

Abstract
Sarcoids are the most prevalent equine skin tumours and remain a therapeutic challenge due to their differing clinical morphology, local aggressive behaviour, and high recurrence following surgical treatment. In vitro, sarcoid derived fibroblasts are invasive and express matrix metalloproteinase (MMP) -1, -2 and -9. It was hypothesised that the MMPs produced by neoplastic cells play a role in both their local invasiveness and interaction with the overlying epidermis (picket fence formation). The objective of this morphological study was to investigate the local behaviour and in situ MMP expression pattern in sarcoids of different clinical types. A total of 43 surgically excised sarcoids were examined by histology, immunohistology for the expression of MMP-1, -2 and -9, and transmission electron microscopy.
Regardless of the clinical type, sarcoids showed local invasion of the dermis and damage to the basement membrane in areas of interaction with the epidermis. This was associated with MMP-1 expression in both neoplastic cells and epidermis. The results suggest a link between MMP-1 expression and the local aggressiveness of sarcoids regardless of the clinical type.
Keywords: Equine sarcoid, Matrix metalloproteinases, Dermal invasion, Picket fence formation, Basement membrane
Introduction
Sarcoids are the most common equine skin tumours worldwide. A prevalence of between 1% and 8% and a predilection for younger animals have been reported (Torrontegui, Reid, 1994, Knottenbelt, 2005). Sarcoids develop after inoculation of fibroblasts with bovine papillomavirus types 1 and 2 (BPV-1, -2) and viral transcription (Bogaert et al, 2010, Hartl et al, 2011).
Sarcoids can typically be recognised clinically and histopathologically (Marti et al, 1993, Knottenbelt, 2005). With inconclusive histopathological features, PCR from superficial swabs, skin scrapings or the tumour mass can be helpful (Martens et al, 2000, Martens et al, 2001a). Six clinical types with individual growth characteristics and local behaviour have been described. These include occult, verrucose (‘warty’), nodular, fibroblastic, mixed and malignant forms (Knottenbelt, 2005). Focal dermal accumulations of fibroblastoid cells, usually in association with hyperplasia, hyperkeratosis and rete peg formation (RPF) of the overlying epidermis are typical histological features. Neoplastic cells often show ‘picket fence’ alignment along the epidermis and are generally locally invasive (Marti et al, 1993, Martens et al, 2000, Scott, Miller, 2003). The latter possibly accounts for the high recurrence rate of up to 50%, and the progression from milder forms to the more aggressive types (Marti et al, 1993, Martens et al, 2001b, Knottenbelt, 2005).
BPV alters the expression of several genes in transformed fibroblasts. Upregulation of the chemokine CXCL5 indicates an inflammatory response which is assumed to facilitate sarcoid growth/invasion. Concurrent downregulation of immune response genes, such as TLR-4 and MHC-I, most likely enables viral infection and persistence (Yuan et al, 2008a, Yuan et al, 2010a). Interestingly, matrix metalloproteinase (MMP) -1 and -9 are also markedly up regulated. MMPs may be responsible for tumour invasiveness since their expression and BPV-1 infection supports fibroblast invasion in vitro (Yuan et al, 2008a, Yuan et al, 2010b). We aimed to assess local tumour behaviour and in vivo MMP expression in naturally occurring sarcoids using histology, immunohistological detection of MMP-1, -2 and -9, and transmission electron microscopy (TEM).
Materials and methods
Animals and tissues
Forty-three naturally occurring surgically excised sarcoids from 25 horses (13 geldings, 4 stallions, 4 mares, 4 unknown gender) of various breeds and ages (2–21 years; average 9.6 years) were studied. The tumours were from various anatomical sites including the face, abdominal wall, axilla and medial thigh. None had been subjected to previous treatment. Individual lesions were classified clinically (Knottenbelt, 2005) and were histologically confirmed.
Immediately after surgical excision representative samples were fixed in 10% non-buffered formalin for 1–3 days and routinely paraffin wax embedded. From selected specimens, a portion was trimmed into 1–2 mm thick slices and fixed in 4% paraformaldehyde (pH 7.4) with 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer at 4 °C for TEM. Control skin samples from two horses euthanased for reasons other than skin disease, were similarly prepared.
Histology and immunohistology
Consecutive sections (3–5 µm) were prepared from paraffin wax embedded specimens and stained with haematoxylin and eosin (HE), Gomori Trichrome stain and Periodic Acid Schiff (PAS) reaction to highlight collagen and the epidermal basement membrane, respectively.
Immunohistology was used to demonstrate MMP-1, -2 and -9 and, in selected cases, cytokeratins (to highlight any potential tumour cell invasion into the epidermis), using cross reacting antibodies: rabbit anti-human MMP-1 (Ab-6), mouse anti-human APMA-activated native MMP-2 (clone A-Gel VC2), and mouse anti-human native MMP-9 (clone IIA5) (Thermo Fisher Scientific Anatomic Pathology), all reacting with both the inactive and the active form of the enzymes, and mouse anti-human pan-cytokeratin (clone AE1/AE3; Dako, Glostrup). The peroxidase anti-peroxidase method with antigen retrieval in heated citrate buffer (pH 6.0; MMP-1, MMP-9) or protease (cytokeratin) and the horseradish peroxidase method (Envision; Dako) without antigen retrieval (MMP-2) were applied (Kipar et al, 1995, Kipar et al, 2005). Antibody binding was visualised with diaminobenzidine as chromogen, followed by Papanicolaou's haematoxylin counterstain.
