

Abstract
We describe a three-color flow cytometry assay for the detection of virus-specific CD4+ and CD8+ T cells in the cat. The assay is based upon detection of intracellular TNFα using the cross-reactive mAb 6401.1111, raised against the human cytokine. Allophycocyanin-conjugated mAb 6401.1111 specifically stained feline TNFα-producing murine cells and also Staphylococcus aureus Enterotoxin B-stimulated feline T cells, thus providing formal evidence for cross-reactivity. By using the anti-TNFα mAb in combination with PE- and FITC-conjugated mAbs against feline CD4 and CD8, respectively, antiviral CD4+ and CD8+ T cells could be identified in the peripheral blood and in the spleens of feline infectious peritonitis virus-infected cats. Moreover, feline calicivirus (FCV)-specific CD4+ T cells were detected in the spleens of FCV-vaccinated cats. As antigen-presenting cells (APCs), we used immortalized autologous fibroblast cell lines, PBMC or splenocytes. A straightforward protocol, in which splenocyte preparations served both as APCs and effector cells, consistently yielded best results. The assay will permit further studies of the cellular immune responses in cats during natural and experimental viral infections. It will contribute to vaccine development against feline viruses by facilitating the identification of T cell antigens and epitopes, and by allowing the quantitative detection of virus-specific T cells after vaccination. Furthermore, the assay will add to the value of those systems in which viral infections of the cat serve as models for human disease.
Keywords: Flow cytometry, Intracellular cytokine staining, Cellular immunity, Coronavirus, Feline infectious peritonitis, Feline calicivirus
1. Introduction
The adaptive cellular immune system is essential for the control of viral infections and plays a key role in the maintenance of antiviral immunity Ahmed and Gray, 1996, Zinkernagel and Hengartner, 1997. During the last decade, various novel methods have been developed to assess T cell responses. These new technologies, which include MHC tetramer staining (Altman et al., 1996), ELISPOT assays and intracellular cytokine staining in combination with flow cytometry Jung et al., 1993, Vikingsson et al., 1994, Prussin and Metcalfe, 1995, Nomura et al., 2000, greatly extend the classical tests such as T cell proliferation and T cell-mediated cytotoxicity (CTL) assays. The full array of assays is available for mouse and human, and is now being developed for rhesus macaques Moretto et al., 2000, Keeney et al., 2003. However, in most host species of veterinary importance the possibilities to measure the specificity and magnitude of the cellular immune response are limited. Clearly, rational vaccine design would benefit from these novel methodologies by allowing (i) the identification of relevant T cell antigens and (ii) the quantitative analysis of the cellular responses following immunization. Moreover, such assays would be of great value in molecular pathogenesis studies, most notably in those instances where viral infections of domestic animals serve as models for human disease. For example, feline immunodeficiency virus infection of cats provides an important and relevant model for HIV pathogenesis (Willet et al., 1997), while feline infectious peritonitis, a lethal coronaviral disease (de Groot and Horzinek, 1995), may serve in many respects as a model for human severe acute respiratory syndrome (SARS).
So far, the tools to monitor feline T cell responses are restricted to the ‘classical assays’. T lymphocyte proliferation tests for the cat have been described by Flynn et al. (1994), but these do not distinguish between T cell subsets. Furthermore, cytotoxicity assays have been established to study T cell responses in cats infected with feline immunodeficiency virus and feline leukemiavirus Tompkins and Tompkins, 1985, Song et al., 1992, Flynn et al., 1994, Flynn et al., 2000, but only provide semi-quantitative information about cytotoxic CD8+ T cells and do not permit a direct ex vivo analysis of antiviral T cell memory.
The development of more sophisticated, quantitative, T cell assays for the cat is hampered by lack of reagents. Moreover, cats represent an outbred population, for which MHC typing is not yet possible and for which syngeneic MHC-matched antigen-presenting cell lines are not readily available. In human and monkey infection models, the problem of haplotype diversity has been solved by using autologous PBMC or transformed B-lymphocytes as antigen-presenting cells Dyer et al., 1999, Larsson et al., 1999, Moretto et al., 2000. Here, we followed a similar approach to detect feline antiviral T cell responses. We describe a three-color flow cytometry assay to identify simultaneously virus-specific Th1 cytokine-producing CD4+ and CD8+ T cells. Stimulated cytokine-producing T cells were detected using a cross-reactive monoclonal antibody against human tumor necrosis factor α. As syngeneic stimulator cells, we used either SV40-immortalized autologous skin fibroblast cell lines, heterologously expressing the dedicated virus-receptor, or autologous PBMC and splenocyte preparations. Using these methods, we were able to demonstrate the presence of virus-specific CD4+ and CD8+ T cells both in the peripheral blood and spleen of cats, which had been infected experimentally with the feline infectious peritonitis coronavirus (FIPV). Moreover, in cats, immunized with a live attenuated feline calicivirus (FCV) vaccine, FCV-specific CD4+ T cells were detected in the spleen.
2. Materials and methods
2.1. Cell lines and viruses
Cell lines were maintained in Dulbecco's modified Eagle's medium (DMEM, Biowhittaker Europe) supplemented with 10% fetal calf serum (FCS, Wisent) and 100 IU penicillin and 100 μg streptomycin per ml. The recombinant vaccinia virus vTF7-3 (Fuerst et al., 1986) was propagated in RK13 cells. Feline infectious peritonitis virus (FIPV) strain 79-1146 (McKeirnan et al., 1981) was propagated in Felis catus whole foetus-D (fcwf) cells (Boyle et al., 1984) as described (de Groot et al., 1987). Feline calicivirus (FCV) strain F9, kindly provided by R.M. Gaskell (University of Liverpool, UK), was grown in Crandell feline kidney cells (American Type Culture Collection) (Crandell et al., 1973).
