

Abstract
Flavonoids are ubiquitous in photosynthesising cells and are commonly found in fruit, vegetables, nuts, seeds, stems, flowers, tea, wine, propolis and honey. For centuries, preparations containing these compounds as the principal physiologically active constituents have been used to treat human diseases. Increasingly, this class of natural products is becoming the subject of anti-infective research, and many groups have isolated and identified the structures of flavonoids possessing antifungal, antiviral and antibacterial activity. Moreover, several groups have demonstrated synergy between active flavonoids as well as between flavonoids and existing chemotherapeutics. Reports of activity in the field of antibacterial flavonoid research are widely conflicting, probably owing to inter- and intra-assay variation in susceptibility testing. However, several high-quality investigations have examined the relationship between flavonoid structure and antibacterial activity and these are in close agreement. In addition, numerous research groups have sought to elucidate the antibacterial mechanisms of action of selected flavonoids. The activity of quercetin, for example, has been at least partially attributed to inhibition of DNA gyrase. It has also been proposed that sophoraflavone G and (−)-epigallocatechin gallate inhibit cytoplasmic membrane function, and that licochalcones A and C inhibit energy metabolism. Other flavonoids whose mechanisms of action have been investigated include robinetin, myricetin, apigenin, rutin, galangin, 2,4,2′-trihydroxy-5′-methylchalcone and lonchocarpol A. These compounds represent novel leads, and future studies may allow the development of a pharmacologically acceptable antimicrobial agent or class of agents.
Keywords: Flavonoids, Antifungal, Antiviral, Antibacterial, Structure–activity, Mechanism of action
1. Introduction
Resistance to antimicrobial agents has become an increasingly important and pressing global problem. Of the 2 million people who acquire bacterial infections in US hospitals each year, 70% of cases now involve strains that are resistant to at least one drug [1]. A major cause for concern in the UK is methicillin-resistant Staphylococcus aureus (MRSA), which was at low levels a decade ago but now accounts for ca. 50% of all S. aureus isolates [2]. Substantial investment and research in the field of anti-infectives are now desperately needed if a public health crisis is to be averted.
Structural modification of antimicrobial drugs to which resistance has developed has proven to be an effective means of extending the lifespan of antifungal agents such as the azoles [3], antiviral agents such as the non-nucleoside reverse transcriptase inhibitors [4], and various antibacterial agents including β-lactams and quinolones [5]. It is not surprising then, that in response to antimicrobial resistance, major pharmaceutical companies have tended to concentrate their efforts on improving antimicrobial agents in established classes [6]. However, with the portfolio of chemotherapeutics currently available, it has been acknowledged that researchers are getting close to the end game in terms of parent structure alterations. A call has therefore been made for the development of new classes of drug that work on different target sites to those in current use [7], [8].
Rational drug design does not always yield effective antimicrobials. In the past, potent enzyme inhibitors have been successfully designed and synthesised but they had only modest antibacterial activity, probably owing to the complex issue of drug uptake by cells. Broad empirical screening of chemical entities for antimicrobial activity represents an alternative strategy for the development of novel drugs. Natural products have been a particularly rich source of anti-infective agents, yielding, for example, the penicillins in 1940, the tetracyclines in 1948 and the glycopeptides in 1955 [9]. The following review will examine the antimicrobial activity of flavonoids, a class of natural products possessing a diverse range of pharmacological properties. Compounds with antifungal, antiviral and antibacterial activity will each be discussed in turn, with particular emphasis on those flavonoids with antibacterial activity.
2. Flavonoids: occurrence, functions, structure and nomenclature
Flavonoids are ubiquitous in photosynthesising cells and therefore occur widely in the plant kingdom [10]. They are found in fruit, vegetables, nuts, seeds, stems and flowers as well as tea, wine [11], propolis and honey [12], and represent a common constituent of the human diet [13]. In the US, the daily dietary intake of mixed flavonoids is estimated to be in the range 500–1000 mg, but this figure can be as high as several grams for people supplementing their diets with flavonoids or flavonoid-containing herbal preparations [14].
The function of flavonoids in flowers is to provide colours attractive to plant pollinators [11], [15]. In leaves, these compounds are increasingly believed to promote physiological survival of the plant, protecting it from, for example, fungal pathogens and UV-B radiation [13], [15]. In addition, flavonoids are involved in photosensitisation, energy transfer, the actions of plant growth hormones and growth regulators, control of respiration and photosynthesis, morphogenesis and sex determination [11], [13].
The basic structural feature of flavonoid compounds is the 2-phenyl-benzo[α]pyrane or flavane nucleus, which consists of two benzene rings (A and B) linked through a heterocyclic pyrane ring (C) (Fig. 1 ) [16]. Flavonoids can be classified according to biosynthetic origin. Some classes, for example chalcones, flavanones, flavan-3-ols and flavan-3,4-diols, are both intermediates in biosynthesis as well as end products that can accumulate in plant tissues. Other classes are only known as end products of biosynthesis, for example anthocyanidins, proanthocyanidins, flavones and flavonols. Two additional classes of flavonoid are those in which the 2-phenyl side chain of flavanone isomerises to the 3 position, giving rise to isoflavones and related isoflavonoids. The neoflavonoid is formed through further isomerisation to the 4 position [13]. Structures of the major classes of flavonoids are given in Fig. 2 . The structures of specific compounds within these classes that possess antimicrobial activity and that are discussed in the present review are summarised in Table 1 .
Fig. 1.
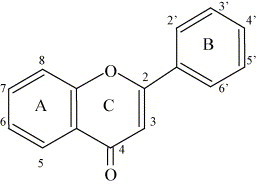
The skeleton structure of the flavones (a class of flavonoids), with rings named and positions numbered [13].
Fig. 2.
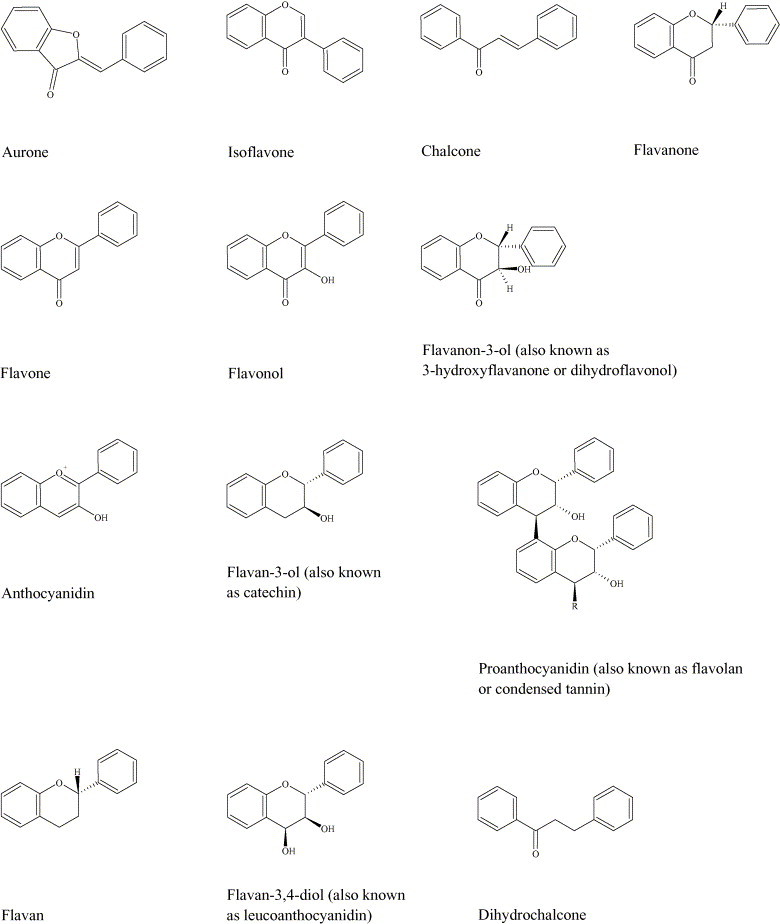
The skeleton structures of the main classes of flavonoids: aurones [17], [18], isoflavones [10], chalcones [13], [19], flavanones [10], [13], flavones [10], flavonols [10], flavanon-3-ols [13], anthocyanidins [13], [20], flavan-3-ols [10], [13], proanthocyanidins (occur as dimers, trimers, tetramers and pentamers; R = 0, 1, 2 or 3 flavan-3-ol structures) [13], flavans [13], flavan-3,4-diols [13] and dihydrochalcones [13].
Table 1.
