

Abstract
The Iberian lynx (Lynx pardinus) is considered the most endangered felid species in the world. To date, less than 200 animals remain in the wild. Low numbers and genetic uniformity may contribute to render this species particularly susceptible to infectious diseases. Different pathogens have been identified in Iberian lynxes; including several feline bacterial and viral agents. Within a 6-month period starting in December 2006, 12 lynxes living in the northern part of the Doñana area were found to be infected with feline leukemia virus (FeLV). Eleven of these animals were antigenemic, and four of them died in the wild in less than 6 months since the first infected animal had been discovered. The remaining viremic lynxes were captured and allocated to a quarantine center to stop the spread of the infection. An additional three animals died shortly in the quarantine center due to acute anemic disease. Sequencing of the envelope surface unit gene revealed a common origin for the FeLV found in all lynxes. The sequences were closely related to FeLV-A/61E, originally isolated from cats in the USA. Our data demonstrate that, similarly to FeLV, the introduction of a new or particularly pathogenic infection brought into the small population of Iberian lynxes by other wild carnivores or feral cats and dogs roaming in the same habitats have severe consequences. It could result in epidemics that have the potential to eradicate the entire lynx population.
Keywords: Lynx pardinus, Feline leukemia virus infection, Feline pathogens, Endangered species, Outbreak
The Iberian lynx (Lynx pardinus) is native to the Iberian peninsula and is considered the most endangered felid species in the world (Nowell and Jackson, 1996, Nowell, 2002, IUCN, 2007). Only two isolated populations live in southern Spain (Doñana and Sierra Morena areas), where only 40–50 and 150–200 individuals, respectively, are estimated to remain (Ferreras et al., 2010). During the last decades, this species has been especially threatened due to the decline of its basic prey (the wild European rabbit, Oryctolagus cuniculus), fragmentation and loss of habitat, and non-natural mortality due to illegal hunting and road kills (Ferreras et al., 2010). In addition, a phylogenetic study has recently documented the limited genetic diversity of the Iberian lynx population (Johnson et al., 2004), and histopathological studies revealed generalized immune depletion in these animals, apparently unrelated to infectious agents or malnutrition (Peña et al., 2006). Low population size, coupled with the low genetic variability, may contribute to render this species particularly susceptible to infectious diseases. Although limited, information is available on the incidence of Cytauxzoon felis (C. felis, a Theileria-like agent), Toxoplasma gondii, hemotropic mycoplasmas, Mycobacterium bovis, Ehrlichia and Anaplasma species, Bartonella henselae, Chlamydophila felis and several parasitic diseases in captive and wild individuals (Briones et al., 2000, Vicente et al., 2004, Luaces et al., 2005, Millán et al., 2007, Millán et al., 2009, Sobrino et al., 2007, Willi et al., 2007, Meli et al., 2009). Occasional exposure to viral infections such as feline herpes virus (FHV), feline calicivirus (FCV), feline parvovirus (FPV), canine distemper virus (CDV) and feline leukemia virus (FeLV) may occur (Luaces et al., 2008, Millán et al., 2009, Roelke et al., 2008, Meli et al., 2009).
Recently, a severe FeLV epidemic struck two small subpopulations of the Doñana area. Within a 6-month period starting in December 2006, 12 lynxes from the northern area of Doñana (10 in a subpopulation called Coto del Rey, and two in Dehesa de Gato, about 12–15 km away) were infected with FeLV. Eleven of these animals were antigenemic (p27-positive), and subsequently seven died, four while still in the wild and three after placement in a quarantine enclosure (López et al., 2009, Meli et al., 2009). As this was the first documented severe epidemic of FeLV to cause a high rate of mortality in Iberian lynxes we focussed on the following actions: (1) characterize FeLV as an isolated event and evaluate the potential for future severe outbreaks of highly pathogenic strains, (2) determine if Iberian lynxes are particularly susceptible to FeLV, and (3) evaluate the potential for other infectious agents to cause such fatal outbreaks and to represent additional threats to the survival of the Iberian lynx. Here, we provide a summary of the steps taken to reduce the threat originating from these infections and to raise public awareness of the critical situation of the Iberian lynx and the necessity to implement a continuous epidemiologic survey and management measures to reduce the infection risks from feral cats and dogs.
