

Abstract
Polycyclic aromatic hydrocarbons (PAHs) are ubiquitous environmental contaminants generated from combustion of carbon-based matter. Upon ingestion these molecules can be bioactivated by cytochrome P450 monooxygenases to oxidized toxic metabolites. Some of these metabolites are potent carcinogens that can form irreversible adducts with DNA and other biological macromolecules. Conjugative enzymes, such as glutathione S-transferases or UDP-glucuronosyltransferases, are responsible for the detoxification and/or facilitate the elimination of these carcinogens. While responses to PAH exposures have been extensively studied for the bioactivating cytochrome P450 enzymes, much less is known regarding the response of glutathione S-transferases in mammalian systems. In this study, we investigated the expression and activity responses of murine hepatic glutathione S-transferases to benzo[a]pyrene exposure using global proteomics and activity-based protein profiling for chemoproteomics, respectively. Using this approach, we identified several enzymes exhibiting increased activity including GSTA2, M1, M2, M4, M6, and P1. The activity of one GST enzyme, GSTA4, was found to be downregulated with increasing B[a]P dose. Activity responses of several of these enzymes were identified as being expression-independent when comparing global and activity-based datasets, possibly alluding to as of yet unknown regulatory post-translational mechanisms.
Keywords: polycyclic aromatic hydrocarbons, benzo[a]pyrene, chemoproteomics, glutathione-S-transferase
Graphical Abstract
Introduction
Polycyclic aromatic hydrocarbons (PAHs) are a group of ubiquitously found environmental contaminants generated by exposing carbon-based materials to very high heat or combustion.1 Many common sources of PAH exposure come from anthropogenic processes such as combustion of fossil fuels, grilling or smoking meat, smoking, and other industrial processes.2–4 Upon ingestion, some relatively non-toxic PAHs are converted to their toxic metabolites by cytochrome P450s. These biotransformation enzymes oxygenate PAHs to a variety of hydroxylated metabolites, diols, epoxides, diol epoxides, and diones.5, 6 One product of the P450 monooxygenation of benzo[a]pyrene (B[a]P) is the prototypical PAH carcinogen, benzo[a]pyrene-7,8-dihydrodiol-9,10-oxide (BPDE). Not only are some PAHs bioactivated to highly carcinogenic metabolites by P450s, they also perpetuate further increases in their own bioactivation via P450 induction7. While P450 bioactivation and induction results in the production of carcinogenic metabolites, detoxifying enzymes convert these metabolites into non-toxic products and facilitate their expulsion. These enzymes include UDP glucuronosyl-transferases, sulfotransferases, and glutathione S-transferases.
Glutathione S-transferases conjugate glutathione to a diverse group of xenobiotics. The active site of GSTs consists of two distinct subsites: the glutathione-binding G site and substrate-binding H site.8 The primary role of GSTs in PAH metabolism is thought to be the glutathione conjugation of PAH metabolites, such as epoxides and diol epoxides.9–15 Induction of GST expression is thought to be caused by changes in redox conditions; for instance, one study showed that the Aryl hydrocarbon receptor (Ahr) mediated little induction of GSTs, and that induction via NFE2-related factor 2 (Nrf2) is much more potent.16 Previously, GST induction by polycyclic aromatic hydrocarbons (PAHs) has been measured by determining mRNA levels, by western blotting for protein abundance, via GSH-agarose column enriched samples, and/or by GST activity via simple colorimetric assays.17–19 Western blotting and measurement of mRNA levels provide information regarding the expression but are not necessarily indicative of enzyme activities. Utilizing GSH-agarose columns to enrich induced samples provides meaningful insight into GSTs that maintain their GSH binding activity, but do not explore the activity of GST H-sites. The studies referenced above measure total GST activity via colorimetric assay and though some isoform and class-specific assays are available, many GST isoforms still lack specific assays in part due to high substrate overlap of different GSTs of the same class.20–23 In a broader sense, the majority of investigations into the role of GSTs following PAH exposure have been performed using mammalian cell lines.24–26 These models of induction do not take into account many complex factors in a mammalian system including absorption, distribution, and excretion, all of which modulate the internal dose observed by individual organs.
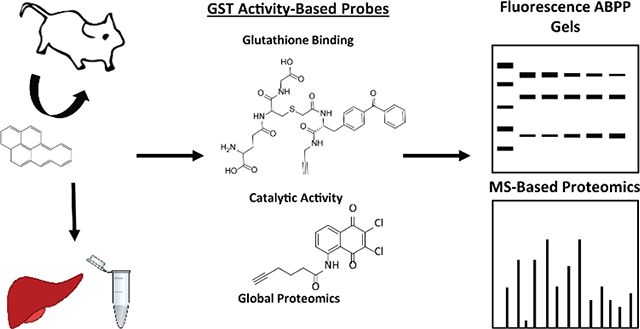
Activity-based protein profiling (ABPP) is a proven method to enrich and quantify enzymes based upon functional activity in biological systems.27 The ABPP approach uses activity-based probes (ABPs) designed to target and covalently bind enzymes of a specific function. Recently, we reported on the synthesis and validation of two ABPs that irreversibly bind and target GSTs and report on their enzymatic activity in mammalian tissues.28 One probe, GSTABP-G, features a glutathione molecule to which benzophenone and alkyne moieties have been appended to the functional thiol of glutathione. This ABP effectively investigates the glutathione binding activities of GSTs. The second probe, GSTABP-H, is based on the known suicide inhibitor of GSTs, 2,3-dicholoro-1,4-naphthoquinone. This probe has been validated to bind active GSTs in substrate binding H sites. Both probes feature an alkyne handle to facilitate click chemistry29 mediated addition of azide appended reporter groups such as TAMRA-azide (for visualization of probe targets by SDS-PAGE and fluorescent detection) or biotin-azide (for streptavidin-biotin enrichment). The use of these probes offers a unique look into the activity of these phase II biotransformation enzymes that is not possible using other methods to measure activity.
The purpose of this study was to investigate B[a]P induction of active hepatic GSTs using chemoproteomic probes. This activity-based proteomics approach results in investigation of activity with improved resolution that allows for a much closer investigation into susceptibilities associated with xenobiotic exposures. While methods such as measurement of GST mRNA levels and western blotting have provided information regarding induction of GST expression by PAHs, they provide no indication of enzyme-specific activity. In addition, mice were used as an in vivo animal model to capture full organismal response, which cannot be replicated in cell culture systems. Herein, using ABPP in tandem with global proteomics analyses we investigated B[a]P-specific GST induction of both active GST G and H sites in the liver.
