

Abstract
Objective
Low back pain is frequently treated with nonsteroidal anti-inflammatory drugs (NSAIDs), but little is known about intervertebral disc metabolism of the prostaglandins that are diminished by these drugs. Hence, this study aimed at delineating prostaglandin actions in cytokine activated disc cells by comparing the response of nucleus pulposus (NP) cells to the pro-inflammatory cytokine interleukin (IL)-1β with and without cyclooxygenase 2 (COX-2) inhibition.
Methods
NP cells cultured in alginate beads were activated with IL-1β ± the COX-2 inhibitor Sc-58125. Media harvested from cultured cells were analyzed for prostaglandin E2 (PGE2), prostaglandin F2 alpha (PGF2α), IL-6, and matrix metalloproteinase (MMP)-3 by enzyme-linked immunosorbent assay and nitric oxide by Griess Reaction. Gene expression along with proteoglycan, collagen, and total protein synthesis were also measured.
Results
IL-1β increased culture media PGE2 and PGF2α, but decreased proteoglycan and collagen syntheses as well as mRNA expression of the matrix genes aggrecan, versican, collagen I, and collagen II. COX-2 inhibition partially rescued proteoglycan and collagen syntheses and collagen I mRNA, but decreased collagen II mRNA IL-1β activated NP cells. COX-2 inhibition initially enhanced and subsequently reduced IL-1β induced inducible nitric oxide synthase, without altering medium nitrite. IL-1β induction of MMP-3 mRNA was increased by COX-2 inhibition at 24 and 48 hours.
Conclusion
COX-2 inhibition alters the response of NP cells to IL-1β, suggesting IL-1β action on disc cells is mediated at least in part through COX-2 and its prostaglandins. COX-2 inhibition produces minimal effects on several key catabolic mediators, with the exception of MMP-3. Blocking COX-2 might be beneficial for maintaining disc matrix since it provides an overall rescue of IL-1 induced loss of matrix protein synthesis.
Keywords: Prostaglandins, COX-2 inhibition, Intervertebral disc cells, IL-1
INTRODUCTION
Low back pain is often associated with intervertebral disc degeneration (IDD) [1]. IDD therapies continue to be palliative and treat the symptoms rather than intervening to slow or reverse the course of IDD. Nonsteroidal anti-inflammatory drugs (NSAIDs) are often prescribed for their analgesic and anti-inflammatory effects. However little work has addressed the actual role in disc metabolism of the prostaglandins whose synthesis and accumulation are diminished by these drugs.
We reported results from microarray analysis, which showed that inflammatory stress increases expression of synthases and receptors for prostaglandin E2 (PGE2) and prostaglandin F2α (PGF2α), resulting in elevated PGE2 and PGF2α production in conditioned media (CM) of disc cells and underscoring the importance of these inflammatory mediators [2,3]. We therefore evaluated the effects of PGE2 and PGF2α on human nucleus pulposus (hNP) cells in culture [2]. PGE2 diminished disc cell proteoglycan synthesis, in a dose-dependent manner. Semiquantitative reverse transcription-polymerase chain reaction (RT-PCR) revealed differential effects of PGE2 and PGF2α on disc cell expression of key matrix structural genes, aggrecan, versican, and collagens type I and II. PGE2 and PGF2α also decreased message for the anabolic factor insulin-like growth factor-1. PGE2 decreased mRNA expression for the anticatabolic factor tissue inhibitor of metalloproteinases 1 (TIMP-1) while PGF2α increased mRNAs for catabolic factors matrix metalloproteinase (MMP)-1 and MMP-3. The data suggest that PGE2 and PGF2α may have an overall negative impact on disc matrix homeostasis, and the use of NSAIDs thus could positively impact disc metabolism while treating back pain.
However, the action of prostaglandins on disc cells under inflammatory conditions, as is often the case when prostaglandins are secreted, may be different from those observed in the studies cited above due to complex interactions of the intracellular signaling pathways in cytokine activated cells. In addition, prostaglandins other than PGE2 and PGF-2α are produced under inflammatory stress, and these prostaglandins could also have specific and differential effects on disc matrix homeostasis.
The potential role of inflammatory mediators in IDD is supported by the increase in cells expressing IL-1 and tumor necrosis factor (TNF)-α in degenerated human disc specimens [4,5]. These cytokines have multiple actions that contribute to matrix loss and disc degeneration. That these cytokines induce COX-2 in disc cells, with subsequent synthesis of prostaglandins, is well documented [6-9]. Given the frequency with which patients with low back pain are treated with NSAIDs, which block COX-2 activity, it is appropriate to test if inhibiting COX-2 activity and thus prostaglandin synthesis, will change the cascade of responses usually elicited by cytokines. Through comparing cell responses in the absence and presence of COX-2 inhibition, we examined the role of prostaglandins in cellular responses in IL-1 activated disc cells. We hypothesized that these effects will differ compared to noninflammatory conditions. This has great clinical relevance since the desired effect in using NSAIDs is an anti-inflammatory response, and not a detrimental effect on the overall disc matrix homeostasis, which could further perpetuate the degenerative cascade. The findings in this study enhanced our understanding of NP cell metabolism which could lead to future therapeutics aimed at targeting the response to inflammatory stimuli.
