

Abstract
Respiratory syncytial virus (RSV) is the most common cause of respiratory infection in infants, and no vaccine is available. In this report, recombinant AIK-C measles vaccines, expressing the RSV G or F protein of subgroup A (MVAIK/RSV/G or F), were investigated as a RSV vaccine candidate. MVAIK/RSV/G or F had the original ts phenotype and expressed RSV/G or F protein. Cross-reactive neutralizing antibodies against RSV subgroups A and B were detected in cotton rats immunized intramuscularly with MVAIK/RSV/F but not MVAIK/RSV/G. In cotton rats infected with RSV, RSV was recovered and lung histopathological finding was compatible with interstitial pneumonia, demonstrating thickening of alveolar walls and infiltration of mononuclear cells. When cotton rats immunized with MVAIK/RSV/F were challenged with homologous RSV subgroup A, no infectious RSV was recovered and very mild inflammation was noted without RSV antigen expression. When they were challenged with subgroup B, protective efficacy decreased. When cotton rats immunized with MVAIK/RSV/G were challenged with RSV subgroup A, low levels of infectious virus were recovered from lung. When challenged with subgroup B, no protective effects was demonstrated, demonstrating large amounts of RSV antigen in bronchial-epithelial cells. MVAIK/RSV/F is promising candidate and protective effects should be confirmed in monkey model.
Keywords: Measles virus (MV), Respiratory syncytial virus (RSV), Cotton rat, Neutralizing antibodies
1. Introduction
Human respiratory syncytial virus (RSV) is a member of the family Paramyxoviridae in the order Mononegavirales. The Paramyxoviridae consist of two subfamilies, Paramyxovirinae and Pneumovirinae [1]. Classified into the genus Pneumovirus, RSV is characterized by a non-segmented, negative sense, single-stranded RNA genome, and has approximately 15,200 nucleotides. All members of the paramyxovirus family are similar in structure and characteristics [2]. Viral particles of RSV are surrounded by a lipid bilayer with two viral glycoproteins, G and F [1], involved in the attachment to, fusion with, and entry into cells during infection. G protein is not always required for infection and cell fusion and the expression of F protein alone leads to cell fusion [3]. RSV was first isolated in 1956 and two antigenically different subgroups, A and B, co-circulate [4]. RSV is the most common cause of lower respiratory infections in infants and young children worldwide, and is responsible for a variety of illnesses, including 20–25% of pneumonia cases and 45–50% of bronchiolitis cases among hospitalized children [5]. The peak of serious RSV infections is at 2–6 months of age and most children experience an RSV infection by two years of age [6]. The infection causes serious illnesses especially in babies born prematurely and having chronic lung diseases, or congenital heart diseases. RSV also causes lower respiratory tract infections in the elderly, and in immunocompromised hosts [7]. The global annual morbidity and mortality for RSV are estimated to be 64 million and 160,000 deaths, respectively [8].
A recent study of the immune response to RSV showed the importance of innate immunity in regulating adaptive immune responses [9]. Adaptive immunity is generally considered effective due to neutralizing antibodies (NT) and cellular immune responses for the clearance of viruses are influenced by innate inflammatory responses. Secretory and NT antibodies were generated after repeated infections with RSV, although the responses were weak in young infants [10]. The presence of IgG antibodies in the lung has been shown to reduce viral load [11]. Even a natural infection did not provide long-term protective immunity against reinfection in young infants, and a humanized monoclonal antibody against the F protein is available as a prophylaxis against RSV, or for reducing serious diseases in high-risk infants during epidemics [12]. However, the high medical costs for monthly administration mean that there is a great need to develop an RSV vaccine [13]. There are several obstacles to developing a RSV vaccine. An aluminium-precipitated formalin-inactivated RSV vaccine (FI-RSV) was developed in the 1960's, but did not prevent infections [14]. In fact, symptoms were exacerbated among recipients subsequently infected with RSV. FI-RSV generated only binding antibodies without neutralizing activity because of the denatured F protein, and did not induce cytotoxic T cell lymphocytes (CTL) activity [15]. Several strategies have been adopted to develop subunit vaccines, live attenuated vaccines through conventional methods of cloning or selecting ts mutants, genetically modified-strain by reverse genetics, and vaccinia virus vector-based recombinant vaccines [16], [17], [18].
Recently, a method for direct manipulation of the genomic RNA of Mononegavirales has been established, known as the infectious cDNA clone system [19]. The transcription and replication of minigenome RNA are driven by viral proteins, which are co-expressed by plasmids or helper viruses. Using this system, the infectious recombinant viruses can be retrieved from the authentic full-size genome cDNA [20], [21]. These “reverse genetics” techniques are powerful tools not only for basic research into viral properties, such as the characteristics of viral proteins, and mechanisms of replication, transcription and pathogenesis, but also for practical purposes, such as the development of new vaccines and viral vectors. As vector-based recombinant vaccines, human parainfluenza virus type III (HPIV III) vector-based, or Sendai virus vector-based vaccines have been evaluated [22], [23].
Current measles vaccines used throughout the world were attenuated from the Edmonston strain, classified as genotype A [24]. The AIK-C strain of the measles vaccine was developed in 1976 in Japan from the Edmonston strain, by plaque cloning through passages in sheep kidney cells and chicken embryonic cells at 33 °C [25]. It shows optimal growth at 33 °C and little or no growth at 39 °C [21]. The safety and immunogenicity of the AIK-C measles vaccine were established through clinical trials [26], [27], [28], [29]. Reverse genetics of the AIK-C live attenuated vaccine was performed and in this study, recombinant AIK-C MV vaccine strains encoding the RSV G or F protein were constructed, and immunogenicity and protective effects against RSV were investigated in cotton rats immunized with recombinant measles vaccines, expressing RSV G or F protein.
2. Materials and methods
2.1. Viral strains and cell cultures
The AIK-C seed strain for vaccine production was used. Wild-type strains of RSV subgroups A and B were isolated in HEp-2 cells from patients. Long and wild-type strains were used for the neutralization test (NT) against RSV subgroups A and B. 293T and HEp-2 cells were maintained in Eagle's MEM (Sigma–Aldrich, Dorset, UK) supplemented with 10% fetal bovine serum (FBS). Vero cells were maintained in Eagle's MEM supplemented with 5% FBS. B95a cells are marmoset B cell line, and maintained in RPMI-1640 medium (Sigma–Aldrich, Dorset, UK) supplemented with 10% FBS [30]. These media were supplemented with 4 mM l-glutamine, 10,000 IU/ml penicillin, and 10,000 μg/ml streptomycin.
2.2. Cloning of the RSV G and F genes
Genomic RNA was extracted from a clinical isolate of subgroup A and B, and the RSV genome was amplified by RT-PCR. The viral RNA was first converted to cDNA using a cDNA primer: 5′-ACACGATTTGCAATCAAACC-3′. The RSV G gene was amplified with 5′-GTTTCCATGGCCAAAACCAAGGACCAA-3′ and 5′-CCAAGCGGCCGCTAGTTTGTTGTGTTGGATGGAGA-3′, which amplified 894 bp. The RSV F gene was amplified with 5′-GTTGCCATGGAGTTGCCAATCCTCAA-3′ and 5′-TGTGGCGGCCGCTAACTAAATGCAATATTATTT-3′, which amplified 1722 bp. The F and G genes were cloned into pMV/20-77 using two restriction enzymes, Nco I and Not I (underlined sequences).
