

Abstract
Microfluidic flow cells are used in single-molecule experiments, enabling measurements to be made with high spatial and temporal resolution. We discuss the fundamental processes affectingflow cell operation and describe the flow cellsin use at present for studying the interactionof optically trapped or mechanically isolated, single DNA molecules with proteins. To assist the experimentalist in flow cell selection, we reviewthe construction techniques and materials used to fabricate both single- and multiple-channel flow cells and the advantages of each design for different experiments.
Observation of protein interactions with individual DNA molecules allows one to measure kinetic and structural changes that are not easy to study using bulk biochemical techniques. In this single-molecule approach, a DNA molecule is bound to a microsphere that is held in place within a microfluidic flow cell using an optical trap or micropipette; or, alternatively, DNA is immobilized on the surface of the flow chamber. Alterations in the DNA and/or protein are monitored by fluorescence or force microscopy. This method has been successfully used to study DNA helicases1–4, DNA packaging motors5,6, dsDNA translocases7,8, nucleases9, DNA polymerases10–12, RNA polymerases13,14, RNA helicases15,16 and DNA and RNA dynamics17,18 and to observe DNA structural changes induced by binding of proteins to DNA19–26. Microfluidic flow cells are key in the above experiments because they allow controlled measurements at the single-molecule level with high spatial and temporal resolution.
The term ‘microfluidic’ refers to the ability to manipulate fluids in one or more channels with dimensions of 5–500 μm (refs. 27,28). All the flow cells discussed in this review can be considered microfluidic and are composed of one or more inlet channels that flow into a common physical channel where experimental measurements take place. The volume of liquid contained in the flow cells is in the range of tens of microliters, so only limited quantities of reagents are used. This, coupled with small-internal-diameter tubing, results in short times for fluid to flow from the fluid delivery device to the observation point within the flow cell at velocities that do not shear DNA.
Flow cells are commonly referred to as single- or multichannel designs. We use the term ‘stream’ to refer to the fluid within a channel and the term ‘channel’ to refer to the physical channel in which fluid streams move. A multichannel design is defined as a flow cell in which two or more entry channels converge into a single, common, physical channel. Within the confines of the common channel, the fluid streams originating from the individual channels flow parallel to one another and remain separate, with only minimal mixing between them occurring from diffusion. Thus it is more accurate to view these designs as ‘multistream’ flow cells. Single-channel designs also have a common, physical channel where reactions are observed, but there is only one fluid stream present. Thus, these designs are ‘single-stream’ flow cells.
We will discuss several flow cell designs, each having distinct advantages for performing specific kinds of measurements. Several components are essential to success, including the material used to construct the cell, the geometry of the design, the number of channels involved, the tubing used to mate the cell with the fluid delivery device and the fluid delivery device itself (Supplementary Table 1 online). There are also other microfluidic flow cells, some quite complex, for studying many other phenomena at the single-molecule level. These studies are not discussed here. Instead, we limit our discussion to those experiments in which the interactions of proteins with DNA have been studied within the confines of laminar flow cells.
Laminar flow
Fluid flow within the channels of the flow cells in this review is laminar27,28 and follows the contour lines of the channel that confines it, requiring no stringent design consideration to route fluid from one point to another within the flow cell (Box 1). An essential aspect of laminar flow in multistream flow cells that is critical to single-molecule experiments is that adjacent streams of miscible fluids flow side by side, with mixing between them occurring only by diffusion at their interface29–33. The distance that molecules diffuse between adjacent streams in the flow cell depends on the molecular weight of the molecule, the viscosity of the buffer, the distance from the channel junction and the velocity of the fluid flow (Fig. 1a). Below, the flow characteristics specific to laminar flow cells are discussed briefly. For a more detailed discussion of microfluidics, the reader is referred to an excellent review by Squires and Quake27.
BOX 1. FLUID FLOW IS LAMINAR IN MULTISTREAM FLOW CELLS.
The Reynolds number (Re) is a dimensionless quantity that describes the behavior of fluids and is defined as
where v is the fluid velocity, l is a characteristic channel dimension, ρ is the fluid density and η is the fluid viscosity. At high Reynolds numbers (that is, Re > 2,000), fluid flow is turbulent. At low Reynolds numbers (that is, Re < ~2,000), fluid flow is laminar.
For the design shown in Figures 4 and 5, channels with widths between 0.25 and 2 mm and depths ranging from 30 to 70 μm have been used1,8,19. For typical fluid flow conditions, v = 30 μm/s, ρ = 1.23 g/cm3, l = 40 μm and η = 15.3 cP (50% sucrose buffer), the resulting Reynolds number is significantly lower than 1, using either the width or the depth of the channel in the above formula. Therefore, the fluid flow within the microchannels is laminar. Increasing the buffer velocity and/or decreasing in buffer viscosity do not significantly affect the Reynolds number, so that fluid flow remains laminar even when water is used (η = 1 cP).
Figure 1 |.