MATERIALS AND METHODS
1. Nucleus Pulposus Cell Isolation and Culture
Disc tissues were harvested from 8 female patients (age 18 to 66 years; mean, 41 years), and 8 males (age 31 to 67 years; mean, 45 years) whose diagnosis was disc degeneration. The discs were classified as grades 1–4 disc degeneration according to the Thompson scale [10], and the average was 2.14 (females) and 2.38 (males); only one grade 4 preparation was used for these cell preparations. The samples came from patients undergoing lumbar discectomy, scoliosis surgery in which discs were removed with anterior approaches, and lumbar interbody fusions for spinal stenosis and/or spondylolisthesis whereby disc tissue was readily acquired as part of the surgical procedure. We excluded patients with tumors, infections or any spine surgeries where disc removal was not part of the planned surgical procedure. No granulation tissue was present, and the nucleus could be clearly defined from the anulus. The nucleus pulposus was dissected, washed with Hank’s balanced salt solution (HBSS), and transported to the laboratory within 30 minutes. The experimental protocol was approved by the human subjects Institutional Review Board at the University of Pittsburgh (No. 0312055), which included informed consent from the patients.
Specimens from individual patients were rinsed with HBSS, minced into small fragments, and cells (hNP) were isolated, expanded in monolayer, and passed to 3-dimensional (3D) culture in alginate beads as previously described [11]. Cells were cultured in F-12/Dulbecco’s Modified Essential Medium containing 10% fetal bovine serum, 1% penicillin-streptomycin, and 25-μg/mL L-ascorbic acid under standard conditions (37°C, 5% CO2, 95% air, bicarbonate buffering to maintain pH 7.2). Cell culture materials were purchased from Invitrogen/Gibco unless otherwise noted, and the hNP cells were used at passage 1 or 2. Experiments began after the cells had been in 3D culture for at least one week, and cells from the same patient were used for matrix synthesis, conditioned medium analysis, and RNA extraction when possible based on cell yield.
2. Response to Cytokines
The response of hNP cells, in low (1%) serum media, to activation by 1-ng/mL recombinant human IL-1β (R&D Systems, Minneapolis, MN, USA) and 5-ng/mL TNF-α (Sigma-Aldrich, St Louis, MO) was followed for 3 days. COX-2 activity was blocked with 0.5 μM of the inhibitor SC-58125 (Sc) (Calbiochem, San Diego, CA, USA) dissolved in dimethyl sulfoxide (DMSO). Sc (0.125%) or DMSO vehicle was added to the cells 30 minutes before addition of IL-1 or TNF-α. Experiments for analysis of gene expression were terminated on days 1, 2, and 3 and total RNA was extracted using the RNeasy Mini Kit (Qiagen, Venlo, The Netherlands), according to the manufacturer’s instructions. CM was harvested at the same time and stored at -40°C until analysis. Proteoglycan synthesis was measured on day 3 and total protein and collagen syntheses were done after 24–48 hours of activation as described below.
Experiments were terminated by harvesting the beads and dissolving the alginate. Cells were harvested, digested in papain buffer at 60°C overnight, and the digest assayed for DNA using Picogreen (Molecular Probes, Eugene, OR, USA).
3. Matrix Protein Synthesis
Proteoglycan synthesis was measured from a six-hour pulse labeling with 35S-sulfate at 20 μCi/mL as previously described [11] and the results calculated as pmol 35S incorporated/μg DNA and expressed relative to Control. Collagenase sensitive and total protein synthesis was measured as 24-hour 3 -H-Proline incorporation as described [12], calculated as dpm/ug DNA, and expressed relative to control.
4. Assay of Conditioned Media
hNP cell production of nitric oxide was assessed as CM nitrite using the Griess Reaction. CM PGE2, MMP-3, TIMP-1, and IL-6 were assayed using enzyme-linked immunosorbent assay kits from R&D Systems (D6050 for IL-6, DMP300 for MMP-3, DTM100 for TIMP-1, and KGE004B for PGE2). CM PGF2α was assayed using a kit from Assay Designs (ADI-900-091). The CM data were normalized to DNA content of the cells in the beads.
5. Gene Expression
Gene expression was analyzed by real-time RT-PCR using an iCycler IQ4 detection system (Bio-Rad, Hercules, CA, USA). The reactions were done with primers designed (Beacon Express) and tested to assure specificity with melt curve analysis performed with each assay [2,13]. Real-time RT-PCR reactions were done in duplicate in 96-well plates in a volume of 25 μL using the reagents and an optimized protocol per the Bio-Rad iScript One-Step RT-PCR Kit. The cycle threshold (Ct) values were obtained, and data normalized to GAPDH (glyceraldehyde 3-phosphate dehydrogenase) expression using the ΔΔCt method to calculate relative mRNA levels of each gene.
6. Statistical Analysis
Data are given as mean±standard error of the n noted in the figure legends. A p-value of Student t-test was calculated, and a p-value of less than 0.05 defined statistically significant differences.
RESULTS
1. Prostaglandin Synthesis
The efficacy of 0.5-μM Sc-58125 to block cytokine-induced prostaglandin synthesis was evaluated from CM assays for PGE2 and PGF2α after 72-hour activation (Fig. 1). Note the substantial production and release of PGF2α as well as PGE2 by cells exposed to IL-1β. TNF-α increased prostaglandins to a lesser extent, but preliminary studies showed that COX-2 inhibition neither influenced matrix protein synthesis nor CM content of MMP-3, IL-6, nitric oxide (NO), or TIMP-1 in these TNF-α activated cells. Thus, subsequent experiments examined NP cell response to IL-1 when prostaglandin accumulation was altered over the higher concentration range seen in response to this agonist.
Fig. 1.