Materials and Methods
Animals
Our group has utilized female B6129SF1/J mice in prior PAH carcinogenicity, toxicokinetic, and in vitro metabolism studies,30–34 and was subsequently chosen for this study. Adult female B6129SF1/J mice were purchased from The Jackson Laboratory (Bar Harbor, ME, USA). Mice were fed Lab Certified rodent chow (PMI Nutrition International Certified Rodent Diet 5002) ad libitum.
Effects of PAHs on GST induction was evaluated using a serial sacrifice design. After acclimation for 1 week, mice were dosed with a water vehicle, a corn oil vehicle, or B[a]P in a corn oil vehicle by oral gavage at 2, 20, or 180 μmol/kg every 24 hr for 3 days (at t = 0 hr, t = 24 hr, t = 48 hr). Four mice were dosed per dosing level. After 72 hr (24 hr after the third consecutive dose), mice were euthanized by CO2 asphyxiation and cervical dislocation. After euthanization, livers were rapidly removed, rinsed in 0.1 M phosphate-buffered saline (PBS, pH 7.4), frozen in liquid nitrogen, and stored at −70 °C. All protocols were approved by the Pacific Northwest National Laboratory Institutional Animal Care and Use Committee.
Preparation of liver cytosolic fraction
Hepatic microsomes were prepared by differential centrifugation. Livers (n=4) from each dosing condition were pooled and minced using a tissue tearor. Minced liver tissue was subsequently homogenized in sucrose (250mM) in PBS solution using a glass dounce homogenizer with 15 pulses each from loose and tight-fitting pestles. Whole liver lysate was centrifuged at 10,000 × g for 25 min at 4 °C. The supernatant S9 fraction was collected and centrifuged again at 100,000 × g to separate the cytosolic (supernatant) and microsomal (pellet) fractions. Cytosolic proteins were quantified via the bicinchoninic acid assay (BCA).35
Activity-based probe labeling and click chemistry
GST enzymes were labeled using two different ABPs. Cytosolic hepatic proteins were normalized to 1 mg/mL protein (2 mg/mL for SDS-PAGE analysis). 100 μg (for SDS-PAGE gels) or 500 μg (for streptavidin enrichment for MS-based chemoproteomics) were incubated with GSTABP-G (20 μM), GSTABP-H (20 μM), or DMSO only controls for 30 min at 37 °C, shaking at 1000 rpm. GSTABP-G labeled proteins were then UV exposed for 7 min to activate the benzophenone for covalent labeling of protein targets. Following probe incubation, click chemistry reactions were conducted on all samples. The following reagents were added to each sample in the following order: (1) TAMRA-azide (60 μM) for SDS-PAGE analysis or biotin-azide (60 μM) for biotin streptavidin enrichment, sodium ascorbate (10 mM), tris-hydroxypropyltriazolylmethylamine (2 mM), and CuSO4 (4 mM). Samples were incubated at room temperature for 90 min. Following click chemistry, ice cold MeOH (60 %) was added to each sample. Samples were then placed in a −70 °C freezer for 1 hr to induce further protein precipitation. Precipitated proteins were pelleted via centrifugation at 14,000 × g for 10 min and the supernatant was discarded. Proteins were resolubilized by adding SDS (1.2 %) in PBS, sonicated with 2 × 1 s pulses at 60 % amplitude. Samples were then heated at 95 °C for 2 min. Protein from samples undergoing streptavidin enrichment were quantified via BCA.
SDS-PAGE
SDS-PAGE was used to visualize activity-based probe labeling. Probe labeled, TAMRA appended proteins were resolved via SDS-PAGE. 2X NuPage Running Buffer and 10X NuPage Reducing Agent were added to resolubilized protein samples. 20 μg protein was loaded into each well of 4 – 12 % Bis-Tris gel cassettes. Gels were run at 150 V, 35 mA for 1 hr in MES buffer. Gels were imaged using a Typhoon FLA 9500 (General Electric). Fluorescent bands were quantified using ImageJ software. After imaging, gels were fixed with a solution of MeOH (50 %), acetic acid (7 %), and water (43 %) and then stained with GelCode Blue. Stained gels were rinsed with water and imaged using GelDocEZ (BioRad Laboratories).
Enrichment of ABP Targets for MS Analysis
Probe labeled, biotin appended proteins were normalized to 450 μg protein per sample, in SDS (1.2 %) in PBS solution. 100 μL streptavidin agarose resin (binding capacity 1–3 mg protein/ 1 mL resin) was washed with SDS (0.5 %) in PBS, urea (6M) in NH4HCO3 (25 mM, pH = 8), and PBS. Beads were incubated with samples in PBS at 37 °C, with rotation, for 1 hr. Following enrichment, beads were transferred to fritted columns (BioRad) and washed under vacuum with SDS (0.5 %) in PBS, urea (6M) in NH4HCO3 (25 mM), PBS, NH4HCO3 (25mM) and MilliQ water. Beads were then suspended in urea (6M) in NH4HCO3 (25 mM). Beads were incubated with tris(2-carboxyethyl)phosphine hydrochloride (5mM) at 37 °C for 30 min to reduce the samples followed by incubation with iodoacetamide (10 mM) at 50 °C for 45 min to alkylate samples. Beads were again placed on the fritted columns and washed under vacuum with PBS and NH4HCO3 (25 mM). On-bead trypsin digestion was carried out overnight with 0.125 μg trypsin (Promega) in NH4HCO3 (25 mM, pH = 8.0) at 37 °C, with rotation. Trypsinized peptides were collected and dried using a speedvac concentrator. Dried peptides were resuspended in NH4HCO3 (25 mM) and analyzed via LC-MS/MS.
Preparation of peptides for global proteomics analysis
Cytosolic proteins from all samples were normalized to 1 mg/mL protein concentration. Urea (6 M) was added to 100 μL normalized protein. Protein was reduced with dithiothreitol (5 mM) at 60 °C for 30 min. Samples were subsequently alkylated with iodoacetamide (40 mM) for 1 hr at 37 °C. Samples were then diluted 10-fold with NH4HCO3 and incubated with 2 μg trypsin (Promega) for 3 hr at 37 °C. Digested peptides were washed on Discovery C18 SPE columns conditioned and equilibrated with methanol and trifluoroacetic acid (0.1%), respectively. After capture on the columns peptides were washed with acetonitrile (5 %) in water and eluted with acetonitrile (80 %) in water. Eluted peptides were dried using a speedvac concentrator, resuspended in NH4HCO3, and normalized to 0.1 mg/mL peptide concentration (via the BCA assay) for LC-MS/MS analysis.
