

Abstract
It has been well established that the formation of reactive metabolites of drugs is associated with drug toxicity. Similarly, there are accumulating data suggesting the role of the formation of reactive metabolites/intermediates through bioactivation in herbal toxicity and carcinogenicity. It has been hypothesized that the resultant reactive metabolites following herbal bioactivation covalently bind to cellular proteins and DNA, leading to toxicity via multiple mechanisms such as direct cytotoxicity, oncogene activation, and hypersensitivity reactions. This is exemplified by aristolochic acids present in Aristolochia spp, undergoing reduction of the nitro group by hepatic cytochrome P450 (CYP1A1/2) or peroxidases in extrahepatic tissues to reactive cyclic nitrenium ion. The latter was capable of reacting with DNA and proteins, resulting in activation of H-ras oncogene, gene mutation and finally carcinogenesis. Other examples are pulegone present in essential oils from many mint species; and teucrin A, a diterpenoid found in germander (Teuchrium chamaedrys) used as an adjuvant to slimming diets. Extensive pulegone metabolism generated p-cresol that was a glutathione depletory, and the furan ring of the diterpenoids in germander was oxidized by CYP3A4 to reactive epoxide which reacts with proteins such as CYP3A and epoxide hydrolase. On the other hand, some herbal/dietary constituents were shown to form reactive intermediates capable of irreversibly inhibiting various CYPs. The resultant metabolites lead to CYP inactivation by chemical modification of the heme, the apoprotein, or both as a result of covalent binding of modified heme to the apoprotein. Some examples include bergamottin, a furanocoumarin of grapefruit juice; capsaicin from chili peppers; glabridin, an isoflavan from licorice root; isothiocyanates found in all cruciferous vegetables; oleuropein rich in olive oil; dially sulfone found in garlic; and resveratrol, a constituent of red wine. CYPs have been known to metabolize more than 95% therapeutic drugs and activate a number of procarcinogens as well. Therefore, mechanism-based inhibition of CYPs may provide an explanation for some reported herb-drug interactions and chemopreventive activity of herbs. Due to the wide use and easy availability of herbal medicines, there is increasing concern about herbal toxicity. The safety and quality of herbal medicine should be ensured through greater research, pharmacovigilance, greater regulatory control and better communication between patients and health professionals.
Keywords: Herbal medicine, Toxicity, Metabolism, Bioactivation
Introduction
There is resurgence in the use of herbal medicines worldwide. An estimated one third of adults in the Western world use alternative therapies, including herbs. These herbs may be used either in their primary forms or combined into mixtures. In contrast to chemical drugs, herbs have sometimes been claimed to be non-toxic, because of their natural origin and long-term use as folk medicines. However, problems may arise due to intrinsic toxicity, adulteration, substitution, contamination, misidentification, drug-herb interactions, and lack of standardizationDe Smet, 1995, Ernst, 2002, Ernst and Pittler, 2002, Koh and Woo, 2000. Both adverse drug reactions and poisonings associated with the use of herbal medicines have increasingly been reported Deng, 2002, Ernst and Pittler, 2002, Klepser and Klepser, 1999. Herbal use has been associated with organ toxicities of heart, liver, blood, kidneys, central nervous system, and skin and carcinogenesis Bensoussan et al., 2002, Ernst, 2000, Greensfelder, 2000, Haller and Benowitz, 2000, Kessler, 2000, McRae et al., 2002, Stedman, 2002, Villegas et al., 2001, Zaacks et al., 1999.
The human cytochrome P450 (CYP) (EC 1.14.14.1) superfamily, containing 57 genes (Nelson, 2003), contributes to the metabolism of a variety of xenobiotics including therapeutic drugs, carcinogens, steroids and eicosanoids Gonzalez, 1990, Nebert and Russell, 2002, Nelson et al., 1996, Rendic, 2002, Rendic and Di Carlo, 1997. The resultant increases in polarity usually facilitate excretion and are considered to be a detoxification process, but in some instances, foreign compounds are converted to products with much greater cytotoxicity, mutagenicity, or carcinogenicity Guengerich and Liebler, 1985, Guengerich and Shimada, 1991, Zhou, 2003. Metabolic activation of procarcinogens is often catalyzed by a limited number of human CYPs including CYP1A1, 1A2, 1B1, 2A6, 2B6 and 3A4 Guengerich, 2001, Guengerich and Liebler, 1985, Guengerich and Shimada, 1991, Zhou, 2003. Among them, CYP1A and 1B enzymes bioactivate polycyclic arylamines, polyaromatic hydrocarbons and aflatoxin B1 (Gonzalez and Gelboin, 1994). These enzymes are regarded as the targets for blocking tumor initiation and specific CYP inhibitors or inactivators could be beneficial for preventing tumor formation Guengerich and Shimada, 1991, Shimada et al., 1996, Shimada et al., 1989. It is likely that constituents in herbal preparations may be substrates, inhibitors, or inducers of CYPs (Zhou et al., 2003), and have an impact on the pharmacokinetics of any coadministered drugs metabolized by this system.
Herbal/dietary constituents may be metabolized by CYPs to nontoxic metabolites and excreted, but the formation of toxic metabolites is possible (Yang et al., 1992). In addition, the inhibition of CYPs by herbal constituents may decrease the formation of toxic metabolites and thus inhibit carcinogenesis, as CYPs play an important role in procarcinogen activation. In some cases, the formation of a reactive intermediate by CYP may also lead to the inactivation of the enzyme. CYP substrates, which are metabolized to reactive intermediates that inactivate the enzyme, are classified as mechanism-based inhibitors. Thus, the inhibition of CYP-mediated carcinogen activation by herbal/dietary constituents has been extensively studied.
The mechanism for herbal toxicity remains elusive, but there are accumulating data suggesting the role of the formation of reactive metabolites/intermediates through the bioactivation of major herbal constituents in herbal toxicity and carcinogenicity. It has been hypothesized that the resultant reactive metabolites following herbal bioactivation covalently bind to cellular proteins and DNA, leading to toxicity via multiple mechanisms such as direct cytotoxicity, oncogene activation, and hypersensitivity reactions. This review highlights the role of formation of reactive intermediates via bioactivation in herbal toxicity using aristolochic acids, pulegone and germander as examples. In addition, mechanism-based inhibition of CYPs by many herbal constituents and the clinical and toxicological relevance are discussed.
Herbal bioactivation: Formation of toxic metabolites
Aristolochic acid nephropathy
Aristolochic acids (AAs), a family of structurally related nitrophenanthrene carboxylic acids, are primarily from herbal medicines such as Aristolochia spp. (e.g. A. fangchi (Guang Fangji), A. clematits, and A. manshuriensis) (Ioset et al., 2003). The predominant AAs are AAI (8-methoxy-6-nitro-phenanthro-(3,4-d)-1,3-dioxolo-5-carboxylic acid) and AAII (6-nitro-phenanthro-(3,4-d)-1,3-dioxolo-5-carboxylic acid). The consumption of herbs containing AAs has been associated with severe nephropathy which is characterized by unique renal fibrosis and development of urothelial cancer Abt et al., 1995, Arlt et al., 2002, Cosyns, 2003, Lord et al., 2001, Nortier and Vanherweghem, 2002, Schwetz, 2001, Zhu, 2002. Approximately 200 cases of AA nephropathy have been reported worldwide, with many of them needing renal replacement therapy (Arlt et al., 2002). A large cohort of these patients with AA nephropathy have developed urothelial cancer Cosyns, 2003, Nortier and Vanherweghem, 2002.
As alkaloids, both AAI and AAII underwent reduction of the nitro group catalyzed by oxidative enzymes to reactive cyclic nitrenium ions (Fig. 1) Schmeiser et al., 1997, Stiborova et al., 2001a, Stiborova et al., 2001b, Stiborova et al., 2002. Hepatic microsomal CYP1A1/2, NADPH:CYP reductase, DT-diaphorase, cyclooxydase-1 and other peroxidases have been found to catalyze the reaction. Addition of inhibitors or inducers of CYP1A1/2 was found to decrease or increase the formation of DNA adducts Stiborova et al., 2001a, Stiborova et al., 2001b. The resultant reactive cyclic nitrenium ions are capable of reacting with DNA and/or proteins, leading to adduct formation. For example, the DNA adducts (e.g. 7-(deoxyadenosin-N6-yl)aristolactam I or II and 7(deoxyguanosin-N2-yl)aristolactam I or II) have been detected in kidney and ureter tissues of patients taking herbs containing AAs, several months or even years after cessation of the herbal consumption Pfau et al., 1990, Schmeiser et al., 1996, Stiborova et al., 1999. The former was the most predominant DNA adduct in human and rat tissues Pfau et al., 1990, Schmeiser et al., 1996, Stiborova et al., 1999.
Fig. 1.
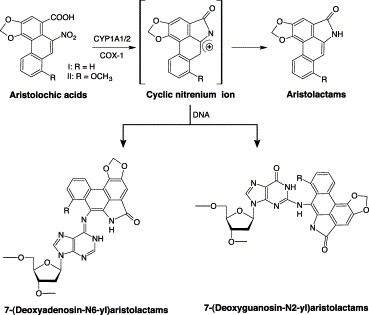
Proposed bioactivation of aristolochic acids.
Studies in the mouse and rat indicated that H-ras protooncogenes were activated with high incidence of an AT→TA transversion mutation in codon 61 of DNA from AAI-induced tumors Schmeiser et al., 1990, Schmeiser et al., 1991. This is consistent with the identification of an N6-deoxyadenosine-AAI adduct formed in vitro and in patients. All these results indicated that deoxyadenosine adduct formation was a critical step in tumor initiation by AAs. A recent study using the λ/lacZ transgenic mice has shown that administration of AAs caused selective AT→TA mutations (Kohara et al., 2002), which is in agreement with the extensive 7-(deoxyadenosin-N6-yl)aristolactam I adduct formation in rats and humans. In rats treated with a single dose of AAI, the 7(deoxyguanosin-N2-yl)aristolactam I adduct was removed continuously from the DNA, whereas the 7-(deoxyadenosin-N6-yl)aristolactam I adduct persisted without proper repair of the DNA (Fernando et al., 1993), leading to mutations and oncogene activation.
Pulegone toxicity
Pulegone is a monoterpene ketone present in essential oils from many mint species (e.g. Hedeoma pulegoides and Mentha pulegium, both called pennyroyal) Budavari, 1996, Grundschober, 1979. Pennyroyal oil has been used as a flavoring agent in foods and beverages, a fragrant constituent and a flea repellent (Hall and Oser, 1965). Pennyroyal has also been used to induce menstruation and abortion. However, pennyroyal oil, especially at high doses, has been reported to cause hepatic failure, central nervous system toxicity, gastritis, renal and pulmonary toxicity, and death (Anderson et al., 1996). Pulegone, constituting >80% of the terpenes in pennyroyal oils, was found to be hepatotoxic and pneumotoxic in mice (Gordon et al., 1982).
The metabolites of pulegone have been considered to be responsible for its toxicity (Gordon et al., 1987). Both menthofuran and p-cresol were potential candidates. Menthofuran is a naturally occurring hepatotoxin (Thomassen et al., 1992) and p-cresol is a known glutathione depletory and toxin (Thompson et al., 1994). In vitro studies have demonstrated CYP-mediated conversion of pulegone to menthofuran which was then bioactivated to reactive γ-ketoenal Gordon et al., 1987, Thomassen et al., 1991. γ-Ketoenal was capable of covalently binding to cellular proteins and caused hepatic injury (Thomassen et al., 1992). The γ-ketoenal can be converted to mintlactones, as well as other reactive intermediates in CYP-mediated oxidation of menthofuran.
The in vivo metabolism of pulegone is very complicated, with about 14 Phase I and 10 Phase II metabolites being identified and characterized in rats Chen et al., 2001, Madyastha and Raj, 1993, McClanahan et al., 1989, Moorthy et al., 1989, Thomassen et al., 1991. These metabolites accounted for only 3% of total radioactivity excreted in bile, and their structures could not be established solely by mass spectral analysis. The metabolism of pulegone in rats involves three major metabolic pathways: a) conjugation by glucuronic acid following C-5 or methyl group hydroxylation or by GSH; with the conjugates being further metabolized; b) the formation of menthofuran; and c) the formation of piperitenone beginning with 5-hydroxylation followed by dehydration (Fig. 2) Chen et al., 2001, Madyastha and Gaikwad, 1999, Madyastha and Raj, 1992, Madyastha and Raj, 1993. The CYP system plays an important role in the metabolism of R-(+)-pulegone Madyastha and Gaikwad, 1999, Madyastha and Raj, 1992. Most of the metabolites of pulegone are derived from menthofuran and piperitenone Madyastha and Gaikwad, 1999, Madyastha and Raj, 1992, Madyastha and Raj, 1993. Studies carried out in vivo have shown 4-methyl-2-cyclohexenone as one of the metabolites of R-(+)-pulegone (Madyastha and Raj, 1993), and conversion of 4-methyl-2-cyclohexenone to p-cresol has been demonstrated in vitro Madyastha and Raj, 1990, Madyastha and Raj, 1993, Madyastha and Raj, 2002.
Fig. 2.
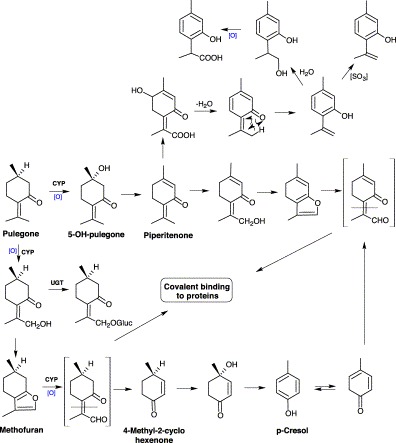
Proposed bioactivation of pulegone.
Germander hepatotoxicity
Germander (Teuchrium chamaedrys), a diterpenoid-containing herbal medicine, was traditionally used as a folk medicine for its alleged choleretic and antiseptic effects. In 1991, germander teas or capsules were promoted as an adjuvant to slimming diets. However, more than 30 cases of hepatotoxicity (mainly cytolytic hepatitis, chronic hepatitis or cirrhosis) after consumption of germander have been reported mainly in France, including cases with positive rechallenge Ben Yahia et al., 1993, Castot and Larrey, 1992, Dao et al., 1993, Laliberte and Villeneuve, 1996, Larrey et al., 1992, Mattei et al., 1992, and a fatality due to fulminant hepatic necrosis (Mostefa-Kara et al., 1992). This medicinal plant has been prohibited in France since 1992.
Germander caused hepatotoxicity in animals Kouzi et al., 1994, Loeper et al., 1994 and humans (Laliberte and Villeneuve, 1996). Germander contains saponins, glycosides, flavonoids and furano neoclerodane diterpenoids (Piozzi et al., 1987). The hepatotoxicity has been ascribed to the diterpenoids including teucrin A and teuchamaedryn A. The furan ring of these diterpenoids is oxidized by CYP3A4 to reactive epoxide which reacts with proteins such as CYP3A and epoxide hydrolase (Fig. 3) Fau et al., 1997, Lekehal et al., 1996, Loeper et al., 2001, Loeper et al., 1994. The reactive epoxide led to mitochondrial permeability transition, caspase activation, and apoptosis in rat and mouse hepatocytes Fau et al., 1997, Lekehal et al., 1996, Loeper et al., 1994. Teucrin A covalently bound to rat hepatocyte proteins, and the furano diterpenoid fraction decreased cell GSH and cytoskeleton-associated protein thiols, but increased cytosolic levels of Ca2+ Fau et al., 1997, Lekehal et al., 1996. All these events led to the formation of plasma membrane blebs, DNA fragmentation, and cell apoptosis.
Fig. 3.
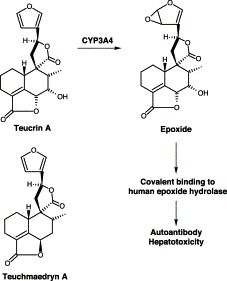
Proposed bioactivation of teucrin A from germander.
The diterpenoid-mediated cytotoxicity was modulated by many factors such as CYP3A inducers or inhibitors. Pretreatment of male rats with troleandomycin (an inhibitor of CYP3A) reduced cellular GSH depletion and cytotoxicity, whereas dexamethasone treatment (an inducer of CYP3A) had opposite effects (Lekehal et al., 1996). Feeding male rats with a sulfur amino acid-deficient diet decreased cellular GSH level and enhanced cytotoxicity, whereas supplementation of the standard diet with cystine had opposite effects (Lekehal et al., 1996). Female rat hepatocytes exhibited little toxicity as they expressed low levels of CYP3A, unless the animals were treated with dexamethasone (Lekehal et al., 1996). Furthermore, the hepatotoxicity in mice was attenuated by pretreatment with butylated hydroxyanisole or clofibrate (two inducers of microsomal epoxide hydrolase) and markedly increased by phorone-induced GSH depletion (Loeper et al., 1994). Pretreatment of mice with either 3-methylcholanthrene or phenobarbital had no effect (Loeper et al., 1994). In addition, genetic factors may be important for the development of germander-induced hepatotoxicity. For example, polymorphism of CYP3A and major histocompability complex molecule can affect the formation of toxic metabolite and antigen presenting to T helper cells, respectively.
Epoxide hydrolase is a plasma membrane-located Phase II microsomal enzyme. It is catalytically competent and may participate in the inactivation of reactive epoxides arising from teucrin A. Thus, it may act as target for reactive metabolites from teucrin A. When incubated with teucrin A, epoxide hydrolase was inactivated in a time-dependent manner, suggesting that a reactive oxide derived from teucrin A could alkylate and inactivate the enzyme (De Berardinis et al., 2000). Interestingly, anti-microsomal epoxide hydrolase autoantibodies were detected in the sera of patients taking germander teas for a long period of time De Berardinis et al., 2000, Loeper et al., 2001, Polymeros et al., 2002. Dose-dependent immunoprecipitation of human epoxide hydrolase by these human sera confirmed human epoxide hydrolase as the autoantigenic target (De Berardinis et al., 2000). It appeared that both direct toxicity and secondary immune reactions were involved in germander-induced heaptotoxicity. The covalent binding of epoxide hydrolase on the outer surface of human hepatocytes by teucrin A might trigger immune responses and induced the formation of autoantibody, leading to cell dealth.