A summary of the structures of antimicrobial flavonoids discussed within the present review article (compiled from The Handbook of Natural Flavonoids [13] and individual research papers)
| Compound | Substituents at carbon position: |
|||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 2′ | 3′ | 4′ | 5′ | 6′ | |
| Flavones and their glycosides | ||||||||||||
| Acacetin | – | – | – | OH | – | OH | – | – | – | OCH3 | – | – |
| Apigenin | – | – | – | OH | – | OH | – | – | – | OH | – | – |
| Baicalin | – | – | – | OH | OH | OR1 | – | – | – | – | – | – |
| Baicalein | – | – | – | OH | OH | OH | – | – | – | – | – | – |
| Chrysin | – | – | – | OH | – | OH | – | – | – | – | – | – |
| Gardenin A (demethylated) | – | – | – | OH | OH | OH | OH | – | OH | OH | OH | – |
| Genkwanin | – | – | – | OH | – | OCH3 | – | – | – | OH | – | – |
| Luteolin | – | – | – | OH | – | OH | – | – | OH | OH | – | – |
| Luteolin 7-glucoside | – | – | – | OH | – | OR2 | – | – | OH | OH | – | – |
| 7,8-Dihydroxyflavone | – | – | – | – | – | OH | OH | – | – | – | – | – |
| 5,5′-Dihydroxy-8,2′,4′-trimethoxyflavone | – | – | – | OH | – | – | OCH3 | OCH3 | – | OCH3 | OH | – |
| 5-Hydroxy-7,4′-dimethoxyflavone | – | – | – | OH | – | OCH3 | – | – | – | OCH3 | – | – |
| 5,7,4′-Trihydroxy-3′,5′-dimethoxyflavone | – | – | – | OH | – | OH | – | – | CH3 | OH | CH3 | – |
| 6,7,4′-Trihydroxy-3′,5′-dimethoxyflavone | – | – | – | – | OH | OH | – | – | CH3 | OH | CH3 | – |
| Isoflavones | ||||||||||||
| 6,8-Diprenylgenistein | – | – | – | OH | R3 | OH | R3 | – | – | OH | – | – |
| Sophoraisoflavone A | – | – | – | OH | – | OH | – | * | * | OH | – | – |
| Flavonols and their glycosides | ||||||||||||
| Galangin | – | OH | – | OH | – | OH | – | – | – | – | – | – |
| Kaempherol | – | OH | – | OH | – | OH | – | – | – | OH | – | – |
| 3-O-methylquercetin | – | OCH3 | – | OH | – | OH | – | – | OH | OH | – | – |
| Morin | – | OH | – | – | – | OH | – | OH | – | OH | OH | – |
| Myricetin | – | OH | – | OH | – | OH | – | – | OH | OH | OH | – |
| Quercetagetin | – | OH | – | OH | OH | OH | – | – | OH | OH | – | – |
| Quercetagetin-7-arabinosyl-galactoside | – | OH | – | OH | OH | OR4 | – | – | OH | OH | – | – |
| Quercetin | – | OH | – | OH | – | OH | – | – | OH | OH | – | – |
| Quercetin-3-O-(2″-galloyl)-α-l-arabinopyranoside | – | OR5 | – | OH | – | OH | – | – | OH | OH | – | – |
| Quercetrin | – | OR6 | – | OH | – | OH | – | – | OH | OH | – | – |
| Robinetin | – | OH | – | – | – | OH | – | – | OH | OH | OH | – |
| Rutin | – | OR7 | – | OH | – | OH | – | – | OH | OH | – | – |
| 3,2′-Dihydroxyflavone | – | OH | – | – | – | – | – | OH | – | – | – | – |
| 3,6,7,3′,4′-Pentahydroxyflavone | – | OH | – | – | OH | OH | – | – | OH | OH | – | – |
| Flavan-3-ols | ||||||||||||
| Catechin | – | OH | OH | – | – | OH | – | – | OH | – | OH | – |
| Epicatechin gallate | – | R8 | – | OH | – | OH | – | – | OH | OH | – | – |
| Epigallocatechin | – | OH | – | OH | – | OH | – | – | OH | OH | OH | – |
| Epigallocatechin gallate | – | R8 | – | OH | – | OH | – | – | OH | OH | OH | – |
| 3-O-octanoyl-(+)-catechin | – | R9 | – | OH | – | OH | – | – | OH | OH | – | – |
| 3-O-octanoyl-(−)-epicatechin | – | R9 | – | OH | – | OH | – | – | OH | OH | – | – |
| Flavanon-3-ols | ||||||||||||
| Dihydrofisetin | – | OH | – | – | – | OH | – | – | OH | OH | – | – |
| Dihydroquercetin | – | OH | – | OH | – | OH | – | – | OH | OH | – | – |
| Flavanones and their glycosides | ||||||||||||
| Lonchocarpol A | – | – | – | OH | R3 | OH | R3 | – | – | OH | – | – |
| Naringenin | – | – | – | OH | – | OH | – | – | – | OH | – | – |
| Naringin | – | – | – | OH | – | OR7 | – | – | – | OH | – | – |
| Pinocembrin | – | – | – | OH | – | OH | – | – | – | – | – | – |
| Ponciretin | – | – | – | OH | – | OH | – | – | – | OCH3 | – | – |
| Sophoraflavanone G | – | – | – | OH | – | OH | R10 | OH | – | OH | – | – |
| 3-Methyleneflavanone | – | CH2 | – | – | – | – | – | – | – | – | – | – |
| 5,7,4′-Trihydroxy-8-methyl-6-(3-methyl-[2-butenyl])-(2S)-flavanone | – | – | – | OH | R3 | OH | CH3 | – | – | OH | – | – |
| Chalcones | ||||||||||||
| Licochalcone A | – | R11 | OH | – | OCH3 | – | – | – | – | OH | – | – |
| Licochalcone C | – | – | OH | R3 | OCH3 | – | – | – | – | OH | – | – |
| 2,4,2′-Trihydroxychalcone | OH | – | OH | – | – | – | – | OH | – | – | – | – |
| 2,4,2′-Trihydroxy-5′-methylchalcone | OH | – | OH | CH3 | – | – | – | OH | – | – | – | – |
| Flavan-3,4-diols and anthocyanidins | ||||||||||||
| Leucocyanidin | – | OH | OH | OH | – | OH | – | – | OH | OH | – | – |
| Pelargonidin chloride | – | Cl | – | OH | – | OH | – | – | – | OH | – | – |
| Flavans | ||||||||||||
| 6,4′-Dichloroflavan | – | – | – | – | Cl | – | – | – | – | Cl | – | – |
| 7-Hydroxy-3′,4′-(methylenedioxy)flavan | – | – | – | – | – | OH | – | – | # | # | – | – |
R1: Glucuronic acid; R2: glucose; R3: prenyl group; R4: arabinose–galactose; R5: (2″-galloyl)-α-l-arabinopyranoside; R6: rhamnose; R7: rhamnose–glucose; R8: gallic acid; R9: octanoyl; R10: lavandulyl; R11: 3-methyl-1-butene.
–, no substitution; *, pyran ring between positions 2′ and 3′; #, O-CH2-O between positions 3′ and 4′.
Note: Hinokiflavone and robustaflavone are biflavonoids (also known as biflavonyls) consisting of two apigenin molecules joined through I-6-O-II-4′ and I-6-II-3′ linkages, respectively.
Individual flavonoids may be assigned names in three different ways. Trivial names are employed extensively and sometimes indicate flavonoid class or plant source. For example, names ending in ‘inidin’ can denote an anthocyanidin, names ending in ‘etin’ generally denote a flavonol, and compounds tricin and hypolaetin have been extracted from plants belonging to the genera Triticum and Hypolaena. Flavonoids may also be named in a semi-systematic manner based on trivial names such as flavone or chalcone as the parent structure, e.g. 3,5,7,3′4′-pentahydroxyflavone or 3,3′,4′,5,7-pentahydroxyflavone. Lastly, flavonoids may be given systematic chemical names, e.g. 3,4-dihydro-2-phenyl-2H-1-benzopyran for flavan, but this method is cumbersome and rarely used [13]. In the present review, trivial names will be used wherever possible.
3. Medicinal properties of flavonoids
Increasingly, flavonoids are becoming the subject of medical research. They have been reported to possess many useful properties, including anti-inflammatory activity, oestrogenic activity, enzyme inhibition, antimicrobial activity [10], [13], antiallergic activity, antioxidant activity [11], vascular activity and cytotoxic antitumour activity [15]. For a group of compounds of relatively homogeneous structure, the flavonoids inhibit a perplexing number and variety of eukaryotic enzymes and have a tremendously wide range of activities. In the case of enzyme inhibition, this has been postulated to be due to the interaction of enzymes with different parts of the flavonoid molecule, e.g. carbohydrate, phenyl ring, phenol and benzopyrone ring [10]. Several reviews have been written on the interaction between flavonoids and mammalian cells, including comprehensive articles by Harborne and Williams [15] and Middleton et al. [20]. An extensive review on the biochemistry and medical significance of flavonoids has also recently been produced by Havsteen [21].
4. History of flavonoid use in antimicrobial treatment
For centuries, preparations that contain flavonoids as the principal physiologically active constituents have been used by physicians and lay healers in attempts to treat human diseases [10]. For example, the plant Tagetes minuta (containing quercetagetin-7-arabinosyl-galactoside) has been used extensively in Argentine folk medicine to treat infectious disease [22]. The healing properties of propolis (or ‘tzori’ in Hebrew) are referred to throughout the Old Testament [23], and this balm was prescribed by Hippocrates (460–377 BC) in Ancient Greece for the treatment of sores and ulcers [24]. The antimicrobial properties of propolis have been attributed to its high flavonoid content and in particular the presence of the flavonoids galangin and pinocembrin [12], [25], [26], [27]. Huang-chin (Scutellaria baicalensis) is yet another example. This herbal medicine has been used systemically and topically for thousands of years in China for the treatment of periodontal abscesses and infected oral wounds. The flavone baicalein is reported to be largely responsible for this plant's antimicrobial effects [28].
5. Toxicity of flavonoids
It has been suggested that because flavonoids are widely distributed in edible plants and beverages and have previously been used in traditional medicine, they are likely to have minimal toxicity. However, this family of compounds has a diverse range of activities in mammalian cells [14], [20] and in vivo confirmation of their side effects would be necessary for a full evaluation of their practical usefulness in the field of modern medicine [29]. Given that the selectivity of flavonoids for eukaryotic enzymes appears to vary from compound to compound [15], [20], such a study would need to assess the toxicity of these phytochemicals on an individual basis.
6. Antifungal activity of flavonoids
Owing to the widespread ability of flavonoids to inhibit spore germination of plant pathogens, they have been proposed for use against fungal pathogens of man [15]. A new prenylated flavanone recently isolated from the shrub Eysenhardtia texana has been identified as 5,7,4′-trihydroxy-8-methyl-6-(3-methyl-[2-butenyl])-(2S)-flavanone and shown to possess activity against the opportunistic pathogen Candida albicans [30]. The flavonoid 7-hydroxy-3′,4′-(methylenedioxy)flavan, isolated from Terminalia bellerica fruit rind, has also been shown to possess activity against C. albicans [31]. Two new flavones from Artemisia giraldi, identified as 6,7,4′-trihydroxy-3′,5′-dimethoxyflavone and 5,5′-dihydroxy-8,2′,4′-trimethoxyflavone, together with 5,7,4′-trihydroxy-3′,5′-dimethoxyflavone have been reported to exhibit activity against Aspergillus flavus [32], a species of fungi that causes invasive disease in immunosuppressed patients [33]. The activity of propolis against dermatophytes and Candida spp. has been attributed at least partially to its high flavonoid content [34]. Galangin, a flavonol commonly found in propolis samples [24], has been shown to have inhibitory activity against Aspergillus tamarii, A. flavus, Cladosporium sphaerospermum, Penicillium digitatum and Penicillium italicum [35].
7. Antiviral activity of flavonoids
A recent area of research that is of particular interest is the apparent inhibitory activity of some flavonoids against human immunodeficiency virus (HIV). To date, most if not all investigations have involved work with the pandemic HIV-1 strain and its enzymes. In vitro studies have shown that baicalin inhibits HIV-1 infection and replication. Inhibition of HIV-1 entry into cells expressing CD4 and chemokine co-receptors [36], and antagonism of HIV-1 reverse transcriptase [37] by the flavone O-glycoside have been demonstrated by Li and colleagues. Baicalein [38], robustaflavone and hinokiflavone [39] have also been shown to inhibit HIV-1 reverse transcriptase, as have several catechins, but catechins inhibit other DNA polymerases and their interaction with the HIV-1 enzyme is therefore thought to be non-specific in nature [40]. In addition, it has been demonstrated that several flavonoids, including demethylated gardenin A and 3,2′-dihydroxyflavone, inhibit HIV-1 proteinase [41]. Robinetin, myricetin, baicalein, quercetagetin [42] and quercetin 3-O-(2″-galloyl)-α-l-arabinopyranoside [43] inhibit HIV-1 integrase, although there are concerns that HIV enzyme inhibition by quercetagetin and myricetin is non-specific [44]. It has also been reported that the flavonoids chrysin, acacetin and apigenin prevent HIV-1 activation via a novel mechanism that probably involves inhibition of viral transcription [45]. Interestingly, in a study by Hu and colleagues, chrysin was reported to have the highest therapeutic index of 21 natural and 13 synthetic flavonoids against HIV-1 [46]. Several research groups have investigated the relationship between flavonoid structure and inhibitory activity against HIV-1 and its enzymes [39], [41], [42], [44], [46]. Furthermore, at least two groups have proposed mechanisms of action for HIV-1 enzyme inhibition [41], [42].
