

Abstract
Background
Ixodes ricinus constitutes the main European vector tick for the Lyme borreliosis pathogen Borrelia burgdorferi (sensu lato), the relapsing fever borrelia Borrelia miyamotoi, as well as Anaplasma phagocytophilum and several Rickettsia species. Under laboratory conditions, a transovarial transmission to the next tick generation is described for Rickettsia spp. and Borrelia spp., especially regarding B. miyamotoi, whereas the efficiency of transovarial transfer under field conditions is largely unstudied.
Methods
In order to better estimate the potential infection risk by tick larvae for humans and animals, 1500 I. ricinus larvae from 50 collected “nests” (larvae adhering to the flag in a clumped manner) were individually examined for Borrelia, Rickettsia and A. phagocytophilum DNA using quantitative real-time PCR (qPCR).
Results
Thirty-nine of 50 nests each (78.0%, 95% CI: 64.0–88.5%) were positive for Borrelia spp. and Rickettsia spp. DNA, and in three nests (6.0%, 95% CI: 1.3–16.5%) A. phagocytophilum DNA was detected. Overall, DNA from at least one pathogen could be detected in 90.0% (45/50, 95% CI: 78.2–96.7%) of the nests. Of the 1500 larvae, 137 were positive for Borrelia spp. DNA (9.1%, 95% CI: 7.7–10.7%), 341 for Rickettsia spp. DNA (22.7%, 95% CI: 20.6–24.9%) and three for A. phagocytophilum DNA (0.2%, 95% CI: 0–0.6%). Quantity of Borrelia spp. and Anaplasma spp. DNA in positive larvae was low, with 2.7 × 100Borrelia 5S-23S gene copies and 2.4 × 101A. phagocytophilum msp2/p44 gene copies detected on average, while Rickettsia-positive samples contained on average 5.4 × 102gltA gene copies. Coinfections were found in 66.0% (33/50, 95% CI: 51.2–78.8%) of the nests and 8.6% (38/443, 95% CI: 6.1–11.6%) of positive larvae. In fact, larvae had a significantly higher probability of being infected with Borrelia spp. or Rickettsia spp. when both pathogens were present in the nest.
Conclusions
This study provides evidence for transovarial transmission of Rickettsia spp. and Borrelia spp. in I. ricinus under field conditions, possibly facilitating pathogen persistence in the ecosystem and reducing the dependence on the presence of suitable reservoir hosts. Further studies are needed to prove transovarial transmission and to explain the surprisingly high proportion of nests containing Rickettsia and/or Borrelia DNA-positive larvae compared to infection rates in adult ticks commonly reported in other studies.
Keywords: Borrelia burgdorferi (sensu lato), Borrelia miyamotoi, Rickettsiales, Anaplasma phagocytophilum, Ticks, Tick-borne diseases
Background
Ixodes ricinus is the most widespread tick species in Europe and acts as a vector for a range of bacterial and viral tick-borne pathogens with relevance for human and animal health. In Germany, different genospecies of the B. burgdorferi (sensu lato) complex, B. miyamotoi, A. phagocytophilum and Rickettsia spp. are among the most important bacterial pathogens transmitted by I. ricinus. These pathogens cause various diseases in humans and animals. Borrelia burgdorferi (s.l.) is the causative agent of Lyme borreliosis (LB), while B. miyamotoi causes febrile illness and has been associated with meningoencephalitis in immunocompromised patients in Europe [1, 2]. Furthermore, several Rickettsia spp. may cause spotted fever or lymphadenopathy in humans [3]. In I. ricinus, R. helvetica, R. monacensis, R. massiliae and R. felis have been detected, with R. helvetica being the most frequent [3–7]. Another member of the order Rickettsiales, A. phagocytophilum, may cause granulocytic anaplasmosis in humans, dogs, horses, goats, sheep and cattle [8–10].
Transmission of these pathogens between or in ticks may occur in various ways, including co-feeding, transstadial, sexual as well as transovarial transmission. Co-feeding transmission, whereby infection spreads from one tick to another feeding in close proximity on the host, seems to be very rare [11–13]. Transstadial transmission, i.e. the transmission of pathogens from one developmental stage to the next, occurs in Borrelia spp. as well as Rickettsia spp. and A. phagocytophilum [14–17]. In contrast, sexual transmission, i.e. transmission from a male to a female tick during copulation, has only been described for Rickettsia species as well as some relapsing-fever borreliae [18, 19]. During transovarial transmission, the offspring of an infected female is affected. An efficient transovarial transmission rate of up to 100% has been shown for Rickettsia spp. under laboratory conditions [17, 20], whereas efficiency is generally believed to be much lower for Borrelia burgdorferi (s.l.), which are mainly transmitted to larvae via a blood meal from an infected host or rarely by co-feeding [21–23]. Borrelia miyamotoi, on the other hand, can be transmitted transovarially from the female tick to more than 90.0% of its larvae [22, 24]. In contrast, a low to inefficient transovarial transmission rate has been described for A. phagocytophilum [25–27]. Transovarial transmission facilitates the persistence of pathogens in the ecosystem and thus reduces dependence on suitable reservoir hosts.
The main hosts of I. ricinus larvae are rodents [28], but humans may also serve as accidental hosts [29]. Many prevalence studies have examined pathogen prevalence in nymphs and adult ticks, but only a few have examined tick larvae. Therefore, the aim of this study was to determine Borrelia spp., Rickettsia spp. and A. phagocytophilum DNA in questing I. ricinus larvae to extrapolate potential transovarial transmission rates of these pathogens under natural conditions and assess the possible human or animal infection risk due to I. ricinus larvae.
Methods
Tick larvae collection and molecular species identification
Questing tick larvae were collected by the flagging method at different sampling sites in northern Germany during 2010–2018. The flag was dragged approximately one meter over the ground, and all larvae adhering to the flag in a clumped manner were defined as a “nest” originating from a single female (Fig. 1). After freezing the flags with the collected larvae overnight at − 20 °C, larvae were picked off the flag and stored individually at − 20 °C until genomic DNA isolation.
Fig. 1.

Representative examples for larvae adhering to the flag in a clumped manner, which were defined as an Ixodes spp. nest
Tick species identification was performed on genomic DNA of two larvae per nest by amplification and sequencing of a part of the 16S rRNA gene, using primers described by Mangold et al. [30]. The reaction mixture and thermoprofile were set up as described by Hauck et al. [31], except that the amount of DNA template was increased to 6 μl and the number of PCR cycles to 41. Amplicons were separated by electrophoresis on 1.5% agarose gels stained with GelRed® (Biotium Inc., Fremont, CA, USA) and visualized under UV light. Obtained PCR products were Sanger-sequenced at Microsynth Seqlab Laboratories (Göttingen, Germany) and aligned with 16S rRNA gene sequences published by Estrada-Peña et al. [32] [GenBank: KM211785, KM211786, KM211787, KM211788 (I. ricinus); KM211789, KM211790 (I. inopinatus)] as well as other selected sequences of I. ricinus, I. inopinatus and I. frontalis available on GenBank [accession nos. GU074592, GU074605 (I. ricinus); KY569415, KY569416, KY569417, KY569418 (I. inopinatus), MF688050 (I. frontalis)] using Clone Manager 9 Professional Edition (Scientific & Educational Software, Denver, USA).