2.3. Construction of recombinant AIK-C
A schematic diagram of the strategy used for the construction of the recombinant cDNA plasmid is shown in Fig. 1 . The full length plasmid was divided from two parts as previously reported. The first half contained the N, P, M and F genes from the leader sequence to the Pac I site at nucleotide position 7238 of the AIK-C genome. The second half contained the H and L regions from the Pac I site from position 7238 of the AIK-C genome to the trailer sequence. The full-length cDNA, pMVAIK, was constructed using these two plasmids [31].
Fig. 1.
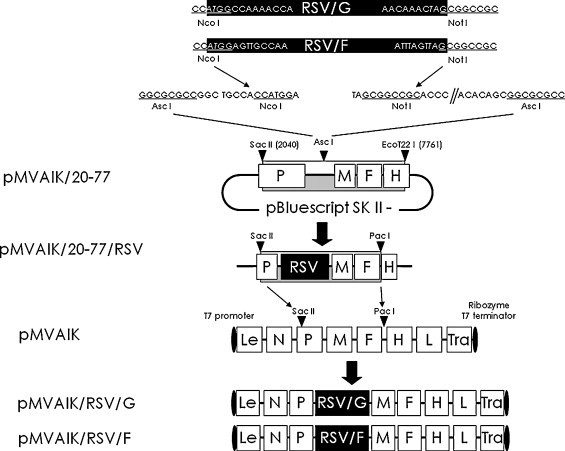
Strategy for the construction of the recombinant AIK-C genome cDNAs having RSV protein genes. The recombinant AIK-C viral cDNAs expressing RSV G or F protein were constructed based on AIK-C cDNA (pMVAIK). pMVAIK/20-77 was constructed for the cloning of foreign genes. The Asc I site was introduced by adding GGCGCG after position 3432 of AIK-C and R1 and R2 sequences were added. The Nco I–Not I fragment of RSV G or F was cloned into pMVAIK/20-77, designed as pMVAIK/20-77/RSV. pMVAIK/20-77/RSV had unique restriction enzyme sites, Sac II and Pac I sites, located in the P gene and between the F and H gene. The DNA fragments between the Sac II and Pac I sites of pMVAIK/20-77/RSV/G and pMVAIK/20-77/RSV/F were inserted into pMVAIK using Sac II and Pac I sites. The recombinant plasmid constructs were designated pMVAIK/RSV/G and pMVAIK/RSV/F, respectively.
The cloning vector for the RSV genome, pMVAIK/20-77, was constructed from positions 2040 (Sac II) to 7761 (EcoT22 I). The RSV G or F PCR product was digested with Nco I and Not I and ligated into pMVAIK/20-77, resulting in pMVAIK/20-77/RSV/G and pMVAIK/20-77/RSV/F, respectively. The pMVAIK/20-77/RSV/G or pMVAIK/20-77/RSV/F was digested with Sac II and Pac I and ligated into pMVAIK. Then, full-length infectious cDNA clones, pMVAIK/RSV/G and pMVAIK/RSV/F, were constructed.
2.4. Rescue of the infectious recombinant virus from cloned cDNA
Monolayers of 293T cells in 6-well plates were infected with the vaccinia virus MVAT7 pol, expressing T7 RNA polymerase. MVAT7 pol was derived from a highly attenuated and host range-restricted vaccinia virus, the Ankara strain [32]. Open reading frames of the N, P, and L genes were cloned downstream of the T7 promoter of pBluescript SK, and the expression plasmids pCIAN01, pCIAP01, and pCIAL01 were constructed [19], [21]. After 1 h of adsorption, the cells were washed with Opti-MEM (GIBCO, Grand Island, NY, US) and transfected with 0.5 μg of pCIAN01, 0.25 μg of pCIAP01, 0.1 μg of pCIAL01, and 1.5 μg of pMVAIK/RSV with TransIT-LT1 Reagent (Mirus Bio Corporation, US). After incubation at 33 °C for 3 h, the medium containing the transfection reagent/plasmid complex was replaced with fresh MEM supplied with 5% FBS. The transfected cells were incubated at 33 °C in 5% CO2 for 3 days. After 3 days, 293T cells were detached and co-cultured with B95a cells. When a demonstrable cytopathic effect (CPE) was observed, the supernatant and cell lysate were harvested and stocked.
2.5. Virus growth
To examine viral growth, B95a cells were infected with MVAIK, MVAIK/RSV/G, and MVAIK/RSV/F (m.o.i. = 0.02) and the plates were placed at temperatures of 33, 35, 37 and 39 °C. The culture fluids were obtained on days 1, 3, 5, and 7 of culture and infective titers were examined and expressed as TCID50/ml in B95a cells.
2.6. Indirect immuno-staining and Western blotting
B95a cells were infected with MVAIK, MVAIK/RSV/G or MVAIK/RSV/F at m.o.i. of 0.01 in 24-well plates and cultured for two days at 33 °C. B95a cells were collected and subjected to indirect immuno-staining without fixation to detect the surface expression. Polyclonal antibodies against RSV raised in goat (Abcam, Cambridge, UK) were used and the cells incubated for 1 h at 37 °C. The cells were washed extensively with phosphate-buffered saline with 0.05% Tween 20 (PBST), and stained with second antibodies against goat IgG conjugated with FITC, raised in rabbit (Vector Laboratories, Burlingame, CA, US), and thereafter, mouse monoclonal antibody against MV HA protein (kindly supplied by Dr. Sato, National Institute of Infectious Diseases, Japan) was used and followed by second antibodies against mouse IgG conjugated with rhodamine raised in goat (Rockland Immunochemicals, Gilbertsville, PA, US).
Vero cells were infected with MVAIK, MVAIK/RSVF, and MVAIK/RSV/G and HEp-2 were infected with RSV subgroup A, Long strain in a 24-well plate. Culture supernatants were collected and cells were freeze-thawed and total protein of 4 μg of supernatants and cell lysate was applied. Samples were subjected to Western blotting. Briefly, after SDS-PAGE, proteins were transferred to membrane (Immobilon; Millipore, Danvers, MA, US). Membranes were washed with PBST, incubated with an RSV polyclonal antibody raised in goats, washed again, and incubated with a donkey anti-goat IgG (H + L) conjugated with horse radish peroxidase (HRP). The final reaction was performed with a DAB SUBSTRATE KIT FOR PEROXIDASE (Vector Laboratories, Burlingame, CA, US) used as recommended by the manufacturer.
Culture medium of Vero cells infected with MVAIK/RSV/G or F was collected and fractionated through sucrose discontinuous gradient ultra-centrifugation. Fraction 1 was obtained at the top of the gradient, 30% sucrose, Fraction 2 between 30% and 45% sucrose, and Fraction 3 between 45% and 60% sucrose. Each fraction was electrophoresed and analyzed by Western blotting, using RSV polyclonal antibodies and monoclonal antibodies against MV N protein.
2.7. Immunogenicity in experimental animals
Six-week-old cotton rats were purchased from Harlan (Indianapolis, IN, US) and Charles River (USA). Five cotton rats for each group were immunized intramuscularly with 1 × 106 TCID50 of MVAIK, MVAIK/RSV/G or MVAIK/RSV/F. Serum samples were obtained immediately before and 1, 3, 5, 8, 12 and 16 weeks after immunization. Cotton rats immunized with MVAIK/RSV/G or F were boosted with the same dose after 16 weeks, and serum samples were collected one week after re-immunization (17 weeks).