Mechanism-based herbal inhibitors of cytochrome P450s
Many herbal constituents have been found to inhibit various CYPs (Zhou et al., 2003). The nature of inhibition may be competitive, non-competitive, or mechanism-based. The latter is characterized by NADPH-, time- and concentration-dependent enzyme inactivation, occurring when some herbal constituents are converted by CYPs to reactive metabolites that are capable of irreversibly binding to CYPs Ortiz de Montellano and Correia, 1995, Osawa and Pohl, 1989, Silverman, 1998. Mechanism-based inhibitors require at least one cycle of the CYP catalytic process to form reactive metabolites. The resultant metabolites lead to CYP inactivation by chemical modification of the heme, the apoprotein, or both as a result of covalent binding of modified heme to the apoprotein.
Capsaicin
Capsaicin (trans-8-methyl-N-vanillyl-6-nonenamide) is a major pungent phenolic constituent present in a variety of capsicum fruits such as hot peppers, and thus represents an important ingredient of the majority of spicy foods Park et al., 1998, Surh and Lee, 1995. It has shown a wide range of pharmacological properties, including pain-killing, antigenotoxic, antimutagenic, and anticarcinogenic effects Surh et al., 1998, Surh and Lee, 1995. Capsaicin has been used to treat various peripheral painful conditions such as rheumatoid arthritis and diabetic neuropathy (Surh and Lee, 1995).However, capsaicin has been reported to be a tumor promoter, carcinogen and potential mutagen Agarwal et al., 1986, Lopez-Carillo et al., 1994, Toth and Gannett, 1992, Ward and Lopez-Carrillo, 1999. It caused strand scission in calf thymus and plasmid DNA in the presence of Cu(II). This breakage is mediated by reactive oxygen species arising from capsaicin, especially the hydroxyl radical Singh et al., 2001a, Singh et al., 2001b.
Capsaicin undergoes bioactivation by CYP2E1 to reactive species (Gannett et al., 1990). The major bioactivation pathways include: a) epoxidation of the vanillyl ring moiety to produce an arene oxide; b) one-electron oxidation of the ring hydroxyl group to a phenoxy radical; and c) O-demethylation at the aromatic ring and subsequent oxidation of the resulting catechol metabolite to semiquinone and quinone derivatives (Fig. 4) . The resultant reactive species are capable of binding covalently to the active site of CYP2E1 as well as DNA (Surh and Lee, 1995). The interaction with target cell DNA would trigger mutagenicity and malignant transformation. However, metabolism of capsaicinoids by CYPs may also represent a detoxification process (in contrast to bioactivation), resulting in a reduction in cytotoxicity (Reilly et al., 2003). For example, a novel dehydrogenation metabolic pathway of capsaicin yields macrocyclic, diene and imide metabolites (Reilly et al., 2003).
Fig. 4.
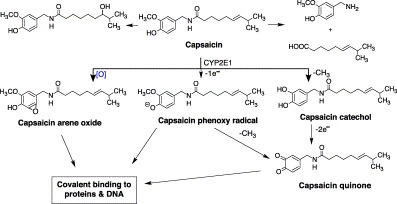
Proposed bioactivation of capsaicin.
Furanocoumarins (bergamottin and 8-methoxypsoralen)
Bergamottin is a major furanocoumarin in grapefruit juice. It reversibly inhibited the activities of CYP1A2, 2A6, 2C9, 2C19, 2D6, 2E1, and 3A4 in human liver microsomes (He et al., 1998). It also inactivated CYP3A4 following metabolic activation in a time- and concentration-dependent manner, with kinact and KI values of 0.3 min−1 and 7.7 μM, respectively (He et al., 1998). The loss of catalytic activity exhibited pseudo-first-order kinetics. During bergamottin-induced inactivation, CYP3A4 retained more than 90% of the heme, but 50% of the apoprotein in the inactivated CYP3A4 could not be recovered. This suggests that the inactivation may involve apoprotein modification in the active site of the enzyme instead of heme adduct formation or heme fragmentation (He et al., 1998). The reactive metabolites of bergamottin remain unknown, but it may undergo oxidation to form a reactive furanoepoxide that covalently binds to CYP3A4 (Fig. 5) .
Fig. 5.
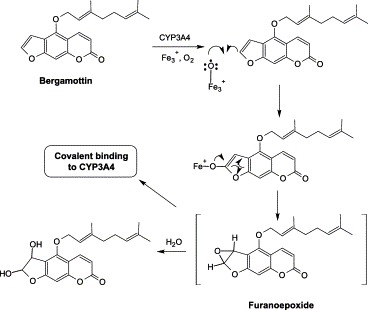
Proposed bioactivation of bergamottin.
8-Methoxypsoralen is a natural furanocoumarin rich in many foodstuffs such as parsnips and parsley. It is used in combination with long wavelength ultraviolet light to treat psoriasis, vitiligo and T cell lymphoma Anderson and Voorhees, 1980, Edelson et al., 1987, Parrish et al., 1974. The usefulness of 8-methoxypsoralen in treating these diseases resides in its ability to be photoactivated to a species capable of binding covalently to nucleic acids and lymphocytes, and thus inhibiting DNA synthesis and cellular proliferation. Early studies indicated that 8-methoxypsoralen was a potent mechanism-based inhibitor of CYPs in rodents Fouin-Fortunet et al., 1986, Labbe et al., 1989, Letteron et al., 1986, Mays et al., 1990 and humans (Tinel et al., 1987). Further studies found that 8-methoxypsoralen was a potent mechanism-based inhibitor of CYP2A6, with KI of 0.8–1.9 μM and kinact of 1–2 min−1 in a reconstituted and native liver microsomes Koenigs et al., 1997, Koenigs and Trager, 1998a, Koenigs and Trager, 1998b, Maenpaa et al., 1993. 8-Methoxypsoralen was also a potent inactivator of CYP2B1, with a KI of 2.9 μM and kinact of 0.34 min−1 Koenigs and Trager, 1998a, Koenigs and Trager, 1998b. However, the reactive species of 8-methoxypsoralen remains unknown, but it is suggested that 8-methoxypsoralen as a furanocoumarin undergoes CYP2A6/2B1-mediated oxidation to form a furanoepoxide (Fig. 6) . The resultant epoxide is then converted to dihydrofuranocoumarin products following hydrolytic attack or direct attack by exogenous nucleophiles Koenigs and Trager, 1998a, Koenigs and Trager, 1998b.
Fig. 6.
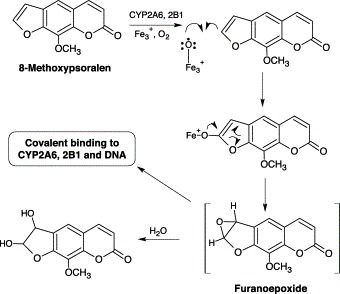
Proposed bioactivation of 8-methoxypsoralen.
Glabridin
The dried roots of the licorice (Glycyrrhiza glabra) have been consumed for the past 6000 years and are used as flavoring and sweating agents, as demulcents and expectorants in the Western world and as antiallergic and anti inflammatory agents in Asian countries including Japan and China (Chandler, 1985). Licorice contains glycyrrhizin (glycyrrhizic acid, a glycoside which is 50 times sweeter than sugar), oleane triterpenoids, glucose, ammonia, polyphenols, flavonoids, and sucrose Hatano et al., 1991a, Hatano et al., 1991b. Flavanoid components of licorice root have been shown to have antitumorigenic, antimicrobial, antiviral, anti inflammatory, estrogen-like, and antioxidative activity Fujisawa et al., 2000, Fukai et al., 2002, Shibata, 2000, Tamir et al., 2001. Licorice root extract, as well as its major flavanoid, the isoflavan glabridin, are potent antioxidants against LDL oxidation in mice and humans (Rosenblat et al., 1999). Glabridin was also shown to inhibit the activity of macrophage NADPH-oxidase presumably by inhibiting protein kinase C and serotonin re-uptake Ofir et al., 2003, Rosenblat et al., 1999. In addition, glycyrrhizin was recently found to be a potent inhibitor of the SARS-associated coronavirus in vitro (Cinatl et al., 2003). However, licorice may cause hypermineralocorticoidism (Nobata et al., 2001), arrhythmia (Bocker and Breithardt, 1991), pseudoaldosteronism (Ferrari et al., 2001), and hypertension (Astrup, 2001). These toxicities have been ascribed to the inhibitory activity of glycyrrhizin and glycyrrhetic acid from licorice on 11-hydroxy-steroid dehydrogenase (Ferrari et al., 2001).
An in vitro study indicated that glabridin inactivated CYP3A4 and 2B6 in a time- and concentration-dependent manner, but CYP2C9 was competitively inhibited (Kent et al., 2002). Metabolism of glabridin by CYP3A4 resulted in the destruction of the heme moiety, as the loss in activity with different concentrations of glabridin correlated well with the loss in the CYP3A4-reduced carbon oxide spectrum and detectable heme at 405 nm (Kent et al., 2002). No other peaks indicative of glabridin-alkylated heme eluting after the native heme were detected at 405 nm. Presumably glabridin undergoes metabolism, forming reactive intermediates causing heme fragmentation. Incubations with 2,4-dimethylglabridin did not lead to a loss in the enzymatic activity of CYP3A4. Incubation of glabridin with CYP3A4 and NADPH resulted in a metabolites, suggesting that the two hydroxyl groups on the 2′ and 4′ position of the flavonoid B ring of glabridin are required for CYP3A4 inactivation (Fig. 7) (Kent et al., 2002). Indeed, these two hydroxyl groups are also believed to be essential for its antioxidative activity (Belinky et al., 1998).
Fig. 7.
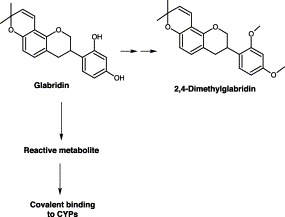
Chemical structure of glabridin.
Isothiocyanates
Isothiocyanates are released upon chewing or maceration of cruciferous vegetables, such as cabbage, cauliflower, and broccoli where they occur as thioglucoside conjugates called glucosinolates (Fenwick et al., 1983). The enzyme myrosinase is released at the same time and hydrolyzes the glucosinolate with rearrangement of intermediates producing the isothiocyanates, hydrogen sulfate, and glucose as the major products. Typical isothiocyanates include benzyl isothiocyanate (BITC) and phenethyl isothiocyanate (PEITC). These compounds have been shown to be potent modulators of CYPs in vitro and in vivo.
Isothiocyanates inhibited CYP1A1, 1A2, 2A1, 2B1, and 2E1 which are involved in carcinogen activation (Yang et al., 1994). BITC and PEITC have been shown to inhibit N-nitrosodimethylamine demethylation activity. BITC is a potent mechanism-based inactivator of rat CYP1A1, 1A2, 2B1, and 2E1, as well as human CYP2B6 and 2D6 Goosen et al., 2000, Moreno et al., 1999. The KI for CYP1A1, 1A2, 2B1, and 2E1 were 35, 28, 16, and 18 μM, respectively; and the corresponding kinact values were 0.26, 0.09, 0.18, and 0.05 min−1, respectively Goosen et al., 2001, Moreno et al., 1999. Human CYP2C9 and rat CYP3A2 were not inactivated by BITC. BITC was also a potent inactivator of CYP2B1, as indicated by a partition ratio of approximately 11:1 (Goosen et al., 2001).
The mechanism of CYP inhibition by isothiocyanates involves both competitive and mechanims-based inhibition. The competitive inhibition involves competition with substrates at the active site of CYP Ishizaki et al., 1990, Smith et al., 1993, while mechanism-based inhibition would involve the metabolic activation of isothiocyanates Goosen et al., 2000, Ishizaki et al., 1990, Lee, 1996, Moreno et al., 1999, Smith et al., 1996. The metabolic activation of isothiocyanates results in reactive intermediates that could ultimately bind to the heme moiety or apoprotein, thereby inactivating the enzyme. For example, BITC was metabolized to a reactive benzyl isocyanate intermediate that is capable of binding covalently to CYP apoprotein or hydrolyzed to form benzylamine (Fig. 8) (Goosen et al., 2001). BITC may be oxidized at the α-carbon to benzaldehyde, instead of being desulfurated to benzyl isocyanate. Alternatively, benzylamine could be deaminated to give benzaldehyde and ammonia. The subsequent oxidation of benzaldehyde would yield benzoic acid. The formation of benzoic acid was also observed in dogs, where administration of BITC resulted in the excretion of hippuric acid, the glycine conjugate of benzoic acid (Brüsewitz et al., 1997). In humans and rats, BITC is metabolized through conjugation with GSH and finally excreted as mercapturic acid. In addition, oxidative desulfuration of the isothiocyanate could also release elemental sulfur, which could further contribute to enzymatic modification and possibly inactivation, as in the case of parathion (Butler and Murray, 1993).
Fig. 8.
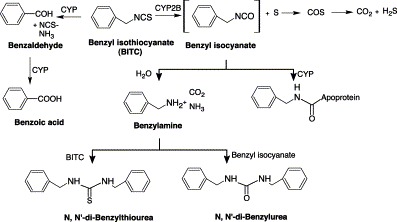
Proposed bioactivation of benzyl isothiocyanate (BITC).
The in vivo administration of isothiocyanates to rodents also modulate CYP content and activity Guo et al., 1992, Smith et al., 1993. The effects appeared to depend on the experimental conditions, the isothiocyanate used and treatment regimen, the target tissue, and the specific CYP evaluated (Zhang and Talalay, 1994). For example, acute administration of PEITC to rats decreased liver CYP2E1 activity, whereas CYP2B1 activity and content increased approximately 10-fold, but without any significant effect on lung CYP1A2 and 2B1 activities (Guo et al., 1992). However, chronic administration of PEITC induced both CYP2B1 and 2E1 with related increases in carcinogenesis (Smith et al., 1993).
Kavalactones (methysticin and dihydromethysticin)
Kava (Piper methysticum, found in Polynesia, Melanesia, and Micronesia) is an effective herbal medicine for the therapy of anxiety and insomnia Pittler and Ernst, 2000, Rouse, 1998, Singh and Singh, 2002, Volz and Kieser, 1997. Clinical studies have shown that kava and kavalactones are effective in the treatment of anxiety at subclinical and clinical levels, anxiety associated with menopause and various other medical conditions Bilia et al., 2002, Pittler and Ernst, 2000, Stevinson et al., 2002, Volz and Kieser, 1997, Wheatley, 2001 The major constituents of kava are pharmacologically active kavalactones which are responsible for about 95% of the total activity of kava (Singh and Singh, 2002). Yangonin, desmethoxyyangonin, methysticin, 7,8-dihydromethysticin, kawain, and 7,8-dihydrokawain are present in the highest levels of kavalactones, accounting for approximately 96% of lipidic extracts (Lebot and Lévesque, 1989).
Both methysticin and dihydromethysticin (Fig. 9) were mechanism-based inhibitors of CYPs, as indicated by the unidentified metabolic intermediate complexes at 455 nm when incubation with human liver microsomes (Mathews et al., 2002). This 455-nm absorbing complex was both NADPH- and time-dependent, with absorption increasing with increasing incubation time. Methysticin analogs in kava extract have been shown to inhibit multiple CYPs after metabolic activation through the formation of metabolic intermediate complexes Murray and Reidy, 1989, Murray et al., 1983. Preincubation of whole kava extract with human liver microsomes inhibited CYP1A2, 2C9, 2C19, 2D6, 3A4, and 4A9/11; whereas CYP2A6, 2C8, and 2E1 were unaffected (Mathews et al., 2002). Various kavalactones inhibited different CYPs. Kava extract and kavalactones inhibited CYP3A4 in cDNA-expressed microsomes (Unger et al., 2002).
Fig. 9.
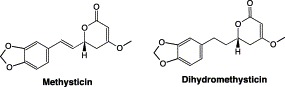
Chemical structures of methysticin and dihydromethysticin from kava.
Oleuropein
The Mediterranean diet has been associated with longevity and low rates of coronary artery disease and cancers despite its high dietary fat content Braga et al., 1998, Owen et al., 2000, Roche et al., 1998. The Mediterranean diet includes the consumption of large amounts of olive oil. Olive oil is a source of at least 30 phenolic compounds (total: 60–1000 mg/kg) (Tuck and Hayball, 2002). The beneficial effect of olive oil has been mainly attributable to oleuropein and its two major metabolites, hydroxytyrosol and tyrosol. Oleuropein is the major phenolic compound in olive fruit, which can be as much as 14% in dried fruit. It is an ester consisting of hydroxytyrosol and elenolic acid which are potent antioxidant agents (Briante et al., 2001).
The phenolic components in olive oil are believed to play a vital role in the prevention of coronary artery disease and atherosclerosis. These compounds, in particular hydroxytyrosol, are potent antioxidants and radical scavengers Tuck and Hayball, 2002, Visiol and Galli, 2002, Visioli and Galli, 1994, Visioli et al., 2002. They exert protective effects on cardiovascular system mainly by inhibiting platelet aggregation and endothelial activation (Carluccio et al., 2003), modulating arachidonic acid metabolism (Kohyama et al., 1997) and preventing the oxidation of low density lipids Visiol and Galli, 2002, Visioli et al., 1995, Visioli et al., 2002. It is presumed that hydroxytyrosol as water and lipid soluble molecule can penetrate in cell membranes and thus inhibit the production of leukotriene B4 effectively from endogenous arachidonic acid (Kohyama et al., 1997).
In vitro studies indicated that oleuropein inactivated androstenedione 6β-hydroxylase (CYP3A4) activity in human liver microsomes, with kinact and KI of 0.09 min−1 and 22.2 μM, respectively Stupans et al., 2001, Stupans et al., 2000. The reactive metabolites of oleuropein remain undetermined, but oleuropein might undergo CYP3A-mediated oxidation to unidentified metabolite(s) capable of binding and inactivating CYP3A4 (Fig. 10) . In addition, hydroxytyrosol and tyrosol and/or their metabolites may be involved. Metabolism studies have indicated that hydroxytyosol is excreted into the urine unchanged and as its glucuronide and sulfate conjugates (Fig. 10) Miro-Casas et al., 2003, Tsarbopoulos et al., 2003, Tuck and Hayball, 2002. Hydroxytyrosol can also be sequentially oxidized to 3,4-dihydroxyphenylacetic acid and 3,4-dihydroxyphenylacetaldehyde by alcohol and aldehyde dehydrogenase, or methylated by catechol methyltransferase to homovanillic acid and then oxidized to homovanillic alcohol (D'Angelo et al., 2001).