2.2. Animal experimentation
Specific pathogen-free cats were purchased from Harlan Netherlands. Cats were housed at the Central Animal Facility of the University of Utrecht. Experiments were performed in accordance with institutional and governmental guidelines after approval of the Animal Ethical Committee of the Faculty of Veterinary Medicine, Utrecht University. Cats 085 and 291 were inoculated oronasally with 1000 PFU of FIPV strain 79-1146 at 9 months of age. Both cats seroconverted and developed a recurring fever for 11 or 19 days concomitant with progressive loss of body weight. Subsequently, the fever subsided and the body weight steadily increased. The animals were sacrificed 4 months after experimental infection. Upon post mortem examination, no FIP lesions were found in the major organs nor in the intestines.
Cats 125 and 136 were vaccinated against FCV strain F9 with Felocell RC® (Pfizer Animal Health) at 6 months of age. Each animal received two doses of the vaccine at 2-week intervals. The vaccine was administered subcutaneously according to the instructions of the manufacturer. No clinical signs were observed. Both cats seroconverted as determined by an immunofluorescence assay. The animals were sacrificed 4 weeks after primo vaccination.
2.3. Plasmids and antibodies
Plasmid pAT153@1, containing the early SV40 coding region from which the large T and small t antigens are expressed Dinsart et al., 1984, Klein et al., 1990, was a gift from R.C. Hoeben (LUMC, Leiden University, The Netherlands). Plasmid pCR-fAPN, containing the feline aminopeptidase N (fAPN, CD13) gene under the control of the IE CMV promoter (Tresnan et al., 1996), was kindly provided by K.V. Holmes (University of Colorado, USA).
Phycoerythrin (PE)-conjugated anti-feline CD4 and FITC-conjugated anti-feline CD8 were purchased from Southern Biotechnology Associates and allophycocyanin-conjugated anti-human TNFα (clone 6401.1111) from Becton Dickinson. Allophycocyanin-conjugated monoclonal antibody (mAb) MOPC-21, which served as an isotype-matched negative control, and mAb PAb 108 against SV40 large T and small t antigens were from BD PharMingen. Ascites fluid A290 from a cat experimentally infected with FIPV strain 79-1146 served as a specific antiserum against FIPV. Cat-anti FCV strain F9 was obtained from the Cornell Feline Health Center, Cornell University, Ithaca, USA.
2.4. Isolation and transformation of feline skin fibroblasts
Skin biopsies of 6 mm diameter were maintained for 14 days in DMEM supplemented with 15% FCS, 50 μg gentamycin, 100 IU penicillin and 100 μg streptomycin per ml (DMEM15). Fibroblasts, cultured from the biopsies, could be propagated for up to passage 6 (P6) in 25-cm2 flasks. For transformation, passage-2 cells were trypsinized and washed once with DMEM. Aliqouts of 5×105 cells in 0.5 ml DMEM were then supplemented with 5 μg each of plasmid pAT153@1 and plasmid pCR-fAPN and electroporated using a Biorad Genepulser II by pulsing once at 250 V and 1170 μF. The fibroblasts were immediately taken up in DMEM15 and passaged at least six times in DMEM, containing 500 μg/ml G418, thus selecting for SV40-immortalized, CD13-expressing cells.
2.5. Indirect immunofluorescence assay
The immunofluorescence assay was performed essentially as described (Mijnes et al., 1996), with minor modifications. Briefly, 105 cells were grown onto 12-mm-diameter glass coverslips. At 16 h after seeding, the cells were either infected with virus at a multiplicity of infection (m.o.i.) of 10 PFU per cell, or harvested immediately to test for the expression of SV40 antigens. The cells were fixed with 100% methanol for 20 min at −20 °C and then incubated for 30 min at room temperature in PBS-5% FCS to reduce aspecific binding of antibodies. For the detection of SV40 antigens, the cells were incubated successively with mAb PAb 108 (diluted 1/50 in PBS-5%FCS), and FITC-conjugated goat anti-mouse IgG (Cappel, diluted 1/150). For the specific detection of FIPV antigens, the cells were incubated with the FIPV-specific antiserum (ascites A290, diluted 1/100) and, subsequently, with goat anti-cat IgG-FITC (Cappel, diluted 1/150). The cells were embedded in FluorSave (Calbiochem) and examined using a Leica DM IRB/TCS SP confocal laser scanning microscope configuration.
2.6. Preparation and stimulation of feline PBMC and splenocytes
Peripheral blood mononuclear cells (PBMC) were purified from heparinized peripheral blood by Ficoll density gradient centrifugation at 2000 rpm. The PBMC were washed twice with PBS and then resuspended in 25 mM HEPES-buffered RPMI 1640 (Invitrogen), supplemented with 10% heat-inactivated FCS, Glutamax I (Invitrogen), 50 μM 2-mercaptoethanol and 100 IU penicillin and 100 μg streptomycin per ml (RPMI10).
Single cell spleen suspensions were prepared by straining spleen homogenates through a 70-μM filter (Cell Strainer, Becton Dickinson). The cells were incubated in ACK buffer (0.15 M NH4Cl, 10 mM KHCO3, 0.1 mM NaEDTA.2H2O) for 10 min to lyse the erythrocytes and subsequently pelleted through an FCS cushion at 1500 rpm for 10 min. The cells were then washed once and resuspended in RPMI10.
For aspecific stimulation of PBMC and splenocytes, 106 cells were incubated for 16 h in RPMI10, containing 1 μg/ml of the superantigen Staphylococcus aureus Enterotoxin B (SEB, Toxin Technology). For virus-specific stimulation, 5×104 fibroblasts, seeded into 96-well flat-bottom microtiter plates (Costar), were either mock-infected or inoculated with FIPV at an m.o.i. of 10 PFU per cell. At 1-h post infectionem (p.i.), the inoculum was replaced by DMEM10 and incubation was continued. At 16-h p.i., 106 autologous splenocytes were added. Alternatively, PBMC and splenocyte preparations served both as APC and effector cells. To this end, 5×105 of these cells were infected at an m.o.i. of 2 PFU per cell with FIPV 79-1146 for 16 h or with FCV F9 for 6 h. Then, 5×105 cells from the same preparation were added. Incubation was continued for 6 h in the presence of either Brefeldin A (Sigma), at a concentration of 10 μg/ml, or GolgiStop (BD PharMingen), containing monensin, according to the instructions of the manufacturer.