1. A FeLV epidemic in Iberian lynxes
As part of a research project to evaluate a supplementary feeding program for Iberian lynxes and determine the health and reproductive condition of free-ranging lynxes of the Doñana National Park, as well as other management activities outside the National Park, free-ranging animals were annually captured starting from December 2005. Samples from these lynxes were regularly tested to determine the presence of feline pathogens (Meli et al., 2009). In early December 2006, an adult male from the Coto del Rey subpopulation of the Doñana National Park was found to be FeLV provirus- and p27-positive. The same animal had been negative in the previous examination in December 2005. However, at time of immobilization in December 2006 this animal showed no signs of disease and was released. In Coto del Rey, another two adult males, three adult females, two young males and two young females were captured in December 2006 and were FeLV-negative. Coto del Rey is a small patch of Mediterranean scrubland habitat with a high lynx density and stable lynx reproduction (Palomares et al., 2001, Palomares et al., 2005). Contemporaneously, another five lynxes (two adult females, one adult male, one young female and one young male) were captured for evaluation in December 2006 in another subpopulation of the center of the Doñana National Park called Reserva Biologica de Doñana (RBD) and were also FeLV-negative. In addition, another adult male, an adult female and a young female, all FeLV-negative, had been captured in December 2006 in another area outside the national park called Dehesa de Gato, situated to 12–15 km to the northeastern from Coto del Rey. Subsequently, between March 13th and May 17th, 2007 all three of the adult males of the Coto del Rey subpopulation were found dead. Another one from Dehesa de Gato was found to be extremely weak with poor body condition, and was euthanized. Necropsies of these lynxes indicated that they apparently died of opportunistic bacterial infections; all of them tested FeLV provirus- and p27-positive.
After these findings, a management control program was started to screen as many free-ranging Iberian lynxes as possible from the Doñana population to remove viremic animals from the field to reduce the infectious pressure and to stop FeLV dissemination in that area. Viremic lynxes were placed in quarantine in a rescue station. Furthermore, vaccination of negative and latently infected individuals was carried out. Lynxes were tested for antigenemia (FeLV ELISA Snap test IDEXX®) upon immobilization, and negative animals were vaccinated using a Canarypox-vectored FeLV vaccine (Merial PureVax FeLV®, kindly donated by Merial France) and released (López et al., 2009).
Still, the epidemic continued with eight additional animals FeLV-infected (seven from Coto del Rey and one from Dehesa de Gato) of which seven were antigenemic. Three more lynxes (two sub-adult males and one young female) succumbed due to a very acute anemic disease in the rescue center. In total, 12 animals were found to be FeLV-infected, seven died: a detailed description of the time course of this FeLV epidemic is depicted in Fig. 1 .
Fig. 1.
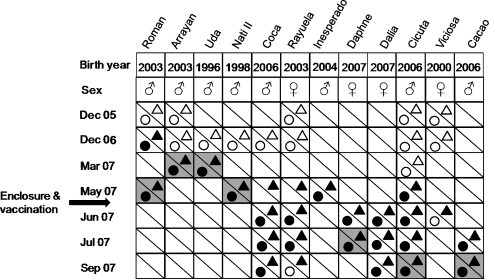
Time course of the FeLV epidemics in the Doñana region (Coto del Rey and Dehesa de Gato). Schematic representation of the FeLV epidemic time course in the Coto del Rey and Dehesa de Gato subpopulations from the Doñana area following the blood collections from December 2005 to September 2007. FeLV-infected lynxes are defined by name, birth date and sex. Collection dates are defined by month and year of collection. Circles: p27 antigen; triangles: provirus. Open symbols: negative result; closed symbols: positive result. Gray shading: animals were found dead or had to be euthanized due to poor conditions. Arrow: time point when the management decided to screen all animals in the region, vaccinate non-viremic animals and report viremic lynxes into the quarantine station. In total 12 animals were found to be infected with FeLV, 11 were found to be viremic and 7 died (4 in the wild, 3 in captivity). All animals but Coca were positive to hemotropic mycoplasmas. Parental relationships: Roman and Rayulela are brother and sister, Viciosa is their mother; Daphne, Dalia, Coca and Cicuta are Rayuela cub‘s. Inesperado escaped from the enclosure and so far nothing is known about his fate.