2.8. Serology
Neutralization tests (NTs) against RSV were performed with the 50% plaque reduction assay, using Long strain and wild-type isolate of subgroup B. Briefly, serum samples were serially diluted by1:4, starting from a 1:10 dilution, and mixed with an equal volume of RSV (100 PFU) in MEM for 1 h at room temperature. The mixtures were inoculated on monolayers of HEp-2 cells in 24-well plates. Plates were incubated for 1 h at 37 °C in 5% CO2 and then overlaid with MEM supplemented with glutamine, antibiotics, 5% fetal bovine serum and 0.5% agar. After incubation for six days at 37 °C in 5% CO2, cells were fixed with 1% formalin. Agar was removed and cells were stained with neutral red. Plaque numbers were counted and NT antibody titers were calculated as the reciprocal of the serum dilutions that showed a 50% reduction of the plaque number.
For the particles agglutination (PA) test, gelatin particles were coated with purified measles virus antigen (Serodia®-Measles, Fuji Rebio, Tokyo, Japan). Sera were serially diluted two-fold, starting from a 1:10 dilution, and each serum dilution was mixed with an equal volume of gelatin particles to detect agglutination, according to the recommendations of the manufacturer. The PA antibody titers were expressed as the reciprocal of the serum dilution which induced particle agglutination.
2.9. Detection of the MV genome
Cotton rats were sacrificed 10 days after immunization with MVAIK/RSV/G and F, and samples of liver, kidney, spleen, lung, thymus, and nasal turbinate were obtained to detect the MV genome. The tissues were homogenized, and total RNA was extracted using an RNeasy® Plus Mini Kit (QIAGEN, MD, US), as recommended. TaqMan PCR was performed in the MV N gene region. Reverse-transcribed real-time PCR was performed using a FastStart TaqMan® Probe Master (Roche Meylan, France), and LightCycler®480 System II (Roche Meylan, France) using 1 μg of extracted mRNA. cDNA was synthesized using an One Step PrimeScript® RT-PCR Kit (TaKaRa Bio, Otsu, Japan). The parameters used were 1 cycle of 95 °C for 10 min, 45 cycles of 95 °C for 10 s, 60 °C for 30 s, and 72 °C for 1 s, and 1 cycle of 40 °C for 30 s. Reactions were performed in triplicate and genome copy numbers were determined by referring to the results of serial dilution of the corresponding plasmid, pCIAN01. The primers used in TaqMan PCR are shown in Table 1 .
Table 1.
Primer and probe sequences for the detection of the MVAIK N gene and RSV N gene by TaqMan real-time PCR.
| Primers | Sequences (5′–3′) | Genomic position |
|---|---|---|
| RSV-Long-N-(+) | aatgctaaaagaaatgggagagg | 411–470 |
| Probe | gctccaga | |
| RSV-Long-N-(−) | ccacaatcaggagaatcatgc | |
| MV-AIK-C-N-(+) | caagatcagtagagcggttgg | 1212–1274 |
| Probe | agcccaag | |
| MV-AIK-C-N-(−) | ctttgatcaccgtgtagaaatga | |
2.10. Protection against RSV
Seven week-old cotton rats were immunized intramuscularly with MVAIK/RSV/F or MVAIK/RSV/G and, five weeks later, challenged with 106 PFU/0.5 ml of RSV subgroups A and B. They were sacrificed four days after the challenge and nasal wash, BAL, nasal turbinate, and lung tissues were obtained. Lung samples were divided into two portions, one for pathological examination, and another for recovering the infective particles and RSV genome.
Tissues were homogenized and 0.1 ml volumes of serial 10-fold dilutions of homogenized samples were placed on HEp-2 cells and overlaid with MEM 5% FBS and 0.5% agar. Plaque numbers were counted after incubation for six days at 37 °C and infectivity was expressed as the number of plaques. RNA was extracted from nasal wash, BAL, nasal turbinate and lung homogenate. cDNA was synthesized and reverse-transcribed real-time PCR was done at position 1212–1274 of the RSV N genome, using the primers and TaqMan probe listed in Table 1. The RSV genome copy number was calculated by referring to a linear regression assay of serial dilutions of the corresponding plasmid.
Lungs were inflated to their normal volumes with 4% formalin and submerged in formalin for overnight fixation. The fixed tissue was embedded in paraffin, sectioned, and stained with hematoxylin–eosin, and immuno-staining was performed using four clone blend monoclonal antibodies against RSV P, F, and N proteins (AdB Serotec, UK), and anti-mouse IgG conjugated with HRP.
3. Results
3.1. Characteristics of recombinant viruses
MVAIK/RSV/G and MVAIK/RSV/F were recovered from full-length recombinant cDNA and MVAIK from vector cDNA. B95a cells were infected with MVAIK, MVAIK/RSV/G and MVAIK/RSV/F at a m.o.i. of 0.02. The culture medium was harvested on days 1, 3, 5, and 7 at 33 °C and the results are shown in Fig. 2 . Infectivity showed a peak titer of 105 TCID50/ml 5 days after infection. MVAIK/RSV/G and MVAIK/RSV/F grew as well as MVAIK in B95a cells. AIK-C has temperature-sensitivity (ts), showing efficient virus growth at 33 °C, but extremely poor at 39 °C, less than 10−4 in comparison with the result at 33 °C. MVAIK/RSV/G and MVAIK/RSV/F were examined for virus growth at 33, 35, 37 and 39 °C. The culture supernatants were harvested on day 7 of the culture and infectivity was examined. Both MVAIK/RSV/G and MVAIK/RSV/F showed 105 TCID50/ml at 33 °C, and MVAIK/RSV/F grew little at 37 °C. But, however, no infectious virus was detected at 39 °C, and the ts phenotype was maintained (Fig. 3 ).
Fig. 2.
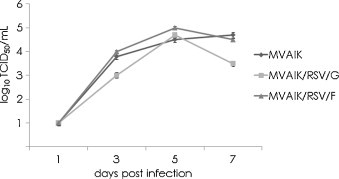
Growth of MVAIK, MVAIK/RSV/G, and MVAIK/RSV/F. B95a cells were infected with MVAIK, MVAIK/RSV/G, and MVAIK/RSV/F at m.o.i. of 0.02. Culture fluid was obtained on days 1, 3, 5, and 7 of culture at 33 °C. Infectivity is shown as mean titers of TCID50/ml assayed in B95a cells. Error bars show 1.0 S.D.
Fig. 3.
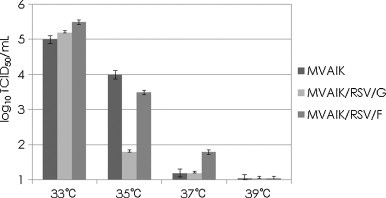
Temperature sensitivity of MVAIK, MVAIK/RSV/G, and MVAIK/RSV/F. B95a cells were infected with MVAIK, MVAIK/RSV/G, and MVAIK/RSV/F at a m.o.i. of 0.02. Culture fluid was obtained on day 5 and the infectivity at 33 °C, 35 °C, 37 °C, and 39 °C is shown as mean infectious titer (TCID50/ml). Error bars show 1.0 S.D.
3.2. Detection of RSV G or F protein
B95a cells were infected with MVAIK/RSV/F, MVAIK/RSV/G, and MVAIK at a m.o.i. of 0.01. Live cells were stained with monoclonal antibodies against measles HA and polyclonal antibodies against RSV and visualized with second antibodies conjugated with rhodamine or FITC, as shown in Fig. 4 . RSV F and MV HA proteins were observed diffusely on the surface of B95a cells infected with MVAIK/RSV/F. RSV G protein was detected in speckled pattern together with MV HA protein on the surface of B95a cells infected with MVAIK/RSV/G.
Fig. 4.
