

Abstract
The aim of this study was to estimate the therapeutic potential of specific egg yolk immunoglobulin (IgY) on dermatophytosis caused by Trichophyton rubrum. The IgY was produced by immunizing hens with cell wall proteins of T. rubrum, extracted from eggs by PEG precipitation and then purified by ammonium sulfate precipitation. The cross-reactivity (CR) with other fungi, growth inhibition on T. rubrum in vitro and therapeutic effect on T. rubrum infection in BALB/C mice of the specific IgY were then evaluated. Anti- T. rubrum cell wall proteins IgY (anti-trCWP IgY) presented a certain degree of cross-reactivity with different fungi. In the in vitro and in vivo activity researches, Anti-trCWP IgY showed a significant dose-dependent growth inhibitory effect on T. rubrum in vitro and a significant dose-dependent therapeutic effect on T. rubrum infection in BALB/C mice.
Keywords: T. rubrum, Dermatophytosis, IgY, Cell wall proteins
Highlights
-
•
Egg yolk immunoglobulin produced using Trichophyton rubrum cell wall proteins as antigen.
-
•
Anti-T. rubrum IgY present cross-reactivity with other fungi.
-
•
Anti- T. rubrum IgY inhibited T. rubrum growth in vitro.
-
•
Anti- T. rubrum IgY alleviated T. rubrum infection in BALB/C mice.
1. Introduction
Dermatophytosis is a global health problem with high morbidity [1,2]. According to the World Health Organization (WHO), dermatophytes affect about 25% of the world population. It is estimated that about 30%–70% of the infected adults are asymptomatic, and the incidence of the infection increases with age [3]. Dermatophytes have been classified into three genera, Trichophyton, Microsporum and,Epidermophyton. T. rubrum is particularly responsible for the major infections of dermatophytes [4,5], which is a kind of filamentous fungus that may cause tinea corporis, tinea cruris, and tinea pedis [6].
At present, most popular treatment options for dermatophytosis are topical use of antifungal chemicals such as amphotericin B, fluconazole, itraconazole, voriconazole, posaconazole, isavuconazole and terbinafine [7,8], oral antifungal drugs such as triazoles, griseofulvin, and terbinafine for systemic therapy of severe cases [9]. Because of the fungal resistance problem of the antifungal chemicals, the research of their suitable alternatives is continuing [10].
Human's regular interactions with fungi rarely result in diseases in persons with normal immunity. However, Those with immune impairment are at high risk of fungal diseases. The recognition that impaired immunity is central to the pathogenesis of many fungal diseases and that antifungal agents are often ineffective in the setting of immune defects, provides a strong rationale for the development of immune-based therapies, or immunotherapy, for fungal diseases [11].
IgY is produced by hens to provide their offspring with an effective humoral immunity against the common avian pathogens until full maturation of their own immune system. Effective protection against Salmonella enteritidis, Salmonella e. Typhimurium, Campylobacter jejuni, Escherichia coli ETEC, murine and bovine rotavirus, and bovine coronavirus infections in mice, pig, and calves has been obtained with the use of passively-administered egg yolk-derived antibodies [12]. Ample data have demonstrated the protection of IgY against different kinds of bacteria, virus, and parasites There are also reports about anti-fungi IgYs [13,14], reminding us that it may also be effective against T. rubrum.
To explore the therapeutic potential of IgY on dermatophytosis caused by T. rubrum, we immunized the hens with cell wall proteins of T. rubrum. The anti- T. rubrum IgY was then extracted from eggs by PEG precipitation and purified by ammonium sulfate precipitation. The purity, titer, and specificity of the IgY were tested to evaluate the IgY production process. A series of researches about cross-reactivity (CR) with other fungi, growth inhibition on T. rubrum in vitro and therapeutic effect on T. rubrum infection in BALB/C mice of the IgY have been carried out to assess the selectivity and therapeutic potential of the IgY.
2. Material and methods
2.1. Antigen preparation
T. rubrum (NBRC5467, provided by Dr. Song-chao Yin from the Third Affiliated Hospital of Sun Yat-sen University) was cultured at 28 °C for 6 days in Sabouraud agar medium. The T. rubrum spores were washed with sterile phosphate-buffered saline (PBS, pH7.4, 0.01 M), and then inoculated to the Liquid Sabouraud Medium, shakingly cultured at 28 °C, 160 rpm for 5 days. After that, T. rubrum hyphae were harvested by centrifugation, washed 3 times with sterile PBS (pH7.4, 0.01 M) and then freeze-dried.
The cell wall proteins were extractedby referring to a reported method [15]. One g of hyphae was ground thoroughly in liquid nitrogen with a mortar and pestle into powder, dissolved with 20 mL PBS (pH7.4, 0.01 M); 200 μL 10 mM PMSF (Beyotime) was added into the solution. The solution was put in an ice-bath and the cells were broken with an Ultrasonic cell breakup instrument (2 s × 300 times, interval 5s). The precipitate was collected by centrifugation (4 °C, 6000 g, 10 min) and washed five times with ice-cold water. After that a cold 0.1 M NaOH solution was added to the precipitate and stirred at 4 °C for 24hr. The supernatant was harvested by centrifugation(4 °C, 12000g, 20min), and pH was adjusted to 7.0 with cold 0.1 M HCl solution. The solution containing cell wall proteins was dialyzed with PBS (pH7.4, 0.01 M) for 72hr and diluted to 1 mg/mL with PBS (pH7.4, 0.01 M). The concentration of the cell wall proteins was measured by BCA assay. The protein components of the cell wall proteins were analyzed by SDS-PAGE (12%) method (Bio-Rad, USA).