With the exception of an adult female, all surviving animals from the quarantine station remained viremic (p27-positive) in each subsequent examination (bimonthly), the last of which was in December 2008. Due to the scarcity of females at breeding age in the region, the animal that ostensibly recovered from FeLV viremia was brought back to the wild. The outbreak was successfully confined to the subpopulations where the virus had appeared and no additional FeLV-related clinical cases/FeLV-positive lynxes were found since August 2007.
2. Molecular characterization of Iberian lynx FeLV strains
In a time frame of about 4 years, from late 2003 to late 2007, 77 free-ranging Iberian lynxes were tested for the presence of FeLV, as well as other infectious agents (Meli et al., 2009). Fourteen lynxes, 13 from the Doñana area (12 animals from the epidemic in 2007 and one animal “Arena” found dead from a road kill accident in 2004) and one from Sierra Morena, tested positive for FeLV-A provirus (Table 1 ), but negative for FeLV-B and FeLV-C (data not shown). FeLV provirus was amplified and sequenced from 11 positive lynxes from the Doñana area. Nucleotide sequence analysis of the surface unit (SU) of the env gene revealed a common origin for the provirus found in Doñana lynxes in 2007 (99.5–100% identity). The sequences clustered with, and were 97.9–98.2% identical to, the FeLV-A/61E strain (Fig. 2A) (Pedersen et al., 1987, Overbaugh et al., 1988), while the sequence obtained from the lynx found to be provirus-positive in 2004 was only 94.8–95% identical to the other sequences and was more related to FeLV-A/Rickard (97.4% identity, Fig. 2A). The predicted amino acid sequences of the SU glycoprotein from animals infected in 2007 presented an identity/similarity of 97.3–97.6%/98.6–98.9% to FeLV-A/61E and of 95.9–96.2%/97.6–97.8% to FeLV-A/Rickard, respectively, while the identity/similarity of the sequence from the animal infected in 2004 was 97.3%/98.6% to FeLV-A/61E, 96.2%/97.8% to FeLV-A/Rickard, and 96.7–97.0%/98.9–99.2% to the 2007 sequences, respectively. At the protein level, all sequences from the Iberian lynxes clustered together with FeLV-A/61E and FeLV-A/Rickard (Fig. 2B). No sequences were found that showed a close relationship to FeLV-A/61C, associated with FeLV-A/61E in cats with severe immunosuppression.
Table 1.
Overview of the prevalence of infections in Iberian lynxes.
| Agent | Doñana |
Sierra Morena |
||
|---|---|---|---|---|
| Serology | PCR | Serology | PCR | |
| Pos/total (%) | Pos/total (%) | Pos/total (%) | Pos/total (%) | |
| Viral infections | ||||
| FeLV | 11/45 (24.4)a | 13/45 (28.9) | 0/30 (0)a | 1/32 (3.1) |
| FIV | 0/44 (0) | 0/45 (0) | 0/30 (0) | 0/32 (0) |
| FCoV | 7/44 (15.9) | 0/45 (0) | 12/30 (40.0) | 0/32 (0) |
| FHV | 7/44 (15.9) | 0/45 (0) | 2/30 (6.7) | 0/30 (0) |
| FPV | 13/44 (29.5) | 2/45 (4.4) | 9/30 (30.0) | 0/30 (0) |
| FCV | 15/44 (34.1) | 0/45 (0) | 14/30 (47.6) | 0/30 (0) |
| CDV | 11/44 (25.0) | 1/45 (2.2) | 1/30 (3.3) | 0/30 (0) |
| Protozoan infections | ||||
| Cytauxzoon felis | 0/45 (0) | 24/32 (75.0) | ||
| Bacterial infections | ||||
| M. haemofelis | 16/45 (35.6) | 9/32 (28.1) | ||
| C.M. haemominutum | 13/45 (28.9) | 14/32 (43.8) | ||
| C.M. turicensis | 6/45 (13.3) | 4/32 (12.5) | ||
| A. phagocytophilum | 1/44 (2.3) | 0/45 (0) | 3/30 (10) | 0/30 (0) |
| B. henselae | 6/45 (13.3) | 10/30 (33.3) | ||
| Chlamydopila felis | 0/45 (0) | 1/30 (3.3) | ||
FeLV: feline leukemia virus; FIV: feline immunodeficiency virus; FCoV: feline coronavirus; FHV: feline herpesvirus; FPV: feline parvovirus; FCV: feline calicivirus; CDV: canine distemper virus; M = mycoplasma; C.M. = Candidatus mycoplasma; A = Anaplasma; B = Bartonella.
p27 antigen.
