

Abstract
Syntax, the structure of sentences, enables humans to express an infinite range of meanings through finite means. The neurobiology of syntax has been intensely studied but with little consensus. Two main candidate regions have been identified: the posterior inferior frontal gyrus (pIFG) and the posterior middle temporal gyrus (pMTG). Integrating research in linguistics, psycholinguistics, and neuroscience, we propose a neuroanatomical framework for syntax that attributes distinct syntactic computations to these regions in a unified model. The key theoretical advances are adopting a modern lexicalized view of syntax in which the lexicon and syntactic rules are intertwined, and recognizing a computational asymmetry in the role of syntax during comprehension and production. Our model postulates a hierarchical lexical-syntactic function to the pMTG, which interconnects previously identified speech perception and conceptual-semantic systems in the temporal and inferior parietal lobes, crucial for both sentence production and comprehension. These relational hierarchies are transformed via the pIFG into morpho-syntactic sequences, primarily tied to production. We show how this architecture provides a better account of the full range of data and is consistent with recent proposals regarding the organization of phonological processes in the brain.
Keywords: Syntax, Dual-Stream model, Broca’s area, posterior temporal lobe, paragrammatism
Introduction
Language is arguably the most uniquely human cognitive trait (von Humboldt 1836; Darwin 1871; Hauser et al. 2002; Tattersall 2004; Bolhuis et al. 2014). Many other abilities once thought to set humans apart from our nearest primate cousins—tool use, culture, theory of mind, altruism, and empathy—are increasingly documented in other species (Whiten et al. 1999; Warneken et al.2007; Call and Tomasello 2008; De Waal 2008; Seed and Byrne,2010). But decades of research on the cognitive capacities of nonhuman apes as well as birds with complex vocalizations has shown that the combinatorial nature of language, with syntactic structure at its core, is beyond their reach (Terrace et al. 1979; Berwick et al. 2011; Beckers et al. 2012; Yang 2013; Poletiek et al.2016). It is no surprise, then, that there has been great interest in the evolution and neural basis of language.
Current models of language evolution, broadly speaking, emphasize its multicomponent nature involving a set of traits, each, perhaps, with its own evolutionary history: traits such as vocal learning, gestural mimicry, and conceptual combination (Fitch 2017). Research on the neural circuits that support language has made significant progress in many of these domains, identifying networks that support vocal motor control (Guenther 2006; Hickok 2012a; Simonyan 2014), imitation (Iacoboni 2009), speech perception and word recognition (Hickok,and Poeppel 2007; Rauschecker and Scott 2009; Chang et al. 2010), and semantics (Binder et al. 2009), including simple forms of conceptual combination (Pylkkannen 2015). But comparatively little progress, measured in terms of agreement in the field, has been made in mapping the computational system that binds all these pieces together and imbues language with its communicative power: syntax.
The dominant view regarding the neurology of syntax over the last 40 years is that Broca’s area—the posterior portion of the inferior frontal gyrus (pIFG) including the pars triangularis (pTri) and pars opercularis—is a critical hub for this capacity (Grodzinsky and Santi 2008; Hagoort 2014; Friederici et al. 2017). The origin of this hypothesis is that people with Broca’s aphasia, and assumed damage to Broca’s area, often suffer from the symptom of expressive “agrammatism”, a tendency to omit grammatical elements and generally simplify sentence structure in connected speech production (Goodglass 1968, 1993). In the 1970s, it was discovered that Broca’s aphasia is also associated with particular sentence comprehension deficits that suggested a fundamental syntactic problem (Caramazza and Zurif 1976), kicking off decades of intense investigation of the role of Broca’s area in syntactic processing, assessed primarily via comprehension tasks (Grodzinsky and Amunts 2006; Matchin and Rogalsky 2017). However, while some large-scale lesion-deficit mapping studies have reported an association between damage to Broca’s area and comprehension of syntactically complex and/or noncanonical sentence structures (Wilson et al. 2010a; Wilson et al. 2011; Magnusdottir et al. 2013; Mesulam et al. 2015; Fridriksson et al. 2018), many others have primarily implicated the posterior temporal lobe in basic syntactic processing and not Broca’s area (Dronkers et al. 2004; Wilson and Saygin 2004; Baldo and Dronkers 2007; Peelle et al. 2008; Pillay et al. 2017; Rogalsky et al. 2018; Wilson et al. 2018b, 2018c).
Some of the strongest evidence for the link between Broca’s area and syntactic comprehension has come from neuroimaging studies of structural processing, which often implicate this region (for reviews, see Friederici 2011, 2017; Hagoort 2014). However, a closer look at the neuroimaging results on sentence comprehension shows that the activation profile of the posterior temporal lobe is almost always coupled with Broca’s area (see Matchin et al. 2017b, for data and a review). This finding weakens the proposed unique associations between Broca’s area and receptive syntactic function and has led to proposals that both frontal and posterior temporal lobe regions participate in syntax (Tyler and Marslen-Wilson 2007; Wilson et al. 2014, 2016; Blank et al. 2016). Other researchers have challenged the imaging results further still, arguing that what appear to be syntactic effects during comprehension may instead be attributed to working memory (Rogalsky et al. 2008a; Rogalsky and Hickok,2011) and/or cognitive control (Novick et al. 2005, 2010; January et al. 2009). This has led to alternative proposals that emphasize the primacy of posterior temporal and/or parietal regions (Thothathiri et al. 2012; Bornkessel-Schlesewsky and,Schlesewsky 2013; Bornkessel-Schlesewsky et al. 2015; Pillay,et al. 2017).
The anterior temporal lobe (ATL) emerged around the turn of the century as a third candidate region to underlie syntactic processing. This is primarily due to its functional response properties: a preference for sentence stimuli over lists of words, music, and environmental sound sequences (Mazoyer et al. 1993; Stowe et al. 1998; Humphries et al. 2001, 2005, 2006; Rogalsky and Hickok 2008; Rogalsky et al. 2011; Matchin et al. 2017b). But on closer examination of the neuroimaging literature, several important observations indicated a semantic, rather than syntactic, function of the ATL. An early study (Mazoyer et al. 1993) showed a weak response to meaning-impoverished but syntactically structured sentences in the ATL, suggesting a function tied to syntax. However, subsequent studies did not replicate this finding, highlighting instead the IFG and pMTG (Pallier et al. 2011; Fedorenko et al. 2012a; Goucha and Friederici 2015; Matchin et al. 2017b). Other neuroimaging studies have revealed that the ATL responds to experimental manipulations of conceptual-semantic or discourse properties regardless of syntax (Fletcher et al. 1995; Maguire et al. 1999; Vandenberghe et al. 2002; Humphries et al. 2006; Westerlund and Pylkkänen 2014), while failing to respond to manipulations of syntax independently of semantics (Rogalsky and Hickok 2008; Del Prato et al. 2014). The strongest evidence comes from neuropsychological studies on patients with ATL degeneration, damage, or resection (see Wilson et al. 2014, for data and review). While such patients have striking conceptual-semantic impairments, basic sentence comprehension in these patients is unimpaired when lexical demands are minimized, and sentence production appears normal, with no evidence of grammatical errors (Hodges et al. 1992; Hodges and Patterson 2007; Kho et al. 2008; Wilson et al. 2012; Mesulam et al. 2015; Rogalsky et al. 2018). In addition, these patients are sensitive to syntactic violations in the same fashion as healthy subjects (Grossman et al. 2005; Cotelli et al. 2007).
Here we propose an alternative model that (1) parcels the syntactic network into two broad computational processes, hierarchical structuring and morpho-syntactic linearization, and (2) acknowledges a task-dependent, “production–comprehension asymmetry” in the recruitment of these computations. This explains the key involvement of inferior frontal regions: predominantly linearization for production, but sometimes co-opted for comprehension as a form of syntactic working memory and syntactic prediction, and posterior temporal regions: hierarchical structuring for both comprehension and production. To preview our proposal, we argue that:
In comprehension, the posterior middle temporal gyrus (pMTG) functions to decode sequences of auditory phonological representations in the posterior superior temporal gyrus (pSTG) into hierarchical structures and link these with two conceptual networks, an entity knowledge hub in the ATL and an event knowledge hub in the angular gyrus (AG) (Binder and Desai 2011).
In production, the computational task is different, to take nonsequential conceptual information, derive hierarchical structures (pMTG), and transform them into sequences of morphemes, which is accomplished via the pTri of the inferior frontal gyrus (IFG, the anterior part of Broca’s area).
This functional architecture extends previous dual-stream proposals regarding perception-production asymmetries at the lexical-phonological level (Hickok and Poeppel 2000, 2004, 2007; Hickok et al. 2012; Hickok 2014a, 2014b) to the syntactic level, thus conserving a basic architectural plan for the language system. In addition, our proposal adopts a modern lexicalized view of syntax in which the lexicon and syntactic rules are intertwined in the same representational system. This allows us to account for the fact that the posterior temporal lobe regions that are associated with syntactic processes are also robustly associated with lexical-level processes (Hickok and Poeppel 2007; Lau et al. 2008; Fedorenko et al. 2018).
The balance of this article is organized as follows. We first define the syntactic domain that we target for neural mapping. Then we lay out our hypotheses along with their conceptual motivation. We then provide a detailed review of the relevant evidence regarding the competing hypotheses for neural organization of syntax, including the present one. We then consider syntax in the context of sentence comprehension and production, arguing for the proposed asymmetry. Finally, we conclude by briefly discussing questions concerning how the syntactic system develops in the human brain.
What is Syntax?
Language is best conceptualized as a form-meaning interface: a system for transforming often nonsequential thoughts into a linear sequence of words and sounds, sign gestures, or orthographic forms and back again. For example, the concept of “two large dogs” is inherently nonsequential; there is no sense in which the concepts “large” or “dog” or a two-element set are ordered. Yet we must communicate these ideas by sequencing elements through a serial channel such as the vocal tract. Similarly, during sentence comprehension, the semantic (meaning) relations among words must be recovered, which is determined through hierarchical structures (Heim and Kratzer 1998). Broadly, then, syntax is the component of language that accomplishes these transformations. Understanding how they are achieved is the research goal of theoretical syntax. This is a rich and technical field of inquiry that we can only highlight in the most cursory manner here. We emphasize those aspects most relevant to the present proposal.
First, it is important to underscore that while syntax critically relates to meaning, it is not reducible to it (Chomsky 1957; Adger 2018). This is illustrated by syntactically unacceptable sentences that nonetheless have an unambiguous semantic interpretation, such as “the child seems sleeping” (cf., “the child is sleeping” or “the child seems to be sleeping”) or “Who did you see Edgar and?” (cf., I saw Edgar and Françoise) (Chomsky 1957). By contrast, there are sentences that have no clear semantic interpretation yet have well-formed syntax, such as “jabberwocky” sentences invented by Carroll (1871), which retain function words/morphemes but replace content words with nonsense words:
‘Twas brillig, and the slithy toves
Did gyre and gimble in the wabe:
All mimsy were the borogoves,
And the mome raths outgrabe.
Interestingly, even though one cannot provide a full meaning to jabberwocky sentences, the prose with its functional elements evokes hierarchically structured relationships between the nonsense words. For example, it is likely that “slithy” is a property of “toves”, which are the agents of “gyre”-ing and “gimble”-ing actions carried out in a “wabe” context. This suggests a correlation between hierarchical syntactic construction and semantic interpretations (Johnson and Goldberg 2013); the syntactic structure constrains the semantic relationships between the elements of a sentence.
This relates to our second point: a central function of syntax is to encode/decode hierarchical grouping relations between elements in a sentence that relate to meaning. To illustrate this, principles of syntactic computation can lead to ambiguity in cases where multiple grouping relations are compatible with the input, as in the sentence (S) “Ursula saw the man with binoculars”. If the prepositional phrase (PP) “with binoculars” is grouped with the verb phrase (VP) “saw the man” (Fig. 1A), the sentence roughly means “Ursula used binoculars to see the man”. However, if the PP “with binoculars” is instead grouped with the noun phrase (NP) “the man”, (Fig. 1B), the sentence roughly means “Ursula saw the man who has binoculars”. In other words, the possible meanings are constrained by the possible groupings. With this background, one can better appreciate that the job of syntax is to translate between a surface phonological sequence and a nonsequential, relational semantic representation via hierarchical syntactic structures.
Figure 1.
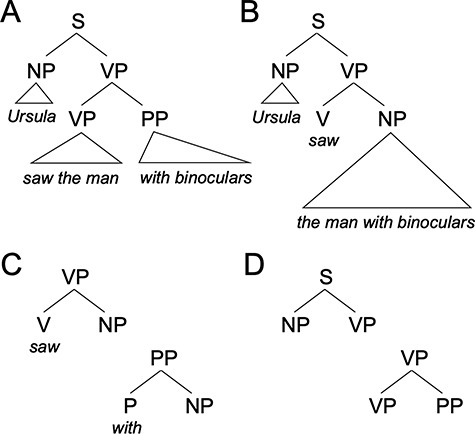
Syntactic structures. (A and B) possible syntactic groupings associated with the sentence Ursula saw the man with binoculars. (A) Structure in which the meaning is roughly “Ursula used binoculars to see the man.” (B) Structure in which the meaning is roughly “Ursula saw the man who has binoculars.” (C) Lexicalized treelets for the verb “saw” and preposition “with”. (D) Abstract lexicalized treelets.
