

Abstract
As most newly characterized emerging infectious diseases are considered to be zoonotic, a modern pre-eminence ascribed within this classification lies clearly within the viral taxonomic realm. In particular, RNA viruses deserve special concern given their documented impact on conservation biology, veterinary medicine and public health, with an unprecedented ability to promote an evolutionary host–pathogen arms race from the ultimate infection and immunity perspective. However, besides the requisite molecular/gross anatomical and physiological bases for infectious diseases to transmit from one host to another, both viral pathogens and their reservoirs/vectors exploit a complex anthropological, cultural, historical, psychological and social suite that specifically defines the phylodynamics within Homo sapiens, unlike any other species. Some of these variables include the ecological benefits of living in groups, decisions on hunting and foraging behaviours and dietary preferences, myths and religious doctrines, health economics, travel destinations, population planning, political decisions on agricultural product bans and many others, in a homo-sociome memetic complex. Taken to an extreme, such complexities elucidate the underpinnings of explanations as to why certain viral zoonoses reside in neglected people, places and things, whereas others are chosen selectively and prioritized for active mitigation. Canine-transmitted rabies serves as one prime example of how a neglected viral zoonosis may transition to greater attention on the basis of renewed advocacy, social media, local champions and vested international community engagement. In contrast, certain bat-associated and arboviral diseases suffer from basic ignorance and perpetuated misunderstanding of fundamental reservoir and vector ecology tenets, translated into failed control policies that only exacerbate the underlying environmental conditions of concern. Beyond applied biomedical knowledge, epidemiological skills and biotechnical abilities alone, if a homo-sociome memetic complex approach is also entertained in a modern transdisciplinary context, neglected viral zoonosis may be better understood, controlled, prevented and possibly eliminated, in a more holistic One Health context.
Keywords: Conservation medicine, ecohealth, emerging infectious diseases, neglected tropical infections, one health, viral zoonoses
Viral zoonotic underpinnings
All major human viral diseases once originated in other animal species and formed unconnected prehistoric biological islands, but now, new pathogens are being characterized at an unprecedented level with the potential for dramatic pandemics, due both to ecological perturbations and to technological improvements in detection [1]. Nevertheless, at a recent international biomedical conference, one speaker reminded the audience of the statistical ‘irrelevance’ of emerging infectious diseases, by categorical comparisons to major causes of human death today. Put simply, even populations in the developing world are approaching levels of cardiovascular disease, stroke, cancer and other chronic maladies, that were previously only found in developed countries. Although this may certainly be true, if an individual evades acute viral diarrhoea, pneumonia, haemorrhagic fever, hepatitis, encephalitis, etc. and survives out of infanthood to adolescence, those more modern facets of chronic illness are largely associated with a personal lifestyle elevated gradually to a community level. In contrast, zoonoses per se, are largely not a matter of conscious public choice, although the exposures that lead to a productive infection are often provoked directly by the individual. Irrespective of surreptitious media bragging rights to causality behind any fictitious ‘zombie apocalypse’ or its preparedness (http://www.cdc.gov/phpr/zombies.htm; http://www.cracked.com/article_15643_5-scientific-reasons-zombie-apocalypse-could-actually-happen.html) in the growing collective consciousness (for which a discerning scientific audience would have to ascribe not to bacteria, fungi or protozoa, but to a viral aetiology objectively), the image of a post-SARS world is only too real in the not-too-distant memory, given the actual extremes of current global headlines of translocated viral infections from one continent to another. Moreover, regardless of the magnitude of long-term lifestyle choices behind major human mortality trends, most health professionals should agree that the probability would be highly unlikely that a traveller today would be screened, detained and possibly quarantined against their will at an international port of entry, because of being a smoker, alcohol-user or obese, as opposed to a suggestion of an infection with a viral infectious aetiology, with obvious immediate consequences for the perceived greater public good. Hence, many would argue that viral zoonoses are indeed still relevant, based upon the evidence at hand.
Why are viral zoonoses relevant, but neglected? Considering the ultimate origins of historical pathogens as diverse as the eradicated smallpox and the much debated 1918 pandemic influenza, viruses are a highly relevant focus group when considering the general concept of zoonoses as a whole, from obvious biodiversity, to the sheer magnitude of their resultant cumulative morbidity and mortality in the animal kingdom writ large and the concomitant host–pathogen arms race [2]. Such viruses are the quintessential obligate microbiological parasites, whose diversity across all life on earth is unmatched. Rates of evolution, particularly among RNA viruses, excel at real-time orders of magnitude [3]. Antibiotics are useless, antiviral drugs remain limited and proper discovery requires a requisite twenty-first century laboratory sophistication, relatively far from the ecosystems where these zoonoses perpetuate, from urban mega-slums to increasingly fragmented Old World rain forests, supporting both viral specialists and generalists within multi-host assemblages [4]. Within the ‘mind's eye’, unlike multicellular organisms and bacterial/fungal colonies, viral existence and growth are typically unseen and often asymptomatic. ‘Out of sight, out of mind’, in the sense of the virome, is taken to an extreme from the perspective of neglect with most viruses in the primary host, unless frank disease and mortality are manifest. To the evolutionary biologist, all extant human viruses rose from more humble origins. Bereft from an appreciation of animal ecology, viruses, by definition, are neglected, in a circumspect fashion, and undergo a pattern of subtle repetition over time (Fig. 1 ). What began as a largely one-sided accident before animal domestication millennia ago proceeds to a ‘reverse-zoonosis’ paradigm, as expanding inter-connected human communities become the drivers back to other animals, often with disastrous consequences for threatened and endangered species [5].
Fig. 1.
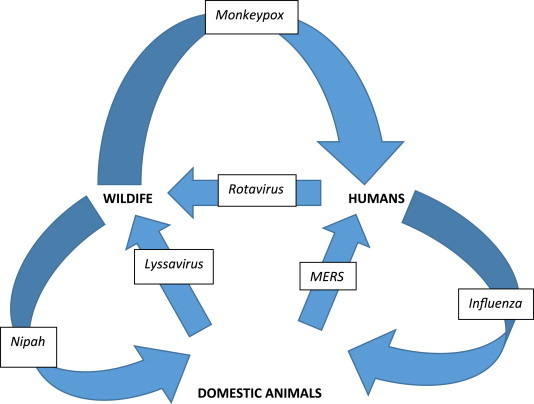
Simplified historical composite and evolution of zoonotic viral infectious threads from simple linear dead-end spillover cases from wildlife, to other eventual amplification into domesticated species, to development of human pandemic states and reverse dynamic spread back among other non-human wild or domestic species, using current examples (for example, although avian influenza may infect domestic poultry or humans, infected humans can also infect domestic animals; similarly, while lyssavirus reservoirs are found among wildlife, rabid domestic dogs can infect wild carnivores).