Fig. 2.
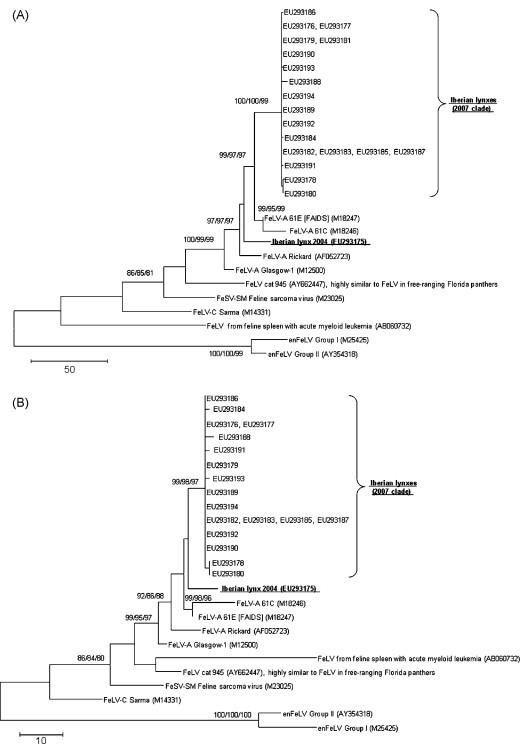
Evolutionary relationships of Iberian lynx FeLV SU (Meli et al., 2009). The maximum parsimony (MP) tree is shown. Trees are drawn to scale; length is in terms of the number of changes over the entire sequence. (A) Relationships at the DNA level. MP tree length = 643, consistency index = (0.744726), retention index = (0.827143), composite index = 0.671491 (0.615995) for all sites and parsimony-informative sites (in parentheses). The codon positions included were 1st + 2nd + 3rd + noncoding. There were a total of 1424 base positions in the final dataset, of which 311 were parsimony-informative. (B) Relationships at the protein level. MP tree length = 253, consistency index = (0.861538), retention index = (0.876712), composite index = 0.783150 (0.755321) for all sites and parsimony-informative sites (in parentheses). There were a total of 473 amino acid positions in the final dataset, of which 108 were parsimony-informative. Phylogenetic trees were constructed using MEGA3 (Kumar et al., 2004). Bootstrap support (1000 replicates) was calculated by the neighbor-joining/minimum evolution/MP methods and considered significant when >70% (Felsenstein, 1985). MP trees were obtained using the close-neighbour-interchange algorithm (Nei and Kumar, 2000) with search level 3 (Felsenstein, 1985), in which initial trees were obtained by random addition of sequences (10 replicates). All alignment gaps were treated as missing data. Branch lengths were calculated using the average pathway method (Nei and Kumar, 2000).
The sequences originating from the epidemic in 2007 form a separate cluster from those originating from the animal found positive in 2004. This may indicate that FeLV had been introduced into the Iberian lynx population on at least at two occasions.
The origin of the virus was most probably a feral or domestic cat. Contact with and killing of other carnivore species, such as domestic cats (Felis catus), has been well documented in the Iberian lynx (Palomares and Caro, 1999). Screening for endogenous FeLV (enFeLV)-related sequences in these Iberian lynxes was essentially negative: only five of the 77 lynxes tested by quantitative real-time PCR displayed weak signals (less than one copy per 10,000 cells), yet incompatible with the presence of enFeLV sequences, which have to be present in at least one copy per cell. These results can only be explained by the presence of either endogenous sequences related to enFeLV of domestic cats, or of a co-infection with another gammaretrovirus, both showing a weak cross-reactivity to the real-time PCR systems used. The absence of FeLV-B in infected Iberian lynxes is not surprising: FeLV-B is the result of a recombination between exogenous and enFeLV, and a relatively frequent mechanism leading to increased pathogenicity of FeLV, with an incidence usually higher than 50% in diseased domestic cats (Coelho et al., 2008). However, it has still to be elucidated whether recombination with as yet to be identified endogenous gammaretroviruses closely related to FeLV might have induced disease in Iberian lynxes through a mechanism analogous to FeLV-B in infected domestic cats (Meli et al., 2009).