Fig. 10.
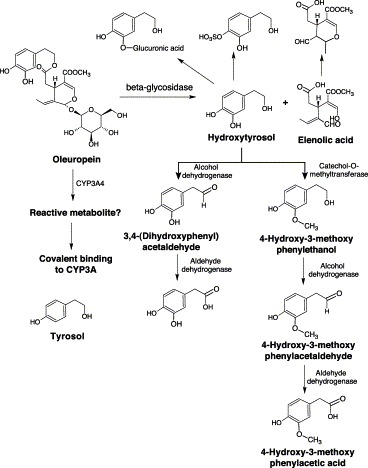
Metabolism of oleuropein.
Organosulfur compounds
Garlic is frequently used as a flavor-enhancing ingredient in food preparation and healthy supplement in the practice of folk medicine. Organosulfur compounds are the major active components of garlic. Garlic also contains numerous flavonoids/isoflavonoids (such as nobiletin, quercetin, rutin, and tangeretin), polysaccharides, prostaglandins, saponins, and terpenes (such as citral, geraniol, linalool, α- and β-phellandrene) Dausch and Nixon, 1990, Singh et al., 2001a, Singh et al., 2001b. Alliin (S-allylcysteine sulfoxide) is one major component of garlic, which is converted by alliinase to allicin. Allicin is further transformed to other garlic compounds, including diallyl sulfide. Organosulfur compounds have been reported to have hypolipidemic, antiplatelet, immune-enhancing, anticancer, chemopreventive, hepatoprotective, antihypertension, and procirculatory effects Rahman, 2001, Spigelski and Jones, 2001.
Organosulfar compounds modulated CYPs in vitro and in vivo (Brady et al., 1991). For example, extracts of fresh garlic, and samples of garlic oil, freeze dried garlic, and aged garlic inhibited human CYP2C9, 2C19, 3A4, 3A5 and 3A7 activity, whereas the CYP2D6 activity was unaltered (Foster et al., 2001). The organosulfur compounds 4,4′-dipyridyl disulfide, di-n-propyl disulfide and diallyl sulfide were also potent competitive inhibitors of coumarin 7-hydroxylase (CYP2A6), with a K i value of 0.06, 1.7 and 2.1 μM respectively (Fujita and Kamataki, 2001). In vivo studies in the mouse indicated that garlic administration increased CYP2E1 and 1A2 levels (Kishimoto et al., 1999), whereas the administration of garlic constituents in the rat decreased the CYP2E1 activity and/or protein level, but increased or did not alter the CYP1A, CYP2B and CYP3A activities Dalvi, 1992, Haber et al., 1995, Haber et al., 1994. A single dose of garlic oil in the rat resulted in a significant inhibition of hepatic CYP-catalyzed reactions including aminopyrine N-demethylase (CYP2C) and aniline hydroxylase (CYP2E1) activity, but administration of garlic for 5 days led to a significant increase in these hepatic CYP activities (Fitzsimmons and Collins, 1997). In addition, administration of garlic oil to healthy subjects for 28 days reduced CYP2E1 activity by 39% when chlorzoxazone was used as a probe compound (Gurley et al., 2002).
Diallyl sulfide is converted by CYP2E1 to diallyl sulfoxide and sequentially to diallyl sulfone (Fig. 11) (Teyssier et al., 1999). All these compounds are competitive inhibitors of CYP2E1 (Brady et al., 1991). A single oral dose of diallyl sulfone inactivated CYP2E1- and 1A-dependent activities in mouse liver microsomes (Lin et al., 1996). Diallyl sulfone is also a mechanism-based inhibitor of CYP2E1, involving the formation of a reactive metabolite which forms a CYP complex leading to autocatalytic destruction Brady et al., 1991, Jin and Baillie, 1997, Premdas et al., 2000. A recent study has demonstrated that diallyl sulfone was oxidized at one of the terminal double bonds, yielding the monoallyleopoxide that inactivated CYP2E1 (Fig. 11) (Premdas et al., 2000). This monoallyleopoxide was conjugated by GSH. It appeared that the reactive intermediate from diallyl sulfone might initially affect the heme and/or apoprotein.
Fig. 11.
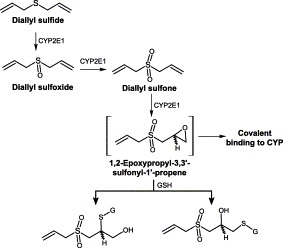
Metabolism of diallyl sulfide by CYP2E1.
Resveratrol
Resveratrol is a polyphenolic phytoalexin found in red wine, grapes and peanuts. It has been reported to have antioxidant, cardioprotective, antimutagenic, antiinflammatory, chemopreventive and antiplatelet effects Aziz et al., 2003, Cal et al., 2003, Gusman et al., 2001, Kimura, 2003, Orallo et al., 2002, Roemer and Mahyar-Roemer, 2002. Resveratrol has also been reported to possess substantial cytotoxic activities in different tumour cell lines El-Mowafy and Alkhalaf, 2003, Ferry-Dumazet et al., 2002, Kuo et al., 2002, Roman et al., 2002. The cytotoxicity may be ascribed to suppressed activation of NF-κB and cell proliferation, inhibition of DNA synthesis (Kuwajerwala et al., 2002), and induction of G2 arrest and apoptosis of tumour cells Estrov et al., 2003, Liang et al., 2003, Niles et al., 2003, Pozo-Guisado et al., 2002, Roman et al., 2002. In addition, resveratrol is able to activate estrogen receptor Bhat et al., 2001, Klinge et al., 2003, Levenson et al., 2003, Mueller et al., 2003, Ratna and Simonelli, 2002, inhibit eicosanoid synthesis and cyclooxygenase-1 and 2 Johnson and Maddipati, 1998, Martinez and Moreno, 2000, Moreno, 2000, Pace-Asciak et al., 1995, activate adenylyl and guanylyl cyclase El-Mowafy, 2002, El-Mowafy and Alkhalaf, 2003, and enhance the expression and activity of endothelial nitric oxide synthase Hattori et al., 2002, Wallerath et al., 2002.
Resveratrol was shown to be an irreversible (mechanism-based) inhibitor for CYP3A4 and a non-competitive reversible inhibitor for CYP2E1, with IC50 values of 4-150 μM in microsomes from rat liver, human liver or cells containing cDNA-expressed CYPs Chan and Delucchi, 2000, Chan et al., 1998, Piver et al., 2001. In a study using Sf9 insect microsomes containing baculovirus-derived human CYP3A4 and NADPH-cytochrome CYP reductase, resveratrol inactivated CYP3A4 in a time- and NADPH-dependent manner, with kinact and KI of 0.20 min−1 and 20 μM, respectively Chan and Delucchi, 2000, Chan et al., 1998. Resveratrol is an electron-rich molecule with two aromatic benzene rings linked by an ethylene bridge. CYP3A-mediated aromatic hydroxylation and epoxidation of this compound are possible, resulting in a reactive p-benzoquinone methide metabolite which is capable of binding covalently to CYP3A4 and inactivating it (Fig. 12) .
Fig. 12.
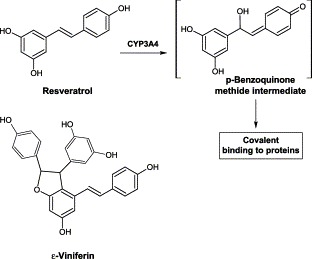
Proposed bioactivation of resveratrol.
Interestingly, ε-viniferin, a dimer of resveratrol, was not a mechanism-based inhibitor of human CYPs (Piver et al., 2003). It showed mixed-type inhibitions for various CYPs with Ki of 0.5–20 μM, except for CYP2E1 (non-competitive). ε-Viniferin was isolated in wine at concentration of 0.5–5 μM (Landrault et al., 2002). It is generated by oxidation from resveratrol, the parent compound and produced by grapevine in response to fungal infection. ε-Viniferin has been shown to have better antifungal and antioxidant activity than resveratrol Baderschneider and Winterhalter, 2000, Bala et al., 2000. ε-Viniferin also had hepatoprotective activity (Oshima et al., 1995).
Mechanism-based inhibition of CYPs and herb-drug interactions
Herbs are often administered in combination with therapeutic drugs, raising the potential of pharmacokinetic and/or pharmacodynamic herb-drug interactions. A number of clinically important herb-drug interactions have been documented and many of them led to altered efficacy and/or adverse events Fugh-Berman, 2000, Fugh-Berman and Ernst, 2001. Pharmacokinetic herb-drug interactions are due to altered absorption, metabolism, distribution and excretion of drugs. Frequently, one of the underlying mechanisms of altered drug concentrations by concomitant herbal medicines is the induction or inhibition of hepatic and intestinal CYPs. CYP3A4 are expressed at high levels in the villus tip of enterocytes, the primary site of absorption for orally administered drugs. Thus, the modulation of intestinal CYP3A represents an important mechanism for the enhanced or reduced bioavailability of coadministered drugs. In particular, the mechanism-based inhibition of CYPs (in particular CYP3A4) by herbal constituents may have important pharmacokinetic implications.
Administration of garlic extract for 3 months in healthy volunteers did not alter the oxidative metabolism of acetaminophen, but caused a slight increase in sulfation (Gwilt et al., 1994). However, a study in mice indicated that diallyl sulfone decreased the plasma levels of oxidative acetaminophen metabolites, but not nonoxidative acetaminophen metabolites (Lin et al., 1996). This is considered to be due to the inhibition of CYP2E1 that is the major enzyme responsible for acetaminophen bioactivation (Manyike et al., 2000). In liver microsomes, diallyl sulfone significantly inhibited the acetaminophen oxidation to N-acetyl-p-benzoquinone imine (the toxic metabolite of acetamoniphen) (Lin et al., 1996). All these results provide an explanation for the protective effect of diallyl sulfone on acetaminophen-induced hepatotoxicity (Lin et al., 1996). When administered orally 1 hr prior to, immediately after, or 20 min after a toxic dose of acetaminophen, diallyl sulfone (25 mg/kg) completely protected mice from development of hepatotoxicity (Lin et al., 1996).
Case reports and pharmacologic studies have indicated a potentiation of the central nervous system effects of benzodiazepines, including alprazolam (Almeida and Grimsley, 1996), in the presence of kava extract and/or kavalactones. Since alprazolam is a substrate of CYP3A4 (Gorski et al., 1999) and kavalactones are potent inhibitors of CYP3A4 (Zou et al., 2002), decreased elimination of alprazolam by kava may contribute to the additive effects reported upon coadministration of kava and alprazolam. Coadministration of kava markedly increased hexobarbital sleep time (Meyer, 1962) and ethanol-induced hypnotic effect (Jamieson and Duffield, 1990) in mice, indicative of inhibition of CYP2C and CYP2E1.
In a crossover study in healthy human volunteers, oral ingestion of aqueous licorice extract for 7 days did not significantly alter the pharmacokinetic parameters and sedative effects of midazolam (Shon et al., 2001). Midazolam is a typical substrate of CYP3A4 (Gorski et al., 1994). The discrepancy between in vitro and animal and human studies may reflect the importance of herbal dosing and regimen in the modulation of CYPs.
Grapefruit juice has been found to significantly increase oral bioavailability of most dihydropyridines (e.g. felodipine), terfenadine, saquinavir, cyclosporine, midazolam, triazolam and verapamil Bailey et al., 2000, Bailey et al., 1998, Bailey et al., 1991, Ducharme et al., 1995, He et al., 1998, Kane and Lipsky, 2000, Mohri and Uesawa, 2001, Yee et al., 1995. The plasma concentrations or area under the concentration-time curve (AUC) of lovastatin, cisapride and astemizole can also be markedly increased by grapefruit juice Bailey et al., 2000, Bailey et al., 1998. As the duration of effect of grapefruit juice can last 24 hr, repeated consumption of grapejuice can lead to a cumulative increase in the AUC and Cmax of coadministered drugs. The inhibition of CYP3A4 activity with no change of CYP3A4 mRNA and P-glycoprotein is believed to be the primary mechanism Bailey et al., 1998, Kane and Lipsky, 2000. However, the pharmacokinetics of many other drugs were not altered by grapefruit juice. For example, grapefruit juice did not alter the bioavailability of digoxin, diltiazem and amlodipine in human volunteers, and indinavir in HIV-positive patients Becquemont et al., 2001, Sigusch et al., 1994, Vincent et al., 2000. Although these drugs undergo presystemic metabolism, CYP3A4 is a minor contributor. These finding indicate the difficulty in predicting herb-drug interactions.
The clinical outcome of herb-drug interactions depends on factors that are related to coadministered drugs (dose, dosing regimen, administration route, pharmacokinetic and therapeutic range), herbs (species, dose, dosing regimen, and administration route) and patients (genetic polymorphism, age, gender and pathological conditions) (Dresser et al., 2000). Generally, a doubling or more in drug plasma concentration has the potential for enhanced adverse effects. However, less marked changes may still be clinically important for drugs with a steep concentration-response relationship or a narrow therapeutic index.
Mechanism-based inhibition of CYPs and herbal chemoprevention
Chemoprevention refers to the application of natural or synthetic compounds to block, reverse, or prevent the development of invasive cancers Shureiqi et al., 2000, Sporn and Suh, 2000, Tamimi et al., 2002, Young and Wilson, 2002. Clinical evidence has indicated the feasibility of this approach in reducing the risk of major human cancers. For example, tamoxifen and raloxifene (both oestrogen receptor modulators) reduce the risk of developing breast cancer in women at increased risk Fisher et al., 1998, Salih and Fentiman, 2001. Results with aspirin and non-steroidal anti-inflammatory agents have proved consistently encouraging in epidemiological studies in lowering the incidence of colorectal cancer (Jolly et al., 2002). However, although these compounds have shown a potential to reduce cancer incidence, several problems have been associated with them. These include severe side effects due to long-term use, high costs and non-compliance. Therefore, safer and more selective chemopreventive agents are needed.
A number of naturally occurring products from herbs have shown chemopreventive properties against carcinogenesis using in vitro and animal models Chen et al., 1998, Fukutake et al., 2000, Fukutake et al., 1998, Lahiri-Chatterjee et al., 1999, Zheng et al., 1997. The mechanisms for the chemopreventive effects of herbal preparations are not fully elucidated, but inhibition of activating enzymes and other enzyme systems, protective effects from toxic xenobiotics, beneficial regulation of cell cycles and cellular signalling pathways have all been suggested (Wargovich et al., 2001).
Naturally occurring isothiocyanates have been shown to be potent and selective inhibitors of carcinogenesis induced by a number of chemical carcinogens Boysen et al., 2003, Hecht et al., 2002, Yang et al., 2002. In most studies, the chemopreventative activity of isothiocyanates required administration either before or during exposure to the carcinogen. PEITC inhibits lung carcinogenesis induced by the tobacco-specific nitrosamine 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone, whereas BITC is ineffective in this regard (Staretz and Hecht, 1995). In turn, BITC effectively inhibits benzo(a)pyrene-induced lung tumors in A/J mice, whereas PEITC is ineffective (Lin et al., 1993). These effects might be related to the inhibitory mechanisms and potencies for CYP2B1 and 1A, respectively. Some isothiocyanates such as PEITC are able to induce CYP2B1 and 2E1 following chronic administration, with increased toxicity (Smith et al., 1993). The extensive conjugation of BITC with GSH might also be important for the in vivo effects of BITC. The conjugation with GSH has been shown to decrease potency, and could contribute to mutagenicity (Raulf-Heimsoth and Baur, 1998). It appears that local release of the conjugated product might also contribute to the toxicity of BITC (Hirose et al., 1998).
Epidemiological studies suggest that the consumption of fruit and vegetables decreases the risk for cancer development (Block et al., 1992). The combination of BITC and PEITC has recently also been proposed for the chemoprevention of lung cancer in individuals resistant to smoking cessation Hecht, 1997, Kelloff et al., 1996. These effects appear to be mediated through modulation of both Phase I and Phase II enzymes involved in carcinogen metabolism by isothiocyanates. In particular, the isothiocyanate-mediated inactivation of CYP1A and 2B responsible for carcinogen activation would inhibit carcinogenesis Boysen et al., 2003, Scharf et al., 2003, Zhang and Talalay, 1994. In addition, the induction of detoxifying Phase II enzymes may also contribute to the beneficial effects of isothiocyanates.
Organosulfur compounds from garlic showed chemopreventive effects at several organ sites in rodents after administration of chemical carcinogens, perhaps by inhibiting CYP2E1-mediated carcinogen activation Reddy et al., 1993, Yang et al., 2001. These compounds have also been shown to reduce the incidence of a multitude of chemically induced tumors in animal models. Pretreatment with aqueous garlic extract significantly reduced the frequencies of N-methyl-N′-nitro-N-nitrosoguanidine-induced micronuclei and chromosomal aberrations (Arivazhagan et al., 2001). These compounds have also been shown to reduce toxicity induced by thioacetamide, carbon tetrachloride, N-nitrosodimethylamine- and acetaminophen (all CYP2E1 substrates) in rodents Ramaiah et al., 2001, Wang et al., 2001, Wang et al., 2000. The protective effect was observed when the organosulfur compounds were given before, during or soon after chemical treatment.
Diallyl sulfide also has been shown to induce other CYP and phase II enzymes as well as decrease hepatic catalase activity (Yang et al., 2001). The preventive effects of garlic extract on bromobenzene-induced hepatotoxicity in precision-cut liver slices was related to an elevation of hepatic glutathione content, and a glutathione sparing effect, possibly due to conjugation of organosulphur compounds in garlic extract with toxic bromobenzene metabolites Guyonnet et al., 2001, Wang et al., 1999. Organosulfur compounds also inhibit the formation of DNA adducts in several target tissues. Antiproliferative activity has been described in several tumor cell lines Hirsch et al., 2000, Nakagawa et al., 2001, and may be due to induction of apoptosis and alterations of the cell cycle Frantz et al., 2000, Kwon et al., 2002. However, all of these effects are observed at concentrations much higher than what is normally ingested by humans and clinical trials will be needed to define the effective dose that has no toxicity in humans.