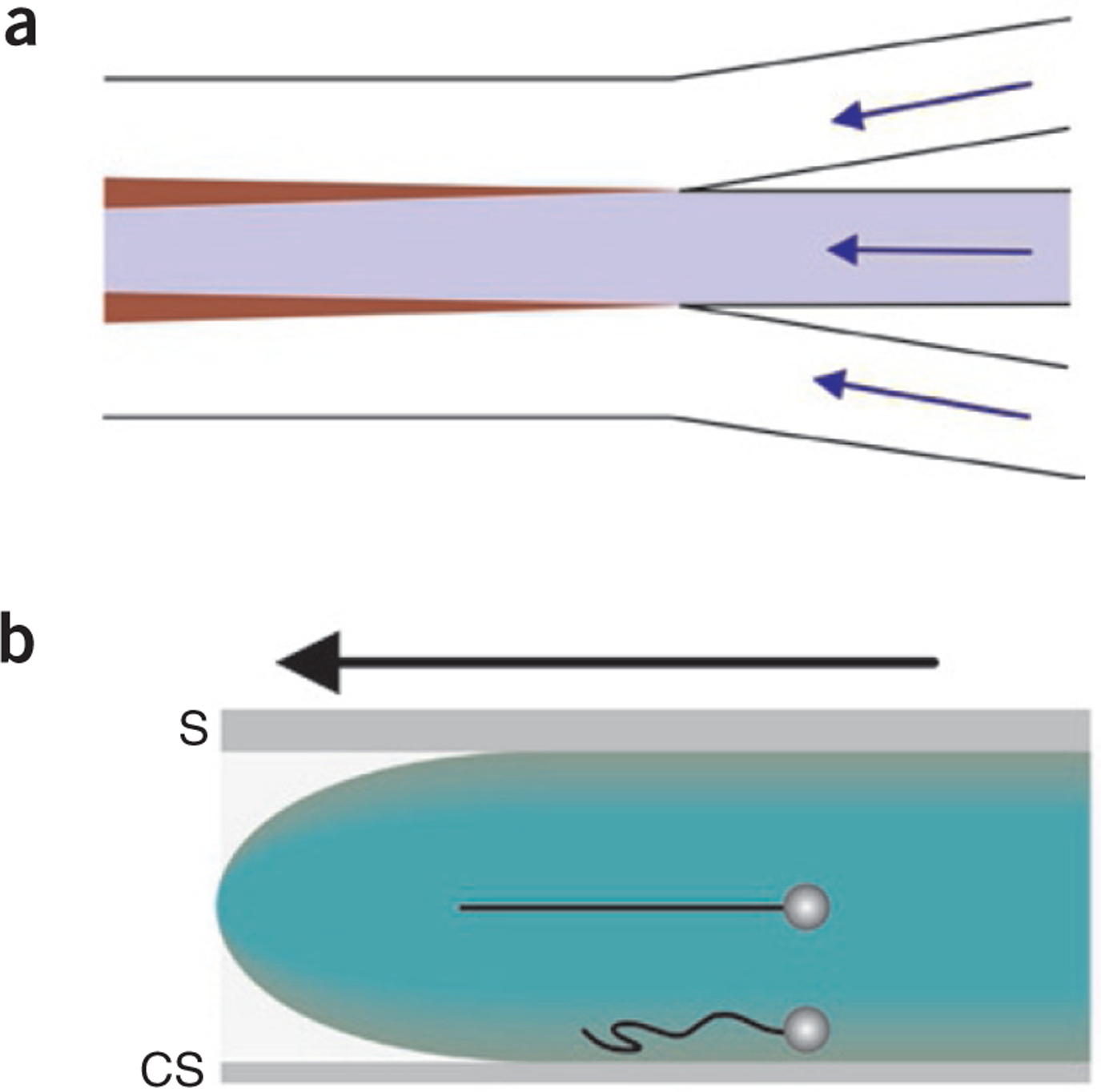
Fluid flow in multistream laminar flow cells. (a) Interchannel diffusion is the main source of mixing between adjacent fluid streams. The flow paths are indicated by blue arrows, and the individual streams are colored white, blue and white. The widening regions of transverse diffusion between channels are indicated in maroon. (b) The flow profile (green) in laminar flow cells is parabolic. The flow cell is viewed from the side with the direction of flow indicated by the black arrow. The fastest fluid velocities occur at the center of the flow cell, and the slowest occur next to the flow cell surfaces. The upper optically trapped DNA-bead complex is positioned ~18 μm from the surface, resulting in full extension of the DNA molecule (black string). Closer to the surface, DNA molecules are not fully stretched (lower DNA molecule). S, slide; CS, coverslip.
Experiments with multistream flow cells demonstrate that the main source of interstream mixing is transverse diffusion (Box 2)29–33. Analysis of mixing between streams carrying the fluorescent, calcium-responsive dye Fluo-3 (molecular mass, 770 Da) and calcium ions in a two-stream flow cell showed that a small amount of mixing occurs within the first few microns of the immediate flow junction; that is, at the dead-volume region of the flow cell31. Beyond this region, transverse diffusion between fluid streams varies as the square root of the distance from the junction and as the inverse of the square root of the flow velocity, except that, near the surfaces of the flow cell, diffusion between the two streams varies as the cube root of the distance from the junction and inversely as the cube root of flow velocity. Transverse diffusion is an important consideration for small molecules such as nucleoside triphosphates and divalent metal ions that modulate protein activity, as well as for proteins themselves, although because of their higher molecular masses, the proteins diffuse less (Box 2).
BOX 2. TRANSVERSE DIFFUSION WITHIN LAMINAR FLOW CELLS.
Mixing of molecules between adjacent streams in multistream flow cells occurs through transverse diffusion. The distance of transverse diffusion, x, is given by <x2> = 2Dt, where <x2> is the mean-square value of x and the diffusion constant D is defined as
where kB is Boltzmann’s constant, T is the temperature in degrees Kelvin, η is the viscosity of the buffer, a is the molecular radius of the protein and t is the time it takes fluid to flow a distance L from the junction of the flow cell with a flow velocity v to the observation site. Substituting for D and inserting t = L/v in the above equation, we find that:
Note that the transverse diffusion distance x varies as the square root of the distance from the microchannel junction, L, and the inverse square root of the flow velocity, v (ref. 31). For η = 15.3 cP, L = 300 μm, v = 30 μm/s, T = 300 K and a = 1.6 nm (a 10-kDa protein), the average transverse diffusion distance x ≈ 10 μm. A flow velocity of v = 30 μm/s implies that measurements made at a distance L = 300 μm or less from the flow cell channel junction will limit the diffusion of proteins (or other molecules with a molecular weight of 10 kDa or greater) into adjacent streams to distances less than 10 μm. Therefore, to minimize mixing resulting from transverse diffusion, experiments are conducted within 200 μm of the channel junction, and approximately 10 μm beyond the dead volume region at the immediate flow junction (Figs. 1a and 5, upper inset)31. As the diffusion constant is related inverselyto the molecular radius of the protein, larger proteins will diffuse less29,30,33. The diffusion constant is also related inversely to the viscosity of the buffer. Therefore, protein transverse diffusion distances for experiments performed with buffer viscosity equalto that of water (1 cP) will be almost four times higher than those performed with 50% sucrose (15.3 cP).
Another important flow factor is the parabolic velocity profile within flow cell channels under laminar flow. The highest flow velocity occurs in the middle of the channel and the slowest occurs next to the surfaces (Fig. 1b)34. The velocity profile within the channel must be taken into account in DNA applications: for example, when one wishes to fully stretch a DNA molecule attached to a bead held in the stream for precise visualization of fluorescently tagged DNA and accurate single-molecule analyses. Typical pump rates of 10 μl/h result in linear flow velocities of 112 μm/s (measured at a depth of 18 μm from the coverslip surface)8. Under these conditions, reduced rates of fluid flow are observed within ~9 μm of the channel surface, resulting in suboptimal stretching of DNA molecules (Fig. 1b). Transverse diffusion is greater in these regions of reduced flow31. In contrast, at greater depths, (that is, between 10 and 20 μm), the flow velocity is more rapid, DNA molecules are extended close to their full contour length and the DNA end distal to the bead in the flow fluctuates less, enabling more precise measurements. For a more detailed discussion of DNA molecules in flow, see ref. 17.
Flow cell materials and channel formation
The materials used for flow cell construction are key for their success. Ideal materials should be mechanically rigid, optically transparent with low fluorescence and impervious to organic solvents. Such materials can provide a stable flow environment for an extended period of time suitable for optical trapping, visualization and force detection. These materials also permit the use of nonaqueous solvents (for example, methanol) to simplify surface wetting and removal of air bubbles and the use of denaturants (for example, 6 M guanidinium hydrochloride) to clean the flow cell.