Critically ignored macroscopic attributes
Broadly speaking, any RNA or DNA virus transmitted to a human from a member of the Kingdom Animalia is open to partitioning as a viral zoonosis. Of the hundreds of potential viral candidates, what constitutes a ‘neglected viral zoonosis’ (NVZ)? To be considered a NVZ, a disease is not equivalent to being merely odd or rare, objectively, from the stand point of concentration, which does not diminish the public health concern for any malady, per se. Hence, as one example, obviously simian herpesvirus is a serious viral infection, but not a NVZ. Similarly, a NVZ does not necessarily mean that no diagnostics or vaccines or practical mitigation options exist or that little is known about pathogen epizootiology. In fact, ironically, many NVZ have pragmatic solutions for investigation, prevention and control—exacerbating the overall philosophy of neglect. Rather, operationally defined subjectively, NVZ should be associated with significant ‘negative’ attributes, including: substantive human morbidity and mortality; broad distribution; serious economic costs; selective opportunity to spread if ignored; a lack of concentrated long-term efforts focused at the source; and a disproportionate burden on the poor [6], [7]. So defined, most NVZ occur in developing countries with a subtropical to tropical occurrence.
To a finite extent, biomedical introspection related to entry, reception, replication, transcription, assembly and exit from one species to a human may provide a great deal of inference to various NVZ. However, beyond the descriptive ‘who, what, where, when and how’ of the original animal virion and the secondary human host, these qualitative insights will always be wanting to a greater epidemiological understanding of the ‘why things are’. Ultimately, besides an anatomical and physiological basis for a viral infection to transmit interspecifically, both pathogens and their reservoirs/vectors must exploit a complex anthropological, cultural, historical, political, psychological and social landscape that uniquely defines Homo sapiens, unlike any other animal population [8], [9], [10], [11], [12]. Some of these facets include the ecological benefits (and limitations) of living in groups as a social species, hunting and gathering behaviours and dietary preferences, myths and religious doctrines, health economics and many others, in a myriad of a homo-sociome memetic complex (HOSMEC). Taken together, such an introspective transdisciplinary approach serves to explain the ultimate success or failure of a zoonosis programme and the basis of clues as to why some NVZ collapse or persist. Moreover, in-depth critical utilization of the HOSMEC idea may provide answers as to why a NVZ may reach a certain critical tipping point to finally receive focused attention among a myriad of outcomes, because of humanitarian extension of a novel intervention from applied research or due in part from a perceived threat to impact non-enzootic developed countries (Table 1 ). The reality of NVZ in situ is not as simple as a random meeting between infectious non-human host X and susceptible human individual Y, but predicated among populations by HOSMEC Z + N factors in space and time, as defined broadly by the human–animal bond (Fig. 1).
Table 1.
Illustrations of various homo-sociome memetic complex (HOSMEC) factors, which may influence emergence and perpetuation of neglected viral zoonoses
| Factor | Example |
|---|---|
| Anthropological | Domestication of certain wild taxa based on recognized utility |
| Cultural | Perceptions of individual utility versus community animal ownership |
| Economic | Differential veterinary care/husbandry based on local value systems |
| Educational | Restriction to only certain groups based on multiple factors |
| Familial | Intrinsic recognition of pets and interactions as family members |
| Historical | Precedent of zoonosis control programmes irrespective of evidence |
| Legal | Animal welfare norms and protections or provisions as chattel |
| Occupational | Degree of engagement is a traditional inheritance or open to free choice |
| Philosophical | Dietary choices from opportunistic to strict veganism unrelated to health |
| Political | Leadership determination of agro-policies, from tribal to national |
| Religious | Interactions with particular species deemed taboo or unclean |
| Social | Age, caste or gender bias to focal interactions dealing with animals |
Using the HOSMEC concept as a backdrop, in this paper we concentrate upon several NVZ that meet our criteria for inclusion, discuss strategies for future prevention and control and draw upon the majority of supportive examples from the recent peer-reviewed literature as to missed opportunities and a way forward in the near term.
The incurable wound revisited as the major NVZ
Recently, one major NVZ selected for greater attention by the international community is human rabies transmitted by dogs, also known as canine rabies, embraced by the triumvirate of FAO (http://www.fao.org/news/story/en/item/198087/icode/), OIE (http://www.oie.int/animal-health-in-the-world/rabies-portal/) and WHO (http://www.who.int/neglected_diseases/WRD_rabies_2013/en/) as a valuable public good, in the mould of the One Health philosophy towards ‘No More Deaths From Rabies’.
A number of pervasive myths served to illustrate the HOSMEC spectrum as to why rabies languished as a NVZ (Table 2 ), despite being one of the oldest zoonoses and having the highest case fatality of any infectious disease. Simply put, a strong deliberate track record of primary evidence coalesced as counter HOSMEC facets to refute such arguments for programmatic obsolescence [13], [14], [15], [16], [17], [18], [19], [20], [21], [22], [23], [24]. Unfathomably, some may question the status of canine rabies as a NVZ, but the disease is a bona fide choice, considering that tens of thousands of people, mostly children, succumb annually, tens of millions of human exposures occur requiring intervention and all of the major tools for elimination were present by the mid-twentieth century [13], [14], [17]. By comparison with rinderpest, vaccination towards herd immunity, sensitive and specific diagnostics and the epidemiological knowledge to apply both to the field were comparable to an equally complex disease for decades [15], [18]. Oddly, canine rabies still exists in a diagnostic cycle of neglect, despite laboratory insights for confirmation since the late 1800s, and is the only infectious disease in which a routine diagnostic test applied to an animal directly determines the need for medical intervention in the bitten person [17]. Clearly, unlike beef, poultry, seafood and swine, dogs are not a conventional market commodity, even if they are still consumed in certain markets. As a result, rabies requires an inter-sectoral approach among professionals for ultimate success [13], [20], [23]. Inarguably, modern human and veterinary biological agents against rabies are pure, potent, safe, efficacious and inexpensive, considering the alternative outcome of a productive encephalitic infection [16], [17], [20]. Long-term lethal culling of dogs has not been demonstrated to be an ethically supported, ecologically based or economically sound strategy to control canine rabies [19], [20]. By contrast, in both the New and Old Worlds, herd immunity is achievable among dynamic canine populations [13], [15], [20]. Although a plethora of wildlife reservoirs maintain a diversity of other rabies virus variants, their existence in developed countries was not a barrier to canine rabies elimination and oral vaccination has proven to be a successful intervention towards a holistic programme approach, when mesocarnivore populations are determined to be important [21]. Although often used as an epidemiological ploy to explain programme failure or to save face, no true rabies virus carriers have been documented [22]. Substantively, rather than indefinite reliance upon outside economic aid, community engagement and social responsibility are a longer-term strategy for programme perpetuation [23]. Another HOSMEC facet to counter naysayers is a designated date for global action (http://rabiesalliance.org/world-rabies-day/) and the international partnering that has evolved among dedicated academic, government, industrial and non-governmental organization stakeholders, who champion the advocacy, communication, education and research towards canine rabies elimination as a realistic cause célèbre [24].