2.7. Intracellular cytokine staining and flow cytometry
Stimulated lymphocytes were pelleted in round-bottom microtiter plates (Costar) by centrifugation at 1200 rpm and washed with PBS, containing 1% protease-free BSA (Sigma) and 0.1% sodium azide (FACS buffer). The cells were surface-stained for 30 min at room temperature by subsequent incubation with anti-feline CD4-PE (diluted 1/25 in FACS buffer) and anti-feline CD8-FITC (diluted 1/17). The cells were washed consecutively with FACS buffer (twice), 1× FACS™ Lysing Solution (Becton Dickinson) (twice), and FACS buffer (once) followed by an incubation in 1× FACS™ Permeabilizing Solution 2 (Becton Dickinson) for 10 min at room temperature. The permeabilized cells were then washed twice with FACS buffer and incubated with allophycocyanin-conjugated anti-human TNFα (diluted 1/25) or with allophycocyanin-conjugated MOPC-21 (diluted 1/400) for 1.5 h. Finally, the cells were washed twice with FACS buffer and resuspended in PBS, containing 2% paraformaldehyde. For each sample, 300,000 cells were analyzed, employing a FACScalibur™ flow cytometer (Becton Dickinson) and the Windows-based WinMDI software (J. Trotter, The Scripps Research Institute, La Jolla, CA, USA), live-gating for lymphocytes in the forward and side scatter.
2.8. cDNA cloning and transient expression of feline TNFα
Total intracellular RNA was isolated from freshly prepared feline PBMC using the RNeasy kit (Qiagen). The RNA was subjected to RT-PCR using Ready-To-Go™ RT-PCR beads (Amersham Biosciences) with primers 2049 (5′ GTTTAAACAAGCTTTACCATGAGCACTGAAAGCATGA 3′) and 2050 (5′ GCGGCCGCAGGATCCTTCACAGGGCAATG ATCCCA 3′). These primers were designed to provide the start codon of the feline TNFα coding sequence with an optimal context for translation-initiation and to create unique 5′ HinDIII and 3′ BamHI sites for convenient cloning. A single RT-PCR product of the anticipated size of 0.8 kb was obtained, which was purified from gel, cut with HinDIII and BamHI and inserted into HinDIII/BamHI digested pcDNA3.1/Myc-His B (Invitrogen), downstream of the bacteriophage T7 promotor. Sequence analysis of the resulting expression vector, pcDNA-fTNF, confirmed that the feline TNFα coding sequence had been inserted correctly and that there were no inadvertent mutations. For transient expression of feline TNFα, 7×105 murine Ost7-1 cells (Elroy-Stein and Moss, 1990) were infected at an m.o.i. of 10 PFU per cell with recombinant vaccinia virus vTF7-3. At 1-h p.i., the cells were either mock-transfected or transfected with 4 μg of pcDNA-fTNF using lipofectine (Invitrogen) as described previously (Mijnes et al., 1996). At 6-h p.i., the cells were washed once with PBS, collected in 500 μl PBS and transferred to a round bottom plate. They were then washed twice with FACS buffer, and permeabilized, stained for intracellular TNFα and analyzed by flow cytometry as described above.
3. Results
3.1. Detection of stimulated, TNFα-producing T cells
Stimulation of T cells results in the induction of cytokine production, characteristically IFNγ and TNFα in the case of a Th1-type response (Slifka and Whitton, 2000). We tested several commercial and non-commercial monoclonal antibodies (mAbs) against feline IFNγ, but none of these detected activated feline T cells in a flow cytometry assay (not shown). So far, mAbs against feline TNFα have not been described. However, in contrast to IFNγ, TNFα is highly conserved among mammalian species. Feline and human TNFα are closely related, sharing 89.7% amino acid sequence identity versus 78% and 78.4% with murine TNFα, respectively. This prompted us to test mouse mAb 6401.1111, which was raised against the human cytokine, for cross-reactivity with TNFα of the cat. We employed the vaccinia virus-based vTF7-3 system (Fuerst et al., 1986) to express feline TNFα heterologously. vTF7-3-infected mouse Ost7-1 cells were transfected at 1-h p.i. with expression vector pcDNA-fTNF, which carries the feline TNFα coding sequence downstream of the bacteriophage T7 promotor. Cells were harvested at 6-h p.i. and stained intracellularly with APC-conjugated mAb 6401.1111. Flow cytometry analysis revealed specific staining of pcDNA-fTNF-transfected cells (∼25%) but not of mock-transfected cells (Fig. 1) . Staining of pcDNA-fTNF-transfected cells was not observed with mAb MOPC-21, which served as an isotype-matched negative control. From these data, we concluded that mAb 6401.1111 indeed cross-reacts with feline TNFα.
Fig. 1.
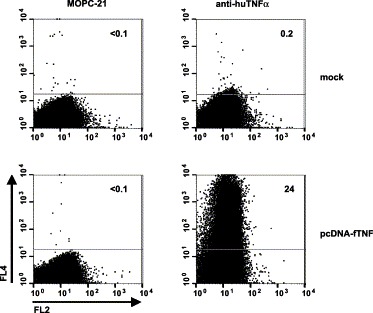
Specific staining of feline TNFα-producing murine cells with allophycocyanin-labeled anti-human TNFα mAb 6401.1111. Mouse Ost7-1 cells were infected with vacinia virus vTF7-3 and, at 1-h p.i., either mock-transfected (upper panels) or transfected with pcDNA-fTNF (lower panels). At 6-h p.i., the cells were formaldehyde-fixed, permeabilized and stained with isotype control mAb MOPC-21 (left panels) or with anti-human TNFα mAb 6401.1111 (right panels). At least 300,000 cells were analyzed. In the graphs, allophycocyanin-fluorescence (FL4 channel) is plotted against autofluorescence in the FL2 channel. The results are representative of two independent experiments.
