

Highlights
-
•
A 2.5 kb chicken MDA5 promoter fragment was cloned and characterized (Accession number KT335979).
-
•
The activity of the promoter is extremely low under basal conditions.
-
•
Viral (IBDV) and non-viral (IFN-β or poly (I:C) inducers could activate the promoter.
-
•
Gene expression driven by exogenous MDA5 promoter is in accordance with endogenous MDA5.
Keywords: MDA5, Promoter, Chicken, Innate immunology, RIG-I-like receptor (RLR), Virus
Abstract
Melanoma differentiation-associated gene 5 (MDA5) is a member of the retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) family and plays a pivotal role in the anti-viral innate immune response. As RIG-I is absent in chickens, MDA5 is hypothesized to be important in detecting viral nucleic acids in the cytoplasm. However, the molecular mechanism of the regulation of chicken MDA5 (chMDA5) expression has yet to be fully elucidated. With this in mind, a ∼2.5 kb chMDA5 gene promoter region was examined and PCR amplified to assess its role in immune response. A chMDA5 promoter reporter plasmid (piggybac-MDA5-DsRed) was constructed and transfected into DF-1 cells to establish a Piggybac-MDA5-DsRed cell line. The MDA5 promoter activity was extremely low under basal condition, but was dramatically increased when cells were stimulated with polyinosinic: polycytidylic acid (poly I:C), interferon beta (IFN-β) or Infectious Bursal Disease Virus (IBDV). The DsRed mRNA level represented the promoter activity and was remarkably increased, which matched the expression of endogenous MDA5. However, Infectious Bronchitis Virus (IBV) and Newcastle disease virus (NDV) failed to increase the MDA5 promoter activity and the expression of endogenous MDA5. The results indicated that the promoter and the Piggybac-MDA5-DsRed cell line could be utilized to determine whether a ligand regulates MDA5 expression. For the first time, this study provides a tool for testing chMDA5 expression and regulation.
1. Introduction
The innate immune system acts as the first defensive barrier against invading pathogens, using pattern recognition receptors to recognize viral pathogen-associated molecular patterns (PAMPs). Pattern recognition receptors consist of toll-like receptor, nucleotide-binding oligomerization domain protein-like receptors and retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) (Janeway and Medzhitov, 2002, Meylan et al., 2006). The RLR family is composed of RIG-I, melanoma differentiation-associated gene 5 (MDA5) and laboratory of genetics and physiology 2, all of which play important roles in recognizing viral RNA in the cytoplasm (Yoneyama and Fujita, 2010). RIG-I and MDA5 both harbor a central DExD/H-box RNA helicase domain, two N-terminal caspase activation and recruitment domains (CARDs), and a C-terminal regulatory domain. The RNA helicase domain is responsible for ATP hydrolysis (Yoneyama et al., 2005, Yoneyama et al., 2004). The C-terminal regulatory domain is essential for binding viral RNA (Cui et al., 2008, Takahasi et al., 2009). The two CARDs function via CARD–CARD interactions with interferon promoter-stimulating factor 1, leading to the activation of interferon regulatory factor 3 and nuclear factor kappa-light chain-enhancer of activated B cells (Belgnaoui et al., 2011), which finally induces type I interferon (IFN-I) production and triggers a series of initial antiviral activities (Stark et al., 1998). During viral infection, swift upregulation of IFN-I is critical to the host antiviral responses (Bracci et al., 2005). Specifically, the expression of RIG-I and MDA5 are greatly induced by IFN-I as a positive-feedback mechanism, which results in signaling cascade to promote antiviral response (Gack et al., 2008).
In mammals, RIG-I and MDA5 recognize distinct, but overlapping, families of RNA viruses (Kawai and Akira, 2007, Ramos and Gale, 2011). However, chickens lack RIG-I and this may provide a rationale for increased susceptibility to certain pathogens in comparison to ducks and mammals, such as Highly Pathogenic Avian Influenza (Barber et al., 2010, Karpala et al., 2011). This suggests that chicken MDA5 (chMDA5) may have an important role in chicken during viral invasion. Unlike ducks and mammals, chMDA5 is able to recognize both long and short poly (I:C) and induce IFN-I beta (IFN-β) expression (Hayashi et al., 2014, Karpala et al., 2011) and the expression of chMDA5 is upregulated and plays a role during infection with influenza. Interestingly, the expression of chMDA5 does not seem to be sufficient to prevent influenza proliferation (Karpala et al., 2011). A plethora of research exists pertaining to MDA5 function in mammals, little is known about the role of chMDA5 and the ligands that interact with chMDA5. Similar to mammalian MDA5, the expression of chMDA5 is enhanced by IFN-β (Kang et al., 2002, Lee et al., 2012), but the molecular mechanism is not yet known. A greater understanding of the transcriptional regulation of chMDA5 is essential to elucidating the role of MDA5 in chicken antiviral response.
