

Abstract
In the present work, a single-step purification of recombinant nucleocapsid protein (NP) of the Newcastle disease virus (NDV) directly from unclarified feedstock using an expanded bed adsorption chromatography (EBAC) was developed. Streamline 25 column (ID = 25 mm) was used as a contactor and Streamline chelating adsorbent immobilized with Ni2+ ion was used as affinity adsorbent. The dynamic binding capacity of Ni2+-loaded Streamline chelating adsorbent for the NP protein in unclarified feedstock was found to be 2.94 mg ml−1 adsorbent at a superficial velocity of 200 cm h−1. The direct purification of NP protein from unclarified feedstock using expanded bed adsorption has resulted in a 31% adsorption and 9.6% recovery of NP protein. The purity of the NP protein recovered was about 70% and the volume of processing fluid was reduced by a factor of 10. The results of the present study show that the IMA-EBAC developed could be used to combine the clarification, concentration and initial purification steps into a single-step operation.
Keywords: NP protein, NDV, IMA-EBAC, Escherichia coli, Dynamic binding capacity
The nucleocapsid (NP) 1 proteins of many viruses are highly immunogenic, which could be further developed as diagnostic reagents. The NP protein of SARS-CoV (a novel coronavirus) is immunodominant and has successfully tested on ELISA for detecting the SARS-CoV NP antibody [1]. The NP proteins of other related viruses, such as Newcastle disease [2], mumps [3], rabies [4], measles [5], and infectious bronchitis [6] viruses, were shown to have the potential to be developed as diagnostic reagents. The NP of Newcastle disease virus has been cloned and expressed as ring-like and herringbone-like structures in Escherichia coli [7]. The former was reported to have the potential to be used as a biocarrier for the development of subunit vaccine [8].
The intracellular recombinant NDV NP is currently purified in a laboratory scale, which involves multiple purification steps. Typically, the feedstock (cell lysate) contains particulate materials, which have to be removed prior to further purification steps. Centrifugation and filtration are the two most widely used techniques for the removal of biomass or cell debris [9]. However, these clarification methods possess several disadvantages such as time consuming, substantial protein loss, and high operational and capital costs. Indeed, our previous study showed that about 80% of the NP protein of NDV was lost during the clarification step using centrifugation [10]. After clarification step, the supernatant has to be concentrated using ammonium sulphate precipitation, dialyzed and purified through sucrose gradient ultracentrifugation. These purification steps are very costly and labour intensive and not scalable. Hence, there is a need to develop a simple and scalable purification process for recombinant NP protein.
Recent advance in expanded bed adsorption (EBA) has shown its promises to be used to capture, concentrate and purify product directly from fermentation broth that contains whole cells, cell debris and other particulate components [11], [12]. EBA has been used for the clarification and purification of biological suspensions containing very small particles such as cell debris [13]. EBA processes employ the flow of fluid through an initially packed bed structure to achieve a contactor with a high-fluid voidage. A stable bed is formed with adsorbent having a well-defined distribution of bead sizes [14]. Under these circumstances, the larger/heavier adsorbent beads will locate near the bottom of the bed, whereas the smaller/lighter adsorbent beads will locate near the top [15]. The high-fluid voidage allows for unhindered passage of cells, cell debris and other particulates during the application of crude feed to the column.
EBA benefits as a process by a combination of clarification, concentration and initial purification into a single, integrated operational procedure for the recovery of proteins from crude feedstock [16]. The advantages of EBA include increased process economy due to a decreased number of process steps, increased process yield, shorter overall processing time, and reducing running and capital costs [13]. EBA has been used successfully with a number of different feedstocks such as bacterial [17], [18], [19], yeast [20], mammalian cell [14] fermentation broths and transgenic milk [21]. Recent work has also demonstrated the application of EBA employing immobilized metal affinity chromatography (IMAC) for the purification of histidine-tag proteins from various sources [22], [23]. The recombinant NP of NDV has been previously tagged with hexa-histidine residues [7] that facilitated its adsorption onto the immobilized metal affinity (IMA) adsorbent. Therefore, the aim of the present study was to develop a simple and rapid technique to recover NP protein from unclarified E. coli lysate using IMA-EBAC.