Tick larvae testing for bacterial pathogen DNA
In total, 1500 larvae originating from 50 larvae nests were examined for Borrelia spp., Rickettsia spp. and A. phagocytophilum. Per nest, 10–40 larvae (average 30 larvae per nest, standard deviation, SD: 11.6) were examined, depending on nest size. In case of a small nest (≤ 40 larvae) all larvae were examined, whereas for larger nests 40 larvae were tested. Larvae were individually homogenized using 0.5 ml polysterene pistils (VWR, Darmstadt, Germany). Genomic DNA was extracted from individual tick larvae using the Nucleo Spin® 8 Blood Core Kit (Macherey-Nagel, Düren, Germany) according to the manufacturer’s specifications, with previously described amendments [5]. Until further use, isolated genomic DNA was stored at − 20 °C.
Testing for Borrelia spp. (B. burgdorferi (s.l.) and B. miyamotoi) and Rickettsia spp. by duplex quantitative real-time PCR (qPCR) was carried out as described previously [33, 34]. For Borrelia spp., the 5S–23S rRNA intergenic spacer (IGS) region was targeted based on a primer-TaqMan™ minor groove binder (MGB) probe combination designed by Strube et al. [35]. For detection of Rickettsia spp., the citrate synthase (gltA) gene was amplified based on a primer-TaqMan™ probe combination by Stenos et al. [36]. The reaction set-up and thermal cycling were performed as described previously [5]. Regarding A. phagocytophilum, larvae nests from 2010 and 2011 were tested by targeting the 16S rRNA gene using a primer-TaqMan™ probe combination by Sirigireddy and Ganta [37]. At the same time, successful DNA isolation was confirmed by duplex amplification of the Ixodes ITS2 region as previously described [5]. For larvae nests collected from 2015 to 2018, the msp2/p44 gene with a primer-TaqMan™ probe combination by Courtney et al. [38] was targeted. Again, successful DNA isolation was confirmed by simultaneous amplification of the Ixodes ITS2 region. The duplex reaction set-up and thermal cycling were carried out according to Blazejak et al. [7].
Borrelia (geno-)species and Rickettsia species identification
To determine the (geno-)species of Borrelia-positive tick larvae, the Reverse Line Blot (RLB) technique was performed for larvae nests from 2010/2011 by amplifying a fragment of the B. burgdorferi (s.l.) 5S-23S rRNA IGS region using biotin-linked forward primer 5SCB and reverse primer 23SN2 as published by Tappe et al. [39]. For larvae collected during 2015–2018, the biotin-linked forward primer B5S was used instead of 5SCB, and a second, hydrolase-23S rRNA region specific biotin-linked forward primer was added for specific amplification of B. miyamotoi as described by Blazejak et al. [40]. Moreover, the RLB setup was modified to include the BisNE1 probe as described by Springer et al. [29].
For Rickettsia species identification, a subset of the Rickettsia-positive samples was subjected to real-time pyrosequencing of a sequence stretch of the rickettsial citrate synthase (gltA) gene as previously described [41].
Statistical analyses
Statistical analyses were conducted in R v. 3.3.1 [42]. To assess factors influencing larval prevalence of Borrelia spp. and Rickettsia spp., generalized linear mixed effect models (GLMMs) with binomial error structure were constructed. Anaplasma phagocytophilum prevalence was not statistically analysed because of the low number of positive larvae. The fixed factors were “location/year” (Hanover 2010, 2015, 2017, 2018, Mellendorf 2018 and Hamburg 2011) and “coinfection of the nest” (yes/no). Due to the small number of nests originating from some locations, only nests from Hanover, Mellendorf and Hamburg were included (n = 1220 larvae from 43 nests). Furthermore, “nest ID” was included as a random effects term. Each model was compared to a null model including only the random effects term in a likelihood ratio test (R function ‘anova’, test = ‘chisq’).
Finally, the predictive variable “location/year” was subjected to post-hoc analysis, computing all pairwise differences between factor levels in a Tukey’s test based on the parameters of the fitted GLMM.
Results
Tick larvae collection and molecular species identification
Total nest size of the 50 collected nests varied between 10 and 1643 larvae. Most nests were collected in Hanover (20/50), Hamburg (12/50) and Mellendorf (11/50). Detailed numbers of larvae per nest including collection sites and dates are presented in Table 1. Molecular species identification revealed that all collected nests belonged to I. ricinus.
Table 1.
Detection of pathogenic microorganisms in I. ricinus larvae nests
| Nest | Location (Sampling site) | Sampling month | No. of examined larvae (total number of larvae) |
Borrelia spp. n/N (%)a |
Rickettsia spp. n/N (%)a |
A. phagocytophilum n/N (%)a |
Coinfection Borrelia spp. + Rickettsia spp. n/N (%) |
Coinfection Rickettsia spp. + A. phagocytophilum n/N (%) |
|---|---|---|---|---|---|---|---|---|
| 1 | Hanover (Misburger Wald) | May 2010 | 20 (20) | 3/20 (15.0) | 1/20 (5.0) | 0/20 (0) | 1/20 (5.0) | 0/20 (0) |
| 2 | Hanover (Bornumer Holz) | Jun 2010 | 28 (28) | 2/28 (7.1) | 10/28 (35.7) | 0/28 (0) | 0/28 (0) | 0/28 (0) |
| 3 | Hanover (Seelhorster Wald) | Jun 2010 | 34 (34) | 2/34 (5.9) | 0/34 (0) | 0/34 (0) | 0/34 (0) | 0/34 (0) |
| 4 | Hanover (Mecklenheide) | Jun 2010 | 12 (12) | 0/12 (0) | 0/12 (0) | 0/12 (0) | 0/12 (0) | 0/12 (0) |
| 5 | Hanover (Misburger Wald) | Aug 2010 | 11 (11) | 0/11 (0) | 0/11 (0) | 0/11 (0) | 0/11 (0) | 0/11 (0) |
| 6 | Hanover (Georgengarten) | Aug 2010 | 10 (10) | 0/10 (0) | 0/10 (0) | 0/10 (0) | 0/10 (0) | 0/10 (0) |
| 7 | Hanover (Alte Heide) | Sep 2010 | 28 (28) | 1/28 (3.6) | 1/28 (3.6) | 0/28 (0) | 0/28 (0) | 0/28 (0) |