Words and rules: lexicalized structure
Essentially, all syntactic theories contain two things: a lexicon of items (i.e., words) and a system for combining them (i.e., rules) (Chomsky 1965; Pinker 1999; Jackendoff 2002). However, theories differ in the degree of separation between words and rules. Older theories in the generative grammar tradition contained an elaborate system of syntactic phrase structure rules (which generate basic hierarchical groupings) and transformational rules (which re-arrange pieces of structure) (Chomsky 1957, 1965, 1981). Most modern syntactic theories typically assume a radically slimmer rule-based component comprised of a single compositional operation (called “Merge” or “Unify”), with much of the structure of sentences represented along with words in the lexicon (Pollard and Sag 1994; Chomsky 1995; Goldberg 1995; Joshi and Schabes 1997; Bresnan 2001; Frank 2002; Jackendoff 2002). In fact, in some theories, this “lexicalization” of grammar is taken even further, replacing traditional syntactic categories (e.g., noun, verb, NP, and VP) with labels derived from the word itself. In such “bare phrase structure” approaches (Chomsky 1994), instead of NPs there are “cat phrases” and “house phrases”, instead of VPs there are “attack phrases” and “walk phrases”, and so forth. Relatedly, a largely separate psycholinguistic literature on word and sentence production has long postulated the existence of “lemmas” (Kempen and Huijbers 1983; Levelt 1993; Dell and O'Seaghdha 1992), which are abstract word forms without phonological or semantic specification, but with syntactic information. This includes syntactic category (e.g., noun and verb), grammatical gender (e.g., feminine, masculine, and neuter), and subcategorization (e.g., the verb “devour” takes a NP complement such as “the cake” while a verb like “reside” takes a PP complement such as “by the river”). Thus, both of these literatures converge on a lexicalist view of syntax, which naturally fits with overlap in the neural systems involved in lexical and syntactic functions (Fedorenko et al. 2018).
In the present work, we assume this modern lexicalist view. Lexicalized approaches posit “treelets” (stored structures) both associated with individual words, that is with links to a specific phonological form and meaning (Fig. 1C), and more abstract treelets without association to a specific word, that is without phonological and/or conceptual links (Fig. 1D). However, all treelets contain syntactic features and hierarchical structures. Numerous parsing models have made heavy use of such treelets (MacDonald 1994; Jurafsky 1996; Vosse and Kempen 2000; Lewis and Vasishth 2005; Demberg et al. 2013). In these models, during sentence comprehension the perception of words activates treelets that are “clipped” together, depending on the stage of the parse, to form a unified structure (Jackendoff 2017).
The role of serial order
Hierarchical structure is intimately related to semantics. Interestingly, while order is a cue to structure, order by itself is irrelevant to meaning (Heim and Kratzer 1998; Everaert et al. 2015). However, order is a key constraint on the perception and production of language. The vocal tract is a serial channel, and so a hierarchically structured sentence must be produced word-by-word. This has led some to speculate that linear properties of language, such as affixation morphology and word order, are driven by the demands of the motor systems (Idsardi and Raimy 2013; Everaert et al. 2015; Berwick and Chomsky 2016). Interestingly, sign languages generally rely on order less than spoken languages (Aronoff et al. 2005). This may be partly due to the fact that the manual systems allow for greater parallel expression (Sandler and Lillo-Martin 2006).
In comprehension, the hierarchical structure of sentences must be determined from sequential inputs. However, it appears that explicit encoding of linear relations is neither necessary nor even possible during structure building in comprehension (parsing) (Lewis 2000; McElree et al. 2003; Lewis and Vasishth 2005; Lewis et al. 2006). In brief, the recall of order relations is very slow, on the order of hundreds of milliseconds (McElree 1993)—too slow to characterize the rapid pace of sentence comprehension. This suggests that the order of elements in the input is used incrementally to drive parsing but is not specifically encoded in the parser. We discuss later how production-related mechanisms can be used to reiterate the linear order of morphemes for reparsing.
Lewis et al. (2006) illustrates how parsing can succeed without explicit order information through an example from Japanese, in which two nominative case-marked nouns occur in successive order in a sentential complement structure:
(1) Mary-ga John-ga butler-o korosita-to omotta
Mary-NOM John-NOM butler-ACC killed-COMP thought.
‘Mary thought that John killed the butler’.
Both “Mary” and “John” are marked with nominative case, meaning that they are each subjects of a different clause. Which one should be associated with the verb “thought” in the main clause, and which one associated with “killed” in the embedded clause? As Lewis et al. (2006) explain, the initial perception of “Mary” with a nominative case marker automatically cues a predictive parser to build a structure in which “Mary” is attached as the subject of the main clause, and then attaching “John” as a subject of the embedded clause. It is unnecessary to explicitly encode the linear order relation between the two NPs.
Another way of looking at this is that the order information is given by the input, having already been sequenced by the interlocutor, and does not need to be redundantly calculated by a predictive parser, which generates structure incrementally given each morphological input. In contrast, the computation of linear order is necessary during production because of the demands of the vocal tract. As we discuss later, we accordingly posit that the hierarchical-to-linear conversion is processed via interaction with frontal motor-related brain systems, particularly Broca’s area, while the core hierarchical syntactic structures are processed in the posterior temporal lobe.
The cortical organization of syntax
Here we put forward the core claim of our proposal starting with what we view as a significant constraint on the localization of syntax: the neuroanatomy of the phonological and semantic networks with which a syntactic system must interface. The last several years have seen substantial convergence in models of the neuroanatomy of these networks. We provide a brief summary of these in turn.
Networks for speech perception strongly implicate mid-to-posterior superior temporal gyrus (STG) regions extending roughly from the lateral aspect of the STG into the dorsal bank of the superior temporal sulcus (STS) (Hickok and Poeppel 2007). (See Wilson et al. 2018a for evidence that the dorsal and ventral banks of the posterior STS are functionally distinguishable). This includes evidence from functional neuroimaging (Binder et al. 2000; Okada and Hickok 2006; Okada et al. 2010; DeWitt and Rauschecker 2012), phonological-level speech decoding via direct cortical recordings (Pasley et al. 2012; Moses et al. 2016), and interference using direct cortical stimulation (Roux et al. 2015). This work has been reviewed extensively in the articles cited here and will not be discussed further. We will refer to the anatomical location of this auditory-based phonological network as the pSTG (Fig. 2, dark blue).
Figure 2.
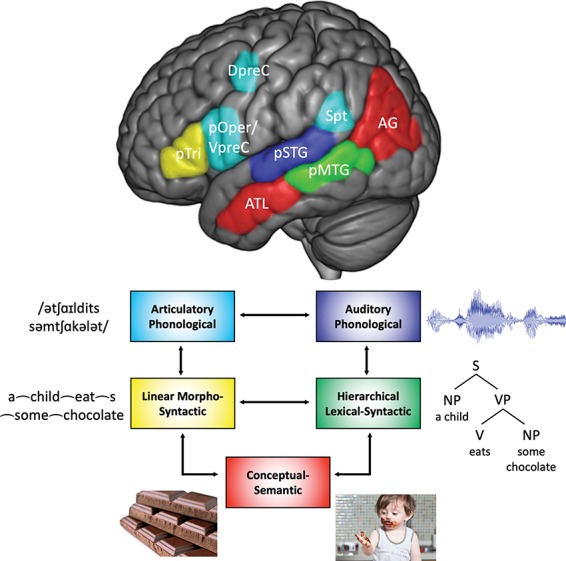
Cortical organization of syntax and the sensorimotor and conceptual-semantic brain systems it interfaces with. pOper/VpreC, inferior frontal gyrus, pars opercularis/ventral precentral gyrus; DpreC, dorsal precentral gyrus; pSTG, posterior superior temporal gyrus (including dorsal bank of STS); pMTG, posterior middle temporal gyrus (including ventral bank of STS). Colored boxes correspond to the same colored brain regions. Arrows indicate bidirectional white matter connections between brain regions.
The auditory-phonological networks in the pSTG also participate in speech production, serving as sound targets for motor-phonological speech planning via a sensorimotor network (Hickok and Poeppel 2007; Rauschecker and Scott 2009; Hickok 2012a, 2012b; Guenther and Hickok, 2015). The sensorimotor network includes the posterior part of Broca’s area, the pars opercularis, the precentral gyrus, the supplementary motor area, and an auditory-motor interface area, Sylvian parietal-temporal (Spt), at the posterior end of the Sylvian fissure (Hickok et al. 2003, 2009; Tourville and Guenther 2011). Thus, phonological networks are asymmetric with respect to task: sensory nodes participate in both perception and production and motor nodes participate primarily in production (Hickok and Poeppel 2000; Hickok et al. 2011; Hickok 2012a) (Fig. 2, cyan).
It has long been postulated that conceptual knowledge representations are widely distributed in cortex, yet organized in one way or another (e.g., by sensorimotor modality and adaptive significance) (Wernicke 1900; Damasio 1989; Mesulam 1990; Caramazza 2000; Martin and Chao 2001; Gage and Hickok 2005; Patterson et al. 2007). Recent work has identified two conceptual knowledge zones that are particularly relevant to the current proposal (Binder et al. 2009; Binder and Desai 2011): one in the ATL (anterior STS/MTG) that is involved in knowledge of entities (e.g., object categories) and another in the vicinity of the AG that is involved in event knowledge (thematic relations between entities) (Fig. 2, red). The distinction between entities and events appears to be a rather fundamental psychological one, revealed in experiments in which adults and children universally tend to group sets of objects together based either on their individual attributes (e.g., grouping CAT and DOG together based on shared features such as ANIMAL, FOUR-LEGGED, and TAIL) or on their relational attributes, that is, how the object interacts with other objects (e.g., grouping DOG with BONE, based on the shared events between them, e.g., WANTING, EATING, and BURYING) (Lin and Murphy 2001; Estes et al. 2011). Fairly direct evidence for this neuroanatomical distinction in the context of a language task comes from Schwartz et al. (2011), who reported that taxonomic lexical substitution errors (e.g., “dog” → “cat”) in stroke patients are associated with damage to the ATL, while thematic errors (e.g., “dog” → “bone”) are associated with damage to the AG. Relatedly, several fMRI studies have shown increased activation with increased verb argument structure complexity (i.e., the number of entities participating in the event) in the AG but not the ATL (Thompson et al. 2007; Thompson et al. 2010; Meltzer-Asscher et al. 2015; Malyutina and den Ouden 2017). Additionally, stimuli containing thematic relations show more robust AG activation than taxonomic relations or nonthematic feature combination (Kalenine et al. 2009; Boylan et al. 2015, 2017; Lewis et al. 2015; Williams et al. 2017). In contrast, increased activation in minimal combinatory contexts that enrich the semantic representation of an entity-based element (e.g., adjective-noun combinations like “red boat”) is consistently observed in the ATL and generally not in the AG (see Pylkkanen 2015, for a review).
Drawing on basic distance minimization principles of neural connectivity (Cherniak 1994; Chklovskii et al. 2002; Chklovskii and Koulakov 2004), a reasonable prior for the location of a network involved in representing hierarchical syntactic information is the cortical zone that sits in between the auditory-phonological and semantic zones. This cortical area corresponds to the pMTG (Fig. 2, green), the location of our proposed hub for hierarchical lexical-syntactic processing. Crucially, this region includes the ventral bank of the STS (Wilson et al. 2018a), which typically shows the most robust syntactic effects in neuroimaging studies (Pallier et al. 2011; Matchin et al. 2017b). This corresponds to the components of theoretical syntax that refer to hierarchical representations, such as phrase structure, C-command relations, movement, and binding constraints (Chomsky 1995, 1981). We define “posterior” as between the lateral extent of Heschl’s gyrus and the end of the Sylvian fissure. This functional anatomical organization converges with that proposed in Wilson et al. (2018a): a syntactic hub situated between semantic and phonological regions of the ventral stream.
We argue further that this pMTG syntactic hub interacts with other networks asymmetrically for comprehension and production, analogous to the perception-production asymmetry in phonological networks. For comprehension, the pMTG converts an incoming linear sequence of morphemes processed in phonological networks into nonlinear hierarchical structures, which are then mapped onto semantic networks. For production, the pMTG syntactic hub converts entity and event concepts processed in the ATL and AG into hierarchical syntactic representations, which are then translated by additional computations into linear morphological sequences for speech output. We argue that the translation from hierarchies to sequences is a function of the interaction between the pMTG syntactic hub and frontal networks. We suggest that the key frontal region is the anterior portion of Broca’s area, the pTri (Fig. 2, yellow), which is connected to the pMTG via the arcuate fasciculus (Yagmurlu et al. 2016) and among the sub-regions of Broca’s area shows the greatest degree of sentence-specific effects in neuroimaging studies (Fedorenko et al. 2012b). These procedures for mapping a hierarchical syntactic representation into a linearized sequence are the objects of inquiry in theoretical syntax and morphology that refer to linear order, such as word order constraints (e.g., the head parameter) and affixation rules (Chomsky 1981; Idsardi and Raimy 2013) (see Bornkessel et al. 2005 and Boeckx et al. 2014 for related proposals regarding linearization and Broca’s area).
Thus, the comprehension-production asymmetry is determined by the difference in computational demands. In comprehension, the task is to convert an input-provided linear sequence of morphemes into a hierarchical representation and thereby to conceptual-semantic representations, whereas for production the task is to transform an intended message first into a nonlinear hierarchy and then into a linear sequence of morphemes. This architecture also provides a natural explanation of the recruitment of frontal circuits in some circumstances of sentence comprehension (Matchin 2017): sequencing mechanisms can be used to maintain information in working memory, which act as a buffer to preserve the input sequence for hierarchical structuring—a “mental rewind button” and may facilitate predictions of upcoming input, via top-down activation of lexical-syntactic representations in pMTG (see Hickok et al. 2011 for a similar proposal in the phonological domain and Hickok 2014, Chapter 10 for some pitfalls with motor-based prediction in perception/comprehension).
Unlike some theorists (e.g., Hagoort 2014; Friederici 2017), we do not propose an anatomical correlate of basic syntactic operations that are promoted in lexicalist theories of syntax (Chomsky 1995; Jackendoff 2002). Instead we focus on broad representation types that are common to all such syntactic theories, leaving the details of specific operations to future work. We speculate that minimal operations such as Merge or Unify are instantiated by subtler biophysical properties within the pMTG such as network connectivity patterns or cortical oscillations (Murphy 2015; Ding et al. 2016) (see Poeppel and Embick 2005; Embick and Poeppel 2015 for discussion of the difficulty in straightforwardly attempting to map basic cognitive operations to brain tissue).