3. Other infectious agents in the Iberian lynx and host immune response
Blood and fecal samples from 77 free-ranging Iberian lynxes collected between late 2003 and September 2007 were tested by serological and molecular methods for the presence of antibodies and antigens of different infectious agents (Meli et al., 2009). Animals were screened for the presence of viral agents such as FHV, FCV, FPV, FCoV, FeLV, FIV, and CDV. These viral infections are known to affect wild felids (Brown et al., 1994, Hofmann-Lehmann et al., 1996, Roelke-Parker et al., 1996). In addition, animals were tested for C. felis, Anaplasma phagocytophilum, hemotropic mycoplasmas, B. henselae, and Chlampydophila felis. In the Doñana area antibody titers to FHV and CDV were found more frequent than in the Sierra Morena area. In contrast, antibody titers to FCoV and to FCV were found more frequent in the Sierra Morena area. CDV- and FPV-antigen positive animals were found only in the Doñana area, while all C. felis-positive lynxes originated from Sierra Morena. Hemotropic mycoplasmas were highly prevalent in both populations. No evidence of active infections with FIV, FCoV, FHV, FCV, or A. phagocytophilum was found (Table 1). Statistically significant associations (based on PCR results) were found between FeLV infection and infections with M. haemofelis (p Fisher = 0.0004) and ‘Candidatus M. turicensis’ (p Fisher = 0.0205). The other pathogens did not show any significant association with FeLV infection.
The high pathogenicity observed in FeLV-infected Iberian lynxes might be due to host, rather than to viral factors. The genetic diversity of the Doñana lynx population is lower than that of the Sierra Morena population (Johnson et al., 2004); FeLV could have a higher pathogenic potential in inbred animals. Cheetahs, another wild felid species with reduced genetic diversity, also demonstrate increased susceptibility to infectious disease agents (Heeney et al., 1990). Recently, an immune-mediated systemic disease of unknown origin has been postulated in the Iberian lynx (Peña et al., 2006). Furthermore, the presence of co-infections could also have contributed to the high pathogenicity of FeLV in the lynxes. A significant association between FeLV infection and haemotropic mycoplasma infections, i.e., M. haemofelis and ‘Candidatus M. turicensis’, was found. Hemotropic mycoplasma infections have been found in the Iberian lynx population prior to the detection of FeLV infection. Thus, it is possible that pre-existing hemoplasma infections may have influenced FeLV infection outcome. The increased frequency of FeLV infection in lynxes positive for hemotropic mycoplasmas could also reflect simultaneous transmission of these agents with FeLV during cat-to-lynx contact.
4. Prospective of the susceptibility of Iberian lynx to future disease outbreaks
In a recently published retrospective study (Luaces et al., 2008), six of 21 lynxes sampled between 1993 and 2003 were found to be FeLV provirus-positive in Doñana and Sierra Morena areas, but not antigenemic. Although no clinical data were reported in this study, there was no indication that FeLV infection could represent a major risk for the lynx populations. From late 2003 to late 2006, FeLV infection (provirus) was detected in one animal. It is likely that FeLV infection in lynxes is rare, not endemic within lynx populations, and most likely originating from feral cats, which are not infrequent in the Doñana area (Palomares and Delibes, 1993). The FeLV antigen prevalence was reported to be ∼16% in Spanish cats (Arjona et al., 2000); it might be even higher around Doñana (G. Lopez, unpublished data). Furthermore, lynxes frequently kill other smaller carnivores including cats (Palomares and Caro, 1999), therefore, a high risk of FeLV transmission from domestic cats to lynxes is likely. Once an Iberian lynx is infected with FeLV it may readily spread the infection through the lynx population: especially males are expected to play an important role in transmitting the virus as they fiercely fight for territory acquisition and access to females, particularly during the mating season (Ferreras et al., 1997). This could be the explanation why all males from the Coto del Rey subpopulation became infected within a few months. Transmission of the infection to females could follow from mating activities, when the tomcat bites the females.
Since the FeLV strain found in Iberian lynx was highly pathogenic, managers tried to prevent the widespread of the diseases by starting a campaign to vaccinate as lynxes as possible using a Canarypox-based FeLV vaccine (Tartaglia et al., 1993, López et al., 2009). Although vaccination did not induce sterilizing immunity in domestic cats, it was nonetheless able to stimulate the immune system to a degree that allowed the cats to overcome the infection rapidly and to clear most of the viral RNA from the blood (Hofmann-Lehmann et al., 2007). Therefore, even if immunization does not completely protect lynxes against transient infection, there is justified hope that the outcome of FeLV infection will be less severe. In addition to vaccination, it is also important to decrease the infectious pressure on lynxes arising from domestic cats. Feral cats should be removed from the natural areas inhabited by lynx, and the domestic cats living in houses and villages close to lynx areas should also be vaccinated against FeLV infection to reduce the risk of FeLV transmission to lynxes.