| 8 | Hamburg (Schwarzberg) | May 2011 | 15 (15) | 1/15 (6.7) | 1/15 (6.7) | 0/15 (0) | 0/15 (0) | 0/15 (0) |
| 9 | Hamburg (Oejendorfer Park) | May 2011 | 20 (20) | 9/20 (45.0) | 5/20 (25.0) | 0/20 (0) | 2/20 (10.0) | 0/20 (0) |
| 10 | Hamburg (Raakmoor) | Jun 2011 | 18 (18) | 5/18 (27.8) | 1/18 (5.6) | 0/18 (0) | 0/18 (0) | 0/18 (0) |
| 11 | Hamburg (Neugrabener Heide) | Jun 2011 | 16 (16) | 0/16 (0) | 0/16 (0) | 0/16 (0) | 0/16 (0) | 0/16 (0) |
| 12 | Hamburg (Schwarzenberg) | Jun 2011 | 13 (13) | 2/13 (15.4) | 0/13 (0) | 0/13 (0 | 0/13 (0) | 0/13 (0) |
| 13 | Hamburg (Gosselers Park) | Jun 2011 | 20 (20) | 7/20 (35.0) | 1/20 (5.0) | 0/20 (0) | 1/20 (5.0) | 0/20 (0) |
| 14 | Hamburg (Stadtpark Winterhude) | Jun 2011 | 19 (19) | 1/19 (5.3) | 15/19 (78.9) | 0/19 (0) | 1/19 (5.3) | 0/19 (0) |
| 15 | Hamburg (Oejendorfer Park) | Jun 2011 | 19 (19) | 1/19 (5.3) | 6/19 (31.6) | 0/19 (0) | 0/19 (0) | 0/19 (0) |
| 16 | Hamburg (Gosselers Park) | Jul 2011 | 16 (16) | 1/16 (6.3) | 1/16 (6.3) | 0/16 (0) | 1/16 (6.3) | 0/16 (0) |
| 17 | Hamburg (Altonaer Volkspark) | Jul 2011 | 15 (15) | 1/15 (6.7) | 1/15 (6.7) | 0/15 (0) | 0/15 (0) | 0/15 (0) |
| 18 | Hamburg (Alster) | Jul 2011 | 16 (16) | 3/16 (18.8) | 7/16 (43.8) | 0/16 (0) | 0/16 (0) | 0/16 (0) |
| 19 | Hamburg (Stadtpark Winterhude) | Jul 2011 | 20 (20) | 2/20 (10.0) | 13/20 (65.0) | 0/20 (0) | 1/20 (5.0) | 0/20 (0) |
| 20 | Hanover (Bothfeld) | May 2015 | 12 (12) | 2/12 (16.7) | 2/12 (16.7) | 1/12 (8.3) | 1/12 (8.3) | 0/12 (0) |
| 21 | Hanover (Bornumer Holz) | May 2015 | 40 (355) | 0/40 (0) | 24/40 (60.0) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 22 | Hanover (Mecklenheide) | May 2017 | 17 (17) | 2/17 (11.8) | 1/17 (5.9) | 1/17 (5.9) | 0/17 (0) | 1/17 (5.9) |
| 23 | Hanover (Mecklenheide) | May 2017 | 40 (154) | 1/40 (2.5) | 2/40 (5.0) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 24 | Hanover (Ricklinger Teiche) | May 2017 | 40 (805) | 2/40 (5.0) | 0/40 (0) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 25 | Hanover (Mecklenheide) | Jun 2017 | 40 (138) | 1/40 (2.5) | 1/40 (2.5) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 26 | Hanover (Georgengarten) | Jun 2017 | 40 (106) | 0/40 (0) | 3/40 (7.5) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 27 | Emen | Sep 2017 | 40 (55) | 2/40 (5.0) | 0/40 (0) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 28 | Lindwedel | Sep 2017 | 40 (73) | 1/40 (2.5) | 3/40 (7.5) | 0/40 (0) | 1/40 (2.5) | 0/40 (0) |
| 29 | Hanover (Misburger Wald) | May 2018 | 40 (87) | 5/40 (12.5) | 0/40 (0) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 30 | Hanover (Mecklenheide) | May 2018 | 17 (36) | 2/17 (11.8) | 0/40 (0) | 0/17 (0) | 0/17 (0) | 0/17 (0) |
| 31 | Hanover (Mecklenheide) | May 2018 | 40 (145) | 1/40 (2.5) | 1/40 (2.5) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 32 | Hanover (Misburger Wald) | Jun 2018 | 40 (44) | 33/40 (82.5) | 4/40 (10.0) | 0/40 (0) | 2/40 (5.0) | 0/40 (0) |
| 33 | Hanover (Misburger Wald) | Jul 2018 | 40 (194) | 2/40 (5.0) | 2/40 (5.0) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 34 | Hanover (Misburger Wald) | Aug 2018 | 24 (24) | 0/24 (0) | 2/24 (8.3) | 0/24 (0) | 0/24 (0) | 0/24 (0) |
| 35 | Mellendorf | May 2018 | 40 (94) | 2/40 (5.0) | 37/40 (92.5) | 0/40 (0) | 1/40 (2.5) | 0/40 (0) |
| 36 | Mellendorf | May 2018 | 40 (341) | 3/40 (7.5) | 3/40 (7.5) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 37 | Mellendorf | May 2018 | 40 (138) | 1/40 (2.5) | 39/40 (97.5) | 1/40 (2.5) | 1/40 (2.5) | 1/40 (2.5) |
| 38 | Mellendorf | May 2018 | 40 (45) | 0/40 (0) | 8/40 (20.0) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 39 | Mellendorf | May 2018 | 40 (128) | 1/40 (2.5) | 14/40 (35.0) | 0/40 (0) | 1/40 (2.5) | 0/40 (0) |
| 40 | Mellendorf | May 2018 | 40 (72) | 1/40 (2.5) | 9/40 (22.5) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 41 | Mellendorf | May 2018 | 40 (91) | 1/40 (2.5) | 2/40 (5.0) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 42 | Mellendorf | May 2018 | 40 (102) | 13/40 (32.5) | 27/40 (67.5) | 0/40 (0) | 9/40 (22.5) | 0/40 (0) |
| 43 | Mellendorf | May 2018 | 40 (83) | 11/40 (27.5) | 28/40 (70.0) | 0/40 (0) | 11/40 (27.5) | 0/40 (0) |
| 44 | Mellendorf | May 2018 | 40 (87) | 2/40 (5.0) | 28/40 (70.0) | 0/40 (0) | 1/40 (2.5) | 0/40 (0) |
| 45 | Mellendorf | May 2018 | 40 (68) | 2/40 (5.0) | 5/40 (12.5) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 46 | Kassel (Erlenloch) | May 2018 | 40 (215) | 0/40 (0) | 1/40 (2.5) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 47 | Kassel (Niederelsungen) | May 2018 | 40 (1643) | 1/40 (2.5) | 2/40 (5.0) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 48 | Kassel (Niederelsungen) | Jun 2018 | 40 (45) | 0/40 (0) | 0/40 (0) | 0/40 (0) | 0/40 (0) | 0/17 (0) |
| 49 | Uchte (Rauher Busch) | Jun 2018 | 40 (65) | 0/40 (0) | 27/40 (67.5) | 0/40 (0) | 0/40 (0) | 0/40 (0) |
| 50 | Wathlingen (Brand) | May 2018 | 40 (43) | 4/40 (10) | 3/40 (7.5) | 0/40 (0) | 1/40 (2.5) | 0/40 (0) |
| Total | 1500 (5855) | 137/1500 (9.1) | 341/1500 (22.7) | 3/1500 (0.2) | 36/1500 (2.4) | 2/1500 (0.1) |
aIncluding coinfections
Abbreviations: n, number of infected larvae; N, number of examined larvae
Detection of bacterial pathogen DNA
Regarding pathogen prevalence, 137/1500 larvae (9.1%, 95% CI: 7.7–10.7%) from 39/50 nests (78.0%, 95% CI: 64.0–88.5%) were positive for Borrelia spp., while 341/1500 (22.7%, 95% CI: 20.6–24.9%) larvae from 39/50 nests (78.0%, 95% CI: 64.0–88.5%) contained Rickettsia spp. DNA. Anaplasma phagocytophilum DNA was detected in 3/1500 larvae (0.2%, 95% CI: 0–0.6%) from three different nests (6.0 %, 95% CI: 1.3–16.5%).