Several materials have found utility in a variety of microfluidic flow cells35–37. Many of these can be excluded from single-molecule work because they either have high autofluorescence, are not optically transparent, or both38. Poly(dimethylsiloxane) or PDMS, poly(methylmethacrylate) or PMMA, glass and quartz all have low autofluorescence, with quartz excelling in this regard38,39. Both PDMS and PMMA have been used successfully in flow cell construction and for fluorescence detection of DNA38,40. The versatility of PDMS makes fabrication of various numbers of channels straightforward and reduces the time, complexity and cost of prototyping and manufacturing flow cells. Consequently, PDMS is widely used in microfluidics28,37,39 and has been combined with glass coverslips to make hybrid flow cells suitable for optical trapping of DNA41,42. Unfortunately, PDMS is not fully compatible with organic solvents. These are absorbed by the PDMS, which leads to swelling39,43,44. The surface properties of PDMS can also be difficult to control, and PDMS is elastomeric, which may lead to shrinking or sagging over time45. Therefore, PDMS is most suitable for prototyping or for construction of flow cells intended for short-term use.
Compared to polymers, glass and quartz have considerably lower autofluorescence, making them ideal for fluorescence microscopy. They are mechanically rigid and resistant to organic solvents and denaturants, and quartz is resistant to scratching. Flow cells constructed entirely of these materials can be rigorously cleaned using denaturants and used repeatedly for periods greater than one year.
A cost-effective alternative to flow cells constructed entirely of quartz or glass is to sandwich pieces of carefully cut Parafilm, acrylic or double-sided tape between glass slides and coverslips. The sandwich material has the channel design cut into it and determines both channel depth and width. Sealing a flow cell containing Parafilm requires gentle heating46, whereas tape already has adhesive attached. Although tape and Parafilm are commonly used to make single-channel flow cells, they have also been used to construct multichannel designs26,47.
The channel patterns for flow cells made of glass, quartz or polymers can be created using a variety of techniques37,48. For PDMS, soft-lithography is straightforward and is performed by casting against a suitable mold, resulting in features with sub-0.1 μm fidelity28,39,42. The fabrication method used by MMR Technologies for creating channels in a glass microscope slide is ‘dry etching’ (essentially micro-sandblasting), which leaves a faint pitted pattern on the surface of the glass. This does not significantly affect flow or background fluorescence. Wet etching of glass or quartz is preferable because of the absolute transparency of the resultant channels49, which is an important consideration for both fluorescence and force microscopy. The angle at which channels join before entering the common channel in a multichannel flow cell is not critical as long as the resulting flow is laminar37,50,51. Many laminar-flow, multichannel devices have been built with angles between channels as large as 90 degrees with no adverse mixing effects in the common channel28.
Fluid delivery into flow cells
Once a flow cell is constructed, it is connected to a fluid delivery device to introduce reaction components. It is possible to pull liquid through a flow chamber by applying vacuum to the outlet while connecting the inlet to a reservoir (or reservoirs) containing reaction components6,11. More frequently, various pressure systems are used to apply positive pressure at the inlet, with the outlet open to atmospheric pressure52. These include systems combining a syringe pump and computer-controlled solenoid valves46, pressure driven pumps53, peristaltic pumps54, a combination of pressure regulators and buffer reservoirs12,42, single syringe pumps with the capability of handling multiple syringes simultaneously1,8,19,41, or several syringe pumps functioning in concert while connected to a single flow cell26.
The use of solenoid valves to control flow is advantageous for force measurements because the manipulation of air pressure provides smooth transitions between flow speeds and overall smoother flow46,47. In contrast, syringe pumps, though straightforward to implement as off-the-shelf components, are often powered by stepper motors, and these can introduce mechanical pulsation into the flow. This can affect the measurement of position and force using beads held by optical traps.
The type of tubing used to connect flow cells with fluid delivery devices and the means by which tubing is attached to the flow cell and pump system are critical. The tubing should have a small internal diameter (that is, ≤ 500 μm) to minimize the use of reagents and should be mechanically rigid, chemically resistant and inert as far as DNA and proteins are considered. Connections must be airtight and stable, and the adhesive or epoxy used to bond the connectors or tubing to the flow cell must be chemically inert once it has cured. PEEK tubing (Upchurch Scientific) meets all of the above criteria and has been used in many studies1,7,8,19,26,47,53. It is easily attached to flow cells using connectors manufactured out of the same material (Supplementary Fig. 1 online).
The type of syringe used to control flow can have large effects on flow stability. Hamilton gastight syringes are superior to any disposable syringe, as they have stiff glass walls and Teflon polytetrafluoroethylene-coated plunger tips, which combine to provide the exceptionally smooth plunger motion that is essential to obtaining smooth flow1,8. Using glass syringes and PEEK tubing, the user can start and stop the flow rapidly because mechanical energy is not stored in the wall of the tubing or the syringes.
The mechanism used to mate the flow cell with the syringes or other fluid delivery device is also important both to fluid control and for eliminating air bubbles in flow channels. In our groups, three-way switching valves are mounted between the syringe pump and flow cell to prevent air bubbles from entering the flow chamber when syringes are removed for refilling or for buffer exchanges (Supplementary Fig. 1). Thereafter, it takes less than 10 min to reestablish stable, laminar flow conditions and resume measurements. Thus, several reaction conditions (as many as ten) can be studied in a single afternoon.
Single-stream flow cells
This design, with single inlet and outlet ports, is widely used and allows the study of a broad range of processes, using techniques including force measurements and fluorescence microscopy2,6,9,14,23,46. A single, 25- to 254-μm-deep channel is created using either two layers of Parafilm (127 μm)20,21,46,55 or one layer of double-sided (25 μm)2,6,56 or acrylic transfer (30 μm) tape23 sandwiched between a microscope slide and coverslip (Fig. 2 and Supplementary Table 1). The channel dimensions can be varied depending on the application and can be as wide as 5 mm and as long as 50 mm (refs. 55,56). To create flow within the channel, holes are drilled into the microscope slide at opposing ends of the channel and attached to tubing that is then connected to the fluid delivery system.
Figure 2 |.

A simple single-stream flow cell. Two different versions of a single channel design are shown. The separate versions are constructed using either double-sided2,6,55 or acrylic tape23 (green) or Parafilm20,46,56 (violet), resulting in channels ranging in depth from 25 to 254 μm. For the tape versions, the open ends can be sealed with epoxy, or alternatively, the channel can be cut out of the tape in a manner identical to that shown for the Parafilm design. The entry and exit ports are then connected to tubing either directly (attached using epoxy to bond directly to the glass slide)6 or through connectors. The purple arrow indicates the flow direction.
Adaptation for force measurements.