Resveratrol seems to be a promising cancer chemopreventive agent and it has been shown to afford protection against several cancer types in several bioassay systems Aziz et al., 2003, Banerjee et al., 2002, Li et al., 2002. The mechanism for these effects is not completely understood, but multiple effects of resveratrol may be involved. Inhibition of Phase I enzymes and induction of Phase II enzymes will contribute to the chemopreventive activity of resveratrol Dubuisson et al., 2002, Jang et al., 1997. It also has activity in the regulation of multiple cellular events associated with carcinogenesis. However, it is not known whether resveratrol inhibits the catalytic activity of CYP1A1, 1A2, and 1A2 in vivo. The potential in vivo effect of resveratrol on the bioactivation of CYP1 substrates may depend not only on the pharmacokinetics of resveratrol but also the tissue of interest (Schwedhelm et al., 2003). It is known that the expression of many CYPs such as CYP1A1, 1A2, and 1B1 is tissue-dependent (Omiecinski et al., 1999).
Conclusions
Bioactivation of herbal constituents appears a critical step for the toxicity induction of some herbs. The resultant reactive intermediates bind covalently to DNA and proteins, leading to organ toxicity and even carcinogenicity. On the other hand, some herbal/dietary constituents were shown to form reactive intermediates capable of irreversibly inhibiting various CYPs. The resultant metabolites lead to CYP inactivation by chemical modification of the heme, the apoprotein, or both as a result of covalent binding of modified heme to the apoprotein. The mechanism-based inhibition of CYPs may provide an explanation for some reported herb-drug interactions. Naturally occurring compounds that inactivate CYPs may represent a novel type of chemopreventive agents with higher selectivity and lower toxicity compared to synthetic compounds.
Herbal medicines often contain multiple active substances and multiple cellular molecules might be the targets of herbal medicine. The identification of these targets may provide molecular evidence for the herb's pharmacological activity and toxicity. Recently, there is an increasing application of genomic approaches (e.g. high-density microarray analysis) to examine the genomic responses to herbal medicines Afshari et al., 1999, Gohil, 2002, Gohil et al., 2000, Owuor and Kong, 2002, Watanabe et al., 2001, Yang et al., 2003. These approaches can identify a huge number (e.g. 1000–12 000) of genes which are repressed or stimulated by a herbal compound of interest. The genomic approaches may offer a powerful tool for defining and predicting pharmacological activity and toxicity of herbal medicines.
Due to the wide use and easy availability of herbal medicines, herbal toxicity has become an issue of concern. The safety and quality of herbal medicine should be ensured through greater research, pharmacovigilance, greater regulatory control and better communication between patients and health professionals.
Acknowledgements
The authors appreciate the support by the National University of Singapore Academic Research Funds.
References
- Abt A., Oh J.Y., Huntington R.A., Burkhart K.K. Chinese herbal medicine induced acute renal failure. Archives of Internal Medicine. 1995;155(2):211–212. [PubMed] [Google Scholar]
- Afshari C.A., Nuwaysir E.F., Barrett J.C. Application of complementary DNA microarray technology to carcinogen identification, toxicology, and drug safety evaluation. Cancer Research. 1999;59(19):4759–4760. [PubMed] [Google Scholar]
- Agarwal R.C., Wiessler M., Hecker E., Bhide S.V. Tumour-promoting effects of chilli extracts in BALB/c mice. International Journal of Cancer. 1986;38(5):689–695. doi: 10.1002/ijc.2910380512. [DOI] [PubMed] [Google Scholar]
- Almeida J.C., Grimsley E.W. Coma from the health food store: interaction between Kava and alprazolam. Annals of Internal Medicine. 1996;125(11):940–941. doi: 10.7326/0003-4819-125-11-199612010-00023. [DOI] [PubMed] [Google Scholar]
- Anderson I.B., Mullen W.H., Meeker J.E., Khojasteh-Bakht S.C., Oishi S., Nelson S.D., Blanc P.D. Pennyroyal toxicity: measurement of toxic metabolite levels in two cases and review of the literature. Annals of Internal Medicine. 1996;124(8):726–734. doi: 10.7326/0003-4819-124-8-199604150-00004. [DOI] [PubMed] [Google Scholar]
- Anderson T.F., Voorhees J.J. Psoralen photochemotherapy of cutaneous disorders. Annual Review of Pharmacology and Toxicology. 1980;20:235–257. doi: 10.1146/annurev.pa.20.040180.001315. [DOI] [PubMed] [Google Scholar]
- Arivazhagan S., Nagini S., Santhiya S.T., Ramesh A. Protection of N-methyl-N′-nitro-N-nitrosoguanidine-induced in vivo clastogenicity by aqueous garlic extract. Asia Pacific Journal of Clinical Nutrition. 2001;10(3):238–241. doi: 10.1046/j.1440-6047.2001.00249.x. [DOI] [PubMed] [Google Scholar]
- Arlt V.M., Stiborova M., Schmeiser H.H. Aristolochic acid as a probable human cancer hazard in herbal remedies: a review. Mutagenesis. 2002;17(4):265–277. doi: 10.1093/mutage/17.4.265. [DOI] [PubMed] [Google Scholar]
- Astrup A.V. Licorice-induced high blood pressure. Ugeskrift for Laeger. 2001;163(51):7284–7285. [PubMed] [Google Scholar]
- Aziz M.H., Kumar R., Ahmad N. Cancer chemoprevention by resveratrol: in vitro and in vivo studies and the underlying mechanisms. International Journal of Oncology. 2003;23(1):17–28. [PubMed] [Google Scholar]
- Baderschneider B., Winterhalter P. Isolation and characterization of novel stilbene derivatives from Riesling wine. Journal of Agricultural and Food Chemistry. 2000;48(7):2681–2686. doi: 10.1021/jf991348k. [DOI] [PubMed] [Google Scholar]
- Bailey D.G., Dresser G.R., Kreeft J.H., Munoz C., Freeman D.J., Bend J.R. Grapefruit-felodipine interaction: effect of unprocessed fruit and probable active ingredients. Clinical Pharmacology and Therapeutics. 2000;68(5):468–477. doi: 10.1067/mcp.2000.110774. [DOI] [PubMed] [Google Scholar]
- Bailey D.G., Malcolm J., Arnold O., Spence J.D. Grapefruit juice-drug interactions. British Journal of Clinical Pharmacology. 1998;46(2):101–110. doi: 10.1046/j.1365-2125.1998.00764.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey D.G., Spence J.D., Munoz C., Arnold J.M.O. Interaction of citrus juices with felodipine and nifedipine. Lancet. 1991;337:268–269. doi: 10.1016/0140-6736(91)90872-m. [DOI] [PubMed] [Google Scholar]
- Bala A.E.A., Kollman A., Ducrot P.-H., Majira A., Kerhoas L., Leroux P., Delorme R., Einhorn J. Cis ε-viniferin: a new antifungal resveratrol dehydrodimer from Cyphostemma crotalariodes roots. Journal of Phytopathology. 2000;148:29–32. [Google Scholar]
- Banerjee S., Bueso-Ramos C., Aggarwal B.B. Suppression of 7,12-dimethylbenz(a)anthracene-induced mammary carcinogenesis in rats by resveratrol: role of nuclear factor-κB, cyclooxygenase 2, and matrix metalloprotease 9. Cancer Research. 2002;62(17):4945–4954. [PubMed] [Google Scholar]
- Becquemont L., Verstuyft C., Kerb R., Brinkmann U., Lebot M., Jaillon P., Funck-Brentano C. Effect of grapefruit juice on digoxin pharmacokinetics in humans. Clinical Pharmacology and Therapeutics. 2001;70(4):311–316. [PubMed] [Google Scholar]
- Belinky P.A., Aviram M., Mahmood S., Vaya J. Structural aspects of the inhibitory effect of glabridin on LDL oxidation. Free Radical Biology and Medicine. 1998;24(9):1419–1429. doi: 10.1016/s0891-5849(98)00006-9. [DOI] [PubMed] [Google Scholar]
- Ben Yahia M., Mavier P., Metreau J.M., Zafrani E.S., Fabre M., Gatineau-Saillant G., Dhumeaux D., Mallat A. [Chronic active hepatitis and cirrhosis induced by wild germander. 3 cases] Gastroenterology and Clinical Biology. 1993;17(12):959–962. [PubMed] [Google Scholar]
- Bensoussan A., Myers S.P., Drew A.K., Whyte I.M., Dawson A.H. Development of a Chinese herbal medicine toxicology database. Journal of Toxicology and Clinical Toxicology. 2002;40(2):159–167. doi: 10.1081/clt-120004404. [DOI] [PubMed] [Google Scholar]
- Bhat K.P., Lantvit D., Christov K., Mehta R.G., Moon R.C., Pezzuto J.M. Estrogenic and antiestrogenic properties of resveratrol in mammary tumor models. Cancer Research. 2001;61(20):7456–7463. [PubMed] [Google Scholar]
- Bilia A.R., Gallon S., Vincieri F.F. Kava-kava and anxiety: growing knowledge about the efficacy and safety. Life Sciences. 2002;70(22):2581–2597. doi: 10.1016/s0024-3205(02)01555-2. [DOI] [PubMed] [Google Scholar]
- Block G., Patterson B., Subar A. Fruit, vegetables, and cancer prevention: a review of the epidemiological evidence. Nutrition and Cancer. 1992;18(1):1–29. doi: 10.1080/01635589209514201. [DOI] [PubMed] [Google Scholar]
- Bocker D., Breithardt G. [Induction of arrhythmia by licorice abuse] Zeitschrift fur Kardiologie. 1991;80(6):389–391. [PubMed] [Google Scholar]
- Boysen G., Kenney P.M., Upadhyaya P., Wang M., Hecht S.S. Effects of benzyl isothiocyanate and 2-phenethyl isothiocyanate on benzo[a]pyrene and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone metabolism in F-344 rats. Carcinogenesis. 2003;24(3):517–525. doi: 10.1093/carcin/24.3.517. [DOI] [PubMed] [Google Scholar]
- Brady J.F., Ishizaki H., Fukuto J.M., Lin M.C., Fadel A., Gapac J.M., Yang C.S. Inhibition of cytochrome P-450 2E1 by diallyl sulfide and its metabolites. Chemical Research and Toxicology. 1991;4(6):642–647. doi: 10.1021/tx00024a008. [DOI] [PubMed] [Google Scholar]
- Braga C., La Vecchia C., Franceschi S., Negri E., Parpinel M., Decarli A., Giacosa A., Trichopoulos D. Olive oil, other seasoning fats, and the risk of colorectal carcinoma. Cancer. 1998;82(3):448–453. doi: 10.1002/(sici)1097-0142(19980201)82:3<448::aid-cncr4>3.0.co;2-l. [DOI] [PubMed] [Google Scholar]
- Briante R., La Cara F., Tonziello M.P., Febbraio F., Nucci R. Antioxidant activity of the main bioactive derivatives from oleuropein hydrolysis by hyperthermophilic beta-glycosidase. Journal of Agricultural and Food Chemistry. 2001;49(7):3198–3203. doi: 10.1021/jf001342r. [DOI] [PubMed] [Google Scholar]
- Brüsewitz G., Cameron B.D., Chasseaud L.F., Görler K., Hawkins D.R., Koch H., Mennicke W.H. The metabolism of benzyl isothiocyanate and its cysteine conjugate. Biochemical Journal. 1997;162:99–107. doi: 10.1042/bj1620099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Budavari S. Merck and Co., Inc; Whitehouse Station, NJ: 1996. The Merck Index 12th ed. Merck Research Laboratories. [Google Scholar]
- Butler A.M., Murray M. Inhibition and inactivation of constitutive cytochromes P450 in rat liver by parathion. Molecular Pharmacology. 1993;43(6):902–908. [PubMed] [Google Scholar]
- Cal C., Garban H., Jazirehi A., Yeh C., Mizutani Y., Bonavida B. Resveratrol and cancer: chemoprevention, apoptosis, and chemo-immunosensitizing activities. Current Medicinal Chemistry and Anti-Cancer Agents. 2003;3(2):77–93. doi: 10.2174/1568011033353443. [DOI] [PubMed] [Google Scholar]
- Carluccio M.A., Siculella L., Ancora M.A., Massaro M., Scoditti E., Storelli C., Visioli F., Distante A., De Caterina R. Olive oil and red wine antioxidant polyphenols inhibit endothelial activation: antiatherogenic properties of mediterranean diet phytochemicals. Arteriosclerosis Thrombosis and Vascular Biology. 2003;23(4):622–629. doi: 10.1161/01.ATV.0000062884.69432.A0. [DOI] [PubMed] [Google Scholar]
- Castot A., Larrey D. [Hepatitis observed during a treatment with a drug or tea containing Wild Germander. Evaluation of 26 cases reported to the Regional Centers of Pharmacovigilance] Gastroenterology and Clinical Biology. 1992;16(12):916–922. [PubMed] [Google Scholar]
- Chan W.K., Delucchi A.B. Resveratrol, a red wine constituent, is a mechanism-based inactivator of cytochrome P450 3A4. Life Sciences. 2000;67(25):3103–3112. doi: 10.1016/s0024-3205(00)00888-2. [DOI] [PubMed] [Google Scholar]
- Chan W.K., Nguyen L.T., Miller V.P., Harris R.Z. Mechanism-based inactivation of human cytochrome P450 3A4 by grapefruit juice and red wine. Life Sciences. 1998;62(10):135–142. doi: 10.1016/s0024-3205(98)00013-7. [DOI] [PubMed] [Google Scholar]
- Chandler R.F. Licorice, more than just a flavour. Can. Pharm. J. 1985;118:420–424. [Google Scholar]
- Chen L.J., Lebetkin E.H., Burka L.T. Metabolism of (R)-(+)-pulegone in F344 rats. Drug Metabolism and Disposition. 2001;29(12):1567–1577. [PubMed] [Google Scholar]
- Chen X.G., Liu H.Y., Lei X.H., Fu Z.D., Li Y., Tao L.H., Han R. Cancer chemopreventive and therapeutic activities of red ginseng. Journal of Ethnopharmacology. 1998;60(1):71–78. doi: 10.1016/s0378-8741(97)00133-5. [DOI] [PubMed] [Google Scholar]
- Cinatl J., Morgenstern B., Bauer G., Chandra P., Rabenau H., Doerr H.W. Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet. 2003;361(9374):2045–2046. doi: 10.1016/S0140-6736(03)13615-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cosyns J.P. Aristolochic acid and ‘Chinese Herbs Nephropathy’: a review of the evidence to date. Drug Safety. 2003;26(1):33–48. doi: 10.2165/00002018-200326010-00004. [DOI] [PubMed] [Google Scholar]
- Dalvi R.R. Alterations in hepatic Phase I and Phase II biotransformation enzymes by garlic oil in rats. Toxicology Letters. 1992;60(3):299–305. doi: 10.1016/0378-4274(92)90288-u. [DOI] [PubMed] [Google Scholar]
- D'Angelo S., Manna C., Migliardi V., Mazzoni O., Morrica P., Capasso G., Pontoni G., Galletti P., Zappia V. Pharmacokinetics and metabolism of hydroxytyrosol, a natural antioxidant from olive oil. Drug Metabolism and Disposition. 2001;29(11):1492–1498. [PubMed] [Google Scholar]
- Dao T., Peytier A., Galateau F., Valla A. [Chronic cirrhogenic hepatitis induced by germander] Gastroenterology and Clinical Biology. 1993;17(8–9):609–610. [PubMed] [Google Scholar]
- Dausch J.G., Nixon D.W. Garlic: a review of its relationship to malignant disease. Preventive Medicine. 1990;19(3):346–361. doi: 10.1016/0091-7435(90)90034-h. [DOI] [PubMed] [Google Scholar]
- De Berardinis V., Moulis C., Maurice M., Beaune P., Pessayre D., Pompon D., Loeper J. Human microsomal epoxide hydrolase is the target of germander-induced autoantibodies on the surface of human hepatocytes. Molecular Pharmacology. 2000;58(3):542–551. doi: 10.1124/mol.58.3.542. [DOI] [PubMed] [Google Scholar]
- de Smet P.A. Health risks of herbal remedies. Drug Safety. 1995;13(2):81–93. doi: 10.2165/00002018-199513020-00003. [DOI] [PubMed] [Google Scholar]
- Deng J.F. Clinical and laboratory investigations in herbal poisonings. Toxicology. 2002;27(181–182):571–576. doi: 10.1016/s0300-483x(02)00484-5. [DOI] [PubMed] [Google Scholar]
- Dresser G.K., Spence J.D., Bailey D.G. Pharmacokinetic-pharmacodynamic consequences and clinical relevance of cytochrome P450 3A4 inhibition. Clinical Pharmacokinetics. 2000;38(1):41–57. doi: 10.2165/00003088-200038010-00003. [DOI] [PubMed] [Google Scholar]
- Dubuisson J.G., Dyess D.L., Gaubatz J.W. Resveratrol modulates human mammary epithelial cell O-acetyltransferase, sulfotransferase, and kinase activation of the heterocyclic amine carcinogen N-hydroxy-PhIP. Cancer Letters. 2002;182(1):27–32. doi: 10.1016/s0304-3835(02)00061-7. [DOI] [PubMed] [Google Scholar]