On average, the mean assumed transovarial transmission rate, i.e. the proportion of positive larvae among the total number of examined larvae of the nest, was 12.3% [SD: 15.4%; min. 2.5% (1/40 larvae), max. 82.5% (33/40 larvae)] in Borrelia-infected nests, for Rickettsia-infected nests 26.5% [SD 29.0%; min. 2.5% (1/40), max. 97.5% (39/40)] and for A. phagocytophilum-infected nests 5.6% [SD 2.9%; min. 2.5% (1/40), max. 8.3% (1/12)]. Prevalences of the different pathogens are pictured in Fig. 2, while detailed data per larvae nest are presented in Table 1. Regarding bacterial abundance in Borrelia-positive larvae, 94.2% (129/137, 95% CI: 88.8–97.4%) of samples contained ≤ 1015S-23S IGS copies and 5.8% (95% CI: 2.6–11.2%) of samples contained between 101 and 102 copies (8/137). On average, 2.7 × 1005S-23S IGS copies were detected in positive DNA samples. For Rickettsia spp., 29.0% (99/341, 95% CI: 24.2–34.2%) of samples contained ≤ 101gltA gene copies, 11.1% (38/341, 95% CI: 8.0–15.0%) of samples contained ≤ 102 copies, 43.4% (148/341, 95% CI: 38.1–48.8%) of samples contained ≤ 102 copies, 16.4% (56/341, 95% CI: 12.7–20.8%) of samples contained between 103 and 104 copies. On average, positive ticks contained 5.4 × 102 copies. The mean copy number of DNA samples of the A. phagocytophilum-infected larvae was 2.4 × 101, with 66.7% (2/3, 95% CI: 9.4–99.2%) samples containing ≤ 101msp2/p44 gene copies and 33.3% (1/3, 95% CI: 0.8–90.6%) between 101 and 102 copies. Copy number distribution for the different pathogens is graphically represented in Fig. 3. Regarding Borrelia prevalence in larvae, no significant differences between the different sampling locations were found (Table 2). In contrast, significantly more larvae were infected with Rickettsia spp. in Mellendorf 2018 than in Hanover 2010, 2017 and 2018 (Table 3).
Fig. 2.
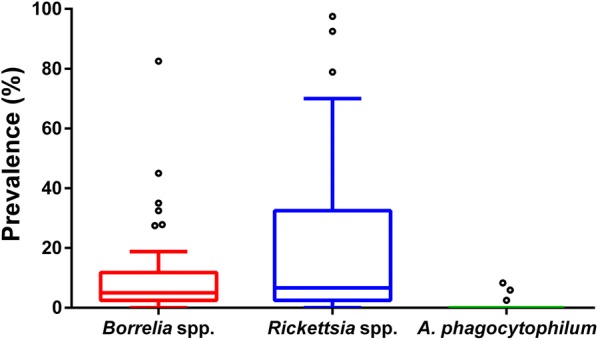
Prevalence of Borrelia spp., Rickettsia spp. and A. phagocytophilum in 50 I. ricinus larvae nests. Boxes extend from the 25th to the 75th percentile, with a line at the median and whiskers extending to 1.5 the interquartile range. Circles represent data points outside of this range (outliers)
Fig. 3.

Distribution of Borrelia spp., Rickettsia spp. and A. phagocytophilum gene copy numbers in DNA samples from positive I. ricinus larvae
Table 2.
Results of the GLMM testing the influence of location/year and coinfection in the nest on Borrelia spp. prevalence in larvae collected in Hanover, Hamburg and Mellendorf (n = 1220 larvae from 43 nests)
| Factor | Estimate | SE | z | P |
|---|---|---|---|---|
| Intercept | − 4.143 | 0.724 | − 5.724 | < 0.001 |
| Hamburg 2011 vs Hanover 2010 | 0.787 | 0.742 | 1.060 | 0.890 |
| Hanover 2015 vs Hamburg 2011 | − 0.846 | 1.203 | − 0.703 | 0.980 |
| Hanover 2017 vs Hamburg 2011 | − 1.206 | 0.783 | − 1.540 | 0.622 |
| Hanover 2018 vs Hamburg 2011 | 0.490 | 0.681 | 0.719 | 0.978 |
| Hanover 2015 vs Hanover 2010 | − 0.059 | 1.286 | − 0.046 | 1.000 |
| Hanover 2017 vs Hanover 2010 | − 0.419 | 0.909 | − 0.461 | 0.997 |
| Hanover 2018 vs Hanover 2010 | 1.277 | 0.811 | 1.574 | 0.599 |
| Hanover 2017 vs Hanover 2015 | − 0.360 | 1.313 | − 0.274 | 1.000 |
| Hanover 2018 vs Hanover 2015 | 1.336 | 1.246 | 1.072 | 0.886 |
| Hanover 2018 vs Hanover 2017 | 1.696 | 0.854 | 1.986 | 0.334 |
| Mellendorf 2018 vs Hanover 2010 | − 0.191 | 0.756 | − 0.253 | 1.000 |
| Mellendorf 2018 vs Hanover 2015 | − 0.132 | 1.211 | − 0.109 | 1.000 |
| Mellendorf 2018 vs Hanover 2017 | 0.228 | 0.796 | 0.286 | 1.000 |
| Mellendorf 2018 vs Hanover 2018 | − 1.468 | 0.699 | − 2.102 | 0.272 |
| Mellendorf 2018 vs Hamburg 2011 | − 0.979 | 0.559 | − 1.750 | 0.481 |
| Coinfection present in the nest | 1.528 | 0.574 | 2.662 | 0.008 |
Notes: The full model was significantly different from a null model containing only the random factor “nest ID” (χ2 = 14.0, df = 6, P = 0.029). Multiple comparisons between levels of the factor “location/year” were performed using Tukeyʼs contrasts with single-step P-value adjustment. Significant P-values (P < 0.05) are shown in bold
Abbreviation: SE, standard error
Table 3.
Results of the GLMM testing the influence of location/year and coinfection in the nest on Rickettsia spp. prevalence in larvae collected in Hanover, Hamburg and Mellendorf (n = 1220 larvae from 43 nests)
| Factor | Estimate | SE | z | P |
|---|---|---|---|---|
| Intercept | − 4.786 | 1.015 | − 4.717 | < 0.001 |
| Hamburg 2011 vs Hanover 2010 | 1.433 | 0.997 | 1.437 | 0.693 |
| Hanover 2015 vs Hamburg 2011 | 1.971 | 1.304 | 1.512 | 0.644 |
| Hanover 2017 vs Hamburg 2011 | − 1.373 | 1.001 | − 1.372 | 0.734 |
| Hanover 2018 vs Hamburg 2011 | − 1.349 | 0.961 | − 1.403 | 0.715 |
| Hanover 2015 vs Hanover 2010 | 3.404 | 1.440 | 2.365 | 0.161 |
| Hanover 2017 vs Hanover 2010 | 0.060 | 1.188 | 0.051 | 1.000 |
| Hanover 2018 vs Hanover 2010 | 0.084 | 1.154 | 0.073 | 1.000 |
| Hanover 2017 vs Hanover 2015 | − 3.344 | 1.427 | − 2.343 | 0.169 |
| Hanover 2018 vs Hanover 2015 | − 3.320 | 1.399 | − 2.373 | 0.158 |
| Hanover 2018 vs Hanover 2017 | 0.024 | 1.148 | 0.021 | 1.000 |
| Mellendorf 2018 vs Hanover 2010 | 2.956 | 0.990 | 2.986 | 0.032 |
| Mellendorf 2018 vs Hanover 2015 | − 0.448 | 1.288 | − 0.348 | 0.999 |
| Mellendorf 2018 vs Hanover 2017 | 2.896 | 0.984 | 2.942 | 0.036 |
| Mellendorf 2018 vs Hanover 2018 | 2.872 | 0.945 | 3.038 | 0.027 |
| Mellendorf 2018 vs Hamburg 2011 | 1.523 | 0.709 | 2.149 | 0.251 |
| Coinfection present in the nest | 1.772 | 0.748 | 2.369 | 0.018 |
Notes: The full model was significantly different from a null model containing only the random factor “nest ID” (χ2 = 28.9, df = 6, P < 0.001). Multiple comparisons between levels of the factor “location/year” were performed using Tukeyʼs contrasts with single-step P-value adjustment. Significant P-values (P < 0.05) are shown in bold
Abbreviation: SE, standard error
Regarding coinfections, 33/50 (66.0%, 95% CI: 51.2–78.8%) nests were positive for Borrelia and Rickettsia spp., while three (6.0%, 95% CI: 1.3–16.5%) nests contained larvae positive for all three pathogens. On the basis of larvae, coinfections were found in 8.6% (38/443, 95% CI: 6.1–11.6%) of the pathogen-positive larvae corresponding to 2.5% (38/1500, 95% CI: 1.8–3.5%) of the total investigated larvae. Rickettsia spp. and Borrelia spp. coinfected larvae were found in 32.0% (16/50, 95% CI: 19.5–46.7%) of nests and in 7.5% (36/478, 95% CI: 5.3–10.3%) of the Rickettsia spp. and Borrelia spp. positive larvae corresponding to 2.4% of all investigated larvae (36/1500, 95% CI: 1.7–3.3%). Coinfection with Rickettsia spp. and A. phagocytophilum was noted in two larvae from 2/50 nests (4.0%, 95% CI: 0.5–13.7%), corresponding to 0.6% (2/344, 95% CI: 0.1–2.8%) of the Rickettsia spp. and A. phagocytophilum positive larvae and 0.1% (2/1500, 95% CI: 0–0.5%) of all investigated larvae. Detailed information on the distribution of coinfections per nest is presented in Table 1. Furthermore, in nests with different pathogens present, larvae had a significantly higher probability of being infected with Borrelia spp. or Rickettsia spp. than in nests with a single pathogen present (Tables 2, 3).