This design was developed for force measurements by the Bustamante group (Fig. 3 and Supplementary Table 1)46. It uses a slide and coverslip with a 254-μm-deep flow channel created with two layers of Parafilm. To permit access to the flow stream for a micropipette, an entry port is created perpendicular to the flow channel by placing between the two layers of Parafilm a quartz catheter tube having an inner diameter of 100 μm (Fig. 3, inset). Typically, micropipettes (pulled from glass capillaries to an inner diameter between 0.5 and 1 μm at the tip) are inserted through the catheter and into the chamber. The flow cell is connected to polyethylene tubing (280-μm internal diameter; Becton Dickinson) using flangeless fittings (Upchurch Scientific) and mated to the fluid delivery device. The flow rate is controlled using pressure, an automatic syringe pump (PSD/2; Hamilton) and a combination of computer-controlled solenoid valves (Clippard Instrument Lab)46. Other experimenters have advanced the design using silicon micromachining techniques to create a silicon-glass hybrid49. This design is reusable and has a smaller volume but is more expensive to produce than the Parafilm-sandwich version.
Figure 3 |.

A single-stream flow cell for force measurements. The flow cell is placed in a chamber holder (tan). Adapted from refs. 21,46. For clarity, the two Parafilm layers are shown as a single layer. One bead is held by the micropipette while the second is held in an optical trap (red, inset). Opposing ends of DNA molecules are attached to separate beads. 1, coverslip; 2, Parafilm layers; 3, microscope slide.
In these designs, each end of a single DNA molecule is sequentially attached to a different bead under flow and is simultaneously held in place by the micropipette and an optical trap, both of which are positioned in the flow channel ~100 μm from the surface (Fig. 3, inset). This is achieved by first flowing streptavidin-coated beads into the flow cell. Once the optical trap has captured a bead, the pipette is brought into close proximity and the bead transferred to its tip. Thereafter, a second bead is captured in the trap, followed by the introduction of DNA biotinylated at both ends. Once a DNA molecule is bound by the pipette bead, it is elongated by the fluid flow, so by moving the pipette relative to the optical trap, the free end of the DNA can be attached to the trap bead. This procedure has been automated so that complete setup from an empty chamber to a stably positioned DNA molecule takes ~40 min (ref. 46), with ~10 min required to initiate reactions or to alter conditions within the flow cell (G. Wuite, Vrije Universiteit Amsterdam, personal communication).
A water-immersion objective, which characteristically has a long working distance, is used to form the optical trap. This kind of objective is less prone to spherical aberration than oil-immersion objectives and can be used to trap objects deep within the flow-cell channel. DNA elasticity measurements can be performed in these flow cells using one of three operational modes: (i) force is applied to the bead held by the optical trap using just the optical trap; (ii) force is applied to this same bead using only laminar flow; or (iii) force is applied to this bead using both the optical trap and laminar flow46. Using any of these approaches, high positional accuracy can be achieved14,16,18,20,21,46.
Adaptation for fluorescence microscopy.
The single-stream design has also been successfully combined with fluorescence microscopy6,23,56. As micropipettes are not required in these experiments, shallower, sandwich-design flow cells can be constructed using either double-sided56 or acrylic transfer tape23. Several groups have combined this design with total internal reflectance fluorescence microscopy (TIRFM)9,23,55,56. This visualization method requires immobilization of biotinylated DNA to the flow cell surface after surface attachment of streptavidin either directly through nonspecific adsorption23 or indirectly through a biotin-PEG linker11,26. After DNA binding, unbound DNA molecules are washed out and replaced with an oxygen-scavenger solution11,26. Thereafter, DNA molecules tethered at one end to the modified microscope-slide surface are stretched at a small angle relative to the surface by fluid flow and reactions initiated9,23,55,56. To enable reaction visualization, either DNA, proteins or both can be fluorescently tagged.
To alter reaction conditions, fluid flow must be turned off, resulting in collapse of the DNA. Once flow is restored, the DNA is stretched again, allowing reaction visualization to resume. To provide more stability to tethered DNA molecules and to prevent their collapse, the free end of the tethered DNA molecule can be attached to a paramagnetic bead53,57. Applying a small magnetic force in combination with flow elevates the beads ~4 μm above the surface, maintaining the DNA in a stretched configuration independent of buffer flow.
Multistream flow cells
Multistream flow cells permit the sequential introduction of different biological components to an optically trapped and flow-stretched DNA molecule without the need for fluid exchange. This design was developed to visualize reactions that occur more rapidly than the time it takes to exchange fluid in a single-stream flow cell and to overcome problems associated with fluid exchanges.
Multistream flow cells combined with optical traps.
Several methods have been described for constructing multistream flow cells1,11,19,26,47. The first reports used a glass coverslip attached to a glass microscope slide in which a Y-shaped pattern of channels with entry and exit ports was formed using a ‘dry’ etching technique (Fig. 4 and Supplementary Table 1)1,19. A glass frit (powdered glass) is used to attach the coverslip to the microscope slide at 660 °C (MMR Technologies). To mate the flow cell to a syringe pump, microtight adapters are attached to the holes at the distal ends of the channels using epoxy tape (Ablestik).
Figure 4 |.