Sign language
Research in the last several decades has revealed that sign languages possess the same core properties of language as spoken ones (Stokoe 1960; Bellugi and Klima 1976; Klima and Bellugi 1979; Petitto 1994; Sandler and Lillo-Martin 2006), as well as similar overall neurobiological organization (Hickok et al. 1996; Neville et al. 1998; Petitto et al. 2000; Macsweeney et al. 2002; Emmorey et al. 2007; Mayberry et al. 2011). In particular, sign languages appear to rely on the same underlying lexical-syntactic and semantic systems as for spoken languages (MacSweeney et al. 2006; Leonard et al. 2012; Newman et al. 2015; Matchin et al. 2017a). While the lexical-syntactic and conceptual-semantic systems appear to be the same between speech and sign, potential differences in cortical organization concerning the frontal morpho-syntactic linearization system may derive from the distinct demands of producing sign and speech and could be investigated in future studies.
The Evidence
We next turn to a detailed examination of the evidence regarding the two major regions that potentially process syntax: the pIFG, or Broca’s area, and the posterior STS/pMTG. Across neuroimaging studies, these regions are most strongly implicated in syntactic processes. However, key pieces of neuroimaging data and the neuropsychological literature provide compelling insight into a functional dissociation of these two areas. Namely, the pMTG (and not pIFG) is consistently implicated in basic receptive syntactic processing, while both regions are implicated in sentence production, albeit distinctly.
Posterior inferior frontal gyrus
Grammatical deficits in aphasia were first documented in speech production (Kussmaul 1877) and are widely known as the syndrome of expressive agrammatism—the reduced use of function words and morphemes and simplified sentence structure (Jakobson 1956; Goodglass and Berko 1960; Alajouanine 1968). Despite this, most modern research and debate on the neurology of syntax has centered on comprehension. Indeed, as we noted in the introduction, the belief in a strong association between Broca’s area and syntax emerged in the 1970s following a study that showed what appeared to be a grammatical deficit in comprehension alongside expressive agrammatism in people with Broca’s aphasia (Caramazza and Zurif 1976). Such patients performed well on sentences that could be comprehended heuristically, but failed on sentences that required a syntactic algorithm. For example, canonical subject-verb-object word order sentences (in English), (1) and (3), could be successfully comprehended by adopting a heuristic that assumes the first noun is the agent of the action, and sentences with semantically nonreversible elements (1 and 2, girls can eat sushi but not the other way around) could be comprehended by a semantic plausibility heuristic. However, neither heuristic gets the right answer for noncanonical, semantically reversible sentences as in (4), which require a syntactic parse (see Grodzinsky 2000 for a review). The co-occurrence of expressive and receptive agrammatism suggested a central deficit in syntax, and the association between agrammatism and Broca’s aphasia yoked syntactic function to Broca’s area.
Nonreversible, subject-relative: “The girl that__devoured the sushi is smart” (good performance)
Nonreversible, object-relative: “The sushi that the girl devoured__is tasty” (good performance)
Reversible, subject-relative: “The girl that__pushed the boy is smart” (good performance)
Reversible, object-relative: “The girl that the boy pushed__is smart” (poor performance)
This initiated a long association between Broca’s area and syntactic processing, which included dozens of studies testing grammatical abilities in Broca’s aphasia (see Matchin and Rogalsky 2017 for a review). Subsequently, much neuroimaging data showed an association between sentence processing and brain activity for this region, along with the pMTG, particularly for sentences with noncanonical word order (such as 2, 4) compared to sentences with canonical word order (such as 1, 3), a contrast that was assumed to tax syntactic resources (see Meyer and Friederici 2016 for a meta-analysis and review).
Counter to this dominant modern narrative, however, we support the classical view—that the function of Broca’s area in syntax is primarily tied to production. There are several problems with the conclusion that damage to Broca’s area causes syntactic comprehension deficits. First, much of the research connecting syntactic deficits with damage to Broca’s area relies on an implicit assumption that patients with Broca’s aphasia have damage to this region. This assumption is problematic because damage restricted only to Broca’s area does not cause Broca’s aphasia; typically, patients with Broca’s aphasia have much larger lesions extending beyond Broca’s area (Mohr et al. 1978), potentially impinging on the temporal lobe (Fridriksson et al. 2015). This means that any putative syntactic comprehension deficits in Broca’s aphasia could derive from damage to other brain regions rather than Broca’s area. Even assuming such a link, people with agrammatic Broca’s aphasia generally succeed at making subtle acceptability judgments about the syntactic well-formedness of sentences (Linebarger et al. 1983; Wulfeck and Bates 1991; Wilson and Saygin 2004; see Matchin and Rogalsky 2017 for a review). Success in making acceptability judgments requires the use of some form of syntactic representations, indicating that so-called “agrammatic comprehension” is not a central loss of syntactic function.
More direct evidence against the proposal that Broca’s area is a core seat of syntactic ability for comprehension comes from lesion studies. Lesion-deficit mapping studies of general sentence comprehension do not highlight Broca’s area, but rather the AG, supramarginal gyrus, the temporal lobe (anterior and posterior portions), and occasionally more anterior parts of the IFG (pars orbitalis) (Dronkers et al. 2004; Baldo and Dronkers 2007; Thothathiri et al. 2012; Magnusdottir et al. 2013; Pillay et al. 2017; Fridriksson et al. 2018; Rogalsky et al. 2018). Intact sentence comprehension and syntactic acceptability judgments in the face of Broca’s area damage together speak strongly against a role for this region in basic syntactic processes.
While Broca’s area is not implicated in “basic” receptive sentence processing, there is variable evidence from the neuropsychological literature for a role in the comprehension of “complex” sentences. For example, some studies have reported an association between Broca’s area damage, or white matter tracts linking posterior temporal regions with Broca’s area, and deficits in comprehending semantically reversible noncanonical sentences (Amici et al. 2007; Wilson et al. 2011; Magnusdottir et al. 2013; Mesulam et al. 2015). Other studies have reported no such association (Dronkers et al. 2004; Race et al. 2012; Thothathiri et al. 2012; Rogalsky et al. 2018). A straightforward summary of the current state of evidence is that a link between Broca’s area and the processing of complex sentence comprehension exists—several studies have yielded positive results backed by relatively consistent functional imaging work—but is weak and variable. Such a pattern is inconsistent with proposals that assign a necessary syntactic computation to Broca’s area for comprehension (Friederici 2011; Hagoort 2014).
The findings are much more robust regarding the link between Broca’s area and language production. Large-scale lesion-deficit mapping studies have identified an association between damage primarily to Broca’s area (including pTri) and agrammatic production (Sapolsky et al. 2010; Wilson et al. 2010b; Wilson et al. 2011; Fridriksson et al. 2015; den Ouden et al. 2019) (Fig. 3, left). Electrical stimulation of Broca’s area, particularly pTri, induces agrammatic production (Chang et al. 2018). And, agrammatic production deficits can be observed following damage to dorsal white matter tracts connecting the IFG to the temporal lobe (Fridriksson et al. 2007; Wilson et al. 2011), suggesting that successful sentence production requires the interaction of processing in the IFG and the posterior temporal lobe.
Figure 3.
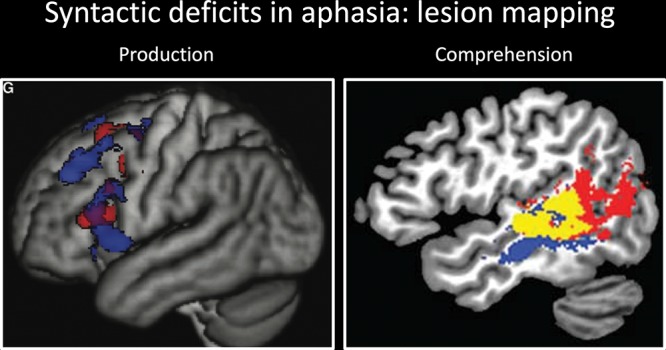
(Left) Results of a voxel-based morphometry study of sentence production (reproduced with permission from Wilson et al. 2010b). Brain regions where atrophy was correlated with lower scores on the composite syntactic measure indicating greater syntactic impairments (red) and reduced numbers of embeddings (blue). (Right) Results of a voxel-based lesion symptom mapping study of sentence comprehension (reproduced with permission from Pillay et al. 2017). Overlap (yellow) between regions associated with impaired auditory sentence comprehension (red) and impaired auditory description naming (blue) with picture naming scores as a covariate.
If the role of the pIFG is primarily tied to production, why does this region consistently activate for structural manipulations in neuroimaging studies, even for relatively simple structures (Pallier et al. 2011; Zaccarella and Friederici 2015; Matchin et al. 2017b; Zaccarella et al. 2017)? We have elsewhere suggested (in addition to other authors) that Broca’s area activation is driven by working memory resources, perhaps specialized for sentences, (Fiebach et al. 2005; Rogalsky and Hickok 2011. Rogalsky et al. 2015; Matchin 2017). Additionally, these frontal resources may be harnessed to facilitate predictions in some situations by preactivating lexical-syntactic representations stored in temporal cortex in a top-down fashion (Lau et al. 2006; Friederici 2012; Bonhage et al. 2015; Matchin et al. 2017b; Rimmele et al. 2018). This supposition is supported by the findings of Wright et al. (2012) who found that the benefit to reaction time of a word monitoring task due to syntactic context depended on tissue integrity of Broca’s area and the arcuate fasciculus (a white matter tract connecting frontal and posterior temporal cortex), as well as Jakuszeit et al. (2013), who found that the speed of the ERP component associated with subject–verb agreement violations was delayed in patients with pIFG lesions. Consistent with these proposals, patients with Broca’s aphasia (and left frontal lesions) show a boost to sentence comprehension performance when the rate of presentation is slowed (Love et al. 2008).
In summary, the evidence for the existence of a syntactic hub in Broca’s area is weak and variable for comprehension but much stronger for production, which primarily implicates the more anterior sector of Broca’s area, the pTri. This production–comprehension asymmetry mirrors a similar state of affairs at the phonological level where there is equivocal evidence at best for a strong role in speech perception (despite many claims for a causal role) (Hickok 2014b), and robust evidence for a critical role in phonological stages of speech production, particularly involving the posterior sector of Broca’s area, the pars opercularis (see Hickok and Poeppel 2007 for review).
Posterior middle temporal gyrus
In the previous section, we argued that the link between syntactic processing and Broca’s area in the neuropsychological literature holds primarily for language production, not comprehension. Here we review evidence that shows strong associations between syntactic processing and the pMTG for both comprehension and production.
With respect to comprehension, neuroimaging studies find that the pMTG and pIFG consistently exhibit effects of structure for semantically impoverished sentences (Pallier et al. 2011; Fedorenko et al. 2012a; Goucha and Friederici 2015; Matchin et al. 2017b). These regions also show effects of structural complexity that cannot be accounted for by verbal working memory (Caplan et al. 2000; Rogalsky et al. 2008a, 2015; Fedorenko et al. 2012b,Matchin et al. 2017b). There are, however, key studies that illustrate asymmetries between these regions—namely, that the posterior temporal lobe is associated with hierarchical processing while Broca’s area is associated with linear processing. A series of fMRI studies by Tyler and colleagues has found that pMTG activates for simple phrasal stimuli, but that the response in pIFG is preferentially driven by inflected word forms, which rely on linear morpho-syntactic computations (Tyler et al. 2004, 2005; Bozic et al. 2015). Additionally, a recent fMRI study by Brennan et al. (2016) localized language-responsive regions of interest in individual subjects and then tested parsing models with an increasing degree of hierarchical information, from linear sequences, to simple hierarchical phrase structure, and finally to more complex transformed syntactic structures. They identified effects of linear sequence in all of the language regions tested, with additional hierarchical effects (both for simple and more complex phrase structure) in pMTG and ATL, but no hierarchical effects at all in pIFG. Finally, an electrocorticography (ECoG) study by Nelson et al. (2017) found that the response of electrodes in the pSTS (but not ATL and IFG) had strong correlations with predictive parsing models that plausibly underlie hierarchical syntactic computations.
In contrast to the weak and variable effects in the pIFG, damage to the pMTG is robustly associated with basic sentence comprehension deficits across many studies (Dronkers et al. 2004; Amici et al. 2007; Baldo and Dronkers 2007; Race et al. 2012; Magnusdottir et al. 2013; Pillay et al. 2017; Fridriksson et al. 2018; Rogalsky et al. 2018; Wilson et al. 2018b, 2018c). Importantly, such deficits cannot be simply attributed to word-level deficits as single word comprehension impairments are rare and typically mild following left unilateral injury (Rogalsky et al. 2008b), even when encompassing posterior temporal cortex (Selnes et al. 1984; Wilson et al. 2018b, 2018c). In contrast, sentence comprehension deficits are more readily detected, even for simple experimental tasks (Dronkers et al. 2004; Wilson et al. 2018b, 2018c) that do not place high demands on working memory or executive function resources.
A recent lesion-deficit mapping study by Pillay et al. (2017) provides compelling evidence (Fig. 3, right). These authors employed two types of tasks. One was a picture naming task, which engages word-level phonological, lexical, and semantic processes as well as articulatory and executive functions. The other was sentence comprehension: first, a naming-to-auditory description task (e.g., stimulus: “what a king wears on his head”, target answer: “crown”), which in addition to processes engaged in single word naming also engages syntactic processes involved in sentence-level comprehension. Second, they also employed an auditory sentence comprehension task, in which subjects decided whether an auditorily presented sentence matched a video animation. Picture naming is known to implicate superior and middle temporal gyri (Indefrey and Levelt 2004; Piras and Marangolo 2007; Indefrey 2011; Baldo et al. 2013), presumably reflecting phonological and lexical processes, respectively (Indefrey and Levelt 2004; Hickok and Poeppel 2007; Indefrey 2011). Pillay et al. looked at the lesions associated with the naming-to-description task and the auditory sentence comprehension task once the effects of picture naming were covaried out from both tasks, thus factoring out single-word processing and isolating sentence-level comprehension. The overlap for the affected areas for sentence processing (with picture naming regressed out) centered on the pMTG. The IFG was not implicated by either task, reinforcing our claims about the lack of a role of the IFG in sentence comprehension. This means that while pMTG is clearly involved in single word lexical processing (see Lau et al. 2008 for a review), it is also involved in sentence comprehension above and beyond single words, presumably reflecting the extra load and/or additional representations involved in sentence processing.