Proteomics data analysis
All peptides were analyzed on a Velos Orbitrap mass spectrometer as outlined in Sadler et. al.36 The resulting peptide spectra were searched against the Uniprot37 mus_musculus database and rescored using MSGF +.38 Following that, an AMT tag analysis was conducted as previously described.39 Only unique peptides with MT FDR threshold ≤ 0.1 and MT Uniqueness ≥ 0.5 were used in further analysis. Peptides meeting these criteria were log2 transformed and rolled up to the protein level using Inferno RRollup software.40 Enrichment of GST targets were validated using paired student’s t-test and fold change analysis of probe vs. no probe samples. Treatment effects were evaluated by testing the slope of a linear regression model and comparing fold changes across treatments. Fold changes >2 and p-values <0.05 were used as thresholds in determining significant dose dependence. Pearson correlation analysis was also conducted to investigate relationships of GSTABP-G and GSTABP-H biotin-streptavidin enriched samples and global proteomics samples.
Results and Discussion
B[a]P induction of GSTs measured via gel electrophoresis
We investigated total probe labeling of GSTs by GSTABP-G and H ABPs in vehicle and B[a]P treated samples. This was measured via fluorescence of probe labeled/TAMRA appended proteins resolved via SDS-PAGE (Figure 1, Figure S1, Figure S2). No apparent differences in probe-measured total GST activities were observed between water and corn oil vehicle. Though no obvious B[a]P dependent changes in probe-determined GST activity were observed, fluorescence was increased over corn oil vehicle dosed samples by 61% in GSTABP-H probed liver cytosol from mice treated with 180 μmol/kg.
Figure 1.

(A) GSTABP-G and (B) GSTABP-H labeling of liver cytosol from mice exposed to water vehicle, corn oil vehicle, 2, 20, or 180 μmol/kg B[a]P. The fluorophore TAMRA was attached to probe-labeled proteins via click and the proteins were separated by electrophoresis and visualized and quantified by fluorescence. The prominent band at roughly 25 kDa (GST molecular weight) was quantified using ImageJ software and charted as the upper bar graphs; values are depicted relative to corn oil vehicle. Error bars represent 95% CI of the average of three technical replicates.
While apparent increases in GST fluorescence observed by both probes were minimal, many GSTs contribute to the total fluorescence at 25 kDa and resolution of individual GSTs is not possible. Due to differences in affinity of individual GSTs for the probes, as well as large differences in expression between them, minimal changes seen via gel don’t necessarily equate to minimal changes in the activities of individual GSTs. Similar method-inherent caveats apply when using commercially-available GST activity assays measuring DNCB or ethacrynic acid glutathione conjugation. Thus, to determine individual GSTs responsible for these observed changes in probe-mediated fluorescence, we enriched probe-targeted GSTs in these B[a]P treated samples.
ABPP of liver cytosolic GSTs
Hepatic cytosolic GSTs from water, corn oil, and B[a]P dosed mice (2, 20, 180 μmol/kg) were enriched in murine liver lysate by labeling with 20 μM of each GSTABP followed by the addition of a biotin molecule via click chemistry. Proteins were then incubated with streptavidin agarose beads and subsequently enriched for LC-MS/MS analysis. Both probes significantly enriched 14 GSTs as depicted in Table 1. These GSTs included GSTA2, A3, A4, K1, M1, M2, M4, M5, M6, M7, P1, T1, T2, and O1. Significant enrichment of these enzymes was determined via fold change analysis and paired student’s t-tests comparing enriched versus corresponding no probe controls for all doses/vehicle controls. Enrichment of GST targets was highly significant with p-values for the majority of enzymes well below 0.001 and fold changes greater than 3 and as high as 1400. All 14 of these enzymes were also detected in global proteomic analysis of the same lysates. In total, the probe enrichment yielded excellent coverage of GSTs (14/19 of the total number of mammalian cytosolic GSTs) for an effective and broad analysis of the inductive capacity of B[a]P in murine liver.
Table 1.
Enrichment (probe vs. DMSO only) of GSTs in liver cytosols from mice treated with B[a]P and vehicle controls
| GSTABP-G | GSTABP-H | |||
|---|---|---|---|---|
| Protein | Fold Change Enrichment | p value (t-test) | Fold Change Enrichment | p value (t-test) |
| GSTA2 | 53.1 | 1.67E-02 | 116.2 | 8.73E-03 |
| GSTA3 | 43.7 | 1.86E-15 | 141.9 | 6.88E-16 |
| GSTA4 | 48.6 | 1.82E-11 | 24.7 | 1.63E-08 |
| GSTK1 | 8.4 | 6.15E-13 | 72.0 | 1.74E-20 |
| GSTM1 | 234.4 | 1.53E-15 | 164.4 | 1.21E-12 |
| GSTM2 | 131.7 | 1.96E-13 | 45.9 | 6.67E-10 |
| GSTM4 | 93.5 | 3.12E-13 | 1251.8 | 3.61E-16 |
| GSTM5 | 8.2 | 5.08E-10 | 370.7 | 2.84E-16 |
| GSTM6 | 29.6 | 6.14E-07 | 171.5 | 6.66E-08 |
| GSTM7 | 40.4 | 2.92E-10 | 61.7 | 3.76E-10 |
| GSTO1 | 3.8 | 3.38E-09 | 84.3 | 3.89E-15 |
| GSTP1 | 19.2 | 1.88E-10 | 699.7 | 7.07E-16 |
| GSTT1 | 10.2 | 4.86E-09 | 545.2 | 3.56E-15 |
| GSTT2 | 6.3 | 5.61E-05 | 1400.9 | 4.29E-13 |
B[a]P induced changes in activity of GST G sites
B[a]P induced active G sites of several enzymes as measured by GSTABP-G. Significantly induced enzymes include GSTA2, GSTM4, and GSTP1. Significance of induction was determined via linear regression analysis (p ≤ 0.05) and fold change abundances of 180 μmol/kg dose versus corn oil controls (fold change ≥ 2). GSTA2 showed 2.7, 2.0, and 3.3-fold increases in 2, 20, and 180 μmol/kg B[a]P treated samples, respectively. GSTP1 was induced 2.3- and 2.5-fold at 20 and 180 μmol/kg B[a]P doses, respectively. Interestingly, GSTA4 showed significant downregulation as determined by fold change and linear regression analysis with 2.5-fold inhibition at 180 μmol/kg B[a]P (Figure 2, Table S1, Figure S3).
Figure 2.

Global proteomics and ABPP results as depicted by volcano plots. Log2 fold changes of relative abundances of GSTs in 180 μmol/kg B[a]P vs. corn oil vehicle samples are plotted on the x axes. –log10 p-values from linear regression of all samples/conditions are plotted on the y axes. Dotted lines are located at p = 0.05 and log2 fold change = 1. Thus, values in the upper right and left quadrants are GSTs that are significantly induced by B[a]P treatment. Analyses are from 3 technical replicates from pooled cytosolic samples from 4 individually exposed mice.