In the present study, we cloned and characterized a 2.5 kb promoter of chMDA5. This promoter was inserted into a reporter plasmid and transfected into chicken cells to generate a cell model which could be used to investigate the promoter activity. The activity of the promoter reflected the expression of endogenous MDA5, indicating that the system could be used to determine potential MDA5 ligands.
2. Materials and methods
2.1. Amplification of the 5′- upstream region of chMDA5 gene
An about 2500 bp of the 5′-flanking region of MDA5 gene was amplified using White Leghorn chicken genomic DNA as the template. Primers (F1 and R1, listed in Table 1 ) were designed according to the sequence published on NCBI GenBank. PCR amplification was performed using KOD-Plus-Neo polymerase (TOYOBO) according to the manufacturer’s instruction. The PCR product was cloned into the pMD19-T vector (Takara) to generate the 19T-MDA5 plasmid and was completely sequenced.
Table 1.
Sequences of primers used.
| Name | Sequence (5′to 3′) | Application |
|---|---|---|
| F1 | TTCTGGAACGCTCTTCTCACA | promoter amplification |
| R1 | GCACCTTGTCCTTCTCCTCT | |
| PciI/BamHIF | GCTCACATGTTTCTGGAACGCTCTTCTCACA | plasmid construction |
| PciI/BamHI R | GGTGGATCCCGCTGCGCTGGCGGCGCCCGGGCCGGCTCTGCTGCT | |
| SfiI/NheI F | GGGGAAAAGGCCTCCAAGGCCTTCTGGAACGCTCTTCTCACA | plasmid construction |
| SfiI/NheI R | ATTCGCTAGCCTTAAGATACATTGATGAGTTTGG | |
| DsRed F | AGTTCCAGTACGGCTCCAAGGT | RT-PCR |
| DsRed R | CAGCCCATGGTCTTCTTCTGCATT | |
| IFN-α F | AACGCCAAAGCCTCCTCAAC | RT-PCR |
| IFN-α R | TTGTCTTGGAGGAAGGTGTG | |
| IFN-β F | TCCTGCAACCATCTTCGTCA | RT-PCR |
| IFN-β R | GCTGCTTGCTTCTTGTCCTT | |
| MDA5 F | CTGGTTGGGATTGAGACTGT | RT-PCR |
| MDA5 R | GTCTTTATCCTCTGTTCCTCCTG | |
| β-actin F | TACGTCGCACTGGATTTCGA | RT-PCR |
| β-actin R | CAGGACTCCATACCCAAGAA |
Seven MDA5 promoter sequences from birds, fish and mammals obtained from the NCBI (http://www.ncbi.nlm.nih.gov/), were compared with the chMDA5 promoter sequence. Turkey was chosen to representative for birds that lack RIG-I and duck for birds that have RIG-I. Zebrafish and the mammals all have RIG-I. The tree was generated using MEGA4.0 by the Bootstrap of Test (neighbor-joining method, 1000 replicates; seed = 58150).
2.2. Construction of chMDA5 promoter reporter plasmid
The promoter was amplified using primers designed with a PciI restriction site on the forward primer and a BamHI restriction site on the reverse primer (Table 1). The PCR reaction was carried out as described above. The amplification product was purified and double digested with restriction enzymes PciI and BamHI. The purified MDA5 promoter was ligated into the pDsRed1-N1 Vector (CLONTECH) that was previously digested with the same restriction enzymes to replace the CMV promoter. This recombinant plasmid is referred to as the MDA5-DsRed plasmid. Primers were designed with a SfiI restriction site on the forward primer and a NheI restriction site on the reverse primer (Table 1) and PCR reaction was carried out using the MDA5-DsRed plasmid as a template to amplify the region containing the MDA5 promoter and the coding region of DsRed. The amplification product was purified and digested with restriction enzymes SfiI and NheI and the purified MDA5-DsRed fragment was ligated into piggybacGFP Plasmid (SBI), which was previously digested with the same restriction enzymes to replace the CMV 7 promoter, generating the piggybac-MDA5-DsRed plasmid.
2.3. Cell culture, transfection and puromycin-selection
DF1, a chicken embryonic fibroblast cell line (Himly et al., 1998), was cultured in Dulbecco’s modified Eagle’s medium (DMEM)(GIBCO) supplemented with 3.7 mg/mL sodium bicarbonate, 1 × MEM Non-Essential Amino Acids (GIBCO), 100 U/mL penicillin, 100 ng/mL streptomycin and 10% fetal bovine serum (FBS) (Hyclone), in a 5% CO2 incubator at 37 °C. The piggybac-MDA5-DsRed plasmids were transfected into DF1 cells using Lipofectamine® LTX & PLUS™ Reagent (Invitrogen) according to the manufacturer’s instructions. 48 h after transfection, 5 μg/mL puromycin was added to the culture media for selection. The continuous cell line was passaged 3–5 times with puromycin selection to obtain a stable cell line containing piggybac-MDA5-DsRed plasmids and that expressed GFP (Piggybac-MDA5-DsRed).