Table 2.
Common prevailing global rabies perceptions that perpetuate a cycle of neglect and programme failure
| Issues |
|---|
| Attitude – ‘It's only rabies!’ |
| Burden – ‘Few to no human fatalities measured’ |
| Monitoring – ‘Just not enough animal cases to justify concern’ |
| Responsibility – ‘Dogs are not a commodity’ |
| Prevention – ‘Local biologicals are ineffective’ |
| Control – ‘Culling will work for any outbreak’ |
| Epizootiology – ‘Uncontrollable stray dogs are impossible to resolve’ |
| Reservoirs – ‘Wildlife maintain infection cycles’ |
| Economics – ‘Vaccines are not affordable’ |
| Perpetuation – ‘Animals are true carriers’ |
| Engagement – ‘Communities simply do not care’ |
| Support – ‘No global partners exist’ |
Besides canine rabies as the epitome of a NVZ and the related HOSMEC applications as background narrative, in consideration of other potential viruses, a divergent ecological and evolutionary perspective can be assumed, as regards potential pathogens and vectors. That is, are there particular major animal hosts that are contenders for discussion? Within the vertebrates, there are no serious NVZ candidates among the fish, amphibians and reptiles. Arguably, with the exception of the ‘avian influenza mixing milieu’ (which is not truly neglected by our above subjective criteria and the considerable budgets available for basic and applied research), no major NVZ are based among the birds. Concentrating upon mammals, with degrees of genetic relationship closest to Homo sapiens, one might be tempted to consider viral pathogens associated with non-human primates. However, unless considering the evolutionary and epidemiological relationships of the simian retroviruses (notwithstanding the consequent radiations that led to human immunodeficiency virus) and their related viral spectrum (which also do not fit the criteria of NVZ), the non-human primates are twenty-first century evolutionary bottlenecks, geographically isolated and largely not in temporal/spatial proximity to most people. This limitation does not apply to other relevant and highly misunderstood mammals—the Chiroptera.
Bats and NVZ
Many of the recent NVZ have been due to human and domestic animal interactions with wildlife and of these, several prominent examples have been associated with bats (Table 3 ). Bats are not simply ‘winged rodents’, so one rationale to consider the bat-associated viral diseases as a single complex pertains to the biological attributes applied objectively to these species as a whole, in a special status [25]. Compared with human and domestic animal populations, wildlife have always been notoriously more difficult to study. Bats are no exception. As typically smaller, inconspicuous, volant animals, relatively less is known about this group than other mammalian taxa. As the second largest mammalian Order, with over 1000 species, their abundance, distribution, biodiversity and lifestyle collectively provide a complication when trying to infer overarching elements from epidemiological insights to prevention and control strategies over their collective virome, as exemplified by numerous NVZ [26], [27], [28], [29], [30], [31]. Besides prominent scientific neglect until the mid-twentieth century, bats have been reviled in many cultures as concerns the HOSMEC principle, despite their obvious utility as the only prominent nocturnal predators of agricultural insect pests and by the ecological services they provide through pollination and reforestation from seed dispersal, which is often forgotten when weighing zoonotic risks in a conservation biology perspective [32].
Table 3.
| Example | Taxa | Distribution | Comment |
|---|---|---|---|
| Lyssaviruses | Rhabdovirus | Global | Thousands of annual deaths |
| Ebola/Marburg | Filovirus | Africa, Asia | Pandemic potential? |
| Henipah | Paramyxovirus | Austral-Asia | Human–human possible |
| SARS/MERS | Coronavirus | Asia/Middle East | Human–human likely |
| Menangle | Paramyxovirus | Australia | Human infection, no deaths |
From a global and zoological perspective, the overwhelming majority of bats require no special consideration for human or domestic animal health. Rather, distribution of the few documented NVZ among the Chiroptera is highly uneven. Major genera appear somewhat restrictive based upon surveillance to date, including: Artibeus, Desmodus, Eidolon, Eptesicus, Hipposideros, Lasiurus, Lasionycteris, Myotis, Perimyotis, Pteropus, Rousettus and Tadarida. Interspecific transmission may be direct, especially in the case of the bat lyssaviruses (such as vampire-transmitted rabies), but often may involve an intermediate mammalian host for amplification: insectivorous bats and civets in the night markets of Asia for SARS; Old World bats and non-human primates and bushmeat exposures for Marburg/Ebola viruses; fruit bats and horses for Hendra virus; pteropid bats and swine for Nipah and Menagle viruses; various bat species and camels for Middle East respiratory syndrome [26], [27], [28], [29], [30], [31]. Besides long-term occupational alternatives for bushmeat acquisition, practices to limit the inappropriate harvesting and mixing of wild mammals for human consumption, curbs on habitat encroachment and degradation, among others, an intermediary link of domestic animals with the bat-associated NVZ offers a benchmark for development and application of relevant veterinary vaccines, as appropriate. Given these comparative points, and the recent flurry of attention over Ebola, a decade from now may be an appropriate time span to review the status of the filoviruses as a whole and their gradual rise to NVZ status and beyond, either towards ascendency approaching avian influenza in prominence or fading to obscurity with the other bat zoonoses.