Materials and methods
Adsorbent
Streamline chelating adsorbent with size distribution of 100–300 μm and approximate density of 1.2 g ml−1 was purchased from Amersham Bioscience (Uppsala, Sweden). It has the metal chelating ligand iminodiacetic acid (IDA) attached to it.
Feedstock preparation
Escherichia coli cells harbouring the recombinant plasmid of the NPcfus gene, pTrcHis2-NPcfus [7], was cultured in Luria–Bertani (LB) broth containing ampicillin (50 μg ml−1) at 37 °C. The protein expression was induced by the addition of isopropyl β-d-thiogalactopyranoside (IPTG, Promega, USA) at a final concentration of 1 mM. The cells were harvested by centrifugation at 3840g (JA 14 rotor, Beckman) for 10 min at 4 °C. It was then disrupted using a sonicator (Labsonic U, B. Braun, Germany) as previously described [10]. The cell lysate was used directly for the expanded bed processes.
Protein purity
The purity of samples was analyzed on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) as previously described [10].
Protein quantification
Following SDS–PAGE, proteins were transferred to nitrocellulose membranes by using the Transblot SD semidry transfer cell (Bio-Rad, Hercules, CA, USA) as previously described [10]. The amount of NP protein was estimated from the intensity of the band on Western blots using the Sophisticated Volume Tools from the Bio-Rad imaging devices supported by Quantity One (Bio-Rad) as described in Tey et al. [24]. The amount of total protein was quantified according to the Bradford assay [25] using bovine serum albumin (BSA) as a standard.
Equilibrium adsorption isotherm
The adsorption isotherm of the Streamline chelating adsorbent was determined by batch-binding experiments. A range of NP protein concentration of unclarified feedstock was prepared in binding buffer (0.02 M sodium phosphate and 0.5 M NaCl, pH 8.0) in separate tubes. The Ni2+-loaded and equilibrated Streamline chelating adsorbents of 0.5 ml were added to each of the protein samples prepared. The adsorbents and protein mixtures were incubated in a roller incubator for 1 h at room temperature to allow equilibration between the solid and the liquid phases. The adsorbents were then separated by gravity sedimentation and the supernatants were assayed for the NP protein concentration.
The effect of pH and the presence of biomass on NP protein adsorption
A set of experiments was performed in batch-binding mode to select an optimal binding pH. A 0.5 ml of settled volume of Streamline chelating adsorbent (pre-loaded with Ni2+ ions and pre-equilibrated in binding buffer at a range of pH values, i.e., pH 6.0–8.5) and 9 ml of clarified and unclarified E. coli feedstock were added to each of the centrifugal tubes. The adsorbent/protein mixtures were mixed well on a roller incubator for 1 h at room temperature. The mixtures were allowed to settle and the supernatants were collected and assayed for the NP protein concentration.
Bed expansion characteristic
The characteristics of bed expansion were measured at room temperature (25 °C) in the Streamline 25 column (ID = 25 mm) (Amersham Bioscience). Nickel ions charged and equilibrated adsorbent particles were loaded into the column, corresponding to a sediment bed height of 13.7 cm. The bed expansion characteristics of the Streamline chelating adsorbents were determined by monitoring the bed height as a function of increasing superficial velocity. The degree of expansion is calculated as the ratio between the expanded bed height (H) and the sediment bed height (H 0).
Dynamic binding capacity
Streamline chelating adsorbents were poured into the Streamline 25 column to give a sediment bed height of 10 cm. The bed was fluidized and charged with Ni2+ ions and then equilibrated with binding buffer (0.02 M sodium phosphate and 0.5 M NaCl, pH 8.0) at a flow velocity of 200 cm h−1 to yield a stable expanded bed height of 20.8 cm. Unclarified E. coli feedstock was pumped onto the expanded bed at the same flow velocity. Protein fractions collected throughout the experiment were subsequently assayed for NP protein concentrations.
