

Abstract
The airway mucosal epithelium is the first site of virus contact with the host, and the main site of infection and inflammation. Nitric oxide (NO) produced by the airway epithelium is vital to antiviral inflammatory and immune defense in the lung. Multiple mechanisms function coordinately to support high-level basal NO synthesis in healthy airway epithelium and further induction of NO synthesis in the infected airway of normal hosts. Hosts deficient in NO synthesis, such as those patients with cystic fibrosis, have impaired antiviral defense and may benefit from therapies to augment NO levels in the airways.
Keywords: Antiviral host defense, Nitric oxide
Introduction
Respiratory virus infection remains a major cause of morbidity and mortality in human populations worldwide with a broad spectrum of clinical disease ranging from asymptomatic infection, rhinitis, to viral pneumonia. A variety of viruses are associated with respiratory tract infections including rhinovirus, coronavirus, human respiratory syncytial virus (HRSV), influenza A and B viruses, human parainfluenza viruses (HPIVs), and adenovirus [1], [2], [3]. Among them, rhinovirus infections are the predominant cause of the common cold and have been associated with exacerbations of asthma, chronic bronchitis, and sinusitis [4], [5], [6], [7]. HRSV is the most common cause of bronchiolitis and other respiratory infections in infants and young children [8], [9], [10]. Although the factors dictating virulence are complex, the interaction between inherent viral properties and host cellular response ultimately determines severity of virus disease and outcome [11], [12], [13].
In this context, the airway mucosal epithelium, a primary target of respiratory viruses, plays an important role in the innate host defense response to virus [14], [15]. A critical early component of the innate host defense in the airway is the ability of respiratory epithelial cells to produce high levels of nitric oxide (NO). NO functions as a signaling molecule in initiation of the inflammatory response to viruses, and also has direct antiviral effects [14], [16], [17], [18]. Here, we review the human airway production of NO and the consequence of NO deficiency on antiviral defense.
NO production in the lung
Nitric oxide synthases
NO is synthesized endogenously by NO synthases (NOS) (EC 1.14.13.39). Three isoforms of NOS, which are products of separate genes [19], [20], have been identified, including neuronal (nNOS or NOS I), endothelial (eNOS or NOS III), and iNOS or NOS II [19], [21], [22]. These enzymes convert L-arginine to NO and L-citrulline in a reaction that requires oxygen, NADPH, and cofactors FAD, FMN, tetrahydrobiopterin, calmodulin, and iron protoporphyrin IX (Fig. 1 ). NOS I and III, originally identified in neuronal and endothelial cells, respectively, depend on increases in calcium to bind calmodulin which results in enzyme activation and picomolar levels of NO production. NOS II is regulated at the level of transcription, mRNA stability, and translation, and is expressed after exposure of cells to specific cytokines and/or endotoxin [19], [23], [24], [25]. NOS II is calcium independent, binding calmodulin even at low calcium concentrations intracellularly, and produces nanomolar levels of NO [19], [26]. NO is relatively unstable and rapidly oxidized in simple buffers to the metabolic end-products nitrite (NO2 −) and nitrate (NO3 −), which can be used as indirect markers to monitor NO formation [19] (Fig. 2 ).
Fig. 1.
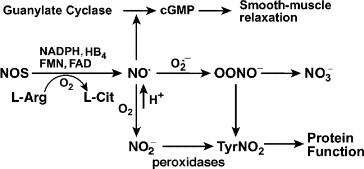
NO metabolism and function. NOS, nitric oxide synthases; NO•, nitric oxide; NO2−, nitrite; NO3−, nitrate; l-Arg, l-arginine; l-Cit, l-citrulline; HB4, tetrahydrobiopterin; cGMP, guanosine 3′,5′-cyclic monophosphate; O2•−, superoxide; OONO−, peroxynitrite; TyrNO2, nitrotyrosine.
Fig. 2.
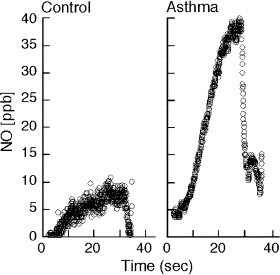
Accumulation of NO in the lower airway during breath hold of healthy control individual shows that NO is formed in the airway. Levels of accumulation are much greater in an asthmatic individual with airway inflammation [26].
NO in the human airway
NO is produced within the human lung, as evidenced by NO detectable in the exhaled breath, and in the lower airway gases of humans (Fig. 2). Furthermore, reaction products of NO (e.g., NO2 −, NO3 −) are detectable at micromolar levels in the bronchoalveolar lavage fluid from human lungs [21], [27]. Immunohistochemical studies reveal the presence of the three isoforms of NOS in the airway. NOS III is primarily localized to pulmonary endothelial cells, and NOS I in nonadrenergic, noncholinergic inhibitory neurons [21], [28]. NOS II is continuously expressed in normal human airway epithelium under basal airway conditions [24], [29], [30], [31]. NO is also produced by the upper respiratory tract epithelium within the nasopharynx and paranasal sinuses, most likely by NOS II [32]. There is evidence that epithelial NOS II activity is a major determinant of NO present in exhaled breath [33].
Traditionally termed inducible NOS, NOS II is constitutively expressed only in select tissues like airway epithelium and is more typically synthesized in response to inflammatory or proinflammatory mediators [31]. Although NOS II expression is relatively abundant, at approximately 5–10 mRNA copies per airway epithelial cell, inflammation related to increased endogenous or exogenous reactive oxygen species (ROS), as in asthma or hyperoxia, leads to marked upregulation of the gene expression in human airways. Basal expression of NOS II is very likely important in the innate mucosal host defense of the airway, with further induction related to host response to pathogen(s) or inflammation. The role of NOS II in host defense of the airway is confirmed by mice genetically deficient in NOS II [34], [35]. On the other hand, increased NOS II expression in response to virus infection may contribute to airway injury [36].
Once produced, NO is freely diffusible and enters target cells. Vasodilatory effects are mediated by signal transduction primarily through activation of soluble guanylate cyclase to produce guanosine 3′,5′-cyclic monophosphate (cGMP) [31]. NO and/or its reaction products also mediate effects through protein modifications, particularly nitration of tyrosine and nitrosylation of thiols in proteins [37], [38], [39]. NO is recognized to play key roles in virtually all aspects of lung biology, and is clearly involved in antimicrobial host defense [20], [21], [23]. Hence, reduction of basal NO levels in the lung is associated with several lung diseases characterized by increased susceptibility to respiratory infections, e.g., cystic fibrosis (CF) (Table 1 ). On the other hand, an excessively high level of NO production is also implicated in the pathophysiology of lung diseases, such as asthma (Fig. 2, Table 1) [29], [40], [41].
Table 1.
Alterations of exhaled NO in lung diseases
| Increased exhaled NO | Decreased exhaled NO |
|---|---|
| Asthma | Pulmonary arterial hypertension |
| Bronchiectasis | Cystic fibrosis |
| Lymphangioleiomyomatosis | α1-antitrypsin deficiency |
| Viral respiratory tract infection | Primary ciliary dyskinesia |
| Active tuberculosis | Acute respiratory distress syndrome |
| Allergen challenge (late response) | Cigarette smoke |
| Hyperoxia | Passive smoking |
| HIV-1 infection |
NO and viral infections
Alterations in exhaled breath concentrations of NO have been described for many pulmonary diseases [21], [42], [43], [44], [45], [46], [47], [48], [49], [50], [51], [52] (Table 1). Exhaled NO is increased in chronic inflammatory airway diseases, such as asthma and bronchiectasis, and in upper respiratory tract infections, including bacterial and viral respiratory illnesses [21], [43], [44], [45]. Epithelial cells at mucosal surfaces have a variety of inflammatory and immune defense mechanisms to deal with viruses, including the induction of NOS II and increased production of NO [14], [15], [16], [17], [18], [29]. For example, experimental influenza virus or rhinovirus 16 infection increase exhaled NO in volunteers [53], [54]. Furthermore, HRSV infection, Hong Kong influenza A (H3N2) virus, or Adenovirus infection in mice in vivo or in primary human airway epithelial cells in vitro increases levels of NOS II mRNA, protein, and activity in respiratory epithelium and increases production of NO (Fig. 3 ) [11], [14], [17], [55]. In contrast, exhaled NO is decreased in subjects infected with the Human immunodeficiency virus (HIV)-1, suggesting that viral mechanisms exist to suppress NO-related host defense [56].
Fig. 3.
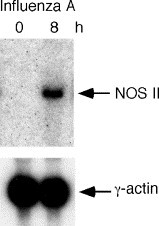
NOS II induction by influenza A. Northern analysis for NOS II was performed using total RNA from A549 cells (20 μg RNA/lane) uninfected or infected with influenza A virus [2 × 105.25 50% of tissue infectious dose (TCID50)/ml for 8 h]. γ-Actin is shown as a control [11].