The glutathione binding sites of cytosolic GSTs are composed of a set of highly conserved residues that non-covalently orient glutathione for optimal nucleophilic attack of electrophiles in adjacent substrate binding sites.8 Glutathione is a tripeptide consisting of glutamine, cysteine, and glycine. Concentration of glutathione in eukaryotic cells typically ranges from 5–10 mM.41 The sulfhydryl from the cysteine residue of glutathione acts as a nucleophile in the attack of electrophilic xeno- and endo-biotic substrates.8 Changes in enrichment of individual GSTs with B[a]P dose, as measured by the glutathione derived activity-based probe, is likely due to either increased expression or could be due to some modification following translation of the enzyme that increases GST affinity for GSH (thus increasing probe binding). Some post-translational modifications of GSTs are known and include the phosphorylation of serines, tyrosines, and threonines of GSTs.37, 42 Phosphorylation of some GSTP1 residues are known to increase metabolic activity.43 Additionally, cysteine residues of GSTP1 are known to be glutathionylated.44 Conversely, these modifications are thought to result in decreases in activity. Distinction between expression-derived G site induction or induction via modification of expressed enzyme can be ascertained by comparing enzyme expression versus probe-determined G site activity (described below).
B[a]P induced changes in activity of GST H sites
B[a]P induced active H sites of several enzymes including GSTA2, GSTM1, GSTM2, GSTM4, GSTM6, and GSTP1. Induction with fold change over 2 was only observed in the highest 180 μmol/kg B[a]P dose. In addition, GSTA4 showed significant 2.31-fold downregulation at the lowest dose (2 μmol/kg), 3.83-fold downregulation at 20 μmol/kg, and 5.13-fold downregulation at 180 μmol/kg B[a]P (Figure 2, Table S1, Figure S3).
In comparison to the highly conserved G sites of individual GST isoenzymes, the substrate binding H sites are much more variable in sequence. The H sites of GSTs are generally electrophilic and/or hydrophobic and attract a wide range of substrates. The variability of GST H sites is thought to provide relative specificity of certain subtypes of xenobiotic substrates. For instance, the alpha and mu classes of human cytosolic GSTs contain a dominantly hydrophobic H site while the enzymes in the pi class contain a half hydrophobic, half hydrophilic H site.8 As the substrate specificity of GSTs comes from distinct and variable H sites, post-translational modification of GSTs could feasibly result in the tuning of H sites for increased affinity toward particular substrates. ABPP using GSTABP-H can be used to discover expression independent induction of active H sites by comparing changes in GSTABP-H enrichment with changes in expression (via global proteomics).
B[a]P induction of GSTs as measured by analysis of the global proteome
Induction of protein expression was measured via global proteomics of unenriched liver cytosolic lysate. Linear regression and fold change analysis of B[a]P induction revealed significant increases in GSTA2, GSTM4, and GSTP1 enzymes. GSTA2 was induced by 3.1-fold, GSTM4 by 2.4-fold, and GSTP1 by 2.7-fold at 180 μmol/kg (Figure 2, Table S1, Figure S3).
GST expression is known to be induced by Nrf2/Keap45, pregnane X receptor (PXR)46, and constitutive androstane receptor (CAR)47 mechanisms. Nrf2/Keap1 is primarily involved in response to oxidative stress. Keap1 attenuates Nrf2 activation via ubiquitination, leading to degradation of Nrf2. In oxidative stress conditions or in the presence of electrophiles, a conformational change occurs in which Keap1 is deactivated, allowing active Nrf2 translocation to the nucleus and subsequently bind the antioxidant response element and elicit an inductive response.48 PXR is a highly important receptor that aids in xenobiotic defense. This receptor is activated by a wide range of hydrophobic substances including toxins, bile acids, antibiotics, prescription drugs, and other chemicals.49 CAR is a receptor that regulates the expression of many genes involved in all three phases of xenobiotic metabolism including oxygenation, conjugation, and excretion. Like PXR, CAR binds and is activated by a wide variety of hydrophobic endo- and xeno-biotics.50 Because of the overlap between these pathways, it is possible that induction of GST expression following B[a]P exposure may be a result of a combination of direct binding of B[a]P and its downstream metabolites to these receptors as well as an oxidative stress response by Nrf2/Keap1.
Correlative analysis of induced changes in active GSTs versus GST protein expression
Following the determination of the specific GSTs induced (or inhibited) after exposure to B[a]P, we investigated the source of induction via comparison of induced or inhibited enzymes between datasets. Overall, we found that when comparing the B[a]P induction of the three proteomic datasets, three major groups of induction were revealed: (1) enzymes with B[a]P induced expression, active G sites, and active H sites, (2) enzymes with increases in active H sites with no expressional or G site induction, and (3) enzymes with reduced active G and H sites with no corresponding changes in expression.
GSTs with increased expression, G and H site activities
Enzymes whose induction was measured across all proteomic analyses include GSTA2, GSTM4, and GSTP1. All of these enzymes were found to show significantly induced expression (via global proteomics), and probe-determined G and H site activities based on fold change (≥2) and linear regression analysis (p < 0.05) (Figure 2, Figure 3A). For GSTP1, results of the Pearson analysis reveal significant (p < 0.05) correlation between proteomics results from each individual probe and global samples. GSTP1 has previously been found to glutathionylate PAH diol epoxides.13 Knocking out this enzyme also has been shown to result in increased PAH-induced skin tumorigenesis in mice.51 Thus, increased GSTP1 activity as well as expression induced by PAH exposure may result in increased metabolism and detoxification of PAHs. GSTM4 induction was correlated between all proteomics datasets (Figure 3A). GSTM4 has not been previously shown to be induced by PAHs; nor has M4 been implicated in the metabolism of P450-bioactivated PAH metabolites.
Figure 3.

Fold change and correlation analyses of GSTs for which significant induction occurred upon B[a]P exposure. For each GST enzyme, fold change induction relative to corn oil vehicle samples is shown (top) as well as results from Pearson Correlation analysis results including matrices of correlation coefficients (r) and p values for indicated comparisons. These correlation comparisons include global vs. GSTABP-G, global vs. GSTABP-H, and GSTABP-G vs. GSTABP-H. (A) Fold change and correlation analysis results of GST enzymes with significant induction as measured by global, GSTABP-G enriched, and GSTABP-H enriched proteomics samples (B) Fold change and correlation analysis results of GST enzymes with significant induction as measured by GSTABP-H enriched proteomics samples, but not global nor GSTABP-G enriched samples (C) Fold change and correlation analysis results of GSTA4, the sole GST in which B[a]P exposure resulted in changes in activity as measured by both probes, with no corresponding changes in expression as measured by global proteomics. Analyses are from 3 technical replicates from pooled cytosolic samples from 4 individually exposed mice. Error bars represent standard error of the mean.