2.2. Immunization of laying hens
The cell wall proteins diluted to 1 mg/mL was mixed thoroughly with an equal amount of Freund's adjuvant (Sigma Chemicals, St Louis, MO, USA) until the emulsion was stable. 4 healthy 20-week-old Roman laying hens housed at an experimental animal center were immunized and boosted via an intramuscular route (i.m. 0.5mL/hen, musculus pectoralis, left and right side) with the antigen emulsified in Freund's complete adjuvant (for the primary immunization) or Freund's incomplete adjuvant (for the subsequent booster injections). The birds were given four booster injections at two-week intervals. Eggs were collected daily from 20d after the first immunization and stored at 4 °C. Eggs collected from unimmunized hens were used for the negative control [16].
2.3. Isolation and purification of IgY
Isolation and purification of IgY were carried out according to reported studies [17]. Eggs were disinfected with 75% alcohol; the egg yolks were then separated from the whites with an egg yolk separator. The collected egg yolks were dissolved with a triple volume of PBS (pH7.4, 0.01 M) containing 3.5% PEG6000 (w/v) and stirred for 40 min; the mixture was centrifuged at 4 °C, 12000g for 20 min. The supernatant was collected, PEG6000 was added to adjust the final polymer concentration to 12% (w/v), and the mixture was stirred for 40 min. The precipitate containing the crude IgY was harvested by centrifugation at 4 °C, 12000g for 20 min. The extracted crude IgY was dissolved into the original volume of yolk in PBS (pH7.4, 0.01 M); the saturated ammonium sulfate solution was added to the equal volume and the mixture was stirred overnight at 4 °C. The precipitate was collected by centrifugation (4 °C, 12000g for 20 min) and washed with a 1:3 saturated ammonium sulfate solution. The precipitate was dissolved and dialyzed against PBS (pH7.4, 0.01 M) and then freeze-dried, the IgY powder obtained was stored at −20 °C.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed to determine the purity of the IgY. A 10% polyacrylamide gel was used (Bio-Rad Laboratories, USA). The analysis was conducted under reduced conditions. The sample(1 mg/mL) was mixed with sample buffer (5 × ) and held for 5–8 min at 100 °C. Ten micro liters of the sample was loaded into each well. Pre-stained protein standard (Fermentas, Lithuania) was used as a molecular weight marker. The protein bands were visualized with Coomassie Brilliant Blue R250 (Sino Pharma Shanghai Chemical Reagent Company). The gel was analyzed by Bio-Rad image analysis software.
2.4. Characterization of IgY by western-blotting
To confirm the specificity of the anti-trCWP IgY, western blotting was conducted according to a previously published method [18]. The extracted T. rubrum cell well protein was subjected to SDS–PAGE with a 12% polyacrylamide gel. After electrophoresis, the gel was equilibrated in the transfer buffer for 15–30 min and the proteins were then electrically transferred onto a PVDF membrane (Millipore, USA) for 3 h at 350 mA, 4 °C. The membrane was stirred in freshly prepared ponceau solution (Beyotime) for 1 min and washed with distilled water, and then cut into 0.5 cm strips, which were blocked with 5% fat-free milk for 2 h at room temperature and washed with Tris buffer solution(0.02 M, pH7.6) containing 0.05% Tween 20 (TBS-T). The strips were then incubated overnight at 4 °C with a 1:100 dilution of anti-trCWP IgY (1 mg/mL). IgY from non-immunized hens was used as a negative control. After incubation, the strips were washed 3 times with TBS-T and incubated with HRP conjugated rabbit anti-chicken IgY (Promega, USA) diluted 1:10000. Following 1 h of incubation at room temperature, the strips were washed three more times. After washing, ECL solution (Millipore, USA) was added to the strips in a dark room with red light as the light source. The strips were photographed and developed.
2.5. Enzyme-linked immunosorbent assay
The titer of the specific IgY was determined by indirect ELISA as described previously by Ref. [19]. Ninety-six-well ELISA plate was coated with T. rubrum whole-cell solution (1.2 × 108 CFU/mL in pH7.4, 0.01 M PBS) at 37 °C for 2 h and 4 °C overnight. The plate was washed 3 times with pH7.4, 0.01 M PBS containing 0.05% Tween 20 (PBST). Blocking was performed by adding 200 μL of 3.3% (w/v) fat-free milk, incubated at 37 °C for 1 h. After washed 3 times with PBST, samples (100μL/well) were added to each well and incubated at 37 °C for 1 h. The IgY (1 mg/mL) was 2-fold serial diluted in PBS (pH7.4, 0.01 M). The IgY from non-immunized hens was used as a negative control. The plate was washed 3 times again and 100μL/well of HRP affiliate rabbit anti-chicken IgY (Promega, USA; 1:5000) was added and the plate was incubated at 37 °C for 1 h. After being washed 3 times, 100 μL of TMB substrate solution (Huamei biological engineering co., Ltd.) was added to each well for 15 min at 37 °C. After that, the reaction was stopped by the addition of 50 μL of 2 M H2SO4 to each well. The optical densities of the wells were determined at 405 nm with a plate reader (Bio-Rad, USA). When ODsample/ODnegative > 2.1 and ODnegative > 0.1, the maximum dilution multiple of the sample was determined as the IgY titer.