To test whether mAb 6401.1111 also detects TNFα-producing feline T cells, PBMC isolated from cats were either mock-stimulated or stimulated with the superantigen S. aureus Enterotoxin B (SEB) for 16 h and incubated for another 6 h in medium containing 10 μg/ml of the protein transport inhibitor Brefeldin A (BFA). The cells were surface-stained with PE-conjugated anti-feline CD4 and FITC-conjugated anti-feline CD8, permeabilized and stained with allophycocyanin-conjugated anti-human TNFα. As shown in Fig. 2A , stimulation of PBMC with SEB resulted in a large increase in the population of mAb 6401.1111-positive CD4+ and CD8+ T cells as compared to mock stimulation. Staining of feline T cells was not observed with mAb MOPC-21. These data showed that mAb 6401.1111, in combination with the mAbs against feline CD4 and CD8, can be employed in a three-color flow cytometry assay to identify stimulated T cells specifically.
Fig. 2.
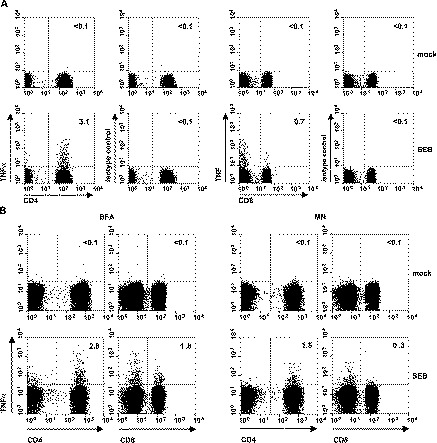
Detection of stimulated TNFα-producing feline T lymphocytes by three-color flow cytometry analysis. (A) Feline PBMC were either mock-stimulated (upper panel) or stimulated with SEB (lower panel) for 16 h and incubated for another 6 h in tissue culture supernatant containing 10 μg/ml of the protein transport inhibitor Brefeldin A. The cells were surface-stained with PE-conjugated anti-feline CD4 and FITC-conjugated anti-feline CD8, permeabilized and stained either with allophycocyanin-conjugated anti-human TNFα or with the allophycocyanin-conjugated isotype-matched control MOPC21. At least 300,000 cells were analyzed. Lymphocytes were selected in the forward and side scatter plot and gated for CD4+ and CD8+ T cells in the side scatter plots. The data are representative of two independent experiments performed with PBMC of two different animals. The numbers indicate the percentages of anti-TNFα-positive or isotype control-positive cells within the CD4+ or CD8+ T cell populations. (B) The effect of protein transport inhibitors on intracellular TNFα staining. PBMC were either mock-stimulated or stimulated with SEB for 16 h prior to a 6-h incubation with either Brefeldin A (BFA, left panel) or monensin (MN, right panel). The cells were surface-stained for CD4 and CD8, permeabilized and stained for intracellular TNFα. Flow cytometry analysis was as described in (A).
Inhibition of cytokine secretion is essential for efficient detection by intracellular staining. The Golgi-disrupting compounds BFA and monensin (MN) are commonly used for this purpose. As reviewed by O'Neil-Andersen and Lawrence (2002), BFA and MN may differentially affect cytokine expression and cell viability depending on cell type and host species. In our model, the percentage of TNFα-expressing CD4+ and CD8+ T cells, detected after SEB-stimulation, was up to 6-fold higher in BFA-treated than in MN-treated PBMC (Fig. 2B). BFA was therefore used in all subsequent experiments.
3.2. Detection of feline infectious peritonitis virus-specific T cells
To assess whether the flow cytometry assay allowed detection of virus-specific feline T cells, we studied CD4+ and CD8+ T cell responses in two cats which had survived an experimental infection with feline infectious peritonitis virus (FIPV) strain 79-1146. Splenocytes and PBMC were isolated to provide a source of antiviral T cells. In the case of CD8+ T cells, specific stimulation requires presentation of the relevant antigenic peptides in the context of the appropriate MHC class I molecules. To circumvent the problem of MHC class I haplotype diversity, we constructed autologous immortalized skin fibroblast cell lines, stably expressing the FIPV type II receptor CD13 (aminopeptidase N), to serve as antigen-presenting cells (APCs). As shown in Fig. 3 , CD13-expression resulted in a dramatic increase in the number of FIPV-permissive cells.
Fig. 3.
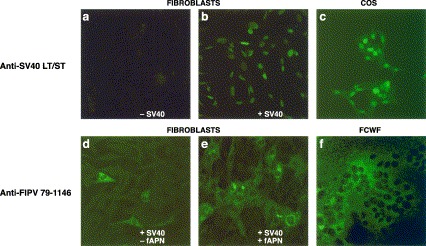
Characterization of immortalized FIPV-susceptible autologous fibroblasts. (A) Expression of SV40 large T and small t antigens by pAT153@1-transfected immortalized fibroblasts. Primary skin fibroblasts (−SV40), pAT153@1-transfected immortalized fibroblasts (+SV40) and COS 7 cells (COS) were grown onto 12-mm-diameter glass coverslips. At 16-h post seeding, the cells were fixed and stained for SV40 large T and small t antigen with mAb PAb 108 and FITC-labeled goat anti-mouse IgG. The cells were examined by confocal laser scanning microscopy. (B) Stable expression of fAPN by SV40-immortalized fibroblasts confers increased permissivity to FIPV infection. Immortalized skin fibroblast cell lines were generated by transfection with pAT153@1 alone (−fAPN) or by co-transfection with expression vector pCR-fAPN (+fAPN). Fibroblasts were grown onto 12-mm-diameter glass coverslips and infected with FIPV strain 79-1146 at an m.o.i. of 10 PFU per cell at 16-h post seeding; FIPV-infected fcwf cells were included as a positive control (FCWF). At 6-h p.i., the cells were fixed and stained using an polyclonal antiserum against FIPV (ascites A290) and FITC-conjugated goat anti-cat IgG. The results are representative of four independent experiments.
The fibroblasts were inoculated with FIPV strain 79-1146 at an m.o.i. of 10 PFU per cell. At 16-h p.i., splenocytes were added and incubation was continued in the presence of BFA for an additional 6 h, after which the cells were surface-stained for CD4, CD8 and stained intracellularly for TNFα. Stimulation of splenocytes with FIPV-infected fibroblasts clearly revealed populations of virus-specific TNFα-producing T cells. As illustrated for cat 085, 1.8% of CD4+ T cells and 0.4% of CD8+ T cells produced TNFα in response to stimulation with FIPV-infected fibroblasts versus 0.6% and <0.1% after mock-stimulation (Fig. 4A) . Similar results were obtained for cat 291 (not shown).