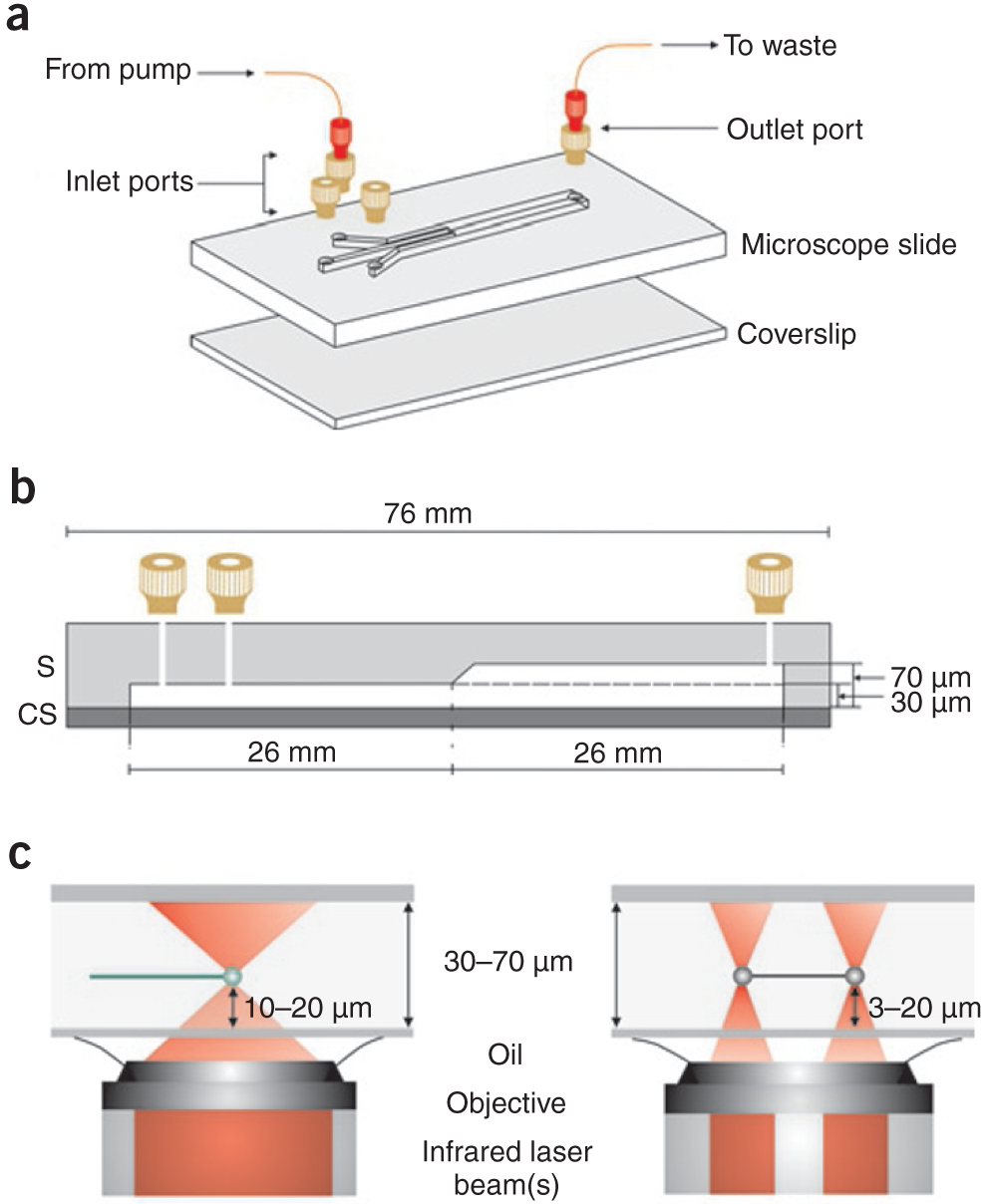
Multistream laminar flow cell for single-molecule studies. (a) A three-stream laminar flow cell constructed from glass8. The designis for an inverted microscope. Inlet channel widths are 250 μm and the common channel width is 750 μm. Machined connectors (tan; Upchurch Scientific) are attached to the slide at the holes. These are then connected to Fingertight connectors (red; Upchurch Scientific) to facilitate the attachment of PEEK tubing. Identical methods may be used to construct two-channel flow cells1,19. (A fully assembled flow cell positioned on a microscope stage is shown in Supplementary Fig. 1a.) (b) A side view of the flow cell in a showing relevant dimensions. (c) A side view of the flow cell demonstrating the trapping height from the coverslip using either one (left) or two (right) optical traps. Optical trapping and DNA visualization are carried out in the deeper portion of the flow cell in our groups.
This assembly technique works well because materials with different coefficients of thermal expansion are firmly attached to one another to create a robust and mechanically stable flow cell (Supplementary Fig. 1). By altering the design and channel layout, two to five entry channels 250 μm wide can be accommodated in a 75 × 25 mm slide. Fluid streams within the flow cell can be visualized using food coloring to assess the performance of newly fabricated devices (Supplementary Fig. 2 online). This allows a measure of the width and location of fluid streams within the single channel.
Techniques used to fabricate channels in glass or quartz are expensive and time-consuming and require specialized facilities. These materials are suitable for flow cells intended for use over an extended period of time (that is, >1 year). Successful multistream designs have also been fabricated using PDMS–glass42 and Parafilm–glass hybrids47. These devices are reusable and easily constructed with standard laboratory tools but suffer from limitations imposed by PDMS or Parafilm and are limited in the number of times that they can be reused.
The volume of fluid in a two-stream flow cell with two 250-μm-wide inlet channels and a 500-μm-wide common channel is ~1.5 μl, increasing to 2.3 μl in a three-stream design (a common channel 750 μm wide). The volume taken up by a 25-cm length of 500-μm internal-diameter PEEK tubing used to link each syringe in the syringe pump with the flow cell is ~60 μl. Syringes with 250 μl of liquid, when combined with flow rates of 10–100 μl/h, result in experiments that last for many hours under conditions of constant flow. To facilitate rapid delivery of fluid to the flow cell at the beginning of an experiment, the syringe pump is run at higher flow speeds (800 μl/h) and then ramped down in 50% increments to ‘trapping flow rates’ once fluid has reached the position of the optical traps. In experiments where reactions are visualized using fluorescence microscopy, linear flow velocities of 20–100 μm/s are used1,3,7,8,19,25,58,59.
In our groups, micromachined flow cells are held in place on a motorized microscope stage and connected to a single syringe pump through three-way switching valves (Supplementary Fig. 1). Asymmetrically biotinylated DNA molecules are attached to streptavidin-coated beads in solution before uptake into a syringe. Individual DNA-bead complexes are optically trapped in the first stream under constant flow by moving the position of the stage, so that the momentary position of a moving DNA-bead complex coincides with the position of the trap19,58,59. Alternatively, optical traps can be positioned at a fixed height above the coverslip surface and the experimentalist can simply wait for complexes to be trapped (a process which occurs every ~5 s)1,8.
Once a full-length DNA molecule is trapped, it is exposed to reaction components present in the adjacent fluid streams by translating the microscope stage perpendicular to the continuously flowing fluid to initiate reactions in a precise and controlled fashion (Fig. 5). If stage translation speed is 50 μm/s and the diffusion overlap between neighboring streams is 10 μm, it takes 0.5 s to move a phage λ DNA molecule across this diffusion boundary and completely into the adjacent stream. For a stream of width 200 μm, it takes 4 s to completely traverse it. Thus, conformational changes in DNA due to protein action that occur on a time scale of only a few seconds can be visualized and the DNA molecule can be rapidly exposed to several reaction conditions, as determined by the number of adjacent fluid streams present.
Figure 5 |.