2.4. Poly (I:C), IFN-β and virus challenge
Piggybac-MDA5-DsRed cells were seeded overnight in 12-well plates to 70–80% confluence prior to treatment. For Poly (I:C) challenge, 2.5 μg/mL of long synthetic dsRNA analog poly (I:C) (size:1.5–8 kb) (Invivogen) or 2.5 μg/mL short synthetic dsRNA analog poly(I:C) (size: <1 kb) (Yuanye) was transfected into cells using Lipofectamine® LTX & PLUS™ Reagent (Invitrogen) according to the manufacturer’s instructions. Cells were harvested 24 h post-transfection for fluorescence assay, fluorescence-activated cell sorting (FACS) and qPCR analysis. For IFN-β treatment, 8 μg/mL chicken IFN-β was added into the culture medium for 48 h before the cells were collected for fluorescence assay, FACS and qPCR analysis. For viral challenge, live vaccines of Infectious Bursal Disease Virus (IBDV) NF8, Newcastle Disease Virus (NDV) Lasota and Infectious Bronchitis Virus (IBV) H52, obtained from Qianyuanhao, were diluted with DMEM without FBS and used to replace the culture medium. For the mock treatment, DMEM without FBS was used to replace the culture medium. After 2 h incubation, the diluted virus was replaced with DMEM supplemented with 2% FBS. Cells were observed and collected for analysis 84 h post infection (hpi).
2.5. Gene expression analysis by real-time quantitative PCR
RNA was extracted using Total RNA kit II (OMEGA) followed by reverse transcription using PrimeScript™ RT Reagent Kit with gDNA Eraser (Perfect Real Time; TaKaRa) according to the manufacturer’s instructions. The relative quantitation of gene expression from the Piggybac-MDA5-DsRed cell line following treatment was performed on a LightCycler® 96 (Roche) to determine transcript levels of DsRed, MDA5, IFN-α, IFN-β and β-actin, using the comparative threshold cycle (ΔΔCT) method. 2 μL of cDNA was amplified in a 25 μL reaction using SYBR® Premix Ex Taq II (TliRNaseH Plus; TaKaRa). The corresponding primer sets are listed in Table 1. The relative expression levels of target genes were expressed in terms of CT value normalized to β-actin as described previously (Lee et al., 2014). The fold changes for each gene were normalized to the β-actin expression in each sample. The relative expression of target gene normalized to β-actin is showed as 2−ΔCT, where ΔCT = CT targetgene- CTβ-actin. The relative expression in the treatment group versus that in the mock group was calculated using the formula 2−ΔΔCT, where ΔΔCT = ΔCT targetgene in the treatment group–ΔCT targetgene in the mock group. The geometric mean of the fold change in expression is shown as log2 scale. The mean of ΔΔCT was based on three samples (n = 3).
2.6. Fluorescence activated cell sorting (FACS) analysis
Cells collected for FACS analysis were fixed with 4% paraformaldehyde for 15 min and washed three times using Dulbecco phosphate-buffered saline (SIGMA), then the cell suspensions were prepared in Dulbecco phosphate-buffered saline for FACS analysis. All evaluations were performed on a Moflo XDP flow cytometer (Beckman Coulter) and analyzed using Flow Jo-V10 software.
2.7. Statistical analysis
Values were expressed as means ± standard deviation and statistical significance among different experimental groups were analyzed by one-way ANOVA using SPSS 18.0 software.
3. Result and discussion
3.1. Sequence analysis of the promoter region of the chMDA5 gene
A ∼2.5 kb genomic fragment upstream of the chMDA5 gene was cloned. The fragment sequence was aligned against the 5′-upstream region of chMDA5 gene using the NCBI BLAST Assembled Genomes (http://blast.ncbi.nlm.nih.gov/Blast), indicating 99% sequence homology with 5′-promoter region. The full length sequence of the MDA5 promoter has been submitted to the GeneBank database (accession number KT335979). Sequence analysis using the TFSEARCH ver.8.3 indicated that the MDA5 promoter region contains several putative transcription factor-binding sites, including binding sites for Sp1, GATA-1, TGGCA-binding protein, AP-1, AP-3, ER-alpha, T3R-alpha, C/EBP alpha, NF-1 (-like proteins) and NF-E4.