Selected continent-jumping mosquito-transmitted NVZ: dengue, chikungunya and Zika viruses
Besides mammals, many viral arthropod-borne contenders meet the criteria of NVZ, considering the myriad of agents transmitted to humans by invertebrates. Among the Arthropoda, ticks and insects are the two major representatives, and of the latter, given the mobility offered by flight (like bats and birds), the mosquitoes are highly effective vectors. Currently, there are at least three different species of arboviruses (dengue virus (DENV), Chikungunya virus (CHIKV) and Zika virus), transmitted predominantly by common vectors, the Aedes mosquitoes, that present similar syndromes and produce rapidly advancing outbreaks, mainly in developing tropical countries. Regardless of the large number of potentially infected individuals and the massive burden these infections pose on local economies, they remain largely ignored. Moreover, despite the emphasis placed on these emerging diseases by organizations such as the WHO, these NVZ are continuing to evolve. Ensuing concomitant human outbreaks are becoming much more frequent. Historically operative and ecologically naive HOSMEC are partly to blame. Not the least of these are referential to the geographic foci of these NVZ and the forgotten opportunities presented by the management of malaria and yellow fever in developed countries, combined with the daily success offered by air conditioning on demand, window screens, insect repellents and environmental mitigation practiced in an ecologically sound philosophy of Western civilization, which naively assumes homogeny abroad.
Prominent arboviral taxa operative among the NVZ
As prominent viral examples, among the Flaviviridae, the genus Flavivirus includes at least 53 recognized species, of which many are human pathogens. These viral species cluster into ecologically distinguishable groups. Sequence analysis provides grouping into distinct lineages that correspond closely to the arthropod groups responsible for transmission. Additionally, the mosquito-borne flaviviruses can be grouped further into a Culex group and an Aedes group. The tick-borne flaviviruses (and the flaviviruses with no vector) cluster into another genetic group. Within the Aedes group are four important clusters: the yellow fever group; the Entebbe virus group; the DENV group (including DENV 1 to 4); and the Spondweni virus group, that includes Zika virus [33].
Recognized as a continuing global threat, more than 2.5 billion people are estimated to be at risk of DENV infection, with 975 million people living in urban areas in South-East Asia, the Pacific and the Americas [34]. Less known than DENV, Zika virus was first isolated in Uganda during 1947, with a natural transmission cycle involving mainly vectors of the Aedes genus and non-human primates, with occasional infection of humans. During 2007, Zika virus was responsible for an epidemic in the Federated States of Micronesia, including the island of Yap [35]. Thereafter, during 2013, the first cases were reported in French Polynesia. This recognition was followed by outbreaks in New Caledonia, the Cook Islands, Easter Island and now almost certainly will spread to other Pacific islands [36].
In contrast to the above Aedes-transmitted flaviviruses, CHIKV is a member of the family Togaviridae, genus Alphavirus. Within Africa, CHIKV circulates in an enzootic cycle involving forest-dwelling mosquitoes and non-human primates. The agent was first isolated in Tanzania during 1953 from the blood of a febrile person during a large outbreak of a disease characterized by severe fever and crippling joint pains (giving CHIKV its name, literally ‘bent over’). Out of Africa, CHIKV was isolated during 1958 from a patient in Bangkok. Sporadic outbreaks were reported in numerous African and Asian countries for decades and then during April 2005, CHIKV was confirmed as the cause of an epidemic of dengue-like illness on the Comoros islands off the east coast of northern Mozambique. Shortly after this outbreak, the disease was reported on Mayotte, Mauritius and the French island of La Réunion [37]. During December 2003, CHIKV was detected in the French West Indies. The virus has spread rapidly to many other Caribbean islands, as well as to South America, and poses a significant threat to both North America and Europe [36].
Morbidity associated with Aedes-transmitted arboviruses
The DENVs produce several syndromes that can be affected by host age and immunological status. Most children experience subclinical infection or mild febrile syndromes. Symptoms and signs include fever, malaise, headache, body pains (break-bone fever) and a rash. Some cases may progress to severe dengue in which there can be bleeding, thrombocytopenia, ascites, continuous abdominal pain, restlessness, persistent vomiting and a sudden reduction in temperature. Severe dengue is more likely to occur when a patient is infected with a second or third case of dengue. There can be substantial differences in the pathogenicity of different strains of virus as well as individual differences in the susceptibility of patients. Previous infection with DENV may be protective to some extent or it may exacerbate the condition [34].
Today, CHIKV is considered to be one of the members of the arthritic alphaviruses, most of which are in the Semliki Forest group. These include Ross River virus, O'nyong-nyong virus and Mayaro virus. The most frequent signs and symptoms include fever, rash, myalgia with arthralgia or arthritis. There may also be fatigue, tenosynovitis, headache, nausea, oedema, vomiting, conjunctivitis and occasional bleeding gums and epistaxis. As apparent to practitioners, a clinical diagnosis of CHIKV infection is complicated, because of the very similar presentation to DENV infection [38].
Similarly, the clinical presentation of Zika virus infection is often described as ‘dengue-like’ with signs and symptoms including arthralgia, oedema of extremities, mild fever, headache, retro-orbital pain, conjunctival hyperaemia and maculopapular rashes, usually spreading downward from the face to the limbs and is frequently pruritic. Additionally, there may be vertigo, myalgia and digestive disorders [39]. During a recent outbreak in French Polynesia, a cluster of patients presented with a Guillain-Barré syndrome that appeared to correlate with disease occurrence [40].
Evolution of the arboviral NVZ
Unlike a well known NVZ such as rabies, one of the basic HOSMEC issues associated with DENV, CHIKV and Zika virus are the previous historical limitations to the subtropical regions, where most Europeans and North Americans never encountered them. In contrast to the other two arboviral NVZ, the DENVs have a long history of human infection, with documented cases of a disease resembling dengue (meaning cramps, seizures, or stiff-gait) as early as the third century. In the late 1700s there appears to have been a widespread geographical distribution of dengue that coincided with the increasing global commerce aided by sailing ships. A second series of dengue-like pandemics were subsequently reported from Africa to India to Oceania and the Americas from 1823 to 1916. Following the Second World War and the dramatic increase in urbanization, with inadequate housing and water distribution systems, the vector Aedes aegypti reached high densities, facilitating the efficient transmission of DENV. Four antigenically distinct serotypes of DENV are recognized at present. In all four cases the transmission of DENV includes a sylvatic, enzootic cycle between non-human primates and arboreal mosquitoes of the genus Aedes. This has led to distinct emergence events presumably involving host shifts from arboreal Aedes mosquitoes to Aedes albopictus and later to Aedes aegypti [41].