- Ducharme M.P., Warbasse L.H., Edwards D.J. Disposition of intravenous and oral cyclosporine after administration with grapefruit juice. Clinical Pharmacology and Therapeutics. 1995;57(5):485–491. doi: 10.1016/0009-9236(95)90032-2. [DOI] [PubMed] [Google Scholar]
- Edelson R., Berger C., Gasparro F., Jegasothy B., Heald P., Wintroub B., Vonderheid E., Knobler R., Wolff K., Plewig G., McKiernan G., Christiansen L., Oster M., Honigsman H., Wilford H., Kokoschka E., Rehle T., Perez M.D., Stingl G., Laroche L. Treatment of cutaneous T-cell lymphoma by extracorporeal photochemotherapy. Preliminary results. New England Journal of Medicine. 1987;316(6):297–303. doi: 10.1056/NEJM198702053160603. [DOI] [PubMed] [Google Scholar]
- El-Mowafy A.M. Resveratrol activates membrane-bound guanylyl cyclase in coronary arterial smooth muscle: a novel signaling mechanism in support of coronary protection. Biochemical and Biophysical Research Communications. 2002;291(5):1218–1222. doi: 10.1006/bbrc.2002.6598. [DOI] [PubMed] [Google Scholar]
- El-Mowafy A.M., Alkhalaf M. Resveratrol activates adenylyl-cyclase in human breast cancer cells: a novel, estrogen receptor-independent cytostatic mechanism. Carcinogenesis. 2003;24(5):869–873. doi: 10.1093/carcin/bgg015. [DOI] [PubMed] [Google Scholar]
- Ernst E. Adverse effects of herbal drugs in dermatology. British Journal of Dermatology. 2000;143(5):923–929. doi: 10.1046/j.1365-2133.2000.03822.x. [DOI] [PubMed] [Google Scholar]
- Ernst E. Toxic heavy metals and undeclared drugs in Asian herbal medicines. Trends in Pharmacological Sciences. 2002;23(3):136–139. doi: 10.1016/S0165-6147(00)01972-6. [DOI] [PubMed] [Google Scholar]
- Ernst E., Pittler M.H. Herbal medicine. Med. Clin. North Am. 2002;86(1):149–161. doi: 10.1016/s0025-7125(03)00077-4. [DOI] [PubMed] [Google Scholar]
- Estrov Z., Shishodia S., Faderl S., Harris D., Van Q., Kantarjian H.M., Talpaz M., Aggarwal B.B. Resveratrol blocks interleukin-1beta-induced activation of the nuclear transcription factor NF-κB, inhibits proliferation, causes S-phase arrest, and induces apoptosis of acute myeloid leukemia cells. Blood. 2003 doi: 10.1182/blood-2002-11-3550. (on line publication) [DOI] [PubMed] [Google Scholar]
- Fau D., Lekehal M., Farrell G., Moreau A., Moulis C., Feldmann G., Haouzi D., Pessayre D. Diterpenoids from germander, an herbal medicine, induce apoptosis in isolated rat hepatocytes. Gastroenterology. 1997;113(4):1334–1346. doi: 10.1053/gast.1997.v113.pm9322529. [DOI] [PubMed] [Google Scholar]
- Fenwick G.R., Heaney R.K., Mullin W.J. Glucosinolates and their breakdown products in food and food plants. Critical Review in Food Sciences and Nutrition. 1983;18(2):123–201. doi: 10.1080/10408398209527361. [DOI] [PubMed] [Google Scholar]
- Fernando R.C., Schmeiser H.H., Scherf H.R., Wiessler M. Formation and persistence of specific purine DNA adducts by 32P-postlabelling in target and non-target organs of rats treated with aristolochic acid I. IARC Scientific Publications. 1993;124:167–171. [PubMed] [Google Scholar]
- Ferrari P., Sansonnens A., Dick B., Frey F.J. In vivo 11β-HSD-2 activity: variability, salt-sensitivity, and effect of licorice. Hypertension. 2001;38(6):1330–1336. doi: 10.1161/hy1101.096112. [DOI] [PubMed] [Google Scholar]
- Ferry-Dumazet H., Garnier O., Mamani-Matsuda M., Vercauteren J., Belloc F., Billiard C., Dupouy M., Thiolat D., Kolb J.P., Marit G., Reiffers J., Mossalayi M.D. Resveratrol inhibits the growth and induces the apoptosis of both normal and leukemic hematopoietic cells. Carcinogenesis. 2002;23(8):1327–1333. doi: 10.1093/carcin/23.8.1327. [DOI] [PubMed] [Google Scholar]
- Fisher B., Costantino J.P., Wickerham D.L., Redmond C.K., Kavanah M., Cronin W.M., Vogel V., Robidoux A., Dimitrov N., Atkins J., Daly M., Wieand S., Tan-Chiu E., Ford L., Wolmark N. Tamoxifen for prevention of breast cancer: Report of the National Surgical Adjuvant Breast and Bowel Project P-1 study. Journal of the National Cancer Institute. 1998;90(18):1371–1388. doi: 10.1093/jnci/90.18.1371. [DOI] [PubMed] [Google Scholar]
- Fitzsimmons M.E., Collins J.M. Selective biotransformation of the human immunodeficiency virus protease inhibitor saquinavir by human small-intestinal cytochrome P450 3A4: potential contribution to high first-pass metabolism. Drug Metabolism and Disposition. 1997;25(2):256–266. [PubMed] [Google Scholar]
- Foster B.C., Foster M.S., Vandenhoek S., Krantis A., Budzinski J.W., Arnason J.T., Gallicano K.D., Choudri S. An in vitro evaluation of human cytochrome P450 3A4 and P-glycoprotein inhibition by garlic. Journal of Pharmacy and Pharmaceutical Sciences. 2001;4(2):176–184. [PubMed] [Google Scholar]
- Fouin-Fortunet H., Tinel M., Descatoire V., Letteron P., Larrey D., Geneve J., Pessayre D. Inactivation of cytochrome P-450 by the drug methoxsalen. Journal of Pharmacology and Experimental Therapeutics. 1986;236(1):237–247. [PubMed] [Google Scholar]
- Frantz D.J., Hughes B.G., Nelson D.R., Murray B.K., Christensen M.J. Cell cycle arrest and differential gene expression in HT-29 cells exposed to an aqueous garlic extract. Nutrition and Cancer. 2000;38(2):255–264. doi: 10.1207/S15327914NC382_15. [DOI] [PubMed] [Google Scholar]
- Fugh-Berman A. Herb-drug interactions. Lancet. 2000;355(9198):134–138. doi: 10.1016/S0140-6736(99)06457-0. [DOI] [PubMed] [Google Scholar]
- Fugh-Berman A., Ernst E. Herb-drug interactions: review and assessment of report reliability. British Journal of Clinical Pharmacology. 2001;52(5):587–595. doi: 10.1046/j.0306-5251.2001.01469.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujisawa Y., Sakamoto M., Matsushita M., Fujita T., Nishioka K. Glycyrrhizin inhibits the lytic pathway of complement—possible mechanism of its anti-inflammatory effect on liver cells in viral hepatitis. Microbiology and Immunology. 2000;44(9):799–804. doi: 10.1111/j.1348-0421.2000.tb02566.x. [DOI] [PubMed] [Google Scholar]
- Fujita K., Kamataki T. Screening of organosulfur compounds as inhibitors of human CYP2A6. Drug Metabolism and Disposition. 2001;29(7):983–989. [PubMed] [Google Scholar]
- Fukai T., Marumo A., Kaitou K., Kanda T., Terada S., Nomura T. Anti-Helicobacter pylori flavonoids from licorice extract. Life Sciences. 2002;71(12):1449–1463. doi: 10.1016/s0024-3205(02)01864-7. [DOI] [PubMed] [Google Scholar]
- Fukutake M., Miura N., Yamamoto M., Fukuda K., Iijima O., Ishikawa H., Kubo M., Okada M., Komatsu Y., Sasaki H., Wakabayashi K., Ishige A., Amagaya S. Suppressive effect of the herbal medicine Oren-gedoku-to on cyclooxygenase-2 activity and azoxymethane-induced aberrant crypt foci development in rats. Cancer Letters. 2000;157(1):9–14. doi: 10.1016/s0304-3835(00)00432-8. [DOI] [PubMed] [Google Scholar]
- Fukutake M., Yokota S., Kamamura H., Iizuka A., Amagaya S., Fukuda K., Komatsu Y. Inhibitory effect of Coptidis Rhizoma and Scutellariae Radix on azoxymethane-induced aberrant crypt foci formation in rat colon. Biological and Pharmaceutical Bulletin. 1998;21(8):814–817. doi: 10.1248/bpb.21.814. [DOI] [PubMed] [Google Scholar]
- Gannett M., Iversen P., Lawson T. The mechanism of inhibition of cytochrome P450IIE1 by dihydrocapsaicin. Bioorganic Chemistry. 1990;18:185–198. [Google Scholar]
- Gohil K. Genomic responses to herbal extracts: lessons from in vitro and in vivo studies with an extract of Ginkgo biloba. Biochemical Pharmacology. 2002;64(5–6):913–917. doi: 10.1016/s0006-2952(02)01163-2. [DOI] [PubMed] [Google Scholar]
- Gohil K., Moy R.K., Farzin S., Maguire J.J., Packer L. mRNA expression profile of a human cancer cell line in response to Ginkgo biloba extract: induction of antioxidant response and the Golgi system. Free Radical Research. 2000;33(6):831–849. doi: 10.1080/10715760000301351. [DOI] [PubMed] [Google Scholar]
- Gonzalez F.J. Evolution of the P450 gene superfamily: animal-plant ‘warfare’, molecular drive and human genetic differences in drug oxidation. Trends in Genetics. 1990;6(6):182–186. doi: 10.1016/0168-9525(90)90174-5. [DOI] [PubMed] [Google Scholar]
- Gonzalez F.J., Gelboin H.V. Role of human cytochromes P450 in the metabolic activation of chemical carcinogens and toxins. Drug Metabolism Reviews. 1994;26(1–2):165–183. doi: 10.3109/03602539409029789. [DOI] [PubMed] [Google Scholar]
- Goosen T.C., Kent U.M., Brand L., Hollenberg P.F. Inactivation of cytochrome P4502B1 by benzyl isothiocyanate, a chemopreventative agent from cruciferous vegetables. Chemical Research and Toxicology. 2000;13(12):1349–1359. doi: 10.1021/tx000133y. [DOI] [PubMed] [Google Scholar]
- Goosen T.C., Mills D.E., Hollenberg P.F. Effects of benzyl isothiocyanate on rat and human cytochromes P450: identification of metabolites formed by P450 2B1. Journal of Pharmacology and Experimental Therapeutics. 2001;296(1):198–206. [PubMed] [Google Scholar]
- Gordon W.P., Forte A.J., McMurtry R.J., Gal J., Nelson S.D. Hepatotoxicity and pulmonary toxicity of pennyroyal oil and its constituent terpenes in the mouse. Toxicology and Applied Pharmacology. 1982;65(3):413–424. doi: 10.1016/0041-008x(82)90387-8. [DOI] [PubMed] [Google Scholar]
- Gordon W.P., Huitric A.C., Seth C.L., McClanahan R.H., Nelson S.D. The metabolism of the abortifacient terpene, (R)-(+)-pulegone, to a proximate toxin, menthofuran. Drug Metabolism and Disposition. 1987;15(5):589–594. [PubMed] [Google Scholar]
- Gorski J.C., Hall S.D., Jones D.R., VandenBranden M., Wrighton S.A. Regioselective biotransformation of midazolam by members of the human cytochrome P450 3A (CYP3A) subfamily. Biochemical Pharmacology. 1994;47(9):1643–1653. doi: 10.1016/0006-2952(94)90543-6. [DOI] [PubMed] [Google Scholar]
- Gorski J.C., Jones D.R., Hamman M.A., Wrighton S.A., Hall S.D. Biotransformation of alprazolam by members of the human cytochrome P4503A subfamily. Xenobiotica. 1999;29(9):931–944. doi: 10.1080/004982599238173. [DOI] [PubMed] [Google Scholar]
- Greensfelder L. Alternative medicine. Herbal product linked to cancer. Science. 2000;288(5473):1946. doi: 10.1126/science.288.5473.1946a. [DOI] [PubMed] [Google Scholar]
- Grundschober F. Literature review of pulegone. Perfumer. 1979:415–417. [Google Scholar]
- Guengerich F.P. Forging the links between metabolism and carcinogenesis. Mutation Research. 2001;488(3):195–209. doi: 10.1016/s1383-5742(01)00059-x. [DOI] [PubMed] [Google Scholar]
- Guengerich F.P., Liebler D.C. Enzymatic activation of chemicals to toxic metabolites. Critical Review in Toxicology. 1985;14(3):259–307. doi: 10.3109/10408448509037460. [DOI] [PubMed] [Google Scholar]
- Guengerich F.P., Shimada T. Oxidation of toxic and carcinogenic chemicals by human cytochrome P-450 enzymes. Chemical Research and Toxicology. 1991;4(4):391–407. doi: 10.1021/tx00022a001. [DOI] [PubMed] [Google Scholar]
- Guo Z., Smith T.J., Wang E., Sadrieh N., Ma Q., Thomas P.E., Yang C.S. Effects of phenethyl isothiocyanate, a carcinogenesis inhibitor, on xenobiotic-metabolizing enzymes and nitrosamine metabolism in rats. Carcinogenesis. 1992;13(12):2205–2210. doi: 10.1093/carcin/13.12.2205. [DOI] [PubMed] [Google Scholar]
- Gurley B.J., Gardner S.F., Hubbard M.A., Williams D.K., Gentry W.B., Cui Y., Ang C.Y. Cytochrome P450 phenotypic ratios for predicting herb-drug interactions in humans. Clinical Pharmacology and Therapeutics. 2002;72(3):276–287. doi: 10.1067/mcp.2002.126913. [DOI] [PubMed] [Google Scholar]
- Gusman J., Malonne H., Atassi G. A reappraisal of the potential chemopreventive and chemotherapeutic properties of resveratrol. Carcinogenesis. 2001;22(8):1111–1117. doi: 10.1093/carcin/22.8.1111. [DOI] [PubMed] [Google Scholar]
- Guyonnet D., Belloir C., Suschetet M., Siess M.H., Le Bon A.M. Antimutagenic activity of organosulfur compounds from Allium is associated with Phase II enzyme induction. Mutation Research. 2001;495(1–2):135–145. doi: 10.1016/s1383-5718(01)00205-4. [DOI] [PubMed] [Google Scholar]
- Gwilt P.R., Lear C.L., Tempero M.A., Birt D.D., Grandjean A.C., Ruddon R.W., Nagel D.L. The effect of garlic extract on human metabolism of acetaminophen. Cancer Epidemiology Biomarkers and Prevention. 1994;3(2):155–160. [PubMed] [Google Scholar]
- Haber D., Siess M.H., Canivenc-Lavier M.C., Le Bon A.M., Suschetet M. Differential effects of dietary diallyl sulfide and diallyl disulfide on rat intestinal and hepatic drug-metabolizing enzymes. Journal of Toxicology and Environmental Health. 1995;44(4):423–434. doi: 10.1080/15287399509531971. [DOI] [PubMed] [Google Scholar]
- Haber D., Siess M.H., De Waziers I., Beaune P., Suschetet M. Modification of hepatic drug-metabolizing enzymes in rat fed naturally occurring allyl sulphides. Xenobiotica. 1994;24(2):169–182. doi: 10.3109/00498259409043230. [DOI] [PubMed] [Google Scholar]
- Hall R.L., Oser B.L. Recent progress in the consideration of flavoring ingredients under the food additives amendment III GRAS substances. Food Technology. 1965;19:253–271. [Google Scholar]
- Haller C.A., Benowitz N.L. Adverse cardiovascular and central nervous system events associated with dietary supplements containing ephedra alkaloids. New England Journal of Medicine. 2000;343(25):1833–1838. doi: 10.1056/NEJM200012213432502. [DOI] [PubMed] [Google Scholar]
- Hatano T., Fukuda T., Liu Y.Z., Noro T., Okuda T. [Phenolic constituents of licorice. IV. Correlation of phenolic constituents and licorice specimens from various sources, and inhibitory effects of licorice extracts on xanthine oxidase and monoamine oxidase] Yakugaku Zasshi. 1991;111(6):311–321. doi: 10.1248/yakushi1947.111.6_311. [DOI] [PubMed] [Google Scholar]
- Hatano T., Fukuda T., Miyase T., Noro T., Okuda T. Phenolic constituents of licorice. III. Structures of glicoricone and licofuranone, and inhibitory effects of licorice constituents on monoamine oxidase. Chemical and Pharmaceutical Bulletin. 1991;39(5):1238–1243. doi: 10.1248/cpb.39.1238. [DOI] [PubMed] [Google Scholar]
- Hattori R., Otani H., Maulik N., Das D.K. Pharmacological preconditioning with resveratrol: role of nitric oxide. American Journal of Physiology and Heart Circulation Physiology. 2002;282(6):H1988–H1995. doi: 10.1152/ajpheart.01012.2001. [DOI] [PubMed] [Google Scholar]
- He K., Iyer K.R., Hayes R.N., Sinz M.W., Woolf T.F., Hollenberg P.F. Inactivation of cytochrome P450 3A4 by bergamottin, a component of grapefruit juice. Chemical Research and Toxicology. 1998;11(4):252–259. doi: 10.1021/tx970192k. [DOI] [PubMed] [Google Scholar]
- Hecht S.S. Approaches to chemoprevention of lung cancer based on carcinogens in tobacco smoke. Environmental Health Perspectives. 1997;105(Suppl.):955–963. doi: 10.1289/ehp.97105s4955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hecht S.S., Kenney P.M., Wang M., Upadhyaya P. Benzyl isothiocyanate: an effective inhibitor of polycyclic aromatic hydrocarbon tumorigenesis in A/J mouse lung. Cancer Letters. 2002;187(1–2):87–94. doi: 10.1016/s0304-3835(02)00410-x. [DOI] [PubMed] [Google Scholar]