Direct purification of NP protein from unclarified feedstock
Process scheme of the purification of the NP protein using EBAC is shown in Fig. 1 . The entire purification process was performed at room temperature (25 °C). The unchanged Streamline chelating adsorbents were first loaded onto a Streamline 25 column. It was then charged with 0.1 M nickel sulphate solution at 100 cm h−1. After charging, the adsorbents were equilibrated with equilibration buffer (0.02 M sodium phosphate and 0.5 M sodium chloride, pH 8.0) at 200 cm h−1. The adsorption of NP protein was performed by loading the unclarified feedstock onto the equilibrated column at 200 cm h−1. After the adsorption, the column was washed with equilibration buffer. The elution of NP protein was performed in the downwards flow packed bed mode, first with elution buffer 1 (50 mM imidazole in 0.02 M sodium phosphate and 0.5 M sodium chloride, pH 8.0) and followed by elution buffer 2 (350 mM imidazole in 0.02 M sodium phosphate and 0.5 M sodium chloride, pH 8.0) at 10 cm h−1. Elution fractions were collected and subjected to protein analysis and quantification.
Fig. 1.

Process scheme of the purification of the NP protein using expanded bed adsorption chromatography. The entire purification process is performed at room temperature (25 °C).
Results and discussion
Adsorption isotherm
The equilibrium adsorption isotherm on the Streamline chelating adsorbent was determined by plotting the NP protein adsorbed per unit mass of adsorbent versus the equilibrium concentration of NP protein in binding buffer. The adsorption isotherm for proteins onto adsorbent is typically modeled by the Langmuir isotherm equation
where Q m is the maximum equilibrium adsorption capacity and K d the dissociation constant. Q and C are the adsorbed and aqueous protein concentration in equilibrium, respectively. The isotherm in Fig. 2 was clearly observed which resembled the linear portion of a Langmuirian isotherm. The maximum adsorption capacity (Q m) of NP protein to the Streamline chelating is 58.48 mg of NP protein per gram of adsorbent and the dissociation constant (K d) is 3.04 mg ml−1. This high binding capacity could be exploited to reduce the amount of adsorbent used and thus decreases the material cost. Therefore, Streamline chelating was chosen to purify NP protein from unclarified E. coli feedstock. Indeed, Clemmitt and Chase [22] have reported that Streamline chelating has very high binding capacity when compared to other similar base matrices with different ligands.
Fig. 2.

Adsorption isotherm of NP protein from unclarified feedstock on Ni2+-loaded Streamline chelating adsorbent.
Effect of pH and biomass on NP protein adsorption
The most important factor in choosing the suitable buffer solution for the adsorption and desorption of proteins depends on the pH and ionic strength to be used. The equilibrium adsorption data in various pH of clarified and unclarified feedstock are summarized in Table 1 . The results show that the optimal binding pH for NP protein onto the Streamline chelating for both feedstocks is pH 8. Indeed, Clemmitt and Chase [26] have reported that the range of pH for the adsorption of proteins onto a metal-IDA adsorbent is between 7 and 8. The adsorption of protein onto the immobilized metals occurs at pH higher than its pK a. Since the pK a of histidine residues is about pH 7 [27], pH 8 has become the optimal pH for its binding. However, further increase of pH to 8.5 has substantially reduced the binding capacity of Streamline chelating adsorbent. The highest equilibrium binding capacity of Ni2+-loaded Streamline chelating adsorbents for NP protein achieved at pH 8.0 was about 13.74 and 10.90 mg ml−1 of adsorbents for unclarified and clarified feedstocks, respectively. The results show that the binding capacity of Streamline chelating adsorbent was not affected by the presence of biomass. Hence, the unclarified feedstock can be directly applied onto the IMAC-EBAC for the purification of NP protein. The direct purification from unclarified feedstock has several advantages such as reduction in processing time, cost, and higher recovery yield. Indeed, substantial amount of NP protein was lost during the clarification process [10]. Only a small fraction of total protein (about 10–20% and most of them were NP protein) bound to the Ni2+-loaded Streamline chelating adsorbents for both clarified and unclarified feedstocks. This suggests that the binding of non-specific proteins to Ni2+-loaded Streamline chelating adsorbent is very little, which is an important feature of IMAC adsorbent.