As the four serotypes of DENV have further evolved, it appears reasonable to regard these as four distinct viral populations with unique and overlapping geographical distributions. Furthermore, within each serotype, distinct clusters of viral strains have been recognized. This was initially achieved through RNA fingerprinting and with the advent of RT-PCR and Sanger sequencing, with additional information on envelope gene sequences, substantial amounts of epidemiological data were produced. With the introduction of next-generation methods, full-length sequencing is rapidly replacing the characterization of single genes and provides a better understanding of the distribution of dynamic quasispecies [41].
The ancestral sylvatic DENV transmission cycles still exist, maintained in the forests of South East Asia and West Africa. Conditions required for the adaptation of these viruses to humans remain operative. Continued spillover and emergence into human populations is likely to occur with strains that are substantially different to those presently circulating, giving rise to further epidemics [42].
In contrast to DENV, CHIKV in Africa evolved with two enzootic lineages. Estimates place introduction into Asia approximately 70–90 years ago. The recent Indian Ocean and Indian subcontinent introductions of CHIKV emerged independently from the east coast of Africa [43]. Currently, three genotypes of CHIKV are recognized as Asian, East Central South African and the West African. The isolates recognized recently in the Caribbean are within the Asian genotype and closely related to strains recently isolated in China and the Philippines. The two isolates recently obtained from Yap, the British Virgin Islands, China and the Philippines form a separate clade within the Asian genotype [44].
Paralleling the translocations of DENV and CHIKV, there have been at least two independent introductions of Zika virus from the Zika Forest region of Uganda into West Africa during the 1950–60s. During the 1940s, Zika virus was introduced into Malaysia and then onto the Pacific Islands [35]. A high serological prevalence was noted in Nigeria during 1979 and in the late 1970s in Indonesia a prevalence of 7.1% was noted. An epidemic has been described in French Polynesia since October 2013 [39]. Following this latter focus, there were subsequent outbreaks in New Caledonia, the Cook Islands and Easter Island [40]. As Zika virus is transmitted by the same mosquitoes as DENV and CHIKV and the clinical signs are relatively similar, there could be substantial incursions in countries where these viruses are already endemic (such as in the Caribbean region), without being readily detected, given the paucity of laboratory-based surveillance.
Economic impacts
Accurate and comprehensive studies that outline the economic and health burdens of most NVZ are relatively sparse, but appear dire when attempted. For example, an estimate of mean health costs of hospitalization in South America for a case of DENV was ∼US$571. One estimate of the annual global burden was 750 000 disability-adjusted life-years. An extrapolation of the annual cost of DENV across the Americas (2000–2007) was US$2,149.8 [45], [46].
In many countries, the burden of DENV is under-reported, as a number of cases are subclinical and in addition the resources available for collecting data are often restricted to government health workers and hospitals. Many of the patients present to private practitioners, who do not have any incentive to report the cases. As CHIKV may have a lower percentage of subclinical cases, the reporting may reflect a slightly more accurate assessment. Unfortunately, most of the estimates fail to take into account the cost of providing vector monitoring and control, and to address all of the additional costs, such as education and lost work days. Significantly, many of the countries where these viruses are endemic, or countries facing epidemics, derive much of their income from tourism. Regardless of the direct impact upon the service sector itself, if a country earns a reputation as an unsafe destination, this would have devastating effects on the tourism industry overall, as a notable HOSMEC incentive towards under-reporting.
By comparison, in the CHIKV outbreak on the island of La Réunion, more than one-third of the 800 000 inhabitants were infected, with an estimated productivity loss of €17.4 million and medical costs of €43.9 million. However, the cost of sequential or concurrent infections with DENV, CHIKV and Zika viruses is unknown [47]. Given the clinical impact, there is an urgent need for additional comprehensive research that reflects the true cost of these collective NVZ and to support vaccination and control programmes [45].
NVZ extrapolation—transmission by blood transfusions
If a person is affected unknowingly by a NVZ, organs, tissues or fluids may be infectious and pose risks for an opportunistic infection. For example, blood collected from arboviraemic patients could be responsible for transfusion-transmitted infections. A survey of Zika virus in donated blood samples in French Polynesia found 2.8% reactors. High rates of asymptomatic infections are a major concern and arbovirus transfusion-transmitted infections have been reported for West Nile virus, DENV and CHIKV. During the outbreaks of CHIKV in Italy during 2007 and in La Réunion 2005–2007, products were imported from blood banks elsewhere. However, this is not an option in most affected developing countries [48].
Threats to non-endemic areas
One of the major concerns behind an interest in NVZ is the risk these aetiological agents and vectors pose to unaffected areas. Large areas of southern Europe, the southern USA and the northern parts of Australia, as well as the subtropical regions of southern Africa and South America, have been colonized by Aedes aegypti and Aedes albopictus is continuing to invade a number of these areas. Aedes albopictus is able to survive at lower temperatures than Aedes aegypti and is less dependent on the urban environment with man-made containers. With the rapid movement of viraemic individuals and the potential of subclinical or unapparent infections, the possibility of transmission of one or all of these viruses is relatively high. If there is a stable population of a suitable vector, autochthonous transmission remains a genuine possibility [36], [49].
Serious prospects for human vaccination?
Unlike rabies, development of safe, efficacious and affordable human vaccines for the arboviral NVZ have lagged. However, efforts to develop effective DENV vaccines have significantly increased over the last decade, and a wide range of vaccine approaches is under current evaluation, largely due to military interests [50]. Regardless of initial proof of concept, such vaccines will need to be tailored to the national requirement of the target countries. Measurement of efficacy may be difficult, especially where subclinical infections are common and the infrastructure for monitoring infection is limited. Antibody-dependent enhancement must not occur in the ideal vaccine candidate and clinical trials must closely monitor the appearance of any such adverse events. To implement any vaccination programme, appropriate cost–benefit analyses are highly desirable. Despite their inherent importance, very few such analyses have been produced to date [46].
Effective vaccines for CHIKV are much more limited than DENV, with no prospect for rapid implementation. Similarly, considering that Zika virus only affects individuals in developing countries, the incentive for any investment in vaccine studies is almost non-existent.