In order to exploit the evolutionary relationship of promoters between chicken and other species, a phylogenetic analysis was performed. The chMDA5 promoter demonstrated closer evolutionary distance to bird MDA promoters, than to that of mammals. The closest evolutionary distance was to MDA5 promoter of turkey, in which RIG-I are also absent (Fig. 1 A). This result was similar to the phylogenetic analysis of the MDA5 gene in previous studies (Huang et al., 2010, Lee et al., 2012, Su et al., 2010) and is likely a reflection of the taxonomic relationship between these vertebrates. These results will be helpful for study of the innate immune system regarding to MDA5 and application of this promoter in expression of antiviral genes, as promoters with high homology would possibly bonded to similar transcription factors.
Fig. 1.
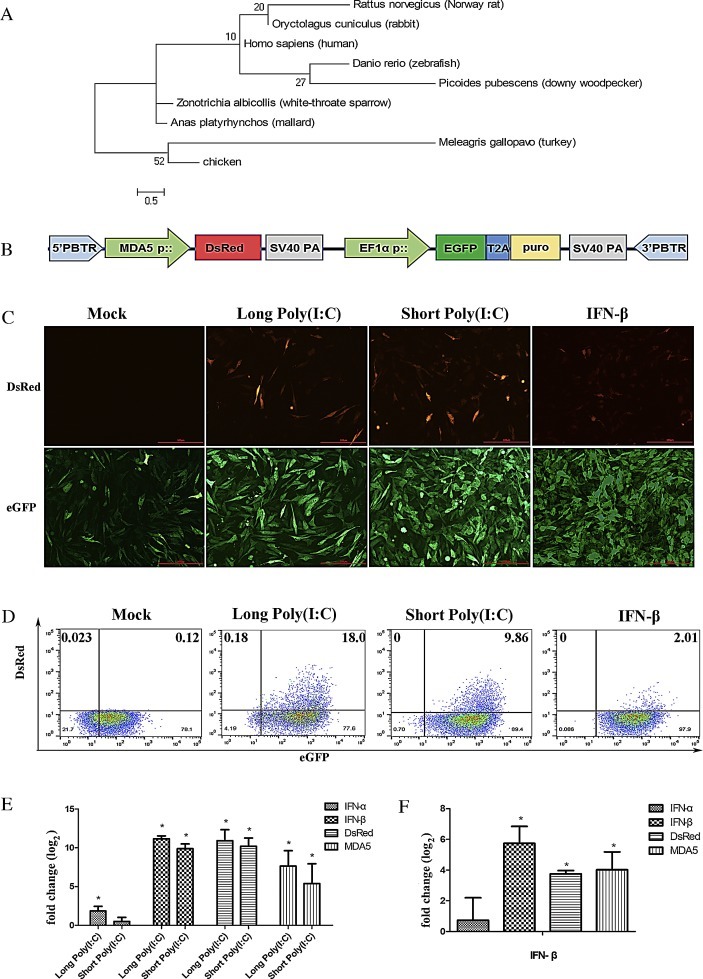
The chMDA5 promoter is dramatically activated in transfected cells that are incubated with poly (I:C) and IFN–β, accompanied by upregulation of endogenous MDA5. (A) Phylogenetic tree of the chMDA5 promoter and MDA5 promoter sequences from other birds, fish and mammals. The tree was generated using MEGA4.0 by bootstrapped phylogenetic analysis (neighbor-joining method, 1000 replicates; seed = 58150). (B) Map of recombinant plasmid piggy-pMDA5-DsRed. (C) Immunofluorescence analysis for DsRed from cells treated with mock, long poly (I:C), short poly (I:C) or IFN–β. Scale bars, 100 μm. (D) FACS analysis of DsRed and eGFP from cells treated with mock, long poly (I:C), short poly (I:C) or IFN–β. The horizontal axis represents eGFP-expressing cells and the vertical represents DsRed-expressing cells. The ratios of DsRed and eGFP positive cells are indicated. (E, F) qRT-PCR analysis of cells treated with poly (I:C) (E) and IFN–β (F). The mRNA expression levels of DsRed, eGFP and endogenous MDA5, IFN-α, IFN-β were analyzed by qRT-PCR. Data are presented as means of three independent experiments. Significance was analyzed with one-way ANOVA. The error bars represent the mean of Standard Error. Asterisk represents significant difference between treatment group and the mock (*p < 0.05).