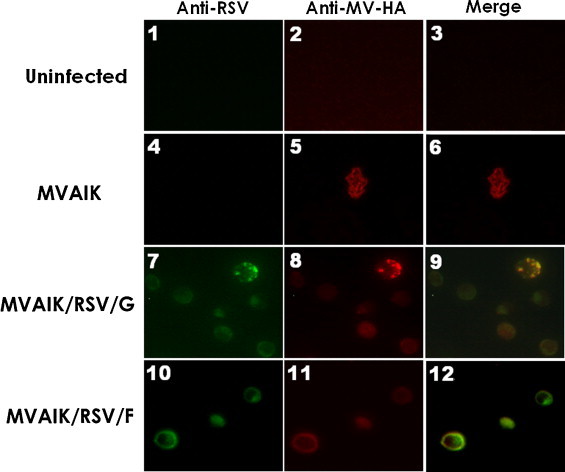
Expression of MV HA and RSV G or F protein. B95a cells were infected with MVAIK (panels 4 and 5), MVAIK/RSV/G (panels 7 and 8) or MVAIK/RSV/F (panels 10 and 11) at a m.o.i. of 0.01 in 24-well plate and cultured for two days at 33 °C. Uninfected B95a cells are shown in panels 1 and 2. B95a cells were collected and subjected to live cell staining without fixation to detect the surface expression. The expression of RSV (panels 1, 4, 7, and 10) and MV HA protein (panels 2, 5, 8, and 11) are shown. Panels 3, 6, 9 and 12 are merged images.
Culture medium and cell lysate were examined for the expression of RSV G and F by Western blotting and the results are shown in Fig. 5 . Live vector virus MVAIK and RSV were used for the negative and positive controls. RSV G and F proteins were detected in both supernatant and cell lysate infected with MVAIK/RSVF, and MVAIK/RSV/G, similar to those infected with RSV (Fig. 5, Panel A).
Fig. 5.
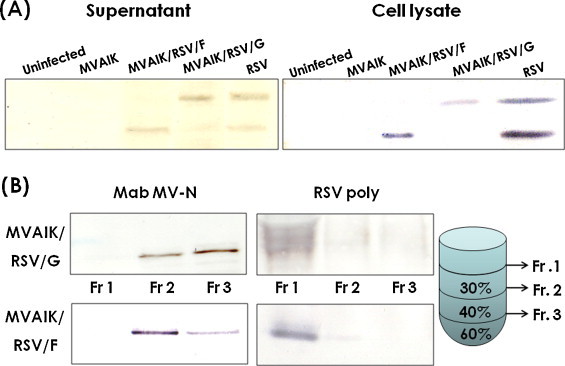
Results of Western blotting of culture supernatant, cell lysate, and purified recombinant measles viral particles. (A) Vero cells were infected with MVAIK, MVAIK/RSVF, and MVAIK/RSV/G and HEp-2 cells were infected with RSV subgroup A, Long strain, and were cultured in 1 ml in a 24-well plate. Just before the appearance of CPE, culture media was replaced with serum free medium (VP-SFM). 1 ml of culture medium was harvested and 100 μl of PBS was added in plate. Cells were freeze-thawed and cell lysate was clarified. As for the Western blotting, 1/30 of initial supernatants and 1/100 of cell lysate were subjected for experiments. They were stained with polyclonal antibodies against RSV. (B) Infectious particles were obtained through sucrose discontinuous gradient ultra-centrifugation. Fraction 1 was obtained at the top of the gradient of 30% sucrose, Fraction 2 between 30% and 45% sucrose, and Fraction 3 between 45% and 60% sucrose. Each fraction was analyzed by Western blotting, using RSV polyclonal antibodies and monoclonal antibodies against MV N protein.
Culture fluid was collected and fractionated through sucrose discontinuous gradient ultra-centrifugation. Fraction 1 was obtained at the top of the gradient, 30% sucrose, Fraction 2 between 30% and 45% sucrose, and Fraction 3 between 45% and 60% sucrose. Each fraction was electrophoresed and analyzed by Western blotting, using RSV polyclonal antibodies and a monoclonal antibody against the MV N protein. RSV G or F was detected in Fraction 1, and, whereas the MV N protein was detected in Fractions 2 and 3 (Fig. 5, Panel B). Accordingly, RSV G or F protein translated from the inserted gene was considered not to be incorporated into MV particles.
3.3. Immunogenicity of recombinant measles viruses
The recombinant viruses, MVAIK/RSV/G and MVAIK/RSV/F, were inoculated into cotton rats to confirm the immunogenicity intramuscularly of the inserted RSV G or F protein. Five cotton rats were immunized with MVAIK, MVAIK/RSV/G, and MVAIK/RSV/F for each study group and serum samples were obtained before and 1, 3, 5, 8, 12, and 16 weeks after immunization. The results are shown in Fig. 6 . PA antibodies against MV were detected three weeks after the immunization in all animals. High levels of PA antibody, 26–8 × 10 (1:640–1:2560), were maintained until 16th week in those immunized with MVAIK, MVAIK/RSV/G, and MVAIK/RSV/F. Two rats were reimmunized 16 weeks after the first immunization and sera were obtained one week after the reimmunization for each group. PA antibodies increased from 27.5 ± 1.5 to 210.5 ± 1.5 × 10 in the MVAIK/RSV/G group, and from 26.5 ± 1.5 to 28.5 ± 1.5 × 10 in the MVAIK/RSV/F group. PA antibodies against MV increased after the reimmunization by four to eight-fold.
Fig. 6.
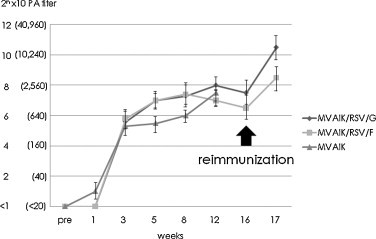
Serological response of PA antibodies against MV. PA antibody titers were examined, using Serodia®-Measles. PA titers are expressed as 2n × 10. Sera were collected before immunization, and 1, 3, 5, 8, 12, 16, and 17 weeks after immunization with MVAIK, MVAIK/RSV/G, and MVAIK/RSV/F. Five cotton rats were immunized and followed for 12 weeks. Mean PA titers ± 1.0 S.D. are shown. Two rats for each were reimmunized at the 16th week.
The results for NT antibodies against RSV are shown in Fig. 7 . In the cotton rats immunized with MVAIK/RSV/G, NT antibodies against RSV subgroup A were detected one week after immunization but the mean titer began to decrease 5 week after immunization. The mean NT titers against RSV subgroup A decreased to undetectable levels 12 weeks after the immunization. In the MVAIK/RSV/F group, NT antibodies against RSV subgroup A were detected one week after the immunization in all animals with a mean titer of 102.0 ± 0.7. High titers were observed at the 5th week with a mean of 102.6 ± 1.0. Levels of these antibodies were maintained until 16th week.
Fig. 7.
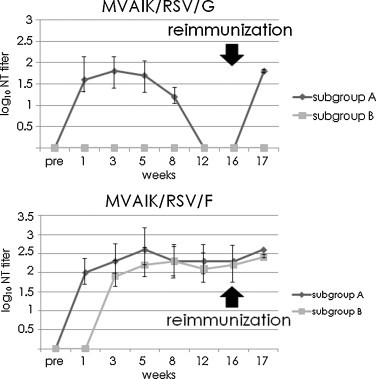
Development of NT antibodies against RSV. NT antibodies were examined using the RSV Long strain (Subgroup A) and wild-type RSV subgroup B strain. Sera were collected before immunization, and 1, 3, 5, 8, 12, 16, and 17 weeks after immunization. 50% plaque reduction NT titers are expressed 10n and are shown as mean NT titers with 1.0 S.D. The upper panel shows the immune response after immunization with MVAIK/RSV/G. The lower panel shows the results after immunization with MVAIK/RSV/F.