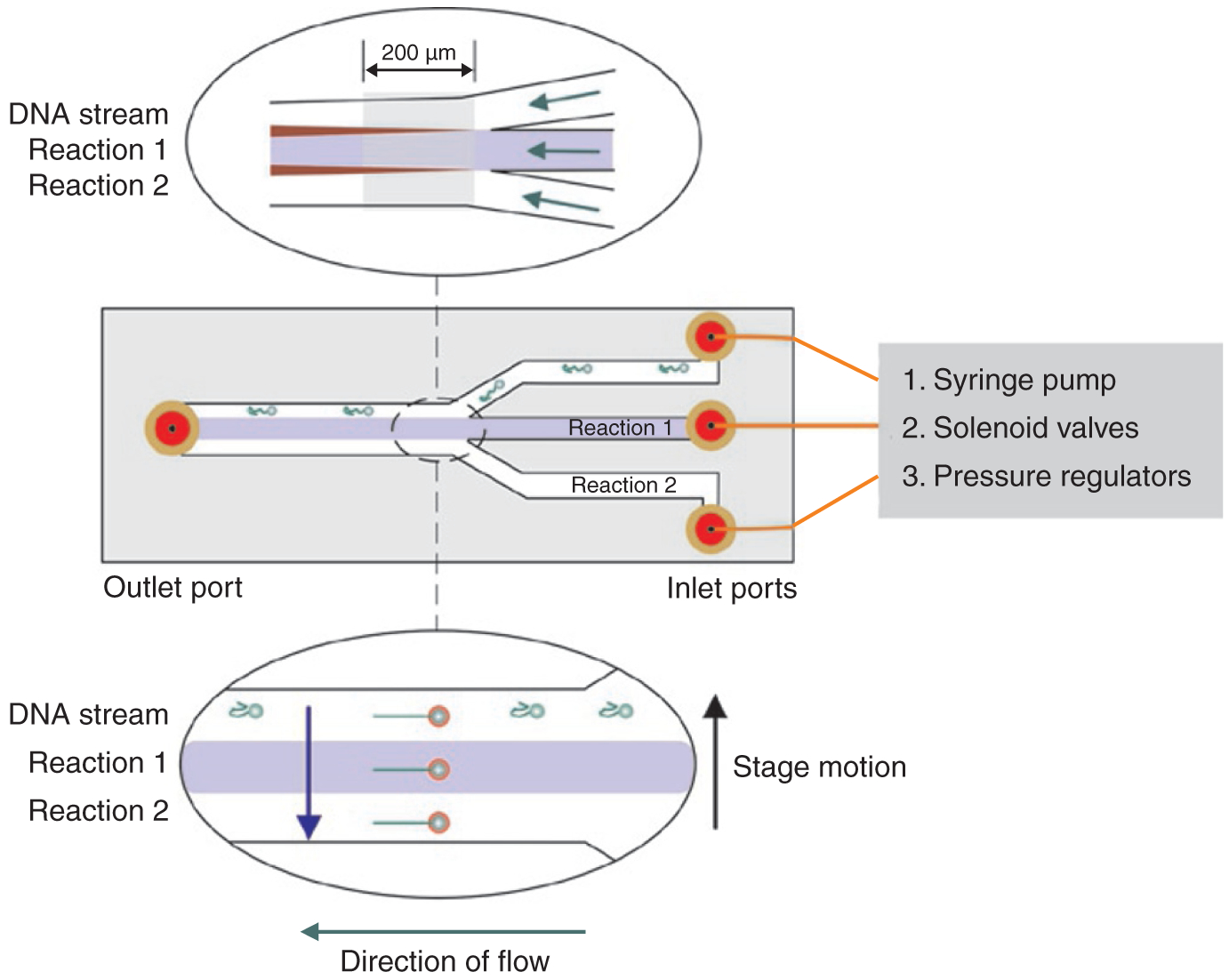
Multistream flow cells facilitate careful single-molecule experiments. A three-channel flow cell, viewed from the top, can be connected to a syringe pump8, solenoid valves (Clippard Europe)47 or pressure regulators42 to introduce buffers. The fluid stream in the middle channel is colored blue for clarity. DNA-bead complexes are introduced into the first channel, where they are optically trapped. Upper inset: the optical trap is positioned in the area (gray shaded box) within 200 μm of the flow junction, where transverse diffusion leading to mixing (maroon) is minimized (see Box 2). The flow paths are indicated by green arrows. Lower inset: an optically trapped DNA-bead complex is translated (blue arrow), perpendicular to the flow, from the DNA stream to the two reaction streams. Translation is facilitated by microscope stage motion (black arrow).
The multistream design combined with single optical traps has been successfully used to study DNA condensation19, dsDNA translocases7,8, DNA helicases1 and protein binding25. It has also been combined with two to four optical traps to study homologous recombination8 and for force measurements24,47. Here, DNA-protein complexes are assembled and trapped by four optical traps in the first few streams of the flow cell under continuous flow at rates of 100 μm/s (linear fluid velocity). Then they are moved into the last stream, where measurements are performed in the absence of flow47. Once a set of measurements is complete, flow is restored to remove interstream contamination that occurs in the absence of flow. The use of multiple optical traps with a multistream flow cell for force measurement24,47 negates potential physical drift that may occur between a micropipette and an optical trap in a single-stream flow cell46,60.
Other uses of multistream flow cells.
Recently, a multistream design was used for hydrodynamic focusing experiments in which interstream diffusion was used to achieve rapid mixing of reaction components11. This permitted measurement of conformational changes of T7 DNA polymerase with rates greater than 1 kHz. In this design, two independent pressure sources are used to push liquid from a central, 10-μm channel and two 40-μm side channels into a single, 40-μm-wide, 20-μm-deep observation channel at flow rates of 2.5 μl/min (linear speed of 75 μm/ms). By decreasing the pressure on the central fluid stream relative to the side streams, a 300-nm-wide stream is produced in the center of the observation channel. This allows mixing between the side and center streams within the observation channel to occur within 50 μs. Kinetic information can be obtained through fluorescence measurements taken at different positions along the flow cell from the point of mixing.
Stopping and starting the flow in single-stream flow cells can cause difficulties that plague TIRFM experiments using stretched DNA. To overcome this problem, the Mizuuchi group developed the laminar boundary-steering flow cell26. This design uses three entry channels that converge into a single physical channel (Fig. 6a and Supplementary Table 1). The entry port to each channel is attached to a separate syringe pump, with each syringe containing a different solution. The three resulting fluid streams within the common channel are separated by laminar boundaries whose positions can be shifted by altering pump flow rates to allow one of the inlets to flow more rapidly into the observation channel (Fig. 6b). Of key importance, the combined flow rate is held constant by simultaneously reducing the flow rates for the remaining channels. Using this approach, a seamless switching of reaction conditions is rapidly achieved within a matter of seconds with minimal disturbance to the DNA attached to the microscope slide surface. This design was successfully used to study MuB-DNA dynamics26. Because one fluid stream predominates within this single channel at any one time, operationally it functions like a single-stream design even though it uses more than one stream.
Figure 6 |.