3.2. Poly (I:C) and IFN-β challenge increases the activity of the chMDA5 promoter and upregulates endogenous MDA5 expression
The chMDA5 promoter reporter plasmid (piggybac-MDA5-DsRed) (Fig. 1B), in which the DsRed expression level represents the promoter activity of chMDA5, was transfected into DF-1 cells to investigate the promoter activity. Fluorescence microscope observations (Fig. 1C) and FACS analysis (Fig. 1D) demonstrated that ∼80% of cultured Piggybac-MDA5-DsRed cells expressed GFP, whereas virtually no DsRed expression could be noted. These results indicated that the MDA5 promoter activity (as observed by DsRed expression) was extremely low under basal conditions.
Previous studies indicated that the chMDA5 recognizes both short and long poly (I:C), with greater preference for short poly (I:C) (Hayashi et al., 2014). However, in the present study, though both short and long poly (I:C) could evoke the activity of the MDA5 promoter, short poly (I:C) caused a lesser increase in expression. DsRed-expressing cells were present 24 h post-transfection with poly (I:C) in Piggybac-MDA5-DsRed cells (Fig. 1C). The proportion of DsRed-expressing cells transfected with long poly (I:C) and short poly (I:C) were 18% and 10%, respectively (Fig. 1D). qRT- PCR results showed that long poly (I:C) dramatically upregulated the DsRed mRNA, which was accompanied by an upregulation of endogenous MDA5, IFN-α and IFN-β. Shor poly (I:C) resulted in a similar effect, although the fold-change was lower for MDA5 and IFN-β and short poly (I:C) stimulation failed to induce IFN-α expression (Fig. 1E). Previous results in mda-5−/− mice demonstrated that IFN-β response to poly (I:C) was abrogated, while IFN-α response was lower than the wild-type mice, suggesting that IFN-α was partially independent of MDA5 (Gitlin et al., 2006). It is possible that the upregulation of MDA5 by short poly (I:C) is not enough to induce IFN-α in chicken, as the results in this study indicate that chMDA5 preferentially senses long poly (I:C).
As MDA5 is an IFN-inducible gene which primarily responds to IFN-β (Kang et al., 2002), the MDA5 promoter response to IFN-β was evaluated. The activity of the chMDA5 promoter was increased after 48 h post-treatment with IFN-β, as observed when cells began to express DsRed (Fig. 1C). The rate of the DsRed-expressing cells was 2% (Fig. 1D). qRT- PCR results showed that the mRNA level of DsRed was significantly enhanced by IFN-β (p < 0.05), as well as endogenous MDA5 (p < 0.05) and IFN-β (p < 0.05) levels. However, IFN-α expression did not change significantly (Fig. 1F). The promoter activity was stimulated by IFN-β, with a concurrent increase in MDA5 expression, which is similar to previous reports (Karpala et al., 2011). This confirms that the promoter responds to non-viral inducers, as an indicator of the changes in endogenous MDA5 expression.
3.3. IBDV challenge activates the chMDA5 promoter along with upregulation of endogenous MDA5
Several different viruses (IBDV, NDV and IBV) were used to infect Piggybac-MDA5-DsRed cells, but only IBDV was able to activate the chMDA5 promoter (1% DsRed expression). Cellular expression of DsRed was coupled with apoptosis that was caused by IBDV infection. However, few cells infected with NDV (0.051%) and IBV (0.12%) infection expressed DsRed (Fig. 2A,B ). The level of DsRed expression was remarkably increased by the infection with IBDV (p < 0.05), as were the mRNA levels of endogenous MDA5 (p < 0.05), IFN-α (p < 0.05) and IFN-β (p < 0.05). In contrast, infection with NDV or IBV had no significant effect on the mRNA levels of DsRed, endogenous MDA5 or IFN-I (Fig. 2C). Moreover, the change fold (log2) of expression of DsRed, MDA5 and IFN-1 in an IBDV infection group (about 5–15) is higher than that in NDV infection group and IBV infection group (mainly under 3). The results could be partially explained by differences between single stranded and double stranded RNA, as previous study demonstrated that the expression of chicken MDA5 was highly upregulated during influenza infection, which is a single stranded RNA virus (Karpala et al., 2011). IBDV is a double-stranded RNA virus that belongs to the Birnaviridae family and causes considerable economical loses in the chicken industry (Muller et al., 1979). In this study, the result supports previous reports that chMDA5 recognizes IBDV and the infection of IBDV (in vitro or in vivo) results in the upregulation of MDA5 and MDA5-related