Neuropsychological evidence for a link between syntax and the pMTG in sentence production comes in the form of a symptom of aphasia known as “paragrammatism”. As early as 1914, Karl Kleist recognized two disturbances of word order (“störungen der wortfolge”), one being agrammatism and the other paragrammatism (Kleist 1914). Here is how he characterized them (translation from Papathanasiou and Coppens 2017, p. 26):
The basic trait of agrammatism is the simplification and coarsening of word sequences. Complicated compound sentences (subordination of clauses) are not built. The patients only speak in small, primitive mini-sentences, if they continue to create sentences at all. All less necessary words, especially pronouns and particles, are reduced or eliminated … .
An example from Goodglass (1993):
Examiner: What brought you to the hospital?
Patient: Yeah … Wednesday, ... Paul and dad … Hospital … yeah … doctors, two … .an’ teeth.
In paragrammatism, Kleist continues,
the ability to create words orders is not abolished, but phrases and sentences are often wrongly chosen and thereby amalgamate and contaminate each other … phrases and sentence constructions are not completed. … The spoken expression is not simplified overall, instead, also conditioned by a strong overproduction of word sequences, it swells to confused sentence monsters (pp. 10–11)
Goodglass (1993) provides some examples of such “sentence monsters”:
“Well, all I know is, somebody is clipping the kreples and some wha, someone here on the kureping arm, … why I don’t know.” (p. 86)
“I feel very well. My hearing, writing been doing well. Things that I couldn’t hear from. In other words, I used to be able to work cigarettes I didn’t know how … . Chesterfeela, for 20 years I can write it.” (p. 86)
Other paragrammatic errors are illustrated by Butterworth and Howard (1987):
“And I want everything to be so talk”
“She was handled to look at the books a bit”
“I’m very want it”
“Isn’t look very dear, is it?”
“But it’s silly, aren’t they?”
The existence of these two disorders suggests that brain injury can disrupt syntactic ability in speech production and can do so in two ways, broadly speaking: one that severely reduces grammatical structure (agrammatism) and another that leaves relatively intact the ability to generate syntactic sequences, but at a higher error rate (paragrammatism). Agrammatism is associated with nonfluent aphasia (e.g., Broca’s aphasia), relatively preserved language comprehension, and frontoparietal lesions (as discussed above), whereas paragrammatism is associated with fluent aphasia (Wernicke’s aphasia, conduction aphasia), relatively poor comprehension, and posterior temporo-parietal lesions (Goodglass 1993; Dronkers and Baldo 2009; Henseler et al. 2014; Fridriksson et al. 2018). Supporting this distinction, Casilio et al. (2019) performed a factor analysis in which paragrammatism was associated with empty speech, semantic paraphasias and neologisms (unattested word forms) but was independent of agrammatism.
While several studies have implicated primarily the pIFG in agrammatic production (Sapolsky et al. 2010; Wilson et al. 2010b; Wilson et al. 2011; Chang et al. 2018; den Ouden et al. 2019), there is comparatively less direct evidence regarding the localization of paragrammatic deficits. Its association with fluent aphasia, however, implicates posterior temporal–parietal regions. Yagata et al. (2017) report two case studies of paragrammatic patients, both with confirmed damage to the posterior temporal and inferior parietal lobes and no frontal lobe damage. In addition, Wilson et al. present several case studies in which paragrammatism and sentence comprehension deficits were associated with posterior temporal damage (see patients included in both Wilson et al. 2018b, 2018c).
To summarize this section, the pMTG is a stronger candidate than the pIFG for a region supporting basic hierarchical syntactic processes. It is consistently implicated in the comprehension of even simple sentences in neuropsychological and neuroimaging studies, shows classic sentence complexity effects, is correlated in activity with parsing operations consistent with hierarchical processing, and when damaged appears to result in a form of syntactic production deficit, paragrammatism.
The Comprehension–Production Asymmetry in Syntactic Processing
The evidence we reviewed converges on the pMTG as a hub for hierarchical lexical-syntactic processing, which plays a role in both sentence comprehension and production. The pIFG, in contrast, is associated primarily with production and plays a critical role, we argue, in morpho-syntactic sequencing. Here we elaborate this aspect of our proposal.
We have characterized the job of syntax as the process of transforming a linear sequence of elements into a nonlinear semantic representation and back again. But as we pointed out in the preceding paragraphs, the computational task of encoding a semantic message into a linguistic form is not the same as that of decoding a message from the speech stream. Specifically, when listening to speech, the job of a parser is to recover the most probable structure given an observed sequence, a set of syntactic constraints (the rules of syntax), and the top-down discourse/semantic constraints that are available. It has been noted that sentence comprehension typically does not strictly require reconstructing all linguistic details of an utterance (Garrett 1980; Bock 1995), requiring only a “good-enough” representation (Ferreira et al. 2002). In the present context, we emphasize that the linear order of morphemes is given and does not need to be explicitly recomputed or maintained in the parse. That is, perceived morpheme order is used in the pMTG to construct a hierarchical representation and is then quickly discarded as the parse proceeds. This mechanism is sufficient for most sentence comprehension situations. However, in some cases, such as when a reanalysis or repair of a parse is needed, the capacity to recompute the linear sequence via interaction with frontal networks (see below) can aid receptive parsing by providing a mental rewind button. Such a mechanism may also serve a predictive coding function.
Producing speech or sign presents a different problem. The starting point is a nonsequential semantic representation that must be translated into a sequence of words and phonemes (Roelofs and Ferreira, 2019; Levelt 1989). In contrast to comprehension, the sequence is unknown and must be computed. In what follows, we propose an architecture for this computation.
We propose that production at the morpho-syntactic level is carried out using an architecture that is homologous to that proposed for the phonological level (Hickok et al. 2011; Hickok 2012a, 2014a), which in turn is homologous to architectures for sensorimotor control generally (Shadmehr and Krakauer 2008). The basic plan for such architectures is that sensory-related cortex codes targets for output planning, which typically involves sequence coding. In visuomotor control, sensory targets might be objects of a particular size, shape, and location. The computational task in motor planning is to transform visual-based features into a motor plan to “hit” those targets (size, shape, and location) with an appropriate sequence of reach and grasp actions of the limb. In phonological output planning, sensory targets are stored memories of the sound pattern of a word. The computational task here is to transform high-level auditory speech features into a sequence of motor speech gestures that will reproduce a sequence of sounds that will “hit” the sound pattern target. In syntactic output planning, we propose that the targets are hierarchical forms (configurations of treelets). The computational task is to transform the hierarchical configurations into a sequence of morphemes that, when decoded by a listener, will reconstruct the same hierarchical configuration. This conservation of computational architectures, we believe, is sensible from an evolutionary standpoint in that it provides a framework for developing hypotheses about how language systems might have evolved from sensorimotor networks. Indeed, we suggest a common “computational ancestor” for these different forms of output planning; the systems are architecturally and computationally homologous.
In Figure 2, note the hierarchical arrangement of the syntactic system (higher level) compared to the phonological system (lower level), which both involve “sensorimotor” loops (where the term sensorimotor is used in the sense of the computational and network homology). The dark blue, green, and red nodes together comprise the comprehension network. We assume that in comprehension, the lexical-phonological network processes each speech input, sequentially activating lexical-syntactic representations in the pMTG that are integrated into a broader nonsequential relational hierarchy and related to the conceptual network for comprehension. The language production network comprises all of the displayed components. Production starts with a conceptual representation in ATL/AG (red) that is transformed into a nonlinear hierarchical representation coded in the pMTG (green). The hierarchical representations in turn serve as the target for a corresponding morphosyntactic sequence (yellow) that is computed via interaction with inferior frontal networks, the pTri in particular. This proposal is consistent with evidence for frontal involvement in sequential processing in artificial grammars, meaningless speech sequences, and music (Gelfand and Bookheimer 2003; Friederici et al. 2006; Flöel et al. 2009; Leaver et al. 2009; Bohland et al. 2010; Petersson et al. 2012; Segawa et al. 2015; Rong et al. 2018).
We propose a direct link between conceptual-semantic representations (red) and the morpho-syntactic sequencing network (yellow) based on the following theoretical and empirical considerations. If the only pathway for activating the frontal sequencing network passes through the hierarchical lexical-syntactic system, then damage to that system should dramatically reduce sentence-level speech output. Instead, when damage occurs to the posterior temporal lobe, one observes grammatical impairments, but output remains fluent (even prodigious). There must be some means to activate morpho-syntactic sequences without involvement of an intact hierarchical lexical-syntactic system. We therefore propose the existence of direct links between the semantic system and the sequence coding network as the culprit. In essence, we suggest the existence of something like “comprehension lemmas” (lexical-syntactic treelets of the pMTG) and “production lemmas” (linear morphological sequence “chunks” of the pTri) (see Caramazza 1986 for neuropsychological evidence regarding the existence of both an input and an output lexicon), and that both systems can be driven by activation in the conceptual-semantic system. Similar arguments have been made for speech production at the phonological level (Hickok et al. 2011; Hickok 2012a).
Why is the direct semantic-to-morpheme sequence pathway insufficient for normal grammatical production? Because it lacks a hierarchical target that constrains the form of the sequence. Why would such a direct pathway exist at all? It would be useful and sufficient for idiomatic or automated expressions (“How are you today?”), and it could improve efficiency by preactivating high-frequency sequence chunks similar to motor sequence chunking generally (a form of predictive coding) that can be fine-tuned or integrated with a sentence context via interaction with the lexical-syntactic network. Again, analogous arguments have been made at the phonological level (Hickok et al. 2011; Hickok 2012a) and have a long history, dating back to the 19th century (Wernicke 1874/1969). Here we extend it to syntactic production (again, see Caramazza 1986).
Consider, now, the distinction between agrammatism and paragrammatism in light of this theoretical framework. Paragrammatism results from impairment of hierarchical lexical-syntactic representations, but intact morpho-syntactic sequencing. Output is fluent because direct conceptual selection of morphological sequences is intact, but it is prone to “sentence monsters” and other syntactic errors because it is unconstrained by hierarchical relationships. An illustration comes from agreement errors in paragrammatic speech, e.g., a mismatch in number feature between the subject and the verb, as in “the birds was a color” (reported in Yagata et al. 2017). Agreement can be arbitrarily linearly far apart, as shown in (1–3):
the man sleep-s
the man that had a dozen hats sleep-s
the man from Tallahassee that had a dozen hats sleep-s
The local hierarchical relation between the subject and the verb (that is preserved regardless of linear distance) is critical for selecting the appropriate agreement morpheme -s. Thus, in English-speaking paragrammatic patients, we hypothesize that the affixation rules for attaching an agreement morpheme onto a verb are intact (relying on the frontal morpho-syntactic sequencing system), but that the hierarchical relationship that constrains the selection of the appropriate agreement feature is impaired. Agrammatism, by contrast, results from damage to the sequencing mechanism itself, thus dramatically reducing the ability to sequence much of anything at all, including a striking reduction in the presence of inflectional morphology. Relatively preserved ability to judge the grammaticality of perceived sentences in agrammatism (Linebarger et al. 1983) is explained on this account by an intact hierarchical lexical-syntactic system, the comprehension side of the network. Consistent with this model, we note evidence linking impairments in grammaticality judgments primarily to posterior temporal lobe damage (Wilson and Saygin 2004).
If paragrammatism results from impaired syntactic hierarchies, shouldn’t sentence output exhibit some degree of reduced syntactic complexity, in contrast to Kleist’s claim of no simplification? Indeed it should and indeed it does, although the reduction appears to be in the form of a reduced frequency of complex structures rather than an absolute inability to generate complex syntax on any attempt (Bates and Devescovi 1989; Goodglass 1993; Bastiaanse et al. 1996). Interestingly, this parallels what has been observed in phonological ability in fluent aphasia (conduction aphasia, in particular): phonological errors are relatively common and exacerbated by phonological complexity or lower frequency forms, yet examples of error-free complex or low frequency phonological forms are produced nonetheless (Goodglass 1992). At the phonological level, this is attributed to damage to an error detection and correction mechanism rather than phonological sequence coding; the assumption is that the probability of a sequence coding error increases with complexity (Hickok et al. 2011; Hickok 2012b). Thus, the deficit leads to greater difficulty with more complex or infrequent forms but does not preclude their occasional correct output. A similar mechanism is at work for syntax, we propose. Morphological sequences can be generated from semantic representations, but the probability of an error increases with greater complexity/lower frequency because the sequences cannot be constrained by interaction with hierarchical representations.
Our account, in which lexical representations and syntactic hierarchies are part of the same system encoded in the pMTG, also explains why paragrammatic speech is often semantically impoverished (e.g., “Things that I couldn’t hear from”) and exhibits a healthy dose of lexical selection errors (e.g., “I used to be able to work cigarettes”) (Bastiaanse et al. 1996; Casilio et al. 2019). Both symptoms could stem from lexical-level problems, which would lead to access “failures” and the overuse of filler words (e.g., “things”), as well as access “errors” leading to a lexical substitution. The co-occurrence of lexical and syntactic errors makes sense in light of the proposed lexical/syntactic hub.