Flavonoids also have inhibitory activity against a variety of other viruses. For example, Selway reports that quercetin, morin, rutin, dihydroquercetin, dihydrofisetin, leucocyanidin, pelargonidin chloride and catechin possess activity against up to seven types of virus, including herpes simplex virus (HSV), respiratory syncytial virus, poliovirus and Sindbis virus [11], [47]. Proposed antiviral mechanisms of action include inhibition of viral polymerase and binding of viral nucleic acid or viral capsid proteins [47]. In addition to the flavonoids mentioned above, three proanthocyanidins from Pavetta owariensis (with structural similarity to proanthocyanidin A2 and cinnamtannin B1 and B2) have been shown to have activity against HSV and coxsackie B virus [48], [49]. It has also been demonstrated that two of the flavonoids found in propolis, chrysin and kaempferol, inhibit viral replication of HSV, human coronavirus and rotavirus [50]. More recently, the flavonol galangin has been reported to have significant antiviral activity against HSV and coxsackie B virus [51].
Although naturally occurring flavonoids with antiviral activity have been recognised since the 1940s, it is only in the last 25 years that attempts have been made to synthetically modify flavonoids for improved antiviral activity. One such synthesised compound is 6,4′-dichloroflavan. However, despite showing strong in vitro activity, this compound proved unsuccessful in clinical trials [11].
Synergism has been demonstrated between various combinations of flavones and flavonols. For example, kaempferol and luteolin show synergy against HSV. It has been suggested that this is why propolis is more active than its individual component compounds [52]. Synergism has also been reported between flavonoids and other antiviral agents. Quercetin, for example, potentiates the effects of 5-ethyl-2′-dioxyuridine [11] and acyclovir [53] against HSV and pseudorabies infection. Apigenin also enhances the antiviral activity of acyclovir against these viruses [53].
8. Antibacterial activity of flavonoids
8.1. Reports of flavonoids possessing antibacterial activity
The antibacterial activity of flavonoids is being increasingly documented. Crude extracts from plants with a history of use in folk medicine have been screened in vitro for antibacterial activity by many research groups. Flavonoid-rich plant extracts from species of Hypericum [54], Capsella [55] and Chromolaena [55] have been reported to possess antibacterial activity. Many other phytochemical preparations with high flavonoid content have also been reported to exhibit antibacterial activity [22], [56], [57], [58], [59], [60], [61], [62], [63]. Propolis has been analysed on many occasions too, and samples containing high concentrations of flavonoids are frequently reported to show antibacterial activity [12], [25], [26], [27], [50], [64].
Many research groups have gone one step further and either isolated and identified the structure of flavonoids that possess antibacterial activity, or quantified the activity of commercially available flavonoids. Examples of such flavonoids are apigenin [65], [66], [67], [68], [69], [70], [71], [72], [73], galangin [35], [74], [75], [76], [77], pinocembrin [78], [79], ponciretin [80], [81], genkwanin [66], [82], sophoraflavanone G and its derivatives [29], [83], [84], [85], naringin and naringenin [29], [60], [86], [87], epigallocatechin gallate and its derivatives [74], [88], [89], [90], [91], [92], [93], [94], [95], luteolin and luteolin 7-glucoside [69], [73], [96], quercetin, 3-O-methylquercetin and various quercetin glycosides [60], [65], [72], [87], [97], [98], [99], [100], [101], [102] and kaempferol and its derivatives [60], [65], [74], [76], [87], [98], [100], [103]. Other flavones [32], [60], [74], [104], [105], [106], [107], flavone glycosides [86], [108], [109], isoflavones [110], [111], flavanones [29], [30], [78], [79], [104], [111], [112], [113], [114], isoflavanones [115], isoflavans [116], flavonols [74], [114], [117], flavonol glycosides [86], [118], [119], [120] and chalcones [79], [104], [111], [121] with antibacterial activity have also been identified.
Some researchers have reported synergy between naturally occurring flavonoids and other antibacterial agents against resistant strains of bacteria. Examples of these include epicatechin gallate [122], [123], [124], [125] and sophoraflavanone G [83], [84]. At least one group has demonstrated synergy between flavonoids with antibacterial activity [126]. Others have synthetically modified natural flavones and analysed them for antibacterial activity [94], [127], [128], [129], [130], [131]. For example, Wang and colleagues have complexed 5-hydroxy-7,4′-dimethoxyflavone with a number of transition metals and shown that this process increases antibacterial activity [130]. Another group reported increased antibacterial activity of 3-methyleneflavanones when the B ring contained bromine or chlorine substituents [131]. Two research groups have described the use of flavonoids in vivo. In a study by Vijaya and Ananthan, oral administration of either 142.9 mg/kg quercetin or 214.3 mg/kg quercetrin protected guinea pigs against an induced Shigella infection that killed untreated control animals [132]. More recently, Dastidar and co-workers reported that intraperitoneal injection of either 1.58 mg/kg sophoraisoflavone A or 3.16 mg/kg 6,8-diprenylgenistein gave significant protection to mice challenged with ∼9.5 × 108 colony-forming units (CFUs) of Salmonella typhimurium [110].
8.2. Discrepancies between reports of flavonoid antibacterial activity
When reports of the antibacterial activity of flavonoids are compared, the results appear widely conflicting (Table 2 ). For example, it was published that apigenin had no activity against S. aureus at concentrations up to 128 μg/mL [72]; a separate study in the same year reported that the flavone inhibited the growth of 15 strains of MRSA and 5 sensitive strains of S. aureus at concentrations between 3.9 μg/mL and 15.6 μg/mL [73]. From Table 2 it can be seen that such discrepancies could perhaps be attributed on occasion to different assays being used (e.g. [65], [70] and [72], [73]). Many different assays are employed in flavonoid research, including the agar dilution technique [29], the paper disk diffusion assay [115], the hole-plate diffusion method [22], the cylinder diffusion method [60], the broth macrodilution technique [71] and the broth microdilution technique [134]. In particular, assays relying on diffusion of test flavonoids may not give a reliable quantitative measure of antibacterial activity because a potent antibacterial flavonoid may have a low rate of diffusion [32]. However, it is clear from Table 2 that additional factors are involved in causing these discrepancies because even groups using the same assay are obtaining conflicting results (e.g. [67], [96] and [67], [72]). Such inconsistencies may be due to variations within each assay. For example, different groups using the agar dilution technique have used different sizes of bacterial inoculum [81], [86]. In a report by the National Committee for Clinical Laboratory Standards (NCCLS), inoculum size was considered the single most important variable in susceptibility testing [135]. It should be noted that many groups assaying flavonoid antibacterial activity have not quantified the test bacterial suspension [60], [115] and others have not even standardised the size of their unenumerated inocula [35], [56], [76], [90], [97]. From the published work it is clear that, in addition to inoculum size, there are many other variable factors for each type of assay. These include volume of broth or agar [90], [116], type of broth or agar [86], [92], size of wells [56], [60], size of paper disks [57], [65], strains of a particular bacterial species used [69], [72] and incubation period [90], [116]. Recently, a set of guidelines was published for standard agar dilution, broth macrodilution and broth microdilution methods [136]. This may help to reduce the number of conflicting reports of flavonoid antibacterial activity in the future. However, it will remain necessary to consider carefully additional variables such as the solvent used to dissolve test flavonoids [116], [118]. It has previously been shown that precipitation occurs when selected flavonoids are dissolved in organic solvents and diluted with neutral polar solutions [75]. Precipitation of flavonoids in a minimum inhibitory concentration (MIC) assay is likely to cause diminished contact between bacterial cells and flavonoid molecules and may lead to false negative reports of antibacterial activity. Also, in improperly controlled experiments, flavonoid precipitation could be misinterpreted as bacterial growth and further false negative results may be recorded as a consequence. The structural alteration of flavonoids such as galangin in alkaline solvents is another matter for consideration [75]. If flavonoid salts are formed and these have increased or decreased potency compared with the parent structure, this may lead to false positive/negative reports of antibacterial activity. Other variables worth noting include whether the test flavonoids are obtained from a commercial or natural source [35], [74] and which companies [74], [75]/natural products [71], [72] the compounds are from.
Table 2.
The inhibitory activity of apigenin against numerous species of Gram-positive and Gram-negative bacteria, as determined by various research groups between 1980 and 2000
| Staphylococcus aureus | MRSA | Staphylococcus albus | Staphylococcus epidermidis | Enterococcus faecalis | Bacillus subtilis | Micrococcus luteus | Escherichia coli | Pseudomonas aeruginosa | Proteus vulgaris | Proteus mirabilis | Klebsiella pneumoniae | Salmonella typhimurium | Enterobacter aerogenes | Enterobacter cloacae | Stenotrophomonas maltophilia | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Khanna et al. (DD) [65] | + | + | − | |||||||||||||
| Miski et al. (DD) [96] | − | − | − | − | − | − | ||||||||||
| Palacios et al. (DD) [66] | − | + | + | |||||||||||||
| Oksuz et al. (DD) [67] | − | − | + | + | + | + | + | |||||||||
| Oksuz et al. (BMAD) [67] | + | + | + | + | + | |||||||||||
| Ohemeng et al. (BMID) [68] | − | + | − | − | − | |||||||||||
| Bashir et al. (NS) [69], [133] | + | + | + | + | ||||||||||||
| Aljancic et al. (AWD) [70] | − | + | + | |||||||||||||
| Basile et al. (BMAD) [71], [72] | − | + | + | − | + | + | + | + | + | |||||||
| Sato et al. (AD) [73] | + | + |
DD, disk diffusion assay; BMAD, broth macrodilution assay; BMID, broth microdilution assay; NS, assay type not stated in report; AWD, agar well diffusion assay; AD, agar dilution assay; +, antibacterial activity detected; −, no antibacterial activity detected.
8.3. Structure–activity relationship for antibacterial activity of flavonoids
The diverse range of cell functions affected by flavonoids in eukaryotic systems is well documented [10], [20]. Although there have been comparatively few studies into the mechanisms underlying flavonoid antibacterial activity, information from published literature indicates that different compounds within this class of phytochemicals may target different components and functions of the bacterial cell [137], [138], [139]. If this is the case, it is surprising that the small number of groups which have investigated the relationship between flavonoid structure and antibacterial activity (summarised below) have been able to identify common structural features among active compounds. However, it may be that individual antibacterial flavonoids have multiple cellular targets, rather than one specific site of action. Alternatively, these common structural features may simply be necessary for flavonoids to gain proximity to or uptake into the bacterial cell.
Tsuchiya and colleagues sought to establish a structure–activity relationship for flavanones by isolating a number of differently substituted compounds and determining their MICs against MRSA [29]. Their study indicated that 2′,4′- or 2′,6′-dihydroxylation of the B ring and 5,7-dihydroxylation of the A ring in the flavanone structure was important for anti-MRSA activity. Substitution at the 6 or 8 position with a long chain aliphatic group such as lavandulyl (5-methyl-2-isopropenyl-hex-4-enyl) or geranyl (trans-3,7-dimethyl-2,6-octadienyl) also enhanced activity [29]. Interestingly, a recent report by Stapleton and colleagues demonstrated that substitution with C8 and C10 chains also enhanced the anti-staphylococcal activity of flavonoids belonging to the flavan-3-ol class [94].
Osawa et al. assessed the activity of a number of structurally different flavonoids including flavones, flavanones, isoflavones and isoflavanones based on the paper disk agar diffusion assay [115]. It was shown that 5-hydroxyflavanones and 5-hydroxyisoflavanones with one, two or three additional hydroxyl groups at the 7, 2′ and 4′ positions inhibited the growth of Streptococcus mutans and Streptococcus sobrinus. These results correlate well with those of Tsuchiya and colleagues [29]. It was also reported by Osawa and colleagues that 5-hydroxyflavones and 5-hydroxyisoflavones with additional hydroxyl groups at the 7 and 4′ positions did not exhibit this inhibitory activity [115]. However, when Sato et al. examined two isoflavones with hydroxyl groups at the 5, 2′ and 4′ positions using an agar dilution assay, intensive inhibitory activity was detected against a wide range of streptococcal species [107]. This may suggest that hydroxylation at position 2′ is important for activity. Alternatively, the lack of activity detected by Osawa et al. may simply be due to the poor diffusion of flavones and isoflavones (compared with flavanones and isoflavanones) through the medium.