innate immune genes, including IFN-α and IFN-β (Hui and Leung, 2015, Lee et al., 2014, Smith et al., 2015). NDV, a negative-stranded RNA virus belonging to paramyxoviruses, which can infect most birds, including chickens (Alexander, 2000), is recognized by RIG-I in mammals. In this study, NDV failed to activate the MDA5 promoter and enhance the expression of endogenous MDA5 and IFN-I, in accordance with previous studies. It has been demonstrated that paramyxovirus V proteins can inactivate MDA5 by disrupting the MDA5 ATP-hydrolysis site and preventing the formation of the RNA-bound MDA5 filament (Motz et al., 2013). Moreover, the V proteins act as IFN-I antagonists inhibiting the increase of IFN-I and blocking binding of IFN to its receptors during NDV infection in DF-1 cells (Jang et al., 2010, Munir et al., 2005). As a result, the antiviral signaling pathway and the positive-feedback mechanism are blocked, which further inhibits the MDA5 promoter activity. Even so, several studies have reported that NDV induced the expression of IFN-I in chicken splenocytes (Rue et al., 2011) and macrophages (Sick et al., 1998) and this discrepancy is likely due to the different cell types used. IBV is a positive-stranded RNA virus belonging to the coronavirus family and causes a highly contagious respiratory disease in poultry (Cavanagh, 2007). Global gene expression profiling has demonstrated that MDA5 expression levels were dramatically increased in chicken kidney tissue infected by Nephropathogenic IBV, while IFN-I expression was not significantly increased (Cong et al., 2013). Furthermore, a previous study found that MDA5 was the primary sensor of IBV and delayed induction of IFN-β in a cell type-independent manner (Kint et al., 2015). In contrast, the expression levels of MDA5 and IFN-I were not upregulated and the MDA5 promoter was not activated by IBV infection in the present study, which may be explained by the differences in virulence and adaptability of the IBV strain. In coronaviruses, 2′-O-methylation of viral mRNA can disrupt recognition of the virus by MDA5. In addition, several proteins encoded by some coronaviruses are involved in counteracting IFN-I (Holmes and Darbyshire, 1978, Liu et al., 2014, Zust et al., 2011). Collectively, the response of the MDA5 promoter to virus inducers was consistent with endogenous MDA5.
Fig. 2.
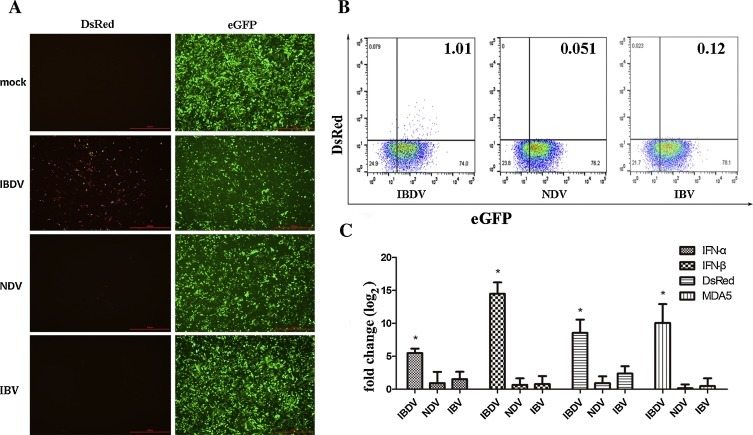
The chMDA5 promoter is dramatically activated in transfected cells that are infected with IBDV, accompanied by upregulation of endogenous MDA5. Piggybac-MDA5-DsRed cells were infected with IBDV, NDV or IBV. (A) Immunofluorescence analysis for DsRed of cells infected with virus. Cells infected with mock, IBDV, NDV and IBV. Scale bars, 200 μm. (B) FACS analysis of DsRed and eGFP-expressing cells. (C) mRNA expression levels of DsRed, eGFP, endogenous MDA5, IFN-α and IFN-β were analyzed by qRT-PCR. Data are presented as means of three independent experiments. Significance was analyzed with one-way ANOVA. The error bars represent the mean of Standard Error. Asterisk represents significant difference between treatment group and the mock (*p < 0.05).
4. Conclusion
In summary, we cloned and characterized the promoter of chMDA5, establishing a system that can be used to determine the effects of viral or non-viral inducer on chMDA5 expression. Importantly, the chMDA5 promoter could be used to express other antiviral genes, as its activity is extremely low under basal conditions and can be modulated by exogenous inducers. To our knowledge, this study is the first to examine the importance of the 2.5 kb promoter of chMDA5. Our results provide a foundation for future research on the direct regulation of MDA5 expression, which could contribute in developing new strategies against viral infection.
Conflict of interest
The authors declare no competing financial interests in relation to the work described.
Author contribution
WXZ designed and performed the vast majority of the experiments presented in this manuscript, wrote most of the paper. EZ assisted in designing and performing the plasmid vector construction. YH assisted to design and perform the virus infection experiment. DYC, XL, MJC and TTL assisted in performing the experiments. XGY, HYX, SSL and MZ assisted in design the experiment and wrote the paper. YQL and KHL are supervisors on this research project. All authors reviewed the results and approved the final version of the manuscript.