- Hirose M., Yamaguchi T., Kimoto N., Ogawa K., Futakuchi M., Sano M., Shirai T. Strong promoting activity of phenylethyl isothiocyanate and benzyl isothiocyanate on urinary bladder carcinogenesis in F344 male rats. International Journal of Cancer. 1998;77(5):773–777. doi: 10.1002/(sici)1097-0215(19980831)77:5<773::aid-ijc17>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- Hirsch K., Danilenko M., Giat J., Miron T., Rabinkov A., Wilchek M., Mirelman D., Levy J., Sharoni Y. Effect of purified allicin, the major ingredient of freshly crushed garlic, on cancer cell proliferation. Nutrition and Cancer. 2000;38(2):245–254. doi: 10.1207/S15327914NC382_14. [DOI] [PubMed] [Google Scholar]
- Ioset J.R., Raoelison G.E., Hostettmann K. Detection of aristolochic acid in Chinese phytomedicines and dietary supplements used as slimming regimens. Food and Chemical Toxicology. 2003;41(1):29–36. doi: 10.1016/s0278-6915(02)00219-3. [DOI] [PubMed] [Google Scholar]
- Ishizaki H., Brady J.F., Ning S.M., Yang C.S. Effect of phenethyl isothiocyanate on microsomal N-nitrosodimethylamine metabolism and other monooxygenase activities. Xenobiotica. 1990;20(3):255–264. doi: 10.3109/00498259009046845. [DOI] [PubMed] [Google Scholar]
- Jamieson D.D., Duffield P.H. Positive interaction of ethanol and kava resin in mice. Clinical and Experimental Pharmacology and Physiology. 1990;17(7):509–514. doi: 10.1111/j.1440-1681.1990.tb01350.x. [DOI] [PubMed] [Google Scholar]
- Jang M., Cai L., Udeani G.O., Slowing K.V., Thomas C.F., Beecher C.W., Fong H.H., Farnsworth N.R., Kinghorn A.D., Mehta R.G., Moon R.C., Pezzuto J.M. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science. 1997;275(5297):218–220. doi: 10.1126/science.275.5297.218. [DOI] [PubMed] [Google Scholar]
- Jin J.X., Baillie T.A. Metabolism of the chemoprotective agent diallyl sulfide to glutathione conjugates in rats. Chemical Research and Toxicology. 1997;10(3):318–327. doi: 10.1021/tx9601768. [DOI] [PubMed] [Google Scholar]
- Johnson J.L., Maddipati K.R. Paradoxical effects of resveratrol on the two prostaglandin H synthases. Prostaglandins. 1998;56(2–3):131–143. doi: 10.1016/s0090-6980(98)00052-5. [DOI] [PubMed] [Google Scholar]
- Jolly K., Cheng K.K., Langman M.J.S. NSAIDs and gastrointestinal cancer prevention. Drugs. 2002;62(6):945–956. doi: 10.2165/00003495-200262060-00006. [DOI] [PubMed] [Google Scholar]
- Kane G.C., Lipsky J.J. Drug-grapefruit juice interactions. Mayo Clinic Proceedings. 2000;75(9):933–942. doi: 10.4065/75.9.933. [DOI] [PubMed] [Google Scholar]
- Kelloff G.J., Crowell J.A., Hawk E.T., Steele V.E., Lubet R.A., Boone C.W., Covey J.M., Doody L.A., Omenn G.S., Greenwald P., Hong W.K., Parkinson D.R., Bagheri D., Baxter G.T., Blunden M., Doeltz M.K., Eisenhauer K.M., Johnson K., Knapp G.G., Longfellow D.G., Malone W.F., Nayfield S.G., Seifried H.E., Swall L.M., Sigman C.C. Strategy and planning for chemopreventive drug development: clinical development plans II. Journal of Cellular Biochemistry. 1996;26(Suppl.):54–57. doi: 10.1002/jcb.240630705. [DOI] [PubMed] [Google Scholar]
- Kent U.M., Aviram M., Rosenblat M., Hollenberg P.F. The licorice root derived isoflavan glabridin inhibits the activities of human cytochrome P450S 3A4, 2B6, and 2C9. Drug Metabolism and Disposition. 2002;30(6):709–715. doi: 10.1124/dmd.30.6.709. [DOI] [PubMed] [Google Scholar]
- Kessler D.A. Cancer and herbs. New England Journal of Medicine. 2000;342(23):1742–1743. doi: 10.1056/NEJM200006083422309. [DOI] [PubMed] [Google Scholar]
- Kimura Y. Pharmacological studies on resveratrol. Methods and Findings in Experimental and Clinical Pharmacology. 2003;25(4):297–310. doi: 10.1358/mf.2003.25.4.727207. [DOI] [PubMed] [Google Scholar]
- Kishimoto R., Ueda M., Yoshinaga H., Goda K., Park S.S. Combined effects of ethanol and garlic on hepatic ethanol metabolism in mice. Journal of Nutrition Sciences and Vitaminology. 1999;45(3):275–286. doi: 10.3177/jnsv.45.275. [DOI] [PubMed] [Google Scholar]
- Klepser T.B., Klepser M.E. Unsafe and potentially safe herbal therapies. American Journal of Health-System Pharmacy. 1999;56(2):125–138. doi: 10.1093/ajhp/56.2.125. [DOI] [PubMed] [Google Scholar]
- Klinge C.M., Risinger K.E., Watts M.B., Beck V., Eder R., Jungbauer A. Estrogenic activity in white and red wine extracts. Journal of Agricultural and Food Chemistry. 2003;51(7):1850–1857. doi: 10.1021/jf0259821. [DOI] [PubMed] [Google Scholar]
- Koenigs L.L., Peter R.M., Thompson S.J., Rettie A.E., Trager W.F. Mechanism-based inactivation of human liver cytochrome P450 2A6 by 8-methoxypsoralen. Drug Metabolism and Disposition. 1997;25(12):1407–1415. [PubMed] [Google Scholar]
- Koenigs L.L., Trager W.F. Mechanism-based inactivation of cytochrome P450 2B1 by 8-methoxypsoralen and several other furanocoumarins. Biochemistry. 1998;37(38):13184–13193. doi: 10.1021/bi981198r. [DOI] [PubMed] [Google Scholar]
- Koenigs L.L., Trager W.F. Mechanism-based inactivation of P450 2A6 by furanocoumarins. Biochemistry. 1998;37(28):10047–10061. doi: 10.1021/bi980003c. [DOI] [PubMed] [Google Scholar]
- Koh H.L., Woo S.O. Chinese proprietary medicine in Singapore: regulatory control of toxic heavy metals and undeclared drugs. Drug Safety. 2000;23(5):351–362. doi: 10.2165/00002018-200023050-00001. [DOI] [PubMed] [Google Scholar]
- Kohara A., Suzuki T., Honma M., Ohwada T., Hayashi M. Mutagenicity of aristolochic acid in the λ/lacZ transgenic mouse (MutaMouse) Mutation Research. 2002;515(1–2):63–72. doi: 10.1016/s1383-5718(01)00350-3. [DOI] [PubMed] [Google Scholar]
- Kohyama N., Nagata T., Fujimoto S., Sekiya K. Inhibition of arachidonate lipoxygenase activities by 2-(3,4-dihydroxyphenyl)ethanol, a phenolic compound from olives. Bioscience Biotechnology and Biochemistry. 1997;61(2):347–350. doi: 10.1271/bbb.61.347. [DOI] [PubMed] [Google Scholar]
- Kouzi S.A., McMurtry R.J., Nelson S.D. Hepatotoxicity of germander (Teucrium chamaedrys L.) and one of its constituent neoclerodane diterpenes teucrin A in the mouse. Chemical Research and Toxicology. 1994;7(6):850–856. doi: 10.1021/tx00042a020. [DOI] [PubMed] [Google Scholar]
- Kuo P.L., Chiang L.C., Lin C.C. Resveratrol-induced apoptosis is mediated by p53-dependent pathway in Hep G2 cells. Life Sciences. 2002;72(1):23–34. doi: 10.1016/s0024-3205(02)02177-x. [DOI] [PubMed] [Google Scholar]
- Kuwajerwala N., Cifuentes E., Gautam S., Menon M., Barrack E.R., Reddy G.P. Resveratrol induces prostate cancer cell entry into S phase and inhibits DNA synthesis. Cancer Research. 2002;62(9):2488–2492. [PubMed] [Google Scholar]
- Kwon K.B., Yoo S.J., Ryu D.G., Yang J.Y., Rho H.W., Kim J.S., Park J.W., Kim H.R., Park B.H. Induction of apoptosis by diallyl disulfide through activation of caspase-3 in human leukemia HL-60 cells. Biochemical Pharmacology. 2002;63(1):41–47. doi: 10.1016/s0006-2952(01)00860-7. [DOI] [PubMed] [Google Scholar]
- Labbe G., Descatoire V., Beaune P., Letteron P., Larrey D., Pessayre D. Suicide inactivation of cytochrome P-450 by methoxsalen: evidence for the covalent binding of a reactive intermediate to the protein moiety. Journal of Pharmacology and Experimental Therapeutics. 1989;250(3):1034–1042. [PubMed] [Google Scholar]
- Lahiri-Chatterjee M., Katiyar S.K., Mohan R.R., Agarwal R. A flavonoid antioxidant, silymarin, affords exceptionally high protection against tumor promotion in the SENCAR mouse skin tumorigenesis model. Cancer Research. 1999;59(3):622–632. [PubMed] [Google Scholar]
- Laliberte L., Villeneuve J.P. Hepatitis after the use of germander, a herbal remedy. Canadian Medical Association Journal. 1996;154(11):1689–1692. [PMC free article] [PubMed] [Google Scholar]
- Landrault N., Larronde F., Delaunay J.C., Castagnino C., Vercauteren J., Merillon J.M., Gasc F., Cros G., Teissedre P.L. Levels of stilbene oligomers and astilbin in French varietal wines and in grapes during noble rot development. Journal of Agricultural and Food Chemistry. 2002;50(7):2046–2052. doi: 10.1021/jf010794g. [DOI] [PubMed] [Google Scholar]
- Larrey D., Vial T., Pauwels A., Castot A., Biour M., David M., Michel H. Hepatitis after germander (Teucrium chamaedrys) administration: another instance of herbal medicine hepatotoxicity. Annals of Internal Medicine. 1992;117(2):129–132. doi: 10.7326/0003-4819-117-2-129. [DOI] [PubMed] [Google Scholar]
- Lebot V., Lévesque J. The origin and distribution of kava (Piper methysticum Forst. f. and Piper wichmannii C. DC., Piperaceae): a phytochemical approach. Allertonia. 1989;5:223–280. [Google Scholar]
- Lee M.-S. Enzyme induction and comparative oxidative desulfuration of isothiocyanates to isocyanates. Chemical Research and Toxicology. 1996;9(7):1072–1078. doi: 10.1021/tx950213f. [DOI] [PubMed] [Google Scholar]
- Lekehal M., Pessayre D., Lereau J.M., Moulis C., Fouraste I., Fau D. Hepatotoxicity of the herbal medicine germander: metabolic activation of its furano diterpenoids by cytochrome P450 3A depletes cytoskeleton-associated protein thiols and forms plasma membrane blebs in rat hepatocytes. Hepatology. 1996;24(1):212–218. doi: 10.1002/hep.510240134. [DOI] [PubMed] [Google Scholar]
- Letteron P., Descatoire V., Larrey D., Tinel M., Geneve J., Pessayre D. Inactivation and induction of cytochrome P-450 by various psoralen derivatives in rats. Journal of Pharmacology and Experimental Therapeutics. 1986;238(2):685–692. [PubMed] [Google Scholar]
- Levenson A.S., Gehm B.D., Pearce S.T., Horiguchi J., Simons L.A., Ward J.E., 3rd, Jameson J.L., Jordan V.C. Resveratrol acts as an estrogen receptor (ER) agonist in breast cancer cells stably transfected with ER-α. International Journal of Cancer. 2003;104(5):587–596. doi: 10.1002/ijc.10992. [DOI] [PubMed] [Google Scholar]
- Li Z.G., Hong T., Shimada Y., Komoto I., Kawabe A., Ding Y., Kaganoi J., Hashimoto Y., Imamura M. Suppression of N-nitrosomethylbenzylamine (NMBA)-induced esophageal tumorigenesis in F344 rats by resveratrol. Carcinogenesis. 2002;23(9):1531–1536. doi: 10.1093/carcin/23.9.1531. [DOI] [PubMed] [Google Scholar]
- Liang Y.C., Tsai S.H., Chen L., Lin-Shiau S.Y., Lin J.K. Resveratrol-induced G2 arrest through the inhibition of CDK7 and p34CDC2 kinases in colon carcinoma HT29 cells. Biochemical Pharmacology. 2003;65(7):1053–1060. doi: 10.1016/s0006-2952(03)00011-x. [DOI] [PubMed] [Google Scholar]
- Lin J.-M., Amin S., Trushin N., Hecht S.S. Effect of isothiocyanates on tumorigenesis by benzo[a]pyrene in murine tumor models. Cancer Letters. 1993;74(3):151–159. doi: 10.1016/0304-3835(93)90237-4. [DOI] [PubMed] [Google Scholar]
- Lin M.C., Wang E.J., Patten C., Lee M.J., Xiao F., Reuhl K.R., Yang C.S. Protective effect of diallyl sulfone against acetaminophen-induced hepatotoxicity in mice. Journal of Biochemical Toxicology. 1996;11(1):11–20. doi: 10.1002/(SICI)1522-7146(1996)11:1<11::AID-JBT2>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- Loeper J., De Berardinis V., Moulis C., Beaune P., Pessayre D., Pompon D. Human epoxide hydrolase is the target of germander autoantibodies on the surface of human hepatocytes: enzymatic implications. Advances in Experimental and Medical Biology. 2001;500:121–124. doi: 10.1007/978-1-4615-0667-6_13. [DOI] [PubMed] [Google Scholar]
- Loeper J., Descatoire V., Letteron P., Moulis C., Degott C., Dansette P., Fau D., Pessayre D. Hepatotoxicity of germander in mice. Gastroenterology. 1994;106(2):464–472. doi: 10.1016/0016-5085(94)90606-8. [DOI] [PubMed] [Google Scholar]
- Lopez-Carillo L., Avila M.H., Dubrow R. Chili pepper consumption and gastric cancer in Mexico: a case control study. American Journal of Epidemiology. 1994;139(3):263–271. doi: 10.1093/oxfordjournals.aje.a116993. [DOI] [PubMed] [Google Scholar]
- Lord G.M., Cook T., Arlt V.M., Schmeiser H.H., Williams G., Pusey C.D. Urothelial malignant disease and Chinese herbal nephropathy. Lancet. 2001;358(9292):1515–1516. doi: 10.1016/s0140-6736(01)06576-x. [DOI] [PubMed] [Google Scholar]
- Madyastha K.M., Gaikwad N.W. Metabolic disposition of a monoterpene ketone, piperitenone, in rats: evidence for the formation of a known toxin, p-cresol. Drug Metabolism and Disposition. 1999;27(1):74–80. [PubMed] [Google Scholar]
- Madyastha K.M., Raj C.P. Biotransformations of R-(+)-pulegone and menthofuran in vitro: chemical basis for toxicity. Biochemical and Biophysical Research Communications. 1990;173(3):1086–1092. doi: 10.1016/s0006-291x(05)80897-6. [DOI] [PubMed] [Google Scholar]
- Madyastha K.M., Raj C.P. Metabolic fate of menthofuran in rats. Novel oxidative pathways. Drug Metabolism and Disposition. 1992;20(2):295–301. [PubMed] [Google Scholar]
- Madyastha K.M., Raj C.P. Studies on the metabolism of a monoterpene ketone, (R)-(+)-pulegonea hepatotoxin in rat: isolation and characterization of new metabolites. Xenobiotica. 1993;23(5):509–518. doi: 10.3109/00498259309059391. [DOI] [PubMed] [Google Scholar]
- Madyastha K.M., Raj C.P. Stereoselective hydroxylation of 4-methyl-2-cyclohexenone in rats: its relevance to R-(+)-pulegone-mediated hepatotoxicity. Biochemical and Biophysical Research Communications. 2002;297(2):202–205. doi: 10.1016/s0006-291x(02)02179-4. [DOI] [PubMed] [Google Scholar]
- Maenpaa J., Sigusch H., Raunio H., Syngelma T., Vuorela P., Vuorela H., Pelkonen O. Differential inhibition of coumarin 7-hydroxylase activity in mouse and human liver microsomes. Biochemical Pharmacology. 1993;45(5):1035–1042. doi: 10.1016/0006-2952(93)90247-t. [DOI] [PubMed] [Google Scholar]
- Manyike P.T., Kharasch E.D., Kalhorn T.F., Slattery J.T. Contribution of CYP2E1 and CYP3A to acetaminophen reactive metabolite formation. Clinical Pharmacology and Therapeutics. 2000;67(3):275–282. doi: 10.1067/mcp.2000.104736. [DOI] [PubMed] [Google Scholar]
- Martinez J., Moreno J.J. Effect of resveratrol, a natural polyphenolic compound, on reactive oxygen species and prostaglandin production. Biochemical Pharmacology. 2000;59(7):865–870. doi: 10.1016/s0006-2952(99)00380-9. [DOI] [PubMed] [Google Scholar]
- Mathews J.M., Etheridge A.S., Black S.R. Inhibition of human cytochrome P450 activities by kava extract and kavalactones. Drug Metabolism and Disposition. 2002;30(22):1153–1157. doi: 10.1124/dmd.30.11.1153. [DOI] [PubMed] [Google Scholar]
- Mattei A., Bizollon T., Charles J.D., Debat P., Fontanges T., Chevallier M., Trepo C. [Liver damage induced by the ingestion of a product of phytotherapy containing wild germander, Four cases] Gastroenterology and Clinical Biology. 1992;16(10):798–800. [PubMed] [Google Scholar]
- Mays D.C., Hilliard J.B., Wong D.D., Chambers M.A., Park S.S., Gelboin H.V., Gerber N. Bioactivation of 8-methoxypsoralen and irreversible inactivation of cytochrome P-450 in mouse liver microsomes: modification by monoclonal antibodies, inhibition of drug metabolism and distribution of covalent adducts. Journal of Pharmacology and Experimental Therapeutics. 1990;254(2):720–731. [PubMed] [Google Scholar]
- McClanahan R.H., Thomassen D., Slattery J.T., Nelson S.D. Metabolic activation of (R)-(+)-pulegone to a reactive enonal that covalently binds to mouse liver proteins. Chemical Research and Toxicology. 1989;2(5):349–355. doi: 10.1021/tx00011a013. [DOI] [PubMed] [Google Scholar]
- McRae C.A., Agarwal K., Mutimer D., Bassendine M.F. Hepatitis associated with Chinese herbs. European Journal of Gastroenterology and Hepatology. 2002;14(5):559–562. doi: 10.1097/00042737-200205000-00015. [DOI] [PubMed] [Google Scholar]