Further studies are needed to determine the implications of induction of GSTM4 for PAH metabolism. A significant positive correlation between global and G site data of GSTA2 was observed (Figure 3A). GSTA2 is also previously known to show induced expression in rat hepatoma cells with PAH exposure,52 however little is known regarding its role in the metabolism and expulsion of bioactivated PAH metabolites. Further studies measuring PAH diol and diol epoxide metabolism should be conducted to determine its role. The similarity in overall inductive trends seen suggest that induction of active G and H sites is likely genetically derived and may constitute a receptor-mediated GST response.
GSTs with expression-independent increases activity unique to the H site
Three mu class GSTs (M1, M2, and M6) showed expression-independent increases in H site activity at 180 μmol/kg B[a]P dose. Interestingly, not only do these enzymes show no increase in protein expression, they also show no increase in G site labeling by GSTABP-G. Correlation analyses reveal no correlation between expression and probe-determined G and H site activities for GSTM1. While expression, active G sites, active H sites are all positively correlated for GSTM2, only probe-determined H site activity significantly increases with dose (from fold change and linear regression analysis). Similarly, while global and GSTABP-H GSTM6 data show a positive correlation, fold change and linear regression analyses indicate that this enzyme shows significant dose dependence solely in GSTABP-H enriched samples and not in global samples (Figure 3B). These expression- and G site-independent increases in active H sites may suggest a post-translational mechanism that does not affect the ability of glutathione to bind the enzymes, but only increases the activity of H sites. We speculate that this post-translational mechanism may cause a conformational change in relation to the catalytic tyrosine residues responsible for stabilizing the glutathione thiolate anion formed in GST active sites. Several GST mu isoenzymes are also known to metabolize PAH diol epoxides.53 In V79 cells, overexpression of GSTM1 resulted in a reduction of DNA adduct formation by dibenzo[a,l]pyrene and B[a]P diol epoxides.54 In BPDE exposed H1355 cells, DNA damage was reduced in cells with exogenously overexpressed GSTM2.55, 56 Due to known roles in the metabolism of the prototypical PAH diol epoxides and prevention of DNA adducts in mammalian cells, this observed GSTM induction may prove important as traditional mRNA and global proteomics would not detect these changes.
GSTA4 expression-independent inhibition of G and H site activities
The enzyme most affected by B[a]P exposure was GSTA4, with significant downregulation observed at the lowest dose tested. GSTABP-H enriched A4 showed over 2-fold downregulation at 2 μmol/kg B[a]P and roughly 4- and 5-fold downregulation at 20 and 180 μmol/kg doses. GSTABP-G enrichment of this enzyme only showed significant downregulation at the 180 μmol/kg dose, while no changes in protein expression were observed. Pearson correlation analysis results showed no significant correlation between any of the three datasets though data from both probes seem to follow a similar trend, albeit at higher doses for GSTABP-G (Figure 3C). This could be suggestive of an inhibitory mechanism or post translational modification. GSTA4 is primarily known to be involved in metabolism of 4-hydroxynonenals and products of lipid peroxidation.57 In addition, increased steady-state levels of aldehydes and decreased ability to conjugate 4-HNE have been observed in GSTA4 null mice.58 The observed reduction in active G and H sites of this enzyme in the liver may lead to an impairment of the liver to detoxify these lipid derived free radicals and could equate to increased oxidative stress.
Conclusions
Exposure to xenobiotics, including PAHs, are characterized by the response of biotransformation enzymes in target tissues. Interindividual differences in detoxifying enzyme family expression and activity is principal to individual susceptibilities that result in PAH-induced toxicities and disease pathology. Our understanding of the oxidative response to PAH exposure is fairly well characterized at the expression and activity levels, particularly in the liver. In contrast, there is little isoenzyme specific activity and expression data for PAH induction of GSTs in mammalian model systems. Thus, in combination with the near complete lack of proteomics data for PAH induction of GSTs and the lack of enzyme-specific activity assays, our investigation improves our understanding of the role of individual GSTs in catalyzing glutathione conjugation in the metabolism of B[a]P.
According to the results described herein, the induction of active G and H sites and expression is moderate. However, large fold change GST induction may not be necessary for the metabolism of the oxidized B[a]P metabolites. Cytochrome P450 enzymes not only catalyze the initial oxidation of PAHs, they also are known to participate in further oxygenation of their products. Products of both P450 monooxygenation and epoxide hydrolase activities result in a much more complex mixture of B[a]P metabolites for conjugating enzymes to detoxify and metabolize.5 GSTs are thought to primarily catalyze the glutathione conjugation of PAH epoxides, diol epoxides, and diones while UGTs and sulfotransferases are thought to primarily catalyze the conjugation of glucuronic acid to PAH diols, for example.5 Thus, GSTs and other conjugating enzymes are likely responsible for the metabolism of small enzyme-family specific subsets of these complex metabolite mixtures. For this reason, doubling or tripling their expression and/or activity, resulting in a matching increase of metabolic capacity, may be ample for their metabolic roles and result in biologically significant increases in detoxification.
Our study reveals B[a]P induced GST activity and provides insight into subsite specific induction of GST activity. Intriguing differences are observed when comparing glutathione-binding and active H sites of the GSTs in this study. At high doses of B[a]P, probe-determined H site activities of three GST mu enzymes show both expression and glutathione binding- independent increases. It is also conceivable that upon certain perturbations, glutathione-binding sites may also show expression-independent activity changes. These findings may generate hypotheses for further investigation into potential post-translational mechanisms of this site-specific induction of activity. In addition, further investigations as to the effect on the rate of GST metabolism of oxidized benzo[a]pyrene metabolites in induced animal models should be conducted in order to determine the role of induced GST enzyme activities in the detoxification of these carcinogens.
Supplementary Material
ACKNOWLEDGMENTS
This research was supported by the National Institutes of Health National Institute of Environmental Health Sciences (P42 ES016465), and employed proteomics capabilities supported by the NIH NIGMS Research Resource for Integrative Biology (P41 GM103493). A portion of the research was performed using EMSL, a DOE Office of Science User Facility sponsored by the Office of Biological and Environmental Research. PNNL is a multiprogram laboratory operated by Battelle for US DOE Contract DE-AC06-76RL01830.
ABBREVIATIONS
- ABPP
activity-based protein profiling
- ABPs
activity-based probes
- Ahr
aryl hydrocarbon receptor
- AMT
accurate mass and time
- B[a]P
benzo[a]pyrene
- BCA
bicinchoninic assay
- BPDE
benzo[a]pyrene diol epoxide
- CAR
constitutive androstane receptor
- DMSO
dimethylsulfoxide
- DNCB 2
4-dinitrochlorobenzene
- GSH
glutathione
- GST
glutathione S-transferase
- LC-MS
liquid chromatography – mass spectrometry
- MT FDR
mass and time false discovery rate
- Nrf2
nuclear factor-like 2
- PAH
polycyclic aromatic hydrocarbon
- PBS
phosphate buffered saline
- PXR
pregnane X receptor
- SDS-PAGE
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
Footnotes
The authors declare no competing financial interests. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE59 partner repository with the dataset identifier PXD010610 and 10.6019/PXD010610.