Fig. 4.
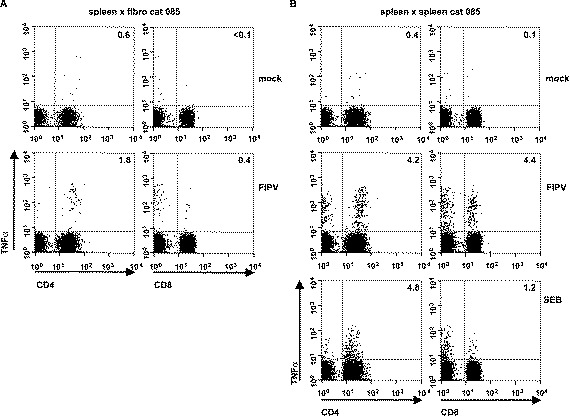
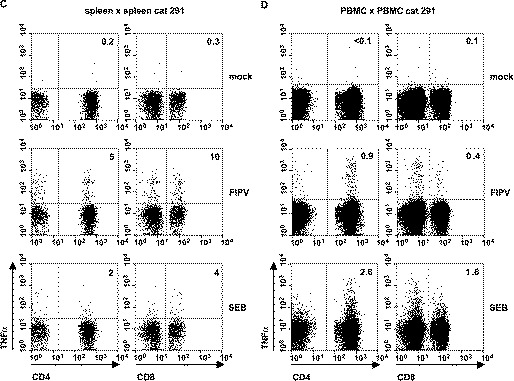
Detection of antiviral T cells in the spleens and PBMC of FIPV-survivors. (A) 5×104 immortalized fAPN-transformed fibroblasts from cat 085 were either mock-infected or infected with FIPV strain 79-1146 at an m.o.i. of 10. At 16-h p.i., 1×106 autologous splenocytes were added and the incubation was continued for 6 h in the presence of BFA. (B and C) 5×105 splenocytes from cats 085 and 291, respectively, were either mock-infected or infected with FIPV strain 79-1146 at an m.o.i. of 2. At 16-h post infection, 5×105 autologous splenocytes were added and the incubation was continued for 6 h in the presence of BFA. As a positive control, 1×106 splenocytes were stimulated with SEB for 16 h prior to the addition of BFA. (D) As in panels B and C, but using PBMC of cat 291 as APCs and effectors. The cells were surface-stained for CD4 and CD8, permeabilized and stained for intracellular TNFα. Three-color flow cytometry analysis was as described in Fig. 2A. The numbers indicate the percentages of TNFα-positive cells within the CD4+ or CD8+ T cell populations.
From these results, we concluded that virus-infected autologous fibroblasts could serve as APCs in this assay. However, the construction of stable virus-permissive cell lines for individual animals is laborious and time-consuming. To explore a more convenient procedure and to corroborate our results independently, we adapted protocols from Larsson et al. (1999) and Moretto et al. (2000) and used autologous splenocytes and PBMC both as APCs and effector cells. This approach exploits the fact that during a natural infection FIPV infects cells of the monocyte/macrophage lineage (de Groot and Horzinek, 1995), which are also commonly present in splenocyte and PBMC preparations.
Splenocytes and PBMC were inoculated at an m.o.i of 2 PFU per cell. At 16-h p.i., an equal amount of the original non-infected cell preparation was added and incubation was continued for 6 h in the presence of BFA. Flow cytometry analysis again revealed the presence of FIPV-specific TNFα-expressing T cells in the spleen. In fact, up to 5% of CD4+ T cells and 10% of CD8+ T cells responded to FIPV-antigens (Fig. 4B,C). The infected splenocyte preparations were clearly more efficient APCs than the immortalized fibroblasts (compare Fig. 3A and B). Importantly, FIPV-specific T cells could not only be detected in the spleen, but also in peripheral blood (Fig. 4D).
3.3. Detection of feline calicivirus-specific T cells in vaccinated cats
Our data show that virus-specific T cells can be detected in FIPV-survivors. However, to be of general use, the assay should also detect T cells against other feline viruses. Furthermore, the assay should be sensitive enough to measure memory T cells after vaccination. To test this, we analyzed the cellular immune responses in two cats which had been immunized subcutaneously with a live attenuated vaccine against feline calicivirus (FCV). FCV has been reported to infect lymphocytes (Kawaguchi et al., 1994). We therefore used PBMC and splenocyte preparations, infected with the homologous FCV strain F9, as APCs. No antiviral T cells were found in the peripheral blood (not shown). However, in the spleens of both vaccinated animals, FCV-specific TNFα-producing CD4+ T cells were observed (∼0.8% of the CD4+ population). FCV-specific CD8+ T cells were not detected (Fig. 5) .
Fig. 5.
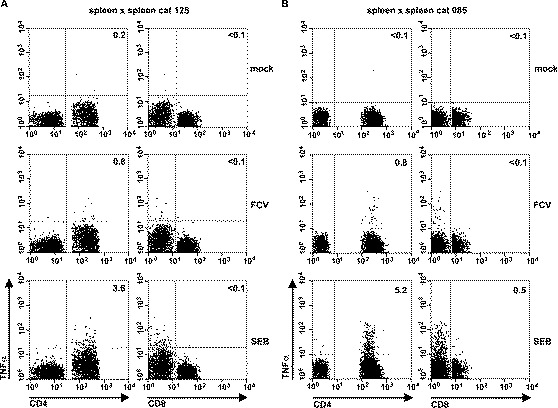
Detection of feline calicivirus-specific T cells in the spleens of vaccinated cats. Splenocytes (5×105 each) from cats 125 (A) and 136 (B) were either mock-infected (mock) or infected with FCV strain F9 (FCV) at an m.o.i. of 2 PFU per cell. At 6-h p.i., 5×105 autologous splenocytes were added and the incubation was continued for 6 h in the presence of BFA. As a positive control, 1×106 splenocytes were stimulated with SEB for 6 h prior to the addition of BFA. The cells were surface-stained for CD4 and CD8, permeabilized and stained for intracellular TNFα. The numbers indicate the percentage of TNFα-positive CD4+ or CD8+ T cells.