In this experiment, RSV source of the recombinant MVAIK/RSV/F or MVAIK/RSV/G was derived from the RSV subgroup A wild type. Cross immunity against RSV subgroup B was further investigated. In cotton rats immunized with MVAIK/RSV/F, NT antibodies against RSV subgroup B were detected at the 3rd week with a mean titer of 150 (102.1) and maintained for 16 weeks. However, cross-reactive antibodies against RSV subgroup B were not detected in the cotton rats immunized with MVAIK/RSV/G.
In the cotton rats immunized with MVAIK/RSV/F, NT antibodies against RSV subgroups A and B increased after reimmunization by two fold, but not significantly. As for the rats immunized with MVAIK/RSV/G, NT antibodies against RSV subgroup A were boosted from an undetectable level before the reimmunization to 101.8 ± 0.1, but those against RSV subgroup B were not detected.
3.4. Protection against RSV challenge
The peak response against RSV was observed five weeks after immunization. Three cotton rats were immunized with MVAIK/RSV/F and MVAIK/RSV/G and challenged with the homologous RSV subgroup A (Long strain) and heterologous subgroup B (wild-type). No infectious virus was recovered from nasal wash and BAL but RSV genome was detected. RSN genome copy number was slightly lower in immunized groups but not significant (data not shown). The recovery of infectious virus and genome copy numbers from lung tissues are shown in Fig. 8 . 105.4 and 104.5 PFU of infectious virus were recovered from 20 mg of lung tissue in two cotton rats of the control group challenged with RSV subgroup A, but no infectious virus was recovered in three cotton rats immunized with MVAIK/RSV/F. Meanwhile, 104.5, 102.8 and 103.3 PFU of infectious virus were recovered in cotton rats immunized with MVAIK/RSV/G.
Fig. 8.
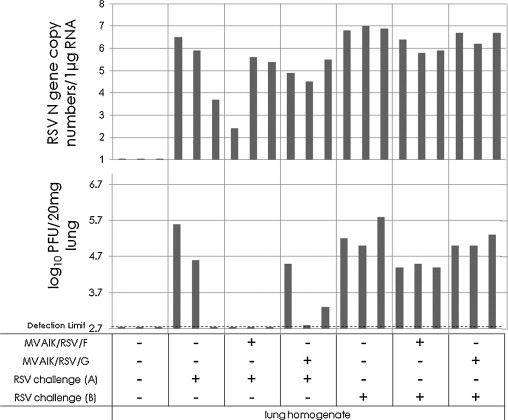
Recovery of RSV infectious virus and genome copy numbers after challenge with RSV subgroups A and B. Three cotton rats were investigated in the normal control group, non-immunized group, and group immunized with MVAIK/RSV/F or MVAIK/RSV/G. Animals non-immunized group, and group immunized with MVAIK/RSV/F or MVAIK/RSV/G were challenged with 1.0 × 106 PFU of the homologous RSV Long strain and wild-type subgroup B five weeks later. Virus infectivity was monitored in lung homogenate, and RSV infectivity is shown as PFU in 20 mg of lung tissue. And 1 μg of total RNA of lung tissue was used for real-time PCR, and each column represents individual result.
As for challenge with RSV subgroup B, 105.0–5.8 PFU of RSV was recovered from lung infected with RSV subgroup B in non-immunized rats. In cotton rats immunized with MVAIK/RSV/F, virus titers were slightly lower, 104.4–4.5 PFU but 105.0–5.3 PFU from their lung tissues in the cotton rats immunized with MVAIK/RSV/G. There was no significant reduction in RSV N gene copy number.
For histopathological examinations, lung tissues were obtained four days after the challenge with RSV subgroups A and B and the results of HE staining and immuno-staining against RSV antigens are shown in Fig. 9 . The non-immunized rat challenged with RSV subgroup A showed prominent interstitial pneumonia (panel 2; thickening of alveolar wall, and infiltration of inflammatory mononuclear cells) with RSV antigens in bronchial epithelial cells (panel 6). In cotton rat immunized with MVAIK/RSV/F showed very mild inflammation (panel 3), though most sections were normal, without RSV antigen in bronchial tissue after RSV challenge with subgroup A (panel 7). In cotton rat immunized with MVAIK/RSV/G, moderate interstitial pneumonia was observed with a small amount of RSV antigen (panels 4 and 8).
Fig. 9.
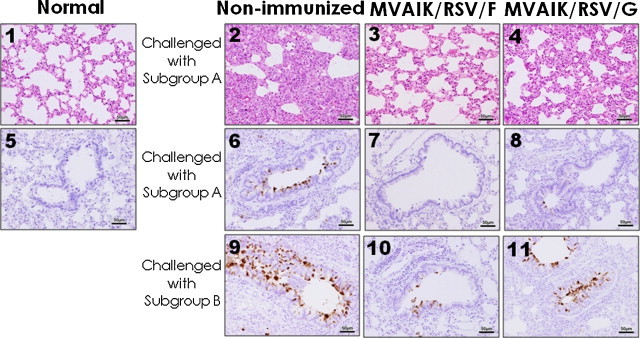
Pulmonary histopathology in cotton rats challenged with RSV subgroups A and B. Cotton rats were immunized intramuscularly with MVAIK/RSV/F (panels 3, 7, and 10) or MVAIK/RSV/G (panels 4, 8, and 11) and then challenged five weeks later, with RSV subgroup A (panels 2, 3, 4, 6, 7, and 8) and subgroups B (panels 9, 10 and 11). They were sacrificed four days after the challenge. Histological examination was performed by HE staining of lung tissues (panels 1, 2, 3, and 4) and the results of immuno-staining of bronchiolar regions are shown in panels 5, 6, 7, 8, 9, 10, and 11. Immuno-staining was performed using four clone blend monoclonal antibodies against RSV P, F, and N proteins and anti-mouse IgG conjugated with HRP. HE staining and immuno-staining of normal control are shown in panels 1 and 5.
As for the challenge with RSV subgroup B, histological findings in non-immunized rat challenged with subgroup B were similar to the results challenged with RSV subgroup A. The results of immuno-staining are shown. Large amounts of RSV antigen were detected in non-immunized rat (panel 9). Small amounts of RSV antigens were detected in MVAIK/RSV/F group (panel 10) in comparison with MVAIK/RSV/G group (panel 11). Finding of RSV antigens were well correlated with the results of the recovery of infectious virus from lung tissues. Inoculated virus would be cleared and demonstrated a mild pathological finding in rats immunized with MVAIK/RSV/F.