Conflict of interest
None of the authors has a financial or personal relationship with other people or organizations that could inappropriately influence or bias the paper.
Acknowledgements
The authors would like to thank Y. Gahler, E. Gönczi, T. Meili Prodan, B. Pineroli, R. Tandon, E. Rogg, E. Schuler, B. Weibel and B. Willi for excellent laboratory assistance and helpful support. The authors are indebted to the Environmental Council of the Government of Andalusia, southern Spain, for providing the Iberian lynx samples. We are grateful to our collaborators in the Iberian Lynx Conservation Life Projects in Andalusia (LIFENAT 02/E/8609 and LIFENAT 06/E/0209), the Doñana National Park, and the ex situ Conservation Program for sampling the free-ranging Iberian lynxes and to Laura Peña from the Departamento de Patologia for her expertise in pathology. Some of the lynxes used in the present study were trapped, radio-tracked and sampled under the research projects CGL2004-00346/BOS of the Spanish Ministry of Education and Science, and 17/2005 of the Spanish Ministry of the Environment under the National Parks research program, with the collaboration of Land Rover España S.A. Laboratory work was performed using the logistics of the Center for Clinical Studies at the Vetsuisse Faculty of the University of Zurich. R.H.L. is the recipient of a professorship by the Swiss National Science Foundation (PP00B 102866 and PP00P3-119136).
References
- Arjona A., Escolar E., Soto I., Barquero N., Martin D., Gómez-Lucia E. Seroepidemiological survey of infection by feline leukemia virus and immunodeficiency virus in Madrid and correlation with some clinical aspects. Journal of Clinical Microbiology. 2000;38:3448–3449. doi: 10.1128/jcm.38.9.3448-3449.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Briones V., de Juan L., Sánchez C., Vela A.I., Galka M., Montero N., Goyache J., Aranaz A., Mateos A., Domínguez L. Bovine tuberculosis and the endangered Iberian lynx. Emerging Infectious Diseases. 2000;6:189–191. doi: 10.3201/eid0602.000214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown E.W., Yuhki N., Packer C., O’Brien S.J. A lion lentivirus related to feline immunodeficiency virus: epidemiologic and phylogenetic aspects. Journal of Virology. 1994;68:5953–5968. doi: 10.1128/jvi.68.9.5953-5968.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coelho F.M., Bomfim M.R., de Andrade Caxito F., Ribeiro N.A., Luppi M.M., Costa E.A., Oliveira M.E., Da Fonseca F.G., Resende M. Naturally occurring feline leukemia virus subgroups A and B infections in urban domestic cats. Journal of General Virology. 2008;89:2799–2805. doi: 10.1099/vir.0.2008/003855-0. [DOI] [PubMed] [Google Scholar]
- Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39:783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x. [DOI] [PubMed] [Google Scholar]
- Ferreras P., Beltran J.F., Aldama J.J., Delibes M. Spatial organization and land tenure system of the endangered Iberian lynx (Lynx pardinus) Journal of Zoology. 1997;243:163–189. [Google Scholar]
- Ferreras, P., Rodríguez, A., Palomares, F., Delibes, M., 2010. Iberian lynx: the uncertain future of a critically endangered cat. In: Macdonald, D.W., Loveridge, A.J. (Eds.), The Biology and Conservation of Wild Felids. Oxford University Press, in press.