2.6. Cross-reactivity with other fungi
To estimate the cross-reactivity (CR) of anti-trCWP IgY against other fungi, indirect ELISAs were performed by the use of antigens from different fungi according to the method described previously. Trichophyton mentagrophytes was cultured in Sabouraud Dextrose Agar at 28 °C for 6 days. Aspergillus flavus (ATCC 11492), Candida albicans (ATCC 10231), Penicillium citrinum (AS 3.2788), Aspergillus niger (ATCC 16404), Rhizopus nigricans (GIM 3.125), Mucor racemosus (GIM 3.86) and Penicillium cyclopium (GIM 3.247) (obtained from the Food Analysis Laboratory Guangdong University of Technology) were cultured in Rose Bengal medium at 28 °C for 6 days [20]. They were suspended and diluted to 1.2 × 108 CFU/mL with pH7.4, 0.01 M PBS to be used as antigens for Elisa test with anti-trCWP IgY. IgY from non-immunized hens was used as a negative control. The cross-reactivity rate was calculated as follows:
| CR(%)=(DRf/DRt) × 100% |
In the formula, DRf was the titer of IgY against other fungi and DRt was the titer of IgY against T. rubrum.
2.7. Growth inhibition on T. rubrum in vitro
The inhibitory effect of specific IgY on T. rubrum growth in vitro was detected according to the M38-A2 method by the Clinical and Laboratory Standards Institute (CLSI) [21]. T. rubrum was cultured at 28 °C for 7 days in Oats agar medium (30 g oats and 20 g agar in 1L water). The T. rubrum spores were washed with sterile PBS (pH7.4, 0.01 M) and diluted to 1.0 × 104 CFU/mL with RPMI1640. The specific IgY was dissolved in sterile PBS (pH7.4, 0.01 M), diluted to 40 mg/mL, 20 mg/mL, 10 mg/mL, 5 mg/mL, 2.5 mg/mL, 1.25 mg/mL and 0.625 mg/mL, and then sterilized by filtration before use. 100 μL of cell suspension and 100 μL of different concentrations of IgY solution were added into one well of the polystyrene plate and incubated at 28 °C for 72 h, each IgY concentration being triplicate. Cell suspension plus PBS (pH7.4, 0.01 M) was used as a blank control and IgY from non-immunized hens diluted with RPMI1640 was used as a negative control. The absorbencies of the mix solutions were then measured at 660 nm (Bio-Rad, USA). The growth-inhibitory curve of the specific IgY on T. rubrum was graphed by using the concentration of IgY as abscissa while OD660 of the mix solution as ordinate.
2.8. Application of specific IgY in a T. rubrum mouse model
The mouse model infected with T. rubrum was built fby referring to a reported method [22]. Experiments were carried out by following an accepted ethical protocol according to the guidelines of the Guangdong University of Technology for animal experiments(GDUT/SBPS/2017060501). The number of mice was reduced to an absolute minimum. Animal suffering was limited to the experiment need and the sacrifice was done with carbon dioxide asphyxiation. BALB/C mice (8 weeks old, purchased from Medical Experimental Animal Center of Guangdong province) were maintained at a 12:12 h light/dark cycle with 18~26 °C and 30–60% relative humidity, and provided ad-lib with food pellets and water. The animals were immunosuppressed with a daily intramuscular injection of triamcinolone acetonide injection (Nanjing Jindun Animal Pharmaceutical Co., Ltd. 0.3 mL per mouse) for 3 days. The back of each animal was depilated with 8% Na2S solution, and the hairless skin was slightly rubbed with a piece of sterilized 1200 mesh sandpaper. The damaged skin (2 cm × 2 cm area) was inoculated with 200 μL T. rubrum conidia suspension (1.0 × 106conidia/mL). After the inoculation, each animal was i.p. injected with dexamethasone sodium phosphate injection (Shanghai Shengguang Animal Health products Co., Ltd., 0.4 mL per mouse) daily for 3 days. After the successful fungal infection, 20 animals were divided randomly into 5 groups and treated with various drugs by smearing the drugs on the infected skin daily for 14 days. Three groups were treated with 50 μl anti-trCWP IgY at concentrations of 5 mg/mL, 10 mg/mL, and 20 mg/mL. The model control (MC) group was treated with 50 μl sterile PBS (pH7.4, 0.01 M). The positive control group was treated with 1 mg/mL fluconazole (Nanjing Zhuopu Biotechnology Co., Ltd.) solution.
The efficacy of specific IgY was evaluated by clinical lesion score and histopathological examination of skin tissues. For clinical evaluation, changes in redness, ulcerative scaling, and hair loss at the inoculation site were visually examined and recorded by use of a lesion score. The cured, improved and aggravated were scored as 0, 1and 2 respectively. The skin lesion score of mice was the sum of the three scoring items. Animals were sacrificed for skin biopsy 24 h after the final drug administration. Infected skin tissues were excised, fixed in neutral buffered formalin, embedded in paraffin, and stained with hematoxylin/eosin (HE) for histopathological examination.
The experimental data were expressed as mean ± SD. Statistical differences between mean values were evaluated by the program SPSS for Windows, v. 17.0 (SPSS Inc.; Chicago, IL, USA). Differences with p < 0.05 were considered significant and those with p < 0.01 were considered highly significant.
3. Results
3.1. Antigen preparation
The CWP of T. rubrum was extracted from T. rubrum hyphae. As shown in Fig. 1 , SDS-PAGE displayed that the CWP had about eight bands ranging from 35KD to 130 KD.
Fig. 1.