- Meyer H.J. [Pharmakologie der wirksamen Prinzipien des Kawa-Rhizoms (Piper methysticum Forst. f.)] Archives in International Pharmacodynamics and Therapeutics. 1962;138:505–536. [PubMed] [Google Scholar]
- Miro-Casas E., Covas M.I., Farre M., Fito M., Ortuno J., Weinbrenner T., Roset P., De La Torre R. Hydroxytyrosol disposition in humans. Clinical Chemistry. 2003;49(6):945–952. doi: 10.1373/49.6.945. [DOI] [PubMed] [Google Scholar]
- Mohri K., Uesawa Y. Effects of furanocoumarin derivatives in grapefruit juice on nifedipine pharmacokinetics in rats. Pharmaceutical Research. 2001;18(2):177–182. doi: 10.1023/a:1011028401189. [DOI] [PubMed] [Google Scholar]
- Moorthy B., Madyastha P., Madyastha K.M. Metabolism of a monoterpene ketone, (R)-(+)-pulegonea hepatotoxin in rat. Xenobiotica. 1989;19(2):217–224. doi: 10.3109/00498258909034694. [DOI] [PubMed] [Google Scholar]
- Moreno J.J. Resveratrol modulates arachidonic acid release, prostaglandin synthesis, and 3T6 fibroblast growth. Journal of Pharmacology and Experimental Therapeutics. 2000;294(1):333–338. [PubMed] [Google Scholar]
- Moreno R.L., Kent U.M., Hodge K., Hollenberg P.F. Inactivation of cytochrome P450 2E1 by benzyl isothiocyanate. Chemical Research and Toxicology. 1999;12(7):582–587. doi: 10.1021/tx9900019. [DOI] [PubMed] [Google Scholar]
- Mostefa-Kara N., Pauwels A., Pines E., Biour M., Levy V.G. Fatal hepatitis after herbal tea. Lancet. 1992;340(8820):674. [PubMed] [Google Scholar]
- Mueller S.O., Kling M., Arifin Firzani P., Mecky A., Duranti E., Shields-Botella J., Delansorne R., Broschard T., Kramer P.J. Activation of estrogen receptor-α and ERβ by 4-methylbenzylidene-camphor in human and rat cells: comparison with phyto- and xenoestrogens. Toxicology Letters. 2003;142(1–2):89–101. doi: 10.1016/s0378-4274(03)00016-x. [DOI] [PubMed] [Google Scholar]
- Murray M., Reidy G.F. In vitro formation of an inhibitory complex between an isosafrole metabolite and rat hepatic cytochrome P-450 PB-B. Drug Metabolism and Disposition. 1989;17(4):449–454. [PubMed] [Google Scholar]
- Murray M., Wilkinson C.F., Marcus C., Dube C.E. Structure-activity relationships in the interactions of alkoxymethylenedioxybenzene derivatives with rat hepatic microsomal mixed-function oxidases in vivo. Molecular Pharmacology. 1983;24(1):129–136. [PubMed] [Google Scholar]
- Nakagawa H., Tsuta K., Kiuchi K., Senzaki H., Tanaka K., Hioki K., Tsubura A. Growth inhibitory effects of diallyl disulfide on human breast cancer cell lines. Carcinogenesis. 2001;22(6):891–897. doi: 10.1093/carcin/22.6.891. [DOI] [PubMed] [Google Scholar]
- Nebert D.W., Russell D.W. Clinical importance of the cytochromes P450. Lancet. 2002;360(9340):1155–1162. doi: 10.1016/S0140-6736(02)11203-7. [DOI] [PubMed] [Google Scholar]
- Nelson, D., 2003. Cytochrome P450 Homepage: http://drnelson.utmem.edu/CytochromeP450.html. [DOI] [PMC free article] [PubMed]
- Nelson D.R., Koymans L., Kamataki T., Stegeman J.J., Feyereisen R., Waxman D.J., Waterman M.R., Gotoh O., Coon M.J., Estabrook R.W., Gunsalus I.C., Nebert D.W. P450 superfamily: update on new sequences, gene mapping, accession numbers and nomenclature. Pharmacogenetics. 1996;6(1):1–42. doi: 10.1097/00008571-199602000-00002. [DOI] [PubMed] [Google Scholar]
- Niles R.M., McFarland M., Weimer M.B., Redkar A., Fu Y.M., Meadows G.G. Resveratrol is a potent inducer of apoptosis in human melanoma cells. Cancer Letters. 2003;190(2):157–163. doi: 10.1016/s0304-3835(02)00676-6. [DOI] [PubMed] [Google Scholar]
- Nobata S., Ohira T., Nagae H., Ushiyama T., Suzuki K., Fujita K. [Licorice-induced pseudoaldosteronism in a patient with a non-functioning adrenal tumor] Hinyokika Kiyo. 2001;47(9):633–635. [PubMed] [Google Scholar]
- Nortier J.L., Vanherweghem J.L. Renal interstitial fibrosis and urothelial carcinoma associated with the use of a Chinese herb (Aristolochia fangchi) Toxicology. 2002;181–182:577–580. doi: 10.1016/s0300-483x(02)00486-9. [DOI] [PubMed] [Google Scholar]
- Ofir R., Tamir S., Khatib S., Vaya J. Inhibition of serotonin re-uptake by licorice constituents. Journal of Molecular Neuroscience. 2003;20(2):135–140. doi: 10.1385/JMN:20:2:135. [DOI] [PubMed] [Google Scholar]
- Omiecinski C.J., Remmel R.P., Hosagrahara V.P. Concise review of the cytochrome P450s and their roles in toxicology. Toxicological Sciences. 1999;48(2):151–156. doi: 10.1093/toxsci/48.2.151. [DOI] [PubMed] [Google Scholar]
- Orallo F., Alvarez E., Camina M., Leiro J.M., Gomez E., Fernandez P. The possible implication of trans-resveratrol in the cardioprotective effects of long-term moderate wine consumption. Molecular Pharmacology. 2002;61(2):294–302. doi: 10.1124/mol.61.2.294. [DOI] [PubMed] [Google Scholar]
- Ortiz de Montellano P.R., Correia M.A. Inhibition of cytochrome P450. In: Ortiz de Montellano P.R., editor. Cytochrome P450: structure, mechanism and biochemistry. Plenum Press; New York: 1995. pp. 305–366. [Google Scholar]
- Osawa Y., Pohl L.R. Covalent bonding of the prosthetic heme to protein: a potential mechanism for suicide inactivation or activation of hemoproteins. Chemical Research and Toxicology. 1989;2(3):131–141. doi: 10.1021/tx00009a001. [DOI] [PubMed] [Google Scholar]
- Oshima Y., Namao K., Kamijou A., Matsuoka S., Nakano M., Terao K., Ohizumi Y. Powerful hepatoprotective and hepatotoxic plant oligostilbenes, isolated from the Oriental medicinal plant Vitis coignetiae (Vitaceae) Experientia. 1995;51(1):63–66. doi: 10.1007/BF01964921. [DOI] [PubMed] [Google Scholar]
- Owen R.W., Giacosa A., Hull W.E., Haubner R., Wurtele G., Spiegelhalder B., Bartsch H. Olive-oil consumption and health: the possible role of antioxidants. Lancet Oncology. 2000;1:107–112. doi: 10.1016/s1470-2045(00)00015-2. [DOI] [PubMed] [Google Scholar]
- Owuor E.D., Kong A.N.T. Antioxidants and oxidants regulated signal transduction pathways. Biochemical Pharmacology. 2002;64(5–6):765–770. doi: 10.1016/s0006-2952(02)01137-1. [DOI] [PubMed] [Google Scholar]
- Pace-Asciak C.R., Hahn S., Diamandis E.P., Soleas G., Goldberg D.M. The red wine phenolics trans-resveratrol and quercetin block human platelet aggregation and eicosanoid synthesis: implications for protection against coronary heart disease. Clinica Chimica Acta. 1995;235(2):207–219. doi: 10.1016/0009-8981(95)06045-1. [DOI] [PubMed] [Google Scholar]
- Park K.K., Chun K.S., Yook J.I., Surh Y.J. Lack of tumor promoting activity of capsaicin, a principal pungent ingredient of red peppers, in mouse skin carcinogens. Anticancer Research. 1998;18(6A):4201–4205. [PubMed] [Google Scholar]
- Parrish J.A., Fitzpatrick T.B., Tanenbaum L., Pathak M.A. Photochemotherapy of psoriasis with oral methoxsalen and longwave ultraviolet light. New England Journal of Medicine. 1974;291(23):1207–1211. doi: 10.1056/NEJM197412052912301. [DOI] [PubMed] [Google Scholar]
- Pfau W., Schmeiser H.H., Wiessler M. Aristolochic acid binds covalently to the exocyclic amino group of purine nucleotides in DNA. Carcinogenesis. 1990;11(2):313–319. doi: 10.1093/carcin/11.2.313. [DOI] [PubMed] [Google Scholar]
- Piozzi F., Rodriguez B., Savona G. Advances in the chemistry of the furano-diterpenoids from Teucrium species. Heterocycles. 1987;25:807–841. [Google Scholar]
- Pittler M.H., Ernst E. Efficacy of kava extract for treating anxiety: systematic review and meta-analysis. Journal of Clinical Psychopharmacology. 2000;20(2):84–89. doi: 10.1097/00004714-200002000-00014. [DOI] [PubMed] [Google Scholar]
- Piver B., Berthou F., Dreano Y., Lucas D. Inhibition of CYP3A, CYP1A and CYP2E1 activities by resveratrol and other non volatile red wine components. Toxicology Letters. 2001;125(1–3):83–91. doi: 10.1016/s0378-4274(01)00418-0. [DOI] [PubMed] [Google Scholar]
- Piver B., Berthou F., Dreano Y., Lucas D. Differential inhibition of human cytochrome P450 enzymes by ε-viniferin, the dimer of resveratrol: comparison with resveratrol and polyphenols from alcoholized beverages. Life Sciences. 2003;73(9):1199–1213. doi: 10.1016/s0024-3205(03)00420-x. [DOI] [PubMed] [Google Scholar]
- Polymeros D., Kamberoglou D., Tzias V. Acute cholestatic hepatitis caused by Teucrium polium (golden germander) with transient appearance of antimitochondrial antibody. Journal of Clinical Gastroenterology. 2002;34(1):100–101. doi: 10.1097/00004836-200201000-00022. [DOI] [PubMed] [Google Scholar]
- Pozo-Guisado E., Alvarez-Barrientos A., Mulero-Navarro S., Santiago-Josefat B., Fernandez-Salguero P.M. The antiproliferative activity of resveratrol results in apoptosis in MCF-7 but not in MDA-MB-231 human breast cancer cells: cell-specific alteration of the cell cycle. Biochemical Pharmacology. 2002;64(9):1375–1386. doi: 10.1016/s0006-2952(02)01296-0. [DOI] [PubMed] [Google Scholar]
- Premdas P.D., Bowers R.J., Forkert P.G. Inactivation of hepatic CYP2E1 by an epoxide of diallyl sulfone. Journal of Pharmacology and Experimental Therapeutics. 2000;293(3):1112–1120. [PubMed] [Google Scholar]
- Rahman K. Historical perspective on garlic and cardiovascular disease. Journal of Nutrition. 2001;131(3S):977S–979S. doi: 10.1093/jn/131.3.977S. [DOI] [PubMed] [Google Scholar]
- Ramaiah S.K., Apte U., Mehendale H.M. Cytochrome P4502E1 induction increases thioacetamide liver injury in diet-restricted rats. Drug Metabolism and Disposition. 2001;29(8):1088–1095. [PubMed] [Google Scholar]
- Ratna W.N., Simonelli J.A. The action of dietary phytochemicals quercetin, catechin, resveratrol and naringenin on estrogen-mediated gene expression. Life Sciences. 2002;70(13):1577–1589. doi: 10.1016/s0024-3205(01)01531-4. [DOI] [PubMed] [Google Scholar]
- Raulf-Heimsoth M., Baur X. Pathomechanisms and pathophysiology of isocyanate-induced diseases: summary of present knowledge. American Journal of Internal Medicine. 1998;34(2):137–143. doi: 10.1002/(sici)1097-0274(199808)34:2<137::aid-ajim6>3.0.co;2-u. [DOI] [PubMed] [Google Scholar]
- Reddy B.S., Rao C.V., Rivenson A., Kelloff G. Chemoprevention of colon carcinogenesis by organosulfur compounds. Cancer Research. 1993;53(15):3493–3498. [PubMed] [Google Scholar]
- Reilly C.A., Ehlhardt W.J., Jackson D.A., Kulanthaivel P., Mutlib A.E., Espina R.J., Moody D.E., Crouch D.J., Yost G.S. Metabolism of capsaicin by cytochrome P450 produces novel dehydrogenated metabolites and decreases cytotoxicity to lung and liver cells. Chemical Research and Toxicology. 2003;16(3):336–349. doi: 10.1021/tx025599q. [DOI] [PubMed] [Google Scholar]
- Rendic S. Summary of information on human CYP enzymes: human P450 metabolism data. Drug Metabolism Reviews. 2002;34(1–2):83–448. doi: 10.1081/dmr-120001392. [DOI] [PubMed] [Google Scholar]
- Rendic S., Di Carlo F.J. Human cytochrome P450 enzyme: a status report summarizing their reactions, substrates, induction, and inhibitors. Drug Metabolism Reviews. 1997;29(1–2):413–580. doi: 10.3109/03602539709037591. [DOI] [PubMed] [Google Scholar]
- Roche M., Zampelas A., Knapper J.M.E., Kafatos A., Webb D., Brooks C., Jackson K.G., Gibney M.J., Williams C.M. Effect of long-term olive oil dietary intervention on postprandial triacylglycerol and factor VII metabolism. American Journal of Clinical Nutrition. 1998;68(3):552–560. doi: 10.1093/ajcn/68.3.552. [DOI] [PubMed] [Google Scholar]
- Roemer K., Mahyar-Roemer M. The basis for the chemopreventive action of resveratrol. Drugs Today. 2002;38(8):571–580. doi: 10.1358/dot.2002.38.8.820097. [DOI] [PubMed] [Google Scholar]
- Roman V., Billard C., Kern C., Ferry-Dumazet H., Izard J.C., Mohammad R., Mossalayi D.M., Kolb J.P. Analysis of resveratrol-induced apoptosis in human B-cell chronic leukaemia. British Journal of Haematology. 2002;117(4):842–851. doi: 10.1046/j.1365-2141.2002.03520.x. [DOI] [PubMed] [Google Scholar]
- Rosenblat M., Belinky P., Vaya J., Levy R., Hayek T., Coleman R., Merchav S., Aviram M. Macrophage enrichment with the isoflavan glabridin inhibits NADPH oxidase-induced cell-mediated oxidation of low density lipoprotein: a possible role for protein kinase C. Journal of Biological Chemistry. 1999;274(20):13790–13799. doi: 10.1074/jbc.274.20.13790. [DOI] [PubMed] [Google Scholar]
- Rouse J. Kava: a South Pacific herb for anxiety, tension and insomnia. Clinical Nutrition Insights. 1998;963:900–3905. [Google Scholar]
- Salih A.K., Fentiman I.S. Breast cancer prevention: Present and future. Cancer Treatment Reviews. 2001;27(5):261–273. doi: 10.1053/ctrv.2001.0235. [DOI] [PubMed] [Google Scholar]
- Scharf G., Prustomersky S., Knasmuller S., Schulte-Hermann R., Huber W.W. Enhancement of glutathione and γ-glutamylcysteine synthetase, the rate limiting enzyme of glutathione synthesis, by chemoprotective plant-derived food and beverage components in the human hepatoma cell line HepG2. Nutrition and Cancer. 2003;45(1):74–83. doi: 10.1207/S15327914NC4501_9. [DOI] [PubMed] [Google Scholar]
- Schmeiser H.H., Bieler C.A., Wiessler M., van Ypersele de Strihou C., Cosyns J.P. Detection of DNA adducts formed by aristolochic acid in renal tissue from patients with Chinese herbs nephropathy. Cancer Research. 1996;56(9):2025–2028. [PubMed] [Google Scholar]
- Schmeiser H.H., Frei E., Wiessler M., Stiborova M. Comparison of DNA adduct formation by aristolochic acids in various in vitro activation systems by 32P-post-labelling: evidence for reductive activation by peroxidases. Carcinogenesis. 1997;18(5):1055–1062. doi: 10.1093/carcin/18.5.1055. [DOI] [PubMed] [Google Scholar]
- Schmeiser H.H., Janssen J.W., Lyons J., Scherf H.R., Pfau W., Buchmann A., Bartram C.R., Wiessler M. Aristolochic acid activates ras genes in rat tumors at deoxyadenosine residues. Cancer Research. 1990;50(17):5464–5469. [PubMed] [Google Scholar]
- Schmeiser H.H., Scherf H.R., Wiessler M. Activating mutations at codon 61 of the c-Ha-ras gene in thin-tissue sections of tumors induced by aristolochic acid in rats and mice. Cancer Letters. 1991;59(2):139–143. doi: 10.1016/0304-3835(91)90178-k. [DOI] [PubMed] [Google Scholar]
- Schwedhelm E., Maas R., Troost R., Boger R. Clinical pharmacokinetics of antioxidants and their impact on systemic oxidative stress. Clinical Pharmacokinetics. 2003;42(5):437–459. doi: 10.2165/00003088-200342050-00003. [DOI] [PubMed] [Google Scholar]
- Schwetz B.A. From the Food and Drug Administration. Journal of American Medical Association. 2001;285(21):2705. [PubMed] [Google Scholar]
- Shibata S. [A drug over the millennia: pharmacognosy, chemistry, and pharmacology of licorice] Yakugaku Zasshi. 2000;120(10):849–862. doi: 10.1248/yakushi1947.120.10_849. [DOI] [PubMed] [Google Scholar]
- Shimada T., Hayes C.L., Yamazaki H., Amin S., Hecht S.S., Guengerich F.P., Sutter T.R. Activation of chemically diverse procarcinogens by human cytochrome P-450 1B1. Cancer Research. 1996;56(13):2979–2984. [PubMed] [Google Scholar]
- Shimada T., Iwasaki M., Martin M.V., Guengerich F.P. Human liver microsomal cytochrome P-450 enzymes involved in the bioactivation of procarcinogens detected by umu gene response in Salmonella typhimurium TA 1535/pSK1002. Cancer Research. 1989;49(12):3218–3223. [PubMed] [Google Scholar]