- Heeney J.L., Evermann J.F., McKeirnan A.J., Marker-Kraus L., Roelke M.E., Bush M., Wildt D.E., Meltzer D.G., Colly L., Lukas J., Manton V.J., Caro T., O’Brien S.J. Prevalence and implications of feline coronavirus infections of captive and free-ranging cheetahs (Acinonyx jubatus) Journal of Virology. 1990;64:1964–1972. doi: 10.1128/jvi.64.5.1964-1972.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hofmann-Lehmann R., Fehr D., Grob M., Elgizoli M., Packer C., Martenson J.S., O’Brien S.J., Lutz H. Prevalence of antibodies to feline parvovirus, calicivirus, herpesvirus, coronavirus, and immunodeficiency virus and of feline leukemia virus antigen and the interrelationship of these viral infections in free-ranging lions in east Africa. Clinical and Diagnostic Laboratory Immunology. 1996;3:554–562. doi: 10.1128/cdli.3.5.554-562.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hofmann-Lehmann R., Cattori V., Tandon R., Boretti F.S., Meli M.L., Riond B., Pepin A.C., Willi B., Ossent P., Lutz H. Vaccination against the feline leukaemia virus: outcome and response categories and long-term follow-up. Vaccine. 2007;25:5531–5539. doi: 10.1016/j.vaccine.2006.12.022. [DOI] [PubMed] [Google Scholar]
- IUCN . 2007. IUCN Red List of Threatened Species.http://www.iucnredlist.org/search/details.php/12520/summ [Google Scholar]
- Johnson W.E., Godoy J.A., Palomares F., Delibes M., Fernandez M., Revilla E., O’Brien S.J. Phylogenetic and phylogeographic analysis of Iberian lynx populations. Journal of Heredity. 2004;95:19–28. doi: 10.1093/jhered/esh006. [DOI] [PubMed] [Google Scholar]
- Kumar S., Tamura K., Nei M. MEGA3: integrated software for molecular evolutionary genetics analysis and sequence alignment. Briefings in Bioinformatics. 2004;5:150–163. doi: 10.1093/bib/5.2.150. [DOI] [PubMed] [Google Scholar]
- López G., López-Parra M., Fernández L., Martínez-Granados C., Martínez F., Meli M.L., Gil-Sánchez J.M., Viqueira N., Díaz-Portero M.A., Cadenas R., Lutz H., Vargas A., Simón M.A. Management measures to control a FeLV outbreak in the endangered Iberian lynx (Lynx pardinus) Animal Conservation. 2009;12:173–182. [Google Scholar]
- Luaces I., Aguirre E., García-Montijano M., Velarde J., Tesouro M.A., Sánchez C., Galka M., Fernández P., Sainz A. First report of an intraerythrocytic small piroplasm in wild Iberian lynx (Lynx pardinus) Journal of Wildlife Diseases. 2005;41:810–815. doi: 10.7589/0090-3558-41.4.810. [DOI] [PubMed] [Google Scholar]
- Luaces I., Domenech A., García-Montijano M., Collado V.M., Sanchez C., Tejerizo J.G., Galka M., Fernández P., Gómez-Lucia E. Detection of feline leukemia virus in the endangered Iberian lynx (Lynx pardinus) Journal of Veterinary Diagnostic Investigation. 2008;20:381–385. doi: 10.1177/104063870802000325. [DOI] [PubMed] [Google Scholar]
- Meli M.L., Cattori V., Martínez F., López G., Vargas A., Simón M.A., Zorrilla I., Muñoz A., Palomares F., López-Bao J.V., Pastor J., Tandon R., Willi B., Hofmann-Lehmann R., Lutz H. Feline leukemia virus and other pathogens as important threats to the survival of the critically endangered Iberian lynx (Lynx pardinus) PLoS ONE. 2009;4:e4744. doi: 10.1371/journal.pone.0004744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Millán J., Naranjo V., Rodrìguez A., Pérez de la Lastra J.M., Mangold A.J., De la Fuente J. Prevalence of infection and 18S rRNA gene sequences of Cytauxzoon species in Iberian lynx (Lynx pardinus) in Spain. Parasitology. 2007;134:995–1001. doi: 10.1017/S003118200700248X. [DOI] [PubMed] [Google Scholar]
- Millán J., Candela M.G., Palomares F., Cubero M.J., Rodríguez A., Barral M., de la Fuente J., Almeria S., Leon-Vizcaino L. Disease threats to the endangered Iberian lynx (Lynx pardinus) Veterinary Journal. 2009;182(1):114–124. doi: 10.1016/j.tvjl.2008.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nei M., Kumar S. Oxford University Press; New York, USA: 2000. Molecular Evolution and Phylogenetics. [Google Scholar]