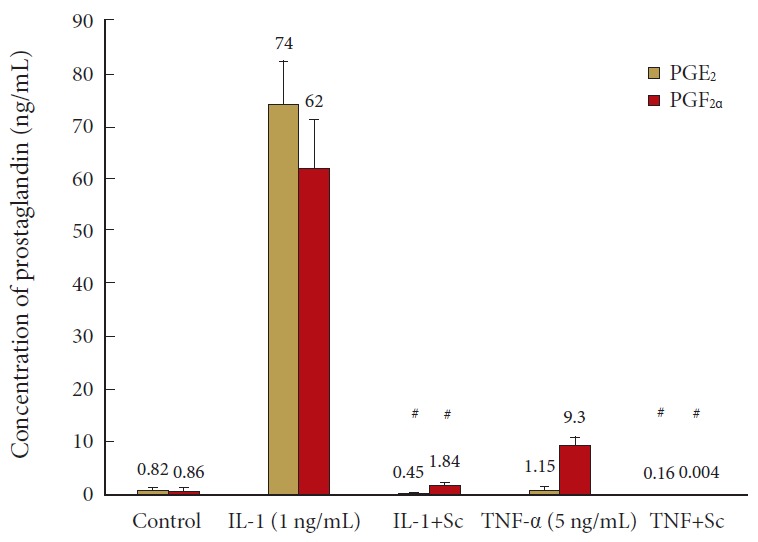
Cyclooxygenase 2 inhibition blunts production of prostaglandin E2 (PGE2) and prostaglandin F2 alpha (PGF2α) in culture media of human nucleus pulposus (hNP) cells activated by interleukin (IL)-1 or tumor necrosis factor (TNF)-α. hNP cells were maintained in alginate beads, activated by IL-1±Sc-58125 (Sc) or TNF-α±Sc and conditioned media harvested after 72 hours for prostaglandin measurement. Values are mean±standard error of n=6. #p<0.05 vs. IL-1 alone.
2. Proteoglycan Synthesis and Gene Expression
IL-1 decreased proteoglycan synthesis to less than 40% of control and blockade of prostaglandin synthesis in these cells counteracted IL-1 action with a modest increase to 48%±4% of control at 72 hours (p = 0.05). In contrast, the decrease in aggrecan mRNA (to 20% of control at 72 hours) was further reduced to ~10% of control by inhibition of prostaglandin synthesis (p = 0.003). Similar results were seen for versican mRNA at day 3 (p = 0.024) (Fig. 2).
Fig. 2.
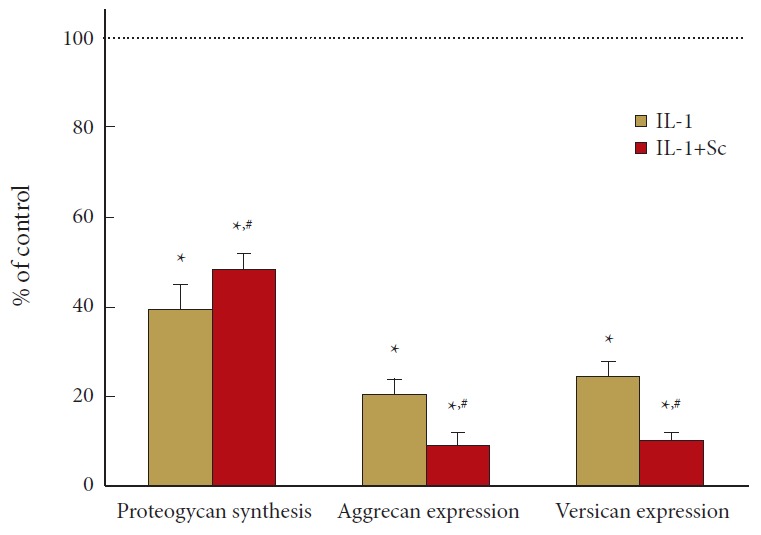
Cyclooxygenase 2 inhibition reverses interleukin (IL)-1 depression of proteoglycan synthesis but decreases aggrecan and versican mRNA in human nucleus pulposus (hNP) cells. hNP cells were exposed to 1-ng/mL IL-1±0.5-μM Sc-58125 (Sc) for 72 hours and proteoglycan synthesis determined as 35-S sulfate incorporation (n=7) during the final 6 hours of culture. Aggrecan and versican mRNA were assayed at 72 hours (n=8). Values are mean±standard error of n=7–8 expressed as % of control, untreated cells. *p<0.05 vs. control. #p<0.05 vs. IL-1 alone.
3. Total and Collagenase Sensitive Protein Synthesis
IL-1 does not change total protein synthesis, however, if prostaglandin production was inhibited in these cells, synthesis actually increased by ~60% (p = 0.003). Collagenase sensitive proline incorporation was similar, with the increase in synthesis effected by COX-2 inhibition 35% above control (p = 0.012). IL-1 decreased collagen I message, however, if prostaglandin synthesis was blocked, this decrease was restored from ~5% of control to ~30% of control (p = 0.0005). mRNA for collagen II was also reduced to ~5% of control in IL-1 activated cells; when prostaglandin synthesis was inhibited, this value further decreased to ~3% (p = 0.017; data is plotted X10 for visualization of these differences in Fig. 3). These results are summarized in Fig. 4.
Fig. 3.
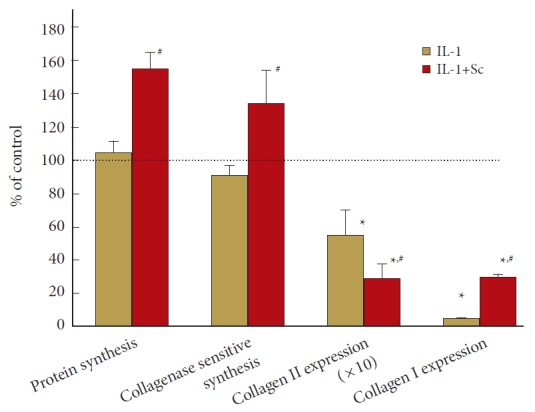
Cyclooxygenase 2 inhibition increases 3-H-proline incorporation into collagenase sensitive and total protein while decreasing collagen II and increasing collagen I mRNA in interleukin (IL)-1 activated human nucleus pulposus (hNP) Cells. hNP in alginate beads were exposed to IL-1±0.5-μM Sc-58125 (Sc). Synthesis was assessed during 24- to 48-hour exposure; mRNA was isolated after 72 hours. n=4 for collagen and total protein synthesis, and n=8 for collagens I and II mRNA expression experiment. Values are mean±standard error of n=4–8. *p<0.05 vs. control. #p<0.05 vs. IL-1 alone. Collagen II data are plotted ×10 for visualization of these differences.