ASSOCIATED CONTENT
Supporting Information. This material is available free of charge via the Internet at http://pubs.acs.org.”
Table S1. Results of linear regression analyses (.xlsx)
Figure S1. Coomassie total protein stain of GSTABP-G labeling of microsomes from B[a]P treated mice (.tif)
Figure S2. Coomassie total protein stain of GSTABP-H labeling of microsomes from B[a]P treated mice (.tif)
Figure S3. Curve fits from linear regression analyses of proteomics data (.pdf)Linear regression curves were fit plotting B[a]P dose (on log2 scaled x-axis) and relative abundance values (plotted on log10 scaled y-axis). Blue represents data from GSTABP-H enriched proteomics samples. Green represents data from GSTABP-G enriched proteomics samples. Red represents data from global proteomics samples.
REFERENCES
- 1.Srogi K, Monitoring of environmental exposure to polycyclic aromatic hydrocarbons: a review. Environmental Chemistry Letters 2007, 5 (4), 169–195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zhong Y; Carmella SG; Upadhyaya P; Hochalter JB; Rauch D; Oliver A; Jensen J; Hatsukami D; Wang J; Zimmerman C; Hecht SS, Immediate Consequences of Cigarette Smoking: Rapid Formation of Polycyclic Aromatic Hydrocarbon Diol Epoxides. Chemical Research in Toxicology 2011, 24 (2), 246–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lin P; Aiona PK; Li Y; Shiraiwa M; Laskin J; Nizkorodov SA; Laskin A, Molecular Characterization of Brown Carbon in Biomass Burning Aerosol Particles. Environmental Science & Technology 2016, 50 (21), 11815–11824. [DOI] [PubMed] [Google Scholar]
- 4.Dipple A; Anita C; Bigger H, Mechanism of action of food-associated polycyclic aromatic hydrocarbon carcinogens. Mutation Research/Genetic Toxicology 1991, 259 (3), 263–276. [DOI] [PubMed] [Google Scholar]
- 5.Shimada T, Xenobiotic-metabolizing enzymes involved in activation and detoxification of carcinogenic polycyclic aromatic hydrocarbons. Drug metabolism and pharmacokinetics 2006, 21 (4), 257–76. [DOI] [PubMed] [Google Scholar]
- 6.Šulc M; Indra R; Moserová M; Schmeiser HH; Frei E; Arlt VM; Stiborová M; White P, The impact of individual cytochrome P450 enzymes on oxidative metabolism of benzo[a]pyrene in human livers. Environmental and Molecular Mutagenesis 2016, 57 (3), 229–235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Shimada T; Sugie A; Yamada T; Kawazoe H; Hashimoto M; Azuma E; Nakajima T; Inoue K; Oda Y, Dose-response studies on the induction of liver cytochromes P4501A1 and 1B1 by polycyclic aromatic hydrocarbons in arylhydrocarbon-responsive C57BL/6J mice. Xenobiotica; the fate of foreign compounds in biological systems 2003, 33 (9), 957–71. [DOI] [PubMed] [Google Scholar]
- 8.Wu B; Dong D, Human cytosolic glutathione transferases: structure, function, and drug discovery. Trends in pharmacological sciences 2012, 33 (12), 656–68. [DOI] [PubMed] [Google Scholar]
- 9.Awasthi YC; Singh SV; Das M; Mukhtar H, Differential activities of rat and human lung glutathione S-transferase isoenzymes towards benzo(a)pyrene epoxides. Biochemical and Biophysical Research Communications 1985, 133 (3), 863–867. [DOI] [PubMed] [Google Scholar]
- 10.Srivastava SK; Hu X; Xia H; Awasthi S; Amin S; Singh SV, Metabolic Fate of Glutathione Conjugate of Benzo[a]pyrene-(7R,8S)-diol (9S,10R)-epoxide in Human Liver. Archives of Biochemistry and Biophysics 1999, 371 (2), 340–344. [DOI] [PubMed] [Google Scholar]
- 11.Eaton DL; Stapleton PL, Simultaneous determination of cytosolic glutathione S-transferase and microsomal epoxide hydrolase activity toward benzo[a]pyrene-4,5-oxide by high-performance liquid chromatography. Analytical Biochemistry 1989, 178 (1), 153–158. [DOI] [PubMed] [Google Scholar]
- 12.Hernandez O; Walker M; Cox RH; Foureman GL; Smith BR; Bend JR, Regiospecificity and stereospecificity in the enzymatic conjugation of glutathione with (±)-benzo(a)pyrene 4,5-oxide. Biochemical and Biophysical Research Communications 1980, 96 (4), 1494–1502. [DOI] [PubMed] [Google Scholar]
- 13.Sundberg K; Johansson AS; Stenberg G; Widersten M; Seidel A; Mannervik B; Jernstrom B, Differences in the catalytic efficiencies of allelic variants of glutathione transferase P1–1 towards carcinogenic diol epoxides of polycyclic aromatic hydrocarbons. Carcinogenesis 1998, 19 (3), 433–6. [DOI] [PubMed] [Google Scholar]
- 14.Dreij K; Sundberg K; Johansson AS; Nordling E; Seidel A; Persson B; Mannervik B; Jernstrom B, Catalytic activities of human alpha class glutathione transferases toward carcinogenic dibenzo[a,l]pyrene diol epoxides. Chemical research in toxicology 2002, 15 (6), 825–31. [DOI] [PubMed] [Google Scholar]
- 15.Sundberg K; Widersten M; Seidel A; Mannervik B; Jernström B, Glutathione Conjugation of Bay- and Fjord-Region Diol Epoxides of Polycyclic Aromatic Hydrocarbons by Glutathione Transferases M1–1 and P1–1. Chemical research in toxicology 1997, 10 (11), 1221–1227. [DOI] [PubMed] [Google Scholar]
- 16.Knight TR; Choudhuri S; Klaassen CD, Induction of Hepatic Glutathione S-Transferases in Male Mice by Prototypes of Various Classes of Microsomal Enzyme Inducers. Toxicological Sciences 2008, 106 (2), 329–338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pushparajah DS; Umachandran M; Plant KE; Plant N; Ioannides C, Up-regulation of the glutathione S-transferase system in human liver by polycyclic aromatic hydrocarbons; comparison with rat liver and lung. Mutagenesis 2008, 23 (4), 299–308. [DOI] [PubMed] [Google Scholar]