A more recent paper [104] also reports the importance of a hydroxyl group at position 5 of flavanones and flavones for activity against MRSA, supporting the earlier findings of Tsuchiya et al. [29]. It further states that chalcones are more effective against MRSA than flavanones or flavones, and that hydroxyl groups at the 2′ position are important for the anti-staphylococcal activity of these compounds. Methoxy groups were reported to drastically decrease the antibacterial activity of flavonoids [104]. The importance of hydroxylation at the 2′ position for antibacterial activity of chalcones is supported by earlier work from Sato and colleagues, who found that 2,4,2′-trihydroxy-5′-methylchalcone and 2,4,2′-trihydroxychalcone inhibited the growth of 15 strains of cariogenic streptococci [140].
As mentioned previously, Ward and colleagues synthesised a number of halogenated derivatives of 3-methyleneflavanone [131]. Substitution of the B ring was found to enhance antibacterial activity, with 3′-chloro, 4′-chloro and 4′-bromo analogues each being approximately twice as effective as their parent compound against S. aureus, and four times more active against Enterococcus faecalis. Also, the 2′,4′-dichloro derivative exhibited a four- to eight-fold improvement in activity against S. aureus and a two- to four-fold improvement against E. faecalis. By contrast, 3-methylene-6-bromoflavanone was less potent than the parent compound and the authors suggested that halogenation of the A ring may diminish activity [131]. Clearly, however, it would be necessary to prepare analogues with substitution at other A-ring positions before this could be said with any certainty. In chalcones, neither fluorination nor chlorination at position 4 of the B ring is reported to affect antibacterial potency significantly [104]. Again, however, other structural analogues of this class of flavonoids would need to be synthesised and examined before the effect of halogenation upon antibacterial activity could be properly assessed.
8.4. Nature of flavonoid activity: bacteriostatic or bactericidal?
Several research groups have attempted to determine whether flavonoid activity is bacteriostatic or bactericidal by conducting time–kill studies. In such experiments, epigallocatechin gallate [89], galangin [75] and 3-O-octanoyl-(+)-catechin [94] have been shown to cause a reduction of 1000-fold or more in viable counts of MRSA-YK, S. aureus NCTC 6571 and EMRSA-16, respectively. This would immediately appear to suggest that flavonoids are capable of bactericidal activity. However, it has recently been demonstrated that 3-O-octanoyl-(−)-epicatechin induces the formation of pseudomulticellular aggregates both in antibiotic-sensitive and antibiotic-resistant strains of S. aureus [94]. If this phenomenon is induced by other compounds within the flavonoid class (and liposomal studies suggest that this is the case for epigallocatechin gallate [88]), questions are raised regarding the interpretation of results from time–kill studies. It may be that flavonoids are not killing bacterial cells but merely inducing the formation of bacterial aggregates and thereby reducing the number of CFUs in viable counts.
8.5. Antibacterial mechanisms of action of various flavonoids
8.5.1. Inhibition of nucleic acid synthesis
In a study using radioactive precursors, Mori and colleagues showed that DNA synthesis was strongly inhibited by flavonoids in Proteus vulgaris, whilst RNA synthesis was most affected in S. aureus [138]. Flavonoids exhibiting this activity were robinetin, myricetin and (−)-epigallocatechin. Protein and lipid synthesis were also affected but to a lesser extent. The authors suggested that the B ring of the flavonoids may play a role in intercalation or hydrogen bonding with the stacking of nucleic acid bases and that this may explain the inhibitory action on DNA and RNA synthesis [138].
Ohemeng et al. screened 14 flavonoids of varying structure for inhibitory activity against Escherichia coli DNA gyrase, and for antibacterial activity against Staphylococcus epidermidis, S. aureus, E. coli, S. typhimurium and Stenotrophomonas maltophilia [68]. It was found that E. coli DNA gyrase was inhibited to different extents by seven of the compounds, including quercetin, apigenin and 3,6,7,3′,4′-pentahydroxyflavone. Interestingly, with the exception of 7,8-dihydroxyflavone, enzyme inhibition was limited to those compounds with B-ring hydroxylation [68], [141]. The authors proposed that the observed antibacterial activity of the seven flavonoids was due in part to their inhibition of DNA gyrase. However, since the level of antibacterial activity and enzyme inhibition did not always correlate, they also suggested that other mechanisms were involved [68].
More recently, Plaper and colleagues reported that quercetin binds to the GyrB subunit of E. coli DNA gyrase and inhibits the enzyme's ATPase activity [142]. Enzyme binding was demonstrated by isolating E. coli DNA gyrase and measuring quercetin fluorescence in the presence and absence of the gyrase subunits. The flavonoid-binding site was postulated to overlap with those of ATP and novobiocin, since addition of these compounds interfered with quercetin fluorescence. Inhibition of GyrB ATPase activity by quercetin was also demonstrated in a coupled ATPase assay. This research is in agreement with the earlier findings of Ohemeng et al. [68] and supports the suggestion that quercetin's antibacterial activity against E. coli may be at least partially attributable to inhibition of DNA gyrase.
When screening natural products for type II topoisomerase inhibitors, Bernard and co-workers found that the glycosylated flavonol rutin was very effective [143]. This compound exhibited antibacterial activity against a permeable E. coli strain (a strain into which the envA1 allele had been incorporated [144], [145]). Using enzyme assays and a technique known as the SOS chromotest, it was shown that rutin selectively promoted E. coli topoisomerase IV-dependent DNA cleavage, inhibited topoisomerase IV-dependent decatenation activity and induced the SOS response of the E. coli strain. The group suggested that since topoisomerase IV is essential for cell survival, the rutin-induced topoisomerase IV-mediated DNA cleavage leads to an SOS response and growth inhibition of E. coli cells [143].
Within our own laboratory, a 4-quinolone-resistant S. aureus strain was shown to have increased susceptibility to the flavonol galangin compared with other 4-quinolone-sensitive and -resistant strains [146]. Interestingly, this strain possesses a distinct amino acid substitution (serine to proline) at position 410 of the GrlB subunit. This may suggest that topoisomerase IV and the relatively homologous gyrase enzyme are involved in the antibacterial mechanism of action of galangin. Clearly, however, further work with mutant strains and purified enzymes would be necessary before this could be verified.
8.5.2. Inhibition of cytoplasmic membrane function
A research team that had previously found sophoraflavanone G to have intensive antibacterial activity against MRSA and streptococci [83], [84], [85] recently reported attempts to elucidate the mechanism of action of this flavanone [139]. The effect of sophoraflavanone G on membrane fluidity was studied using liposomal model membranes and compared with the less active flavanone naringenin, which lacks 8-lavandulyl and 2′-hydroxyl groups. At concentrations corresponding to the MIC values, sophoraflavanone G was shown to increase fluorescence polarisation of the liposomes significantly. These increases indicated an alteration of membrane fluidity in hydrophilic and hydrophobic regions, suggesting that sophoraflavanone G reduced the fluidity of outer and inner layers of membranes. Naringenin also exhibited a membrane effect but at much higher concentrations. This correlation between antibacterial activity and membrane interference was suggested to support the theory that sophoraflavanone G demonstrates antibacterial activity by reducing membrane fluidity of bacterial cells [139].
Another group, Ikigai and colleagues, carried out research on (−)-epigallocatechin gallate, a strongly antibacterial catechin found in green tea. Catechins are a group of flavonoids that appear to have greater activity against Gram-positive than Gram-negative bacteria [88]. In this study, liposomes were again used as model bacterial membranes, and it was shown that epigallocatechin gallate induced leakage of small molecules from the intraliposomal space. Aggregation was also noted in liposomes treated with the compound. The group therefore concluded that catechins primarily act on and damage bacterial membranes. It was not known how this damage occurred but two theories were put forward. First, catechins may perturb the lipid bilayers by directly penetrating them and disrupting the barrier function. Alternatively, catechins may cause membrane fusion, a process that results in leakage of intramembranous materials and aggregation. Interestingly, the group also demonstrated that leakage induced by epigallocatechin gallate was significantly lower when liposome membranes were prepared containing negatively charged lipids. It was therefore suggested that the low catechin susceptibility of Gram-negative bacteria may be at least partially attributable to the presence of lipopolysaccharide acting as a barrier [88].
As mentioned previously, Stapleton and colleagues found that substitution with C8 and C10 chains increased the antibacterial activity of selected flavan-3-ols (catechins). The group went on to show that cells of an MRSA clinical isolate treated with (−)-epicatechin gallate and 3-O-octanoyl-(+)-catechin, respectively, exhibited moderately and highly increased levels of labelling with the selectively permeable fluorescent stain propidium iodide. In addition, when S. aureus cells were grown in the presence of either (−)-epicatechin gallate or 3-O-octanoyl-(−)-epicatechin and examined by transmission electron microscopy, they were shown to form pseudomulticellular aggregates [94]. This work constitutes a substantial advance in the development of catechins as antibacterial agents and lends support to Ikigai's argument that catechins act on and damage bacterial membranes.
It has also been demonstrated by Sato and colleagues that the chalcone 2,4,2′-trihydroxy-5′-methylchalcone induces leakage of 260 nm absorbing substances from S. mutans. This observation generally indicates leakage of intracellular material such as nucleotide, and the authors suggested that 2,4,2′-trihydroxy-5′-methylchalcone exerts its antibacterial effect by changing the permeability of the cellular membrane and damaging membrane function [140].
In addition, the effect of galangin upon cytoplasmic integrity in S. aureus has been investigated by measuring loss of internal potassium [147]. When high cell densities of S. aureus were incubated for 12 h in media containing 50 μg/mL of the flavonol, a 60-fold decrease in the number of CFUs was noted and cells lost ca. 20% more potassium than untreated control bacteria. These data strongly suggest that galangin induces cytoplasmic membrane damage and potassium leakage. Whether galangin damages the membrane directly, or indirectly as a result of autolysis or cell wall damage and osmotic lysis, remains to be established however [147].
In an investigation into the antimicrobial action of propolis, Mirzoeva and colleagues showed that one of its constituent flavonoids, quercetin, caused an increase in permeability of the inner bacterial membrane and a dissipation of the membrane potential [148]. The electrochemical gradient of protons across the membrane is essential for bacteria to maintain capacity for ATP synthesis, membrane transport and motility. Mirzoeva et al. suggested that the effect of propolis on membrane permeability and membrane potential may contribute enormously to its overall antibacterial activity and may decrease the resistance of cells to other antibacterial agents. It was thought that this might explain the synergistic effect that occurs between propolis and other antibiotics such as tetracycline [148] and ampicillin [149]. The group also demonstrated that the flavonoids quercetin and naringenin significantly inhibited bacterial motility, providing further evidence that the proton motive force is disrupted. Bacterial motility and chemotaxis are thought to be important in virulence as they guide bacteria to their sites of adherence and invasion. Mirzoeva et al. suggested that the antimotility action of propolis components may have an important role in inhibition of bacterial pathogenesis and the development of infection [148]. The cytoplasmic membrane activity detected for quercetin by Mirzoeva and co-workers may represent one of the additional mechanisms of antibacterial action that was suspected to be present among the seven DNA gyrase-inhibiting flavonoid compounds tested by Ohemeng and colleagues [68].