Conclusions, Future Directions, and Loose Ends
The neuroanatomical model of syntax we have proposed here integrates research in a wide variety of domains: theoretical linguistics, psycholinguistics, computational modeling, neuroimaging, and neuropsychology. This ambition leaves many important data points and theoretical perspectives unaddressed. However, approaching the problem of syntax in the brain with conceptual and theoretical motivations regarding the fundamental nature of syntax as connecting sound and meaning, and as connecting distinct semantic systems with each other, has helped to piece together many disparate empirical findings across domains. One of the biggest challenges in the cognitive neuroscience of syntax is reconciling theories of syntax in the brain, usually motivated by neuroimaging data, with conflicting neuropsychology data. We have shown here that a role for the posterior temporal lobe in hierarchical lexical-syntactic processing and inferior frontal cortex in linear morpho-syntactic sequencing successfully resolves this conflict. This distinction allows us to make the following empirical predictions:
Large-scale lesion-deficit mapping studies should reveal a double dissociation between agrammatic and paragrammatic production: agrammatic production follows from damage to Broca’s area, while paragrammatic production follows from posterior temporal damage.
Neuroimaging studies of sentence production, using computational models that specify distinct stages of hierarchical and linear syntactic processing, should identify correlations between hierarchical planning processes with activation in pMTG and linear planning processes with activation in pTri.
Neuroimaging studies of syntactic comprehension using tasks that encourage subjects to parse linear morpho-syntactic information vs. simple tasks that focus on semantic interpretation without requiring explicit linear parsing will differentially recruit pTri, but more equally recruit pMTG.
An additional question that we have not so far addressed is how to reconcile the focal picture of syntactic processing painted by the neuroimaging literature with the coarser picture painted by the neuropsychological literature. Earlier we noted that chronic linguistic deficits in aphasia typically do not result from focal damage to the IFG, requiring broader lesions to produce full-blown Broca’s aphasia (Mohr 1978; Fridriksson et al. 2015). We have claimed that linearization processes in syntax are tied to a subregion of Broca’s area, the pTri, and one may wonder whether the lesion evidence is a problem for our claim: if damage restricted to Broca’s area does not produce Broca’s aphasia, then presumably it does not produce agrammatism, as we would expect given our localization claims. We have two comments on this issue. One is that restricted damage to Broca’s area may have an effect on syntax, if not profoundly. Commenting on the effects of lesions restricted to Broca’s area, Benson and Ardila (1996) write, “Although these patients can produce an occasional long phrase, phrase length is usually shortened; syntax is restricted (simplified) and occasionally incorrect; a full picture of agrammatism, however, is not observed.” (p. 127). The fact that “some” degree of syntactic difficulty is evident even with focal lesions to Broca’s area is consistent with our proposal. What remains to be explained is why the deficit is not severe. One likely possibility is that the computational machinery we ascribe to the pTri is not as focal as the functional imaging evidence suggests. A limitation of hemodynamic methods is that across studies they will tend to identify only the most robust and temporally sustained effects, perhaps missing key contributions from other regions that have activations that are subthreshold or more variable localization from one participant to the next. Indeed, one study that used ECoG (affording precise spatial and temporal resolution) identified a much broader network of cortical regions engaged by syntactic mechanisms (Nelson et al. 2017). This is an important issue to be addressed in future research.
Finally, our model raises questions about the evolution and development of syntax and semantics in the human brain. The data appear to suggest that the syntactic function of the pMTG may be in part innate. The STS, as part of pMTG, is a general anatomical region that is greatly expanded in humans relative to nonhuman primates (Hill et al. 2010), as are white matter tracts connecting the frontal lobe to pMTG (Rilling et al. 2008). The specific morphology of the STS, namely its hemispheric asymmetry in depth, is unique to humans (Leroy et al. 2015) and is present in 3-month-old infants (Glasel et al. 2011), suggesting a genetic component. Also, if the lexical-syntactic system derives its properties solely through the emergent connections between sensory and semantic systems, then one would expect a distinct cortical locus between auditory-vocal and visual-manual languages. However, syntactic effects for spoken languages and sign languages occur in apparently the same portions of the ventral STS (Pallier et al. 2011; Matchin et al. 2017a). How these evolutionary changes relate to the syntactic function of pMTG remains a tangible mystery to be explored. This does not suggest that the entire system is genetically determined—the specific hierarchical structures in a given language vary across languages, and must be acquired through experience. Late environmental exposure to language in development is correlated with severe language deficits (Mayberry et al. 2002) and reduced activations in posterior temporal and inferior frontal brain regions for sentence comprehension in adulthood (Mayberry et al. 2011). This suggests a crucial interaction of genes and environment for the syntactic properties of the pMTG to emerge.
Similar questions concern the morpho-syntactic sequencing function of the pIFG, pTri (Matchin 2017). The dorsal white matter tract connecting the pMTG to the pIFG, the arcuate fasciculus, is relatively immature in young children relative to adults compared to other white matter pathways (Dubois et al. 2006, 2008), suggesting that the link between these regions is constructed in development and may drive the sentence-specificity of activations in the pTri (Fedorenko et al. 2012b). However, without direct evidence, such proposals remain speculative and need to be confirmed with research examining the degree of sentence-specificity occurring in both of these regions across the lifespan.
Funding
W.M. was supported by the National Institutes of Health (grant DC012797 to Rachel Mayberry); G.H. was supported by the National Institutes of Health (grant DC014664).
Notes
For feedback on this work, we would like to thank Stephen Wilson, Shota Momma, Dirk den Ouden, Julius Fridriksson, Rachel Mayberry, Eric Halgren, Ellen Lau, Norbert Hornstein, Robert Berwick, the University of Maryland Linguistics Dept., and the audience of the 2017 Society for the Neurobiology of Language meeting held in Baltimore, and an anonymous reviewer. Any errors or omissions are entirely ours. Conflict of Interest: None declared.
References
- Adger D. 2018. The autonomy of syntax In: Hornstein N, Lasnik H, Patel-Grosz P, Yang C, editors. Syntactic structures after 60 years: the impact of the chomskyan revolution in linguistics. Berlin: Walter de Gruyter GmbH. Vol 129, p. 153.
- Alajouanine T. 1968. L’aphasie et le langage pathologique. Paris: J.B. Balliere. [Google Scholar]
- Amici S, Brambati SM, Wilkins DP, Ogar J, Dronkers NL, Miller BL, Gorno-Tempini ML. 2007. Anatomical correlates of sentence comprehension and verbal working memory in neurodegenerative disease. J Neurosci. 27:6282–6290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldo JV, Dronkers NF. 2007. Neural correlates of arithmetic and language comprehension: a common substrate? Neuropsychologia. 45:229–235. [DOI] [PubMed] [Google Scholar]
- Baldo JV, Arevalo A, Patterson JP, Dronkers NF. 2013. Grey and white matter correlates of picture naming: evidence from a voxel-based lesion analysis of the Boston naming test. Cortex. 49:658–667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bastiaanse R, Edwards S, Kiss K. 1996. Fluent aphasia in three languages: aspects of spontaneous speech. Aphasiology. 10:561–575. [Google Scholar]
- Bates E, Devescovi A. 1989. Crosslinguistics studies of sentence production In: MacWhinney B, Bates E, editors. The crosslinguistics study of sentence processing. Cambridge: Cambridge University Press, pp. 225–253. [Google Scholar]
- Beckers GJL, Bolhuis JJ, Okanoya K, Berwick RC. 2012. Birdsong neurolinguistics: songbird context-free grammar claim is premature. NeuroReport. 23:139–145. [DOI] [PubMed] [Google Scholar]
- Bellugi U, Klima ES. 1976. Two faces of sign: iconic and abstract. Annals New York Acad Sci. 280:514–538. [DOI] [PubMed] [Google Scholar]
- Benson DF, Ardila A. 1996. Aphasia: a clinical perspective. Oxford: Oxford University Press. [Google Scholar]
- Berwick RC, Okanoya K, Beckers GJ, Bolhuis JJ. 2011. Songs to syntax: the linguistics of birdsong. Trends Cogn Sci. 15:113–121. [DOI] [PubMed] [Google Scholar]
- Berwick RC, Chomsky N. 2016. Why only us: language and evolution. Cambridge (MA): MIT press. [Google Scholar]
- Binder JR, Desai RH. 2011. The neurobiology of semantic memory. Trends Cogn Sci. 15:527–536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Binder JR, Desai RH, Graves WW, Conant LL. 2009. Where is the semantic system? A critical review and metaanalysis of 120 functional neuroimaging studies. Cereb Cortex. 19:2767–2796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Binder JR, Frost JA, Hammeke TA, Bellgowan PS, Springer JA, Kaufman JN, Possing ET. 2000. Human temporal lobe activation by speech and nonspeech sounds. Cereb Cortex. 10:512–528. [DOI] [PubMed] [Google Scholar]
- Blank I, Balewski Z, Mahowald K, Fedorenko E. 2016. Syntactic processing is distributed across the language system. NeuroImage. 127:307–323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bock K. 1995. Sentence production: from mind to mouth In: Miller JL, Eimas PD, editors. Speech, language, and communication. London: Academic Press. [Google Scholar]
- Boeckx C, Martinez-Alvarez A, Leivada E. 2014. The functional neuroanatomy of serial order in language. J Neurolinguis. 32:1–5. [Google Scholar]
- Bohland JW, Bullock D, Guenther FH. 2010. Neural representations and mechanisms for the performance of simple speech sequences. J Cogn Neurosci. 22:1504–1529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolhuis JJ, Tattersall I, Chomsky N, Berwick RC. 2014. How could language have evolved? PLoS Biol. 12(8):e1001934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonhage CE, Mueller JL, Friederici AD, Fiebach CJ. 2015. Combined eye tracking and fMRI reveals neural basis of linguistic predictions during sentence comprehension. Cortex. 68:33–47. [DOI] [PubMed] [Google Scholar]
- Bornkessel I, Zysset S, Friederici AD, Von Cramon DY, Schlesewsky M. 2005. Who did what to whom? The neural basis of argument hierarchies during language comprehension. Neuroimage. 26:221–233. [DOI] [PubMed] [Google Scholar]
- Bornkessel-Schlesewsky I, Schlesewsky M. 2013. Reconciling time, space and function: a new dorsal–ventral stream model of sentence comprehension. Brain Lang. 125:60–76. [DOI] [PubMed] [Google Scholar]
- Bornkessel-Schlesewsky I, Schlesewsky M, Small SL, Rauschecker JP. 2015. Neurobiological roots of language in primate audition: common computational properties. Trends Cogn Sci. 19:142–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boylan C, Trueswell JC, Thompson-Schill SL. 2015. Compositionality and the angular gyrus: a multi-voxel similarity analysis of the semantic composition of nouns and verbs. Neuropsychologia. 78:130–141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boylan C, Trueswell JC, Thompson-Schill SL. 2017. Relational vs. attributive interpretation of nominal compounds differentially engages angular gyrus and anterior temporal lobe. Brain Lang. 169:8–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bozic M, Fonteneau E, Su L, Marslen-Wilson WD. 2015. Grammatical analysis as a distributed neurobiological function. Hum Brain Mapp. 36:1190–1201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brennan JR, Stabler EP, Van Wagenen SE, Luh WM, Hale JT. 2016. Abstract linguistic structure correlates with temporal activity during naturalistic comprehension. Brain Lang. 157:81–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bresnan J. 2001. Lexical-functional syntax. Malden (MA): Blackwell. [Google Scholar]
- Butterworth B, Howard D. 1987. Paragrammatisms. Cognition. 26(1):1–37. [DOI] [PubMed] [Google Scholar]
- Call J, Tomasello M. 2008. Does the chimpanzee have a theory of mind? 30 years later. Trends Cogn Sci. 12:187–192. [DOI] [PubMed] [Google Scholar]
- Caplan D, Alpert N, Waters G, Olivieri A. 2000. Activation of Broca's area by syntactic processing under conditions of concurrent articulation. Hum Brain Mapp. 9:65–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caramazza A, Zurif EB. 1976. Dissociation of algorithmic and heuristic processes in language comprehension: evidence from aphasia. Brain Lang. 3:572–582. [DOI] [PubMed] [Google Scholar]
- Caramazza A. 1986. The structure of the lexical system: evidence from acquired language disorders. Clin Aphasiol. 16:291–301. [Google Scholar]
- Caramazza A. 2000. The organization of conceptual knowledge in the brain In: Gazzaniga MS, editor. The new cognitive neurosciences. 2nd ed. Cambridge (MA): MIT Press, pp. 1037–1046. [Google Scholar]
- Casilio M, Rising K, Beeson PM, Bunton K, Wilson SM. 2019. Auditory-perceptual rating of connected speech in aphasia. Am J Speech Lang Pathol. 28(2):550–568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang EF, Rieger JW, Johnson K, Berger MS, Barbaro NM, Knight RT. 2010. Categorical speech representation in human superior temporal gyrus. Nat Neurosci. 13:1428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang EF, Kurteff G, Wilson SM. 2018. Selective interference with syntactic encoding during sentence production by direct Electrocortical stimulation of the inferior frontal gyrus. J Cogn Neurosci. 30:411–420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cherniak C. 1994. Component placement optimization in the brain. J Neurosci. 14:2418–2427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chklovskii DB, Koulakov AA. 2004. Maps in the brain: what can we learn from them? Annu Rev Neurosci. 27:369–392. [DOI] [PubMed] [Google Scholar]
- Chklovskii DB, Schikorski T, Stevens CF. 2002. Wiring optimization in cortical circuits. Neuron. 34:341–347. [DOI] [PubMed] [Google Scholar]
- Carroll L. 1871. Through the looking glass: and what Alice found there. Rand: McNally. [Google Scholar]
- Chomsky N. 1957. Syntactic structures. The Hague: Mouton. [Google Scholar]
- Chomsky N. 1965. Aspects of the theory of syntax. Cambridge (MA): MIT Press. [Google Scholar]
- Chomsky N. 1981. Lectures on government and binding. Dordrecht: Foris. [Google Scholar]
- Chomsky N. 1994. Bare phrase structure. Vol 8 Cambridge (MA): MIT Press. [Google Scholar]
- Chomsky N. 1995. The minimalist program. Cambridge (MA): MIT Press. [Google Scholar]
- Cotelli M, Borroni B, Manenti R, Ginex V, Calabria M, Moro A et al. 2007. Universal grammar in the frontotemporal dementia spectrum: evidence of a selective disorder in the corticobasal degeneration syndrome. Neuropsychologia. 45:3015–3023. [DOI] [PubMed] [Google Scholar]
- Damasio AR. 1989. The brain binds entities and events by multiregional activation from convergence zones. Neural Comput. 1:123–132. [Google Scholar]
- Darwin C. 1871. The Descent of Man, and Selection in Relation to Sex (second edition). London: John Murray. Reprinted in Penguin Classics 2004, London: Penguin.
