPD) analysis. Vet. Microbiol. 2001;80:77–83. doi: 10.1016/s0378-1135(00)00380-1. [DOI] [PubMed] [Google Scholar]
- Cheng D.R., Sun H.C., Xu J.S., Gao S. PCR detection of virulence factor genes in Escherichia coli isolates from weaned piglets with edema disease and/or diarrhea in China. Vet. Microbiol. 2006;115:320–328. doi: 10.1016/j.vetmic.2006.02.013. [DOI] [PubMed] [Google Scholar]
- De Moura A.C., Irino K., Vidotto M.C. Genetic variability of avian Escherichia coli isolates evaluated by enterobacterial reptitive intergenic consensus and repetitive extragenic palindromic polymerase chain reation. Avian. Dis. 2001;45:173–181. [PubMed] [Google Scholar]
- Dombek P.E., Johnson L.K., Zimmerley S.T., Sadowsky M. Use of repetitive DNA sequences and the PCR to differentiate Escherichia coli isolates from human and animal sources. Appl. Environ. Microbiol. 2000;66:2572–2577. doi: 10.1128/aem.66.6.2572-2577.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donaldson A. Airborne spread of foot-and-mouth disease. Microbiol. Today. 1999;26:118–119. [Google Scholar]
- Donaldson A.I., Gloster J., Harvey L.D., Deans D.H. Use of prediction models to forecast and analyse airborne spread during the foot-and-mouth disease outbreaks in Brittany, Jersey and the Isle of Wight in 1981. Vet. Record. 1982;110:53–57. doi: 10.1136/vr.110.3.53. [DOI] [PubMed] [Google Scholar]
- Donham K.J., Popendorf W., Palmgren U., Larsson L. Characterization of dusts collected from swine confinement buildings. Am. J. Ind. Med. 1986;10:294–297. doi: 10.1002/ajim.4700100318. [DOI] [PubMed] [Google Scholar]
- Duan H.Y., Chai T.J., Wang Y.X., Mueller W., Zucker B.A. Concentration of airborne endotoxins and airborne bacteria in Chinese rabbit houses. Berl. Münch Tierärztl Wochenschr. 2006;119:40–44. [PubMed] [Google Scholar]
- Eubank S., Guclu H., Anil Kumar V.S., Marathe M.V., Srinivasan A., Toroczkai Z., Wang N. Modelling disease outbreaks in realistic urban social networks. Nature. 2004;429:180–184. doi: 10.1038/nature02541. [DOI] [PubMed] [Google Scholar]
- Georghiou P.R., Hamill R.J., Wright C.E., Versalovic J., Koeuth T., Watson D.A., Lupski J.R. Molecular epidemiology of infections due to Enterobacter aerogenes, identification of hospital-associated strains by molecular techniques. Clin. Infect. Dis. 1995;20:84–94. doi: 10.1093/clinids/20.1.84. [DOI] [PubMed] [Google Scholar]
- Gilson E., Clement J.M., Brutlag D., Hofnung M. A family of dispersed repetitive extragenic palindromic DNA sequence in E. coli. EMBO J. 1984;3(6):1417–1421. doi: 10.1002/j.1460-2075.1984.tb01986.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffith J.F., Weisberg S.B., McGee C.D. Evaluation of microbial source tracking methods using mixed fecal sources in aqueous test samples. J. Water Health. 2003;01(4):141–151. [PubMed] [Google Scholar]
- Gyles C.L., Fairbrother J.M. Escherichia coli. In: Gyles C.L., Prescott J.F., Songer J.G., Theon C.O., editors. Pathogenesis of Bacterial Infections in Animals. Blackwell Publishing Professional; Ames, Iowa: 2004. pp. 193–214. [Google Scholar]
- Hamill R.J., Houston E.D., Georgjiou P.R., Wright C.E., Koza M.A., Cadle R.M., Goepfert P.A., Lewis D.A., Zenon G.J., Clarridge J.E. An outbreak of Burkholderia (formerly Pseudomonas) cepacia respiratory tract colonization and infection associated with nebulized albuterol therapy. Ann. Intern. Med. 1995;122:762–766. doi: 10.7326/0003-4819-122-10-199505150-00005. [DOI] [PubMed] [Google Scholar]