Consecutive sections incubated with TBS-Tween and/or a non-reacting antibody against feline coronavirus and/or Toxoplasma gondii instead of the primary antibody served as negative controls. Constitutive MMP expression in the normal skin was assessed on the two control skin specimens stained for the MMPs. A biopsy from an inflamed skin wound served as positive control for the markers.
Grading and statistical analysis
For MMP-1, the staining intensity of the neoplastic cells was scored over a scale from faint (+; 0.5) to strong (+++; 3). Where staining intensity varied between tumour areas, an average was calculated from the range of scores (Table 1 ). The association between MMP-1 expression intensity and sarcoid clinical type was analysed using the Kruskal–Wallis test since the data were not normally distributed and there was no appropriate transformation. The Mann–Whitney test was used to compare the sarcoid types pair-wise and the associations of MMP-1 expression with histopathological findings (picket fence formation [PFF], epidermal hyperplasia [EH], RPF). A Bonferroni correction was applied post-hoc for the three pairwise comparisons. The two types only existing as single samples (mixed and occult) were excluded from the analysis. Stata 13 (StataCorp) software was used for statistical analyses.
Table 1.
(A) Sarcoid cases with clinical type, relevant histological features and intensity of MMP-1 expression. (B) Summary results of the examination for the presence of relevant histopathological features of the epidermis and its interaction with the neoplastic cells in the different sarcoid types.
| A | ||||||
|---|---|---|---|---|---|---|
| Case No. | Sarcoid type | Morphological features | MMP-1 expression intensity | |||
| Ulceration | EH | RPF | PFF | |||
| 1 | Fibroblastic | Y | Y | Y | Y | ++/+++ |
| 2 | Fibroblastic | Y | Y | Y | Y | ++/+++ |
| 3 | Mixed | Y (chronic) | ENI | ENI | ENI | (+) – +++* |
| 4 | a.–c. Nodular | ENI | ENI | ENI | ENI | ++/+++ |
| d. Nodular | Y | N | N | N | (+) – ++ | |
| 5 | Fibroblastic | Y (chronic) | Y | Y | Y | (+) – ++ |
| 6 | Verrucose | N | Y | N | Y | (+) – ++ |
| 7 | Fibroblastic | Y | Y | Y | Y | (+) – ++ |
| 8 | a. Verrucose | N | Y | N | Y | ++ |
| b. Verrucose | N | Y | Y | Y | ++ | |
| 9 | a.–c. Nodular | N | Y | Y | Y | ++ |
| 10 | Fibroblastic | Y (chronic) | Y | N | Y | (+)/+ |
| 11 | Nodular | Y | Y | Y | Y | ++ |
| 12 | Fibroblastic | N | Y | N | Y | (+) |
| 13 | a. Verrucose | N | Y | N | Y | ++/+++ |
| b. Nodular | N | N | N | N | ++/+++ | |
| 14 | a. Nodular | ENI | ENI | ENI | ENI | ++/+++ |
| b., d. Nodular | N | N | N | N | ++/+++ | |
| c. Nodular | N | Y | Y | Y | ++/+++ | |
| e., f. Nodular | N | N | Y | Y | ++/+++ | |
| g. Verrucose | Y | Y | Y | Y | ++/+++ | |
| 15 | a. Nodular | Y | Y | Y | Y | (+) |
| b. Verrucose | N | N | N | Y | (+) | |
| 16 | Nodular | Y | Y | Y | Y | ++/+++ |
| 17 | Nodular | N | N | N | N | ++/+++ |
| 18 | Verrucose | N | N | N | Y | (+) – ++ |
| 19 | Fibroblastic | Y | Y | Y | Y | ++/+++ |
| 20 | Verrucose | N | N | N | Y | (+) – ++ |
| 21 | Verrucose | N | Y | N | Y | (+) – ++ |
| 22 | a. Nodular | Y | Y | Y | Y | (+) – ++ |
| b., c. Verrucose | N | N | N | Y | (+) – ++ | |
| 23 | Fibroblastic | Y (chronic) | Y | Y | Y | (+) – ++ |
| 24 | Occult | Y | N | N | Y | (+) – ++ |
| 25 | a. Fibroblastic | ENI | ENI | ENI | ENI | (+) – ++ |
| b. Fibroblastic | Y | Y | Y | Y | ++/+++ | |
| c. Fibroblastic | Y | Y | Y | Y | ++ | |
| B | ||||||
|---|---|---|---|---|---|---|
| Type of sarcoid | Epidermal hyperplasia | Epidermal rete peg formation | Picket fence formation | |||
| Present (%) | Absent (%) | Present (%) | Absent (%) | Present (%) | Absent (%) | |
| Verrucous | 6 (54.55) | 5 (45.45) | 2 (18.18) | 9 (81.82) | 11 (100.00) | 0 (0.00) |
| Nodular | 8 (53.33) | 7 (46.67) | 10 (66.67) | 5 (33.33) | 10 (66.67) | 5 (33.33) |
| Fibroblastic | 10 (100) | 0 (0) | 8 (80.00) | 2 (20.00) | 10 (100.00) | 0 (0.00) |
EH, epidermal hyperplasia; RPF, rete peg formation; PFF, picket fence formation; ENI, epidermis not included.
MMP-1 expression score: (+) faint, +weak, ++ moderate, +++ strong.
Indicates the range of scores in different tumour areas.