Fig. 4.
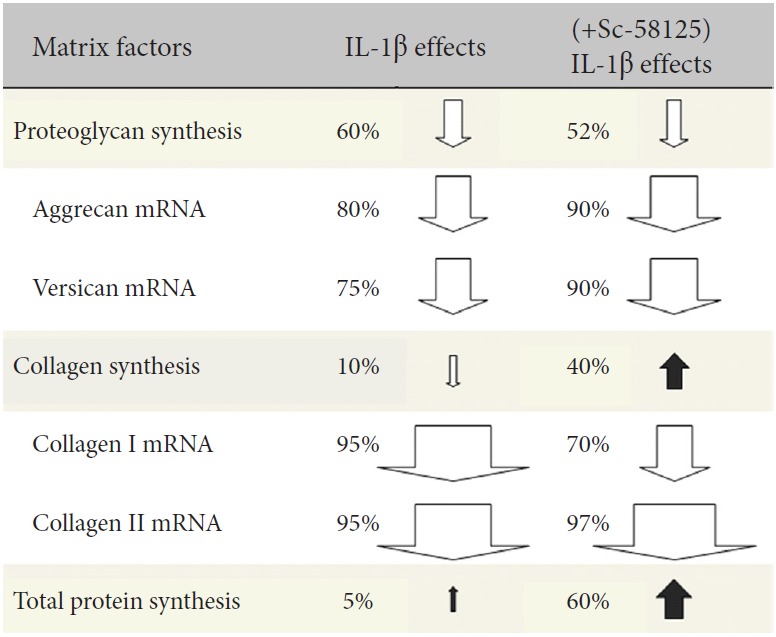
Summary of effects of COX-2 inhibition on matrix gene expression and protein synthesis in nucleus pulposus cells under inflammatory stress. COX-2, cyclooxygenase 2; IL, interleukin.
Figs. 5-6 show 3 different patterns of time-dependent response to COX-2 inhibition in IL-1 activated NP cells. As shown in Fig. 5, the 6,000-fold increase in iNOS mRNA is almost doubled by COX-2 inhibition to 11,000 at 24 hours (p = 0.021). However, by 48-hour iNOS mRNA expression is unchanged in cells treated with IL-1 alone but dropped to 2,500 when prostaglandin synthesis is inhibited by Sc (p = 0.035). By day 3, iNOS mRNA had declined to 1,700- and 816-time control, with the value in cells exposed to IL-1 + Sc still significantly less that that in cells exposed to IL-1 alone (p = 0.015). Interestingly, the accumulation of Nitrite in the CM is not significantly changed by COX-2 inhibition at any of the timepoints examined (Fig. 6, inset).
Fig. 5.
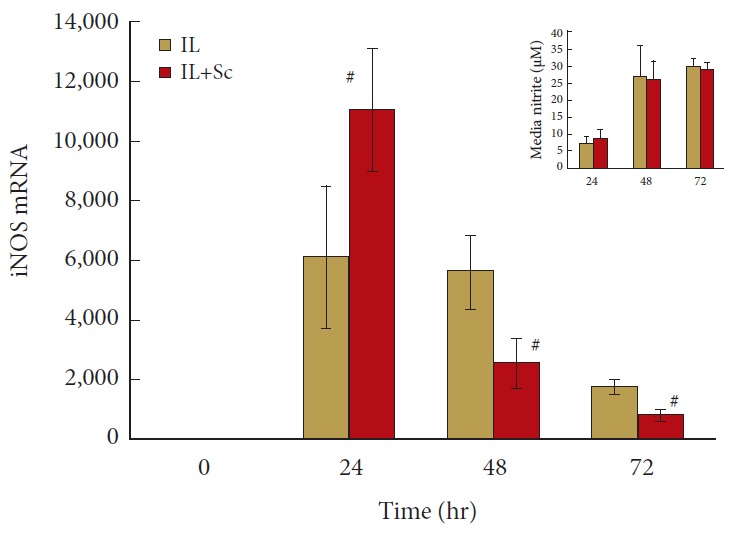
Cyclooxygenase 2 inhibition has biphasic effects on inducible nitric oxide synthase (iNOS) expression by interleukin (IL)-1 activated human nucleus pulposus (hNP) cells. hNP cells were maintained in alginate beads, activated by IL-1±Sc-58125 (Sc), and conditioned media harvested and RNA isolated after 24, 48, and 72 hours. mRNA level on the y-axis indicates fold changes relative to control untreated cells. Values are mean±standard error of n=8. #p<0.05 vs. IL-1 alone.
Fig. 6.

Cyclooxygenase 2 inhibition increases matrix metalloproteinase (MMP)-3 gene expression and accumulation in the media of interleukin (IL)-1 activated human nucleus pulposus (hNP) cells. hNP cells were treated as described for Fig. 3. n=8 for MMP-3 protein level measurement in culture media and n=12 for MMP-3 mRNA expression. mRNA level on the y-axis indicates fold changes relative to control untreated cells. Values are mean±standard error of n=8–12. Sc, Sc-58125. #p<0.05 vs. IL-1 alone.
The pattern of MMP-3 mRNA induction (Fig. 6) differs from that of iNOS. In this case, COX-2 inhibition enhances MMP-3 message in IL-1 activated cells at both 24 (from 3,000- to 11,000-fold of control, p = 0.05) and 48 hours (8,800- to 16,000-fold, p = 0.008). By 72 hours levels of message are similar to cells treated with IL-1 alone. The conditioned medium accumulation of MMP-3 reflects the changes in mRNA, with the pattern suggesting an approximately 24-hour lag time between mRNA expression changes and MMP protein synthesis.