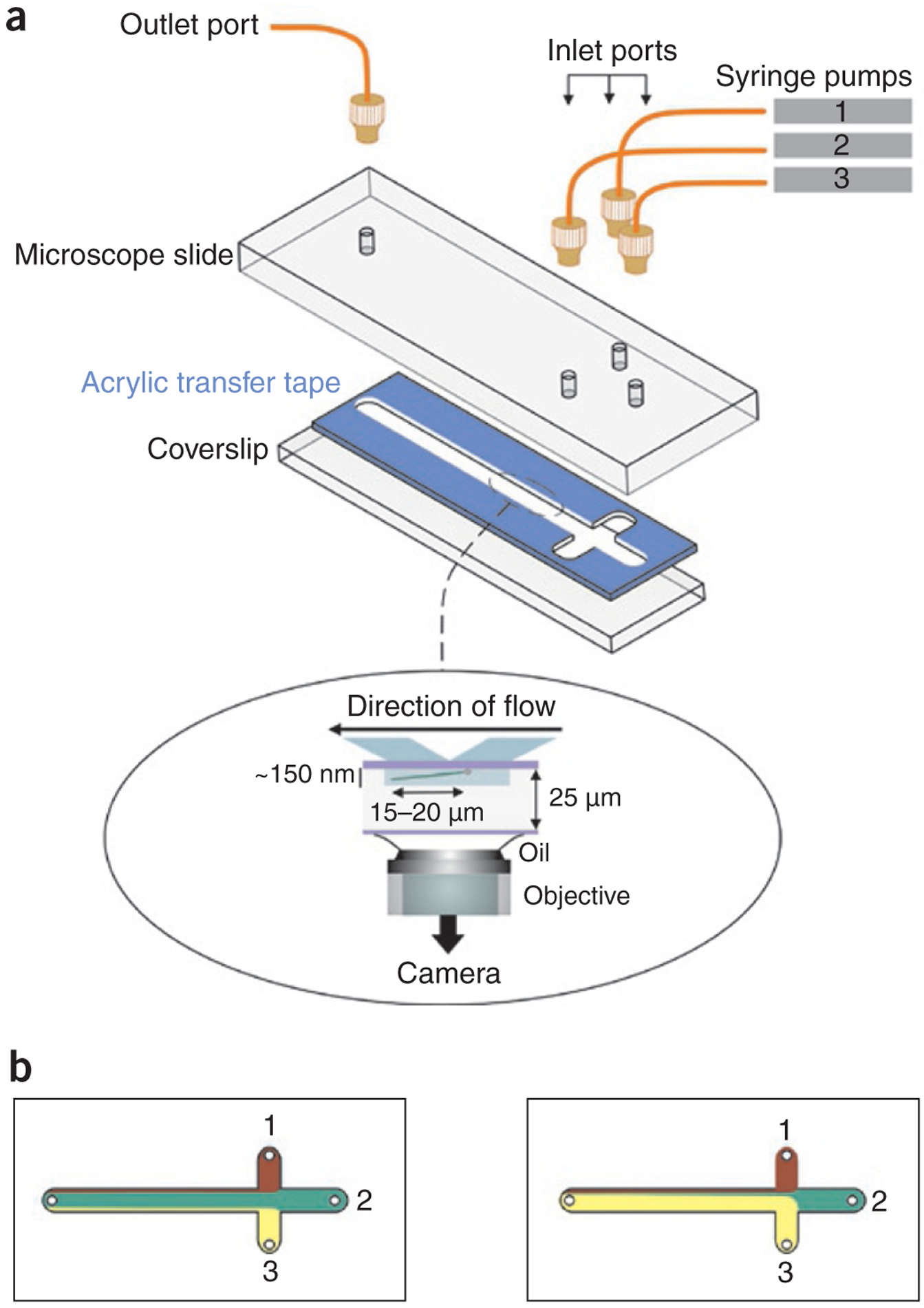
A laminar boundary-steering flow cell for TIRFM. (a) The flow cell design and assembly26. The width of all channels is 3 mm. Nanoport connectors (tan) are used to secure PEEK tubing to the flow cell. Inset, side view of flow cell being used for TIRFM26. A single DNA molecule (green) attached to the surface through streptavidin (sphere) is shown stretched by the constant flow. The DNA is visible within the evanescent wave (~150 nm depth; gray rectangle). (b) Laminar flow boundaries are steered by altering flow rates of individual pumps while maintaining the net flow rate constant. The numbers indicate the three syringe pumps used to control flow. The colors indicate different buffer solutions. Left, the dominant pump is syringe pump 2; right, the dominant pump is pump 3. Panel b adapted from ref. 26.
Selecting an appropriate flow cell
Although there are many possibilities for flow cell designs27,28,37, only a few have become widely used in the study of DNA-protein interactions (Supplementary Table 1). What factors determine which flow cell is selected? Initial experiments while a system is being developed could take advantage of sandwich designs assembled from Parafilm or tape. Construction of these flow cells does not require microfabrication facilities, and different channel configurations can be easily cut into the channel-forming material using a razor blade47 or laser cutting6,26. Nanoport connectors (Upchurch Scientific) can be bonded to the inlet and outlet ports, PEEK tubing attached and the assembled flow cell mated to the fluid delivery device. In addition to initial experiments, these designs are suitable for fluorescence microscopy6,9,23 and for force measurements using optical tweezers2,47. If force measurements are pursued using the Bustamante approach, then a thicker channel-forming material must be used to accommodate the micropipette46.
The reusability of flow cells is an important issue because most experiments use DNA, beads, fluorescent dyes and protein. These must be removed from the flow cell surfaces to prevent contamination of subsequent experiments and to reduce fluorescence background. Removal requires rigorous cleaning using denaturants. Sandwich materials may not be compatible with rigorous cleaning, thereby limiting the number of times a flow cell can be used. Most TIRFM experiments use surface-tethered reaction components, and it is critical to avoid unintended adsorption of fluorophores to the flow cell surface. Used TIRFM flow cells are typically discarded or are disassembled with the connectors, slide and coverslip being reused (K. Mizuuchi, US National Institute of Diabetes and Digestive and Kidney Diseases, personal communication). If it is difficult to manufacture, replace and realign disposable flow cells, reusable flow cell designs must be considered. Single- or multistream designs manufactured from glass or quartz are chemically inert and can be rigorously cleaned at the end of each experiment, allowing long-term use1,8,19. Repeatedly using a single flow cell eliminates the complication of having to frequently remake flow cells.
An important issue to consider when selecting a flow cell design is the rate of the reaction being investigated. Because of the need to replace the solution in the entire channel in single-stream designs, the reactions to be studied must induce changes in the DNA molecule slowly compared to the time it takes to fill the flow cell with a given reaction component. Multistream flow cells allow an optically trapped DNA molecule to be rapidly exposed to several biological components sequentially for experiments that take less than 1–2 min to complete1,8,58,59. For experiments that require measurements of reactions with rates ≥1 kHz, hydrodynamic-focusing flow cells are necessary.
Cost-effective multistream designs providing stable flow can be constructed from Parafilm or tape26,47. Although the laminar boundary-steering flow cell is operated like a single-fluid-stream design, it can be also serve as a limited-use multistream flow cell. To create parallel streams, fluid velocity in each channel must be identical. Here, the flow cell can be connected either to three separate syringe pumps, with each operating at an identical pump rate, or to a single syringe pump housing three syringes.
Laminar flow cells provide a carefully controlled environment for performing single-molecule studies. A number of beautiful studies have already been done using a variety of flow cells designs, and as time progresses the number should continue to increase.