Continued emergence of highly opportunistic vector species
New sylvatic cycles frequently rely on host shifts and vector species that are not currently involved in the transmission and maintenance of these NVZ. Bats, mesocarnivores, rodents and arthropod vectors are highly successful as invasive taxa. These vector species may have the potential to be transferred to other locations, where they can thrive. For example, in some limited areas, there have been additional species responsible for the transmission of CHIKV and Zika viruses. Relocation of these vector species may be likely, especially with impending climate change scenarios. Given the reality of modern globalization, these vector–virus translocations are becoming a routine event.
Challenges in the monitoring, prevention and control of NVZ
A holistic appreciation of the ecology of infectious diseases should provide solutions towards practical interventions. Clearly, from the rapid and ongoing increase in prevalence of these highlighted neglected taxa, control measures presently being used are ineffective. However, in some resource-poor environments, it may be possible to at least monitor the presence and relative abundance of insect vectors as a start, if technical staff with the requisite knowledge, skills and abilities are nurtured appropriately. Nevertheless, implementation of effective control programmes appears to be beyond the scope of the present resources. Some future innovative control programmes may have the potential to make a difference in regions where only one mosquito is the sole vector, but similar approaches have yet to be successful with all potential vectors and NVZ pathogens.
Prevention by vaccination would require a high level of human herd immunity and is likely to be only successful if applied as part of a very integrated approach using vector control and education to supplement such vaccination.
Vector management is an integral part of the control of any NVZ. Significantly, during the 1960s, a campaign to eradicate Aedes aegypti from South America had a dramatic influence on the transmission of DENV. When this campaign was abandoned, there was a massive increase in vector abundance that contributed significantly to the high rates of DENV infection today [51]. Even greater challenges are posed with Aedes albopictus [52], [53]. An integrated One Health programme should engage community participation in vector control, including elimination of larvae by reducing potential breeding containers and treatment with larvicides, insecticidal spraying, especially targeted spraying in potential resting areas (fogging with large amounts of insecticide has minimal effect when it fails to reach the target insects) and biological control using larvivorous organisms or bioinsecticides. There is potential opportunity in environmental release of genetically modified insects, including those infected with Wolbachia species. However, this has yet to be developed for Aedes albopictus [52], [53]. Additionally, the implementation of autocidal gravid ovitraps may substantially reduce the populations of vectors [54]. Post-application, vector monitoring is an extremely useful tool and is absolutely essential to direct effective control programmes and to gauge success.
Conclusions
Viral zoonoses span a gamut from widespread global distributions and dramatic human mortality to more chronic perpetuation and existence in relative obscurity. Rabies, the bat-associated viral zoonoses and the arthropod-borne viruses transmitted by Aedes mosquitoes provide a contrasting panoramic cross-section of the opportunities and challenges posed by such concomitant NVZ in economically disadvantaged tropical regions. By historical illustration, as one of the oldest appreciated infectious diseases, canine-transmitted rabies is the epitome of a NVZ, with a strong probability for continued success in substantial prevention, control and elimination over the next 10–20 years. In contrast, although the bat-associated zoonotic viruses have received increasing attention over the last decade and appear to be emergent for similar reasons regardless of viral pathogen, there are no major options for vaccine intervention at the wildlife source, in comparison with canine rabies. Considering arboviruses, the DENV, CHIKV and Zika viruses represent a similar clinical syndrome in toto and may obfuscate surveillance and appropriate medical response, yet more simplified monitoring systems and mitigation focused upon the primary mosquito vectors should be advantageous, in lieu of available human vaccination.
Besides these selected examples of NVZ, additional salient representatives could have included Rift Valley fever, the rodent-associated zoonoses such as the hantaviruses, hepatitis E virus, tick-borne encephalitis and multiple others. Nevertheless, regardless of pathogen or vector, consideration of all of the NVZ (as well as other non-viral agents) in a One Health context is the best option for a community-based approach to the long-term detection of, response to and mitigation of zoonoses as an integrated unit, rather than as an individual agent or disease [55]. Finally, beyond the classical microbiological and biomedical aspects of human, veterinary and conservation medicine, the basic underpinnings to these neglected entities are often more deeply rooted in a complex suite of anthropological, cultural, religious and social factors, which also may persist or change abruptly over time as the agents and vectors in kind. Failing to appreciate such a transdisciplinary HOSMEC milieu may do little to alter the history associated in the complex causality at the root of zoonotic disease emergence and perpetuation, within the evolutionary play on an increasingly small global ecological stage.
Transparency declaration
The authors declare that they have no conflicts of interest.
Acknowledgements
We thank our many colleagues around the global village, in academia, government, industry and non-governmental organizations who have helped contribute substantially to the surveillance, diagnosis, ecology, prevention, control and treatment of NVZ over the past several decades in this view of life—you know who you are …!