- Hanes N.B., Fragala R. Effect of seawater concentration on survival of indicator bacteria. Res. J. Water Pollut. Control Fed. 1967;39:97–104. [PubMed] [Google Scholar]
- Hermann J.R., Hoff S.J., Yoon K.J., Roof M., Burkhardt A., Zimmerman J. Effect of temperature and relative humidity on the stability of infectious porcine reproductive and respiratory syndrome virus in aerosols. Vet. Res. 2007;38:81–93. doi: 10.1051/vetres:2006044. [DOI] [PubMed] [Google Scholar]
- Hojovec J., Fiser A., Kubicek K.Z. Die Rolle von Indikator keimen fuer die Beurteilung der Stalluft. Mh. Vet. Med. 1977;32:766–769. [Google Scholar]
- Hulton C.S., Higgins C.F., Sharp P.M. ERIC sequences: a novel family of repetitive elements in the genomes of Escherichia coli, Salmonella typhimuirum and other enterobacteria. Molec. Microbiol. 1991;5:825–834. doi: 10.1111/j.1365-2958.1991.tb00755.x. [DOI] [PubMed] [Google Scholar]
- Imberechts H., De Greve H., Lintermans P. The pathogenesis of edema disease in pigs. A review. Mol. Microbiol. 1992;31:221–233. doi: 10.1016/0378-1135(92)90080-d. [DOI] [PubMed] [Google Scholar]
- Indest K.J., Betts K., Furey J.S. Application of oligonucleotide microarrays for bacterial source tracking of environmental Enterococcus sp. isolates. Int. J. Environ. Res. Public Health. 2005;2:175–185. doi: 10.3390/ijerph2005010175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johson J.R., Calbots C. Improved repetitive-element PCR fingerprinting of Salmonella enterica with the use of extremely elevated annealing temperatures. Clin. Diag. Lab. Immunol. 2000;7:258–264. doi: 10.1128/cdli.7.2.258-264.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones A.M., Harrison R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—a review. Sci. Total Environ. 2004;326:151–180. doi: 10.1016/j.scitotenv.2003.11.021. [DOI] [PubMed] [Google Scholar]
- Keswick B.H., Gerba C.B., Secor S.L., Cech I. Survival of enteric viruses and indicator bacteria in groundwater. J. Environ. Sci. Health A. 1982;17:903–912. [Google Scholar]
- Kim K.Y., Kim C.N. Airborne microbiological characteristics in public buildings of Korea. Build. Environ. 2007;42:2188–2196. [Google Scholar]
- Klontz K.C., Hynes N.A., Gunn R.A., Wilder M.H., Harmon M.W., Kendal A.P. An outbreak of influenza a/Taiwan/1/86 (H1N1) infections at a naval base and its association with airplane travel. Am. J. Epidemiol. 1989;129:341–348. doi: 10.1093/oxfordjournals.aje.a115137. [DOI] [PubMed] [Google Scholar]
- Landman W.J., Schrier C.C. Eradication from commercial poultry is still not in sight. Avian Influenza. 2004;129:782–796. [PubMed] [Google Scholar]
- Lee A.K.Y., Lau A.P.S., Cheng J.Y.W., Fang M., Chan C.K. Source identification analysis for the airborne bacteria and fungi using a biomarker approach. Atmos. Environ. 2007;41:2831–2843. [Google Scholar]