Antiviral effects of NO
NOS II is sufficient for antiviral defense in human airway epithelial cells [16], [34], [35], [57], [58], [59], [60], [61]. Replication of a wide range of DNA and RNA viruses are inhibited in vitro by the addition of chemical donors of NO, or by the overexpression or induction of NOS II. NO inhibits both virus replication and latency of viruses, including coxsackievirus, influenza A and B, rhinovirus, cytomegalovirus, vaccinia virus, ectromelia virus, human herpesvirus-1, and HPIV3 [18], [34], [57], [58], [59], [61], [62], [63], [64], [65]. Likewise, loss of NO synthesis leads to increase of virus titers with infection, and significantly more severe clinical outcomes [34], [35], [60], [66], [67]. For example, mice genetically deficient in NOS II have significantly higher mortality, and decreased virus clearance, with cytomegalovirus (CMV) [35] or coxsackievirus B4 [34] as compared to wild-type. Similarly, in mice treated with a specific inhibitor of NOS II, CMV replication is moderately enhanced as evidenced by 5-fold increase in viral titers, and this results in viral persistence and latency [60].
High-level NO synthesis results in a large variety of reactive products which inhibits viral protein and RNA synthesis, and viral replication by modifying target molecules essential for replication [68], [69]. Two specific virus targets of NO, ribonucleotide reductase, the rate-limiting enzyme in DNA synthesis, and viral protease, have been identified on the basis of in vitro exposure of viral protein to NO donors in cell-free systems [62], [64], [70]. However, targets of NO modification also include host cell proteins, particularly thiol groups and tyrosine [37], [38], [39]. NO may also bind to heme iron in proteins [71]. In lung epithelial cells exposed to inflammatory cytokines to induce NOS II, over 40 cellular proteins are modified by tyrosine nitration, with consequences on activity and function [37]. In this context, NO production can be beneficial as an antiviral effector mechanism against viruses [61], [62], [72], [73], but also may be detrimental by contributing to inflammation and injury of the respiratory tract through formation of toxic reactive nitrogen intermediates and cellular protein modifications [72], [73], [74]. For example, in murine models of virus-induced pneumonia, NOS II activity and mRNA expression are greatly increased within lung tissue [72], [73], [74], but inhibition of NO production improves the histopathologic changes and the survival of virus-infected mice without exerting an effect on the pulmonary viral titer. Therefore, in some cases, pulmonary injury may be attenuated by administration of NO inhibitors without significant effect on viral clearance [72], [73], [74].
Abnormalities of NO and virus infection in cystic fibrosis
Cystic fibrosis (CF) is a hereditary disease characterized by dysfunction of the exocrine glands, but respiratory disease is the major cause of morbidity and mortality. Chronic endobronchial infection and inflammation ultimately result in lung destruction and death [75]. Bacterial colonization of CF lung is usually established in the first decade of life and, once established, is difficult to eradicate [76]. Little is known about the natural history of bacterial respiratory tract infection and factors associated with initial colonization in CF lung, but studies show a clear relationship between acute viral respiratory tract infection, including HRSV, HPIV, influenza virus, and adenovirus, and pulmonary exacerbation and disease progression in children with CF [77], [78], [79], [80], [81]. In fact, CF children have increased susceptibility to virus infections as compared to non-CF siblings. In one study of CF children with early diagnosis at neonatal screening, hospitalizations in the first year of life were due to respiratory virus infection in over 50% of cases [82]. Surprisingly, despite the inflammatory nature and chronic infections of the lung in CF, NO levels in exhaled breath are lower while nitrite in bronchoalveolar lavage fluid (BALF) is higher from CF patients as compared to healthy controls or individuals with other inflammatory lung diseases [83], [84], [85]. Notably, infants with CF before the onset of respiratory symptoms have mean levels of exhaled NO threefold lower than healthy control infants, suggesting that the lower levels of NO in CF are not the result but, rather, a precursor of pulmonary disease [86]. Recent studies support that loss of NO synthesis is a primary pathophysiologic event in the susceptibility of CF patients to virus and other microbial pathogens, through defects in signal transduction to NOS II [16], [87], [88], [89].
Interacting determinants in NO–antiviral host defense
Other components and/or pathways are also involved directly or indirectly in the antiviral host defense mechanisms, in part through interactions with NO. ROS, including superoxide (O2 -) and hydrogen peroxide (H2O2), are critical in antimicrobial defense but also contribute to human lung inflammation and injury [90], [91], [92]. Increased generation of ROS is intrinsic to viral infections and can be caused by direct effects of virus on cells and/or inflammatory responses of the host [90], [91], [93], [94]. NO and ROS, particularly O2 −, rapidly form the reactive nitrogen oxide, peroxynitrite (ONOO−), that may cause oxidation and nitration of amino acid residues of proteins or guanine of DNA, lipid peroxidation, and DNA cleavage, all of which can cause cellular dysfunction and injury leading to cell death [91], [93], [95]. Myeloperoxidase (MPO), eosinophil peroxidase (EPO), and lactoperoxidase (LPO) are members of the mammalian peroxidase superfamily and play specific and complementary roles in host defense through oxidative pathways [90], [92], [95], [96], [97], [98], [99]. MPO is highly expressed in neutrophils, monocytes, and certain subpopulations of tissue macrophages [100], and LPO is present in human airway epithelium [101]. Virus-infected cells produce peroxide that drives the virucidal peroxidase–H2O2–halide system to catalyze the H2O2-dependent peroxidation of halides and pseudohalides to produce antimicrobial hypohalous acids. Hypochlorous acids (HOCl) play an important role in killing microorganisms including virus but also injure normal tissues [90], [96], [97], [98], [99]. Recent studies show that members of the mammalian peroxidase superfamily can reduce NO levels [101], [102], [103], [104], [105]. NO consumption may be a direct consequence of peroxidase reaction since NO is a peroxidase substrate, or occur through NO reactions with peroxidase-generated free radical species (Fig. 1) [101], [102], [103], [104], [105], [106].
Reduced glutathione (GSH), an important water-soluble antioxidant, is present at varying levels in the airway lining fluid [90], [107], [108]. Systemic deficiency of extracellular GSH is present in CF patients, and related to mutation of the CFTR protein, as CFTR is a channel permeant to chloride anions, and other larger organic anions, including reduced GSH [108], [109], [110]. GSH deficiency may decrease overall NO production, through decreasing NOS II expression or by the parallel loss of S-nitrosoglutathione (GSNO), an important storage form and source of NO [107], [108].
Regulation of NO synthesis
NO biosynthesis is regulated at multiple levels in cells, i.e., NOS gene transcription, mRNA processing, protein expression and dimerization, and enzyme reaction kinetics [19]. All NOSs convert L-arginine to NO and L-citrulline in a reaction that requires oxygen, NADPH, and cofactors FAD, FMN, tetrahydrobiopterin, calmodulin, and iron protoporphyrin IX. Thus, pathways that alter substrate L-arginine availability or cofactor levels are key mechanisms for regulation of NO synthesis [27], [49], [111]. Further, the concentration of NO in any biologic system is the sum of its rate of enzymatic formation and consumption by other biological molecules, such as in reactions with ROS. Nevertheless, NOS II is largely subject to predominantly transcriptional regulation [19], [25], [26], [112], [113], [114]. Induction of NOS II mRNA expression varies in different human cell types, but typically is increased by cytokines [22], [25], [113], [114]. IFNγ is necessary for induction of NOS II in human airway epithelial cells [114], [115], [116], [117], [118], [119], [120] (Fig. 4 ). NOS II expressed constitutively by airway epithelium is identical to the NOS II that is induced by cytokines or endotoxin [29]. The V max of NOS II assures a high level of NO synthesis and is 10-fold greater than the traditional constitutive NOSs, such as NOS III [121].
Fig. 4.
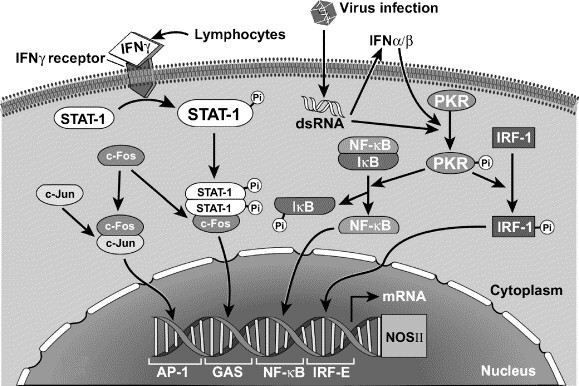
Mechanisms of viral-induced NOS II expression in human airway epithelial cells. STAT, signal transducer and activator of transcription; dsRNA, double-stranded RNA; PKR, dsRNA-activated protein kinase; NF-κB, nuclear factor kappa B; IRF-1, interferon regulatory factor 1; AP-1, activator protein-1; GAS, γ-activated site; IRF-E, interferon regulatory factor element.
The basal ability to produce high NO levels indicates that airway epithelial cells are primary effector cells of the host defense system. Healthy human airway epithelium has abundant expression of NOS II due to continuous transcriptional activation of the gene in vivo. Basal NOS II expression in airway epithelium is linked to STAT1 activation, and is related to physiologic levels of IFNγ in vivo. Removal of airway epithelial cells from the airway milieu results in rapid loss of NOS II transcription within a few hours of placing cells into culture. IFNγ induces NOS II gene expression in cells in vitro, with gene response in a dose-dependent manner from a range of 10 to 10,000 U IFNγ/ml, which encompasses healthy physiologic to pathologic levels of IFNγ found during virus infection [25], [26], [74]. Overall, constitutive NOS II expression in the airway is related to airway factors, including physiologic levels of IFNγ with STAT1 activation, and is important for mucosal host defense under basal conditions. Virus infection may further induce NOS II gene expression, resulting in more NOS II protein and NO synthesis that may function to inhibit virus proliferation, but also may contribute to the oxidative injury of the airway.