Table 1.
Influence of pH on the binding of NP protein from (a) clarified and (b) unclarified feedstock [10% biomass wet mass (w/v)] to Ni2+-loaded Streamline chelating adsorbent
| pH | Equilibrium binding capacity (mg NP protein/ml adsorbent) |
|
|---|---|---|
| Unclarified feedstock | Clarified feedstock | |
| 6.0 | 8.82 | 4.94 |
| 7.5 | 10.70 | 9.64 |
| 8.0 | 13.74 | 10.90 |
| 8.5 | 8.54 | 10.26 |
Bed expansion
The bed expansion characteristics of the Streamline chelating adsorbent with the binding buffer and feedstock suspension [8% biomass wet mass (w/v)] are shown in Fig. 3 . The bed height increased linearly with the superficial velocity and can be described by the Richardson–Zaki equation
where U is the superficial velocity (cm h−1), ε is the bed voidage, U t is the terminal settling velocity (cm h−1), and n is the Richardson–Zaki exponent. The voidage of the expanded bed depends on the degree of expansion, which is the ratio of the expanded and sediment bed height, H/H 0. The bed voidage of the expanded bed was calculated from equation
where ε is the expanded bed voidage and ε 0 is the settled bed voidage. A value of 0.4 was assumed for ε 0 [28], [29]. Bed expansion characteristics are dependent on the viscosity and density of process liquid and certain physical properties of adsorbent. As such, the expanded bed was found to expand to a great degree at a particular velocity in the flow in the presence of 8% (w/v) cell lysate due to the increase of feedstock viscosity and density. The n and U t values were determined from the linear regression of the Richardson–Zaki equation. The value of the n for the buffer and the presence of biomass was 5.23 and 5.17, respectively (Table 2 ). These values correspond to the theoretical value of 4.8, which is normally used in the laminar flow regime [30]; thus, the bed was assumed to be stable upon expansion in the presence of biomass. The U t for the adsorbent fluidized in the feedstock suspension, 865.4 cm h−1, is less than the value in the buffer because of the greater viscosity and density of feedstock suspension than the buffer. The linear flow rate intended to be used for bed equilibration and adsorption, 200 cm h−1, is far lower than the U t. If the linear flow rate is higher than the U t, the adsorbent bead will be “blow out” from the column.
Fig. 3.

Bed expansion characteristic of the Streamline chelating adsorbent in binding buffer (■) and unclarified feedstock [8% biomass wet mass (w/v)] (♦).
Table 2.
Results of the linear regression of the Richardson–Zaki plots
| Binding buffer | Feedstock [8% biomass wet mass (w/v)] | |
|---|---|---|
| Richardson–Zaki coefficient, n | 5.23 | 5.17 |
| Terminal settling velocity, Ut (cm h−1) | 1104.71 | 865.35 |
Dynamic binding capacity
Frontal analysis was performed to determine the dynamic binding capacity of an expanded bed of Ni2+-loaded Streamline chelating for NP protein in unclarified feedstock. The dynamic binding capacity (mg of protein adsorbed per ml of sedimented adsorbent) can be calculated as Q B [31]
where V b is the volume at 10% breakthrough (ml), C 0 is the initial concentration of the feedstock (mg ml−1), and V s is the settled volume of the adsorbent (ml). The estimated dynamic binding capacity of adsorbent in unclarified feedstock could be used to determine the maximal loading of unclarified feedstock that could be applied onto the expanded bed. Fig. 4 shows the breakthrough curve for the NP protein from unclarified feedstock onto Ni2+-loaded Streamline chelating adsorbent in an expanded bed. The NP protein was efficiently captured from the unclarified feedstock. In this study, the dynamic binding capacity of Ni2+-loaded Streamline chelating adsorbent for the NP protein in unclarified feedstock was found to be 2.94 mg ml−1 adsorbent at a superficial velocity of 200 cm h−1. Therefore, the optimal loading of unclarified E. coli feedstock (0.98 mg ml−1 NP protein) that could be applied onto the 100 ml of adsorbent is about 300 ml. However, 300 ml of feedstock is somehow difficult to handle for the present EBA column setup. Instead, 400 ml of feedstock was used, which resulted in the reasonable amount of NP lose during the breakthrough and washing (Table 3 ).