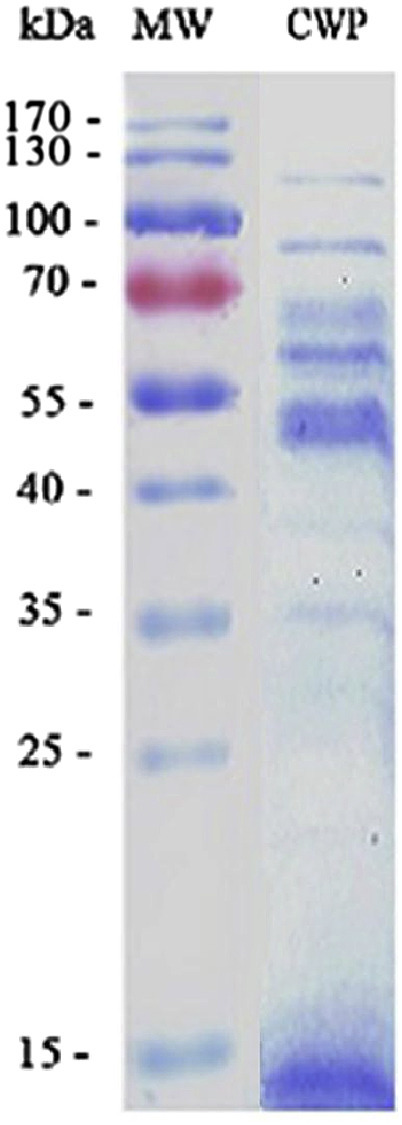
SDS-PAGE result of extracted antigen protein. Extracted cell wall proteins of T. rubrum were subjected to SDS-PAGE with a 12% separation gel. Line Mw: Molecular weight markers; Line CWP: Extracted cell wall proteins of T. rubrum.
3.2. Isolation and purification of IgY
IgY was extracted from the egg yolk through the two-step PEG precipitation method and purified with saturated ammonium sulfate. As shown in Fig. 2 , SDS–PAGE revealed that the IgY preparation was pure and dissociated into two protein bands with molecular weights of 68KD and 27KD, which was consistent with other reports (da Silva and Tambourgi 2010). Calculated with the protein content test results, the yield of specific IgY (freeze-dried powder) extracted and purified from egg yolkwas 8.7–9.1 mg/mL with a purity of more than 90%.
Fig. 2.
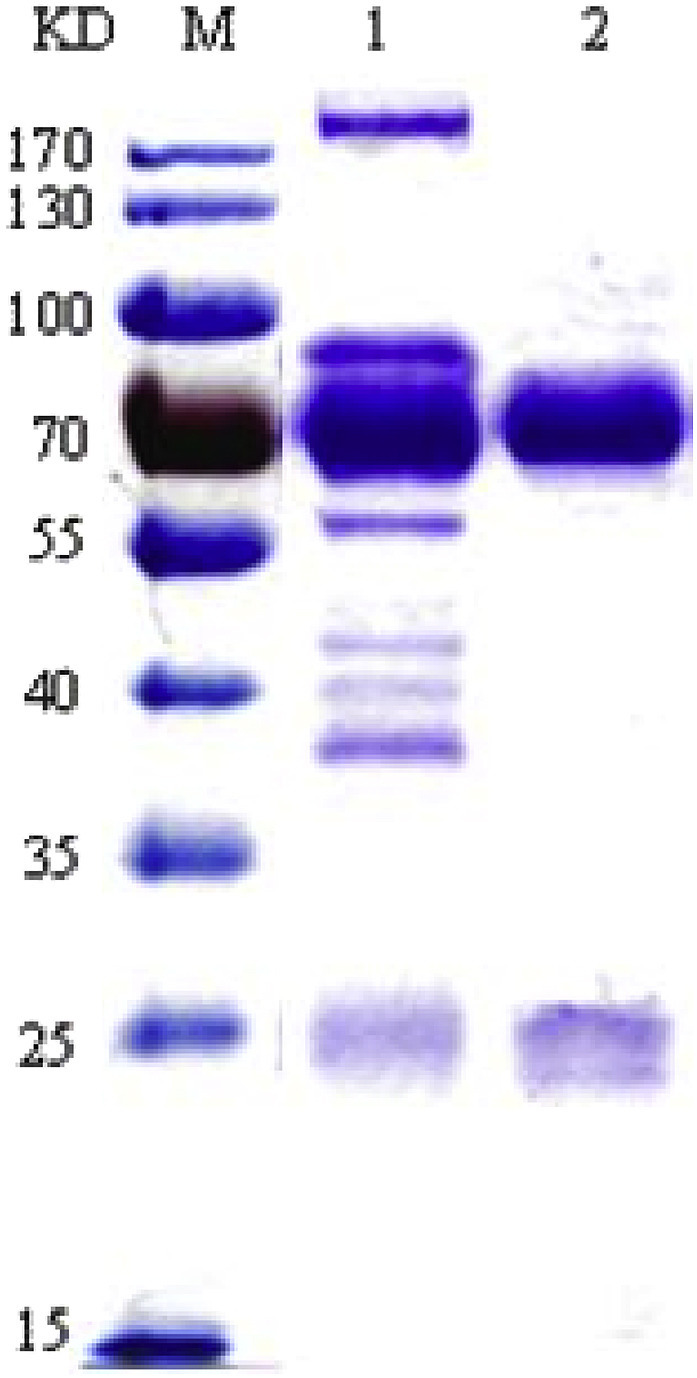
SDS-PAGE result of extracted IgY and purified IgY. Crude anti-trCWP IgY extracted by PEG precipitation from egg yolks and anti-trCWP IgY purified by ammonium sulfate precipitation were subjected to SDS-PAGE with a 10% separation gel. Lane M: Molecular weight markers; Lane 1: Crude anti-trCWP IgY; Lane 2: Purified anti-trCWP IgY.