4. Discussion
RSV is a clinically important cause of respiratory tract infections, especially among high-risk infants, immunocompromised hosts, and the elderly. Despite a serious disease burden, there is no licensed vaccine for RSV. Initial efforts to develop a vaccine involved FI-RSV which unexpectedly enhanced the disease in clinical trials in RSV-naive children [33]. FI-RSV failed to induce a secretory IgA response after pareteral administration without inducing a CTL response, which was a serious drawback of the inactivated vaccine. The defeated F protein would not induce Th1 response and the aluminium-precipitated vaccine induced only Th2 response. The allergic reaction to this vaccine would be caused by the Th2-prone reaction [34]. Several subunit vaccines were investigated, but failed to generate effective antibodies. A live attenuated vaccine has the advantage of inducing humoral and cellular immune responses similar to a natural infection. Temperature-sensitive (ts) and cold-adapted (ca) RSV vaccine strains have been developed by conventional attenuation methods. Over the last 40 years, cautious and deliberate progress has been made toward developing a RSV vaccine using various experimental approaches, including live attenuated strains and vector-based and viral protein subunit vaccine candidates. But the balance between the safety and immunogenicity is a key issue to the development of a live attenuated vaccine, and the (ts) RSV vaccine candidate resulted in insufficient attenuation, causing similar respiratory illness [35]. Based on a vaccine candidate having the ts phenotype, several recombinant vaccine candidates were developed by deletion of the SH gene or NS1 gene or mutation by reverse genetics. These recombinant RSV vaccines induced sufficient immune response in chimpanzees [36]. Another approach involved the application of reverse genetics to express RSV protein in a recombinant vector-based vaccine. The first vector-based candidate was evaluated using vaccinia virus. Recombinants expressing RSV F or G was highly immunogenic, induced protection in mouse but provided inconsistent protection in chimpanzees [37]. MVA strain of vaccinia-based recombinants expressing RSV G and F protein were immunogenic in rodent but not in rhesus monkey model [38]. Several vector-based live vaccine platforms were established using HPIV-III and Sendai virus [23], [39]. Through preceding experiments, the F protein is known to be more effective than G. But there were no experiences for clinical usage and the HPIV-III-based recombinant vaccine was poorly immunogenic in human clinical trials.
In this report, reverse genetics using the AIK-C live attenuated measles vaccines were developed. A recombinant measles virus vector-based vaccine was established using the Schwartz strain, expressing the West Nile virus [40]. As well as the Schwartz strain, the AIK-C measles vaccine is a further attenuated vaccine strain having the ts phenotype, and its safety and immunogenicity has been confirmed [26], [29]. Thus, in this report, AIK-C was used for a live virus vaccine-vector. Expression of the RSV G or F protein was confirmed by indirect immuno-staining of B95a cells infected with MVAIK/RSV/G or F with polyclonal and monoclonal antibodies against the F protein. By Western blotting, the G or F protein was detected in culture medium and cell lysate of B95a cells infected with MVAIK/RSV/G or F. The RSV G and F proteins were considered not to be incorporated into MV particles because theoretically they had no binding site for the MV M protein. MV envelop proteins bound the M protein [41]. The genetic stability of the vaccine candidate was examined and inserted genes for RSV G and F were stable even after 15 passages.
The recombinant measles virus (MVAIK) triggered an immune response three weeks after vaccination in cotton rats. Levels of these antibodies were maintained for 16 weeks. The same was observed after immunization with MVAIK/RSV/G or F. To investigate the viral growth, samples of nasal turbinate, lung, thymus, spleen, liver, kidney, and bone marrow were obtained 10 days after immunization, but no infectious virus was recovered. Total RNA was extracted and RT-real time PCR was performed to detect the measles N gene by real-time PCR. The MV genome was detected only in thymus in cotton rats immunized with MVAIK, MVAIK/RSV/G, and MVAIK/RSV/F (data not shown). Infectious virus was recovered from inguinal superficial lymph nodes three days after infection in the previous study [42]. NT antibody titers against RSV were investigated, using RSV Long (subgroup A) and wild-type subgroup B. MVAIK/RSV/G or MVAIK/RSV/F induced the production of NT antibodies against RSV subgroup A from one week after vaccination in cotton rats. Antibody titers were higher after immunization with MVAIK/RSV/F than with MVAIK/RSV/G. RSV has distinctly different subgroups, A and B. The G or F gene of subgroup A was used in this study. Therefore, the cross reaction of NT against subgroup B was investigated. MVAIK/RSV/G did not generate NT antibodies against RSV subgroup B, but MVAIK/RSV/F induced production of cross-reactive NT antibodies against RSV subgroups A and B. The predicted amino acid sequence of the RSV F protein used in this study exhibited 98.6% homology among F proteins of subgroup A strains and 90.8% in comparison with those of subgroup B strains. The predicted amino acid sequence of RSV G protein has 86.9% homology among subgroup A strains but 49.7% in comparison with subgroup B. Thus, F protein was relatively conserved between subgroups A and B but the G protein of RSV was variable and thought not suitable as a vaccine antigen. Recently, a humanized monoclonal antibody against the RSV F protein was used for prevention of serious RSV infections in young infants having cardiac and pulmonary disorders, with a low birth weight, or born prematurely. In this study, recombinant MVAIK/RSV/G or F was administered intramuscularly and induced sufficient NT antibodies. Secretory IgA antibodies and CTL response were not examined but it protected against the challenge with homologous RSV subgroup A. In non-immunized cotton rats, 105.4 and 104.5 PFU of infectious virus were recovered from 20 mg of lung tissue four days after the RSV challenge. But those immunized with MVAIK/RSV/F were protected, without recovery of infectious virus from the lung tissues. And they did not demonstrate interstitial pneumonia. Cross reactive NT antibodies were demonstrated after immunization with MVAIK/RSV/F but its protective effect is not sufficient against subgroup B, demonstrating slightly lower levels (approximately 1/10 of non-immunized control) of the recovery of infectious virus. Protective effects of MVAIK/RSV/G were poor in comparison with MVAIK/RSV/F similar to the serological responses.
As for the experimental animal models, transgenic mice expressing human CD46 with the knock out of type I interferon (IFN) receptor gene were used to evaluate the immunogenicity of a recombinant MV vaccine candidate produced using the West Nile virus [40], SARS corona virus [43], hepatitis B virus [44] and HIV [45]. Efficient immune responses were reported, but the IFN system is the most important signal for innate immunity. In the case of the RSV vaccine candidate, innate immunity modified the adaptive immunity, and, therefore, cotton rats without gene manipulation were used in these experiments [46].
Recombinant MV vaccine-based vectors have practical limitation for timing of immunization. In young infants, maternal conferred immunity would interfere with vaccine effects. In field trials, AIK-C gave efficient sero-conversion and induction of cell-mediated immunity even when the vaccine was given at the age of six months [26], [27]. They demonstrated more than 80% sero-conversion rate to overcome maternal conferred immunity and the safety was similarly confirmed, suggesting no evidence of immune-suppression. RSV infection was observed even after six months of age, and, therefore, MVAIK/RSV/F would be applicable for six months to provide protective immunity both against RSV and measles especially for developing countries.
As for the effective protection against RSV infection, intranasal administration is desired. But we have no experience of intranasal administration of AIK-C vaccine, and, in our previous experiments, the recombinant MVAIK did not induce serum NT against MV through intranasal administration because of the strict ts phenotype in cotton rat model, having high body temperature [21], [42]. Therefore, the comparative studies are planning to investigate the immunogenicity and challenge tests in monkeys immunized with MVAIK/RSV/F.
In conclusion, a new MV vaccine-strain-based RSV vaccine candidate was demonstrated to confer protection against RSV in cotton rats. The xenogeneic recombinant might induce simultaneously protective immunity against backbone-MV and inserted-RSV infections. Recombinant MVAIK expressing RSV F protein is a promising candidate and protective effects should be confirmed in monkey model, considering the immunization routes.
Acknowledgements
We would like to thank Dr. H. Ueki, Dr. M. Ikeda and S. Maruyama of the Kitasato Institute, Pathology Laboratory, Research & Development Division, Research Center for Biologicals, for investigating histopathological examinations, and M. Maeda for her help in cotton rat experiments.