Transmission electron microscopy
Local tumour cell behaviour was assessed by TEM on case 14b with epidermal RPF, cases 12 and 16 with PFF and case 13b with dermal infiltrative growth. Tissue blocks (2 mm3) were prepared from the fixed specimens, post-fixed in 1% osmium tetroxide(aq) and routinely resin embedded. Toluidine blue stained semi-thin sections (0.75 µm) identified areas of interest from which ultrathin sections (80 nm) were subsequently prepared and examined.
Results
Clinical information, classification and histological features of sarcoids
Sarcoids were clinically classified as nodular (n = 19), verrucose (n = 11), fibroblastic (n = 11), occult (n = 1) or mixed (n = 1) (Table 1). They presented typical histological features and were variably cellular, non-demarcated aggregates of spindle shaped fibroblastoid cells in an irregular collagenous stroma (Martens et al, 2000, Scott, Miller, 2003).
In 37 cases, an intact epidermis at least partly covered the neoplastic infiltrate. It exhibited epidermal hyperplasia (EH; n = 24), RPF (n = 20), and/or PFF (n = 32), and appeared unaltered in only three nodular sarcoids. PFF was always present in the verrucose and fibroblastic sarcoids (Table 1). In verrucose sarcoids, RPF occurred only alongside EH (2/6). Nodular sarcoids always showed EH together with both RPF and PFF, but either both (2/6) or none (4/6) of the latter without EH (Table 1).
Expression of MMP-1, -2 and -9 in the skin and in sarcoids
The normal skin showed intense constitutive MMP-1 expression, represented by a diffuse cytoplasmic reaction in epithelial cells of epidermis, hair follicles and adnexal glands, in vascular and erector pili smooth muscle cells, and in endothelial cells (see Appendix: Supplementary Fig. S1). In contrast, MMP-2 and MMP-9 expression was very weak and limited to epithelial cells (see Appendix: Supplementary Fig. S1). Similar MMP expression was generally found in the skin over and around sarcoid (Fig. 1A ). However, epithelial MMP-2 expression was variable, often lacking (n = 22) and sometimes (n = 6) slightly more intense than in the control skin.
Fig. 1.
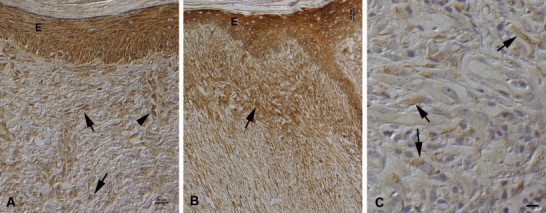
MMP-1 and -9 expression in neoplastic cells. PAP method. (A) Nodular sarcoid (case 14c) with epidermal hyperplasia. The epidermis (E) shows moderate, the neoplastic cells (arrows) weak MMP-1 expression. Activated endothelial cells in a vein are also strongly MMP-1 positive (arrowhead). Bar, 20 µm. (B) Nodular sarcoid with picket fence formation (case 9b). MMP-1 is strongly expressed by keratinocytes in the epidermis (E). Neoplastic cells are diffusely positive, but show the strongest expression intensity close to the epidermis (arrow). Bar, 50 µm. (C) Fibroblastic sarcoid (case 23), exhibiting weak MMP-9 expression by neoplastic cells (arrows). Bar, 10 µm.
In all tumours, neoplastic cells expressed MMP-1, evident as a diffuse cytoplasmic staining (Fig. 1A). Staining intensities varied, both between specimens and within individual tumours. Comparison of the overall staining intensity in the larger tumour cohorts showed the highest mean expression in nodular (2.16) and the lowest in verrucose sarcoids (1.55), fibroblastic sarcoids showed only a slightly higher average (1.66). There were significant differences overall between the different sarcoid types (P < 0.05) while pair-wise testing identified differences between the verrucose and nodular sarcoids (P < 0.05).
In many cases, there was evidence of an expression gradient in association with PFF (most intense close to epidermis; Fig. 1B), (focal) ulceration (strongest reaction in proximity to ulceration) and granulation tissue formation (strongest reaction in neoplastic cells intermingled with new vessels; see Appendix: Supplementary Fig. S2) though this spatial association was not reflected in the association between the overall presence of PFF and the intensity of MMP-1 expression. In some cases, a weak reaction was also seen in pre-existing fibrocytes and nerve fibres in the tumour-free dermis, and in the serum.
Univariable non-parametric analysis using Mann–Whitney test identified no significant association between the intensity of MMP-1 expression and the presence of EH, RPF and PFF (P > 0.05). Because of the lack of normal distribution the analysis does not consider the effect that the different histopathological features may have on the expression of the MMP-1 or the clustering effect of the origin of some sarcoids from the same animal. Multivariable analysis on a larger sample could help clarify such associations. Neoplastic cells were generally MMP-2 negative. In 10 cases (occult, 2 nodular, 3 verrucose, 4 fibroblastic sarcoids), however, they exhibited very weak MMP-9 expression (Fig. 1C).
Many cases (17/38 with natural surface; Table 1), and in particular the fibroblastic (9/10) and nodular (5/15) types exhibited variably extensive ulceration. In four tumours, this was chronic, with marked granulation tissue formation. Occasionally, mild peripheral perivascular mononuclear infiltration was also seen. Macrophages within infiltrates expressed MMP-1, MMP-2 and, occasionally, MMP-9. In areas of early ulceration, keratinocytes were strongly MMP-2 positive (see Appendix: Supplementary material).