- Dell GS, O'Seaghdha PG. 1992. Stages of lexical access in language production. Cognition. 42:287–314. [DOI] [PubMed] [Google Scholar]
- Del Prato P, Pylkkänen L. 2014. MEG evidence for conceptual combination but not numeral quantification in the left anterior temporal lobe during language production. Front Psychol. 5:524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demberg V, Keller F, Koller A. 2013. Incremental, predictive parsing with psycholinguistically motivated tree-adjoining grammar. Computat Linguis. 39:1025–1066. [Google Scholar]
- den Ouden DB, Malyutina S, Basilakos A, Bonilha L, Gleichgerrcht E, Yourganov G, Hillis AE, Hickok G, Rorden C, Fridriksson J. 2019. Cortical and structural-connectivity damage correlated with impaired syntactic processing in aphasia. Human brain mapping, 40(7): 2153–2173. [DOI] [PMC free article] [PubMed]
- De Waal FB. 2008. Putting the altruism back into altruism: the evolution of empathy. Annu Rev Psychol. 59:279–300. [DOI] [PubMed] [Google Scholar]
- DeWitt I, Rauschecker JP. 2012. Phoneme and word recognition in the auditory ventral stream. Proc Natl Acad Sci. 109:E505–E514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding N, Melloni L, Zhang H, Tian X, Poeppel D. 2016. Cortical tracking of hierarchical linguistic structures in connected speech. Nat Neurosci. 19:158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dronkers N, Baldo J. 2009. Language: aphasia In: Squire LR, editor. Encyclopedia of neuroscience. Vol 5 Oxford: Academic Press, pp. 343–348. [Google Scholar]
- Dronkers NF, Wilkins DP, Valin RD, Redfern BB, Jaeger JJ. 2004. Lesion analysis of the brain areas involved in language comprehension. Cognition. 92:145–177. [DOI] [PubMed] [Google Scholar]
- Dubois J, Hertz-Pannier L, Dehaene-Lambertz G, Cointepas Y, Le Bihan D. 2006. Assessment of the early organization and Matura- tion of infants' cerebral white matter fiber bundles: a feasibility study using quantitative diffusion tensor imaging and tractography. NeuroImage. 30(4):1121–1132. [DOI] [PubMed] [Google Scholar]
- Dubois J, Dehaene-Lambertz G, Perrin M, Mangin JF, Cointepas Y, Duchesnay E et al. 2008. Asynchrony of the early maturation of white matter bundles in healthy infants: quantitative landmarks revealed noninvasively by diffusion tensor imaging. Human Brain Mapping. 29(1):14–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Embick D, Poeppel D. 2015. Towards a computational (ist) neurobiology of language: correlational, integrated and explanatory neurolinguistics. Lang Cogn Neurosci. 30:357–366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emmorey K, Mehta S, Grabowski TJ. 2007. The neural correlates of sign versus word production. Neuroimage. 36:202–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Estes Z, Golonka S, Jones LL. 2011. Thematic thinking: the apprehension and consequences of thematic relations. Psychol Learn Motiv. 54:249–294. [Google Scholar]
- Everaert MB, Huybregts MA, Chomsky N, Berwick RC, Bolhuis JJ. 2015. Structures, not strings: linguistics as part of the cognitive sciences. Trends Cogn Sci. 19:729–743. [DOI] [PubMed] [Google Scholar]
- Fedorenko E, Duncan J, Kanwisher N. 2012b. Language-selective and domain-general regions lie side by side within Broca’s area. Curr Biol. 22:2059–2062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fedorenko E, Nieto-Castañon A, Kanwisher N. 2012a. Lexical and syntactic representations in the brain: an fMRI investigation with multi-voxel pattern analyses. Neuropsychologia. 50:499–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fedorenko E, Mineroff Z, Siegelman M, Blank I. 2018 (preprint). Word meanings and sentence structure recruit the same set of fronto-temporal regions during comprehension. BioRxiv. [Google Scholar]
- Ferreira F, Bailey KG, Ferraro V. 2002. Good-enough representations in language comprehension. Curr Dir Psychol Sci. 11:11–15. [Google Scholar]
- Fiebach CJ, Schlesewsky M, Lohmann G, Von Cramon DY, Friederici AD. 2005. Revisiting the role of Broca's area in sentence processing: syntactic integration versus syntactic working memory. Hum Brain Mapp. 24:79–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fitch WT. 2017. Empirical approaches to the study of language evolution. Psychon Bull Rev. 24:3–33. [DOI] [PubMed] [Google Scholar]
- Fletcher PC, Happe F, Frith U, Baker SC, Dolan RJ, Frackowiak RS, Frith CD. 1995. Other minds in the brain: a functional imaging study of “theory of mind” in story comprehension. Cognition. 57:109–128. [DOI] [PubMed] [Google Scholar]
- Flöel A, de Vries MH, Scholz J, Breitenstein C, Johansen-Berg H. 2009. White matter integrity in the vicinity of Broca’s area predicts grammar learning success. Neuroimage. 47:1974–1981. [DOI] [PubMed] [Google Scholar]
- Frank R. 2002. Phrase structure composition and syntactic dependencies. Cambridge (MA): MIT Press. [Google Scholar]
- Fridriksson J, Bonilha L, Rorden C. 2007. Severe Broca’s aphasia without Broca’s area damage. Behav Neurol. 18:237–238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fridriksson J, Fillmore P, Guo D, Rorden C. 2015. Chronic Broca’s aphasia is caused by damage to Broca’s and Wernicke’s areas. Cereb Cortex. 25:4689–4696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fridriksson J, den Ouden DB, Hillis AE, Hickok G, Rorden C, Basilakos A, Yourganov G, Bonilha L. 2018. Anatomy of aphasia revisited. Brain, 141(3): 848–862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friederici AD, Bahlmann J, Heim S, Schubotz RI, Anwander A. 2006. The brain differentiates human and non-human grammars: functional localization and structural connectivity. Proc Natl Acad Sci. 103:2458–2463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friederici AD. 2011. The brain basis of language processing: from structure to function. Physiolog Rev. 91:1357–1392. [DOI] [PubMed] [Google Scholar]
- Friederici AD. 2012. The cortical language circuit: from auditory perception to sentence comprehension. Trends Cogn Sci. 16:262–268. [DOI] [PubMed] [Google Scholar]
- Friederici AD. 2017. Language in our brain: the origins of a uniquely human capacity. Cambridge (MA): MIT Press. [Google Scholar]
- Gage N, Hickok G. 2005. Multiregional cell assemblies, temporal binding and the representation of conceptual knowledge in cortex: a modern theory by a "classical" neurologist, Carl Wernicke. Cortex. 41:823–832. [DOI] [PubMed] [Google Scholar]
- Garrett M. 1980. Levels of processing in sentence production In Language production Vol. 1: Speech and talk (pp. 177–220). London: Academic Press. [Google Scholar]
- Gelfand JR, Bookheimer SY. 2003. Dissociating neural mechanisms of temporal sequencing and processing phonemes. Neuron. 38:831–842. [DOI] [PubMed] [Google Scholar]
- Glasel H, Leroy F, Dubois J, Hertz-Pannier L, Mangin JF, Dehaene-Lambertz G. 2011. A robust cerebral asymmetry in the infant brain: the rightward superior temporal sulcus. Neuroimage. 58:716–723. [DOI] [PubMed] [Google Scholar]
- Goldberg AE. 1995. Constructions: a construction grammar approach to argument structure. Chicago (IL): University of Chicago Press. [Google Scholar]
- Goodglass H, Berko J. 1960. Agrammatism and inflectional morphology in English. J Speech Hear Res. 3:257–267. [DOI] [PubMed] [Google Scholar]
- Goodglass H. 1968. Studies in the grammar of aphasics In: Rosenberg S, Koplin J, editors. Developments in applied psycholinguistic research. New York (NY): MacMillan. [Google Scholar]
- Goodglass H. 1992. Diagnosis of conduction aphasia In: Kohn SE, editor. Conduction aphasia. Hillsdale (NJ): Lawrence Erlbaum Associates, pp. 39–49. [Google Scholar]
- Goodglass H. 1993. Understanding aphasia. San Diego (CA): Academic Press. [Google Scholar]
- Goucha T, Friederici AD. 2015. The language skeleton after dissecting meaning: a functional segregation within Broca's area. Neuroimage. 114:294–302. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y. 2000. The neurology of syntax: language use without Broca's area. Behav Brain Sci. (1):1–21. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y, Amunts K e. 2006. Broca's region. New York (NY): Oxford University Press. [Google Scholar]
- Grodzinsky Y, Santi A. 2008. The battle for Broca’s region. Trends Cogn Sci. 12:474–480. [DOI] [PubMed] [Google Scholar]
- Grossman M, Rhee J, Moore P. 2005. Sentence processing in frontotemporal dementia. Cortex. 41:764–777. [DOI] [PubMed] [Google Scholar]
- Guenther FH. 2006. Cortical interactions underlying the production of speech sounds. J Commun Disord. 39:350–365. [DOI] [PubMed] [Google Scholar]
- Guenther FH, Hickok G. 2015. Role of the auditory system in speech production In: Celesia GG, Hickok G, editors. Handbook of clinical neurology. Vol 129 Elsevier, pp. 161–175. [DOI] [PubMed] [Google Scholar]
- Hagoort P. 2014. Nodes and networks in the neural architecture for language: Broca's region and beyond. Curr Opin Neurobiolog. 28:136–141. [DOI] [PubMed] [Google Scholar]
- Hauser MD, Chomsky N, Fitch WT. 2002. The faculty of language: what is it, who has it, and how did it evolve? Science. 298:1569–1579. [DOI] [PubMed] [Google Scholar]
- Henseler I, Regenbrecht F, Obrig H. 2014. Lesion correlates of patholinguistic profiles in chronic aphasia: comparisons of syndrome-, modality- and symptom-level assessment. Brain. 137:918–930. [DOI] [PubMed] [Google Scholar]
- Heim I, Kratzer A. 1998. Semantics in generative grammar. Oxford: Blackwell. [Google Scholar]
- Hickok G, Bellugi U, Klima ES. 1996. The neurobiology of sign language and its implications for the neural basis of language. Nature. 381:699. [DOI] [PubMed] [Google Scholar]
- Hickok G, Poeppel D. 2000. Towards a functional neuroanatomy of speech perception. Trends Cogn Sci. 4:131–138. [DOI] [PubMed] [Google Scholar]
- Hickok G, Buchsbaum B, Humphries C, Muftuler T. 2003. Auditory-motor interaction revealed by fMRI: speech, music, and working memory in area Spt. J Cogn Neurosci. 15:673–682. [DOI] [PubMed] [Google Scholar]
- Hickok G, Poeppel D. 2004. Dorsal and ventral streams: a framework for understanding aspects of the functional anatomy of language. Cognition. 92:67–99. [DOI] [PubMed] [Google Scholar]
- Hickok G, Poeppel D. 2007. The cortical organization of speech processing. Nat Rev Neurosci. 8:393–402. [DOI] [PubMed] [Google Scholar]
- Hickok G, Okada K, Serences JT. 2009. Area Spt in the human planum temporale supports sensory-motor integration for speech processing. Journal of neurophysiology. 101(5):2725–2732. [DOI] [PubMed] [Google Scholar]
- Hickok G, Houde J, Rong F. 2011. Sensorimotor integration in speech processing: computational basis and neural organization. Neuron. 69:407–422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickok G. 2012a. Computational neuroanatomy of speech production. Nat Rev Neurosci. 13:135–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickok G. 2012b. The cortical organization of speech processing: feedback control and predictive coding the context of a dual-stream model. J Commun Disord. 45:393–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickok G. 2014a. The architecture of speech production and the role of the phoneme in speech processing. Lang Cogn Process. 29:2–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickok G. 2014b. The myth of mirror neurons: the real neuroscience of communication and cognition. New York (NY): W.W. Norton & Company. [Google Scholar]
- Hill J, Dierker D, Neil J, Inder T, Knutsen A, Harwell J, Coalson T, Van Essen D. 2010. A surface-based analysis of hemispheric asymmetries and folding of cerebral cortex in term-born human infants. J Neurosci. 30:2268–2276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hodges JR, Patterson K, Oxbury S, Funnell E. 1992. Semantic dementia: progressive fluent aphasia with temporal lobe atrophy. Brain. 115:1783–1806. [DOI] [PubMed] [Google Scholar]
- Hodges JR, Patterson K. 2007. Semantic dementia: a unique clinicopathological syndrome. Lancet Neurol. 6:1004–1014. [DOI] [PubMed] [Google Scholar]
- Humphries C, Willard K, Buchsbaum B, Hickok G. 2001. Role of anterior temporal cortex in auditory sentence comprehension: an fMRI study. Neuroreport. 12:1749–1752. [DOI] [PubMed] [Google Scholar]
- Humphries C, Binder JR, Medler D, Liebenthal E. 2006. Syntactic and semantic modulation of neural activity during auditory sentence comprehension. J Cogn Neurosci. 18:665–679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphries C, Love T, Swinney D, Hickok G. 2005. Response of anterior temporal cortex to syntactic and prosodic manipulations during sentence processing. Hum Brain Mapp. 26:128–138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iacoboni M. 2009. Imitation, empathy, and mirror neurons. Ann Rev Psychol. 60:653–670. [DOI] [PubMed] [Google Scholar]
- Idsardi W, Raimy E. 2013. Three types of linearization and the temporal aspects of speech In: Biberauer MT, Roberts I, editors. Challenges to linearization. Berlin: Mouton de Gruyter, p. 31e56. [Google Scholar]
- Indefrey P. 2011. The spatial and temporal signatures of word production components: a critical update. Front Psychol. 2:255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Indefrey P, Levelt WJ. 2004. The spatial and temporal signatures of word production components. Cognition. 92:101–144. [DOI] [PubMed] [Google Scholar]
- Jackendoff R. 2002. Foundations of language: how language connects to the brain, the world, evolution, and thinking. New York (NY): Oxford University Press. [Google Scholar]
- Jackendoff R. 2017. In defense of theory. Cogn Sci. 41:185–212. [DOI] [PubMed] [Google Scholar]