3.3. Characterization of IgY by western-blotting
As shown in Fig. 3 , through immunoblotting, extracted IgY could be bound to the second antibody. The specific IgYwas found specificallybound to related antigen protein, while a negative IgY was not.
Fig. 3.
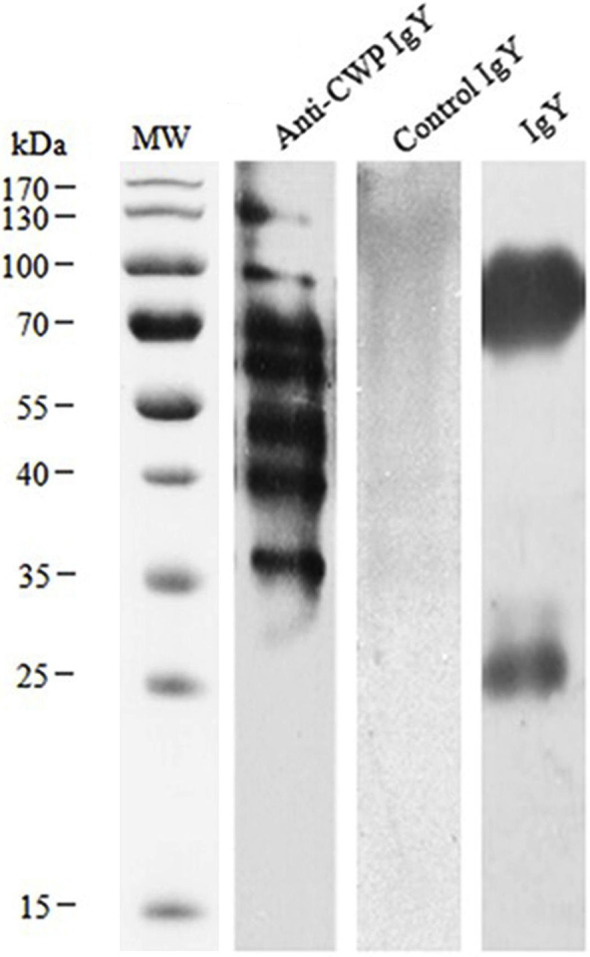
Western blotting analysis of anti-trCWP IgY response to antigen protein of T. rubrum. The antigen protein was subjected to SDS–PAGE (12%) under reduced condition, electrically transferred onto PVDF membrane and probed with 1:100 diluted IgYs from immunized and non-immunized hens. Lane MW: molecular marker; Lane Anti-CWP IgY: CWP with Anti-CWP IgY; Lane Control IgY: CWP with non-immunized IgY; Lane IgY: anti-CWP IgY with the second antibody.
3.4. Titer of specific IgY
Eggs were collected 20d after the first immunization; ELISA was performed to assess the immune response of the purified IgY. The titer of specific IgY began to increase on the 20th day after the first immunization, as shown in Fig. 4 , and reached the highest level on the 40th day after the first immunization. The highest titer of specific IgY could reach 1:16 000.and was maintained for 30 days before declining (Fig. 4).
Fig. 4.

The titer of anti-trCWP IgY in egg yolks increasing over time. Egg yolks were collected from immunized hens over a course of 100 days after the first immunization.
3.5. Cross-reactivity with other fungi
Cross-reactivity (CR) of anti-T. rubrum IgY with other Trichophyton strain or other fungi were tested by Elisa by using antigens from different fungi. As shown in Fig. 5 , specific IgY showed a certain degree of cross-reactivity with different fungi. The most significant cross-reactivity was with T. mentagrophytes, a Trichophyton strain (Fig. 5).
Fig. 5.

Cross-reactivity of anti-trCWP IgY with other fungi. Eight kinds of fungi were used as coating antigens (A. flavus, C. albicans, P. citrinum, A. niger, R. nigricans, M. racemosus, P. cyclopium, and T. mentagrophytes). The CR (%) of anti-CWP IgY with T. rubrum was considered to be 100%.
3.6. Growth inhibition on T. rubrum in vitro
As shown in the growth inhibitory curve (Fig. 6 ), compared with the control IgY, specific IgY showed a significant dose-dependent inhibitory effect on cell growth of T. rubrum.
Fig. 6.

The growth inhibitory effect of specific IgY to T. rubrum. The T. rubrum cells were diluted in RPMI1640 medium and 1:1 mixed with PBS (blank control), 0.625, 1.25, 2.5, 5, 10, 20 or 40 mg/mL of specific IgY or IgY from non-immunized hens (negative control). Data are presented as means ± SD (n = 3).
3.7. Application of specific IgY in a T. rubrum mouse model
Skin lesion scores after 14 days of treatment of 5 experimental groups were shown in Fig. 7 . In comparison with the model control (MC) group treated with sterile PBS (pH7.4, 0.01 M), scores of specific IgY groups decreased significantly and the treatment effect increased following the increase of dosages(P < 0.05,P < 0.05,P < 0.01)(Fig. 7).
Fig. 7.

Skin lesion scores in T. rubrum-infected BALB/C mice. After being infected, the mice were treated with PBS (Model control), 5 mg/mL, 10 mg/mL, or 20 mg/mL specific IgY or 1 mg/mL fluconazole (Positive control). Changes in redness, ulcerative scaling, and hair loss at the inoculation site were visually examined and recorded by using a lesion score. The cured, improved and aggravated were scored as 0, 1and 2 respectively. The skin lesion score of mice was the sum of the three scoring items. The data were presented as means ± SD (n = 4).*p < 0.05, **p < 0.01 in comparison with MC group.