Fig. 4.

Breakthrough curve for the NP protein from unclarified feedstock onto Ni2+-loaded Streamline chelating adsorbent in an expanded bed.
Table 3.
IMA-EBAC purification of NP protein from unclarified feedstock
| Purification stages | Volume (ml) | NP protein concentration (mg ml−1) | NP protein yield (%) | Purity (%) |
|---|---|---|---|---|
| Unclarified feedstock | 400 | 0.98 | 100 | 10.0 |
| Flow through | 450 | 0.38 | 44.0a | 10.2 |
| Washing | 1040 | 0.09 | 25.1a | 7.5 |
| NP protein bound | — | — | 30.9 | — |
| Elution 1 (50 mM imidazole) | 80 | 0.11 | 2.2 | 6.2 |
| Elution 2 (350 mM imidazole) | 40 | 0.94 | 9.6 | 70.0 |
It indicates protein lost.
Direct purification of the NP protein from unclarified feedstock
Streamline chelating (100 ml) was used and corresponds to a sediment bed height of 20 cm. The unclarified feedstock was applied to the expanded bed at a flow velocity of 200 cm h−1, which corresponded to an expanded bed height of 41 cm. The stepwise elution is often selected for large preparative applications because it is technically simpler and reproducible compared to gradient elution [32]. Table 3 shows that the direct purification from unclarified feedstock using IMAC-EBAC has resulted in a 31% adsorption and 9.6% recovery of NP protein. It was a great improvement compared to the 1.3% final yield obtained from the multistep conventional methods involving centrifugation, precipitation, and ultracentrifugation [10].
Unbound and weakly bound proteins were eliminated with elution buffer containing low imidazole concentration (50 mM). The NP protein was subsequently eluted from the column by applying elution buffer containing high imidazole concentration (350 mM). Fig. 7 shows the SDS–PAGE analysis of protein from elution fractions collected from the second elution, while lane 5 in Fig. 5 is the pool of these elution fractions. Fig. 6 shows the elution profile of both elution 1 and elution 2. Peak 1 represents the proteins eluted from elution 1 (50 mM imidazole) and peak 2 represents the proteins eluted from elution 2 (350 mM imidazole). Peak 1 of Fig. 6 and lane 4 of Fig. 5 show that eluate 1 contains higher amount of contaminating proteins and only about 2% of NP proteins were present. Approximately, 38 mg or about 31% of bound NP protein has been eluted from the adsorbent by elution buffer with high imidazole concentration. Substantial amount of protein with molecular weight lower than 60 kDa was found in the eluate 2 (Fig. 5, lane 5 and Fig. 7). These impurities could be either the contaminating protein that was not removed during elution 1 or the degraded NP protein. Indeed, Wizemann and von Brunn [33] have also reported the presence of protein slightly below the size of target protein in the eluate during the purification of hepatitis B virus nucleocapsid protein using Ni2+-chelate affinity chromatography and regarded it as an N-terminal degradation product of the target protein. The present results show that the NP protein has been efficiently purified from unclarified feedstock by using IMA-EBAC without the need of any clarification and concentration steps. In addition, the purity of the NP protein recovered is reasonably high (about 70%) and most importantly the volume of processing fluid was reduced by a factor of 10 (400 ml of feedstock to 40 ml of eluate; Table 3).
Fig. 7.

SDS–PAGE of the NP protein fractions collected in Elution 2 (350 mM imidazole). Lane M: molecular mass markers in kDa; lanes 1–27: fractions collected in elution 2. The arrow shows the location of NP protein (58 kDa).
Fig. 5.