Two other factors that might affect matrix degradation were evaluated. After 3 days of exposure, IL-1 increases mRNA for MMP-13 to 8.9±0.6-fold of control, and when COX-2 was inhibited, this induction was similar at 10.1±0.77 (n = 6). mRNA for TIMP-1, an inhibitor of MMP activity, was not changed by COX-2 inhibition in IL-1 activated cells, except at day 1 when the values are modestly increased from 1.12±0.16 to 1.72±0.26 times control. By day 3, message in IL-1 and IL-1 +Sc cells was decreased to 0.26- and 0.28-time control, respectively, and CM accumulation of TIMP-1 was not significantly different at any time (data not shown).
Fig. 7 shows that IL-1 induction of IL-6 mRNA is decreased by COX-2 inhibition, suggesting a role for prostaglandins in modulating IL-6 expression under IL-1 induced inflammatory stress. Although the differences in mRNA are significant at 48 hours (1,500 vs. 860, p = 0.029) and 72 hours (951 vs. 480, p = 0.002), there are no significant differences in CM accumulation of IL-6 at the times evaluated.
Fig. 7.
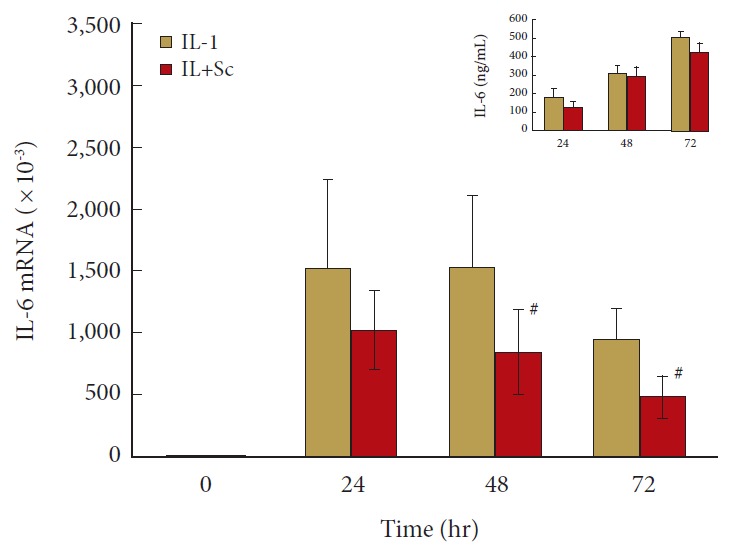
Cyclooxygenase 2 inhibition blunts human nucleus pulposus (hNP) interleukin (IL)-6 gene expression at 48 and 72 hours after IL-1 activation. hNP cells were treated as described for Fig. 3. n=4 for IL-6 protein level measurement in culture media and n=8 for IL-6 mRNA expression. mRNA level on the y-axis indicates fold changes relative to control untreated cells. Values are mean±standard error of n=4–8. Sc, Sc-58125. #p<0.05 vs. IL-1 alone.
DISCUSSION
This study was designed to determine whether any of the observed IL-1β actions on disc cells are through induction of COX-2 and subsequent prostaglandin-mediated effects on matrix metabolism. We did not observe complete abolition of IL-1β effects by COX-2 inhibition, as would be expected if COX-2 and prostaglandin activity was the only mediator of IL-1 action. However, the observation of several modifying effects by COX2 inhibition demonstrates that IL-1β actions are partially dependent on COX-2/prostaglandin actions. This is not unexpected given the numerous effects of IL-1β on cells, and the results reveal complex adaptive responses under IL-1β -induced inflammatory stress when COX-2 is inhibited.
The relationship between iNOS/NO and prostaglandins in articular cartilage has been the subject of intense study and speculation as to NO’s relevance to cartilage degeneration (recently reviewed by Weinberg et al. [14]). In general, although exogenous NO can inhibit prostaglandin production (and decreasing endogenous NO can increase prostaglandins) there has been no conclusive evidence for “cross-talk” in the reverse direction, i.e., that prostaglandins affect NO production in disc cells. This contrasts with the well-known action of prostaglandins to modulate iNOS/NO synthesis in other cell types [14,15]. We did observe significant effects of decreasing prostaglandins on iNOS message in IL-1 activated hNP with an initial increase, followed by decreases at both 48 and 72 hours. Although the accumulation of nitrite in the CM at 24-hour intervals was not significantly different, we cannot know whether there may be transient differences in tissue exposure to NO that could modulate cell function. Regardless, the data suggests that prostaglandins can affect IL-1 induction of iNOS in hNP; the consequences for disc matrix homeostasis remain to be explored.
Sc almost completely inhibits both PGE2 and PGF2α accumulation, thus diminishing NP cell exposure to these and potentially other prostaglandins whose syntheses are mediated by COX-2 after IL-1 activation. Proteoglycan, collagen, and total protein synthesis were all enhanced by COX-2 inhibition in IL-1β activated cells. These changes were seen in spite of the fact that COX-2 inhibition decreased aggrecan, versican, and collagen II mRNA, and are in contrast to the observations in cells not exposed to inflammatory stimuli [2]. The increase in message for collagen I in IL-1β activated cells might account in part for the increase in collagen synthesis, i.e., collagenase sensitive 3H proline incorporation. However, as shown in Fig. 2, in spite of the effect of prostaglandin synthesis inhibition to increase collagen I message, the values are still significantly less than control during the time that synthesis was evaluated, e.g., 24–48 hours, when Col I mRNA was between 55% and 35% of control. These data suggest that when prostaglandin levels are decreased, hNP cells experience a general, enhanced protein synthesis. Although extrapolation to the in vivo situation from the response in isolated cell experiments cannot be done with certainty, the results suggest that prostaglandins produced by cells of the NP could diminish matrix protein synthesis in IL-1β activated hNP, and thus therapy to minimize prostaglandins may be beneficial to the anabolic balance of the disc when the disc is in a pro-inflammatory state.