Skin biopsies of infected areas of the experimental groups were subjected to histological analysis. In comparison with the normal untreated controls (Fig. 8 A), the MC group showed fungal elements, damage of epithelium, dermis granulomas, and infiltration of inflammatory cells into the dermis, indicating the successful infection (Fig. 8B). The fluconazole group showed no obvious inflammation or tissue destruction (Fig. 8C). Fewer inflammatory cells, weakened epithelium damage and hyperplastic tissue of cuticle were observed in the 5 mg/mL, 10 mg/mL, and 20 mg/mL specific IgY groups (Fig. 8D, E, and 8F). Dermis granulomas decreased when the concentration of specific IgY increases (Fig. 8). (Color should be used in print).
Fig. 8.

Histopathological examination of skin sections from T. rubrum-infected BALB/C mice. (A) Normal group. (B) Model control group. (C) Fluconazole group. (D–F) 5 mg/mL, 10 mg/mL, and 20 mg/mL anti-CWP IgY groups.
4. Discussion
Some studies have proved that specific IgYs were effective to bovine mastitis caused by E. coli and Staphylococcus aureus [[23], [24], [25]]. In those studies, inactivated strains were used as antigens to immunize hens. The studies showed that S. aureus specific IgYs could inhibit the growth of S. aureus and the internalization of S. aureus by bovine mammary epithelial cells, and could decrease the bacterial count in milk and cure experimental mastitis (83.3%) and clinical mastitis (50%). IgYs were observed ineffective to lyse Gram-positive bacteria, especially the encapsulated strain, but presented some inhibitory effect under growth conditions. E. coli specific IgYs could inhibit the growth of E. coli bybeing bound with it in a dose-dependent manner and enhance phagocytosis of E. coli by milk macrophages and polymorphonuclear neutrophil leukocytes.
There were also other studies about antifungal IgYs such as anti-C. albicans IgYs [14,26,27]. One of the studies evaluated in vitro and in vivo effectiveness of IgY against C. albicans by using proteins from sonicated cells of C. albicans as antigen to immunized hens. In the study, anti- C. albicans IgY significantly reduced the adherence capacity of C. albicans to human pharynx carcinoma cells. Oral administration of anti- C. albicans IgY by mice significantly reduced the number of C. albicans and the scores of tongue lesions; anti- C. albicans IgY also reduced colonization of C. albicans in mouse organs, indicating that anti- C. albicans IgY had a protective effect against the oral candidiasis of experimentally infected mice and reduced the dissemination of C. albicans [26].
In this study, we investigated the growth inhibition activity of anti-trCWP IgY to T. rubrum and the protective effect of anti-trCWP IgY against dermatophytosis of experimentally infected mice. We immunized the hens with cell wall proteins extracted from T. rubrum. The cell wall was considered to be the major part of the deposition of fungal immune response products. Many studies have shown that cell wall proteins have good immunogenicity and can induce specific antibodies [[28], [29], [30]]. In our study, the cell wall proteins showed good immunogenicity, highest titer of anti-trCWP IgY reaching 1:16000. The anti-trCWP IgY expressed significant inhibitory effect on T. rubrum in a dose-dependent manner compared with IgY from unimmunized hens,and significantly reduced lesion scores of mice experimentally infected with T. rubrum. Moreover, the anti-trCWP IgY reduced infiltration of inflammatory cells, epithelium damage and hyperplastic tissue of cuticle.
Cell wall proteins of pathogenic fungi are crucial for growth, virulence, and pathogenicity as well as important in mediating the host-fungus interaction. The cell wall provides both adhesive properties critical for the invasion of host tissue and protection against the host defense mechanisms [31]. The first step of dermatophytes to infect the host is to inoculate the germinating fungus into the skin, which adheres to the cuticle. The occlusion and impregnation of the cuticle accelerate the adhesion process. The fungal spores sprout after being adhered to the cuticle for a long time; the mycelia continue to spread, especially in the lower layer of the cuticle [32]. Anti-trCWP IgY may be directly bound to the proteins on the fungal cell wall, inhibit the proliferation of T. rubrum, reduce the adherence capacity of T. rubrum to host cells and prevent further invasion of the fungal hypha to the host.
Anti-trCWP IgY showed some degree of cross-reactivity with other fungi, most significantly with T. mentagrophytes, which indicated that the cell wall protein components of T. rubrum and T. mentagrophytes were quite similar. The anti-trCWP IgY may also be effective to other fungal pathogens of dermatophytosis. This result was similar to another study. In that study, anti- E. coli O111 IgY showed strong growth inhibition activity to both E. coli O111 and five other E. coli strains isolated from mastitis cows [23].
Owing to the effect on pathogenic microorganisms, IgYs have been widely added into foods to prevent microbial infection of animals or aquatic organisms and widely added to cosmetics to resist bacteria and viruses to protect skin. They have also been added to mouthwash and toothpaste to prevent oral infection, to yogurt or food to prevent and treat Helicobacter pylori and gastrointestinal virus infection. Anti-trCWP IgY can be used in a detergent to prevent dermatophytosis, and can be used in combination with antifungal chemicals to improve the therapeutic effect of dermatophytosis and reduce the generation of drug resistance.
5. Conclusion
In conclusion, we presented evidence for the activity of anti-trCWP IgY against T. rubrum. Anti-trCWP IgY showed a significant dose-dependent growth inhibitory effect on T. rubrum in vitro and a significant dose-dependent treatment effect on T. rubrum infection in BALB/C mice. These results suggested that anti-trCWP IgY could be used as preventive immunotherapy against T. rubrum.
Conflicts of interest
The authors have no conflict of interest to declare.