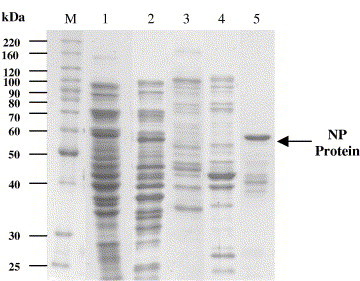
SDS–PAGE analysis of the IMA-EBAC purification of NP protein from unclarified feedstock. Lane M: Molecular mass markers in kDa. Lane 1: unclarified feedstock; lane 2: flow through; lane 3: wash; lane 4: elution 1 (50 mM imidazole); lane 5: elution 2 (350 mM imidazole). The arrow shows the position of NP protein (58 kDa).
Fig. 6.

Elution profile of the purification of NP protein from unclarified E. coli feedstock using IMA-EBAC. [Peak 1: elution 1 (50 mM imidazole); peak 2: elution 2 (350 mM imidazole).]
Conclusion
A procedure for the IMA-EBAC for the direct recovery of the NP protein from unclarified E. coli lysates has been developed. The results of the present study showed that it is possible to recover NP protein directly from unclarified feedstock utilizing IMA-EBAC. A two-step elution protocol was used to elute NP protein from the adsorbent. The protocol employed elution buffer containing 50 mM imidazole that is capable of eliminating high amount of contaminating proteins and elution buffer containing 350 mM imidazole that efficiently elutes large amount of NP protein from the adsorbent. The elution of NP protein was performed in packed bed mode in order to reduce fraction volumes and to concentrate the NP protein. The combination of clarification, capture and purification in a single-step in IMA-EBAC has substantially reduced the downstream processing time, volume of processing fluid, as well as capital and operational costs involved.
Acknowledgments
This study was supported by IRPA Grants 09-02-04-0766-EA001 and 01-02-04-003 BTK/ER/006 from the Ministry of Science, Technology and the Innovation of Malaysia.
Footnotes
Abbreviations used: NP, nucleocapsid; EBA, expanded bed adsorption; IMAC, immobilized metal affinity chromatography; LB, Luria–Bertani; IPTG, isopropyl β-d-thiogalactopyranoside; SDS–PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis; BSA, bovine serum albumin.
References
- 1.Timani K.A., Ye L., Ye L.B., Zhu Y., Wu Z.H., Gong Z.J. Cloning, sequencing, expression, and purification of SARS-associated coronavirus nucleocapsid protein for serodiagnosis of SARS. J. Clin. Virol. 2004;30:309–312. doi: 10.1016/j.jcv.2004.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Errington W., Steward M., Emmerson P. A diagnostic immunoassay for Newcastle disease virus based on the nucleocapsid protein expressed by a recombinant baculovirus. J. Virol. Methods. 1995;55:357–365. doi: 10.1016/0166-0934(95)00074-7. [DOI] [PubMed] [Google Scholar]
- 3.Slibinskas R., Samuel D., Gedvilaite A., Staniulis J., Sasnauskas K. Synthesis of the measles virus nucleoprotein in yeast Pichia pastoris and Saccharomyces cerevisiae. J. Biotechnol. 2004;107:115–124. doi: 10.1016/j.jbiotec.2003.10.018. [DOI] [PubMed] [Google Scholar]
- 4.Reid-Sanden F., Sumner J.W., Smith J.S., Fekadu M., Shaddock J.H., Bellini W.J. Rabies diagnostic reagents prepared from a rabies N gene recombinant expressed in Baculovirus. J. Clin. Microbiol. 1990;28:858–863. doi: 10.1128/jcm.28.5.858-863.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Samuel D., Sasnauskas K., Jin L., Gedvilaite A., Slibinskas R., Beard S., Zvirbliene A., Oliveira S.A., Staniulis J., Cohen B., Brown D. Development of a measles specific IgM ELISA for use with serum and oral fluid samples using recombinant measles nucleoprotein produced in Saccharomyces cerevisiae. J. Clin. Virol. 2003;28:121–129. doi: 10.1016/s1386-6532(02)00273-1. [DOI] [PubMed] [Google Scholar]