8.5.3. Inhibition of energy metabolism
Haraguchi and colleagues recently carried out an investigation into the antibacterial mode of action of two retrochalcones (licochalcone A and C) from the roots of Glycyrrhiza inflata [137]. These flavonoids demonstrated inhibitory activity against S. aureus and Micrococcus luteus but not against E. coli, and in preliminary tests licochalcone A inhibited incorporation of radioactive precursors into macromolecules (DNA, RNA and protein). The group hypothesised that the licochalcones may be interfering with energy metabolism in a similar way to respiratory-inhibiting antibiotics, since energy is required for active uptake of various metabolites and for biosynthesis of macromolecules [137]. Interestingly, the licochalcones were found to inhibit strongly oxygen consumption in M. luteus and S. aureus but not in E. coli, which correlated well with the observed spectrum of antibacterial activity. The group further demonstrated that licochalcones A and C effectively inhibited NADH-cytochrome c reductase, but not cytochrome c oxidase or NADH-CoQ reductase. It was therefore suggested that the inhibition site of these retrochalcones was between CoQ and cytochrome c in the bacterial respiratory electron transport chain [137].
Merck Research Laboratories recently reported that the flavanone lonchocarpol A inhibits macromolecular synthesis in Bacillus megaterium. Using radioactive precursors, it was demonstrated that RNA, DNA, cell wall and protein synthesis were all inhibited at concentrations similar to the MIC value [150]. This may represent another example of a flavonoid that interferes with energy metabolism.
9. Concluding remarks
With regard to natural products, it is generally accepted that phytochemicals are less potent anti-infectives than agents of microbial origin, i.e. antibiotics [48]. However, new classes of antimicrobial drug are urgently required and the flavonoids represent a novel set of leads. Future optimisation of these compounds through structural alteration may allow the development of a pharmacologically acceptable antimicrobial agent or group of agents. Existing structure–activity data suggest that it might be possible, for example, to prepare a potent antibacterial flavanone by synthesising a compound with halogenation of the B ring as well as lavandulyl or geranyl substitution of the A ring. Also, it is worth noting that the rapid progress which is being made toward elucidation of flavonoid biosynthetic pathways [151] may soon allow the production of structural analogues of active flavonoids through genetic manipulation. Screening of these analogues might lead to the identification of compounds that are sufficiently potent to be useful as antifungal, antiviral or antibacterial chemotherapeutics. In addition to the structural alteration of weak and moderately active antimicrobial flavonoids, investigation into the mechanisms of action of these compounds is likely to be a productive area of research. Such information may assist in the optimisation of a lead compound's activity, provide a focus for toxicological attention and aid in the anticipation of resistance. Also, characterisation of the interaction between antimicrobial flavonoids and their target sites could potentially allow the design of second-generation inhibitors.
Acknowledgments
The authors are very grateful to Dr Paul Kong and Dr Satyajit Sarker for critiquing preliminary drafts of the manuscript and for advice on flavonoid classification and structure. Thanks are extended to Dr Peter Taylor for insightful comments regarding interpretation of data from time–kill studies with flavonoids. Thanks also to Dr Derek Chapman, Miss Vivienne Hamilton, Dr Bruce Thomson and Mrs Amina Al-Mossawi for their kind support and encouragement.
References
- 1.Infectious Diseases Society of America. Statement of the IDSA concerning ‘Bioshield II: Responding to an ever-changing threat’. Alexandria, VA: IDSA; 2004.
- 2.Adcock H. Pharmageddon: is it too late to tackle growing resistance to anti-infectives? Pharm J. 2002;269:599–600. [Google Scholar]
- 3.Jeu L., Piacenti F.J., Lyakhovetskiy A.G., Fung H.B. Voriconazole. Clin Ther. 2003;25:1321–1381. doi: 10.1016/s0149-2918(03)80126-1. [DOI] [PubMed] [Google Scholar]
- 4.De Clercq E. New developments in anti-HIV chemotherapy. Farmaco. 2001;56:3–12. doi: 10.1016/s0014-827x(01)01007-2. [DOI] [PubMed] [Google Scholar]
- 5.Poole K. Overcoming antimicrobial resistance by targeting resistance mechanisms. J Pharm Pharmacol. 2001;53:283–294. doi: 10.1211/0022357011775514. [DOI] [PubMed] [Google Scholar]
- 6.Taylor P.W., Stapleton P.D., Paul Luzio J. New ways to treat bacterial infections. Drug Discov Today. 2002;7:1086–1091. doi: 10.1016/s1359-6446(02)02498-4. [DOI] [PubMed] [Google Scholar]
- 7.Anonymous. The global threat of antibiotic resistance (British Pharmaceutical Conference 2000). Pharm J 2000;265:692–4.
- 8.Kimberlin D.W., Whitley R.J. Antiviral resistance: mechanisms, clinical significance, and future implications. J Antimicrob Chemother. 1996;37:403–421. doi: 10.1093/jac/37.3.403. [DOI] [PubMed] [Google Scholar]
- 9.Silver L., Bostian K. Screening of natural products for antimicrobial agents. Eur J Clin Microbiol Infect Dis. 1990;9:455–461. doi: 10.1007/BF01964283. [DOI] [PubMed] [Google Scholar]
- 10.Havsteen B. Flavonoids, a class of natural products of high pharmacological potency. Biochem Pharmacol. 1983;32:1141–1148. doi: 10.1016/0006-2952(83)90262-9. [DOI] [PubMed] [Google Scholar]
- 11.Middleton E., Jr., Chithan K. The impact of plant flavonoids on mammalian biology: implications for immunity, inflammation and cancer. In: Harborne J.B., editor. The flavonoids: advances in research since 1986. Chapman and Hall; London, UK: 1993. [Google Scholar]
- 12.Grange J.M., Davey R.W. Antibacterial properties of propolis (bee glue) J R Soc Med. 1990;83:159–160. doi: 10.1177/014107689008300310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Harborne J.B., Baxter H. Vols 1 and 2. John Wiley and Sons; Chichester, UK: 1999. (The handbook of natural flavonoids). [Google Scholar]
- 14.Skibola C.F., Smith M.T. Potential health impacts of excessive flavonoid intake. Free Radic Biol Med. 2000;29:375–383. doi: 10.1016/s0891-5849(00)00304-x. [DOI] [PubMed] [Google Scholar]
- 15.Harborne J.B., Williams C.A. Advances in flavonoid research since 1992. Phytochemistry. 2000;55:481–504. doi: 10.1016/s0031-9422(00)00235-1. [DOI] [PubMed] [Google Scholar]
- 16.Brown J.P. A review of the genetic effects of naturally occurring flavonoids, anthraquinones and related compounds. Mutat Res. 1980;75:243–277. doi: 10.1016/0165-1110(80)90029-9. [DOI] [PubMed] [Google Scholar]
- 17.Muziol T., Cody V., Wojtczak A. Comparison of binding interactions of dibromoflavonoids with transthyretin. Acta Biochim Pol. 2001;48:885–892. [PubMed] [Google Scholar]
- 18.Villemin D., Martin B., Bar N. Application of microwave in organic synthesis; dry synthesis of 2-arylmethylene-3(2)-naphthofuranones. Molecules. 1998;3:88–93. [Google Scholar]
- 19.Xu H.X., Lee S.F. Activity of plant flavonoids against antibiotic-resistant bacteria. Phytother Res. 2001;15:39–43. doi: 10.1002/1099-1573(200102)15:1<39::aid-ptr684>3.0.co;2-r. [DOI] [PubMed] [Google Scholar]
- 20.Middleton E., Jr., Kandaswami C., Theoharides T.C. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol Rev. 2000;52:673–751. [PubMed] [Google Scholar]
- 21.Havsteen B.H. The biochemistry and medical significance of the flavonoids. Pharmacol Ther. 2002;96:67–202. doi: 10.1016/s0163-7258(02)00298-x. [DOI] [PubMed] [Google Scholar]
- 22.Tereschuk M.L., Riera M.V., Castro G.R., Abdala L.R. Antimicrobial activity of flavonoids from leaves of Tagetes minuta. J Ethnopharmacol. 1997;56:227–232. doi: 10.1016/s0378-8741(97)00038-x. [DOI] [PubMed] [Google Scholar]
- 23.The Bible, Jeremiah 8, verse 22; Jeremiah 46, verse 11; Jeremiah 51, verse 8.