- Shon J.H., Park J.Y., Kim M.S., Cha I.J., Chun B.H., Shin J.G. Effect of licorice (Radix glycyrrhizae) on the pharmacokinetics and pharmacodynamics of midazolam in healthy subjects (abstract) Clinical Pharmacology and Therapeutics. 2001;69:P78. [Google Scholar]
- Shureiqi I., Reddy P., Brenner D.E. Chemoprevention: General perspective. Critical Review in Oncology and Hematology. 2000;33(3):157–167. doi: 10.1016/s1040-8428(99)00072-4. [DOI] [PubMed] [Google Scholar]
- Sigusch H., Henschel L., Kraul H., Merkel U., Hoffmann A. Lack of effect of grapefruit juice on diltiazem bioavailability in normal subjects. Pharmazie. 1994;49(9):675–679. [PubMed] [Google Scholar]
- Silverman R.B. CRC Press; Boca Raton, FL: 1998. Mechanism-based enzyme inactivation: chemistry and enzymology. [Google Scholar]
- Singh S., Asad S.F., Ahmad A., Khan N.U., Hadi S.M. Oxidative DNA damage by capsaicin and dihydrocapsaicin in the presence of Cu(II) Cancer Letters. 2001;169(2):139–146. doi: 10.1016/s0304-3835(01)00544-4. [DOI] [PubMed] [Google Scholar]
- Singh U.P., Prithiviraj B., Sarma B.K., Singh M., Ray A.B. Role of garlic (Allium sativum L.) in human and plant diseases. Indian Journal of Experimental Biology. 2001;39(4):310–322. [PubMed] [Google Scholar]
- Singh Y.N., Singh N.N. Therapeutic potential of kava in the treatment of anxiety disorders. CNS Drugs. 2002;16(11):731–743. doi: 10.2165/00023210-200216110-00002. [DOI] [PubMed] [Google Scholar]
- Smith T.J., Guo Z., Guengerich F.P., Yang C.S. Metabolism of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) by human cytochrome P450 1A2 and its inhibition by phenethyl isothiocyanate. Carcinogenesis. 1996;17(4):809–813. doi: 10.1093/carcin/17.4.809. [DOI] [PubMed] [Google Scholar]
- Smith T.J., Guo Z., Li C., Ning S.M., Thomas P.E., Yang C.S. Mechanisms of inhibition of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone bioactivation in mouse by dietary phenethyl isothiocyanate. Cancer Research. 1993;53(14):3276–3282. [PubMed] [Google Scholar]
- Spigelski D., Jones P.J. Efficacy of garlic supplementation in lowering serum cholesterol levels. Nutrition Reviews. 2001;59(7):236–241. doi: 10.1111/j.1753-4887.2001.tb07015.x. [DOI] [PubMed] [Google Scholar]
- Sporn M.B., Suh N. Chemoprevention of cancer. Carcinogenesis. 2000;21:525–530. doi: 10.1093/carcin/21.3.525. [DOI] [PubMed] [Google Scholar]
- Staretz M.E., Hecht S.S. Effects of phenethyl isothiocyanate on the tissue distribution of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone and metabolites in F344 rats. Cancer Research. 1995;55(23):5580–5588. [PubMed] [Google Scholar]
- Stedman C. Herbal hepatotoxicity. Seminars in Liver Disease. 2002;22(2):195–206. doi: 10.1055/s-2002-30104. [DOI] [PubMed] [Google Scholar]
- Stevinson C., Huntley A., Ernst E. A systematic review of the safety of kava extract in the treatment of anxiety. Drug Safety. 2002;25(4):251–261. doi: 10.2165/00002018-200225040-00003. [DOI] [PubMed] [Google Scholar]
- Stiborova M., Frei E., Breuer A., Bieler C.A., Schmeiser H.H. Aristolactam I a metabolite of aristolochic acid I upon activation forms an adduct found in DNA of patients with Chinese herbs nephropathy. Experimental Toxicology and Pathology. 1999;51(4–5):421–427. doi: 10.1016/S0940-2993(99)80033-5. [DOI] [PubMed] [Google Scholar]
- Stiborova M., Frei E., Breuer A., Wiessler M., Schmeiser H.H. Evidence for reductive activation of carcinogenic aristolochic acids by prostaglandin H synthase—32P-postlabeling analysis of DNA adduct formation. Mutation Research. 2001;493(1–2):149–160. doi: 10.1016/s1383-5718(01)00171-1. [DOI] [PubMed] [Google Scholar]
- Stiborova M., Frei E., Sopko B., Wiessler M., Schmeiser H.H. Carcinogenic aristolochic acids upon activation by DT-diaphorase form adducts found in DNA of patients with Chinese herbs nephropathy. Carcinogenesis. 2002;23(4):617–625. doi: 10.1093/carcin/23.4.617. [DOI] [PubMed] [Google Scholar]
- Stiborova M., Frei E., Wiessler M., Schmeiser H.H. Human enzymes involved in the metabolic activation of carcinogenic aristolochic acids: evidence for reductive activation by cytochromes P450 1A1 and 1A2. Chemical Research and Toxicology. 2001;14(8):1128–1137. doi: 10.1021/tx010059z. [DOI] [PubMed] [Google Scholar]
- Stupans I., Murray M., Kirlich A., Tuck K.L., Hayball P.J. Inactivation of cytochrome P450 by the food-derived complex phenol oleuropein. Food and Chemical Toxicology. 2001;39(11):1119–1124. doi: 10.1016/s0278-6915(01)00060-6. [DOI] [PubMed] [Google Scholar]
- Stupans I., Stretch G., Hayball P. Olive oil phenols inhibit human hepatic microsomal activity. Journal of Nutrition. 2000;130(9):2367–2370. doi: 10.1093/jn/130.9.2367. [DOI] [PubMed] [Google Scholar]
- Surh Y.J., Lee E., Lee J.M. Chemopreventive properties of some pungent ingredients present in red pepper and ginger. Mutation Research. 1998;402(1–2):259–267. doi: 10.1016/s0027-5107(97)00305-9. [DOI] [PubMed] [Google Scholar]
- Surh Y.J., Lee S.S. Capsaicin, a double edged sword: toxicity, metabolism, and chemopreventative potential (Review) Life Sciences. 1995;56(22):1845–1855. doi: 10.1016/0024-3205(95)00159-4. [DOI] [PubMed] [Google Scholar]
- Tamimi R.M., Lagiou P., Adami H.O., Trichopoulos D. Prospects for chemoprevention of cancer. Journal of Internal Medicine. 2002;251(4):286–300. doi: 10.1046/j.1365-2796.2002.00969.x. [DOI] [PubMed] [Google Scholar]
- Tamir S., Eizenberg M., Somjen D., Izrael S., Vaya J. Estrogen-like activity of glabrene and other constituents isolated from licorice root. Journal of Steroid Biochemistry and Molecular Biology. 2001;78(3):291–298. doi: 10.1016/s0960-0760(01)00093-0. [DOI] [PubMed] [Google Scholar]
- Teyssier C., Guenot L., Suschetet M., Siess M.H. Metabolism of diallyl disulfide by human liver microsomal cytochromes P-450 and flavin-containing monooxygenases. Drug Metabolism and Disposition. 1999;27(7):835–841. [PubMed] [Google Scholar]
- Thomassen D., Knebel N., Slattery J.T., McClanahan R.H., Nelson S.D. Reactive intermediates in the oxidation of menthofuran by cytochromes P-450. Chemical Research and Toxicology. 1992;5(1):123–130. doi: 10.1021/tx00025a021. [DOI] [PubMed] [Google Scholar]
- Thomassen D., Pearson P.G., Slattery J.T., Nelson S.D. Partial characterization of biliary metabolites of pulegone by tandem mass spectrometry. Drug Metabolism and Disposition. 1991;19(5):997–1003. [PubMed] [Google Scholar]
- Thompson D.C., Perera K., Fisher R., Brendel K. Cresol Isomers: comparison of toxic potency in rat liver slices. Toxicology and Applied Pharmacology. 1994;125(1):51–58. doi: 10.1006/taap.1994.1048. [DOI] [PubMed] [Google Scholar]
- Tinel M., Belghiti J., Descatoire V., Amouyal G., Letteron P., Geneve J., Larrey D., Pessayre D. Inactivation of human liver cytochrome P-450 by the drug methoxsalen and other psoralen derivatives. Biochemical Pharmacology. 1987;36(6):951–955. doi: 10.1016/0006-2952(87)90190-0. [DOI] [PubMed] [Google Scholar]
- Toth B., Gannett P. Carcinogenicity of lifelong administration of capsaicin of hot pepper in mice. In Vivo. 1992;6(1):59–63. [PubMed] [Google Scholar]
- Tsarbopoulos A., Gikas E., Papadopoulos N., Aligiannis N., Kafatos A. Simultaneous determination of oleuropein and its metabolites in plasma by high-performance liquid chromatography. Journal of Chromatography B - Analytical Technologies in the Biomedical and Life Sciences. 2003;785(1):157–164. doi: 10.1016/s1570-0232(02)00905-4. [DOI] [PubMed] [Google Scholar]
- Tuck K.L., Hayball P.J. Major phenolic compounds in olive oil: metabolism and health effects. Journal of Nutritional Biochemistry. 2002;13(11):636–644. doi: 10.1016/s0955-2863(02)00229-2. [DOI] [PubMed] [Google Scholar]
- Unger M., Holzgrabe U., Jacobsen W., Cummins C., Benet L.Z. Inhibition of cytochrome P450 3A4 by extracts and kavalactones of Piper methysticum (Kava-Kava) Planta Medica. 2002;68(12):1055–1058. doi: 10.1055/s-2002-36360. [DOI] [PubMed] [Google Scholar]
- Villegas J.F., Barabe D.N., Stein R.A., Lazar E. Adverse effects of herbal treatment of cardiovascular disease: what the physician must know. Heart Disease. 2001;3(3):169–175. doi: 10.1097/00132580-200105000-00007. [DOI] [PubMed] [Google Scholar]
- Vincent J., Harris S.I., Foulds G., Dogolo L.C., Willavize S., Friedman H.L. Lack of effect of grapefruit juice on the pharmacokinetics and pharmacodynamics of amlodipine. British Journal of Clinical Pharmacology. 2000;50(5):455–463. doi: 10.1046/j.1365-2125.2000.00283.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visiol F., Galli C. Biological properties of olive oil phytochemicals. Critical Review in Food Sciences and Nutrition. 2002;42(3):209–221. doi: 10.1080/10408690290825529. [DOI] [PubMed] [Google Scholar]
- Visioli F., Bellomo G., Montedoro G., Galli C. Low density lipoprotein oxidation is inhibited in vitro by olive oil constituents. Atherosclerosis. 1995;117(1):25–32. doi: 10.1016/0021-9150(95)05546-9. [DOI] [PubMed] [Google Scholar]
- Visioli F., Galli C. Oleuropein protects low density lipoprotein from oxidation. Life Sciences. 1994;55(24):1965–1971. doi: 10.1016/0024-3205(94)00529-x. [DOI] [PubMed] [Google Scholar]
- Visioli F., Poli A., Gall C. Antioxidant and other biological activities of phenols from olives and olive oil. Medical Research Reviews. 2002;22(1):65–75. doi: 10.1002/med.1028. [DOI] [PubMed] [Google Scholar]
- Volz H.P., Kieser M. Kava-kava extract WS 1490 versus placebo in anxiety disorders—a randomized placebo-controlled 25-week outpatient trial. Pharmacopsychiatry. 1997;30(1):1–5. doi: 10.1055/s-2007-979474. [DOI] [PubMed] [Google Scholar]
- Wallerath T., Deckert G., Ternes T., Anderson H., Li H., Witte K., Forstermann U. Resveratrol, a polyphenolic phytoalexin present in red wine, enhances expression and activity of endothelial nitric oxide synthase. Circulation. 2002;106(13):1652–1658. doi: 10.1161/01.cir.0000029925.18593.5c. [DOI] [PubMed] [Google Scholar]
- Wang B.H., Zuzel K.A., Rahman K., Billington D. Treatment with aged garlic extract protects against bromobenzene toxicity to precision cut rat liver slices. Toxicology. 1999;132(2–3):215–225. doi: 10.1016/s0300-483x(99)00004-9. [DOI] [PubMed] [Google Scholar]
- Wang T., Shankar K., Bucci T.J., Warbritton A., Mehendale H.M. Diallyl sulfide inhibition of CYP2E1 does not rescue diabetic rats from thioacetamide-induced mortality. Toxicology and Applied Pharmacology. 2001;173(1):27–37. doi: 10.1006/taap.2001.9165. [DOI] [PubMed] [Google Scholar]
- Wang T., Shankar K., Ronis M.J.J., Mehendale H.M. Potentiation of thioacetamide liver injury in diabetic rats is due to induced CYP2E1. Journal of Pharmacology and Experimental Therapeutics. 2000;294(2):473–479. [PubMed] [Google Scholar]
- Ward M.H., Lopez-Carrillo L. Dietary factors and the risk of gastric cancer in Mexico City. American Journal of Epidemiology. 1999;149(10):925–932. doi: 10.1093/oxfordjournals.aje.a009736. [DOI] [PubMed] [Google Scholar]
- Wargovich M.J., Woods C., Hollis D.M., Zander M.E. Herbals, cancer prevention and health. Journal of Nutrition. 2001;131(11 Suppl. S):3034S–3036S. doi: 10.1093/jn/131.11.3034S. [DOI] [PubMed] [Google Scholar]
- Watanabe C.M.H., Wolffram S., Ader P., Rimbach G., Packer L., Maguire J.J., Schultz P.G., Gohil K. The in vivo neuromodulatory effects of the herbal medicine Ginkgo biloba. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(12):6577–6580. doi: 10.1073/pnas.111126298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheatley D. Stress-induced insomnia treated with kava and valerian: singly and in combination. Human Psychopharmacology. 2001;16(4):353–356. doi: 10.1002/hup.299. [DOI] [PubMed] [Google Scholar]
- Yang C.S., Brady J.F., Hong J.-Y. Dietary effects on cytochromes P450, xenobiotic metabolism, and toxicity. FASEB (Federation of American Societies of Experimental Biology) Journal. 1992;6(2):737–744. doi: 10.1096/fasebj.6.2.1537464. [DOI] [PubMed] [Google Scholar]
- Yang C.S., Chhabra S.K., Hong J.Y., Smith T.J. Mechanisms of inhibition of chemical toxicity and carcinogenesis by diallyl sulfide (DAS) and related compounds from garlic. Journal of Nutrition. 2001;131(3S):S1041–S1045. doi: 10.1093/jn/131.3.1041S. [DOI] [PubMed] [Google Scholar]
- Yang C.S., Smith T.J., Hong J.-Y. Cytochrome P-450 enzymes as targets for chemoprevention against chemical carcinogenesis and toxicity: opportunities and limitations. Cancer Research. 1994;54(7 Suppl.):S1982–S1986. [PubMed] [Google Scholar]
- Yang S.H., Kim J.S., Oh T.J., Kim M.S., Lee S.W., Woo S.K., Cho H.S., Choi Y.H., Kim Y.H., Rha S.Y., Chung H.C., An S.W. Genome-scale analysis of resveratrol-induced gene expression profile in human ovarian cancer cells using a cDNA microarray. International Journal of Oncology. 2003;22(4):741–750. [PubMed] [Google Scholar]
- Yang Y.M., Conaway C.C., Chiao J.W., Wang C.X., Amin S., Whysner J., Dai W., Reinhardt J., Chung F.L. Inhibition of benzo[a]pyrene-induced lung tumorigenesis in A/J mice by dietary N-acetylcysteine conjugates of benzyl and phenethyl isothiocyanates during the postinitiation phase is associated with activation of mitogen-activated protein kinases and p53 activity and induction of apoptosis. Cancer Research. 2002;62(1):2–7. [PubMed] [Google Scholar]
- Yee G.C., Stanley D.L., Pessa J.L., Costa T.D., Beltz S.E., Ruiz J., Lowenthal D.T. Effect of grapefruit juice on blood cyclosporin concentration. Lancet. 1995;345(8955):955–956. doi: 10.1016/s0140-6736(95)90700-9. [DOI] [PubMed] [Google Scholar]
- Young R.C., Wilson C.M. Cancer prevention: Past, present, and future. Clinical Cancer Research. 2002;8(1):11–16. [PubMed] [Google Scholar]
- Zaacks S.M., Klein L., Tan C.D., Rodriguez E.R., Leikin J.B. Hypersensitivity myocarditis associated with ephedra use. Journal of Toxicology and Clinical Toxicology. 1999;37(4):485–489. doi: 10.1081/clt-100102440. [DOI] [PubMed] [Google Scholar]
- Zhang Y., Talalay P. Anticarcinogenic activities of organic isothiocyanates: chemistry and mechanisms. Cancer Research. 1994;54(7 Suppl.):S1976–S1981. [PubMed] [Google Scholar]
- Zheng S., Yang H., Zhang S., Wang X., Yu L.L., Lu J.Q., Li J. Initial study on naturally occurring products from traditional Chinese herbs and vegetables for chemoprevention. Journal of Cellular Biochemistry. 1997;27(Suppl.):106–112. [PubMed] [Google Scholar]
- Zhou S.F. Drug bioactivation and covalent binding to target proteins and toxicity relevance. Drug Metabolism Reviews. 2003 doi: 10.1081/dmr-200028812. (in press) [DOI] [PubMed] [Google Scholar]
- Zhou S.F., Gao Y.H., Wen Q.J., Huang M., Xu A.L., Paxton J.W. Interactions of herbs with cytochrome P450. Drug Metabolism Reviews. 2003;35(1):35–98. doi: 10.1081/dmr-120018248. [DOI] [PubMed] [Google Scholar]
- Zhu Y.P. Toxicity of the Chinese herb Mu tong (Aristolochia manshuriensis): What history tells us. Adverse Drug Reactions and Toxicology Reviews. 2002;21(4):171–177. doi: 10.1007/BF03256194. [DOI] [PubMed] [Google Scholar]
- Zou L., Harkey M.R., Henderson G.L. Effects of herbal components on cDNA-expressed cytochrome P450 enzyme catalytic activity. Life Sciences. 2002;71(13):1579–1589. doi: 10.1016/s0024-3205(02)01913-6. [DOI] [PubMed] [Google Scholar]