- 6.Seah J.N., Yu L., Kwang J. Localization of linear B-cell epitopes on infectious bronchitis virus nucleocapsid protein. Vet. Microbiol. 2000;75:11–16. doi: 10.1016/s0378-1135(00)00202-9. [DOI] [PubMed] [Google Scholar]
- 7.Kho C.L., Tan W.S., Yusoff K. Production of the nucleocapsid protein of Newcastle disease virus in Escherichia coli and its assembly into ring- and nucleocapsid-like particles. J. Microbiol. 2001;39:293–299. [Google Scholar]
- 8.Rabu A., Tan W.S., Kho C.L., Omar A.R., Yusoff K. Chimeric Newcastle disease virus nucleocapsid with parts of viral hemagglutinin-neuraminidase and fusion proteins. Acta virol. 2002;46:211–217. [PubMed] [Google Scholar]
- 9.Scope R.K. Springer-Verlag; New York: 1994. Protein Purification—Principle and Practice. pp. 1–21. [Google Scholar]
- 10.Tan Y.P., Ling T.C., Yusoff K., Tan W.S., Tey B.T. Comparative evaluation of three purification methods for the nucleocapsid protein of Newcastle disease virus from Escherichia coli homogenates. J. Microbiol. 2005;43:295–300. [PubMed] [Google Scholar]
- 11.Bruce L.J., Chase H.A. Evaluation of the effect of in-bed sampling on expanded bed adsorption. Bioseparation. 1999;8:77–83. [PubMed] [Google Scholar]
- 12.Fernandez-Lahore H.M., Kleef R., Kula M.-R., Thommes J. The influence of complex biological feedstock on the fluidization and bed stability in expanded bed adsorption. Biotechnol. Bioeng. 1999;64:484–496. doi: 10.1002/(sici)1097-0290(19990820)64:4<484::aid-bit11>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- 13.Anspach F.B., Curbelo D., Hartmann R., Garke G., Deckwer W.-D. Expanded-bed chromatography in primary protein purification. J. Chromatogr. A. 1999;865:129–144. doi: 10.1016/s0021-9673(99)01119-x. [DOI] [PubMed] [Google Scholar]
- 14.Thommes J., Halfar M., Lenz S., Kula M.R. Purification of monoclonal antibodies from whole hybridoma fermentation broth by fluidized bed adsorption. Biotechnol. Bioeng. 1995;45:205–211. doi: 10.1002/bit.260450304. [DOI] [PubMed] [Google Scholar]
- 15.Bruce L.J., Chase H.A. Hydrodynamics and adsorption behavior within an expanded bed adsorption column studied using in-bed sampling. Chem. Eng. Sci. 2001;56:3149–3162. [Google Scholar]
- 16.Fernandez-Lahore H.M., Geilenkirchen S.K., Boldt A., Nagel, Kula M.-R., Thommes J. The influence of cell adsorbent interactions on protein adsorption in expanded beds. J. Chromatogr. A. 2000;873:195–208. doi: 10.1016/s0021-9673(99)01308-4. [DOI] [PubMed] [Google Scholar]
- 17.Johansson H.J., Jagersten C., Shiloach J. Large scale recovery and purification of periplasmic recombinant protein from E. coli using expanded bed adsorption chromatography followed by new ion exchange media. J. Biotechnol. 1996;48:9–14. doi: 10.1016/0168-1656(96)01390-9. [DOI] [PubMed] [Google Scholar]
- 18.Lee Y.S., Suh C.W., Park S.K., Lee E.K. Purification of soluble human epidermal growth factor (hEGF) for recombinant Escherichia coli culture broth by using expanded bed adsorption chromatography. Biotechnol. Appl. Biochem. 2003;37:1–5. doi: 10.1042/BA20020113. [DOI] [PubMed] [Google Scholar]
- 19.Ling T.C., Loong C.K., Tan W.S., Tey B.T., Abdullah W.M.W., Ariff A. Purification of filamentous bacteriophage M13 by expanded bed anion exchange chromatography. J. Microbiol. 2004;42:228–232. [PubMed] [Google Scholar]