Acknowledgements
This research was jointly supported by National Natural Science Foundation of China (31560631), Guangxi Natural Science Foundation (2015GXNSFCB139009, 2014GXNSFBA118096), Guangxi International Collaborative Project (1347004-6) and Guangxi High Education Innovative Research Group and Eminent Scholar Project (2014-7).
Footnotes
Supplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/j.molimm.2016.06.006.
Appendix A. Supplementary data
The following are Supplementary data to this article:
References
- Alexander D.J. Newcastle disease and other avian paramyxoviruses. Rev. Sci. Tech. Oie. 2000;19:443–462. doi: 10.20506/rst.19.2.1231. [DOI] [PubMed] [Google Scholar]
- Barber M.R.W. Association of RIG-I with innate immunity of ducks to influenza. Proc. Natl. Acad. Sci. U.S.A. 2010;107:5913–5918. doi: 10.1073/pnas.1001755107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belgnaoui S.M. Orchestrating the interferon antiviral response through the mitochondrial antiviral signaling (MAVS) adapter. Curr. Opin. Immunol. 2011;23:564–572. doi: 10.1016/j.coi.2011.08.001. [DOI] [PubMed] [Google Scholar]
- Bracci L. Type I IFN is a powerful mucosal adjuvant for a selective intranasal vaccination against influenza virus in mice and affects antigen capture at mucosal level. Vaccine. 2005;23:2994–3004. doi: 10.1016/j.vaccine.2004.12.006. [DOI] [PubMed] [Google Scholar]
- Cavanagh D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007;38:281–297. doi: 10.1051/vetres:2006055. [DOI] [PubMed] [Google Scholar]
- Cong F. Transcriptome analysis of chicken kidney tissues following coronavirus avian infectious bronchitis virus infection. BMC Genomics. 2013;14:743. doi: 10.1186/1471-2164-14-743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui S. The C-terminal regulatory domain is the RNA 5′-triphosphate sensor of RIG-I. Mol. Cell. 2008;29:169–179. doi: 10.1016/j.molcel.2007.10.032. [DOI] [PubMed] [Google Scholar]
- Gack M.U. Roles of RIG-I N-terminal tandem CARD and splice variant in TRIM25-mediated antiviral signal transduction. Proc. Natl. Acad. Sci. U.S.A. 2008;105:16743–16748. doi: 10.1073/pnas.0804947105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gitlin L. Essential role of mda-5 in type I IFN responses to polyriboinosinic:polyribocytidylic acid and encephalomyocarditis picornavirus. Proc. Natl. Acad. Sci. U.S.A. 2006;103:8459–8464. doi: 10.1073/pnas.0603082103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi T. Chicken MDA5 senses short double-stranded RNA with implications for antiviral response against avian influenza viruses in chicken. J. Innate Immun. 2014;6:58–71. doi: 10.1159/000351583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Himly M. The DF-1 chicken fibroblast cell line: transformation induced by diverse oncogenes and cell death resulting from infection by avian leukosis viruses. Virology. 1998;248:295–304. doi: 10.1006/viro.1998.9290. [DOI] [PubMed] [Google Scholar]
- Holmes H.C., Darbyshire J.H. Induction of chicken interferon by avian infectious bronchitis virus. Res. Vet. Sci. 1978;25:178–181. [PubMed] [Google Scholar]
- Huang T. Identification and expression profiling analysis of grass carp Ctenopharyngodon idella LGP2 cDNA. Fish Shellfish Immun. 2010;29:349–355. doi: 10.1016/j.fsi.2010.04.001. [DOI] [PubMed] [Google Scholar]
- Hui R.K., Leung F.C. Differential expression profile of chicken embryo fibroblast DF-1Cells infected with cell-adapted infectious bursal disease virus. PLoS One. 2015;10:e0111771. doi: 10.1371/journal.pone.0111771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janeway C.A., Medzhitov R. Innate immune recognition. Annu. Rev. Immunol. 2002;20:197–216. doi: 10.1146/annurev.immunol.20.083001.084359. [DOI] [PubMed] [Google Scholar]
- Jang J. Overexpression of newcastle disease virus (NDV) V protein enhances NDV production kinetics in chicken embryo fibroblasts. Appl. Microbiol. Biotechnol. 2010;85:1509–1520. doi: 10.1007/s00253-009-2189-z. [DOI] [PubMed] [Google Scholar]
- Kang D.C. mda-5: an interferon-inducible putative RNA helicase with double-stranded RNA-dependent ATPase activity and melanoma growth-suppressive properties. Proc. Natl. Acad. Sci. U.S.A. 2002;99:637–642. doi: 10.1073/pnas.022637199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karpala A.J. Characterization of chicken Mda5 activity: regulation of IFN-beta in the absence of RIG-I functionality. J. Immunol. 2011;186:5397–5405. doi: 10.4049/jimmunol.1003712. [DOI] [PubMed] [Google Scholar]