Local infiltrative growth and interaction between neoplastic cells and epidermis in sarcoids
Tumour infiltrates were not demarcated. Instead, neoplastic fibroblasts infiltrated the pre-existing dermal collagenous matrix (Fig. 2A ). Within the tumour mass, collagen bundles were irregular and less densely packed than in the normal dermis (Fig. 2B) and stained positively for MMP-1 (Fig. 2C). TEM indicated that they were loosened around neoplastic cells (Fig. 2D).
Fig. 2.
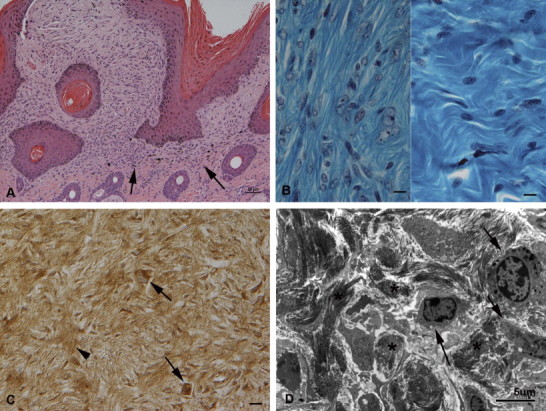
Dermal infiltrative growth. (A) Nodular sarcoid (case 11) with picket fence formation. In the dermis, the neoplastic infiltrate is not demarcated and neoplastic fibroblastoid cells infiltrate into the adjacent pre-existing collagen rich extracellular matrix (arrows). HE stain. Bar, 50 µm. (B) Left: Nodular sarcoid (case 14c). The cells are embedded in loosely arranged thin collagen strands. Right: Normal dermis, comprised of densely packed wavy collagen bundles with a few embedded unaltered fibrocytes. Gomori Trichrome stain. Bars = 10 µm. (C) Nodular sarcoid (case 4b) with intense MMP-1 expression in neoplastic cells (arrows). There is also a moderate reaction in the extracellular matrix (arrowhead). PAP method. Bar, 10 µm. (D) Nodular sarcoid (case 13b). Transmission electron micrograph of the dermal neoplastic infiltrate. Several tumour cells (arrows) with prominent cell projections, infiltrating the pre-existing collagen rich extracellular matrix that contains dense bundles of short collagen fibres (asterisks) which appear loosened around the neoplastic cells. Bar, 5 µm.
Interaction between epidermis and neoplastic cells was indicated by PFF, the perpendicular alignment of tumour cells along the epidermis (Fig. 3A ) without infiltration of the latter, as confirmed by immunohistology for pan-cytokeratin. PFF was identified in 86% of cases (32/37) with intact epidermis and in all clinical types (Table 1). In PFF areas, the dermo–epidermal junction was often obscured, as neoplastic cells and keratinocytes exhibited equally strong MMP-1 expression (Fig. 3B). The basement membrane (BM) appeared thickened and/or loosened in the PAS reaction, and in semi-thin sections, interdigitation of neoplastic cells and keratinocytes in the basal epidermal layer became obvious (Fig. 3C). TEM showed that the BM was absent in these areas (Fig. 3D). In contrast, the BM was continuous when PFF was not observed (Figs. 3E, F). In all cases, the dermal collagen appeared disintegrated where neoplastic cells were present (Figs. 3D, F).
Fig. 3.
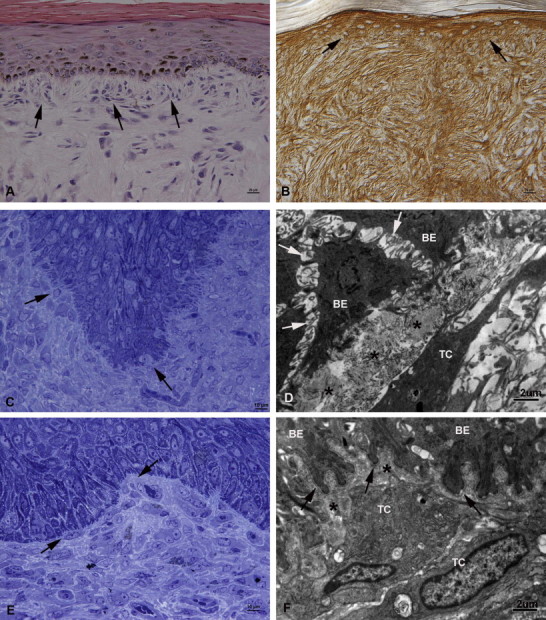
Interaction between epidermis and neoplastic cells. (A) Fibroblastic sarcoid (case 7) with picket fence formation (PFF), represented by the perpendicular arrangement and interdigitation of the subepithelial neoplastic cells with the basal cells of the epidermis (arrows). HE stain. Bar, 20 µm. (B) Verrucose sarcoid with PFF (case 8a) with strong MMP-1 expression of both epidermis and neoplastic cells, obscuring the dermo–epidermal junction (arrows). PAP method. Bar, 20 µm. (C) Nodular sarcoid with PFF (case 16). The epidermal basement membrane (BM) appears multifocally lost due to the interdigitation of neoplastic cells with the basal cells of the epidermis (arrows). Semi-thin section, Toluidine blue stain. Bar = 10 µm. (D) Fibroblastic sarcoid with PFF (case 12). Transmission electron micrograph (TEM). The basal epithelial cells (BE) of the epidermis are devoid of the BM and show loss of cell–cell contact (acantholysis; arrows). The subepithelial collagen (asterisks) is disintegrated and closely apposed to a tumour cell (TC) with its long cell projections. Bar, 2 µm. (E, F) Nodular sarcoid (case 14b) with rete peg formation, but no PFF. (E) Neoplastic cells are positioned close to the BM, but do not interdigitate with the epidermis (arrows). Semi-thin section, Toluidine blue stain. Bar, 10 µm. (F) TEM. The basal epithelial cells (BE) of the epidermis exhibit a well-defined continuous BM (arrows). The subepithelial collagen (asterisks) is disintegrated and closely apposed to tumour cells (TC). Bar, 2 µm.