Borrelia (geno-)species and Rickettsia species identification
Borrelia (geno-)species determination by RLB revealed presence of B. spielmanii in one larva (1.4 × 1005S-23S IGS copies). Unfortunately, the Borrelia (geno-)species of the remaining 136 Borrelia-positive larvae could not be determined by RLB. Due to the high prevalence of R. helvetica [5, 6], only a random sample of 63 Rickettsia-positive larvae from 19 nests was subjected to pyrosequencing. R. helvetica was identified in 73.0% (46/63, 95% CI: 60.3–83.4%) of these larvae, while species discrimination failed for the remaining 17 Rickettsia-positive samples. Successfully identified samples had a mean gltA copy number of 6.6 × 102, while the 17 unidentified samples contained 3.2 × 102 copies on average. Of the successfully pyrosequenced larvae, 2.2% (1/46, 95% CI: 0.1–11.5%) contained ≤ 101 copies, 2.2% (1/46, 95% CI: 0.1–11.5%) ≤ 102 copies, 71.7% (33/46, 95% CI: 56.5–84.0%) ≤ 103 copies and 23.9% (11/46, 95% CI: 12.6–38.8%) between 103 and 104 copies. Among the samples which were not successfully sequenced, 47.1% (8/17, 95% CI: 23.0–72.2%) contained ≤ 101 copies, 17.6% (3/17, 95% CI: 3.8–43.4%) contained ≤ 102 copies, 17.6% (3/17, 95% CI: 3.8–43.4%) contained ≤ 103 and 17.6% (3/17, 95% CI: 3.8–43.4%) between 103 and 104 copies.
Discussion
This study aimed to provide an insight into transovarial transmission of Borrelia spp., Rickettsia spp. and A. phagocytophilum in I. ricinus under field conditions by extrapolating from DNA detection rates in questing tick larvae and, by extension, to estimate the potential infection risk for humans and animals by tick larvae bites. Since prevalence studies in central Europe on questing nymphal and adult I. ricinus report higher numbers of Rickettsia- and Borrelia- than Anaplasma-positive ticks [7, 40, 43], correspondingly higher numbers of Rickettsia- and Borrelia-positive nests were expected, regardless of the transovarial transmission efficiency. Indeed, both Rickettsia spp. and Borrelia spp. were detected in 78.0% of nests, while only 6.0% of nests contained A. phagocytophilum-positive larvae. The proportion of nests with Borrelia- and Rickettsia-infected larvae was considerably higher than expected based on the prevalence of these pathogens in questing adult ticks. In studies from Hamburg 2011 and Hanover 2010 and 2015, Borrelia spp. DNA was detected in a total of 34.1% (30.0% adults, 34.5% nymphs) [34], 22.7% (33.3% adults, 20.3% nymphs) [33, 39] and 24.1% (35.4% adults, 19.8% nymphs) [40] of ticks. Rickettsia spp. DNA was detected in a total of 52.5% (56.0% adults, 52.1% nymphs) [6], 26.2% (30.4% adults, 25.5% nymphs) [5, 44] and 50.8% (54.1% adults, 49.5% nymphs) [7]. The discrepancy between the number of positive nests, and consequently the number of infected female ticks these nests originated from, compared to the prevalence in questing adult ticks may be due to several factors. For example, prevalences measured in questing adult ticks exclude those infections that arise during the blood meal of the adult I. ricinus female before oviposition or, in the case of Rickettsia spp., during mating [19]. For A. phagocytophilum, a prevalence of 86.1% has been found in engorged adult ticks collected from roe deer compared to only 8.9% in questing adult ticks, while Rickettsia spp. were detected in 16.6% of engorged adult ticks and 13.9% of questing adult ticks [45]. These infections may be transmitted transovarially, although not as effectively as infections that were acquired earlier [20]. Furthermore, infected larvae might show a higher questing activity than non-infected larvae, e.g. due to higher need of energy as a consequence of infection, or a pathogen-tick-interaction facilitating questing and in turn facilitating pathogen transmission. Thus, infected larvae may be disproportionately represented in the examined larvae collected by the flagging method. Further studies are needed to investigate these explanatory approaches.
The overall infection rate for unfed Ixodes spp. ticks with A. phagocytophilum in the prevalence studies in Hamburg 2011, Hanover 2010 and 2015 was 3.6% (2.1% adults, 3.8% nymphs), 3.2% (1.9% adults, 3.6% nymphs) and 3.8% (7.2% adults, 2.4% nymphs), respectively [5–7, 44]. In comparison, 6% of the nests in this study were infected. Possibly, the same assumptions as for Rickettsia spp. and Borrelia spp. are valid to explain this discrepancy.
Within nests, larval pathogen prevalence differed considerably, ranging between 2.5% and 97.5% for Rickettsia spp., 2.5% and 82.5% for Borrelia spp., as well as 2.5% and 8.3% for A. phagocytophilum. Except for Rickettsia-infections in Hanover (2010, 2015, 2018) vs Mellendorf (2018), no statistically significant regional differences in larval pathogen prevalence were detected. Additionally, to this influence of region, the differences in Rickettsia prevalence within nests might be explained by the degree of rickettsial development in the ovarian tissues of the mother tick at the time of oviposition. Burgdorfer and Brinton [20] described that experimentally infected female ticks with generalised massive infections transmitted Rickettsia spp. to 100% of their offspring, and this was also observed regarding R. helvetica [46]. However, females with a mild rickettsial infection or in the initial phase, had a lower percentage of infected larvae [20].