Analysis of the 5′-flanking region of the human NOSII shows that cooperative DNA binding of multiple signal transduction proteins is involved in activation of the NOS II promoter. A region of human NOS II 4.9 kb upstream of the transcription start site (GenBank Accession No. AF017634) [122] contains several sites that serve as composite binding elements, i.e., region encompassing the γ-activated site (GAS) and binding sites for activator protein (AP)-1 and nuclear factor kappa B (NF-κB). Prior work has emphasized the importance of AP-1 binding sites in activation of NOS II promoter in response to cytokine combinations [113], [119]. AP-1 is a complex composed of proteins of the Fos (c-Fos, FosB, Fra-1 and Fra-2) and Jun (c-Jun, JunB, and JunD) proto-oncogene families [123], [124], [125]. In general, Fos and Jun family proteins function as dimeric transcription factors that bind to AP-1 regulatory elements in the promoter and enhancer regions of genes [123], [125]. Other studies show that NF-κB activation and binding to κB DNA elements in the 5′-flanking region of the NOS II gene play a role in the cytokine induction of NOS II in A549 lung epithelial cell line in vitro [114]. We have shown that NOS II gene expression requires a physical interaction between c-Fos and signal transducer and activator of transcription (STAT)-1 to participate in cooperative transcriptional activation of the gene [122].
Viral mechanisms of airway inflammation and NO induction
IFN-γ, produced by lymphocytes in the airway mucosa within 24 to 36 h following viral infection, is almost certainly involved in NOS II induction in the course of viral infection through the activation of STAT-1. However, NOS II induction by virus early in infection is mediated by proteins responsive to the viral replicative intermediate, double-stranded RNA (dsRNA) [11]. Most viruses induce the synthesis of dsRNA at some time during their replication cycle [126]. For single-stranded RNA (ssRNA) viruses, dsRNA is the replicative intermediate in infected cells. For DNA viruses, dsRNA accumulates as the result of overlapping convergent transcription. Intracellular dsRNA formed during viral replication [126] binds to and activates a serine/threonine kinase, dsRNA-activated protein kinase (PKR), which activates signal transduction that leads to the activation of NOS II [11], [16], [17], [126], [127], [128], [129], [130].
PKR is critical for host antiviral mechanisms and for the activation of NOS II, as assessed by experiments in murine embryo fibroblast cells derived from mice genetically deficient in PKR [11], [129]. PKR exhibits two distinct kinase activities upon activation by dsRNA, autophosphorylation/activation and phosphorylation of substrates [131], which regulate transcriptional events by phosphorylation of proteins related to signal transduction pathways. For example, NF-κB activation by dsRNA in human airway epithelial cells is due to PKR activation and phosphorylation of the inhibitor of NF-κB (IκB). PKR also leads to transcriptional and posttranslational effects on interferon regulatory factor 1 (IRF-1), which may subsequently activate NOS II gene expression [132], [133], [134], [135]. IRF-1 protein is induced and activated by dsRNA in human airway epithelial cells [11]. Thus, PKR is essential in the signaling pathway for dsRNA induction of NOS II. Intriguingly, PKR is also essential for endotoxin-induction of NOS II in murine cells, confirming a central role for PKR in a generalized microbial-response signaling pathway to NOS II [11].
Defects in signal transduction to NOS II in CF
Unlike normal airway epithelium, NOS II expression under basal conditions of the CF airway epithelium is absent in vivo. Further, NOS II expression is not inducible in CF airway cells in vitro, despite multiple stimuli including HPIV3, influenza virus, dsRNA mimic (poly(IC)) or combinations of cytokines (IFNγ, IL-1β, and TNFα) and endotoxin [16], [89]. Loss of expression is linked to impaired activation of STAT-1 [16], [89]. PKR, IRF-1, and NF-κB activation induced by poly(IC) are similar in CF and healthy control cells [16]. Loss of constitutive and inducible gene expression and the subsequent absence of high-level NO synthesis have been linked to host susceptibility to microbial invasion of the airway.
STAT-1 is required for IFN signal transduction in the cell and essential for the host response to virus infection [136], [137]. Despite numerous downstream targets of STAT-1 activation, loss of NOS II has been identified as one primary factor in the susceptibility of STAT-1 null animals to virus [138], [139], [140]. Previous studies have shown that nonfunctional JAK1 or Tyk2, receptor-associated kinases in the IFN signaling pathway, also result in decreased STAT-1 protein and activation, and a defective antiviral state, although the response to IFNα or γ is intact [139], [140]. The Tyk2-deficient cells display a phenotype remarkably analogous to CF airway epithelial cells: impairment of STAT-1 activation, increased virus replication in cells, and lack of NOS II induction, but most all other IFN-dependent pathways are intact. Altogether, these studies support that NOS II is one of the central mediators of the antiviral effects of STAT-1 activation in the human airway.
Therapies with NO as antiviral agent
Chemical donors of NO or induction of NOS II inhibit virus replication in vitro [16], [34], [35], [57], [58], [59], [60], [61], [62], [63], [64]. Since loss of NO synthesis is a primary pathophysiologic event in the susceptibility of CF patients to virus and other microbial pathogens [16], [87], overexpression of NOS II or NO donors is rational strategy to protect from viral infection. These approaches are successful in reducing virus infection of CF airway epithelial cells in culture [16] (Fig. 5 ). Important for application to patient care, providing NO donor after virus infection also significantly reduces virus replication [58], [59], [62], [63], [141], [142]. Thus, exogenous application of NO to the lung is a prospect for future antiviral therapy especially in the CF neonate in whom virus infection is a life-threatening event.
Fig. 5.
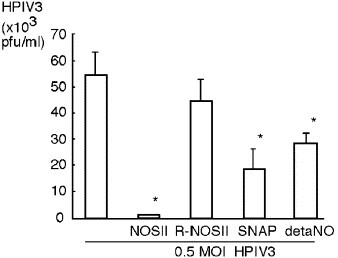
NOS II overexpression or NO donors protect CF cells from HPIV3 infection. Plaque assay of media overlying CF cells 24 h after HPIV3 infection (0.5 multiplicity of infections). Twenty-four hours prior to infection, CF cells were transfected with NOS II expression construct (NOSII), reverse sequence NOS II expression construct (R-NOSII), or left untreated. At the time of infection, some untreated CF cells were exposed to NO donors [SNAP, or deta NONOate (detaNO)]. Untreated cells have higher titer of infectious virus production than cells with NOS II-transgene or with NO donors [16]. Reprinted from S. Zheng et al., Immunity, Vol. 18, pp. 619–630, copyright (2003) with permission from Elsevier.
Conclusion
NO is a critical component of the antiviral host defense mechanisms in airway epithelium. Healthy human airway epithelium has abundant expression of NOS II due to continuous transcriptional activation of the gene in vivo. NOS II gene expression increases rapidly with viral infection in response to the viral replicative intermediate dsRNA activation of PKR signaling events. Multiple mechanisms function to support NO synthesis later in the course of virus infection including the IFN/STAT-1 pathways (Fig. 4). Thus, the airway epithelium has highly efficient NO synthetic machinery which is amplified in viral infection. Loss of NO synthesis in lung diseases such as CF predisposes one to increased virus/microbe infection due to defects in signal transduction through STAT-1. Clinical epidemiology suggests that virus infections impact on early morbidity and tempo of disease progression in CF. In fact, although extension of CF children’s life expectancy occurred with the introduction of antibiotics, vaccination against virus such as measles was one of the earliest interventions that led to improved survival in the 1960s [143]. Prior to viral vaccinations, CF neonates who contracted measles, a virus like HPIV3 in the paramyxovirus family, had progressive decline with worsening of pulmonary infections and early death [144], [145]. Thus, therapies aimed at improving antiviral host defense may further improve and extend the life of CF individuals. On the other hand, chronic high levels of NO lead to oxidative stress and airway injury, so the optimal timing, duration, and amount of NO supplementation which may benefit patients will be critical in planning therapy.
Acknowledgments
W.X. and S.C.E. were supported by AI70649, HL60917, and M01 RR018390 from the National Center for Research Resources. R.A.D. was supported by HL-68863.
Biographies
Weiling Xu, M.D., obtained her M.D. from Faculty of Medicine, Shanghai Medical University, Shanghai, China, and postdoctoral training at Cleveland Clinic Foundation, Cleveland. Her research focuses on basic mechanisms of airway inflammation in asthma and pulmonary hypertension.
Shuo Zheng, Ph.D., obtained her B.E. in Analytical Chemistry from Beijing University of Chemical Technology, Beijing, China, and Ph.D. in Clinical-Bioanalytical Chemistry from Cleveland State University, Cleveland. Her research focuses on gene regulation and airway epithelial biology.
Raed A. Dweik, M.D., is a graduate of The University of Jordan Faculty of Medicine. He completed his residency at Wright State University and his pulmonary fellowship at the Cleveland Clinic. His research focuses on studying the role of nitric oxide in lung physiology and in the pathophysiology of lung diseases.
Serpil Erzurum, M.D., obtained her M.D. from Northeastern Ohio Universities College of Medicine, and postdoctoral training at Baylor College of Medicine, University of Colorado and the National Institutes of Health. Her research focuses on investigation of airway and pulmonary vascular diseases.
Footnotes
This article is part of a series of reviews on “Redox signaling in immune function and cellular responses in lung injury and diseases.” The full list of papers may be found on the home page of the journal.