Discussion
Equine sarcoids originate from fibroblasts transformed by BPV that acquire the capacity to invade the extracellular matrix (ECM) through the expression of MMPs (Yuan et al, 2008b, Yuan et al, 2010b, Hartl et al, 2011). In the present in vivo study, histology, immunohistology for MMP-1, -2 and -9, and TEM were used to assess local behaviour and MMP expression patterns in sarcoids. Our study confirms that histology does not allow sub-classification, since all relevant morphological features are observed in all clinical types (Pascoe, Knottenbelt, 1999, Martens et al, 2000, Knottenbelt, 2005). Hyperplasia and RPF of the overlying epidermis and interaction of neoplastic cells with the epidermis (PFF) were also typically found in all types, to varying extents.
Sarcoids are locally aggressive, recur frequently and can progress rapidly from clinically benign to more aggressive forms (Knottenbelt, 2005, Martens et al, 2001b, Marti et al, 1993, Tarwid et al, 1985). This is reflected histologically by local infiltrative growth of the neoplastic cells (Martens et al., 2000) which we found in association with apparent loosening of the dermal collagen matrix. We observed strong MMP-1 expression by neoplastic cells and MMP-1 deposition in the ECM suggesting that MMP-1 contributes to the ECM changes.
MMP-1 is an interstitial collagenase and is expressed by several cell types, including fibroblasts, keratinocytes and macrophages; it predominantly degrades collagen III. Expression is regulated at transcriptional level, and induced, for example, by cytokines and growth factors (Shapiro, 1998, Westermarck, Kähäri, 1999, Fields, 2013). Produced in an inactive form, MMP-1 is activated by several enzymes (Kähäri, 1997, Westermarck, Kähäri, 1999). In equine sarcoids, MMP-1 is present in its active form (Yuan et al., 2010b), and its release by neoplastic cells may be responsible for infiltrative dermal tumour growth, via partial destruction of the ECM, i.e. the cleaving of collagen III. This indicates increased ECM turnover, and indeed, sarcoids have been shown to exhibit an overall lower amount of collagen than the normal equine dermis, through an increased and more organised, fibrous collagen III network (Williams et al., 1982). The fate of the cleaved collagen III remains unclear, since further degrading requires the gelatinases, MMP-2 and -9 (Shapiro, 1998, Visse, Nagase, 2003, Fields, 2013), neither of which were found to be up regulated in the present nor a previous study (Yuan et al., 2010b).
We demonstrated MMP-1 expression in sarcoids by both neoplastic fibroblasts and epithelial cells, in particular epidermal keratinocytes, and with similar intensity. Assuming that the active form is present, MMP-1, which can degrade several BM components, could also be responsible for PFF in sarcoids since it accompanies loss of BM continuity. In support of this, an in vitro study showed that sarcoid fibroblast-derived cells invaded a 3D Matrigel model composed of several BM components (Yuan et al., 2010b).
The epidermal BM is a thin (<100 nm), but vital interface between basal keratinocytes and dermis. Its upper lamina lucida and lower lamina densa solidly attach the epidermis to the dermis. This anchoring system is composed of hemidesmosomes at the basal pole of the basal keratinocytes, thin anchoring filaments that traverse the lamina lucida, and anchoring fibrils (composed of collagen VII loops) originating from the lamina densa and extending towards the dermis, as well as collagen IV and laminin networks which provide the basic scaffolding of the BM and are linked via nidogen. The keratin filament of the hemidesmosomes binds to laminin 5 which in turn binds to, among others, collagen VII (McMillan et al, 2003, Breitkreutz et al, 2013). MMP-1 can degrade laminins and collagen VII; with the destruction of these two proteins, i.e. the anchoring fibrils and one of the two main networks (Shapiro, 1998, Villone et al, 2008), the BM would lose its normal stable architecture. This could account for the BM destruction seen with PFF in sarcoids and might also render the skin overlying the neoplastic infiltrate more vulnerable and liable to ulceration.
The local behaviour of sarcoids differs from malignant tumours that invade the epidermis, such as human cutaneous melanomas in which MMP-2 and -9 are expressed by neoplastic cells and degrade collagen IV, thereby destroying the BM scaffolding, (Väisäinen et al, 1996, Chen et al, 2012, Breitkreutz et al, 2013). This is not encountered in sarcoids. However, MMP-2 is also expressed by hyperproliferative keratinocytes (Krengel et al., 2002) which are found in the epidermis overlying equine sarcoids (Martens et al., 2000). This may account for the slight increase in epidermal MMP-2 expression in some of our cases. The latter was particularly obvious in association with PFF, suggesting that direct contact between the epidermis and fibroblasts is a prerequisite of proliferation (Martens et al., 2000). Alternatively, BPV-1, found to productively infect the epidermis overlying natural sarcoids (Brandt et al., 2011), could induce keratinocyte hyperproliferation.