- Lighthart B. Microbial aerosols: estimated contribution of combine harvesting to an airshed. Appl. Environ. Microbiol. 1984;47:430–432. doi: 10.1128/aem.47.2.430-432.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lupsiki J.R., Weinstock G.M. Short, interspersed repetitive DNA sequence in prokaryotic genomes. J. Bacteriol. 1992;174:4525–4529. doi: 10.1128/jb.174.14.4525-4529.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marthi B., Fieland V.P., Walter M., Seidler R.J. Survival of bacteria during aerosolization. Appl. Environ. Microbiol. 1990;56:3463–3467. doi: 10.1128/aem.56.11.3463-3467.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin B., Humbert O., Camara M., Guenzi E., Walker J., Mitchell T., Andrew P., Prudhomme M., Alloing G., Hakenbeck R., Morrison D.A., Boulnois G.J., Claverys J.P. A highly conserved repeated DNA element located in the chromosome of Streptococcus pneumoniae. Nucleic Acids Res. 1992;20:3479–3483. doi: 10.1093/nar/20.13.3479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McFeters G.A., Bissonnette G.K., Jezeski J.J., Thomoson C.A., Stuart D.G. Comparative survival of indicator bacteria and enteric pathogens in well water. Appl. Microbiol. 1974;27:823–829. doi: 10.1128/am.27.5.823-829.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLellan S.L., Daniels A.D., Salmore A.K. Genetic characterization of Escherichia coli populations from host sources of fecal pollution by using DNA fingerprinting. Appl. Environ. Microbiol. 2003;69:2587–2594. doi: 10.1128/AEM.69.5.2587-2594.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meays C.L., Broersma K., Nordin R., Mazumder A. Source tracking fecal bacteria in water: a critical review of current methods. J. Environ. Manage. 2004;73:71–79. doi: 10.1016/j.jenvman.2004.06.001. [DOI] [PubMed] [Google Scholar]
- Mehta Y.R., Mehta A., Rosato Y.B. ERIC and REP banding patterns and sequence analysis of the internal transcribed spacer region of Stemphylium solani of cotton. Curr. Microbiol. 2002;44:323–328. doi: 10.1007/s00284-001-0026-4. [DOI] [PubMed] [Google Scholar]
- Mims S.A., Mims F.M., III Fungal spores are transported long distances in smoke from biomass fires. Atmos. Environ. 2004;38:651–655. [Google Scholar]
- Northcutt J.K., Jones D.R., Musgrove M.T. Airborne microorganisms during the commercial production and processing of Japanese Quail. Int. J. Poultry Sci. 2004;3:242–247. [Google Scholar]
- Olive D.M., Bean P. Principles and applications of methods for DNA-based typing of microbial organisms. J. Clin. Microbiol. 1999;37:1661–1669. doi: 10.1128/jcm.37.6.1661-1669.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Omura I., Omura M., Hashmoto Y. Viability and adaptability of E. coli and Enterococcus group to salt water with high concentration of sodium chloride. Water Sci. Technol. 1982;14:115–126. [Google Scholar]
- Prazmo Z., Dutkiewicz J., Skorska C., Sitkowska J., Cholewa G. Exposure to airborne Gram-negative bacteria, dust, and endotoxin in paper factories. Ann. Agric. Environ. Med. 2003;10:93–100. [PubMed] [Google Scholar]
- Rodriguez-Barradas M.C., Hamill R.J., Houston E.D., Georghiou P.R., Clarridge J.E., Regnery R.L., Koehler J.E. Genomic fingerprinting of Bartonella species by repetitive element PCR for distinguishing species and isolates. J. Clin. Microbiol. 1995;33:1089–1093. doi: 10.1128/jcm.33.5.1089-1093.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rohlf, F.J., 2000. NTSYS-pc. Numerical Taxonomy and Multi-variate Analysis System, Version 2.1. Exeter Software, Setauket, New York.