References
- 1.Makela M.J., Puhakka T., Ruuskanen O., Leinonen M., Saikku P., Kimpimaki M., Blomqvist S., Hyypia T., Arstila P. Viruses and bacteria in the etiology of the common cold. J. Clin. Microbiol. 1998;36(2):539–542. doi: 10.1128/jcm.36.2.539-542.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jacoby D.B. Virus-induced asthma attacks. JAMA. 2002;287(6):755–761. doi: 10.1001/jama.287.6.755. [DOI] [PubMed] [Google Scholar]
- 3.File T.M. Viral respiratory tract infections: increasing importance and a new pathogen. Curr. Opin. Infect. Dis. 2003;16(2):125–127. doi: 10.1097/00001432-200304000-00008. [DOI] [PubMed] [Google Scholar]
- 4.Gwaltney J.M., Jr., Phillips C.D., Miller R.D., Riker D.K. Computed tomographic study of the common cold. N. Engl. J. Med. 1994;330(1):25–30. doi: 10.1056/NEJM199401063300105. [DOI] [PubMed] [Google Scholar]
- 5.Pattemore P.K., Johnston S.L., Bardin P.G. Viruses as precipitants of asthma symptoms: I. Epidemiology. Clin. Exp. Allergy. 1992;22(3):325–336. doi: 10.1111/j.1365-2222.1992.tb03094.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hayden F.G. Rhinovirus and the lower respiratory tract. Rev. Med. Virol. 2004;14(1):17–31. doi: 10.1002/rmv.406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Fendrick A.M. Viral respiratory infections due to rhinoviruses: current knowledge, new developments. Am. J. Ther. 2003;10(3):193–202. doi: 10.1097/00045391-200305000-00006. [DOI] [PubMed] [Google Scholar]
- 8.Shay D.K., Holman R.C., Newman R.D., Liu L.L., Stout J.W., Anderson L.J. Bronchiolitis-associated hospitalizations among US children, 1980-1996. JAMA. 1999;282(15):1440–1446. doi: 10.1001/jama.282.15.1440. [DOI] [PubMed] [Google Scholar]
- 9.Holman R.C., Shay D.K., Curns A.T., Lingappa J.R., Anderson L.J. Risk factors for bronchiolitis-associated deaths among infants in the United States. Pediatr. Infect. Dis. J. 2003;22(6):483–490. doi: 10.1097/01.inf.0000069765.43405.3b. [DOI] [PubMed] [Google Scholar]
- 10.Peebles R.S., Jr., Graham B.S. Pathogenesis of respiratory syncytial virus infection in the murine model. Proc. Am. Thorac. Soc. 2005;2(2):110–115. doi: 10.1513/pats.200501-002AW. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Uetani K., Der S.D., Zamanian-Daryoush M., de La Motte C., Lieberman B.Y., Williams B.R., Erzurum S.C. Central role of double-stranded RNA-activated protein kinase in microbial induction of nitric oxide synthase. J. Immunol. 2000;165(2):988–996. doi: 10.4049/jimmunol.165.2.988. [DOI] [PubMed] [Google Scholar]
- 12.Garcia-Sastre A., Durbin R.K., Zheng H., Palese P., Gertner R., Levy D.E., Durbin J.E. The role of interferon in influenza virus tissue tropism. J. Virol. 1998;72(11):8550–8558. doi: 10.1128/jvi.72.11.8550-8558.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Holtzman M.J., Tyner J.W., Kim E.Y., Lo M.S., Patel A.C., Shornick L.P., Agapov E., Zhang Y. Acute and chronic airway responses to viral infection: implications for asthma and chronic obstructive pulmonary disease. Proc. Am. Thorac. Soc. 2005;2(2):132–140. doi: 10.1513/pats.200502-015AW. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kao Y.J., Piedra P.A., Larsen G.L., Colasurdo G.N. Induction and regulation of nitric oxide synthase in airway epithelial cells by respiratory syncytial virus. Am. J. Respir. Crit. Care Med. 2001;163(2):532–539. doi: 10.1164/ajrccm.163.2.9912068. [DOI] [PubMed] [Google Scholar]
- 15.Folkerts G., Busse W.W., Nijkamp F.P., Sorkness R., Gern J.E. Virus-induced airway hyperresponsiveness and asthma. Am. J. Respir. Crit. Care Med. 1998;157(6 Pt 1):1708–1720. doi: 10.1164/ajrccm.157.6.9707163. [DOI] [PubMed] [Google Scholar]
- 16.Zheng S., De B.P., Choudhary S., Comhair S.A., Goggans T., Slee R., Williams B.R., Pilewski J., Haque S.J., Erzurum S.C. Impaired innate host defense causes susceptibility to respiratory virus infections in cystic fibrosis. Immunity. 2003;18(5):619–630. doi: 10.1016/s1074-7613(03)00114-6. [DOI] [PubMed] [Google Scholar]
- 17.Stark J.M., Khan A.M., Chiappetta C.L., Xue H., Alcorn J.L., Colasurdo G.N. Immune and functional role of nitric oxide in a mouse model of respiratory syncytial virus infection. J. Infect. Dis. 2005;191(3):387–395. doi: 10.1086/427241. [DOI] [PubMed] [Google Scholar]
- 18.Sanders S.P., Siekierski E.S., Porter J.D., Richards S.M., Proud D. Nitric oxide inhibits rhinovirus-induced cytokine production and viral replication in a human respiratory epithelial cell line. J. Virol. 1998;72(2):934–942. doi: 10.1128/jvi.72.2.934-942.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Stuehr D.J., Griffith O.W. Mammalian nitric oxide synthases. Adv. Enzymol. Relat. Areas Mol. Biol. 1992;65:287–346. doi: 10.1002/9780470123119.ch8. [DOI] [PubMed] [Google Scholar]
- 20.Nathan C., Xie Q.W. Nitric oxide synthases: roles, tolls, and controls. Cell. 1994;78(6):915–918. doi: 10.1016/0092-8674(94)90266-6. [DOI] [PubMed] [Google Scholar]
- 21.Gaston B., Drazen J.M., Loscalzo J., Stamler J.S. The biology of nitrogen oxides in the airways. Am. J. Respir. Crit. Care Med. 1994;149(2 Pt 1):538–551. doi: 10.1164/ajrccm.149.2.7508323. [DOI] [PubMed] [Google Scholar]
- 22.Nathan C. Nitric oxide as a secretory product of mammalian cells. FASEB J. 1992;6(12):3051–3064. [PubMed] [Google Scholar]
- 23.Schmidt H.H., Walter U. NO at work. Cell. 1994;78(6):919–925. doi: 10.1016/0092-8674(94)90267-4. [DOI] [PubMed] [Google Scholar]
- 24.Guo F.H., Erzurum S.C. Characterization of inducible nitric oxide synthase expression in human airway epithelium. Environ. Health Perspect. 1998;106(Suppl. 5):1119–1124. doi: 10.1289/ehp.98106s51119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Guo F.H., Uetani K., Haque S.J., Williams B.R., Dweik R.A., Thunnissen F.B., Calhoun W., Erzurum S.C. Interferon gamma and interleukin 4 stimulate prolonged expression of inducible nitric oxide synthase in human airway epithelium through synthesis of soluble mediators. J. Clin. Invest. 1997;100(4):829–838. doi: 10.1172/JCI119598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Guo F.H., Comhair S.A., Zheng S., Dweik R.A., Eissa N.T., Thomassen M.J., Calhoun W., Erzurum S.C. Molecular mechanisms of increased nitric oxide (NO) in asthma: evidence for transcriptional and post-translational regulation of NO synthesis. J. Immunol. 2000;164(11):5970–5980. doi: 10.4049/jimmunol.164.11.5970. [DOI] [PubMed] [Google Scholar]
- 27.Dweik R.A., Laskowski D., Abu-Soud H.M, Kaneko R., Hutte D.J., Stuehr S.C. Nitric oxide synthesis in the lung. Regulation by oxygen through a kinetic mechanism. J. Clin. Invest. 1998;101(3):660–666. doi: 10.1172/JCI1378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Barnes P.J. Nitric oxide and airway disease. Ann. Med. 1995;27(3):389–393. doi: 10.3109/07853899509002592. [DOI] [PubMed] [Google Scholar]
- 29.Guo F.H., De Raeve H.R., Rice T.W., Stuehr D.J., Thunnissen F.B., Erzurum S.C. Continuous nitric oxide synthesis by inducible nitric oxide synthase in normal human airway epithelium in vivo. Proc. Natl. Acad. Sci. USA. 1995;92(17):7809–7813. doi: 10.1073/pnas.92.17.7809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kobzik L., Bredt D.S., Lowenstein C.J., Drazen J., Gaston B., Sugarbaker D., Stamler J.S. Nitric oxide synthase in human and rat lung: immunocytochemical and histochemical localization. Am. J. Respir. Cell. Mol. Biol. 1993;9(4):371–377. doi: 10.1165/ajrcmb/9.4.371. [DOI] [PubMed] [Google Scholar]
- 31.Stuehr D.J. Mammalian nitric oxide synthases. Biochim. Biophys. Acta. 1999;1411(2−3):217–230. doi: 10.1016/s0005-2728(99)00016-x. [DOI] [PubMed] [Google Scholar]
- 32.Lundberg J.O., Farkas-Szallasi T., Weitzberg E., Rinder J., Lidholm J., Anggaard A., Hokfelt T., Lundberg J.M., Alving K. High nitric oxide production in human paranasal sinuses. Nat. Med. 1995;1(4):370–373. doi: 10.1038/nm0495-370. [DOI] [PubMed] [Google Scholar]