- Jakobson R. 1956. Two aspects of language and two types of aphasic disturbances In: Jakobson R, Halle M, editors. Fundamentals of language. The Hague, Netherlands: Mouton. [Google Scholar]
- Jakuszeit M, Kotz SA, Hasting AS. 2013. Generating predictions: lesion evidence on the role of left inferior frontal cortex in rapid syntactic analysis. Cortex. 49:2861–2874. [DOI] [PubMed] [Google Scholar]
- January D, Trueswell JC, Thompson-Schill SL. 2009. Co-localization of Stroop and syntactic ambiguity resolution in Broca's area: implications for the neural basis of sentence processing. J Cogn Neurosci. 21:2434–2444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson MA, Goldberg AE. 2013. Evidence for automatic accessing of constructional meaning: jabberwocky sentences prime associated verbs. Lang Cogn Process. 28:1439–1452. [Google Scholar]
- Joshi AK, Schabes Y. 1997. Tree-adjoining grammars In: Handbook of formal languages. Heidelberg: Springer Berlin, pp. 69–123. [Google Scholar]
- Jurafsky D. 1996. A probabilistic model of lexical and syntactic access and disambiguation. Cogn Sci. 20:137–194. [Google Scholar]
- Kalénine S, Peyrin C, Pichat C, Segebarth C, Bonthoux F, Baciu M. 2009. The sensory-motor specificity of taxonomic and thematic conceptual relations: a behavioral and fMRI study. NeuroImage. 44:1152–1162. [DOI] [PubMed] [Google Scholar]
- Kempen G, Huijbers P. 1983. The lexicalization process in sentence production and naming: indirect election of words. Cognition. 14:185–209. [Google Scholar]
- Kho KH, Indefrey P, Hagoort P, van Veelen CWM, van Rijen PC, Ramsey NF. 2008. Unimpaired sentence comprehension after anterior temporal cortex resection. Neuropsychologia. 46:1170–1178. [DOI] [PubMed] [Google Scholar]
- Kleist K. 1914. Aphasie und geisteskrankheit. Münch Med Wochenschr. 6:8–12. [Google Scholar]
- Klima ES, Bellugi U. 1979. The signs of language. Cambridge (MA): Harvard University Press. [Google Scholar]
- Kussmaul A. 1877. Die Störungen der Sprache: Versuch einer Pathologie der Sprache. Leipzig: Vogel. [Google Scholar]
- Lau E, Stroud C, Plesch S, Phillips C. 2006. The role of structural prediction in rapid syntactic analysis. Brain Lang. 98:74–88. [DOI] [PubMed] [Google Scholar]
- Lau EF, Phillips C, Poeppel D. 2008. A cortical network for semantics: (de) constructing the N400. Nat Rev Neurosci. 9:920–933. [DOI] [PubMed] [Google Scholar]
- Leaver AM, Van Lare J, Zielinski B, Halpern AR, Rauschecker JP. 2009. Brain activation during anticipation of sound sequences. J Neurosci. 29:2477–2485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leonard MK, Ramirez NF, Torres C, Travis KE, Hatrak M, Mayberry RI, Halgren E. 2012. Signed words in the congenitally deaf evoke typical late lexicosemantic responses with no early visual responses in left superior temporal cortex. J Neurosci. 32:9700–9705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leroy F, Cai Q, Bogart SL, Dubois J, Coulon O, Monzalvo K, Fischer C, Glasel H, Van der Haegen L, Bénézit A et al. 2015. New human-specific brain landmark: the depth asymmetry of superior temporal sulcus. Proc Nat Acad Sci. 112:1208–1213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levelt WJ. 1993. Speaking: from intention to articulation. Cambridge (MA): MIT Press. [Google Scholar]
- Levelt WJ. 1989. Speaking: From intention to articulation. Cambridge (MA): MIT Press. [Google Scholar]
- Lewis RL. 2000. Specifying architectures for language processing: process, control, and memory in parsing and interpretation In: Crocker MW, et al., editors. Architectures and mechanisms for language processing. Cambridge (MA): Cambridge University Press. [Google Scholar]
- Lewis RL, Vasishth S. 2005. An activation-based model of sentence processing as skilled memory retrieval. Cogn Sci. 29:375–419. [DOI] [PubMed] [Google Scholar]
- Lewis RL, Vasishth S, Van Dyke JA. 2006. Computational principles of working memory in sentence comprehension. Trends Cogn Sci. 10:447–454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis GA, Poeppel D, Murphy GL. 2015. The neural bases of taxonomic and thematic conceptual relations: an MEG study. Neuropsychologia. 68:176–189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin EL, Murphy GL. 2001. Thematic relations in adults’ concepts. J Exp Psychol Gen. 130:3–28. [DOI] [PubMed] [Google Scholar]
- Linebarger MC, Schwartz MF, Saffran EM. 1983. Sensitivity to grammatical structure in so-called agrammatic aphasics. Cognition. 13:361–392. [DOI] [PubMed] [Google Scholar]
- Love T, Swinney D, Walenski M, Zurif E. 2008. How left inferior frontal cortex participates in syntactic processing: evidence from aphasia. Brain Lang. 107:203–219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacDonald MC, Pearlmutter NJ, Seidenberg MS. 1994. The lexical nature of syntactic ambiguity resolution. Psych Rev. 101:676. [DOI] [PubMed] [Google Scholar]
- MacSweeney M, Woll B, Campbell R, McGuire PK, David AS, Williams SC, Suckling J, Calvert GA, Brammer MJ. 2002. Neural systems underlying British sign language and audio-visual English processing in native users. Brain. 125:1583–1593. [DOI] [PubMed] [Google Scholar]
- MacSweeney M, Campbell R, Woll B, Brammer MJ, Giampietro V, David AS, Calvert GA, McGuire PK. 2006. Lexical and sentential processing in British sign language. Hum Brain Mapp. 27:63–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magnusdottir S, Fillmore P, Den Ouden DB, Hjaltason H, Rorden C, Kjartansson O, Bonilha L, Fridriksson J. 2013. Damage to left anterior temporal cortex predicts impairment of complex syntactic processing: a lesion-symptom mapping study. Hum Brain Mapp. 34:2715–2723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maguire EA, Frith CD, Morris RGM. 1999. The functional neuroanatomy of comprehension and memory: the importance of prior knowledge. Brain. 122:1839–1850. [DOI] [PubMed] [Google Scholar]
- Malyutina S, den Ouden DB. 2017. Task-dependent neural and behavioral effects of verb argument structure features. Brain Lang. 168:57–72. [DOI] [PubMed] [Google Scholar]
- Martin A, Chao LL. 2001. Semantic memory and the brain: structure and processes. Curr Opin Neurobiol. 11:194–201. [DOI] [PubMed] [Google Scholar]
- Matchin W. 2017. A neuronal retuning hypothesis of sentence-specificity in Broca’s area. Psychonom Bull Rev. 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matchin W, Hammerly C, Lau E. 2017b. The role of the IFG and pSTS in syntactic prediction: evidence from a parametric study of hierarchical structure in fMRI. Cortex. 88:106–123. [DOI] [PubMed] [Google Scholar]
- Matchin W, Villwock A, Roth A, Ilkbasaran D, Hatrak H, Davenport T, Halgren E, Mayberry R. 2017a. The cortical organization of syntactic processing in American Sign Language: evidence from a parametric manipulation of constituent structure in fMRI and MEG Poster presented at the 2017 meeting of the Society for the Neurobiology of Language in Baltimore.
- Matchin W, Rogalsky C. in press. Aphasia & syntax In: Sprouse J, editor. The Oxford Handbook of Experimental Syntax. Oxford (UK): Oxford University Press.
- Mayberry RI, Lock E, Kazmi H. 2002. Linguistic ability and early language exposure. Nature. 417:38–38. [DOI] [PubMed] [Google Scholar]
- Mayberry RI, Chen JK, Witcher P, Klein D. 2011. Age of acquisition effects on the functional organization of language in the adult brain. Brain Lang. 119:16–29. [DOI] [PubMed] [Google Scholar]
- Mazoyer BM, Tzourio N, Frak V, Syrota A, Murayama N, Levrier O, Salamon G, Dehaene S, Cohen L, Mehler J. 1993. The cortical representation of speech. J Cogn Neurosci. 5:467–479. [DOI] [PubMed] [Google Scholar]
- McElree B. 1993. The locus of lexical preference effects in sentence comprehension: a time-course analysis. J Mem Lang. 32:536–571. [Google Scholar]
- McElree B, Foraker S, Dyer L. 2003. Memory structures that subserve sentence comprehension. Journal of memory and language. 48(1):67–91. [Google Scholar]
- Meltzer-Asscher A, Mack JE, Barbieri E, Thompson CK. 2015. How the brain processes different dimensions of argument structure complexity: evidence from fMRI. Brain Lang. 142:65–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mesulam MM. 1990. Large-scale neurocognitive networks and distributed processing for attention, language, and memory. Annals Neurol. 28:597–613. [DOI] [PubMed] [Google Scholar]
- Mesulam MM, Thompson CK, Weintraub S, Rogalski EJ. 2015. The Wernicke conundrum and the anatomy of language comprehension in primary progressive aphasia. Brain. 138:2423–2437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer L, Friederici AD. 2016. Neural systems underlying the processing of complex sentences In: Hickok G, Small SA, editors. The neurobiology of language. In: Elsevier, London: Academic Press, pp. 597–606. [Google Scholar]
- Mohr JP, Pessin MS, Finkelstein S, Funkenstein HH, Duncan GW, Davis KR. 1978. Broca aphasia pathologic and clinical. Neurol. 28:311–311. [DOI] [PubMed] [Google Scholar]
- Moses DA, Mesgarani N, Leonard MK, Chang EF. 2016. Neural speech recognition: continuous phoneme decoding using spatiotemporal representations of human cortical activity. J Neural Eng. 13:056004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy E. 2015. The brain dynamics of linguistic computation. Front Psychol. 6:1515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson MJ, El Karoui I, Giber K, Yang X, Cohen L, Koopman H, Cash SS, Naccache L, Hale JT, Pallier C et al. 2017. Neurophysiological dynamics of phrase-structure building during sentence processing. Proc Natl Acad Sci. 114:E3669–E3678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neville HJ, Bavelier D, Corina D, Rauschecker J, Karni A, Lalwani A, Braun A, Clark V, Jezzard P, Turner R. 1998. Cerebral organization for language in deaf and hearing subjects: biological constraints and effects of experience. Proc Nat Acad Sci. 95:922–929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newman AJ, Supalla T, Fernandez N, Newport EL, Bavelier D. 2015. Neural systems supporting linguistic structure, linguistic experience, and symbolic communication in sign language and gesture. Proc Nat Acad Sci. 112:11684–11689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Novick JM, Trueswell JC, Thompson-Schill SL. 2005. Cognitive control and parsing: Reexamining the role of Broca’s area in sentence comprehension. Cogn Affect Behav Neurosci. 5:263–281. [DOI] [PubMed] [Google Scholar]
- Novick JM, Trueswell JC, Thompson-Schill SL. 2010. Broca’s area and language processing: evidence for the cognitive control connection. Lang Linguis Compass. 4:906–924. [Google Scholar]
- Okada K, Hickok G. 2006. Identification of lexical-phonological networks in the superior temporal sulcus using fMRI. Neuroreport. 17:1293–1296. [DOI] [PubMed] [Google Scholar]
- Okada K, Rong F, Venezia J, Matchin W, Hsieh IH, Saberi K, Hickok G. 2010. Hierarchical organization of human auditory cortex: evidence from acoustic invariance in the response to intelligible speech. Cereb Cortex. 20:2486–2495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pallier C, Devauchelle A, Dehaene S. 2011. Cortical representation of the constituent structure of sentences. Proc Natl Acad Sci. 108:2522–2527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papathanasiou I, Coppens P. 2017. Aphasia and related neurogenic communication disorders. 2nd ed. Burlington (MA): Jones & Bartlett Learning. [Google Scholar]
- Pasley BN, David SV, Mesgarani N, Flinker A, Shamma SA, Crone NE, Chang EF. 2012. Reconstructing speech from human auditory cortex. PLoS Biol. 10:e1001251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patterson K, Nestor PJ, Rogers TT. 2007. Where do you know what you know? The representation of semantic knowledge in the human brain. Nat Rev Neurosci. 8:976–987. [DOI] [PubMed] [Google Scholar]
- Peelle JE, Troiani V, Gee J, Moore P, McMillan C, Vesely L, Grossman M. 2008. Sentence comprehension and voxel-based morphometry in progressive nonfluent aphasia, semantic dementia, and nonaphasic frontotemporal dementia. Journal of Neurolinguistics. 21(5):418–432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petersson KM, Folia V, Hagoort P. 2012. What artificial grammar learning reveals about the neurobiology of syntax. Brain Lang. 120:83–95. [DOI] [PubMed] [Google Scholar]
- Petitto LA. 1994. Are signed languages ‘real’ languages . Evidence from American Sign Language and Langue des Signes Québécoise Signpost. International Quarterly of the Sign Linguistics Association, 7(3):1–10. [Google Scholar]
- Petitto LA, Zatorre RJ, Gauna K, Nikelski EJ, Dostie D, Evans AC. 2000. Speech-like cerebral activity in profoundly deaf people processing signed languages: implications for the neural basis of human language. Proc Nat Acad Sci. 97:13961–13966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pillay SB, Binder JR, Humphries C, Gross WL, Book DS. 2017. Lesion localization of speech comprehension deficits in chronic aphasia. Neurol. 88:970–975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinker S. 1999. Words and rules: the ingredients of language. New York (NY): Basic Books. [Google Scholar]
- Piras F, Marangolo P. 2007. Noun-verb naming in aphasia: a voxel-based lesion-symptom mapping study. Neuroreport. 18:1455–1458. [DOI] [PubMed] [Google Scholar]
- Poeppel D, Embick D. 2005Defining the relation between linguistics and neuroscience In: Twenty-first century psycholinguistics: four cornerstones, ed. by Cutler Anne, Mahwah (NJ): Lawrence Erlbaum Associates Publishers, 103–118.