- 18.McLellan LI; Kerr LA; Cronshaw AD; Hayes JD, Regulation of mouse glutathione S-transferases by chemoprotectors. Molecular evidence for the existence of three distinct Alpha-class glutathione S-transferase subunits, Ya1, Ya2, and Ya3, in mouse liver. Biochemical Journal 1991, 276 (2), 461–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Vos RM; Snoek MC; van Berkel WJ; Muller F; van Bladeren PJ, Differential induction of rat hepatic glutathione S-transferase isoenzymes by hexachlorobenzene and benzyl isothiocyanate. Comparison with induction by phenobarbital and 3-methylcholanthrene. Biochemical pharmacology 1988, 37 (6), 1077–82. [DOI] [PubMed] [Google Scholar]
- 20.Banger KK; Lock EA; Reed CJ, The characterization of glutathione S-transferases from rat olfactory epithelium. The Biochemical journal 1993, 290 ( Pt 1) (Pt 1), 199–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Meyer DJ; Coles B; Pemble SE; Gilmore KS; Fraser GM; Ketterer B, Theta, a new class of glutathione transferases purified from rat and man. Biochemical Journal 1991, 274 (2), 409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cmarik JL; Inskeep PB; Meredith MJ; Meyer DJ; Ketterer B; Guengerich FP, Selectivity of rat and human glutathione S-transferases in activation of ethylene dibromide by glutathione conjugation and DNA binding and induction of unscheduled DNA synthesis in human hepatocytes. Cancer research 1990, 50 (9), 2747–52. [PubMed] [Google Scholar]
- 23.Hayes JD; Pulford DJ, The Glut athione S-Transferase Supergene Family: Regulation of GST and the Contribution of the lsoenzymes to Cancer Chemoprotection and Drug Resistance Part I. Critical Reviews in Biochemistry and Molecular Biology 1995, 30 (6), 445–520. [DOI] [PubMed] [Google Scholar]
- 24.Kushman ME; Kabler SL; Fleming MH; Ravoori S; Gupta RC; Doehmer J; Morrow CS; Townsend AJ, Expression of human glutathione S-transferase P1 confers resistance to benzo[a]pyrene or benzo[a]pyrene-7,8-dihydrodiol mutagenesis, macromolecular alkylation and formation of stable N2-Gua-BPDE adducts in stably transfected V79MZ cells co-expressing hCYP1A1. Carcinogenesis 2007, 28 (1), 207–14. [DOI] [PubMed] [Google Scholar]
- 25.Naspinski C; Gu X; Zhou GD; Mertens-Talcott SU; Donnelly KC; Tian Y, Pregnane X receptor protects HepG2 cells from BaP-induced DNA damage. Toxicological sciences : an official journal of the Society of Toxicology 2008, 104 (1), 67–73. [DOI] [PubMed] [Google Scholar]
- 26.Gelhaus SL; Harvey RG; Penning TM; Blair IA, Regulation of benzo[a]pyrene-mediated DNA- and glutathione-adduct formation by 2,3,7,8-tetrachlorodibenzo-p-dioxin in human lung cells. Chemical research in toxicology 2011, 24 (1), 89–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cravatt BF; Wright AT; Kozarich JW, Activity-Based Protein Profiling: From Enzyme Chemistry to Proteomic Chemistry. Annual Review of Biochemistry 2008, 77 (1), 383–414. [DOI] [PubMed] [Google Scholar]
- 28.Stoddard EG; Killinger BJ; Nair RN; Sadler NC; Volk RF; Purvine SO; Shukla AK; Smith JN; Wright AT, Activity-Based Probes for Isoenzyme- and Site-Specific Functional Characterization of Glutathione S-Transferases. Journal of the American Chemical Society 2017, 139 (45), 16032–16035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Speers AE; Cravatt BF, Activity-Based Protein Profiling (ABPP) and Click Chemistry (CC)-ABPP by MudPIT Mass Spectrometry. Current protocols in chemical biology 2009, 1, 29–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Smith JN; Mehinagic D; Nag S; Crowell SR; Corley RA, In vitro metabolism of benzo[a]pyrene-7,8-dihydrodiol and dibenzo[def,p]chrysene-11,12 diol in rodent and human hepatic microsomes. Toxicology letters 2017, 269, 23–32. [DOI] [PubMed] [Google Scholar]
- 31.Crowell SR; Sharma AK; Amin S; Soelberg JJ; Sadler NC; Wright AT; Baird WM; Williams DE; Corley RA, Impact of Pregnancy on the Pharmacokinetics of Dibenzo[def,p]chrysene in Mice. Toxicological Sciences 2013, 135 (1), 48–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Crowell SR; Amin SG; Anderson KA; Krishnegowda G; Sharma AK; Soelberg JJ; Williams DE; Corley RA, Preliminary physiologically based pharmacokinetic models for benzo[a]pyrene and dibenzo[def,p]chrysene in rodents. Toxicology and Applied Pharmacology 2011, 257 (3), 365–376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Crowell SR; Hanson-Drury S; Williams DE; Corley RA, In vitro metabolism of benzo[a]pyrene and dibenzo[def,p]chrysene in rodent and human hepatic microsomes. Toxicology letters 2014, 228 (1), 48–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Shorey LE; Castro DJ; Baird WM; Siddens LK; Löhr CV; Matzke MM; Waters KM; Corley RA; Williams DE, Transplacental carcinogenesis with dibenzo[def,p]chrysene (DBC): Timing of maternal exposures determines target tissue response in offspring. Cancer Letters 2012, 317 (1), 49–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Walker JM, The Bicinchoninic Acid (BCA) Assay for Protein Quantitation In Basic Protein and Peptide Protocols, Walker JM, Ed. Humana Press: Totowa, NJ, 1994; pp 5–8. [DOI] [PubMed] [Google Scholar]
- 36.Sadler NC; Bernstein HC; Melnicki MR; Charania MA; Hill EA; Anderson LN; Monroe ME; Smith RD; Beliaev AS; Wright AT, Dinitrogenase-Driven Photobiological Hydrogen Production Combats Oxidative Stress in Cyanothece sp. Strain ATCC 51142. Applied and environmental microbiology 2016, 82 (24), 7227–7235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chen C; Huang H; Wu CH, Protein Bioinformatics Databases and Resources. Methods in molecular biology (Clifton, N.J.) 2017, 1558, 3–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kim S; Pevzner PA, MS-GF+ makes progress towards a universal database search tool for proteomics. Nature Communications 2014, 5, 5277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zimmer JSD; Monroe ME; Qian W-J; Smith RD, Advances in Proteomics Data Analysis and Display Using an Accurate Mass and Time Tag Approach. Mass spectrometry reviews 2006, 25 (3), 450–482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Polpitiya AD; Qian W-J; Jaitly N; Petyuk VA; Adkins JN; Camp DG; Anderson GA; Smith RD, DAnTE: a statistical tool for quantitative analysis of -omics data. Bioinformatics (Oxford, England) 2008, 24 (13), 1556–1558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wu G; Fang Y-Z; Yang S; Lupton JR; Turner ND, Glutathione Metabolism and Its Implications for Health. The Journal of Nutrition 2004, 134 (3), 489–492. [DOI] [PubMed] [Google Scholar]