- 33.Lane C., Knight D., Burgess S., Franklin P., Horak F., Legg J., Moeller A., Stick S. Epithelial inducible nitric oxide synthase activity is the major determinant of nitric oxide concentration in exhaled breath. Thorax. 2004;59(9):757–760. doi: 10.1136/thx.2003.014894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Flodstrom M., Horwitz M.S., Maday A., Balakrishna D., Rodriguez E., Sarvetnick N. A critical role for inducible nitric oxide synthase in host survival following coxsackievirus B4 infection. Virology. 2001;281(2):205–215. doi: 10.1006/viro.2000.0801. [DOI] [PubMed] [Google Scholar]
- 35.Noda S., Tanaka K., Sawamura S., Sasaki M., Matsumoto T., Mikami K., Aiba Y., Hasegawa H., Kawabe N., Koga Y. Role of nitric oxide synthase type 2 in acute infection with murine cytomegalovirus. J. Immunol. 2001;166(5):3533–3541. doi: 10.4049/jimmunol.166.5.3533. [DOI] [PubMed] [Google Scholar]
- 36.Karupiah G., Chen J.H., Mahalingam S., Nathan C.F., MacMicking J.D. Rapid interferon gamma-dependent clearance of influenza A virus and protection from consolidating pneumonitis in nitric oxide synthase 2-deficient mice. J. Exp. Med. 1998;188(8):1541–1546. doi: 10.1084/jem.188.8.1541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Aulak K.S., Miyagi M., Yan L., West K.A., Massillon D., Crabb J.W., Stuehr D.J. Proteomic method identifies proteins nitrated in vivo during inflammatory challenge. Proc. Natl. Acad. Sci. USA. 2001;98(21):12056–12061. doi: 10.1073/pnas.221269198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Aulak K.S., Koeck T., Crabb J.W., Stuehr D.J. Dynamics of protein nitration in cells and mitochondria. Am. J. Physiol. Heart Circ. Physiol. 2004;286(1):H30–H38. doi: 10.1152/ajpheart.00743.2003. [DOI] [PubMed] [Google Scholar]
- 39.Gow A.J., Farkouh C.R., Munson D.A., Posencheg M.A., Ischiropoulos H. Biological significance of nitric oxide-mediated protein modifications. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004;287(2):L262–L268. doi: 10.1152/ajplung.00295.2003. [DOI] [PubMed] [Google Scholar]
- 40.Titheradge M.A. Nitric oxide in septic shock. Biochim. Biophys. Acta. 1999;1411(2−3):437–455. doi: 10.1016/s0005-2728(99)00031-6. [DOI] [PubMed] [Google Scholar]
- 41.Lyons C.R. The role of nitric oxide in inflammation. Adv. Immunol. 1995;60:323–371. doi: 10.1016/s0065-2776(08)60589-1. [DOI] [PubMed] [Google Scholar]
- 42.Yates D.H. Role of exhaled nitric oxide in asthma. Immunol. Cell Biol. 2001;79(2):178–190. doi: 10.1046/j.1440-1711.2001.00990.x. [DOI] [PubMed] [Google Scholar]
- 43.Barnes P.J., Belvisi M.G. Nitric oxide and lung disease. Thorax. 1993;48(10):1034–1043. doi: 10.1136/thx.48.10.1034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wildhaber J.H., Hall G.L., Stick S.M. Measurements of exhaled nitric oxide with the single-breath technique and positive expiratory pressure in infants. Am. J. Respir. Crit. Care Med. 1999;159(1):74–78. doi: 10.1164/ajrccm.159.1.9805021. [DOI] [PubMed] [Google Scholar]
- 45.Baraldi E., Dario C., Ongaro R., Scollo M., Azzolin N.M., Panza N., Paganini N., Zacchello F. Exhaled nitric oxide concentrations during treatment of wheezing exacerbation in infants and young children. Am. J. Respir. Crit. Care Med. 1999;159(4 Pt. 1):1284–1288. doi: 10.1164/ajrccm.159.4.9807084. [DOI] [PubMed] [Google Scholar]
- 46.Ischiropoulos H., Mendiguren I., Fisher D., Fisher A.B., Thom S.R. Role of neutrophils and nitric oxide in lung alveolar injury from smoke inhalation. Am. J. Respir. Crit. Care Med. 1994;150(2):337–341. doi: 10.1164/ajrccm.150.2.8049812. [DOI] [PubMed] [Google Scholar]
- 47.Ozkan M., Dweik R.A., Laskowski D., Arroliga A.C., Erzurum S.C. High levels of nitric oxide in individuals with pulmonary hypertension receiving epoprostenol therapy. Lung. 2001;179(4):233–243. doi: 10.1007/s004080000064. [DOI] [PubMed] [Google Scholar]
- 48.Kaneko F.T., Arroliga A.C., Dweik R.A., Comhair S.A., Laskowski D., Oppedisano R., Thomassen M.J., Erzurum S.C. Biochemical reaction products of nitric oxide as quantitative markers of primary pulmonary hypertension. Am. J. Respir. Crit. Care Med. 1998;158(3):917–923. doi: 10.1164/ajrccm.158.3.9802066. [DOI] [PubMed] [Google Scholar]
- 49.Xu W., Kaneko F.T., Zheng S., Comhair S.A., Janocha A.J., Goggans T., Thunnissen F.B., Farver C., Hazen S.L., Jennings C., Dweik R.A., Arroliga A.C., Erzurum S.C. Increased arginase II and decreased NO synthesis in endothelial cells of patients with pulmonary arterial hypertension. FASEB J. 2004;18(14):1746–1748. doi: 10.1096/fj.04-2317fje. [DOI] [PubMed] [Google Scholar]
- 50.Persson M.G., Zetterstrom O., Agrenius V., Ihre E., Gustafsson L.E. Single-breath nitric oxide measurements in asthmatic patients and smokers. Lancet. 1994;343(8890):146–147. doi: 10.1016/s0140-6736(94)90935-0. [DOI] [PubMed] [Google Scholar]
- 51.Schilling J., Holzer P., Guggenbach M., Gyurech D., Marathia K., Geroulanos S. Reduced endogenous nitric oxide in the exhaled air of smokers and hypertensives. Eur. Respir. J. 1994;7(3):467–471. doi: 10.1183/09031936.94.07030467. [DOI] [PubMed] [Google Scholar]
- 52.Kharitonov S.A., Robbins R.A., Yates D., Keatings V., Barnes P.J. Acute and chronic effects of cigarette smoking on exhaled nitric oxide. Am. J. Respir. Crit. Care Med. 1995;152(2):609–612. doi: 10.1164/ajrccm.152.2.7543345. [DOI] [PubMed] [Google Scholar]
- 53.de Kluijver J., Evertse C.E., Sont J.K., Schrumpf J.A., van Zeijl-van der Ham C.J., Dick C.R., Rabe K.F., Hiemstra P.S., Sterk P.J. Are rhinovirus-induced airway responses in asthma aggravated by chronic allergen exposure? Am. J. Respir. Crit. Care Med. 2003;168(10):1174–1180. doi: 10.1164/rccm.200212-1520OC. [DOI] [PubMed] [Google Scholar]
- 54.Murphy A.W., Platts-Mills T.A., Lobo M., Hayden F. Respiratory nitric oxide levels in experimental human influenza. Chest. 1998;114(2):452–456. doi: 10.1378/chest.114.2.452. [DOI] [PubMed] [Google Scholar]
- 55.Zsengeller Z.K., Ross G.F., Trapnell B.C., Szabo C., Whitsett J.A. Adenovirus infection increases iNOS and peroxynitrite production in the lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2001;280(3):L503–L511. doi: 10.1152/ajplung.2001.280.3.L503. [DOI] [PubMed] [Google Scholar]
- 56.Loveless M.O., Phillips C.R., Giraud G.D., Holden W.E. Decreased exhaled nitric oxide in subjects with HIV infection. Thorax. 1997;52(2):185–186. doi: 10.1136/thx.52.2.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Karupiah G., Chen J.H., Nathan C.F., Mahalingam S., MacMicking J.D. Identification of nitric oxide synthase 2 as an innate resistance locus against ectromelia virus infection. J. Virol. 1998;72(9):7703–7706. doi: 10.1128/jvi.72.9.7703-7706.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Karupiah G., Harris N. Inhibition of viral replication by nitric oxide and its reversal by ferrous sulfate and tricarboxylic acid cycle metabolites. J. Exp. Med. 1995;181(6):2171–2179. doi: 10.1084/jem.181.6.2171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zaragoza C., Ocampo C.J., Saura M., McMillan A., Lowenstein C.J. Nitric oxide inhibition of coxsackievirus replication in vitro. J. Clin. Invest. 1997;100(7):1760–1767. doi: 10.1172/JCI119702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kosugi I., Kawasaki H., Arai Y., Tsutsui Y. Innate immune responses to cytomegalovirus infection in the developing mouse brain and their evasion by virus-infected neurons. Am. J. Pathol. 2002;161(3):919–928. doi: 10.1016/S0002-9440(10)64252-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Harris N., Buller R.M., Karupiah G. Gamma interferon-induced, nitric oxide-mediated inhibition of vaccinia virus replication. J. Virol. 