- Nowell K., Jackson P. IUCN/SSC; Gland, Switzerland: 1996. Wild Cats: Status Survey and Conservation Action Plan. [Google Scholar]
- Nowell K. IUCN/SSC Cat Specialist Group, Authority (ISRLF); 2002. Revision of the Felidae Red List of Threatened Species.http://lynx.uio.no/lynx/catsgportal/red-list/03_cats-and-red-list/nowell-cat_news_no37_2002.pdf [Google Scholar]
- Overbaugh J., Donahue P.R., Quackenbush S.L., Hoover E.A., Mullins J.I. Molecular cloning of a feline leukemia virus that induces fatal immunodeficiency disease in cats. Science. 1988;239:906–910. doi: 10.1126/science.2893454. [DOI] [PubMed] [Google Scholar]
- Palomares F., Delibes M. Note on the movements of a free-ranging male domestic cat in southwestern Spain. Hystrix. 1993;5:11–15. [Google Scholar]
- Palomares F., Caro T.M. Interspecific killing among mammalian carnivores. The American Naturalist. 1999;153:492–508. doi: 10.1086/303189. [DOI] [PubMed] [Google Scholar]
- Palomares F., Delibes M., Revilla E., Calzada J., Fedriani J.M. vol. 148. Wildlife Monographs-Wildlife Society; Washington: 2001. (Spatial Ecology of Iberian Lynx and Abundance of European Rabbits in Southwestern Spain). [Google Scholar]
- Palomares F., Revilla E., Calzada J., Fernández N., Delibes M. Reproduction and pre-dispersal survival of Iberian lynx in a subpopulation of the Doñana National Park. Biological Conservation. 2005;122:53–59. [Google Scholar]
- Pedersen N.C., Ho E.W., Brown M.L., Yamamoto J.K. Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome. Science. 1987;235:790–793. doi: 10.1126/science.3643650. [DOI] [PubMed] [Google Scholar]
- Peña L., García P., Jiménez M.A., Benito A., Alenza M.D.P., Sánchez B. Histopathological and immunohistochemical findings in lymphoid tissues of the endangered Iberian lynx (Lynx pardinus) Comparative Immunology, Microbiology and Infectious Diseases. 2006;29:114–126. doi: 10.1016/j.cimid.2006.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roelke-Parker M.E., Munson L., Packer C., Kock R., Cleaveland S., Carpenter M., O’Brien S.J., Pospischil A., Hofmann-Lehmann R., Lutz H., Mwamengele G.L.M., Mgasa M.N., Machange G.A., Summers B.A., Appel M.J.G. A canine distemper virus epidemic in Serengeti lions (Panthera leo) Nature. 1996;379:441–445. doi: 10.1038/379441a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roelke M.E., Johnson W.E., Millán J., Palomares F., Revilla E., Rodriguez A., Calzada J., Ferreras P., Leon-Vizcaino L., Delibes M., O’Brien S.J. Exposure to disease agents in the endangered Iberian lynx (Lynx pardinus) European Journal of Wildlife Research. 2008;54:171–178. doi: 10.1007/s10344-007-0122-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sobrino R., Cabezon O., Millán J., Pabon M., Arnal M.C., Luco D.F., Gortazar C., Dubey J.P., Almeria S. Seroprevalence of Toxoplasma gondii antibodies in wild carnivores from Spain. Veterinary Parasitology. 2007;148:187–192. doi: 10.1016/j.vetpar.2007.06.038. [DOI] [PubMed] [Google Scholar]
- Tartaglia J., Jarrett O., Neil J.C., Desmettre P., Paoletti E. Protection of cats against feline leukemia virus by vaccination with a canarypox virus recombinant, ALVAC-FL. Journal of Virology. 1993;67:2370–2375. doi: 10.1128/jvi.67.4.2370-2375.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vicente J., Palomares F., Ruíz de Ibañez R., Ortiz J. Epidemiology of Ancylostoma spp. in the endangered Iberian lynx (Lynx pardinus) in the Doñana National Park, south-west Spain. Journal of Helminthology. 2004;78:179–183. doi: 10.1079/joh2003216. [DOI] [PubMed] [Google Scholar]
- Willi B., Filoni C., Catao-Dias J.L., Cattori V., Meli M.L., Vargas A., Martínez F., Roelke M.E., Ryser-Degiorgis M.-P., Leutenegger C.M., Lutz H., Hofmann-Lehmann R. Worldwide occurrence of feline hemoplasma infections in wild felid species. Journal of Clinical Microbiology. 2007;45:1159–1166. doi: 10.1128/JCM.02005-06. [DOI] [PMC free article] [PubMed] [Google Scholar]