The observed changes in collagen I and collagen II mRNA are consistent with the well-documented actions of PGE2 on chondrocyte expression of these matrix proteins [15,16]. Blocking prostaglandin accumulation increases collagen I message and decreases message for collagen II, thus we infer that prostaglandins in IL-1β activated hNP decrease collagen I and conversely increase message for collagen II, as PGE2 does in chondrocytes [16]. Addition of exogenous PGE2 decreased message for both collagen I and collagen II in hNP cells [2], suggesting that regulation of these matrix components, especially collagen II, is determined by signal pathways activated by IL-1β, which over-ride those activated by PGE2 alone in the cells under a noncytokine activated state.
The effects on hNP cell collagen and proteoglycan synthesis, however, are not consistent with some previous data from cartilage [17] and a rat chondrocyte cell line [18] where PGE2 actually stimulated matrix protein synthesis. However, our observation that COX-2 inhibition increases hNP proteoglycan synthesis is consistent with recent studies showing that selective COX-2 inhibitors which depress PGE2 can actually increase proteoglycan synthesis by chondrocytes in situ in cartilage explants [19,20]. More germane to the actions of prostaglandins in hNP, the increase in proteoglycan synthesis when prostaglandin synthesis is inhibited is consistent with the decrease in synthesis seen in hNP exposed to exogenous PGE2 [2]. This again suggests that therapy, which blunts ecoisanoid accumulation, could enhance matrix proteoglycan synthesis. However, a limitation of our study is that disc cells were cultured in normoxia instead of hypoxia, which is known to influence matrix production to some extent.
The matrix synthesis results do not directly correlate with the gene expression data. Intuitively one would expect gene expression to be reflected in matrix synthesis and production of conditioned medium factors. However, the translation of genes into proteins, especially the complex proteins of the disc matrix, involves multiple regulated steps in synthesis, cellular transport, and export to the extracellular space. Another obstacle to reconciling the dissociation between matrix protein gene expression and synthesis is that we have evaluated mRNA for only a few of the genes that may contribute to total “synthesis.” For example, microarray analysis of hNP showed high levels of expression for multiple proteoglycans other than aggrecan and versican in these cells [21] with mRNA for Decorin > Syndecan 2 > Mimecan > Versican > Aggrecan. Others have shown that relative abundance of the small leucine-rich proteoglycans decorin, biglycan, and lumican in the disc may vary with mechanical force as well as stage of degeneration [22]. Microarray analysis identified multiple forms of collagen as well, with mRNA for collagen I> XV> VI> III> XI> II [21]. Eyre et al. [23] have reviewed the collagens of the disc, which included those listed above as well as collagen IIIAI and collagen VIA1-3, and confirmed that, regardless of the relative abundance of message, the bulk of normal disc matrix is composed collagen types I and II. Polymorphisms in collagens are associated with susceptibility to disc degeneration [24,25], thus it may be important to determine how different collagens, as well as proteoglycans, are modulated by cytokines/prostaglandins, and how changes in their relative abundance affect the structure and function of the disc.
A time-dependent modulation of message and protein of catabolic mediators by COX-2 inhibition in IL-1β activated cells is clearly shown in the results with iNOS, MMP-3, and IL-6. iNOS mRNA was initially elevated and then depressed at a later time (Fig. 5). MMP-3 mRNA was also elevated but returned to IL-1β alone levels by 72 hours (Fig. 6), while IL-6 mRNA consistently decreased throughout the 3-day period evaluated (Fig. 7). Thus COX-2 mediates IL-1β action on disc NP cells through complex temporal regulation of these catabolic factors. Whether COX-2 inhibition modulates net matrix loss from the catabolic action of IL-1β on disc tissue requires further investigation.
Blocking prostaglandin synthesis actually increased both message and synthesis/conditioned medium accumulation of MMP-3, suggesting that the prostaglandins normally produced by IL-1 activated hNP could limit disc matrix exposure to this metalloproteinase. This contrasts with the results seen when hNP were exposed to either PGE2 or PGF2α alone, with PGF2α increasing both MMP-1 and -3 message while PGE2 diminished MMP1 message without affecting that for MMP-3. The differences may be explained by alterations in COX-2-mediated synthesis of prostaglandin products other than PGE2 and PGF2α, or by the complex interplay of signaling pathways in hNP exposed to prostaglandins alone versus those activated in IL-1 stimulated cells.
The relevance of any influence of COX-2 inhibition on IL-6 awaits further investigation. The data suggest a direct relationship between IL-1 induced prostaglandin concentration and induction of IL-6. This is consistent with studies showing PGE2 is necessary for IL-1 induction of IL-6 in human articular chondrocytes [26] and synovium [27]. IL-6 can have anti- and pro-inflammatory actions in arthritis [28] and Legendre et al. [29,30] showed IL-6 down regulation of matrix protein transcription and up regulation of MMP and ADAMTs gene expression in chondrocytes and Flannery et al. [31] reported that IL-6 augments aggrecanase-mediated proteoglycan catabolism in cartilage. Cells of degenerating discs can produce high concentrations of IL-6 [6], however, little is know concerning the effects, if any, of IL-6 to change disc cell metabolism. IL-6 in combination with its soluble receptor can decrease proteoglycan synthesis and acutely enhance IL-1 and TNF-α mediated inhibition of proteoglycan synthesis in human NP [13]. However, in the current studies the average IL-6 in CM was not changed by COX-2 inhibition in IL-1β activated cells, in spite of the decrease in message. Whether there could be discrete, localized differences in IL-6 production and autocrine action remain questions for future experiments.