- 20.Owen R.O., McCreath G.E., Chase H.A. A new approach to continuous counter-current protein chromatography: direct purification of malate dehydrogenase from a Saccharomyces cerevisiae homogenate as a model system. Biotechnol. Bioeng. 1997;53:427–441. doi: 10.1002/(SICI)1097-0290(19970220)53:4<427::AID-BIT11>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- 21.Ozyurt S., Kirdar B., Ulgen K.O. Recovery of antithrombin III from milk by expanded bed chromatography. J. Chromatogr. A. 2002;944:203–210. doi: 10.1016/s0021-9673(01)01102-5. [DOI] [PubMed] [Google Scholar]
- 22.Clemmitt R.H., Chase H.A. Facilitated downstream processing of a histidine-tagged protein from unclarified Escherichia coli homogenates using immobilized metal affinity expanded-bed adsorption. Biotechnol. Bioeng. 2000;67:206–216. doi: 10.1002/(sici)1097-0290(20000120)67:2<206::aid-bit10>3.0.co;2-x. [DOI] [PubMed] [Google Scholar]
- 23.Noubhani A.M., Dieryck W., Chevalier S., Santarelli X. On-line purification of His-tag enhanced green fluorescent protein taken directly from a bioreactor by continuous ultrasonic homogenization coupled with immobilized metal affinity expanded bed adsorption. J. Chromatogr. A. 2002;968:113–120. doi: 10.1016/s0021-9673(02)00794-x. [DOI] [PubMed] [Google Scholar]
- 24.Tey B.T., Yong K.H., Ong H.P., Ling T.C., Ong S.T., Tan Y.P., Ariff A., Tan W.S. Optimal conditions for Hepatitis B core Antigen production in shaked flask fermentation. Biotechnol. Bioprocess Eng. 2004;9:374–378. [Google Scholar]
- 25.Bradford M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 26.Clemmitt R.H., Chase H.A. Immobilized metal affinity chromatography of β-galactosidase from unclarified Escherichia coli homogenate using expanded bed adsorption. J. Chromatogr. A. 2000;874:27–43. doi: 10.1016/s0021-9673(00)00087-x. [DOI] [PubMed] [Google Scholar]
- 27.Yip T.-T., Nakagawa Y., Porath J. Evaluation of the interaction of peptides with Cu2+, Ni2+, Zn2+ by high performance immobilized metal ion affinity chromatography. Anal. Biochem. 1989;183:159–171. doi: 10.1016/0003-2697(89)90184-x. [DOI] [PubMed] [Google Scholar]
- 28.Nayak D.P., Ponrathnam S., Rajan C.R. Macroporous copolymer matrix IV. Expanded bed adsorption application. J. Chromatogr. A. 2001;922:63–76. doi: 10.1016/s0021-9673(01)00923-2. [DOI] [PubMed] [Google Scholar]
- 29.Theodossiou I., Elsner H.D., Thomas O.R.T., Hobley T.J. Fluidization and dispersion behaviour of small high density pellicular expanded bed adsorbents. J. Chromatogr. A. 2002;964:77–89. doi: 10.1016/s0021-9673(02)00590-3. [DOI] [PubMed] [Google Scholar]
- 30.Chang Y.K., Chase H.A. Ion exchange purification of G6PDH from unclarified yeast cell homogenates using expanded bed adsorption. Biotechnol. Bioeng. 1996;49:204–216. doi: 10.1002/(SICI)1097-0290(19960120)49:2<204::AID-BIT9>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- 31.Güzeltunç E., Ülgen K.Ö. Recovery of actinorhodin from fermentation broth. J. Chromatogr. A. 2001;914:67–76. doi: 10.1016/s0021-9673(00)01245-0. [DOI] [PubMed] [Google Scholar]
- 32.Kalil S.J., Maugeri-Filho F., Rodrigues M.I. Ion exchange expanded bed chromatographic for the purification of an extracellular inulinase from Kluyveromyces marxianus. Process Biochem. 2005;40:581–586. [Google Scholar]
- 33.Wizemann H., von Brunn A. Purification of E. coli-expressed HIS-tagged hepatitis B core antigen by Ni2+-chelate affinity chromatography. J. Virol. Methods. 1999;77:189–197. doi: 10.1016/s0166-0934(98)00152-9. [DOI] [PubMed] [Google Scholar]