- 24.Fearnley J. Souvenir Press Ltd.; London, UK: 2001. Bee propolis. [Google Scholar]
- 25.Bosio K., Avanzini C., D’Avolio A., Ozino O., Savoia D. In vitro activity of propolis against Streptococcus pyogenes. Lett Appl Microbiol. 2000;31:174–177. doi: 10.1046/j.1365-2672.2000.00785.x. [DOI] [PubMed] [Google Scholar]
- 26.Hegazi A.G., Abd El Hady F.K., Abd Allah F.A. Chemical composition and antimicrobial activity of European propolis. Z Naturforsch [C] 2000;55:70–75. doi: 10.1515/znc-2000-1-214. [DOI] [PubMed] [Google Scholar]
- 27.Pepeljnjak S., Jalsenjak I., Maysinger D. Growth inhibition of Bacillus subtilis and composition of various propolis extracts. Pharmazie. 1982;37:864–865. [PubMed] [Google Scholar]
- 28.Tsao T.F., Newman M.G., Kwok Y.Y., Horikoshi A.K. Effect of Chinese and western antimicrobial agents on selected oral bacteria. J Dent Res. 1982;61:1103–1106. doi: 10.1177/00220345820610091501. [DOI] [PubMed] [Google Scholar]
- 29.Tsuchiya H., Sato M., Miyazaki T. Comparative study on the antibacterial activity of phytochemical flavanones against methicillin-resistant Staphylococcus aureus. J Ethnopharmacol. 1996;50:27–34. doi: 10.1016/0378-8741(96)85514-0. [DOI] [PubMed] [Google Scholar]
- 30.Wachter G.A., Hoffmann J.J., Furbacher T., Blake M.E., Timmermann B.N. Antibacterial and antifungal flavanones from Eysenhardtia texana. Phytochemistry. 1999;52:1469–1471. doi: 10.1016/s0031-9422(99)00221-6. [DOI] [PubMed] [Google Scholar]
- 31.Valsaraj R., Pushpangadan P., Smitt U.W. New anti-HIV-1, antimalarial, and antifungal compounds from Terminalia bellerica. J Nat Prod. 1997;60:739–742. doi: 10.1021/np970010m. [DOI] [PubMed] [Google Scholar]
- 32.Zheng W.F., Tan R.X., Yang L., Liu Z.L. Two flavones from Artemisia giraldii and their antimicrobial activity. Planta Med. 1996;62:160–162. doi: 10.1055/s-2006-957841. [DOI] [PubMed] [Google Scholar]
- 33.Prescott L.M., Harley J.P., Klein D.A. WCB/McGraw-Hill; London, UK: 1999. Microbiology. [Google Scholar]
- 34.Cafarchia C., De Laurentis N., Milillo M.A., Losacco V., Puccini V. Antifungal activity of Apulia region propolis. Parassitologia. 1999;41:587–590. [PubMed] [Google Scholar]
- 35.Afolayan A.J., Meyer J.J. The antimicrobial activity of 3,5,7-trihydroxyflavone isolated from the shoots of Helichrysum aureonitens. J Ethnopharmacol. 1997;57:177–181. doi: 10.1016/s0378-8741(97)00065-2. [DOI] [PubMed] [Google Scholar]
- 36.Li B.Q., Fu T., Dongyan Y., Mikovits J.A., Ruscetti F.W., Wang J.M. Flavonoid baicalin inhibits HIV-1 infection at the level of viral entry. Biochem Biophys Res Commun. 2000;276:534–538. doi: 10.1006/bbrc.2000.3485. [DOI] [PubMed] [Google Scholar]
- 37.Li B.Q., Fu T., Yan Y.D., Baylor N.W., Ruscetti F.W., Kung H.F. Inhibition of HIV infection by baicalin — a flavonoid compound purified from Chinese herbal medicine. Cell Mol Biol Res. 1993;39:119–124. [PubMed] [Google Scholar]
- 38.Ono K., Nakane H., Fukushima M., Chermann J.C., Barre-Sinoussi F. Inhibition of reverse transcriptase activity by a flavonoid compound, 5,6,7-trihydroxyflavone. Biochem Biophys Res Commun. 1989;160:982–987. doi: 10.1016/s0006-291x(89)80097-x. [DOI] [PubMed] [Google Scholar]
- 39.Lin Y.M., Anderson H., Flavin M.T. In vitro anti-HIV activity of biflavonoids isolated from Rhus succedanea and Garcinia multiflora. J Nat Prod. 1997;60:884–888. doi: 10.1021/np9700275. [DOI] [PubMed] [Google Scholar]
- 40.Moore P.S., Pizza C. Observations on the inhibition of HIV-1 reverse transcriptase by catechins. Biochem J. 1992;288:717–719. doi: 10.1042/bj2880717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Brinkworth R.I., Stoermer M.J., Fairlie D.P. Flavones are inhibitors of HIV-1 proteinase. Biochem Biophys Res Commun. 1992;188:631–637. doi: 10.1016/0006-291x(92)91103-w. [DOI] [PubMed] [Google Scholar]
- 42.Fesen M.R., Pommier Y., Leteurtre F., Hiroguchi S., Yung J., Kohn K.W. Inhibition of HIV-1 integrase by flavones, caffeic acid phenethyl ester (CAPE) and related compounds. Biochem Pharmacol. 1994;48:595–608. doi: 10.1016/0006-2952(94)90291-7. [DOI] [PubMed] [Google Scholar]
- 43.Kim H.J., Woo E.R., Shin C.G., Park H. A new flavonol glycoside gallate ester from Acer okamotoanum and its inhibitory activity against human immunodeficiency virus-1 (HIV-1) integrase. J Nat Prod. 1998;61:145–148. doi: 10.1021/np970171q. [DOI] [PubMed] [Google Scholar]
- 44.Ono K., Nakane H., Fukushima M., Chermann J.C., Barre-Sinoussi F. Differential inhibitory effects of various flavonoids on the activities of reverse transcriptase and cellular DNA and RNA polymerases. Eur J Biochem. 1990;190:469–476. doi: 10.1111/j.1432-1033.1990.tb15597.x. [DOI] [PubMed] [Google Scholar]
- 45.Critchfield J.W., Butera S.T., Folks T.M. Inhibition of HIV activation in latently infected cells by flavonoid compounds. AIDS Res Hum Retroviruses. 1996;12:39–46. doi: 10.1089/aid.1996.12.39. [DOI] [PubMed] [Google Scholar]
- 46.Hu C.Q., Chen K., Shi Q., Kilkuskie R.E., Cheng Y.C., Lee K.H. Anti-AIDS agents, 10. Acacetin-7-O-beta-D-galactopyranoside, an anti-HIV principle from Chrysanthemum morifolium and a structure–activity correlation with some related flavonoids. J Nat Prod. 1994;57:42–51. doi: 10.1021/np50103a006. [DOI] [PubMed] [Google Scholar]
- 47.Selway J.W.T. Antiviral activity of flavones and flavans. In: Cody V., Middleton E., Harborne J.B., editors. Plant flavonoids in biology and medicine: biochemical, pharmacological, and structure–activity relationships. Alan R. Liss, Inc.; New York, NY: 1986. [Google Scholar]
- 48.Yamada H. Natural products of commercial potential as medicines. Curr Opin Biotechnol. 1991;2:203–210. doi: 10.1016/0958-1669(91)90011-s. [DOI] [PubMed] [Google Scholar]
- 49.Balde A.M., Van Hoof L., Pieters L.A., Vanden Berghe D.A., Vlietinck A.J. Plant antiviral agents. VII. Antiviral and antibacterial proanthocyanidins from the bark of Pavetta owariensis. Phytother Res. 1990;4:182–188. [Google Scholar]
- 50.Cheng P.C., Wong G. Honey bee propolis: prospects in medicine. Bee World. 1996;77:8–15. [Google Scholar]
- 51.Meyer J.J., Afolayan A.J., Taylor M.B., Erasmus D. Antiviral activity of galangin isolated from the aerial parts of Helichrysum aureonitens. J Ethnopharmacol. 1997;56:165–169. doi: 10.1016/s0378-8741(97)01514-6. [DOI] [PubMed] [Google Scholar]
- 52.Amoros M., Simoes C.M., Girre L., Sauvager F., Cormier M. Synergistic effect of flavones and flavonols against herpes simplex virus type 1 in cell culture. Comparison with the antiviral activity of propolis. J Nat Prod. 1992;55:1732–1740. doi: 10.1021/np50090a003. [DOI] [PubMed] [Google Scholar]
- 53.Mucsi I., Gyulai Z., Beladi I. Combined effects of flavonoids and acyclovir against herpesviruses in cell cultures. Acta Microbiol Hung. 1992;39:137–147. [PubMed] [Google Scholar]
- 54.Dall’Agnol R., Ferraz A., Bernardi A.P. Antimicrobial activity of some Hypericum species. Phytomedicine. 2003;10:511–516. doi: 10.1078/094471103322331476. [DOI] [PubMed] [Google Scholar]
- 55.El-Abyad M.S., Morsi N.M., Zaki D.A., Shaaban M.T. Preliminary screening of some Egyptian weeds for antimicrobial activity. Microbios. 1990;62:47–57. [PubMed] [Google Scholar]
- 56.Aladesanmi A.J., Sofowora A., Leary J.D. Preliminary biological and phytochemical investigation of two Nigerian medicinal plants. Int J Crude Drug Res. 1986;24:147–153. [Google Scholar]
- 57.Al-Saleh F.S., Gamal El-Din A.Y., Abbas J.A., Saeed N.A. Phytochemical and biological studies of medicinal plants in Bahrain: family Chenopodiaceae. Part 2. Int J Pharmacogn. 1997;35:38–42. [Google Scholar]
- 58.Mahmoud M.J., Jawad A.L., Hussain A.M., Al-Omari M., Al-Naib A. In vitro antimicrobial activity of Salsola rosmarinus and Adiantum capillus-veneris. Int J Crude Drug Res. 1989;27:14–16. [Google Scholar]
- 59.Quarenghi M.V., Tereschuk M.L., Baigori M.D., Abdala L.R. Antimicrobial activity of flowers from Anthemis cotula. Fitoterapia. 2000;71:710–712. doi: 10.1016/s0367-326x(00)00229-x. [DOI] [PubMed] [Google Scholar]
- 60.Rauha J.P., Remes S., Heinonen M. Antimicrobial effects of Finnish plant extracts containing flavonoids and other phenolic compounds. Int J Food Microbiol. 2000;56:3–12. doi: 10.1016/s0168-1605(00)00218-x. [DOI] [PubMed] [Google Scholar]
- 61.Singh R.K., Nath G. Antimicrobial activity of Elaeocarpus sphaericus. Phytother Res. 1999;13:448–450. doi: 10.1002/(sici)1099-1573(199908/09)13:5<448::aid-ptr480>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 62.Tarle D., Dvorzak I. Antimicrobial activity of the plant Cirsium oleraceum (L.) Scop. Acta Pharm Jugosl. 1990;40:569–571. [Google Scholar]
- 63.Torrenegra R.D., Ricardo A.A., Pedrozo J.P., Fuentes O.C. Flavonoids from Gnaphalium gracile H.B.K. Int J Crude Drug Res. 1989;27:22–24. [Google Scholar]
- 64.Park Y.K., Ikegaki M. Preparation of water and ethanolic extracts of propolis and evaluation of the preparations. Biosci Biotechnol Biochem. 1998;62:2230–2232. doi: 10.1271/bbb.62.2230. [DOI] [PubMed] [Google Scholar]
- 65.Khanna P., Sharma O.P., Sehgal M. Antimicrobial principles from tissue culture of some plant species. Indian J Pharm Sci. 1980;42:113–117. [Google Scholar]
- 66.Palacios P., Gutkind G., Rondina R.V., de Torres R., Coussio J.D. Genus Baccharis. II. Antimicrobial activity of B. crispa and B. notosergila. Planta Med. 1983;49:128. [PubMed] [Google Scholar]
- 67.Oksuz S., Ayyildiz H., Johansson C. 6-Methoxylated and C-glycosyl flavonoids from Centaurea species. J Nat Prod. 1984;47:902–903. doi: 10.1021/np50035a034. [DOI] [PubMed] [Google Scholar]