4. Discussion
Here, we describe a three-color flow cytometry assay for the detection of virus-specific CD4+ and CD8+ T cells in cats. Usually, in this type of assay, stimulated T cells are identified by intracellular staining for IFNγ Vikingsson et al., 1994, Prussin and Metcalfe, 1995, Murali-Krishna et al., 1998. It is well established, however, that stimulated Th1-type T cells also produce TNFα Slifka et al., 1999, Appay et al., 2000, Badovinac and Harty, 2000, Slifka and Whitton, 2000 and that staining for this cytokine provides a good alternative for the identification of activated T cells in mice, humans and macaques Badovinac and Harty, 2000, Nomura et al., 2000, Keeney et al., 2003, Nomura et al., 2003. As our initial attempts to detect SEB-stimulated T cells using various commercial and non-commercial antisera against feline IFNγ were unsuccessful, we explored the use of a commercially available mAb, raised against human TNFα. MAb 6401.1111 specifically stained feline TNFα-producing murine cells and also S. aureus Enterotoxin B-stimulated feline T cells, thus providing formal evidence for cross-reactivity. Cross-reactivity between a mAb against human TNFα with the feline homologue has also recently been reported by Aguirre et al. (2003). By using allophycocyanin-conjugated mAb 6401.1111 in combination with PE- and FITC-conjugated mAbs against feline CD4 and CD8, respectively, antiviral T cells could be specifically identified in the peripheral blood and/or the spleens of FIPV-infected and of FCV-vaccinated cats.
One particular hurdle in the development of T cell assays for cats is that these animals constitute an out-bred population for which MHC-typing has not yet been developed and for which virus-permissive, MHC-matched target cells are not readily available. Previously, both primary and SV40-immortalized autologous skin fibroblasts have been successfully used in 51Cr-release CTL assays for feline leukemia virus and feline immunodeficiency virus Flynn et al., 1994, Flynn et al., 2000, Köksoy et al., 2001. Hence, the use of immortalized autologous fibroblasts as APCs for T cell stimulation seemed an obvious choice. The fibroblast cultures were fully permissive to FCV (not shown), but only 1% to 5% of the cells could be infected by FIPV strain 79-1146. To ensure that the cells were also FIPV-permissive, we generated continuous fibroblast cell lines, stably expressing the FIPV receptor CD13. The construction of cell lines for each individual animal is, however, labor-intensive. Therefore, a comparison was made with less elaborate approaches, in which PBMC or splenocytes served both as APC and effector cells. Virus-specific CD4+ and CD8+ T cells were identified with all three procedures, but assays performed with splenocyte preparations consistently yielded the best results. As compared to stimulation with infected autologous fibroblast cell lines, stimulation with FIPV-infected splenocytes resulted in up to 3- and 11-fold increases in the number of stimulated TNFα-producing CD4+ and CD8+ T cells, respectively. One possible explanation for this difference is that fibroblasts are non-professional antigen presenting cells which, presumably, lack co-stimulatory molecules such as B7-1 and B7-2. In contrast, the FIPV-permissive dendritic cells and macrophages, present in splenocyte preparations, are specialized APCs fully competent to provide the required co-stimulation.
FIPV-specific T cells could readily be detected in the peripheral blood using PBMC preparations both as APCs and effector cells, although the percentages of stimulated T cells observed were generally lower than those measured when using splenocytes. It is possible that in hosts which have cleared an FIPV infection the numbers of virus-specific (memory) T cells within the spleen exceed those present in the periphery. In addition, the numbers of FIPV-permissive APCs, i.e. CD13+ monocytes and macrophages, may be lower in PBMC than in splenocyte preparations. Be that as it may, the fact that virus-specific T cells can be detected in peripheral blood makes it possible to monitor the dynamics of the adaptive cellular immune response ex vivo in longitudinal acute and chronic infection studies.
Antiviral CD4+ T cells were also identified in FCV-vaccinated animals, although exclusively in the spleen. FCV-specific CD8+ T cells were not detected. Although FCV can be isolated from the blood (Gillespie and Scott, 1973) and reportedly infect macrophages (Langloss et al., 1978) and lymphocytes (Kawaguchi et al., 1994), we cannot exclude the possibility that a lack of FCV-permissive APCs in the PBMC and splenocyte preparations precluded efficient MHC class I presentation. It is of note, however, that stimulation of splenocytes with FCV-infected autologous fibroblasts also failed to reveal antiviral CD8+ T cells (not shown). Most likely, the observed quality and quantity of the FCV-specific T cell responses are pertinent to the vaccine or to the vaccination route. It will be interesting to determine the T cell responses in vaccinated and non-vaccinated animals upon intranasal infection with virulent FCV.
The flow cytometry assay described in this paper will allow us to gain further insight into cellular immune function in cats during natural and experimental infections. It will contribute to vaccine development against feline viruses by facilitating the identification of T cell antigens and epitopes and by allowing the quantitative detection of virus-specific T cells after vaccination; the magnitude, bias and specificity of the cellular immune responses may provide new correlates of protection. Last but not least, we believe that the assay will also add to the value of those systems in which viral infections in the cat serve as models for human disease.
Acknowledgments
We thank the personnel of the Infection Unit of the Central Animal Facility of the University of Utrecht for excellent technical assistance. We are grateful to Dr. G.J.A. Arkesteijn (Flow Cytometry Facility, Faculty of Veterinary Medicine, University of Utrecht) and Ing. L. van Weeren (Center of Cell Imaging, Faculty of Veterinary Medicine, University of Utrecht) for helpful suggestions concerning the FACScalibur™ and the confocal laser scanning microscope, respectively. The research of J.D.F. de Groot-Mijnes was supported by Intervet International.