Acknowledgments
We thank Dr. Songchao Yin (The Third Affiliated Hospital of Sun Yat-sen University) for providing fungi strain and technical guidance.
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Footnotes
Supplementary data to this article can be found online at https://doi.org/10.1016/j.micpath.2019.103741.
Contributor Information
Yire Xiao, Email: 7952315@qq.com.
Qingqing Hu, Email: hqq2218@126.com.
Luoying Jiao, Email: jiaoluoying@163.com.
Xiping Cui, Email: cuixiping1989@163.com.
Panpan Wu, Email: wyuwpp@mail2.gdut.edu.cn.
Pan He, Email: 1076913953@qq.com.
Nana Xia, Email: nanaxia6947@163.com.
Rui Lv, Email: m13424004193@163.com.
Yuxin Liang, Email: lyx617022@126.com.
Suqing Zhao, Email: sqzhao@gdut.edu.cn.
Appendix A. Supplementary data
The following is the Supplementary data to this article:
References
- 1.Borman A.M., Campbell C.K., Fraser M., Johnson E.M. Analysis of the dermatophyte species isolated in the british isles between 1980 and 2005 and review of worldwide dermatophyte trends over the last three decades. Med. Mycol. 2007;45(2):131–141. doi: 10.1080/13693780601070107. [DOI] [PubMed] [Google Scholar]
- 2.Rosen T. Assessment of dermatophytosis treatment studies: interpreting the data. J. Drugs Dermatol. JDD. 2015;14(10):S48–S54. [PubMed] [Google Scholar]
- 3.Rochjadi A., Leuthold J.H. Dermatophytes: host-pathogen interaction and antifungal resistance. An. Bras. Dermatol. 2010;85(5):657–667. doi: 10.1590/s0365-05962010000500009. [DOI] [PubMed] [Google Scholar]
- 4.Ghannoum M., Isham N. Fungal nail infections (onychomycosis): a never-ending story? PLoS Pathog. 2014;10(6):e1004105. doi: 10.1371/journal.ppat.1004105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Aoun K., Bouratbine A., Mokni M., Chatti S., Ismail R.B., Osman A.B. Tinea capitis caused by trichophyton rubrum in two children with extensive dermatophytosis. J. De Mycol. Mã©Dicale/J. Med. Mycol. 1998;8(4):200–202. [Google Scholar]
- 6.Costa J.E., Neves R.P., Delgado M.M., Lima-Neto R.G., Morais V.M., Coêlho M.R. Dermatophytosis in patients with human immunodeficiency virus infection: clinical aspects and etiologic agents. Acta Trop. 2015;150:111–115. doi: 10.1016/j.actatropica.2015.07.012. [DOI] [PubMed] [Google Scholar]
- 7.Badali H., Mohammadi R., Mashedi O., de Hoog G.S., Meis J.F. In vitro susceptibility patterns of clinically important trichophyton and epidermophyton species against nine antifungal drugs. Mycoses. 2015;58(5):303–307. doi: 10.1111/myc.12315. [DOI] [PubMed] [Google Scholar]
- 8.Bhatia V.K., Sharma P.C. Determination of minimum inhibitory concentrations of itraconazole, terbinafine and ketoconazole against dermatophyte species by broth microdilution method. Indian J. Med. Microbiol. 2015;33(4):533. doi: 10.4103/0255-0857.167341. [DOI] [PubMed] [Google Scholar]
- 9.Koroishi A.M., Foss S.R., Cortez D.A.G., Ueda-Nakamura T., Nakamura C.V., Filho B.P.D. In vitro antifungal activity of extracts and neolignans from piper regnellii, against dermatophytes. J. Ethnopharmacol. 2008;117(2):270–277. doi: 10.1016/j.jep.2008.01.039. [DOI] [PubMed] [Google Scholar]
- 10.Ghannoum M.A., Hajjeh R.A., Scher R., Konnikov N., Gupta A.K., Summerbell R. A large-scale north american study of fungal isolates from nails: the frequency of onychomycosis, fungal distribution, and antifungal susceptibility patterns. J. Am. Acad. Dermatol. 2000;43(4):641–648. doi: 10.1067/mjd.2000.107754. [DOI] [PubMed] [Google Scholar]
- 11.Datta K., Pirofski L.A. Immunotherapy of fungal infections. Ref. Modul. Life Sci. 2017;44(8):738–776. doi: 10.3109/08820139.2015.1093913. [DOI] [PubMed] [Google Scholar]
- 12.Chalghoumi R., Beckers Y., Portetelle D., Théwis A. Hen egg yolk antibodies (igy), production and use for passive immunization against bacterial enteric infections in chicken: a review. Biotechnol. Agronomie Soc. Et Environ. 2009;13(2):295–308. [Google Scholar]
- 13.Ziglari T., Khosravi A.R., Tadjbdkhsh H., Asadi F., Modirsaneii M., Shokri H. Evaluation of the igy production against hsp90 of candida albicans. J. Mycol. Med. 2009;19(2):87–93. [Google Scholar]
- 14.Wang X.Z., Fan B., Liu L.G., Hu X.Y., Li R.Y., Wei Y. In vitro inhibition of oral candida albicans, by chicken egg yolk antibody (igy) Mycopathologia. 2008;165(6):381–387. doi: 10.1007/s11046-008-9097-0. [DOI] [PubMed] [Google Scholar]