- 68.Ohemeng K.A., Schwender C.F., Fu K.P., Barrett J.F. DNA gyrase inhibitory and antibacterial activity of some flavones (1) Bioorg Med Chem Lett. 1993;3:225–230. [Google Scholar]
- 69.Bashir A.K., Abdalla A.A., Wasfi I.A., Hassan E.S., Amiri M.H., Crabb T.A. Flavonoids of Limonium axillare. Int J Pharmacogn. 1994;32:366–372. [Google Scholar]
- 70.Aljancic I., Vajs V., Menkovic N. Flavones and sesquiterpene lactones from Achillea atrata subsp. multifida: antimicrobial activity. J Nat Prod. 1999;62:909–911. doi: 10.1021/np980536m. [DOI] [PubMed] [Google Scholar]
- 71.Basile A., Giordano S., Lopez-Saez J.A., Cobianchi R.C. Antibacterial activity of pure flavonoids isolated from mosses. Phytochemistry. 1999;52:1479–1482. doi: 10.1016/s0031-9422(99)00286-1. [DOI] [PubMed] [Google Scholar]
- 72.Basile A., Sorbo S., Giordano S. Antibacterial and allelopathic activity of extract from Castanea sativa leaves. Fitoterapia. 2000;71:S110–S116. doi: 10.1016/s0367-326x(00)00185-4. [DOI] [PubMed] [Google Scholar]
- 73.Sato Y., Suzaki S., Nishikawa T., Kihara M., Shibata H., Higuti T. Phytochemical flavones isolated from Scutellaria barbata and antibacterial activity against methicillin-resistant Staphylococcus aureus. J Ethnopharmacol. 2000;72:483–488. doi: 10.1016/s0378-8741(00)00265-8. [DOI] [PubMed] [Google Scholar]
- 74.Nishino C., Enoki N., Tawata S., Mori A., Kobayashi K., Fukushima M. Antibacterial activity of flavonoids against Staphylococcus epidermidis, a skin bacterium. Agric Biol Chem. 1987;51:139–143. [Google Scholar]
- 75.Cushnie T.P.T., Hamilton V.E.S., Lamb A.J. Assessment of the antibacterial activity of selected flavonoids and consideration of discrepancies between previous reports. Microbiol Res. 2003;158:281–289. doi: 10.1078/0944-5013-00206. [DOI] [PubMed] [Google Scholar]
- 76.Pomilio A.B., Buschi C.A., Tomes C.N., Viale A.A. Antimicrobial constituents of Gomphrena martiana and Gomphrena boliviana. J Ethnopharmacol. 1992;36:155–161. doi: 10.1016/0378-8741(92)90016-k. [DOI] [PubMed] [Google Scholar]
- 77.Pepeljnjak S., Kosalec I. Galangin expresses bactericidal activity against multiple-resistant bacteria: MRSA, Enterococcus spp. and Pseudomonas aeruginosa. FEMS Microbiol Lett. 2004;240:111–116. doi: 10.1016/j.femsle.2004.09.018. [DOI] [PubMed] [Google Scholar]
- 78.Fukui H., Goto K., Tabata M. Two antimicrobial flavanones from the leaves of Glycyrrhiza glabra. Chem Pharm Bull (Tokyo) 1988;36:4174–4176. doi: 10.1248/cpb.36.4174. [DOI] [PubMed] [Google Scholar]
- 79.Hufford C.D., Lasswell W.L. Antimicrobial activities of constituents of Uvaria chamae. Lloydia. 1978;41:156–160. [PubMed] [Google Scholar]
- 80.Bae E.A., Han M.J., Kim D.H. In vitro anti-Helicobacter pylori activity of some flavonoids and their metabolites. Planta Med. 1999;65:442–443. doi: 10.1055/s-2006-960805. [DOI] [PubMed] [Google Scholar]
- 81.Kim D.H., Bae E.A., Han M.J. Anti-Helicobacter pylori activity of the metabolites of poncirin from Poncirus trifoliata by human intestinal bacteria. Biol Pharm Bull. 1999;22:422–444. doi: 10.1248/bpb.22.422. [DOI] [PubMed] [Google Scholar]
- 82.Cottiglia F., Loy G., Garau D. Antimicrobial evaluation of coumarins and flavonoids from the stems of Daphne gnidium L. Phytomedicine. 2001;8:302–305. doi: 10.1078/0944-7113-00036. [DOI] [PubMed] [Google Scholar]
- 83.Sakagami Y., Mimura M., Kajimura K. Anti-MRSA activity of sophoraflavanone G and synergism with other antibacterial agents. Lett Appl Microbiol. 1998;27:98–100. doi: 10.1046/j.1472-765x.1998.00386.x. [DOI] [PubMed] [Google Scholar]
- 84.Sato M., Tsuchiya H., Takase I., Kureshiro H., Tanigaki S., Iinuma M. Antibacterial activity of flavanone isolated from Sophora exigua against methicillin-resistant Staphylococcus aureus and its combination with antibiotics. Phytother Res. 1995;9:509–512. [Google Scholar]
- 85.Tsuchiya H., Sato M., Iinuma M. Inhibition of the growth of cariogenic bacteria in vitro by plant flavanones. Experientia. 1994;50:846–849. doi: 10.1007/BF01956469. [DOI] [PubMed] [Google Scholar]
- 86.Ng T.B., Ling J.M., Wang Z.T., Cai J.N., Xu G.J. Examination of coumarins, flavonoids and polysaccharopeptide for antibacterial activity. Gen Pharmacol. 1996;27:1237–1240. doi: 10.1016/0306-3623(95)02143-4. [DOI] [PubMed] [Google Scholar]
- 87.Ramaswamy A.S., Jayaraman S., Sirsi M., Rao K.H. Antibacterial action of some naturally occurring citrus bioflavonoids. Indian J Exp Biol. 1972;10:72–73. [PubMed] [Google Scholar]
- 88.Ikigai H., Nakae T., Hara Y., Shimamura T. Bactericidal catechins damage the lipid bilayer. Biochim Biophys Acta. 1993;1147:132–136. doi: 10.1016/0005-2736(93)90323-r. [DOI] [PubMed] [Google Scholar]
- 89.Kono K., Tatara I., Takeda S., Arakawa K., Hara Y. Antibacterial activity of epigallocatechin gallate against methicillin-resistant Staphylococcus aureus. Kansenshogaku Zasshi. 1994;68:1518–1522. doi: 10.11150/kansenshogakuzasshi1970.68.1518. [DOI] [PubMed] [Google Scholar]
- 90.Sakanaka S., Kim M., Taniguchi M., Yamamoto T. Antibacterial substances in Japanese green tea extract against Streptococcus mutans, a cariogenic bacterium. Agric Biol Chem. 1989;53:2307–2311. [Google Scholar]
- 91.Yam T.S., Shah S., Hamilton-Miller J.M. Microbiological activity of whole and fractionated crude extracts of tea (Camellia sinensis), and of tea components. FEMS Microbiol Lett. 1997;152:169–174. doi: 10.1111/j.1574-6968.1997.tb10424.x. [DOI] [PubMed] [Google Scholar]
- 92.Yee Y.K., Koo M.W. Anti-Helicobacter pylori activity of Chinese tea: in vitro study. Aliment Pharmacol Ther. 2000;14:635–638. doi: 10.1046/j.1365-2036.2000.00747.x. [DOI] [PubMed] [Google Scholar]
- 93.Zhao W.H., Hu Z.Q., Okubo S., Hara Y., Shimamura T. Mechanism of synergy between epigallocatechin gallate and beta-lactams against methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2001;45:1737–1742. doi: 10.1128/AAC.45.6.1737-1742.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Stapleton P.D., Shah S., Hamilton-Miller J.M.T. Anti-Staphylococcus aureus activity and oxacillin resistance modulating capacity of 3-O-acyl-catechins. Int J Antimicrob Agents. 2004;24:374–380. doi: 10.1016/j.ijantimicag.2004.03.024. [DOI] [PubMed] [Google Scholar]
- 95.Taguri T., Tanaka T., Kouno I. Antimicrobial activity of 10 different plant polyphenols against bacteria causing food-borne disease. Biol Pharm Bull. 2004;27:1965–1969. doi: 10.1248/bpb.27.1965. [DOI] [PubMed] [Google Scholar]
- 96.Miski M., Ulubelen A., Johansson C., Mabry T.J. Antibacterial activity studies of flavonoids from Salvia palaestina. J Nat Prod. 1983;46:874–875. doi: 10.1021/np50030a007. [DOI] [PubMed] [Google Scholar]
- 97.Arima H., Danno G. Isolation of antimicrobial compounds from guava (Psidium guajava L.) and their structural elucidation. Biosci Biotechnol Biochem. 2002;66:1727–1730. doi: 10.1271/bbb.66.1727. [DOI] [PubMed] [Google Scholar]
- 98.el-Gammal A.A., Mansour R.M. Antimicrobial activities of some flavonoid compounds. Zentralbl Mikrobiol. 1986;141:561–565. doi: 10.1016/s0232-4393(86)80010-5. [DOI] [PubMed] [Google Scholar]
- 99.Gutkind G., Norbedo C., Mollerach M., Ferraro G., Coussio J.D., de Torres R. Antibacterial activity of Achyrocline flaccida. J Ethnopharmacol. 1984;10:319–321. doi: 10.1016/0378-8741(84)90019-9. [DOI] [PubMed] [Google Scholar]
- 100.Jit S., Nag T.N. Antimicrobial principles from in vitro tissue culture of Tribulus alatus. Indian J Pharm Sci. 1985;47:101–103. [Google Scholar]
- 101.Van Puyvelde L., De Kimpe N., Costa J. Isolation of flavonoids and a chalcone from Helichrysum odoratissimum and synthesis of helichrysetin. J Nat Prod. 1989;52:629–633. doi: 10.1021/np50063a025. [DOI] [PubMed] [Google Scholar]
- 102.Waage S.K., Hedin P.A. Quercetin 3-O-galactosyl-(1-6)-glucoside, a compound from narrowleaf vetch with antibacterial activity. Phytochemistry. 1985;24:243–245. [Google Scholar]
- 103.Sakar M.K., Engelshowe R., Tamer A.U. Isolation and antimicrobial activity of flavonoids from Prunus spinosa L. flowers. Hacettepe Universitesi Eczacilik Fakultesi Dergisi. 1992;12:59–63. [Google Scholar]
- 104.Alcaraz L.E., Blanco S.E., Puig O.N., Tomas F., Ferretti F.H. Antibacterial activity of flavonoids against methicillin-resistant Staphylococcus aureus strains. J Theor Biol. 2000;205:231–240. doi: 10.1006/jtbi.2000.2062. [DOI] [PubMed] [Google Scholar]
- 105.Encarnacion R., Ochoa N., Anthoni U., Christophersen C., Nielsen P.H. Two new flavones from Calliandra californica. J Nat Prod. 1994;57:1307–1309. doi: 10.1021/np50111a023. [DOI] [PubMed] [Google Scholar]
- 106.Iniesta-Sanmartin E., Tomas-Barberan F.A., Guirado A., Tomas-Lorente F. Antibacterial flavonoids from Helichrysum picardii and H. italicum. Planta Med. 1990;56:648–649. [Google Scholar]
- 107.Sato M., Fujiwara S., Tsuchiya H. Flavones with antibacterial activity against cariogenic bacteria. J Ethnopharmacol. 1996;54:171–176. doi: 10.1016/s0378-8741(96)01464-x. [DOI] [PubMed] [Google Scholar]
- 108.el-Lakany A.M., Abdel-Kader M.S., Hammoda H.M., Ghazy N.M., Mahmoud Z.F. A new flavone glycoside with antimicrobial activity from Carduus pycnocephalus L. Pharmazie. 1997;52:78–79. [PubMed] [Google Scholar]
- 109.Verma D.K., Singh S.K., Tripathi V. A rare antibacterial flavone glucoside from Lantana camara. Indian Drugs. 1997;34:32–35. [Google Scholar]
- 110.Dastidar S.G., Manna A., Kumar K.A. Studies on the antibacterial potentiality of isoflavones. Int J Antimicrob Agents. 2004;23:99–102. doi: 10.1016/j.ijantimicag.2003.06.003. [DOI] [PubMed] [Google Scholar]