In contrast, the strong MMP-2 expression that we observed in keratinocytes in early ulceration could result from cytokine and MMP-1 release by inflammatory cells (Kähäri, 1997). Similarly, the pronounced MMP-1 expression of neoplastic cells might result from local cytokine release with ulceration, granulation tissue formation and PFF (Westermarck and Kähäri, 1999). Our observation that normal equine epidermis and follicular and glandular epithelium constitutively express MMP-1 differs from a previous study (Miragliotti et al., 2008). This could be due to differences in the detection sensitivity since different antibodies and detection systems were used.
The equine sarcoid remains a clinical challenge and there remains a high risk of failure with all treatment methods (Knottenbelt and Kelly, 2000). Invasion is the major constraint to therapy. The likelihood of recurrence is highest with surgical methods that do not include a safe margin but the difficulty in defining a safe margin for any individual lesion is a significant surgical challenge. The likelihood of recurrence is seemingly correlated with the presence of BPV DNA in the surgical margins (Martens et al., 2001c). Topical and intralesional chemotherapy options may fail because the tumour extends beyond the reach of the chemotherapeutic agent (Knottenbelt and Kelly, 2000). This study suggests that, excluding the malignant form which we have not examined, the risks of invasion are not significantly different in any particular sarcoid subtype. Early, pre-emptive, use of medications that could alter the tendency to invasion through manipulation of MMP-1 expression might provide improved outcomes when they are subsequently treated either surgically or medically.
Conclusions
The present in situ study demonstrates a correlation between the expression of MMP-1 by neoplastic and epithelial cells and the local behaviour of sarcoids. The release of MMP-1 could be responsible for the consistently dermal invasion of the neoplastic cells, through the degradation of collagen III, and could, through partial destruction of the BM, mediate PFF. These results which were obtained in all clinical types of sarcoids suggest potential new approaches for the therapy of sarcoids.
Conflict of interest statement
None of the authors of this paper has a financial or personal relationship with other people or organisations that could inappropriately influence or bias the content of the paper.
Acknowledgements
The authors wish to thank the technicians in the Histology Laboratory and the Electron Microscopy Unit, Veterinary Laboratory Services, School of Veterinary Sciences, University of Liverpool, for excellent technical support.
Footnotes
Supplementary data associated with this article can be found in the online version at doi:10.1016/j.tvjl.2014.07.026.
Appendix. Supplementary material
The following are the supplementary data to this article:
Expression of MMP-1, -2 and -9 in the normal equine skin (control case). (A) MMP-1 is moderately to strongly expressed by epithelial cells in epidermis (E), hair follicles (HF) and sebaceous (SG) and apocrine glands (arrows). In addition, smooth muscle and endothelial cells in blood vessels (V) as well as the smooth muscle cells of the arrector pili muscles (arrowheads) show a positive reaction. PAP method. Bar, 50 µm. (B) MMP-2 expression is generally very weak and restricted to epithelial cells in epidermis (E), hair follicles (HF) and sebaceous (SG) and apocrine glands (arrows). HRP method. Bar, 50 µm. (C) MMP-9 expression is generally weak and seen in epithelial cells in epidermis (E), hair follicles (HF) and sebaceous (SG) and apocrine glands (arrows). PAP method. Bar, 50 µm.
Ulceration and granulation tissue formation in sarcoids. (A) Fibroblastic sarcoid (case 5) with chronic ulceration, with underlying granulation tissue with numerous new vessels (arrowheads). Inset: Occult sarcoid (case 24) with focal mononuclear perivascular infiltration. Bar, 50 µm (inset: Bar, 20 µm). HE stain. (B) Mixed sarcoid (case 3) with extensive ulceration and granulation tissue formation. MMP-1 is expressed by inflammatory cells and proliferating fibroblasts in the granulation tissue (arrows) and by endothelial cells forming the new vessels (arrowheads). Neoplastic cells also exhibit moderate MMP-1 expression. Inset: Occult sarcoid (case 24). Leukocytes in the mononuclear perivascular infiltrate exhibit moderate MMP-1 expression. PAP method. Bar, 50 µm (inset: Bar, 20 µm). (C) Fibroblastic sarcoid (case 5) with ulceration and granulation tissue formation. MMP-2 expression is seen in infiltrating macrophages in the inflammatory process. Endothelial cells in new vessels (arrowheads) and neoplastic cells are MMP-2 negative. HRP method. Bar, 50 µm. (D) Verrucose sarcoid (case 6) with early ulceration, exhibiting strong MMP-2 expression by epithelial cells (arrowheads). HRP method. Bar, 50 µm.