Overall, Rickettsia prevalence in the collected tick larvae was 22.7%. This is comparable to previous studies, as prevalence of Rickettsia spp. in field-collected Ixodes larvae in Hanover 2005 and 2010 amounted to 27.3% (24/88) and 16.1% (5/31) [4, 5]. In Germany, R. helvetica is the most frequent Rickettsia spp. in I. ricinus [3–7], and was the only Rickettsia spp. that could be defined in larvae in the present study. The proportion of R. helvetica-infected female ticks producing at least one positive egg or larva was described as 100% [17]. The high Rickettsia DNA detection rates in the present study and the identification of R. helvetica in the entire subset of sequenced samples provide evidence for a high transovarial transmission rate for R. helvetica in I. ricinus under field conditions. This is also supported by the high number of Rickettsia gltA copies (average 5.4 × 102 copies) found in the positive larva samples, compared to Borrelia spp. (average 2.7 × 100 copies) or A. phagocytophilum (average 2.4 × 101 copies). Furthermore, the calculated copy numbers may be underestimated, because the tick DNA samples were tested against a plasmid standard. The plasmid standard represents an ideal template, while the tick DNA template is more complex; therefore, the copy numbers of standard versus tick DNA are not fully comparable. However, it should be kept in mind that DNA detection does not necessarily imply that viable bacteria were present in the larvae. The pathogens could have died in the egg, during development from egg to larva or during larval hatching. Furthermore, it is also possible that DNA from dead and lysed pathogens in the mother tick may have entered the eggs and was detected.
In contrast to Rickettsia spp., a rather inefficient transovarial transmission is assumed for B. burgdorferi (s.l.). Nevertheless, van Duijvendijk et al. [47] showed that flagged larvae can transmit B. afzelii and B. miyamotoi to rodents. Thus, larvae also pose a potential infection risk. Borrelia-prevalence in individual, field-collected unfed I. ricinus larvae varied from 0% to 25.8% in previous studies [33, 47–49]. This is in line with the overall larval Borrelia spp. prevalence of 9.1% determined in the present study. Within positive nests, 12.3% of larvae were positive on average, but rates of up to 82.5% were detected. Likewise, Burgdorfer et al. [50] described a transovarial transmission rate of 60.0% and 100.0% in two I. ricinus females infected with Borrelia spirochetes. However, in another study, only one of sixteen B. burgdorferi (s.l.)-infected I. persulcatus females could transmit the pathogen via eggs to the larvae. The infection rate of these larvae was 7.0% [51]. In contrast, an efficient transovarial transmission from female ticks to more than 90.0% of their larvae has been described for the relapsing fever borrelia B. miyamotoi [22, 24]. In individual field-collected larvae, B. miyamotoi showed a prevalence of 0–4.2% [22, 47, 52]. Among Borrelia-positive I. ricinus ticks collected in 2015 in Hanover, B. miyamotoi showed a prevalence of 18.2% in successfully differentiated Borrelia-positive ticks, subdivided into 20.0% in nymphs, 14.0% in females and 18.2% in males [31]. Thus, larvae infected with B. miyamotoi were expected. However, species differentiation using RLB was only successful in one case due to the low number of 5S-23S IGS copies in Borrelia-positive larvae. In this larva originating from a nest with a transovarial transmission rate of 82.5%, B. spielmanii was identified. For the remaining nests, it was not possible to determine whether the DNA originated from B. burgdorferi (s.l.) or B. miyamotoi, and thus not possible to investigate whether there was a correlation between rather low or high transmission rates and the two Borrelia species. Furthermore, the nests may have been infected with several Borrelia (geno-)species, including B. miyamotoi. Additionally, transovarial transmission rates of B. burgdorferi (s.l.) might vary between genospecies. Further laboratory studies examining transovarial transmission in ticks infected with known Borrelia (geno-)species are needed to achieve a clearer picture and to assess the potential Borrelia infection risk posed by larvae.
Due to the low A. phagocytophilum prevalence, there were mainly coinfections with Borrelia spp. and Rickettsia species. In total, 66.0% of all nests and 2.5% of all larvae were coinfected with Rickettsia spp. and Borrelia spp., while for nymphs and adult ticks, coinfection rates between 7.3% and 22.9% have been described in northern Germany [4, 37, 40]. Furthermore, larvae had a significantly higher probability of being infected with Borrelia spp. or Rickettsia spp. when the nest was coinfected, leading to the hypothesis that coinfection of the mother tick promotes transovarial transmission efficiency.
Transovarial transmission of A. phagocytophilum is described as absent or inefficient in the literature [16, 25]. A study by Baldridge et al. [53] showed transovarial transmission rates of 10.0–40.0% in Dermacentor albipictus, while Jahfari et al. [27] and Hagedorn [54] detected A. phagocytophilum in 1.3% of field-collected I. ricinus larvae. In Hanover, positive A. phagocytophilum larvae have not been detected in previous studies [5, 44]. The detection of A. phagocytophilum DNA in three unfed larvae in three different nests in the present study shows that transovarial transmission may be possible, but that it is inefficient. Therefore, a potential risk of A. phagocytophilum infection by I. ricinus larvae appears to be low.
Conclusions
Detection of pathogen DNA in questing I. ricinus larvae provides evidence for transovarial transmission of Rickettsia spp. and Borrelia spp. under field conditions. In consequence, I. ricinus larvae might serve as a source of human or animal infection with these pathogens. However, further studies investigating the percentage of viable transovarially transmitted Rickettsia and Borrelia species are necessary. As expected, transovarial transmission of A. phagocytophilum was rare.
Acknowledgements
We thank Julia Tappe, Kathrin May, Katrin Blazejak, Steffen Knoll, Katrin Liebig and Mathias Boelke for contribution of larvae nest.
Authors’ contributions
DH and DJ collected larvae nests and performed molecular analyses. DH performed statistical analyses and drafted the manuscript. AS supported statistical analyses. VF provided Borrelia positive controls. CS designed and coordinated the study. DH, DJ, AS, BS, SP, VF and CS participated in data analysis and interpretation, and reviewed the manuscript draft. All authors read and approved the final manuscript.
Funding
The study was supported by funding from Bayer Vital GmbH and Bayer Animal Health GmbH.
Availability of data and materials
The data analysed during this study are included in the article.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
SP is an employee of Bayer Vital GmbH, BS is an employee of Bayer Animal Health GmbH. Study data collection and interpretation is completely independent from the company’s opinion and the authors declare that there is no conflict with commercial interests. DH, DJ, AS, VF and CS declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Daniela Hauck, Email: daniela.hauck@tiho-hannover.de.
Daniela Jordan, Email: daniela.jordan@tiho-hannover.de.
Andrea Springer, Email: andrea.springer@tiho-hannover.de.
Bettina Schunack, Email: bettina.schunack@bayer.com.
Stefan Pachnicke, Email: stefan.pachnicke@bayer.com.
Volker Fingerle, Email: volker.fingerle@lgl.bayern.de.
Christina Strube, Email: christina.strube@tiho-hannover.de.