1995;69(2):910–915. doi: 10.1128/jvi.69.2.910-915.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Croen K.D. Evidence for antiviral effect of nitric oxide. Inhibition of herpes simplex virus type 1 replication. J. Clin. Invest. 1993;91(6):2446–2452. doi: 10.1172/JCI116479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Rimmelzwaan G.F., Baars M.M., de Lijster P., Fouchier R.A., Osterhaus A.D. Inhibition of influenza virus replication by nitric oxide. J. Virol. 1999;73(10):8880–8883. doi: 10.1128/jvi.73.10.8880-8883.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Saura M., Zaragoza C., McMillan A., Quick R.A., Hohenadl C., Lowenstein J.M., Lowenstein C.J. An antiviral mechanism of nitric oxide: inhibition of a viral protease. Immunity. 1999;10(1):21–28. doi: 10.1016/S1074-7613(00)80003-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Sanders S.P., Kim J., Connolly K.R., Porter J.D., Siekierski E.S., Proud D. Nitric oxide inhibits rhinovirus-induced granulocyte macrophage colony-stimulating factor production in bronchial epithelial cells. Am. J. Respir. Cell. Mol. Biol. 2001;24(3):317–325. doi: 10.1165/ajrcmb.24.3.4131. [DOI] [PubMed] [Google Scholar]
- 66.Zhou A., Paranjape J.M., Der S.D., Williams B.R., Silverman R.H. Interferon action in triply deficient mice reveals the existence of alternative antiviral pathways. Virology. 1999;258(2):435–440. doi: 10.1006/viro.1999.9738. [DOI] [PubMed] [Google Scholar]
- 67.Xiang Y., Condit R.C., Vijaysri S., Jacobs B., Williams B.R., Silverman R.H. Blockade of interferon induction and action by the E3L double-stranded RNA binding proteins of vaccinia virus. J. Virol. 2002;76(10):5251–5259. doi: 10.1128/JVI.76.10.5251-5259.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Biron C.A. Initial and innate responses to viral infections–pattern setting in immunity or disease. Curr. Opin. Microbiol. 1999;2(4):374–381. doi: 10.1016/s1369-5274(99)80066-6. [DOI] [PubMed] [Google Scholar]
- 69.Akerstrom S., Mousavi-Jazi M., Klingstrom J., Leijon M., Lundkvist A., Mirazimi A. Nitric oxide inhibits the replication cycle of severe acute respiratory syndrome coronavirus. J. Virol. 2005;79(3):1966–1969. doi: 10.1128/JVI.79.3.1966-1969.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Lepoivre M., Fieschi F., Coves J., Thelander L., Fontecave M. Inactivation of ribonucleotide reductase by nitric oxide. Biochem. Biophys. Res. Commun. 1991;179(1):442–448. doi: 10.1016/0006-291x(91)91390-x. [DOI] [PubMed] [Google Scholar]
- 71.Grisham M.B., Jourd'Heuil D., Wink D.A. Nitric oxide. I. Physiological chemistry of nitric oxide and its metabolites: implications in inflammation. Am. J. Physiol. 1999;276(2 Pt. 1):G315–G321. doi: 10.1152/ajpgi.1999.276.2.G315. [DOI] [PubMed] [Google Scholar]
- 72.Adler H., Beland J.L., Del-Pan N.C., Kobzik L., Brewer J.P., Martin T.R., Rimm I.J. Suppression of herpes simplex virus type 1 (HSV-1)-induced pneumonia in mice by inhibition of inducible nitric oxide synthase (iNOS, NOS2) J. Exp. Med. 1997;185(9):1533–1540. doi: 10.1084/jem.185.9.1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Tanaka K., Nakazawa H., Okada K., Umezawa K., Fukuyama N., Koga Y. Nitric oxide mediates murine cytomegalovirus-associated pneumonitis in lungs that are free of the virus. J. Clin. Invest. 1997;100(7):1822–1830. doi: 10.1172/JCI119710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Akaike T., Noguchi Y., Ijiri S., Setoguchi K., Suga M., Zheng Y.M., Dietzschold B., Maeda H. Pathogenesis of influenza virus-induced pneumonia: involvement of both nitric oxide and oxygen radicals. Proc. Natl. Acad. Sci. US A. 1996;93(6):2448–2453. doi: 10.1073/pnas.93.6.2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Konstan M.W.a.B., Melvin . In: Cystic fibrosis. Davis P.B.,, editor. 1993. Infection and inflammation of the lung in cystic fibrosis; p. 223. [Google Scholar]
- 76.Rosenfeld M., Ramsey B. Evolution of airway microbiology in the infant with cystic fibrosis: role of nonpseudomonal and pseudomonal pathogens. Semin. Respir. Infect. 1992;7(3):158–167. [PubMed] [Google Scholar]
- 77.Hordvik N.L., Konig P., Hamory B., Cooperstock M., Kreutz C., Gayer D., Barbero G. Effects of acute viral respiratory tract infections in patients with cystic fibrosis. Pediatr. Pulmonol. 1989;7(4):217–222. doi: 10.1002/ppul.1950070406. [DOI] [PubMed] [Google Scholar]
- 78.Asano K., Chee C.B., Gaston B., Lilly C.M., Gerard C., Drazen J.M., Stamler J.S. Constitutive and inducible nitric oxide synthase gene expression, regulation, and activity in human lung epithelial cells. Proc. Natl. Acad. Sci. USA. 1994;91(21):10089–10093. doi: 10.1073/pnas.91.21.10089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hiatt P.W., Grace S.C., Kozinetz C.A., Raboudi S.H., Treece D.G., Taber L.H., Piedra P.A. Effects of viral lower respiratory tract infection on lung function in infants with cystic fibrosis. Pediatrics. 1999;103(3):619–626. doi: 10.1542/peds.103.3.619. [DOI] [PubMed] [Google Scholar]
- 80.Wang E.E., Prober C.G., Manson B., Corey M., Levison H. Association of respiratory viral infections with pulmonary deterioration in patients with cystic fibrosis. N. Engl. J. Med. 1984;311(26):1653–1658. doi: 10.1056/NEJM198412273112602. [DOI] [PubMed] [Google Scholar]
- 81.Petersen N.T., Hoiby N., Mordhorst C.H., Lind K., Flensborg E.W., Bruun B. Respiratory infections in cystic fibrosis patients caused by virus, chlamydia and mycoplasma—Possible synergism with Pseudomonas aeruginosa. Acta Paediatr. Scand. 1981;70(5):623–628. doi: 10.1111/j.1651-2227.1981.tb05757.x. [DOI] [PubMed] [Google Scholar]
- 82.Armstrong D., Grimwood K., Carlin J.B., Carzino R., Hull J., Olinsky A., Phelan P.D. Severe viral respiratory infections in infants with cystic fibrosis. Pediatr. Pulmonol. 1998;26(6):371–379. doi: 10.1002/(sici)1099-0496(199812)26:6<371::aid-ppul1>3.0.co;2-n. [DOI] [PubMed] [Google Scholar]
- 83.Balfour-Lynn I.M., Laverty A., Dinwiddie R. Reduced upper airway nitric oxide in cystic fibrosis. Arch. Dis. Child. 1996;75(4):319–322. doi: 10.1136/adc.75.4.319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Lundberg J.O., Nordvall S.L., Weitzberg E., Kollberg H., Alving K. Exhaled nitric oxide in paediatric asthma and cystic fibrosis. Arch Dis. Child. 1996;75(4):323–326. doi: 10.1136/adc.75.4.323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Cetin I., Ozcelik U., Gocmen A., Kiper N., Dogru D., Yalcin E. BALF nitrite as an indicator of inflammation in children with cystic fibrosis. Respiration. 2004;71(6):625–629. doi: 10.1159/000081764. [DOI] [PubMed] [Google Scholar]
- 86.Elphick H.E., Demoncheaux E.A., Ritson S., Higenbottam T.W., Everard M.L. Exhaled nitric oxide is reduced in infants with cystic fibrosis. Thorax. 2001;56(2):151–152. doi: 10.1136/thorax.56.2.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kelley T.J., Drumm M.L. Inducible nitric oxide synthase expression is reduced in cystic fibrosis murine and human airway epithelial cells. J. Clin. Invest. 1998;102(6):1200–1207. doi: 10.1172/JCI2357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Meng Q.H., Springall D.R., Bishop A.E., Morgan K., Evans T.J., Habib S., Gruenert D.C., Gyi K.M., Hodson M.E., Yacoub M.H., Polak J.M. Lack of inducible nitric oxide synthase in bronchial epithelium: a possible mechanism of susceptibility to infection in cystic fibrosis. J. Pathol. 1998;184(3):323–331. doi: 10.1002/(SICI)1096-9896(199803)184:3<323::AID-PATH2>3.0.CO;2-2. [DOI] [PubMed] [Google Scholar]
- 89.Zheng S., Xu W., Bose S., Banerjee A.K., Haque S.J., Erzurum S.C. Impaired nitric oxide synthase-2 signaling pathway in cystic fibrosis airway epithelium. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004;287(2):L374–L381. doi: 10.1152/ajplung.00039.2004. [DOI] [PubMed] [Google Scholar]