- 111.Chacha M., Bojase-Moleta G., Majinda R.R. Antimicrobial and radical scavenging flavonoids from the stem wood of Erythrina latissima. Phytochemistry. 2005;66:99–104. doi: 10.1016/j.phytochem.2004.10.013. [DOI] [PubMed] [Google Scholar]
- 112.Biyiti L., Pesando D., Puiseux-Dao S. Antimicrobial activity of two flavanones isolated from the Cameroonian plant Erythrina sigmoidea. Planta Med. 1988;54:126–128. doi: 10.1055/s-2006-962368. [DOI] [PubMed] [Google Scholar]
- 113.Deng Y., Lee J.P., Tianasoa-Ramamonjy M. New antimicrobial flavanones from Physena madagascariensis. J Nat Prod. 2000;63:1082–1089. doi: 10.1021/np000054m. [DOI] [PubMed] [Google Scholar]
- 114.Kuroyanagi M., Arakawa T., Hirayama Y., Hayashi T. Antibacterial and antiandrogen flavonoids from Sophora flavescens. J Nat Prod. 1999;62:1595–1599. doi: 10.1021/np990051d. [DOI] [PubMed] [Google Scholar]
- 115.Osawa K., Yasuda H., Maruyama T., Morita H., Takeya K., Itokawa H. Isoflavanones from the heartwood of Swartzia polyphylla and their antibacterial activity against cariogenic bacteria. Chem Pharm Bull (Tokyo) 1992;40:2970–2974. doi: 10.1248/cpb.40.2970. [DOI] [PubMed] [Google Scholar]
- 116.Li W., Asada Y., Yoshikawa T. Antimicrobial flavonoids from Glycyrrhiza glabra hairy root cultures. Planta Med. 1998;64:746–747. doi: 10.1055/s-2006-957571. [DOI] [PubMed] [Google Scholar]
- 117.Simin K., Ali Z., Khaliq-Uz-Zaman S.M., Ahmad V.U. Structure and biological activity of a new rotenoid from Pongamia pinnata. Nat Prod Lett. 2002;16:351–357. doi: 10.1080/10575630290033114. [DOI] [PubMed] [Google Scholar]
- 118.Faizi S., Ali M. Shamimin: a new flavonol C-glycoside from leaves of Bombax ceiba. Planta Med. 1999;65:383–385. doi: 10.1055/s-2006-960796. [DOI] [PubMed] [Google Scholar]
- 119.Liu H., Orjala J., Sticher O., Rali T. Acylated flavonol glycosides from leaves of Stenochlaena palustris. J Nat Prod. 1999;62:70–75. doi: 10.1021/np980179f. [DOI] [PubMed] [Google Scholar]
- 120.Yadava R.N., Reddy K.I. A new bio-active flavonol glycoside from the stems of Butea superba Roxb. J Asian Nat Prod Res. 1998;1:139–145. doi: 10.1080/10286029808039856. [DOI] [PubMed] [Google Scholar]
- 121.Gafner S., Wolfender J.L., Mavi S., Hostettmann K. Antifungal and antibacterial chalcones from Myrica serrata. Planta Med. 1996;62:67–69. doi: 10.1055/s-2006-957804. [DOI] [PubMed] [Google Scholar]
- 122.Hamilton-Miller J.M.T., Shah S. Activity of the tea component epicatechin gallate and analogues against methicillin-resistant Staphylococcus aureus. J Antimicrob Chemother. 2000;46:852–853. doi: 10.1093/jac/46.5.852. [DOI] [PubMed] [Google Scholar]
- 123.Shiota S., Shimizu M., Mizushima T. Marked reduction in the minimum inhibitory concentration (MIC) of beta-lactams in methicillin-resistant Staphylococcus aureus produced by epicatechin gallate, an ingredient of green tea (Camellia sinensis) Biol Pharm Bull. 1999;22:1388–1390. doi: 10.1248/bpb.22.1388. [DOI] [PubMed] [Google Scholar]
- 124.Yam T.S., Hamilton-Miller J.M.T., Shah S. The effect of a component of tea (Camellia sinensis) on methicillin resistance, PBP2′ synthesis, and beta-lactamase production in Staphylococcus aureus. J Antimicrob Chemother. 1998;42:211–216. doi: 10.1093/jac/42.2.211. [DOI] [PubMed] [Google Scholar]
- 125.Stapleton P.D., Shah S., Anderson J.C., Hara Y., Hamilton-Miller J.M.T., Taylor P.W. Modulation of beta-lactam resistance in Staphylococcus aureus by catechins and gallates. Int J Antimicrob Agents. 2004;23:462–467. doi: 10.1016/j.ijantimicag.2003.09.027. [DOI] [PubMed] [Google Scholar]
- 126.Arima H., Ashida H., Danno G. Rutin-enhanced antibacterial activities of flavonoids against Bacillus cereus and Salmonella enteritidis. Biosci Biotechnol Biochem. 2002;66:1009–1014. doi: 10.1271/bbb.66.1009. [DOI] [PubMed] [Google Scholar]
- 127.Ayhan-Kilcigil G., Bozdag O., Tuncbilek M., Altanlar N., Ertan R. Synthesis and antimicrobial activity of flavone-6-carboxaldehyde oxime ether derivatives. Pharmazie. 1999;54:228–229. [PubMed] [Google Scholar]
- 128.Bozdag-Dundar O., Tuncbilek M., Altanlar N., Ertan R. Synthesis and antimicrobial activity of flavone-3′-carboxaldehyde oxime ether derivatives. Arzneimittelforschung. 2003;53:522–525. doi: 10.1055/s-0031-1297143. [DOI] [PubMed] [Google Scholar]
- 129.Tuncbilek M., Bozdag O., Ayhan-Kilcigil G., Altanlar N., Buyukbingol E., Ertan R. Synthesis and antimicrobial activity of some new flavonyl oxime ether derivatives. Arzneimittelforschung. 1999;49:853–857. doi: 10.1055/s-0031-1300514. [DOI] [PubMed] [Google Scholar]
- 130.Wang S.X., Zhang F.J., Feng Q.P., Li Y.L. Synthesis, characterization, and antibacterial activity of transition metal complexes with 5-hydroxy-7,4′-dimethoxyflavone. J Inorg Biochem. 1992;46:251–257. doi: 10.1016/0162-0134(92)80035-t. [DOI] [PubMed] [Google Scholar]
- 131.Ward F.E., Garling D.L., Buckler R.T., Lawler D.M., Cummings D.P. Antimicrobial 3-methyleneflavanones. J Med Chem. 1981;24:1073–1077. doi: 10.1021/jm00141a011. [DOI] [PubMed] [Google Scholar]
- 132.Vijaya K., Ananthan S. Therapeutic efficacy of medicinal plants against experimentally induced shigellosis in guinea pigs. Indian J Pharm Sci. 1996;58:191–193. [Google Scholar]
- 133.Mariee N.K., Khalil A.A., Nasser A.A., al-Hiti M.M., Ali W.M. Isolation of the antimicrobial alkaloid stemmadenine from Iraqi Rhazya stricta. J Nat Prod. 1988;51:186–187. doi: 10.1021/np50055a036. [DOI] [PubMed] [Google Scholar]
- 134.Liu I.X., Durham D.G., Richards RM.E. Baicalin synergy with beta-lactam antibiotics against methicillin-resistant Staphylococcus aureus and other beta-lactam-resistant strains of S. aureus. J Pharm Pharmacol. 2000;52:361–366. doi: 10.1211/0022357001773922. [DOI] [PubMed] [Google Scholar]
- 135.National Committee for Clinical Laboratory Standards. Methods for determining bactericidal activity of antimicrobial agents; Approved guideline (M26-A), Vol. 19 (18). Wayne, PA: NCCLS; 2000.
- 136.National Committee for Clinical Laboratory Standards. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; Approved guideline (M7-A5), Vol. 20 (2). Wayne, PA: NCCLS; 2000.
- 137.Haraguchi H., Tanimoto K., Tamura Y., Mizutani K., Kinoshita T. Mode of antibacterial action of retrochalcones from Glycyrrhiza inflata. Phytochemistry. 1998;48:125–129. doi: 10.1016/s0031-9422(97)01105-9. [DOI] [PubMed] [Google Scholar]
- 138.Mori A., Nishino C., Enoki N., Tawata S. Antibacterial activity and mode of action of plant flavonoids against Proteus vulgaris and Staphylococcus aureus. Phytochemistry. 1987;26:2231–2234. [Google Scholar]
- 139.Tsuchiya H., Iinuma M. Reduction of membrane fluidity by antibacterial sophoraflavanone G isolated from Sophora exigua. Phytomedicine. 2000;7:161–165. doi: 10.1016/S0944-7113(00)80089-6. [DOI] [PubMed] [Google Scholar]
- 140.Sato M., Tsuchiya H., Akagiri M., Takagi N., Iinuma M. Growth inhibition of oral bacteria related to denture stomatitis by anti-candidal chalcones. Aust Dent J. 1997;42:343–346. doi: 10.1111/j.1834-7819.1997.tb00141.x. [DOI] [PubMed] [Google Scholar]
- 141.Hilliard J.J., Krause H.M., Bernstein J.I. A comparison of active site binding of 4-quinolones and novel flavone gyrase inhibitors to DNA gyrase. Adv Exp Med Biol. 1995;390:59–69. doi: 10.1007/978-1-4757-9203-4_5. [DOI] [PubMed] [Google Scholar]
- 142.Plaper A., Golob M., Hafner I., Oblak M., Solmajer T., Jerala R. Characterization of quercetin binding site on DNA gyrase. Biochem Biophys Res Commun. 2003;306:530–536. doi: 10.1016/s0006-291x(03)01006-4. [DOI] [PubMed] [Google Scholar]
- 143.Bernard F.X., Sable S., Cameron B. Glycosylated flavones as selective inhibitors of topoisomerase IV. Antimicrob Agents Chemother. 1997;41:992–998. doi: 10.1128/aac.41.5.992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Normark S. Transduction and dominance studies of the envA gene present in a chain-forming mutant of Escherichia coli K12. J Gen Microbiol. 1969;57 [PubMed] [Google Scholar]
- 145.Normark S., Boman H.G., Matsson E. Mutant of Escherichia coli with anomalous cell division and ability to decrease episomally and chromosomally mediated resistance to ampicillin and several other antibiotics. J Bacteriol. 1969;97:1334–1342. doi: 10.1128/jb.97.3.1334-1342.1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Cushnie TPT, Lamb AJ. Assessment of the antibacterial activity of galangin against 4-quinolone resistant strains of Staphylococcus aureus. Phytomedicine, in press. [DOI] [PubMed]
- 147.Cushnie T.P.T., Lamb A.J. Detection of galangin-induced cytoplasmic membrane damage in Staphylococcus aureus by measuring potassium loss. J Ethnopharmacol. 2005;101:243–248. doi: 10.1016/j.jep.2005.04.014. [DOI] [PubMed] [Google Scholar]
- 148.Mirzoeva O.K., Grishanin R.N., Calder P.C. Antimicrobial action of propolis and some of its components: the effects on growth, membrane potential and motility of bacteria. Microbiol Res. 1997;152:239–246. doi: 10.1016/S0944-5013(97)80034-1. [DOI] [PubMed] [Google Scholar]
- 149.Stepanovic S., Antic N., Dakic I., Svabic-Vlahovic M. In vitro antimicrobial activity of propolis and synergism between propolis and antimicrobial drugs. Microbiol Res. 2003;158:353–357. doi: 10.1078/0944-5013-00215. [DOI] [PubMed] [Google Scholar]
- 150.Salvatore M.J., King A.B., Graham A.C. Antibacterial activity of lonchocarpol A. J Nat Prod. 1998;61:640–642. doi: 10.1021/np9703961. [DOI] [PubMed] [Google Scholar]
- 151.Dixon R.A., Howles P.A., Lamb C., He X.Z., Reddy J.T. Prospects for the metabolic engineering of bioactive flavonoids and related phenylpropanoid compounds. Adv Exp Med Biol. 1998;439:55–66. doi: 10.1007/978-1-4615-5335-9_5. [DOI] [PubMed] [Google Scholar]