CONCLUSION
In conclusion, COX-2 inhibition significantly affects the response of hNP cells to IL-1β. The dissociation between iNOS message/nitrite accumulation, aggrecan and versican message/proteoglycan synthesis, and collagen message/synthesis suggests actions downstream of transcription or actions affecting other proteoglycans and collagens that should be investigated. The data suggest that blocking prostaglandin synthesis in cytokine activated hNP cells could have both anabolic and catabolic effects; determining the specific mechanisms involved and whether these changes can alter the course of IDD requires further study.
Acknowledgments
The authors gratefully acknowledge the following funding sources that enabled the success of this project: the National Institute of Health (AG044376 to Nam Vo), VA Rehabilitation Research and Development Grant # B4-3894RA to Studer R, and the Albert B Ferguson Jr. MD Orthopaedic Fund of the Pittsburgh Foundation to Nam Vo, Gwendolyn Sowa, and James Kang.
The authors acknowledge the technical assistance of SuLan Huang, Elizabeth Christian, Geoffrey Sirockman, and the support of the personnel of the Ferguson Lab for Orthopaedic Research. The authors would like the thank Jessa Darwin for providing editorial assistance on the manuscript.
Footnotes
The authors have nothing to disclose.
REFERENCES
- 1.Freemont AJ. The cellular pathobiology of the degenerate intervertebral disc and discogenic back pain. Rheumatology (Oxford) 2009;48:5–10. doi: 10.1093/rheumatology/ken396. [DOI] [PubMed] [Google Scholar]
- 2.Vo NV, Sowa GA, Kang JD, et al. Prostaglandin E2 and prostaglandin F2α differentially modulate matrix metabolism of human nucleus pulposus cells. J Orthop Res. 2010;28:1259–66. doi: 10.1002/jor.21157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hiyama A, Yokoyama K, Nukaga T, et al. Response to tumor necrosis factor-α mediated inflammation involving activation of prostaglandin E2 and Wnt signaling in nucleus pulposus cells. J Orthop Res. 2015;33:1756–68. doi: 10.1002/jor.22959. [DOI] [PubMed] [Google Scholar]
- 4.Le Maitre CL, Hoyland JA, Freemont AJ. Catabolic cytokine expression in degenerate and herniated human intervertebral discs: IL-1beta and TNFalpha expression profile. Arthritis Res Ther. 2007;9:R77. doi: 10.1186/ar2275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Weiler C, Nerlich AG, Bachmeier BE, et al. Expression and distribution of tumor necrosis factor alpha in human lumbar intervertebral discs: a study in surgical specimen and autopsy controls. Spine (Phila Pa 1976) 2005;30:44–53. doi: 10.1097/01.brs.0000149186.63457.20. discussion 54. [DOI] [PubMed] [Google Scholar]
- 6.Kang JD, Stefanovic-Racic M, McIntyre LA, et al. Toward a biochemical understanding of human intervertebral disc degeneration and herniation. Contributions of nitric oxide, interleukins, prostaglandin E2, and matrix metalloproteinases. Spine (Phila Pa 1976) 1997;22:1065–73. doi: 10.1097/00007632-199705150-00003. [DOI] [PubMed] [Google Scholar]
- 7.Miyamoto H, Saura R, Doita M, et al. The role of cyclooxygenase-2 in lumbar disc herniation. Spine (Phila Pa 1976) 2002;27:2477–83. doi: 10.1097/00007632-200211150-00011. [DOI] [PubMed] [Google Scholar]
- 8.Jimbo K, Park JS, Yokosuka K, et al. Positive feedback loop of interleukin-1beta upregulating production of inflammatory mediators in human intervertebral disc cells in vitro. J Neurosurg Spine. 2005;2:589–95. doi: 10.3171/spi.2005.2.5.0589. [DOI] [PubMed] [Google Scholar]
- 9.O’Donnell JL, O'Donnell AL. Prostaglandin E2 content in herniated lumbar disc disease. Spine (Phila Pa 1976) 1996;21:1653–5. doi: 10.1097/00007632-199607150-00007. discussion 1655-6. [DOI] [PubMed] [Google Scholar]
- 10.Thompson JP, Pearce RH, Schechter MT, et al. Preliminary evaluation of a scheme for grading the gross morphology of the human intervertebral disc. Spine (Phila Pa 1976) 1990;15:411–5. doi: 10.1097/00007632-199005000-00012. [DOI] [PubMed] [Google Scholar]
- 11.Studer RK, Aboka AM, Gilbertson LG, et al. p38 MAPK inhibition in nucleus pulposus cells: a potential target for treating intervertebral disc degeneration. Spine (Phila Pa 1976) 2007;32:2827–33. doi: 10.1097/BRS.0b013e31815b757a. [DOI] [PubMed] [Google Scholar]
- 12.Gilbertson L, Ahn SH, Teng PN, et al. The effects of recombinant human bone morphogenetic protein-2, recombinant human bone morphogenetic protein-12, and adenoviral bone morphogenetic protein-12 on matrix synthesis in human annulus fibrosis and nucleus pulposus cells. Spine J. 2008;8:449–56. doi: 10.1016/j.spinee.2006.11.006. [DOI] [PubMed] [Google Scholar]
- 13.Studer RK, Vo N, Sowa G, et al. Human nucleus pulposus cells react to IL-6: independent actions and amplification of response to IL-1 and TNF-α. Spine (Phila Pa 1976) 2011;36:593–9. doi: 10.1097/BRS.0b013e3181da38d5. [DOI] [PubMed] [Google Scholar]