References
- 1.Lamb R.A., Parks G.D. Paramyxoviridae: the viruses and their replication. In: Knipe D.M., Howley P.M., editors. Fields virology. 5th ed. Lippincott Williams & Wilkins; Philadelphia: 2007. pp. 1449–1496. [Google Scholar]
- 2.Collins P.L., Hill M.G., Camargo E., Grosfeld H., Chanock R.M., Murphy B.R. Production of infectious human respiratory syncytial virus from cloned cDNA confirms an essential role for the transcription elongation factor from the 5′ proximal open reading frame of the M2 mRNA in gene expression and provides a capability for vaccine development. Proc Natl Acad Sci USA. 1995;92(25):11563–11567. doi: 10.1073/pnas.92.25.11563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chanock R., Finberg L. Recovery from infants with respiratory illness of a virus related to chimpanzee coryza agent (CCA). II. Epidemiologic aspects of infection in infants and young children. Am J Hyg. 1957;66(3):291–300. doi: 10.1093/oxfordjournals.aje.a119902. [DOI] [PubMed] [Google Scholar]
- 4.Anderson L.J., Hierholzer J.C., Tsou C., Hendry R.M., Fernie B.F., Stone Y. Antigenic characterization of respiratory syncytial virus strains with monoclonal antibodies. J Infect Dis. 1985;151(4):626–633. doi: 10.1093/infdis/151.4.626. [DOI] [PubMed] [Google Scholar]
- 5.La Via W.V., Grant S.W., Stutman H.R., Marks M.I. Clinical profile of pediatric patients hospitalized with respiratory syncytial virus infection. Clin Pediatr (Phila) 1993;32(8):450–454. doi: 10.1177/000992289303200801. [DOI] [PubMed] [Google Scholar]
- 6.Henderson F.W., Collier A.M., Clyde W.A., Jr., Denny F.W. Respiratory-syncytial-virus infections, reinfections and immunity. A prospective, longitudinal study in young children. N Engl J Med. 1979;300(10):530–534. doi: 10.1056/NEJM197903083001004. [DOI] [PubMed] [Google Scholar]
- 7.Ebbert J.O., Limper A.H. Respiratory syncytial virus pneumonitis in immunocompromised adults: clinical features and outcome. Respiration. 2005;72(3):263–269. doi: 10.1159/000085367. [DOI] [PubMed] [Google Scholar]
- 8.Anderson L.J., Parker R.A., Strikas R.L. Association between respiratory syncytial virus outbreaks and lower respiratory tract deaths of infants and young children. J Infect Dis. 1990;161(4):640–646. doi: 10.1093/infdis/161.4.640. [DOI] [PubMed] [Google Scholar]
- 9.González P.A., Bueno S.M., Riedel C.A., Kalergis A.M. Impairment of T cell immunity by the respiratory syncytial virus: targeting virulence mechanisms for therapy and prophylaxis. Curr Med Chem. 2009;16(34):4609–4625. doi: 10.2174/092986709789760724. [DOI] [PubMed] [Google Scholar]
- 10.Becker Y. Respiratory syncytial virus (RSV) evades the human adaptive immune system by skewing the Th1/Th2 cytokine balance toward increased levels of Th2 cytokines and IgE, markers of allergy—a review. Virus Genes. 2006;33(2):235–252. doi: 10.1007/s11262-006-0064-x. [DOI] [PubMed] [Google Scholar]
- 11.Fisher R.G., Crowe J.E., Jr., Johnson T.R., Tang Y.W., Graham B.S. Passive IgA monoclonal antibody is no more effective than IgG at protecting mice from mucosal challenge with respiratory syncytial virus. J Infect Dis. 1999;180(4):1324–1327. doi: 10.1086/315037. [DOI] [PubMed] [Google Scholar]
- 12.Piedra P.A., Cron S.G., Jewell A., Hamblett N., McBride R., Palacio M.A. Immunogenicity of a new purified fusion protein vaccine to respiratory syncytial virus: a multi-center trial in children with cystic fibrosis. Vaccine. 2003;21(19–20):2448–2460. doi: 10.1016/s0264-410x(03)00098-7. [DOI] [PubMed] [Google Scholar]
- 13.Lofland J.H., O’Connor J.P., Chatterton M.L., Moxey E.D., Paddock L.E., Nash D.B. Palivizumab for respiratory syncytial virus prophylaxis in high-risk infants: a cost-effectiveness analysis. Clin Ther. 2000;22(11):1357–1369. doi: 10.1016/s0149-2918(00)83032-5. [DOI] [PubMed] [Google Scholar]
- 14.Murphy B.R., Prince G.A., Walsh E.E., Kim H.W., Parrott R.H., Hemming V.G. Dissociation between serum neutralizing and glycoprotein antibody responses of infants and children who received inactivated respiratory syncytial virus vaccine. J Clin Microbiol. 1986;24(2):197–202. doi: 10.1128/jcm.24.2.197-202.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Murphy B.R., Walsh E.E. Formalin-inactivated respiratory syncytial virus vaccine induces antibodies to the fusion glycoprotein that are deficient in fusion-inhibiting activity. J Clin Microbiol. 1988;26(8):1595–1597. doi: 10.1128/jcm.26.8.1595-1597.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wright P.F., Belshe R.B., Kim H.W., Van Voris L.P., Chanock R.M. Administration of a highly attenuated, live respiratory syncytial virus vaccine to adults and children. Infect Immun. 1982;37(1):397–400. doi: 10.1128/iai.37.1.397-400.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Whitehead S.S., Firestone C.Y., Karron R.A., Crowe J.E., Jr., Elkins W.R., Collins P.L. Addition of a missense mutation present in the L gene of respiratory syncytial virus (RSV) cpts530/1030 to RSV vaccine candidate cpts248/404 increases its attenuation and temperature sensitivity. J Virol. 1999;73(2):871–877. doi: 10.1128/jvi.73.2.871-877.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Collins P.L., Purcell R.H., London W.T., Lawrence L.A., Chanock R.M., Murphy B.R. Evaluation in chimpanzees of vaccinia virus recombinants that express the surface glycoproteins of human respiratory syncytial virus. Vaccine. 1990;8:164–168. doi: 10.1016/0264-410x(90)90141-8. [DOI] [PubMed] [Google Scholar]
- 19.Nakayama T., Komase K., Uzuka R., Hoshi A., Okafuji T. Leucine at position 278 of the AIK-C measles virus vaccine strain fusion protein is responsible for reduced syncytium formation. J Gen Virol. 2001;82:2143–2150. doi: 10.1099/0022-1317-82-9-2143. [DOI] [PubMed] [Google Scholar]
- 20.Ballart I., Eschle D., Cattaneo R., Schmid A., Metzler M., Chan J. Infectious measles virus from cloned cDNA. EMBO J. 1990;9(2):379–384. doi: 10.1002/j.1460-2075.1990.tb08121.x. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 21.Komase K., Nakayama T., Iijima M., Miki K., Kawanishi R., Uejima H. The phosphoprotein of attenuated measles AIK-C vaccine strain contributes to its temperature-sensitive phenotype. Vaccine. 2006;24(6):826–834. doi: 10.1016/j.vaccine.2005.06.036. [DOI] [PubMed] [Google Scholar]
- 22.Tang R.S., MacPhail M., Schickli J.H., Kaur J., Robinson C.L., Lawlor H.A. Parainfluenza virus type 3 expressing the native or soluble fusion (F) protein of respiratory syncytial virus (RSV) confers protection from RSV infection in African green monkeys. J Virol. 2004;78(20):11198–11207. doi: 10.1128/JVI.78.20.11198-11207.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhan X., Hurwitz J.L., Krishnamurthy S., Takimoto T., Boyd K., Scroggs R.A. Respiratory syncytial virus (RSV) fusion protein expressed by recombinant Sendai virus elicits B-cell and T-cell responses in cotton rats and confers protection against RSV subtypes A and B. Vaccine. 2007;25(52):8782–8793. doi: 10.1016/j.vaccine.2007.10.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Rima B.K., Earle J.A., Yeo R.P., Herlihy L., Baczko K., ter Meulen V. Temporal and geographical distribution of measles virus genotypes. J Gen Virol. 1995;76(Pt 5):1173–1180. doi: 10.1099/0022-1317-76-5-1173. [DOI] [PubMed] [Google Scholar]