References
- 1.Cutler S.J., Fooks A.R., van der Poel W.H. Public health threat of new, reemerging, and neglected zoonoses in the industrialized world. Emerg Infect Dis. 2010;16:1–7. doi: 10.3201/eid1601.081467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Daugherty M.D., Malik H.S. Rules of engagement: molecular insights from host-virus arms races. Annu Rev Genet. 2012;46:677–700. doi: 10.1146/annurev-genet-110711-155522. [DOI] [PubMed] [Google Scholar]
- 3.Domingo E., Escarmís C., Sevilla N., Moya A., Elena S.F., Quer J. Basic concepts in RNA virus evolution. FASEB J. 1996;10:859–864. doi: 10.1096/fasebj.10.8.8666162. [DOI] [PubMed] [Google Scholar]
- 4.Elena S.F., Agudelo-Romero P., Lalić J. The evolution of viruses in multi-host fitness landscapes. Open Virol J. 2009;3:1–6. doi: 10.2174/1874357900903010001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Messenger A.M., Barnes A.N., Gray G.C. Reverse zoonotic disease transmission (zooanthroponosis): a systematic review of seldom-documented human biological threats to animals. PLoS One. 2014;9:e89055. doi: 10.1371/journal.pone.0089055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Seimenis A. Zoonoses and poverty—a long road to the alleviation of suffering. Vet Ital. 2012;48:5–13. [PubMed] [Google Scholar]
- 7.World Health Organization Research priorities for zoonoses and marginalized infections. World Health Organ Tech Rep Ser. 2012;971:ix–xi. 1–119. [PubMed] [Google Scholar]
- 8.Atran S. Folk biology and the anthropology of science: cognitive universals and cultural particulars. Behav Brain Sci. 1998;21:547–569. doi: 10.1017/s0140525x98001277. [DOI] [PubMed] [Google Scholar]
- 9.Wallace R., Wallace R.G. Organisms, organizations and interactions: an information theory approach to biocultural evolution. Biosystems. 1999;51:101–119. doi: 10.1016/s0303-2647(99)00023-4. [DOI] [PubMed] [Google Scholar]
- 10.Kamhi A.G. A meme's eye view of speech-language pathology. Lang Speech Hear Serv Sch. 2004;35:105–111. doi: 10.1044/0161-1461(2004/012). [DOI] [PubMed] [Google Scholar]
- 11.Cardoso G.C., Atwell J.W. Directional cultural change by modification and replacement of memes. Evolution. 2011;65:295–300. doi: 10.1111/j.1558-5646.2010.01102.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.McNamara A. Can we measure memes? Front Evol Neurosci. 2011;3:1. doi: 10.3389/fnevo.2011.00001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lankester F., Hampson K., Lembo T., Palmer G., Taylor L., Cleaveland S. Implementing Pasteur's vision for rabies elimination. Science. 2014;345:1562–1564. doi: 10.1126/science.1256306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Knobel D.L., Cleaveland S., Coleman P.G., Fèvre E.M., Meltzer M.I., Miranda M.E. Re-evaluating the burden of rabies in Africa and Asia. Bull World Health Organ. 2005;83:360–368. [PMC free article] [PubMed] [Google Scholar]
- 15.Zinsstag J., Dürr S., Penny M.A., Mindekem R., Roth F., Menendez Gonzalez S. Transmission dynamics and economics of rabies control in dogs and humans in an African city. Proc Natl Acad Sci U S A. 2009;106:14996–15001. doi: 10.1073/pnas.0904740106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Anderson A., Shwiff S.A. The cost of canine rabies on four continents. Transbound Emerg Dis. 2013 doi: 10.1111/tbed.12168. [DOI] [PubMed] [Google Scholar]
- 17.Meslin F.X., Briggs D.J. Eliminating canine rabies, the principal source of human infection: what will it take? Antiviral Res. 2013;98:291–296. doi: 10.1016/j.antiviral.2013.03.011. [DOI] [PubMed] [Google Scholar]
- 18.Townsend S.E., Lembo T., Cleaveland S., Meslin F.X., Miranda M.E., Putra A.A. Surveillance guidelines for disease elimination: a case study of canine rabies. Comp Immunol Microbiol Infect Dis. 2013;6:249–261. doi: 10.1016/j.cimid.2012.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Townsend S.E., Sumantra I.P., Pudjiatmoko, Bagus G.N., Brum E., Cleaveland S. Designing programs for eliminating canine rabies from islands: Bali, Indonesia as a case study. PLoS Negl Trop Dis. 2013;7:e2372. doi: 10.1371/journal.pntd.0002372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Vigilato M.A., Clavijo A., Knobl T., Silva H.M., Cosivi O., Schneider M.C. Progress towards eliminating canine rabies: policies and perspectives from Latin America and the Caribbean. Philos Trans R Soc Lond B Biol Sci. 2013;368:20120143. doi: 10.1098/rstb.2012.0143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Slate D., Algeo T.P., Nelson K.M., Chipman R.B., Donovan D., Blanton J.D. Oral rabies vaccination in North America: opportunities, complexities, and challenges. PLoS Negl Trop Dis. 2009;3:e549. doi: 10.1371/journal.pntd.0000549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhang Y.Z., Fu Z.F., Wang D.M., Zhou J.Z., Wang Z.X., Lv T.F. Investigation of the role of healthy dogs as potential carriers of rabies virus. Vector Borne Zoonotic Dis. 2008;8:313–319. doi: 10.1089/vbz.2007.0209. [DOI] [PubMed] [Google Scholar]
- 23.Lapiz S.M., Miranda M.E., Garcia R.G., Daguro L.I., Paman M.D., Madrinan F.P. Implementation of an intersectoral program to eliminate human and canine rabies: the Bohol Rabies Prevention and Elimination Project. PLoS Negl Trop Dis. 2012;6:e1891. doi: 10.1371/journal.pntd.0001891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lembo T., Attlan M., Bourhy H., Cleaveland S., Costa P., de Balogh K. Renewed global partnerships and redesigned roadmaps for rabies prevention and control. Vet Med Int. 2011;2011:923149. doi: 10.4061/2011/923149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Luis A.D., Hayman D.T., O'Shea T.J., Cryan P.M., Gilbert A.T., Pulliam J.R. A comparison of bats and rodents as reservoirs of zoonotic viruses: are bats special? Proc Biol Sci. 2013;280:20122753. doi: 10.1098/rspb.2012.2753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Banyard A.C., Evans J.S., Luo T.R., Fooks A.R. Lyssaviruses and bats: emergence and zoonotic threat. Viruses. 2014;6:2974–2990. doi: 10.3390/v6082974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Johnson N., Aréchiga-Ceballos N., Aguilar-Setien A. Vampire bat rabies: ecology, epidemiology and control. Viruses. 2014;6:1911–1928. doi: 10.3390/v6051911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Olival K.J., Hayman D.T. Filoviruses in bats: current knowledge and future directions. Viruses. 2014;6:1759–1788. doi: 10.3390/v6041759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Raj V.S., Osterhaus A.D., Fouchier R.A., Haagmans B.L. MERS: emergence of a novel human coronavirus. Curr Opin Virol. 2014;5:58–62. doi: 10.1016/j.coviro.2014.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hahn M.B., Gurley E.S., Epstein J.H., Islam M.S., Patz J.A., Daszak P. The role of landscape composition and configuration on Pteropus giganteus roosting ecology and Nipah virus spillover risk in Bangladesh. Am J Trop Med Hyg. 2014;90:247–255. doi: 10.4269/ajtmh.13-0256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Barr J.A., Smith C., Marsh G.A., Field H., Wang L.F. Evidence of bat origin for Menangle virus, a zoonotic paramyxovirus first isolated from diseased pigs. J Gen Virol. 2012;93(Pt 12):2590–2594. doi: 10.1099/vir.0.045385-0. [DOI] [PubMed] [Google Scholar]