- 90.Miller R.A., Britigan B.E. Role of oxidants in microbial pathophysiology. Clin. Microbiol. Rev. 1997;10(1):1–18. doi: 10.1128/cmr.10.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Akaike T., Suga M., Maeda H. Free radicals in viral pathogenesis: molecular mechanisms involving superoxide and NO. Proc. Soc. Exp. Biol. Med. 1998;217(1):64–73. doi: 10.3181/00379727-217-44206. [DOI] [PubMed] [Google Scholar]
- 92.Andreadis A.A., Hazen S.L., Comhair S.A., Erzurum S.C. Oxidative and nitrosative events in asthma. Free Radic. Biol. Med. 2003;35(3):213–225. doi: 10.1016/s0891-5849(03)00278-8. [DOI] [PubMed] [Google Scholar]
- 93.Valyi-Nagy T., Dermody T.S. Role of oxidative damage in the pathogenesis of viral infections of the nervous system. Histol. Histopathol. 2005;20(3):957–967. doi: 10.14670/HH-20.957. [DOI] [PubMed] [Google Scholar]
- 94.Kato M., Hayashi Y., Kimura H. Oxygen radicals in inflammation and allergy related to viral infections. Curr. Drug Targets Inflamm. Allergy. 2005;4(4):497–501. doi: 10.2174/1568010054526377. [DOI] [PubMed] [Google Scholar]
- 95.Brennan M.L., Wu W., Fu X., Shen Z., Song W., Frost H., Vadseth C., Narine L., Lenkiewicz E., Borchers M.T., Lusis A.J., Lee J.J., Lee N.A., Abu-Soud H.M., Ischiropoulos H., Hazen S.L. A tale of two controversies: defining both the role of peroxidases in nitrotyrosine formation in vivo using eosinophil peroxidase and myeloperoxidase-deficient mice, and the nature of peroxidase-generated reactive nitrogen species. J. Biol. Chem. 2002;277(20):17415–17427. doi: 10.1074/jbc.M112400200. [DOI] [PubMed] [Google Scholar]
- 96.Spalteholz H., Wenske K., Panasenko O.M., Schiller J., Arnhold J. Evaluation of products upon the reaction of hypohalous acid with unsaturated phosphatidylcholines. Chem. Phys. Lipids. 2004;129(1):85–96. doi: 10.1016/j.chemphyslip.2003.12.005. [DOI] [PubMed] [Google Scholar]
- 97.Kawai Y., Morinaga H., Kondo H., Miyoshi N., Nakamura Y., Uchida K., Osawa T. Endogenous formation of novel halogenated 2′-deoxycytidine. Hypohalous acid-mediated DNA modification at the site of inflammation. J. Biol. Chem. 2004;279(49):51241–51249. doi: 10.1074/jbc.M408210200. [DOI] [PubMed] [Google Scholar]
- 98.El Messaoudi K., Verheyden A.M., Thiry L., Fourez S., Tasiaux N., Bollen A., Moguilevsky N. Human recombinant myeloperoxidase antiviral activity on cytomegalovirus. J. Med. Virol. 2002;66(2):218–223. [PubMed] [Google Scholar]
- 99.Chochola J., Yamaguchi Y., Moguilevsky N., Bollen A., Strosberg A.D., Stanislawski M. Virucidal effect of myeloperoxidase on human immunodeficiency virus type 1-infected T cells. Antimicrob. Agents Chemother. 1994;38(5):969–972. doi: 10.1128/aac.38.5.969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Klebanoff S.J. Oxygen metabolism and the toxic properties of phagocytes. Ann. Intern. Med. 1980;93(3):480–489. doi: 10.7326/0003-4819-93-3-480. [DOI] [PubMed] [Google Scholar]
- 101.Abu-Soud H.M., Khassawneh M.Y., Sohn J.T., Murray P., Haxhiu M.A., Hazen S.L. Peroxidases inhibit nitric oxide (NO) dependent bronchodilation: development of a model describing NO-peroxidase interactions. Biochemistry. 2001;40(39):11866–11875. doi: 10.1021/bi011206v. [DOI] [PubMed] [Google Scholar]
- 102.Galijasevic S., Saed G.M., Diamond M.P., Abu-Soud H.M. Myeloperoxidase up-regulates the catalytic activity of inducible nitric oxide synthase by preventing nitric oxide feedback inhibition. Proc. Natl. Acad. Sci. USA. 2003;100(25):14766–14771. doi: 10.1073/pnas.2435008100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Eiserich J.P., Baldus S., Brennan M.L., Ma W., Zhang C., Tousson A., Castro L., Lusis A.J., Nauseef W.M., White C.R., Freeman B.A. Myeloperoxidase, a leukocyte-derived vascular NO oxidase. Science. 2002;296(5577):2391–2394. doi: 10.1126/science.1106830. [DOI] [PubMed] [Google Scholar]
- 104.Abu-Soud H.M., Hazen S.L. Nitric oxide is a physiological substrate for mammalian peroxidases. J. Biol. Chem. 2000;275(48):37524–37532. doi: 10.1074/jbc.275.48.37524. [DOI] [PubMed] [Google Scholar]
- 105.Abu-Soud H.M., Hazen S.L. Nitric oxide modulates the catalytic activity of myeloperoxidase. J. Biol. Chem. 2000;275(8):5425–5430. doi: 10.1074/jbc.275.8.5425. [DOI] [PubMed] [Google Scholar]
- 106.Galijasevic S., Saed G.M., Hazen S.L., Abu-Soud H.M. Myeloperoxidase metabolizes thiocyanate in a reaction driven by nitric oxide. Biochemistry. 2006;45(4):1255–1262. doi: 10.1021/bi051438k. [DOI] [PubMed] [Google Scholar]
- 107.Comhair S.A., Erzurum S.C. Antioxidant responses to oxidant-mediated lung diseases. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002;283(2):L246–L255. doi: 10.1152/ajplung.00491.2001. [DOI] [PubMed] [Google Scholar]
- 108.Hudson V.M. Rethinking cystic fibrosis pathology: the critical role of abnormal reduced glutathione (GSH) transport caused by CFTR mutation. Free Radic. Biol. Med. 2001;30(12):1440–1461. doi: 10.1016/s0891-5849(01)00530-5. [DOI] [PubMed] [Google Scholar]
- 109.Roum J.H., Buhl R., McElvaney N.G., Borok Z., Crystal R.G. Systemic deficiency of glutathione in cystic fibrosis. J. Appl. Physiol. 1993;75(6):2419–2424. doi: 10.1152/jappl.1993.75.6.2419. [DOI] [PubMed] [Google Scholar]
- 110.Hull J., Vervaart P., Grimwood K., Phelan P. Pulmonary oxidative stress response in young children with cystic fibrosis. Thorax. 1997;52(6):557–560. doi: 10.1136/thx.52.6.557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Luzzi S.D., Marletta M.A. L-arginine analogs as alternate substrates for nitric oxide synthase. Bioorg. Med. Chem. Lett. 2005;15(17):3934–3941. doi: 10.1016/j.bmcl.2005.05.088. [DOI] [PubMed] [Google Scholar]
- 112.Geller D.A., Nussler A.K., Di Silvio M., Lowenstein C.J., Shapiro R.A., Wang S.C., Simmons R.L., Billiar T.R. Cytokines, endotoxin, and glucocorticoids regulate the expression of inducible nitric oxide synthase in hepatocytes. Proc. Natl. Acad. Sci. USA. 1993;90(2):522–526. doi: 10.1073/pnas.90.2.522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Marks-Konczalik J., Chu S.C., Moss J. Cytokine-mediated transcriptional induction of the human inducible nitric oxide synthase gene requires both activator protein 1 and nuclear factor kappaB-binding sites. J. Biol. Chem. 1998;273(35):22201–22208. doi: 10.1074/jbc.273.35.22201. [DOI] [PubMed] [Google Scholar]
- 114.de Vera M.E., Shapiro R.A., Nussler A.K., Mudgett J.S., Simmons R.L., Morris S.M., Jr., Billiar T.R., Geller D.A. Transcriptional regulation of human inducible nitric oxide synthase (NOS2) gene by cytokines: initial analysis of the human NOS2 promoter. Proc. Natl. Acad. Sci. USA. 1996;93(3):1054–1059. doi: 10.1073/pnas.93.3.1054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Gao J., Morrison D.C., Parmely T.J., Russell S.W., Murphy W.J. An interferon-gamma-activated site (GAS) is necessary for full expression of the mouse iNOS gene in response to interferon-gamma and lipopolysaccharide. J. Biol. Chem. 1997;272(2):1226–1230. doi: 10.1074/jbc.272.2.1226. [DOI] [PubMed] [Google Scholar]