- 15.Pitarch A., Sánchez M., Nombela C., Gil C. Sequential fractionation and two-dimensional gel analysis unravels the complexity of the dimorphic fungus candida albicans cell wall proteome. Mol. Cell. Proteom. 2002;1(12):967–982. doi: 10.1074/mcp.m200062-mcp200. [DOI] [PubMed] [Google Scholar]
- 16.Yang Y.E., Wen J., Zhao S., Zhang K., Zhou Y. Prophylaxis and therapy of pandemic h1n1 virus infection using egg yolk antibody. J. Virol Methods. 2014;206(7):19–26. doi: 10.1016/j.jviromet.2014.05.016. [DOI] [PubMed] [Google Scholar]
- 17.Wen J., Zhao S., He D., Yang Y., Li Y., Zhu S. Preparation and characterization of egg yolk immunoglobulin y specific to influenza b virus. Antivir. Res. 2012;93(1):154–159. doi: 10.1016/j.antiviral.2011.11.005. [DOI] [PubMed] [Google Scholar]
- 18.Zhai Z., Cheng L., Tang F., Lu Y., Shao J., Liu G. Immunoproteomic identification of 11 novel immunoreactive proteins of riemerella anatipestifer serotype 2. Pathog. Dis. 2012;65(1):84–95. doi: 10.1111/j.1574-695X.2012.00947.x. [DOI] [PubMed] [Google Scholar]
- 19.Shin J.H., Nam S.W., Kim J.T., Yoon J.B., Bang W.G., Roe I.H. Identification of immunodominant helicobacter pylori proteins with reactivity to h. pylori-specific egg-yolk immunoglobulin. J. Med. Microbiol. 2003;52(Pt 3):217–222. doi: 10.1099/jmm.0.04978-0. [DOI] [PubMed] [Google Scholar]
- 20.Beuchat L.R., Mann D.A. Comparison of new and traditional culture-dependent media for enumerating foodborne yeasts and molds. J. Food Prot. 2016;79(1):95. doi: 10.4315/0362-028X.JFP-15-357. [DOI] [PubMed] [Google Scholar]
- 21.Robledo-Leal E., Elizondo-Zertuche M., González G.M. Susceptibility of dermatophytes to thiabendazole using clsi broth macrodilution. Isrn Dermatol. 2012;2012(2):351842. doi: 10.5402/2012/351842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mei Y.X., Dai X.Y., Yang W., Xu X.W., Liang Y.X. Antifungal activity of chitooligosaccharides against the dermatophyte trichophyton rubrum. Int. J. Biol. Macromol. 2015;77:330–335. doi: 10.1016/j.ijbiomac.2015.03.042. [DOI] [PubMed] [Google Scholar]
- 23.Zhen Y.H., Jin L.J., Guo J., Li X.Y., Lu Y.N., Chen J. Characterization of specific egg yolk immunoglobulin (igy) against mastitis-causing escherichia coli. Vet. Microbiol. 2008;130(1):126–133. doi: 10.1016/j.vetmic.2007.12.014. [DOI] [PubMed] [Google Scholar]
- 24.Wang L.H., Li X.Y., Jin L.J., You J.S., Zhou Y., Li S.Y. Characterization of chicken egg yolk immunoglobulins (igys) specific for the most prevalent capsular serotypes of mastitis-causing staphylococcus aureus. Vet. Microbiol. 2011;149(3–4):415–421. doi: 10.1016/j.vetmic.2010.11.029. [DOI] [PubMed] [Google Scholar]
- 25.Zhen Y.H., Jin L.J., Li X.Y., Guo J., Li Z., Zhang B.J. Efficacy of specific egg yolk immunoglobulin (igy) to bovine mastitis caused by staphylococcus aureus. Vet. Microbiol. 2009;133(4):317–322. doi: 10.1016/j.vetmic.2008.07.016. [DOI] [PubMed] [Google Scholar]
- 26.El-Sm I., Rahman A.K., Isoda R., Umeda K., Van S.N., Kodama Y. In vitro and in vivo effectiveness of egg yolk antibody against candida albicans (anti-ca igy) Vaccine. 2008;26(17):2073–2080. doi: 10.1016/j.vaccine.2008.02.046. [DOI] [PubMed] [Google Scholar]
- 27.Kamikawa Y., Fujisaki J., Nagayama T., Kawasaki K., Hirabayashi D., Hamada T. Use of candida-specific chicken egg yolk antibodies to inhibit the adhering of candida to denture base materials: prevention of denture stomatitis. Gerodontology. 2016;33(3):342–347. doi: 10.1111/ger.12163. [DOI] [PubMed] [Google Scholar]
- 28.Holden C.A., Hay R.J., Macdonald D.M. A method for identification of dermatophyte antigens in situ by an immunoperoxidase technique in light and electron microscopy. Clin. Exp. Dermatol. 1981;6(3):311–316. doi: 10.1111/j.1365-2230.1981.tb02310.x. [DOI] [PubMed] [Google Scholar]
- 29.Mescon H., Grots I.A. Fluorescence microscopy in dermatology. J. Investig. Dermatol. 1963;41:181. doi: 10.1038/jid.1963.95. [DOI] [PubMed] [Google Scholar]
- 30.Hussin Z., Smith J.M.B. Vaccination procedures and the infectivity of dermatophyte lesions. Mycopathologia. 1983;81(2):71–76. doi: 10.1007/BF00436982. [DOI] [PubMed] [Google Scholar]
- 31.Sui X., Yan L., Jiang Y.Y. The vaccines and antibodies associated with als3p for treatment of candida albicans infections. Vaccine. 2017;35(43):5786. doi: 10.1016/j.vaccine.2017.08.082. [DOI] [PubMed] [Google Scholar]
- 32.Rashid A. Arthroconidia as vectors of dermatophytosis. Cutis. 2001;67(5 Suppl):23. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.