References
- Bogaert L., Martens A., Wijbe M., van Marck E., de Cock H. Bovine papillomavirus DNA can be detected in keratinocytes of equine sarcoid tumors. Veterinary Microbiology. 2010;146:269–275. doi: 10.1016/j.vetmic.2010.05.032. [DOI] [PubMed] [Google Scholar]
- Brandt S., Tober R., Carteggio A., Burger S., Sabitzer S., Walter I., Kainzbauer C., Steinborn R., Nasir L., Borzacchiello G. BPV-1 infection is not confined to the dermis but also involves the epidermis in equine sarcoids. Veterinary Microbiology. 2011;150:35–40. doi: 10.1016/j.vetmic.2010.12.021. [DOI] [PubMed] [Google Scholar]
- Breitkreutz D., Kozholt I., Thiemann K., Nischt R. Skin basement membrane: The foundation of epidermal integrity – BM functions and diverse roles of bridging molecules nidogen and perlacan. BioMed Research International. 2013;2013 doi: 10.1155/2013/179784. 179784; Epub 2013 Mar 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y., Chen Y., Huang L., Yu J. Evaluation of heparanase and matrix metalloproteinase-9 in patients with cutaneous malignant melanoma. The Journal of Dermatology. 2012;39:339–343. doi: 10.1111/j.1346-8138.2011.01441.x. [DOI] [PubMed] [Google Scholar]
- Fields G.B. Interstitial collagen catabolism. The Journal of Biological Chemistry. 2013;288:8783–8793. doi: 10.1074/jbc.R113.451211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartl B., Hainisch E.K., Shafti-Keramat S., Kirnbauer R., Corteggio A., Borzacchiello G., Tober R., Kainzbauer C., Practscher B., Brandt S. Inoculation of young horses with bovine papillomavirus type 1 virions leads to early infection of PBMCs prior to pseudo-sarcoid formation. Journal of General Virology. 2011;92:2437–2445. doi: 10.1099/vir.0.033670-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kähäri V.-M. Matrix metalloproteinases in skin. Experimental Dermatology. 1997;6:199–213. doi: 10.1111/j.1600-0625.1997.tb00164.x. [DOI] [PubMed] [Google Scholar]
- Kipar A., Baumgärtner W., Burkhardt E. Round cell sarcomas of possible myelomonocytic origin localized at the lip of aged dogs. Journal of Veterinary Medicine Series A. 1995;42:185–200. doi: 10.1111/j.1439-0442.1995.tb00370.x. [DOI] [PubMed] [Google Scholar]
- Kipar A., May H., Menger S., Weber M., Leukert W., Reinacher M. Morphological features and development of granulomatous vasculitis in feline infectious peritonitis. Veterinary Pathology. 2005;42:321–330. doi: 10.1354/vp.42-3-321. [DOI] [PubMed] [Google Scholar]
- Knottenbelt D.C. A suggested clinical classification for the equine sarcoid. Clinical Techniques in Equine Practice. 2005;4:278–295. [Google Scholar]
- Knottenbelt D.C., Kelly D.F. The diagnosis and treatment of periorbital sarcoid in the horse: 445 cases from 1974 to 1999. Veterinary Ophthalmology. 2000;3:169–191. doi: 10.1046/j.1463-5224.2000.00119.x. [DOI] [PubMed] [Google Scholar]
- Krengel S., Alexander M., Brinckmann J., Tronnier M. MMP-2, TIMP-2 and MT1-MMP are differentially expressed in lesional skin of melanocytic nevi and their expression is modulated by UVB-light. Journal of Cutaneous Pathology. 2002;29:390–396. doi: 10.1034/j.1600-0560.2002.290702.x. [DOI] [PubMed] [Google Scholar]
- Martens A., De Moor A., Demeulemeester J., Ducatelle R. Histopathological characteristics of five clinical types of equine sarcoid. Research in Veterinary Science. 2000;69:295–300. doi: 10.1053/rvsc.2000.0432. [DOI] [PubMed] [Google Scholar]
- Martens A., de Moor A., Ducatelle R. PCR detection of bovine papilloma virus DNA in superficial swabs and scrapings from equine sarcoids. The Veterinary Journal. 2001;161:280–286. doi: 10.1053/tvjl.2000.0524. [DOI] [PubMed] [Google Scholar]
- Martens A., de Moor A., Vlaminck L., Pille F., Sternhaut M. Evaluation of excision, cryosurgery and local BCG vaccination for the treatment of equine sarcoids. Veterinary Record. 2001;149:665–669. doi: 10.1136/vr.149.22.665. [DOI] [PubMed] [Google Scholar]
- Martens A., de Moor A., Demeulemeester J., Peelman L. Polymerase chain reaction analysis of the surgical margins of equine sarcoids for bovine papilloma virus DNA. Veterinary Surgery. 2001;30:460–467. doi: 10.1053/jvet.2001.25874. [DOI] [PubMed] [Google Scholar]
- Marti E., Lazary S., Antczak D.F., Gerber H. Report of the first international workshop on equine sarcoid. Equine Veterinary Journal. 1993;25:397–407. doi: 10.1111/j.2042-3306.1993.tb02981.x. [DOI] [PubMed] [Google Scholar]
- McMillan J.R., Akiyama M., Shimizu H. Epidermal basement membrane zone components: Ultrastructural distribution and molecular interactions. Journal of Dermatological Science. 2003;31:169–177. doi: 10.1016/s0923-1811(03)00045-8. [DOI] [PubMed] [Google Scholar]
- Miragliotti V., Lefebvre-Lavoie J., Lussier J.G., Theoret C.L. Equine ANXA2 and MMP1 expression analyses in an experimental model of normal and pathological wound repair. Journal of Dermatological Science. 2008;51:103–112. doi: 10.1016/j.jdermsci.2008.03.008. [DOI] [PubMed] [Google Scholar]
- Pascoe R.R.R., Knottenbelt D.C. Neoplastic conditions. In: Pascoe R.R.R., Knottenbelt D.C., editors. Manual of Equine Dermatology. Saunders; London: 1999. pp. 27–31. [Google Scholar]