- 25.Makino S., Sasaki K., Nakamura N., Nakagawa M., Nakajima S. Studies on the modification of the live AIK measles vaccine. II. Development and evaluation of the live AIK-C measles vaccine. Kitasato Arch Exp Med. 1974;47(1–2):13–21. [PubMed] [Google Scholar]
- 26.Bolotovski V.M., Grabowsky M., Clements C.J., Albrecht P., Brenner E.R., Zargaryantzs A.I. Immunization of 6 and 9 month old infants with AIK-C, Edmonston-Zagreb, Leningrad-16 and Schwarz strains of measles vaccine. Int J Epidemiol. 1994;23(5):1069–1077. doi: 10.1093/ije/23.5.1069. [DOI] [PubMed] [Google Scholar]
- 27.Pabst H.F., Spady D.W., Carson M.M., Krezolek M.P., Barreto L., Wittes R.C. Cell-mediated and antibody immune responses to AIK-C and Connaught monovalent measles vaccine given to 6 month old infants. Vaccine. 1999;17(15–16):1910–1918. doi: 10.1016/s0264-410x(98)00472-1. [DOI] [PubMed] [Google Scholar]
- 28.Cutts F.T., Grabowsky M., Markowitz L.E. The effect of dose and strain of live attenuated measles vaccines on serological responses in young infants. Biologicals. 1995;23(1):95–106. doi: 10.1016/1045-1056(95)90018-7. [DOI] [PubMed] [Google Scholar]
- 29.Nakayama T., Onoda K. Vaccine adverse events reported in post-marketing study of the Kitasato Institute from 1994 to 2004. Vaccine. 2007;25(3):570–576. doi: 10.1016/j.vaccine.2006.05.130. [DOI] [PubMed] [Google Scholar]
- 30.Kobune F., Sakata H., Sugiura A. Marmoset lymphoblastoid cells as a sensitive host for isolation of measles virus. J Virol. 1990;64(February (2)):700–705. doi: 10.1128/jvi.64.2.700-705.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kumada A., Komase K., Nakayama T. Recombinant measles AIK-C strain expressing current wild-type hemagglutinin protein. Vaccine. 2004;22(3–4):309–316. doi: 10.1016/j.vaccine.2003.08.015. [DOI] [PubMed] [Google Scholar]
- 32.Sutter G., Ohlmann M., Erfle V. Non-replicating vaccinia vector efficiently expresses bacteriophage T7 RNA polymerase. FEBS Lett. 1995;371(1):9–12. doi: 10.1016/0014-5793(95)00843-x. [DOI] [PubMed] [Google Scholar]
- 33.Kapikian A.Z., Mitchell R.H., Chanock R.M., Shvedoff R.A., Stewart C.E. An epidemiologic study of altered clinical reactivity to respiratory syncytial (RS) virus infection in children previously vaccinated with an inactivated RS virus vaccine. Am J Epidemiol. 1969;89(4):405–421. doi: 10.1093/oxfordjournals.aje.a120954. [DOI] [PubMed] [Google Scholar]
- 34.Polack F.P., Teng M.N., Collins P.L., Prince G.A., Exner M., Regele H. A role for immune complexes in enhanced respiratory syncytial virus disease. J Exp Med. 2002;196(6):859–865. doi: 10.1084/jem.20020781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Karron R.A., Wright P.F., Crowe J.E., Jr., Clements-Mann M.L., Thompson J., Makhene M. Evaluation of two live, cold-passaged, temperature-sensitive respiratory syncytial virus vaccines in chimpanzees and in human adults, infants, and children. J Infect Dis. 1997;176(6):1428–1436. doi: 10.1086/514138. [DOI] [PubMed] [Google Scholar]
- 36.Teng M.N., Whitehead S.S., Bermingham A., St Claire M., Elkins W.R., Murphy B.R. Recombinant respiratory syncytial virus that does not express the NS1 or M2-2 protein is highly attenuated and immunogenic in chimpanzees. J Virol. 2000;74(19):9317–9321. doi: 10.1128/jvi.74.19.9317-9321.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wyatt L.S., Whitehead S.S., Venanzi K.A., Murphy B.R., Moss B. Priming and boosting immunity to respiratory syncytial virus by recombinant replication-defective vaccinia virus MVA. Vaccine. 1999;18(5–6):392–397. doi: 10.1016/s0264-410x(99)00257-1. [DOI] [PubMed] [Google Scholar]
- 38.de Wall L., Wyatt L.S., Yuksel S., Van Amerongen G., Moss B., Niester H.G. Vaccination of infant macaques with a recombinant modified vaccinia virus Ankara expressing the respiratory syncytial virus F and G genes does not predispose for immunopathology. Vaccine. 2004;22:923–926. doi: 10.1016/j.vaccine.2003.10.010. [DOI] [PubMed] [Google Scholar]
- 39.Jones B., Zhan X., Mishin V., Slobod K.S., Surman S., Russell C.J. Human PIV-2 recombinant Sendai virus (rSeV) elicits durable immunity and combines with two additional rSeVs to protect against hPIV-1, hPIV-2, hPIV-3, and RSV. Vaccine. 2009;27(12):1848–1857. doi: 10.1016/j.vaccine.2009.01.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Desprès P., Combredet C., Frenkiel M.P., Lorin C., Brahic M., Tangy F. Live measles vaccine expressing the secreted form of the West Nile virus envelope glycoprotein protects against West Nile virus encephalitis. J Infect Dis. 2005;191(2):207–214. doi: 10.1086/426824. [DOI] [PubMed] [Google Scholar]
- 41.Wild T.F., Buckland R. Functional aspects of envelope-associated measles virus proteins. Curr Top Microbiol Immunol. 1995;191:51–64. doi: 10.1007/978-3-642-78621-1_4. [DOI] [PubMed] [Google Scholar]
- 42.Haga T., Murayama N., Shimizu Y., Saito A., Sakamoto T., Morita T. Analysis of antibody response by temperature-sensitive measles vaccine strain in the cotton rat model. Comp Immunol Microbiol Infect Dis. 2009;32(5):395–406. doi: 10.1016/j.cimid.2007.11.011. [DOI] [PubMed] [Google Scholar]
- 43.Liniger M., Zuniga A., Tamin A., Azzouz-Morin T.N., Knuchel M., Marty R.R. Induction of neutralising antibodies and cellular immune responses against SARS coronavirus by recombinant measles viruses. Vaccine. 2008;26(17):2164–2174. doi: 10.1016/j.vaccine.2008.01.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.del Valle J.R., Devaux P., Hodge G., Wegner N.J., McChesney M.B., Cattaneo R. A vectored measles virus induces hepatitis B surface antigen antibodies while protecting macaques against measles virus challenge. J Virol. 2007;81(19):10597–10605. doi: 10.1128/JVI.00923-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Liniger M., Zuniga A., Morin T.N., Combardiere B., Marty R., Wiegand M. Recombinant measles viruses expressing single or multiple antigens of human immunodeficiency virus (HIV-1) induce cellular and humoral immune responses. Vaccine. 2009;27(25–26):3299–3305. doi: 10.1016/j.vaccine.2009.01.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Boukhvalova M.S., Prince G.A., Blanco F.C.G. The cotton rat model of respiratory viral infections pathogenesis and immunity. Biologicals. 2009;37:152–159. doi: 10.1016/j.biologicals.2009.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]