- Kawai T., Akira S. Antiviral signaling through pattern recognition receptors. J. Biochem. 2007;141:137–145. doi: 10.1093/jb/mvm032. [DOI] [PubMed] [Google Scholar]
- Kint J. Activation of the chicken type I interferon response by infectious bronchitis coronavirus. J. Virol. 2015;89:1156–1167. doi: 10.1128/JVI.02671-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee C.C. Characterization of chicken melanoma differentiation-associated gene 5 (MDA5) from alternative translation initiation. Comp. Immunol. Microbiol. Infect. Dis. 2012;35:335–343. doi: 10.1016/j.cimid.2012.02.004. [DOI] [PubMed] [Google Scholar]
- Lee C.-C. Chicken melanoma differentiation-associated gene 5 (MDA5) recognizes infectious bursal disease virus infection and triggers MDA5-related innate immunity. Arch. Virol. 2014;159:1671–1686. doi: 10.1007/s00705-014-1983-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu D.X. Accessory proteins of SARS-CoV and other coronaviruses. Antiviral Res. 2014;109:97–109. doi: 10.1016/j.antiviral.2014.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meylan E. Intracellular pattern recognition receptors in the host response. Nature. 2006;442:39–44. doi: 10.1038/nature04946. [DOI] [PubMed] [Google Scholar]
- Motz C. Paramyxovirus V proteins disrupt the fold of the RNA sensor MDA5 to inhibit antiviral signaling. Science. 2013;339:690–693. doi: 10.1126/science.1230949. [DOI] [PubMed] [Google Scholar]
- Muller H. The genome of infectious bursal disease virus consists of two segments of double-stranded RNA. J. Virol. 1979;31:584–589. doi: 10.1128/jvi.31.3.584-589.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munir S. Transcriptional response of avian cells to infection with Newcastle disease virus. Virus Res. 2005;107:103–108. doi: 10.1016/j.virusres.2004.07.001. [DOI] [PubMed] [Google Scholar]
- Ramos H.J., Gale M. RIG-I like receptors and their signaling crosstalk in the regulation of antiviral immunity. Curr. Opin. Virol. 2011;1:167–176. doi: 10.1016/j.coviro.2011.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rue C.A. Virulent Newcastle disease virus elicits a strong innate immune response in chickens. J. Gen. Virol. 2011;92:931–939. doi: 10.1099/vir.0.025486-0. [DOI] [PubMed] [Google Scholar]
- Sick C. Promoter structures and differential responses to viral and nonviral inducers of chicken type I interferon genes. J. Biol. Chem. 1998;273:9749–9754. doi: 10.1074/jbc.273.16.9749. [DOI] [PubMed] [Google Scholar]
- Smith J. Analysis of the early immune response to infection by infectious bursal disease virus in chickens differing in their resistance to the disease. J. Virol. 2015;89:2469–2482. doi: 10.1128/JVI.02828-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stark G.R. How cells respond to interferons. Annu. Rev. Biochem. 1998;67:227–264. doi: 10.1146/annurev.biochem.67.1.227. [DOI] [PubMed] [Google Scholar]
- Su J.G. Molecular cloning and immune responsive expression of MDA5 gene, a pivotal member of the RLR gene family from grass carp Ctenopharyngodon idella. Fish Shellfish Immun. 2010;28:712–718. doi: 10.1016/j.fsi.2010.01.009. [DOI] [PubMed] [Google Scholar]
- Takahasi K. Solution structures of cytosolic RNA sensor MDA5 and LGP2 C-terminal domains identification of the rna recognition loop in rig-I-like receptors. J. Biol. Chem. 2009;284:17465–17474. doi: 10.1074/jbc.M109.007179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoneyama M., Fujita T. Recognition of viral nucleic acids in innate immunity. Rev. Med. Virol. 2010;20:4–22. doi: 10.1002/rmv.633. [DOI] [PubMed] [Google Scholar]
- Yoneyama M. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 2004;5:730–737. doi: 10.1038/ni1087. [DOI] [PubMed] [Google Scholar]
- Yoneyama M. Shared and unique functions of the DExD/H-box helicases RIG-I, MDA5, and LGP2 in antiviral innate immunity. J. Immunol. 2005;175:2851–2858. doi: 10.4049/jimmunol.175.5.2851. [DOI] [PubMed] [Google Scholar]
- Zust R. Ribose 2′-O-methylation provides a molecular signature for the distinction of self and non-self mRNA dependent on the RNA sensor Mda5. Nat. Immunol. 2011;12:137–143. doi: 10.1038/ni.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.