Note added in proof
While our manuscript was under review, a publication appeared in which antibodies against feline IFN-γ were used to detect feline leukemia virus-specific CD4+ T cells (Graham, Jarrett and Flynn, 2003 Development of antibodies to feline IFN-γ as tools to elucidate the cellular immune responses to FELV. Journal of Immunological Methods 279, 69–78).
References
- Aguirre A., Escobar A., Ferreira V., Molina M.C., Ferreira A., Aguillon J.C. An anti-human recombinant tumor necrosis factor alpha (TNF alpha) monoclonal antibody recognizes an epitope in feline TNF alpha. Vet. Rec. 2003;34:177. doi: 10.1051/vetres:2002064. [DOI] [PubMed] [Google Scholar]
- Ahmed R., Gray D. Immunological memory and protective immunity: understanding their relation. Science. 1996;272:54. doi: 10.1126/science.272.5258.54. [DOI] [PubMed] [Google Scholar]
- Altman J.D., Moss P.A., Goulder P.J., Barouch D.H., McHeyzer-Williams M.G., Bell J.I., McMichael A.J., Davis M.M. Phenotypic analysis of antigen-specific T lymphocytes. Science. 1996;274:94. doi: 10.1126/science.274.5284.94. [DOI] [PubMed] [Google Scholar]
- Appay V., Nixon D.F., Donahoe S.M., Gillespie G.M., Dong T., King A., Ogg G.S., Spiegel H.M., Conlon C., Spina C.A., Havlir D.V., Richman D.D., Waters A., Easterbrook P., McMichael A.J., Rowland-Jones S.L. HIV-specific CD8(+) T cells produce antiviral cytokines but are impaired in cytolytic function. J. Exp. Med. 2000;192:63. doi: 10.1084/jem.192.1.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Badovinac V.P., Harty J.T. Intracellular staining for TNF and IFN-gamma detects different frequencies of antigen-specific CD8(+) T cells. J. Immunol. Methods. 2000;238:107. doi: 10.1016/s0022-1759(00)00153-8. [DOI] [PubMed] [Google Scholar]
- Boyle J.F., Pedersen N.C., Evermann J.F., McKeirnan A.J., Ott R.L., Black J.W. Plaque assay, polypeptide composition and immunochemistry of feline infectious peritonitis virus and feline enteric coronavirus isolates. Adv. Exp. Med. Biol. 1984;173:133. doi: 10.1007/978-1-4615-9373-7_12. [DOI] [PubMed] [Google Scholar]
- Crandell R.A., Fabricant C.G., Nelson-Rees W.A. Development, characterization, and viral susceptibility of a feline (Felis catus) renal cell line (CRFK) In Vitro. 1973;9:176. doi: 10.1007/BF02618435. [DOI] [PubMed] [Google Scholar]
- de Groot R.J., Horzinek M.C. Feline infectious peritonitis. In: Siddell S.G., editor. The Coronaviridae. Plenum; New York: 1995. p. 239. [Google Scholar]
- de Groot R.J., ter Haar R.J., Horzinek M.C., van der Zeijst B.A. Intracellular RNAs of the feline infectious peritonitis coronavirus strain 79-1146. J. Gen. Virol. 1987;68:995. doi: 10.1099/0022-1317-68-4-995. [DOI] [PubMed] [Google Scholar]
- Dinsart C., Cornelis J.J., Klein B., van der Eb A.J., Rommelaere J. Transfection with extracellularly UV-damaged DNA induces human and rat cells to express a mutator phenotype towards parvovirus H-1. Mol. Cell. Biol. 1984;4:324. doi: 10.1128/mcb.4.2.324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dyer W.B., Ogg G.S., Demoitie M.A., Jin X., Geczy A.F., Rowland-Jones S.L., McMichael A.J., Nixon D.F., Sullivan J.S. Strong human immunodeficiency virus (HIV)-specific cytotoxic T-lymphocyte activity in Sidney Blood Bank Cohort patients infected with nef-defective HIV type 1. J. Virol. 1999;73:436. doi: 10.1128/jvi.73.1.436-443.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elroy-Stein O., Moss B. Cytoplasmic expression system based on constitutive synthesis of bacteriophage T7 RNA polymerase in mammalian cells. Proc. Natl. Acad. Sci. U. S. A. 1990;87:6743. doi: 10.1073/pnas.87.17.6743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flynn J.N., Cannon C.A., Beatty J.A., Mackett M., Rigby M.A., Neil J.C., Jarrett C. Induction of feline immunodeficiency virus-specific cytotoxic T cells in vivo with carrier-free synthetic peptide. J. Virol. 1994;68:5835. doi: 10.1128/jvi.68.9.5835-5844.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flynn J.N., Hanlon L., Jarrett O. Feline leukaemia virus: protective immunity is mediated by virus-specific cytotoxic T lymphocytes. Immunology. 2000;101:120. doi: 10.1046/j.1365-2567.2000.00089.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuerst T.R., Niles E.G., Studier F.W., Moss B. Eukaryotic transient-expression system based on recombinant vaccinia virus that synthesizes bacteriophage T7 RNA polymerase. Proc. Natl. Acad. Sci. U. S. A. 1986;83:8122. doi: 10.1073/pnas.83.21.8122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillespie J.H., Scott F.W. Feline viral infections. Adv. Vet. Sci. Comp. Med. 1973;17:163. [PubMed] [Google Scholar]
- Jung T., Schauer U., Heusser C., Neumann C., Rieger C. Detection of intracellular cytokines by flow cytometry. J. Immunol. Methods. 1993;159:197. doi: 10.1016/0022-1759(93)90158-4. [DOI] [PubMed] [Google Scholar]
- Kawaguchi Y., Tohya Y., Horimoto T., Maeda K., Miyazawa T., Mikami T. Carrier-state infection of feline T-lymphoblastoid cells with feline calicivirus. Vet. Microbiol. 1994;40:379. doi: 10.1016/0378-1135(94)90125-2. [DOI] [PubMed] [Google Scholar]