References
- 1.Hovius JWR, de Wever B, Sohne M, Brouwer MC, Coumou J, Wagemakers A, et al. A case of meningoencephalitis by the relapsing fever spirochaete Borrelia miyamotoi in Europe. Lancet. 2013;382:658. doi: 10.1016/S0140-6736(13)61644-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Boden K, Lobenstein S, Hermann B, Margos G, Fingerle V. Borrelia miyamotoi-associated neuroborreliosis in immunocompromised person. Emerg Infect Dis. 2016;22:1617–1620. doi: 10.3201/eid2209.152034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dobler G, Wölfel R. Typhus and other rickettsioses: emerging infections in Germany. Dtsch Arztebl Int. 2009;106:348–354. doi: 10.3238/arztebl.2009.0348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Schicht S, Schnieder T, Strube C. Rickettsia spp. and coinfections with other pathogenic microorganisms in hard ticks from northern Germany. J Med Entomol. 2012;49:766–771. doi: 10.1603/ME11204. [DOI] [PubMed] [Google Scholar]
- 5.Tappe J, Strube C. Anaplasma phagocytophilum and Rickettsia spp. infections in hard ticks (Ixodes ricinus) in the city of Hanover (Germany): revisited. Ticks Tick Borne Dis. 2013;4:432–438. doi: 10.1016/j.ttbdis.2013.04.009. [DOI] [PubMed] [Google Scholar]
- 6.May K, Strube C. Prevalence of Rickettsiales (Anaplasma phagocytophilum and Rickettsia spp.) in hard ticks (Ixodes ricinus) in the city of Hamburg, Germany. Parasitol Res. 2014;113:2169–2175. doi: 10.1007/s00436-014-3869-x. [DOI] [PubMed] [Google Scholar]
- 7.Blazejak K, Janecek E, Strube C. A 10-year surveillance of Rickettsiales (Rickettsia spp. and Anaplasma phagocytophilum) in the city of Hanover, Germany, reveals Rickettsia spp. as emerging pathogens in ticks. Parasit Vectors. 2017;10:588. doi: 10.1186/s13071-017-2537-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Thomas RJ, Birtles RJ, Radford AD, Woldehiwet Z. Recurrent bacteraemia in sheep infected persistently with Anaplasma phagocytophilum. J Comp Pathol. 2012;147:360–367. doi: 10.1016/j.jcpa.2012.02.005. [DOI] [PubMed] [Google Scholar]
- 9.Tegtmeyer P, Ganter M, von Loewenich FD. Simultaneous infection of cattle with different Anaplasma phagocytophilum variants. Ticks Tick Borne Dis. 2019;10:1051–1056. doi: 10.1016/j.ttbdis.2019.05.011. [DOI] [PubMed] [Google Scholar]
- 10.Dumler JS, Choi K-S, Garcia-Garcia JC, Barat NS, Scorpio DG, Garyu JW, et al. Human granulocytic anaplasmosis and Anaplasma phagocytophilum. Emerg Infect Dis. 2005;11:1828–1834. doi: 10.3201/eid1112.050898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Levin ML, Fish D. Immunity reduces reservoir host competence of Peromyscus leucopus for Ehrlichia phagocytophila. Infect Immun. 2000;68:1514–1518. doi: 10.1128/IAI.68.3.1514-1518.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ogden N, Nuttall P, Randolph S. Natural Lyme disease cycles maintained via sheep by co-feeding ticks. Parasitology. 1997;115:591–599. doi: 10.1017/S0031182097001868. [DOI] [PubMed] [Google Scholar]
- 13.Raoult D, Roux V. Rickettsioses as paradigms of new or emerging infectious diseases. Clin Microbiol Rev. 1997;10:694–719. doi: 10.1128/CMR.10.4.694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Humair P-F, Gern LJM. The wild hidden face of Lyme borreliosis in Europe. Infection. 2000;2:915–922. doi: 10.1016/s1286-4579(00)00393-2. [DOI] [PubMed] [Google Scholar]
- 15.Monin R, Gern L, Aeschlimann A. A study of the different modes of transmission of Borrelia burgdorferi by Ixodes ricinus. Zentralbl Bakteriol. 1989;2(Suppl. 18):14–20. [Google Scholar]
- 16.Rikihisa Y. Mechanisms of obligatory intracellular infection with Anaplasma phagocytophilum. Clin Microbiol Rev. 2011;24:469–489. doi: 10.1128/CMR.00064-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Socolovschi C, Mediannikov O, Raoult D, Parola P. The relationship between spotted fever group Rickettsiae and ixodid ticks. Vet Res. 2009;40:1–20. doi: 10.1051/vetres/2009017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gaber MS, Khalil GM, Hoogstraal H. Borrelia crocidurae: venereal transfer in Egyptian Ornithodoros erraticus ticks. Exp Parasitol. 1982;54:182–184. doi: 10.1016/0014-4894(82)90125-4. [DOI] [PubMed] [Google Scholar]
- 19.Hayes S, Burgdorfer W, Aeschlimann A. Sexual transmission of spotted fever group Rickettsiae by infected male ticks: detection of Rickettsiae in immature spermatozoa of Ixodes ricinus. Infect Immun. 1980;27:638–642. doi: 10.1128/IAI.27.2.638-642.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Burgdorfer W, Brinton LP. Mechanisms of transovarial infection of spotted fever rickettsiae in ticks. Ann N Y Acad Sci. 1975;266:61–72. doi: 10.1111/j.1749-6632.1975.tb35088.x. [DOI] [PubMed] [Google Scholar]
- 21.Piesman J, Sinsky RJ. Ability of Ixodes scapularis, Dermacentor variabilis, and Amblyomma americanum (Acari: Ixodidae) to acquire, maintain, and transmit Lyme disease spirochetes (Borrelia burgdorferi) J Med Entomol. 1988;25:336–339. doi: 10.1093/jmedent/25.5.336. [DOI] [PubMed] [Google Scholar]
- 22.Richter D, Debski A, Hubalek Z, Matuschka FR. Absence of Lyme disease spirochetes in larval Ixodes ricinus ticks. Vector Borne Zoon Dis. 2012;12:21–27. doi: 10.1089/vbz.2011.0668. [DOI] [PubMed] [Google Scholar]
- 23.Voordouw MJ. Co-feeding transmission in Lyme disease pathogens. Parasitology. 2015;142:290–302. doi: 10.1017/S0031182014001486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Scoles GA, Papero M, Beati L, Fish D. A relapsing fever group spirochete transmitted by Ixodes scapularis ticks. Vector Borne Zoon Dis. 2001;1:21–34. doi: 10.1089/153036601750137624. [DOI] [PubMed] [Google Scholar]
- 25.Ogden N, Bown K, Horrocks B, Woldehiwet Z, Bennett M. Granulocytic Ehrlichia infection in ixodid ticks and mammals in woodlands and uplands of the UK. Med Vet Entomol. 1998;12:423–429. doi: 10.1046/j.1365-2915.1998.00133.x. [DOI] [PubMed] [Google Scholar]
- 26.Krücken J, Schreiber C, Maaz D, Kohn M, Demeler J, Beck S, et al. A novel high-resolution melt PCR assay discriminates Anaplasma phagocytophilum and “Candidatus Neoehrlichia mikurensis”. J Clin Microbiol. 2013;51:1958–1961. doi: 10.1128/JCM.00284-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jahfari S, Coipan EC, Fonville M, Van Leeuwen AD, Hengeveld P, Heylen D, et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasit Vectors. 2014;7:365. doi: 10.1186/1756-3305-7-365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gray J, Kahl O, Lane R, Levin M, Tsao J. Diapause in ticks of the medically important Ixodes ricinus species complex. Ticks Tick Borne Dis. 2016;7:992–1003. doi: 10.1016/j.ttbdis.2016.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Springer A, Raulf MK, Fingerle V, Strube C. Borrelia prevalence and species distribution in ticks removed from humans in Germany, 2013–2017. Ticks Tick Borne Dis. 2020;11:101363. doi: 10.1016/j.ttbdis.2019.101363. [DOI] [PubMed] [Google Scholar]