- Scott D.W., Miller W.H., Jr. Neoplastic and non-neoplastic tumours. In: Scott D.W., Miller W.H., editors. Equine Dermatology. Elsevier Science (USA); St. Louis: 2003. pp. 719–731. [Google Scholar]
- Shapiro S.D. Matrix metalloproteinase degradation of extracellular matrix: Biological consequences. Current Opinion in Cell Biology. 1998;10:602–608. doi: 10.1016/s0955-0674(98)80035-5. [DOI] [PubMed] [Google Scholar]
- Tarwid J.N., Fretz P.B., Clark E.G. Equine sarcoids: A study with emphasis on pathologic diagnosis. The Compendium of Continuing Education. 1985;7:293–300. [Google Scholar]
- Torrontegui B.O., Reid S.W.J. Clinical and pathological epidemiology of the equine sarcoid in a referral population. Equine Veterinary Education. 1994;6:85–88. [Google Scholar]
- Väisäinen A., Tuominen H., Kallioinen M., Turpeenniemi-Hujanen T. Matrix metalloproteinase-2 (72kD type IV collagenase) expression occurs in the early stage of human melanocytic tumor progression and may have prognostic value. The Journal of Pathology. 1996;180:283–289. doi: 10.1002/(SICI)1096-9896(199611)180:3<283::AID-PATH662>3.0.CO;2-3. [DOI] [PubMed] [Google Scholar]
- Villone D., Fritsch A., Koch M., Hansen U., Bruckner P. Supramolecular interactions at the dermo-epidermal junction zone. Anchoring fibril-collagen VII tightly binds to banded collagen fibrils. The Journal of Biological Chemistry. 2008;283:24506–24513. doi: 10.1074/jbc.M802415200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visse R., Nagase H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circulatory Research. 2003;92:827–839. doi: 10.1161/01.RES.0000070112.80711.3D. [DOI] [PubMed] [Google Scholar]
- Westermarck J., Kähäri V.-M. Regulation of matrix metalloproteinases in tumor invasion. The FASEB Journal. 1999;13:781–792. [PubMed] [Google Scholar]
- Williams I.F., Heaton A., McCullagh K.G. Connective tissue composition of the equine sarcoid. Equine Veterinary Journal. 1982;14:305–310. doi: 10.1111/j.2042-3306.1982.tb02436.x. [DOI] [PubMed] [Google Scholar]
- Yuan Z.Q., Nicolson L., Marchetti B., Gault E.A., Campo M.S., Nasir L. Transcriptional changes induced by bovine papillomavirus type 1 in equine fibroblasts. Journal of Virology. 2008;82:6481–6491. doi: 10.1128/JVI.00429-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan Z.Q., Gault E.A., Goeil P., Nixon C., Campo M.S., Nasir L. Establishment and characterization of equine fibroblast cell lines transformed in vivo and in vitro by BOV-1: Model systems for equine sarcoids. Virology. 2008;373:352–361. doi: 10.1016/j.virol.2007.11.037. [DOI] [PubMed] [Google Scholar]
- Yuan Z.Q., Bennett L., Campo M.S., Nasir L. Bovine papillomavirus type 1 E2 and E7 proteins down-regulate toll like receptor 4 (TLR4) expression in equine fibroblasts. Virus Research. 2010;149:124–127. doi: 10.1016/j.virusres.2010.01.008. [DOI] [PubMed] [Google Scholar]
- Yuan Z.Q., Gobeil A.M., Campo M.S., Nasir L. Equine sarcoid fibroblasts over-express matrix metalloproteinases and are invasive. Virology. 2010;396:143–151. doi: 10.1016/j.virol.2009.10.010. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Expression of MMP-1, -2 and -9 in the normal equine skin (control case). (A) MMP-1 is moderately to strongly expressed by epithelial cells in epidermis (E), hair follicles (HF) and sebaceous (SG) and apocrine glands (arrows). In addition, smooth muscle and endothelial cells in blood vessels (V) as well as the smooth muscle cells of the arrector pili muscles (arrowheads) show a positive reaction. PAP method. Bar, 50 µm. (B) MMP-2 expression is generally very weak and restricted to epithelial cells in epidermis (E), hair follicles (HF) and sebaceous (SG) and apocrine glands (arrows). HRP method. Bar, 50 µm. (C) MMP-9 expression is generally weak and seen in epithelial cells in epidermis (E), hair follicles (HF) and sebaceous (SG) and apocrine glands (arrows). PAP method. Bar, 50 µm.
Ulceration and granulation tissue formation in sarcoids. (A) Fibroblastic sarcoid (case 5) with chronic ulceration, with underlying granulation tissue with numerous new vessels (arrowheads). Inset: Occult sarcoid (case 24) with focal mononuclear perivascular infiltration. Bar, 50 µm (inset: Bar, 20 µm). HE stain. (B) Mixed sarcoid (case 3) with extensive ulceration and granulation tissue formation. MMP-1 is expressed by inflammatory cells and proliferating fibroblasts in the granulation tissue (arrows) and by endothelial cells forming the new vessels (arrowheads). Neoplastic cells also exhibit moderate MMP-1 expression. Inset: Occult sarcoid (case 24). Leukocytes in the mononuclear perivascular infiltrate exhibit moderate MMP-1 expression. PAP method. Bar, 50 µm (inset: Bar, 20 µm). (C) Fibroblastic sarcoid (case 5) with ulceration and granulation tissue formation. MMP-2 expression is seen in infiltrating macrophages in the inflammatory process. Endothelial cells in new vessels (arrowheads) and neoplastic cells are MMP-2 negative. HRP method. Bar, 50 µm. (D) Verrucose sarcoid (case 6) with early ulceration, exhibiting strong MMP-2 expression by epithelial cells (arrowheads). HRP method. Bar, 50 µm.