- 32.Wood J.L., Leach M., Waldman L., Macgregor H., Fooks A.R., Jones K.E. A framework for the study of zoonotic disease emergence and its drivers: spillover of bat pathogens as a case study. Philos Trans R Soc Lond B Biol Sci. 2012;367:2881–2892. doi: 10.1098/rstb.2012.0228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Grard G., Moureau G., Charrel R.N., Holmes E.C., Gould E.A., de Lamballerie X. Genomics and evolution of Aedes-borne flaviviruses. J Gen Virol. 2010;91:87–94. doi: 10.1099/vir.0.014506-0. [DOI] [PubMed] [Google Scholar]
- 34.Guzman M.G., Halstead S.B., Artsob H., Buchy P., Farrar J., Gubler D.J. Dengue: a continuing global threat. Nat Rev Microbiol. 2010;8:S7–S16. doi: 10.1038/nrmicro2460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Faye O., Freire C.C., Iamarino A., Faye O., de Oliveira J.V., Diallo M. Molecular evolution of Zika virus during its emergence in the 20(th) century. PLoS Negl Trop Dis. 2014;8:e2636. doi: 10.1371/journal.pntd.0002636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mansuy J.M., Grouteau E., Mengelle C., Claudet I., Izopet J. Chikungunya in the Caribbean–threat for Europe. Emerg Infect Dis. 2014;20:1423–1425. doi: 10.3201/eid2008.140650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Morrison T.E. Reemergence of chikungunya virus. J Virol. 2014;88:11644–11647. doi: 10.1128/JVI.01432-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Suhrbier A., Jaffar-Bandjee M.C., Gasque P. Arthritogenic alphaviruses—an overview. Nat Rev Rheumatol. 2012;8:42–49. doi: 10.1038/nrrheum.2012.64. [DOI] [PubMed] [Google Scholar]
- 39.Ioos S., Mallet H.P., Leparc Goffart I., Gauthier V., Cardoso T., Herida M. Current Zika virus epidemiology and recent epidemics. Med Mal Infect. 2014;44:302–307. doi: 10.1016/j.medmal.2014.04.008. [DOI] [PubMed] [Google Scholar]
- 40.Musso D., Nilles E.J., Cao-Lormeau V.M. Rapid spread of emerging Zika virus in the Pacific area. Clin Microbiol Infect. 2014 Oct;20(10):O595–O596. doi: 10.1111/1469-0691.12707. PubMed PMID: 24909208. [DOI] [PubMed] [Google Scholar]
- 41.Weaver S.C., Vasilakis N. Molecular evolution of dengue viruses: contributions of phylogenetics to understanding the history and epidemiology of the preeminent arboviral disease. Infect Genet Evol. 2009;9:523–540. doi: 10.1016/j.meegid.2009.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Vasilakis N., Cardosa J., Hanley K.A., Holmes E.C., Weaver S.C. Fever from the forest: prospects for the continued emergence of sylvatic dengue virus and its impact on public health. Nat Rev Microbiol. 2011;9:532–541. doi: 10.1038/nrmicro2595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Volk S.M., Chen R., Tsetsarkin K.A., Adams A.P., Garcia T.I., Sall A.A. Genome-scale phylogenetic analyses of chikungunya virus reveal independent emergences of recent epidemics and various evolutionary rates. J Virol. 2010;84:6497–6504. doi: 10.1128/JVI.01603-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lanciotti R.S., Valadere A.M. Transcontinental movement of Asian genotype chikungunya virus. Emerg Infect Dis. 2014;20:1400–1402. doi: 10.3201/eid2008.140268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Beatty M.E., Beutels P., Meltzer M.I., Shepard D.S., Hombach J., Hutubessy R. Health economics of dengue: a systematic literature review and expert panel's assessment. Am J Trop Med Hyg. 2011;84:473–488. doi: 10.4269/ajtmh.2011.10-0521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lam S.K., Burke D., Capeding M.R., Chong C.K., Coudeville L., Farrar J. Preparing for introduction of a dengue vaccine: recommendations from the 1st Dengue v2V Asia-Pacific Meeting. Vaccine. 2011;29:9417–9422. doi: 10.1016/j.vaccine.2011.08.047. [DOI] [PubMed] [Google Scholar]
- 47.Roth A., Hoy D., Horwood P.F., Ropa B., Hancock T., Guillaumot L. Preparedness for threat of chikungunya in the Pacific. Emerg Infect Dis. 2014 Aug;20(8) doi: 10.3201/eid2008.130696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Musso D., Nhan T., Robin E., Roche C., Bierlaire D., Zisou K. Potential for Zika virus transmission through blood transfusion demonstrated during an outbreak in French Polynesia, November 2013 to February 2014. Euro Surveill. 2014 Apr 10;19(14) doi: 10.2807/1560-7917.es2014.19.14.20761. pii: 20761. [DOI] [PubMed] [Google Scholar]
- 49.Grange L., Simon-Loriere E., Sakuntabhai A., Gresh L., Paul R., Harris E. Epidemiological risk factors associated with high global frequency of inapparent dengue virus infections. Front Immunol. 2014;5:280. doi: 10.3389/fimmu.2014.00280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Thisyakorn U., Thisyakorn C. Latest developments and future directions in dengue vaccines. Ther Adv Vaccines. 2014;2:3–9. doi: 10.1177/2051013613507862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Gubler D.J. The changing epidemiology of yellow fever and dengue, 1900 to 2003: full circle? Comp Immunol Microbiol Infect Dis. 2004;27:319–330. doi: 10.1016/j.cimid.2004.03.013. [DOI] [PubMed] [Google Scholar]
- 52.Bonizzoni M., Gasperi G., Chen X., James A.A. The invasive mosquito species Aedes albopictus: current knowledge and future perspectives. Trends Parasitol. 2013;29:460–468. doi: 10.1016/j.pt.2013.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Paupy C., Delatte H., Bagny L., Corbel V., Fontenille D. Aedes albopictus, an arbovirus vector: from the darkness to the light. Microbes Infect. 2009;11:1177–1185. doi: 10.1016/j.micinf.2009.05.005. [DOI] [PubMed] [Google Scholar]
- 54.Barrera R., Amador M., Acevedo V., Hemme R.R., Felix G. Sustained, area-wide control of Aedes aegypti using CDC autocidal gravid ovitraps. Am J Trop Med Hyg. 2014 Dec;91(6):1269–1276. doi: 10.4269/ajtmh.14-0426. pii:14–0426. PubMed PMID: 25223937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Rist C.L., Arriola C.S., Rubin C. Prioritizing zoonoses: a proposed one health tool for collaborative decision-making. PLoS One. 2014;9:e109986. doi: 10.1371/journal.pone.0109986. [DOI] [PMC free article] [PubMed] [Google Scholar]