- 30.Mangold A, Bargues M, Mas-Coma S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae) Parasitol Res. 1998;84:478–484. doi: 10.1007/s004360050433. [DOI] [PubMed] [Google Scholar]
- 31.Hauck D, Springer A, Pachnicke S, Schunack B, Fingerle V, Strube C. Ixodes inopinatus in northern Germany: occurrence and potential vector role for Borrelia spp., Rickettsia spp., and Anaplasma phagocytophilum in comparison with Ixodes ricinus. Parasitol Res. 2019;118:3205–3216. doi: 10.1007/s00436-019-06506-4. [DOI] [PubMed] [Google Scholar]
- 32.Estrada-Peña A, Nava S, Petney T. Description of all the stages of Ixodes inopinatus n. sp. (Acari: Ixodidae) Ticks Tick Borne Dis. 2014;5:734–743. doi: 10.1016/j.ttbdis.2014.05.003. [DOI] [PubMed] [Google Scholar]
- 33.Tappe J, Jordan D, Janecek E, Fingerle V, Strube C. Erratum to: Revisited: Borrelia burgdorferi sensu lato infections in hard ticks (Ixodes ricinus) in the city of Hanover (Germany) Parasit Vectors. 2016;9:249. doi: 10.1186/s13071-016-1516-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.May K, Jordan D, Fingerle V, Strube C. Borrelia burgdorferi sensu lato and co-infections with Anaplasma phagocytophilum and Rickettsia spp. in Ixodes ricinus in Hamburg, Germany. Med Vet Entomol. 2015;29:425–429. doi: 10.1111/mve.12125. [DOI] [PubMed] [Google Scholar]
- 35.Strube C, Montenegro VM, Epe C, Eckelt E, Schnieder T. Establishment of a minor groove binder-probe based quantitative real time PCR to detect Borrelia burgdorferi sensu lato and differentiation of Borrelia spielmanii by osp A-specific conventional PCR. Parasit Vectors. 2010;3:69. doi: 10.1186/1756-3305-3-69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Stenos J, Graves SR, Unsworth NB. A highly sensitive and specific real-time PCR assay for the detection of spotted fever and typhus group rickettsiae. Am J Trop Med Hyg. 2005;73:1083–1085. doi: 10.4269/ajtmh.2005.73.1083. [DOI] [PubMed] [Google Scholar]
- 37.Sirigireddy KR, Ganta RR. Multiplex detection of Ehrlichia and Anaplasma species pathogens in peripheral blood by real-time reverse transcriptase-polymerase chain reaction. J Mol Diagn. 2005;7:308–316. doi: 10.1016/S1525-1578(10)60559-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Courtney JW, Kostelnik LM, Zeidner NS, Massung RF. Multiplex real-time PCR for detection of anaplasma phagocytophilum and Borrelia burgdorferi. J Clin Microbiol. 2004;42:3164–3168. doi: 10.1128/JCM.42.7.3164-3168.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tappe J, Jordan D, Janecek E, Fingerle V, Strube C. Revisited: Borrelia burgdorferi sensu lato infections in hard ticks (Ixodes ricinus) in the city of Hanover (Germany) Parasit Vectors. 2014;7:441. doi: 10.1186/1756-3305-7-441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Blazejak K, Raulf MK, Janecek E, Jordan D, Fingerle V, Strube C. Shifts in Borrelia burgdorferi (s.l.) geno-species infections in Ixodes ricinus over a 10-year surveillance period in the city of Hanover (Germany) and Borrelia miyamotoi-specific Reverse Line Blot detection. Parasit Vectors. 2018;11:304. doi: 10.1186/s13071-018-2882-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Janecek E, Streichan S, Strube C. SNP-based real-time pyrosequencing as a sensitive and specific tool for identification and differentiation of Rickettsia species in Ixodes ricinus ticks. BMC Infect Dis. 2012;12:261. doi: 10.1186/1471-2334-12-261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.R Development Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2018. https://www.R-project.org/.
- 43.Rizzoli A, Silaghi C, Obiegala A, Rudolf I, Hubalek Z, Foldvari G, et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: new hazards and relevance for public health. Front Public Health. 2014;2:251. doi: 10.3389/fpubh.2014.00251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tappe J, Strube C. Corrigendum to “Anaplasma phagocytophilum and Rickettsia spp. infections in hard ticks (Ixodes ricinus) in the city of Hanover (Germany): Revisited” [Ticks Tick Borne Dis. 4 (2013) 432-438] Ticks Tick Borne Dis. 2016;7:1059–1060. doi: 10.1016/j.ttbdis.2016.03.004. [DOI] [PubMed] [Google Scholar]
- 45.Overzier E, Pfister K, Herb I, Mahling M, Bock G, Jr, Silaghi C. Detection of tick-borne pathogens in roe deer (Capreolus capreolus), in questing ticks (Ixodes ricinus), and in ticks infesting roe deer in southern Germany. Ticks Tick Borne Dis. 2013;4:320–328. doi: 10.1016/j.ttbdis.2013.01.004. [DOI] [PubMed] [Google Scholar]
- 46.Burgdorfer W, Barbour AG, Hayes SF, Péter O, Aeschlimann A. Ixodes ricinus: vector of a hitherto undescribed spotted fever group agent in Switzerland. Acta Trop. 1979;36:357–367. [PubMed] [Google Scholar]
- 47.van Duijvendijk G, Coipan C, Wagemakers A, Fonville M, Ersoz J, Oei A, et al. Larvae of Ixodes ricinus transmit Borrelia afzelii and B. miyamotoi to vertebrate hosts. Parasit Vectors. 2016;9:97. doi: 10.1186/s13071-016-1389-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Mejlon HA, Jaenson TG. Seasonal prevalence of Borrelia burgdorferi in Ixodes ricinus in different vegetation types in Sweden. Scand J Infect Dis. 1993;25(4):449–456. doi: 10.3109/00365549309008526. [DOI] [PubMed] [Google Scholar]
- 49.Rijpkema S, Nieuwenhuijs J, Franssen F, Jongejan F. Infection rates of Borrelia burgdorferi in different instars of Ixodes ricinus ticks from the Dutch North Sea Island of Ameland. Exp Appl Acarol. 1994;18:531–542. doi: 10.1007/BF00058936. [DOI] [PubMed] [Google Scholar]
- 50.Burgdorfer W, Barbour A, Hayes S, Peter O, Aeschlimann A. Erythema chronicum migrans - a tickborne spirochetosis. Acta Trop. 1983;40:79–83. [PubMed] [Google Scholar]
- 51.Nefedova VV, Korenberg EI, Gorelova NB, Kovalevskii YV. Studies on the transovarial transmission of Borrelia burgdorferi sensu lato in the taiga tick Ixodes persulcatus. Folia Parasitol. 2004;51:67–71. doi: 10.14411/fp.2004.010. [DOI] [PubMed] [Google Scholar]
- 52.Kjelland V, Rollum R, Korslund L, Slettan A, Tveitnes D. Borrelia miyamotoi is widespread in Ixodes ricinus ticks in southern Norway. Ticks Tick Borne Dis. 2015;6:516–521. doi: 10.1016/j.ttbdis.2015.04.004. [DOI] [PubMed] [Google Scholar]
- 53.Baldridge GD, Scoles G, Burkhardt NY, Schloeder B, Kurtti TJ, Munderloh UG. Transovarial transmission of Francisella-like endosymbionts and Anaplasma phagocytophilum variants in Dermacentor albipictus (Acari: Ixodidae) J Med Entomol. 2009;46:625–632. doi: 10.1603/033.046.0330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hagedorn P. Untersuchung von Zecken als Marker für die Gefährdung durch von ihnen übertragene Krankheiten: Die Prävalenz von Borrelien, Babesien, Rickettsien und Anaplasmen in Ixodes ricinus, Dermacentor reticulatus und Haemaphysalis concinna. Thesis, University of Berlin, Germany; 2013.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data analysed during this study are included in the article.