- 116.Lowenstein C.J., Alley E.W., Raval P., Snowman A.M., Snyder S.H., Russell S.W., Murphy W.J. Macrophage nitric oxide synthase gene: two upstream regions mediate induction by interferon gamma and lipopolysaccharide. Proc. Natl. Acad. Sci. USA. 1993;90(20):9730–9734. doi: 10.1073/pnas.90.20.9730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Xie Q.W., Whisnant R., Nathan C. Promoter of the mouse gene encoding calcium-independent nitric oxide synthase confers inducibility by interferon gamma and bacterial lipopolysaccharide. J. Exp. Med. 1993;177(6):1779–1784. doi: 10.1084/jem.177.6.1779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ganster R.W., Taylor B.S., Shao L., Geller D.A. Complex regulation of human inducible nitric oxide synthase gene transcription by Stat 1 and NF-kappa B. Proc. Natl. Acad. Sci. USA. 2001;98(15):8638–8643. doi: 10.1073/pnas.151239498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Chu S.C., Marks-Konczalik J., Wu H.P., Banks T.C., Moss J. Analysis of the cytokine-stimulated human inducible nitric oxide synthase (iNOS) gene: characterization of differences between human and mouse iNOS promoters. Biochem. Biophys. Res. Commun. 1998;248(3):871–878. doi: 10.1006/bbrc.1998.9062. [DOI] [PubMed] [Google Scholar]
- 120.Zhang X., Laubach V.E., Alley E.W., Edwards K.A., Sherman P.A., Russell S.W., Murphy W.J. Transcriptional basis for hyporesponsiveness of the human inducible nitric oxide synthase gene to lipopolysaccharide/interferon-gamma. J. Leukoc. Biol. 1996;59(4):575–585. doi: 10.1002/jlb.59.4.575. [DOI] [PubMed] [Google Scholar]
- 121.Rusche K.M., Spiering M.M., Marletta M.A. Reactions catalyzed by tetrahydrobiopterin-free nitric oxide synthase. Biochemistry. 1998;37(44):15503–15512. doi: 10.1021/bi9813936. [DOI] [PubMed] [Google Scholar]
- 122.Xu W., Comhair S.A., Zheng S., Chu S.C., Marks-Konczalik J., Moss J., Haque S.J., Erzurum S.C. STAT-1 and c-Fos interaction in nitric oxide synthase-2 gene activation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003;285(1):L137–L148. doi: 10.1152/ajplung.00441.2002. [DOI] [PubMed] [Google Scholar]
- 123.van Dam H., Castellazzi M. Distinct roles of Jun : Fos and Jun : ATF dimers in oncogenesis. Oncogene. 2001;20(19):2453–2464. doi: 10.1038/sj.onc.1204239. [DOI] [PubMed] [Google Scholar]
- 124.Shaulian E., Karin M. AP-1 in cell proliferation and survival. Oncogene. 2001;20(19):2390–2400. doi: 10.1038/sj.onc.1204383. [DOI] [PubMed] [Google Scholar]
- 125.Chinenov Y., Kerppola T.K. Close encounters of many kinds: Fos-Jun interactions that mediate transcription regulatory specificity. Oncogene. 2001;20(19):2438–2452. doi: 10.1038/sj.onc.1204385. [DOI] [PubMed] [Google Scholar]
- 126.Jacobs B.L., Langland J.O. When two strands are better than one: the mediators and modulators of the cellular responses to double-stranded RNA. Virology. 1996;219(2):339–349. doi: 10.1006/viro.1996.0259. [DOI] [PubMed] [Google Scholar]
- 127.Majde J.A. Viral double-stranded RNA, cytokines, and the flu. J. Interferon Cytokine Res. 2000;20(3):259–272. doi: 10.1089/107999000312397. [DOI] [PubMed] [Google Scholar]
- 128.Kumar A., Krieger A., Symeoneides S., Parrillo J.E. Myocardial dysfunction in septic shock. Part II. Role of cytokines and nitric oxide. J. Cardiothorac. Vasc. Anesth. 2001;15(4):485–511. doi: 10.1053/jcan.2001.25003. [DOI] [PubMed] [Google Scholar]
- 129.Yang Y.L., Reis L.F., Pavlovic J., Aguzzi A., Schafer R., Kumar A., Williams B.R., Aguet M., Weissmann C. Deficient signaling in mice devoid of double-stranded RNA-dependent protein kinase. EMBO J. 1995;14(24):6095–6106. doi: 10.1002/j.1460-2075.1995.tb00300.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Suliman H.B., Ryan L.K., Bishop L., Folz R.J. Prevention of influenza-induced lung injury in mice overexpressing extracellular superoxide dismutase. Am. J. Physiol. Lung Cell. Mol. Physiol. 2001;280(1):L69–L78. doi: 10.1152/ajplung.2001.280.1.L69. [DOI] [PubMed] [Google Scholar]
- 131.Meurs E., Chong K., Galabru J., Thomas N.S., Kerr I.M., Williams B.R., Hovanessian A.G. Molecular cloning and characterization of the human double-stranded RNA-activated protein kinase induced by interferon. Cell. 1990;62(2):379–390. doi: 10.1016/0092-8674(90)90374-n. [DOI] [PubMed] [Google Scholar]
- 132.Kumar A., Yang Y.L., Flati V., Der S., Kadereit S., Deb A., Haque J., Reis L., Weissmann C., Williams B.R. Deficient cytokine signaling in mouse embryo fibroblasts with a targeted deletion in the PKR gene: role of IRF-1 and NF-kappaB. EMBO J. 1997;16(2):406–416. doi: 10.1093/emboj/16.2.406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Harada H., Fujita T., Miyamoto M., Kimura Y., Maruyama M., Furia A., Miyata T., Taniguchi T. Structurally similar but functionally distinct factors, IRF-1 and IRF-2, bind to the same regulatory elements of IFN and IFN-inducible genes. Cell. 1989;58(4):729–739. doi: 10.1016/0092-8674(89)90107-4. [DOI] [PubMed] [Google Scholar]
- 134.Watanabe N., Sakakibara J., Hovanessian A.G., Taniguchi T., Fujita T. Activation of IFN-beta element by IRF-1 requires a posttranslational event in addition to IRF-1 synthesis. Nucleic Acids Res. 1991;19(16):4421–4428. doi: 10.1093/nar/19.16.4421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Kirchhoff S., Koromilas A.E., Schaper F., Grashoff M., Sonenberg N., Hauser H. IRF-1 induced cell growth inhibition and interferon induction requires the activity of the protein kinase PKR. Oncogene. 1995;11(3):439–445. [PubMed] [Google Scholar]
- 136.Durbin J.E., Hackenmiller R., Simon M.C., Levy D.E. Targeted disruption of the mouse Stat1 gene results in compromised innate immunity to viral disease. Cell. 1996;84(3):443–450. doi: 10.1016/s0092-8674(00)81289-1. [DOI] [PubMed] [Google Scholar]
- 137.Meraz M.A., White J.M., Sheehan K.C., Bach E.A., Rodig S.J., Dighe A.S., Kaplan D.H., Riley J.K., Greenlund A.C., Campbell D., Carver-Moore K., DuBois R.N., Clark R., Aguet M., Schreiber R.D. Targeted disruption of the Stat1 gene in mice reveals unexpected physiologic specificity in the JAK-STAT signaling pathway. Cell. 1996;84(3):431–442. doi: 10.1016/s0092-8674(00)81288-x. [DOI] [PubMed] [Google Scholar]
- 138.Karupiah G., Xie Q.W., Buller R.M., Nathan C., Duarte C., MacMicking J.D. Inhibition of viral replication by interferon-gamma-induced nitric oxide synthase. Science. 1993;261(5127):1445–1448. doi: 10.1126/science.7690156. [DOI] [PubMed] [Google Scholar]
- 139.Briscoe J., Rogers N.C., Witthuhn B.A., Watling D., Harpur A.G., Wilks A.F., Stark G.R., Ihle J.N., Kerr I.M. Kinase-negative mutants of JAK1 can sustain interferon-gamma-inducible gene expression but not an antiviral state. EMBO J. 1996;15(4):799–809. [PMC free article] [PubMed] [Google Scholar]
- 140.Karaghiosoff M., Neubauer H., Lassnig C., Kovarik P., Schindler H., Pircher H., McCoy B., Bogdan C., Decker T., Brem G., Pfeffer K., Muller M. Partial impairment of cytokine responses in Tyk2-deficient mice. Immunity. 2000;13(4):549–560. doi: 10.1016/s1074-7613(00)00054-6. [DOI] [PubMed] [Google Scholar]
- 141.Mannick J.B., Asano K., Izumi K., Kieff E., Stamler J.S. Nitric oxide produced by human B lymphocytes inhibits apoptosis and Epstein-Barr virus reactivation. Cell. 1994;79(7):1137–1146. doi: 10.1016/0092-8674(94)90005-1. [DOI] [PubMed] [Google Scholar]
- 142.Zell R., Markgraf R., Schmidtke M., Gorlach M., Stelzner A., Henke A., Sigusch H.H., Gluck B. Nitric oxide donors inhibit the coxsackie virus B3 proteinases 2A and 3C in vitro, virus production in cells, and signs of myocarditis in virus-infected mice. Med. Microbiol. Immunol. (Berl.) 2004;193(2−3):91–100. doi: 10.1007/s00430-003-0198-6. [DOI] [PubMed] [Google Scholar]
- 143.Huang N.N., Macri C.N., Girone J., Sproul A. Survival of patients with cystic fibrosis. Am. J. Dis. Child. 1970;120(4):289–295. doi: 10.1001/archpedi.1970.02100090063002. [DOI] [PubMed] [Google Scholar]
- 144.Enders J.F., Mc C.K., Mitus A., Cheatham W.J. Isolation of measles virus at autopsy in cases of giant-cell pneumonia without rash. N. Engl. J. Med. 1959;261:875–881. doi: 10.1056/NEJM195910292611801. [DOI] [PubMed] [Google Scholar]
- 145.Shwachman H., Katz S., Kulczycki L.L. Attenuated measles vaccine in cystic fibrosis. Am. J. Dis. Child. 1962;103:405–409. doi: 10.1001/archpedi.1962.02080020417048. [DOI] [PubMed] [Google Scholar]