- Poletiek FH, Fitz H, Bocanegra BR. 2016. What baboons can (not) tell us about natural language grammars. Cognition. 151:108–112. [DOI] [PubMed] [Google Scholar]
- Pollard C, Sag IA. 1994. Head-driven phrase structure grammar. Chicago (IL): University of Chicago Press. [Google Scholar]
- Pylkkänen L. 2015. Composition of complex meaning: interdisciplinary perspectives on the left anterior temporal lobe In: Hickok G, Small S, editors. Neurobiology of language. Amsterdam: Academic Press, pp. 621–631. [Google Scholar]
- Race DS, Ochfeld E, Leigh R, Hillis AE. 2012. Lesion analysis of cortical regions associated with the comprehension of nonreversible and reversible yes/no questions. Neuropsychologia. 50:1946–1953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rauschecker JP, Scott SK. 2009. Maps and streams in the auditory cortex: nonhuman primates illuminate human speech processing. Nat Neurosci. 12:718–724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rilling JK, Glasser MF, Preuss TM, Ma X, Zhao T, Hu X, Behrens TE. 2008. The evolution of the arcuate fasciculus revealed with comparative DTI. Nat Neurosci. 11:426. [DOI] [PubMed] [Google Scholar]
- Rimmele JM, Morillon B, Poeppel D, Arnal LH. 2018. Proactive sensing of periodic and aperiodic auditory patterns. Trends Cogn Sci. 22:870–882. [DOI] [PubMed] [Google Scholar]
- Roelofs A, Ferreira VS. 2019. The architecture of speaking In Hagoort P. (Ed.). Human language: from genes and brains to behavior. Cambridge (MA): MIT Press.
- Rogalsky C, Hickok G. 2008. Selective attention to semantic and syntactic features modulates sentence processing networks in anterior temporal cortex. Cereb Cortex. 19:786–796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogalsky C, Matchin W, Hickok G. 2008a. Broca's area, sentence comprehension, and working memory: an fMRI study. Front Hum Neurosci. 6:2–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogalsky C, Pitz E, Hillis AE, Hickok G. 2008b. Auditory word comprehension impairment in acute stroke: relative contribution of phonemic versus semantic factors. Brain Lang. 107:167–169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogalsky C, Hickok G. 2011. The role of Broca's area in sentence comprehension. J Cogn Neurosci. 23:1664–1680. [DOI] [PubMed] [Google Scholar]
- Rogalsky C, Rong F, Saberi K, Hickok G. 2011. Functional anatomy of language and music perception: temporal and structural factors investigated using functional magnetic resonance imaging. J Neurosci. 31:3843–3852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogalsky C, Almeida D, Sprouse J, Hickok G. 2015. Sentence processing selectivity in Broca's area: evident for structure but not syntactic movement. Lang Cogn Neurosci. 30:1326–1338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogalsky C, LaCroix AN, Chen KH, Anderson SW, Damasio H, Love T, Hickok G. 2018. The neurobiology of Agrammatic sentence comprehension: a lesion study. J Cogn Neurosci. 30:234–255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rong F, Isenberg AL, Sun E, Hickok G. 2018. The neuroanatomy of speech sequencing at the syllable level. PloS One. 13(10):e0196381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roux FE, Miskin K, Durand JB, Sacko O, Rehault E, Tanova R, Demonet JF. 2015. Electrostimulation mapping of comprehension of auditory and visual words. Cortex. 71:398–408. [DOI] [PubMed] [Google Scholar]
- Sandler W, Lillo-Martin D. 2006. Sign language and linguistic universals. Cambridge (MA): Cambridge University Press. [Google Scholar]
- Sapolsky D, Bakkour A, Negreira A, Nalipinski P, Weintraub S, Mesulam MM, Caplan D, Dickerson BC. 2010. Cortical neuroanatomic correlates of symptom severity in primary progressive aphasia. Neurology. 75(4):358–366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz MF, Kimberg DY, Walker GM, Brecher A, Faseyitan OK, Dell GS, Mirman D, Coslett HB. 2011. Neuroanatomical dissociation for taxonomic and thematic knowledge in the human brain. Proceedings of the National Academy of Sciences. 108(20):8520–8524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seed A, Byrne R. 2010. Animal tool-use. Curr Biol. 20:R1032–R1039. [DOI] [PubMed] [Google Scholar]
- Segawa JA, Tourville JA, Beal DS, Guenther FH. 2015. The neural correlates of speech motor sequence learning. J Cogn Neurosci. 27:819–831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selnes OA, Niccum N, Knopman DS, Rubens AB. 1984. Recovery of single word comprehension: CT-scan correlates. Brain Lang. 21:72–84. [DOI] [PubMed] [Google Scholar]
- Shadmehr R, Krakauer JW. 2008. A computational neuroanatomy for motor control. Exper Brain Res. 185:359–381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simonyan K. 2014. The laryngeal motor cortex: its organization and connectivity. Curr Opinion Neurobiol. 28:15–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stokoe WC. 1960. Sign language structure: an outline of the visual communication systems of the American deaf In: Studies in linguistics, occasional papers Vol 8 Buffalo (NY): University of Buffalo. [DOI] [PubMed] [Google Scholar]
- Stowe LA, Broere CA, Paans AM, Wijers AA, Mulder G, Vaalburg W et al. 1998. Localizing components of a complex task: sentence processing and working memory. Neuroreport. 9:2995–2999. [DOI] [PubMed] [Google Scholar]
- Tattersall I. 2004. What happened in the origin of human consciousness? Anat Rec B: New Anatomist (ARB). 276:19–26. [DOI] [PubMed] [Google Scholar]
- Terrace HS, Petitto LA, Sanders RJ, Bever TG. 1979. Can an ape create a sentence? Science. 206:891–902. [DOI] [PubMed] [Google Scholar]
- Thompson CK, Bonakdarpour B, Fix SC, Blumenfeld HK, Parrish TB, Gitelman DR, Mesulam M. 2007. Neural correlates of verb argument structure processing. J Cogn Neurosci. 19:1753–1767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson CK, Bonakdarpour B, Fix SF. 2010. Neural mechanisms of verb argument structure processing in agrammatic aphasic and healthy age-matched listeners. J Cogn Neurosci. 22:1993–2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thothathiri M, Kimberg DY, Schwartz MF. 2012. The neural basis of reversible sentence comprehension: evidence from voxel-based lesion symptom mapping in aphasia. J Cogn Neurosci. 24:212–222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tourville JA, Guenther FH. 2011. The DIVA model: a neural theory of speech acquisition and production. Language and cognitive processes. 26(7):952–981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tyler LK, Bright P, Fletcher P, Stamatakis EA. 2004. Neural processing of nouns and verbs: the role of inflectional morphology. Neuropsychologia. 42:512–523. [DOI] [PubMed] [Google Scholar]
- Tyler LK, Stamatakis EA, Post B, Randall B, Marslen-Wilson W. 2005. Temporal and frontal systems in speech comprehension: an fMRI study of past tense processing. Neuropsychologia. 43:1963–1974. [DOI] [PubMed] [Google Scholar]
- Tyler LK, Marslen-Wilson W. 2007. Fronto-temporal brain systems supporting spoken language comprehension. Philos Trans Royal Soc B: Biolog Sci. 363:1037–1054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandenberghe R, Nobre AC, Price CJ. 2002. The response of left temporal cortex to sentences. J Cogn Neurosci. 14:550–560. [DOI] [PubMed] [Google Scholar]
- von Humboldt W. 1836. On language (Reproduced) New York (NY): Cambridge University Press, p. 1988. [Google Scholar]
- Vosse T, Kempen G. 2000. Syntactic structure assembly in human parsing: a computational model based on competitive inhibition and a lexicalist grammar. Cognition. 75:105–143. [DOI] [PubMed] [Google Scholar]
- Warneken F, Hare B, Melis AP, Hanus D, Tomasello M. 2007. Spontaneous altruism by chimpanzees and young children. PLoS Biol. 5:e184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wernicke C. 1874/1969. The symptom complex of aphasia: a psychological study on an anatomical basis In: Cohen RS, Wartofsky MW, editors. Boston studies in the philosophy of science. Dordrecht: D. Reidel Publishing Company, pp. 34–97. [Google Scholar]
- Wernicke C. 1900. Grundriss der Psychiatrie. Leipzig: Verlag von Georg Thieme. [Google Scholar]
- Westerlund M, Pylkkänen L. 2014. The role of the left anterior temporal lobe in semantic composition vs. semantic memory. Neuropsychologia. 57:59–70. [DOI] [PubMed] [Google Scholar]
- Whiten A, Goodall J, McGrew WC, Nishida T, Reynolds V, Sugiyama Y, Tutin CE, Wrangham RW, Boesch C. 1999. Cultures in chimpanzees. Nature. 99:682. [DOI] [PubMed] [Google Scholar]
- Williams A, Reddigari S, Pylkkänen L. 2017. Early sensitivity of left perisylvian cortex to relationality in nouns and verbs. Neuropsychologia. 100:131–143. [DOI] [PubMed] [Google Scholar]
- Wilson SM, Saygın AP. 2004. Grammaticality judgment in aphasia: deficits are not specific to syntactic structures, aphasic syndromes, or lesion sites. J Cogn Neurosci. 16:238–252. [DOI] [PubMed] [Google Scholar]
- Wilson SM, Dronkers NF, Ogar JM, Jang J, Growdon ME, Agosta F, Henry ML, Miller BL, Gorno-Tempini ML. 2010a. Neural correlates of syntactic processing in the nonfluent variant of primary progressive aphasia. J Neurosci. 30:16845–16854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson SM, Henry ML, Besbris M, Ogar JM, Dronkers NF, Jarrold W, Miller BL, Gorno-Tempini ML. 2010b. Connected speech production in three variants of primary progressive aphasia. Brain. 133:2069–2088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson SM, Galantucci S, Tartaglia MC, Rising K, Patterson DK, Henry ML, Gorno-Tempini ML. 2011. Syntactic processing depends on dorsal language tracts. Neuron. 72:397–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson SM, DeMarco AT, Henry ML, Gesierich B, Babiak M, Mandelli ML, Miller BL, Gorno-Tempini ML. 2014. What role does the anterior temporal lobe play in sentence-level processing? Neural correlates of syntactic processing in semantic variant primary progressive aphasia. J Cogn Neurosci. 26:970–985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson SM, DeMarco AT, Henry ML, Gesierich B, Babiak M, Miller BL, Gorno-Tempini ML. 2016. Variable disruption of a syntactic processing network in primary progressive aphasia. Brain. 139:2994–3006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson SM, Bautista A, McCarron A. 2018a. Convergence of spoken and written language processing in the superior temporal sulcus. NeuroImage. 171:62–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson SM, Eriksson DK, Schneck SM, Lucanie JM. 2018b. A quick aphasia battery for efficient, reliable, and multidimensional assessment of language function. PloS One. 13:e0192773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson SM, Yen M, Eriksson DK. 2018c. An adaptive semantic matching paradigm for reliable and valid language mapping in individuals with aphasia. Hum Brain Mapp. 39:3285–3307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright P, Stamatakis EA, Tyler LK. 2012. Differentiating hemispheric contributions to syntax and semantics in patients with left-hemisphere lesions. J Neurosci. 32:8149–8157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wulfeck B, Bates E. 1991. Differential sensitivity to errors of agreement and word order in Broca's aphasia. J Cogn Neurosci. 3:258–272. [DOI] [PubMed] [Google Scholar]
- Yagata SA, Yen M, McCarron A, Bautista A, Lamair-Orosco G, Wilson SM. 2017. Rapid recovery from aphasia after infarction of Wernicke’s area. Aphasiol. 31:951–980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yagmurlu K, Middlebrooks EH, Tanriover N, Rhoton AL. 2016. Fiber tracts of the dorsal language stream in the human brain. Journal of neurosurgery. 124(5):1396–1405. [DOI] [PubMed] [Google Scholar]
- Yang C. 2013. Ontogeny and phylogeny of language. Proc Nat Acad Sci. 110(16):6324–6327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaccarella E, Friederici AD. 2015. Merge in the human brain: a sub-region based functional investigation in the left pars opercularis. Front Psychol. 6:1818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaccarella E, Meyer L, Makuuchi M, Friederici AD. 2017. Building by syntax: the neural basis of minimal linguistic structures. Cereb Cortex. 27:411–421. [DOI] [PubMed] [Google Scholar]