- 14.Weinberg JB, Fermor B, Guilak F. Nitric oxide synthase and cyclooxygenase interactions in cartilage and meniscus: relationships to joint physiology, arthritis, and tissue repair. Subcell Biochem. 2007;42:31–62. doi: 10.1007/1-4020-5688-5_2. [DOI] [PubMed] [Google Scholar]
- 15.Amin AR, Dave M, Attur M, et al. COX-2, NO, and cartilage damage and repair. Curr Rheumatol Rep. 2000;2:447–53. doi: 10.1007/s11926-000-0019-5. [DOI] [PubMed] [Google Scholar]
- 16.Goldring MB, Berenbaum F. The regulation of chondrocyte function by proinflammatory mediators: prostaglandins and nitric oxide. Clin Orthop Relat Res. 2004;(427 Suppl):S37–46. doi: 10.1097/01.blo.0000144484.69656.e4. [DOI] [PubMed] [Google Scholar]
- 17.Di Battista JA, Doré S, Martel-Pelletier J, et al. Prostaglandin E2 stimulates incorporation of proline into collagenase digestible proteins in human articular chondrocytes: identification of an effector autocrine loop involving insulin-like growth factor I. Mol Cell Endocrinol. 1996;123:27–35. doi: 10.1016/0303-7207(96)03887-7. [DOI] [PubMed] [Google Scholar]
- 18.Lowe GN, Fu YH, McDougall S, et al. Effects of prostaglandins on deoxyribonucleic acid and aggrecan synthesis in the RCJ 3.1C5.18 chondrocyte cell line: role of second messengers. Endocrinology. 1996;137:2208–16. doi: 10.1210/endo.137.6.8641167. [DOI] [PubMed] [Google Scholar]
- 19.El Hajjaji H, Marcelis A, Devogelaer JP, et al. Celecoxib has a positive effect on the overall metabolism of hyaluronan and proteoglycans in human osteoarthritic cartilage. J Rheumatol. 2003;30:2444–51. [PubMed] [Google Scholar]
- 20.Mastbergen SC, Jansen NW, Bijlsma JW, et al. Differential direct effects of cyclo-oxygenase-1/2 inhibition on proteoglycan turnover of human osteoarthritic cartilage: an in vitro study. Arthritis Res Ther. 2006;8:R2. doi: 10.1186/ar1846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vo N, Seidel C, Coelho P, et al. Identification of novel TNF-α regulated extracellular matrix and cell-matrix adhesion genes in intervertebral disc cells: a microarray gene profiling of human nucleus pulposus cells [abstract]. National Asian American Survey; 2008. [Google Scholar]
- 22.Chen J, Yan W, Setton LA. Static compression induces zonal-specific changes in gene expression for extracellular matrix and cytoskeletal proteins in intervertebral disc cells in vitro. Matrix Biol. 2004;22:573–83. doi: 10.1016/j.matbio.2003.11.008. [DOI] [PubMed] [Google Scholar]
- 23.Eyre DR, Matsui Y, Wu JJ. Collagen polymorphisms of the intervertebral disc. Biochem Soc Trans. 2002;30(Pt 6):844–8. doi: 10.1042/bst0300844. [DOI] [PubMed] [Google Scholar]
- 24.Aladin DM, Cheung KM, Chan D, et al. Expression of the Trp2 allele of COL9A2 is associated with alterations in the mechanical properties of human intervertebral discs. Spine (Phila Pa 1976) 2007;32:2820–6. doi: 10.1097/BRS.0b013e31815b75c5. [DOI] [PubMed] [Google Scholar]
- 25.Mio F, Chiba K, Hirose Y, et al. A functional polymorphism in COL11A1, which encodes the alpha 1 chain of type XI collagen, is associated with susceptibility to lumbar disc herniation. Am J Hum Genet. 2007;81:1271–7. doi: 10.1086/522377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Clausen PA, Flechtenmacher J, Haeuselmann HJ, et al. Evidence of an eicosanoid contribution to IL-1 induction of IL-6 in human articular chondrocytes. Am J Ther. 1996;3:101–8. doi: 10.1097/00045391-199602000-00002. [DOI] [PubMed] [Google Scholar]
- 27.Bukata SV, Gelinas J, Wei X, et al. PGE2 and IL-6 production by fibroblasts in response to titanium wear debris particles is mediated through a Cox-2 dependent pathway. J Orthop Res. 2004;22:6–12. doi: 10.1016/S0736-0266(03)00153-0. [DOI] [PubMed] [Google Scholar]
- 28.Wong PK, Campbell IK, Egan PJ, et al. The role of the interleukin-6 family of cytokines in inflammatory arthritis and bone turnover. Arthritis Rheum. 2003;48:1177–89. doi: 10.1002/art.10943. [DOI] [PubMed] [Google Scholar]
- 29.Legendre F, Dudhia J, Pujol JP, et al. JAK/STAT but not ERK1/ERK2 pathway mediated IL-6/soluble IL-6R down-regulation of type II collagen, aggrecan core, and link protein transcription in articular chondrocytes. Association with a downregulation of SOX9 expression. J Biol Chem. 2003;278:2093–12. doi: 10.1074/jbc.M110773200. [DOI] [PubMed] [Google Scholar]
- 30.Legendre F, Bogdanowicz P, Boumediene K, et al. Role of interleukin 6 (IL-6)/IL-6R-induced signal tranducers and activators of transcription and mitogen-activated protein kinase/extracellular. J Rheumatol. 2005;32:1307–16. [PubMed] [Google Scholar]
- 31.Flannery CR, Little CB, Hughes CE, et al. IL-6 and its soluble receptor augment aggrecanase-mediated proteoglycan catabolism in articular cartilage. Matrix Biol. 2000;19:549–53. doi: 10.1016/s0945-053x(00)00111-6. [DOI] [PubMed] [Google Scholar]