- Keeney T.S., Nomura L.E., Maecker H.T., Sastry K.J. Flow cytometric analysis of macaque whole blood for antigen-specific intracellular cytokine production by T lymphocytes. J. Med. Primatol. 2003;32:23. doi: 10.1034/j.1600-0684.2003.00005.x. [DOI] [PubMed] [Google Scholar]
- Klein B., Pastink A., Odijk H., Westerveld A., van der Eb A.J. Transformation and immortalization of diploid xeroderma pigmentosum fibroblasts. Exp. Cell Res. 1990;191:256. doi: 10.1016/0014-4827(90)90012-y. [DOI] [PubMed] [Google Scholar]
- Köksoy S., Phipps A.J., Hayes K.A., Mathes L.E. SV40 immortalization of feline fibroblasts as targets for MHC-restricted cytotoxic T-cell assays. Vet. Immunol. Immunopathol. 2001;79:285. doi: 10.1016/s0165-2427(01)00272-0. [DOI] [PubMed] [Google Scholar]
- Langloss J.M., Hoover E.A., Kahn D.E., Kniazeff A.J. In vitro interaction of alveolar macrophages and pneumocytes with feline respiratory viruses. Infect. Immun. 1978;20:836. doi: 10.1128/iai.20.3.836-841.1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larsson M., Jin X., Ramratnam B., Ogg G.S., Engelmayer J., Demoitie M.A., McMichael A.J., Cox W.I., Steinman R.M., Nixon D., Bhardwaj N. A recombinant vaccinia virus based ELISPOT assay detects high frequencies of Pol-specific CD8 T cells in HIV-1-positive individuals. AIDS. 1999;13:767. doi: 10.1097/00002030-199905070-00005. [DOI] [PubMed] [Google Scholar]
- McKeirnan A.J., Evermann J.F., Hargis A., Ott R.L. Isolation of feline coronaviruses from two cats with diverse disease manifestations. Feline Pract. 1981;11:17. [Google Scholar]
- Mijnes J.D., van der Horst L.M., van Anken E., Horzinek M.C., Rottier P.J., de Groot R.J. Biosynthesis of glycoproteins E and I of feline herpesvirus: gE–gI interaction is required for intracellular transport. J. Virol. 1996;70:5466. doi: 10.1128/jvi.70.8.5466-5475.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moretto W.J., Drohan L.A., Nixon D.F. Rapid quantification of SIV-specific CD8 T cell responses with recombinant vaccinia virus ELISPOT or cytokine flow cytometry. AIDS. 2000;14:2625. doi: 10.1097/00002030-200011100-00034. [DOI] [PubMed] [Google Scholar]
- Murali-Krishna K., Altman J.D., Suresh M., Sourdive D.J., Zajac A.J., Miller J.D., Slansky J., Ahmed R. Counting antigen-specific CD8 T cells: a reevaluation of bystander activation during viral infection. Immunity. 1998;8:177. doi: 10.1016/s1074-7613(00)80470-7. [DOI] [PubMed] [Google Scholar]
- Nomura L.E., Walker J.M., Maecker H.T. Optimization of whole blood antigen-specific cytokine assays for CD4(+) T cells. Cytometry. 2000;40:60. doi: 10.1002/(sici)1097-0320(20000501)40:1<60::aid-cyto8>3.0.co;2-j. [DOI] [PubMed] [Google Scholar]
- Nomura L.E., DeHaro E.D., Martin L.N., Maecker H.T. Optimal preparation of rhesus macaque blood for cytokine flow cytometric analysis. Cytometry. 2003;53A:28. doi: 10.1002/cyto.a.10038. [DOI] [PubMed] [Google Scholar]
- O'Neil-Andersen N.J., Lawrence D.A. Differential modulation of surface and intracellular protein expression by T cells after stimulation in the presence of monensin or brefeldin A. Clin. Diagn. Lab. Immunol. 2002;9:243. doi: 10.1128/CDLI.9.2.243-250.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prussin C., Metcalfe D.D. Detection of intracytoplasmic cytokine using flow cytometry and directly conjugated anti-cytokine antibodies. J. Immunol. Methods. 1995;188:117. doi: 10.1016/0022-1759(95)00209-x. [DOI] [PubMed] [Google Scholar]
- Slifka M.K., Whitton J.L. Activated and memory CD8+ T cells can be distinguished by their cytokine profiles and phenotypic markers. J. Immunol. 2000;164:208. doi: 10.4049/jimmunol.164.1.208. [DOI] [PubMed] [Google Scholar]
- Slifka M.K., Rodriguez F., Whitton J.L. Rapid on/off cycling of cytokine production by virus-specific CD8+ T cells. Nature. 1999;401:76. doi: 10.1038/43454. [DOI] [PubMed] [Google Scholar]
- Song W., Collisson E.W., Billingsley P.M., Brown W.C. Induction of feline immunodeficiency virus-specific cytolytic T-cell responses from experimentally infected cats. J. Virol. 1992;66:5409. doi: 10.1128/jvi.66.9.5409-5417.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tompkins M.B., Tompkins W.A. Stimulation of a cell-mediated cytotoxic response to FeLV-induced T cell lymphomas in the cat. J. Immunol. 1985;135:2817. [PubMed] [Google Scholar]
- Tresnan D.B., Levis R., Holmes K.V. Feline aminopeptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I. J. Virol. 1996;70:8669. doi: 10.1128/jvi.70.12.8669-8674.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vikingsson A., Pederson K., Muller D. Enumeration of IFN-gamma producing lymphocytes by flow cytometry and correlation with quantitative measurement of IFN-gamma. J. Immunol. Methods. 1994;173:219. doi: 10.1016/0022-1759(94)90300-x. [DOI] [PubMed] [Google Scholar]
- Willet B.J., Flynn J.N., Hosie M.J. FIV infection of the domestic cat: an animal model for AIDS. Immunol. Today. 1997;18:182. doi: 10.1016/s0167-5699(97)84665-8. [DOI] [PubMed] [Google Scholar]
- Zinkernagel R.M., Hengartner H. Antiviral immunity. Immunol. Today. 1997;18:258. doi: 10.1016/s0167-5699(97)80017-5. [DOI] [PubMed] [Google Scholar]