- Scharek L., Guth J., Reiter K., Weyrauch K.D., Taras D., Schwerk P., Schierack P., Schmidt M.F., Wieler L.H., Tedin K. Influence of a probiotic Enterococcus faecium strain on development of the immune system of sows and piglets. Vet. Immunol. Immunopathol. 2005;105:151–161. doi: 10.1016/j.vetimm.2004.12.022. [DOI] [PubMed] [Google Scholar]
- Schierack P., Steinrück H., Kleta S., Vahjen W. Virulence factor gene profiles of Escherichia coli isolates from clinically healthy pigs. Appl. Environ. Microbiol. 2006;72:6680–6686. doi: 10.1128/AEM.02952-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selander R.K., Musser J.M., Caugant D.A., Gilmour M.N., Whittam T.S. Population genetics of pathogenic bacteria. Microb. Pathog. 1987;3:1–7. doi: 10.1016/0882-4010(87)90032-5. [DOI] [PubMed] [Google Scholar]
- Sharples G.J., Lloyd R.G. A novel repeated DNA sequence located in the intergenic regiens of bacterial chromosomes. Nucleic Acids Res. 1990;18:6503–6508. doi: 10.1093/nar/18.22.6503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinn E.A., Smith G.W., Prospero J.M., Betzer P., Hayes M.L., Garrison V., Barber R.T. African dust and the demise of Caribbean coral reefs. Geophys. Res. Lett. 2000;27:3029–3032. [Google Scholar]
- Stern M.J., Ames G.F., Smith N.H., Robinson E.C., Higgins C.F. Repetitive extragenic palindromic sequence: a major component of the bacterial genome. Cell. 1984;37:1015–1026. doi: 10.1016/0092-8674(84)90436-7. [DOI] [PubMed] [Google Scholar]
- Stewart S.L., Grinshpun S.A., Willeke K., Terzieva S., Ulevicius V., Donnelly J. Effect of impact stress on microbial recovery on an agar surface. Appl. Environ. Microbiol. 1995;61:1232–1239. doi: 10.1128/aem.61.4.1232-1239.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Belkum A. DNA fingerprinting of medically important microorganisms by use of PCR. J. Clin. Microb. Rev. 1994;7:174–184. doi: 10.1128/cmr.7.2.174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Belkum A., Sluijuter M., Groot R.d., Verbrugh H., Hermans P. Novel BOX repeat PCR assay for high-resolution typing of strepotococcus pneumoniae strains. J. Clin. Microb. 1996;34:1176–1179. doi: 10.1128/jcm.34.5.1176-1179.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Versalovic J., Koeuth T., Lupski J.R. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nuclic Acids Res. 1991;19:6823–6831. doi: 10.1093/nar/19.24.6823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Versalovic J., Scheider M., de Bruijn F.J., Lupski J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Meth. Cell. Mol. Biol. 1994;5:25–40. [Google Scholar]
- Wait D.A., Sobsey M.D. Comparative survival of enteric viruses and bacteria in Atlantic Ocean seawater. Water Sci. Technol. 2001;43:139–142. [PubMed] [Google Scholar]
- Weiss R.A., McMichael A.J. Social and environmental risk factors in the emergence of infectious diseases. Nat. Med. 2004;10:S70–S76. doi: 10.1038/nm1150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wieler L.H., Ilieff A., Herbst W., Bauer C., Vieler E., Bauerfeind R., Failing K., Klos H., Wengert D., Baljer G., Zahner H. Prevalence of enteropathogens in suckling and weaned piglets with diarrhoea in southern. J. Vet. Med. B. 2001;48:151–159. doi: 10.1111/j.1439-0450.2001.00431.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woods C.R., Versalovic J., Koeuth T., Lupski J.R. Analysis of relationships among isolates of Citrobacter diversus by using DNA fingerprints generated by repetitive-sequence based polymerase chain reaction. J. Clin. Microbiol. 1992;30:2921–2929. doi: 10.1128/jcm.30.11.2921-2929.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao M.L., Zhang B., Chai T.J. Antibiotic resistance of airborne Escherichia coli from hen houses and rabbitry and their spreading to surroundings. J. Northwest A & F Univ. (Nat. Sci. Ed.) 2007;35:60–64. (in Chinese) [Google Scholar]
- Zucker B.A., Trojan S., Mueller W. Airborne gram-negative bacterial flora in animal houses. J. Vet. Med. B. 2000;47:37–46. doi: 10.1046/j.1439-0450.2000.00308.x. [DOI] [PubMed] [Google Scholar]