- 42.Huttlin EL; Jedrychowski MP; Elias JE; Goswami T; Rad R; Beausoleil SA; Villen J; Haas W; Sowa ME; Gygi SP, A tissue-specific atlas of mouse protein phosphorylation and expression. Cell 2010, 143 (7), 1174–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lo HW; Antoun GR; Ali-Osman F, The human glutathione S-transferase P1 protein is phosphorylated and its metabolic function enhanced by the Ser/Thr protein kinases, cAMP-dependent protein kinase and protein kinase C, in glioblastoma cells. Cancer research 2004, 64 (24), 9131–8. [DOI] [PubMed] [Google Scholar]
- 44.Townsend DM; Manevich Y; He L; Hutchens S; Pazoles CJ; Tew KD, Novel role for glutathione S-transferase pi. Regulator of protein S-Glutathionylation following oxidative and nitrosative stress. The Journal of biological chemistry 2009, 284 (1), 436–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kwak MK; Itoh K; Yamamoto M; Sutter TR; Kensler TW, Role of transcription factor Nrf2 in the induction of hepatic phase 2 and antioxidative enzymes in vivo by the cancer chemoprotective agent, 3H-1, 2-dimethiole-3-thione. Molecular Medicine 2001, 7 (2), 135–145. [PMC free article] [PubMed] [Google Scholar]
- 46.Gong H; Singh SV; Singh SP; Mu Y; Lee JH; Saini SP; Toma D; Ren S; Kagan VE; Day BW; Zimniak P; Xie W, Orphan nuclear receptor pregnane X receptor sensitizes oxidative stress responses in transgenic mice and cancerous cells. Molecular endocrinology (Baltimore, Md.) 2006, 20 (2), 279–90. [DOI] [PubMed] [Google Scholar]
- 47.Maglich JM; Stoltz CM; Goodwin B; Hawkins-Brown D; Moore JT; Kliewer SA, Nuclear pregnane x receptor and constitutive androstane receptor regulate overlapping but distinct sets of genes involved in xenobiotic detoxification. Mol Pharmacol 2002, 62 (3), 638–46. [DOI] [PubMed] [Google Scholar]
- 48.Fan Z; Wirth AK; Chen D; Wruck CJ; Rauh M; Buchfelder M; Savaskan N, Nrf2-Keap1 pathway promotes cell proliferation and diminishes ferroptosis. Oncogenesis 2017, 6, e371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kliewer SA; Goodwin B; Willson TM, The nuclear pregnane X receptor: a key regulator of xenobiotic metabolism. Endocrine reviews 2002, 23 (5), 687–702. [DOI] [PubMed] [Google Scholar]
- 50.di Masi A; Marinis ED; Ascenzi P; Marino M, Nuclear receptors CAR and PXR: Molecular, functional, and biomedical aspects. Molecular Aspects of Medicine 2009, 30 (5), 297–343. [DOI] [PubMed] [Google Scholar]
- 51.Henderson CJ; Smith AG; Ure J; Brown K; Bacon EJ; Wolf CR, Increased skin tumorigenesis in mice lacking pi class glutathione S-transferases. Proceedings of the National Academy of Sciences of the United States of America 1998, 95 (9), 5275–5280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Kaisarevic S; Dakic V; Hrubik J; Glisic B; Lubcke-von Varel U; Pogrmic-Majkic K; Fa S; Teodorovic I; Brack W; Kovacevic R, Differential expression of CYP1A1 and CYP1A2 genes in H4IIE rat hepatoma cells exposed to TCDD and PAHs. Environmental toxicology and pharmacology 2015, 39 (1), 358–68. [DOI] [PubMed] [Google Scholar]
- 53.Funk M; Gath I; Seidel A; Oesch F; Platt KL, Conjugation of anti-dihydrodiol epoxides of benzo[a]pyrene, chrysene, benzo[c]phenanthrene and dibenz[a,h]anthracene with glutathione catalyzed by cytosol and by the Mu-class glutathione transferase HTP II from rat liver. Chemico-Biological Interactions 1995, 95 (1), 189–201. [DOI] [PubMed] [Google Scholar]
- 54.Sundberg K; Dreij K; Seidel A; Jernstrom B, Glutathione conjugation and DNA adduct formation of dibenzo[a,l]pyrene and benzo[a]pyrene diol epoxides in V79 cells stably expressing different human glutathione transferases. Chem Res Toxicol 2002, 15 (2), 170–9. [DOI] [PubMed] [Google Scholar]
- 55.Weng MW; Hsiao YM; Chiou HL; Yang SF; Hsieh YS; Cheng YW; Yang CH; Ko JL, Alleviation of benzo[a]pyrene-diolepoxide-DNA damage in human lung carcinoma by glutathione S-transferase M2. DNA repair 2005, 4 (4), 493–502. [DOI] [PubMed] [Google Scholar]
- 56.Tang SC; Sheu GT; Wong RH; Huang CY; Weng MW; Lee LW; Hsu CP; Ko JL, Expression of glutathione S-transferase M2 in stage I/II non-small cell lung cancer and alleviation of DNA damage exposure to benzo[a]pyrene. Toxicology letters 2010, 192 (3), 316–23. [DOI] [PubMed] [Google Scholar]
- 57.Hubatsch I; Ridderström M; Mannervik B, Human glutathione transferase A4–4: an alpha class enzyme with high catalytic efficiency in the conjugation of 4-hydroxynonenal and other genotoxic products of lipid peroxidation. Biochemical Journal 1998, 330 (Pt 1), 175–179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Engle MR; Singh SP; Czernik PJ; Gaddy D; Montague DC; Ceci JD; Yang Y; Awasthi S; Awasthi YC; Zimniak P, Physiological role of mGSTA4–4, a glutathione S-transferase metabolizing 4-hydroxynonenal: generation and analysis of mGsta4 null mouse. Toxicol Appl Pharmacol 2004, 194 (3), 296–308. [DOI] [PubMed] [Google Scholar]
- 59.Vizcaino JA; Csordas A; del-Toro N; Dianes JA; Griss J; Lavidas I; Mayer G; Perez-Riverol Y; Reisinger F; Ternent T; Xu QW; Wang R; Hermjakob H, 2016 update of the PRIDE database and its related tools. Nucleic acids research 2016, 44 (D1), D447–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.