Supplementary Material
ACKNOWLEDGMENTS
We would like to thank C. Bustamante, Kiyoshi Mizuuchi, G. Wuite, H. Gao and M. Gao for critical reading of the manuscript. We would also like to thank K. Mizuuchi and G. Wuite for sharing unpublished flow cell parameters and flow velocities. Funding for the experimental work in the Brewer group was provided by US National Institutes of Health grant HD01387 to L.R.B. Funding for the experimental work in the Bianco group was from US National Institutes of Health grant GM66831 and Susan G. Komen Breast Cancer Foundation grant BCTR0601350 to P.R.B.
Footnotes
Note: Supplementary information is available on the Nature Methods website.
Published online at http://www.nature.com/naturemethods/
References
- 1.Bianco PR et al. Processive translocation and DNA unwinding by individual RecBCD enzyme molecules. Nature 409, 374–378 (2001). [DOI] [PubMed] [Google Scholar]
- 2.Perkins TT et al. Forward and reverse motion of single RecBCD molecules on DNA. Biophys. J 86, 1640–1648 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Handa N, Bianco PR, Baskin RJ & Kowalczykowski S Direct visualization of RecBCD movement reveals cotranslocation of the RecD motor after χ recognition. Mol. Cell 17, 745–750 (2005). [DOI] [PubMed] [Google Scholar]
- 4.Spies M et al. A molecular throttle: the recombination hotspot chi controls DNA translocation by the RecBCD helicase. Cell 114, 647–654 (2003). [DOI] [PubMed] [Google Scholar]
- 5.Chemla YR et al. Mechanism of force generation of a viral DNA packaging motor. Cell 122, 683–692 (2005). [DOI] [PubMed] [Google Scholar]
- 6.Grayson P, Han L, Winther T & Phillips R Real-time observations of single bacteriophage lambda DNA ejections in vitro. Proc. Natl. Acad. Sci. USA 104, 14652–14657 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Amitani I, Baskin RJ & Kowalczykowski SC Visualization of Rad54, a chromatin remodeling protein, translocating on single DNA molecules. Mol. Cell 23, 143–148 (2006). [DOI] [PubMed] [Google Scholar]
- 8.Bianco PR, Bradfield JJ, Castanza LR & Donnelly AN Rad54 oligomers translocate and cross-bridge double-stranded DNA to stimulate synapsis. J. Mol. Biol 374, 618–640 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.van Oijen AM et al. Single-molecule kinetics of lambda exonuclease reveal base dependence and dynamic disorder. Science 301, 1235–1238 (2003). [DOI] [PubMed] [Google Scholar]
- 10.Wuite GJL, Smith SB, Young M, Keller D & Bustamante C Single-molecule studies of the effect of template tension on T7 DNA polymerase activity. Nature 404, 103–106 (2000). [DOI] [PubMed] [Google Scholar]
- 11.Luo G, Wang M, Konigsberg WH & Xie XS Single-molecule and ensemble fluorescence assays for a functionally important conformational change in T7 DNA polymerase. Proc. Natl. Acad. Sci. USA 104, 12610–12615 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tanner NA et al. Single-molecule studies of fork dynamics in Escherichia coli DNA replication. Nat. Struct. Mol. Biol 15, 170–176 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Harada Y et al. Single-molecule imaging of RNA polymerase-DNA interactions in real time. Biophys. J 76, 709–715 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Davenport RJ, Wuite GJ, Landick R & Bustamante C Single-molecule study of transcriptional pausing and arrest by E. coli RNA polymerase. Science 287, 2497–2500 (2000). [DOI] [PubMed] [Google Scholar]
- 15.Cheng W, Dumont S, Tinoco I Jr. & Bustamante C NS3 helicase actively separates RNA strands and senses sequence barriers ahead of the opening fork. Proc. Natl. Acad. Sci. USA 104, 13954–13959 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dumont S et al. RNA translocation and unwinding mechanism of HCV NS3 helicase and its coordination by ATP. Nature 439, 105–108 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shaqfeh ES The dynamics of single-molecule DNA in flow. J. Non-Newt. Fluid Mech 130, 1–28 (2005). [Google Scholar]
- 18.Bustamante C Unfolding single RNA molecules: bridging the gap between equilibrium and non-equilibrium statistical thermodynamics. Q. Rev. Biophys 38, 291–301 (2005). [DOI] [PubMed] [Google Scholar]
- 19.Brewer LR, Corzett M & Balhorn R Protamine-induced condensation and decondensation of the same DNA molecule. Science 286, 120–123 (1999). [DOI] [PubMed] [Google Scholar]
- 20.Bennink ML et al. Single-molecule manipulation of double-stranded DNA using optical tweezers: interaction studies of DNA with RecA and YOYO-1. Cytometry 36, 200–208 (1999). [DOI] [PubMed] [Google Scholar]
- 21.Bennink ML et al. Unfolding individual nucleosomes by stretching single chromatin fibers with optical tweezers. Nat. Struct. Biol 8, 606–610 (2001). [DOI] [PubMed] [Google Scholar]
- 22.Bustamante C, Bryant Z & Smith SB Ten years of tension: single-molecule DNA mechanics. Nature 421, 423–427 (2003). [DOI] [PubMed] [Google Scholar]
- 23.Greene EC & Mizuuchi K Direct observation of single MuB polymers: evidence for a DNA-dependent conformational change for generating an active target complex. Mol. Cell 9, 1079–1089 (2002). [DOI] [PubMed] [Google Scholar]
- 24.Dame RT, Noom MC & Wuite GJ Bacterial chromatin organization by H-NS protein unravelled using dual DNA manipulation. Nature 444, 387–390 (2006). [DOI] [PubMed] [Google Scholar]
- 25.Galletto R, Amitani I, Baskin RJ & Kowalczykowski SC Direct observation of individual RecA filaments assembling on single DNA molecules. Nature 443, 875–878 (2006). [DOI] [PubMed] [Google Scholar]
- 26.Tan X, Mizuuchi M & Mizuuchi K DNA transposition target immunity and the determinants of the MuB distribution patterns on DNA. Proc. Natl. Acad. Sci. USA 104, 13925–13929 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Squires TM & Quake SR Microfluidics: fluid physics at the nanoliter scale. Rev. Mod. Phys 77, 977–1026 (2005). [Google Scholar]
- 28.Weibel DB & Whitesides GM Applications of microfluidics in chemical biology. Curr. Opin. Chem. Biol 10, 584–591 (2006). [DOI] [PubMed] [Google Scholar]
- 29.Levin S & Tawil G Effect of surfactants on the diffusion coefficients of proteins, measured by analytical SPLITT fractionation (ASF) in the diffusion mode. J. Pharm. Biomed. Anal 12, 499–507 (1994). [DOI] [PubMed] [Google Scholar]
- 30.Fuh CB, Levin S & Giddings JC Rapid diffusion coefficient measurements using analytical SPLITT fractionation: application to proteins. Anal. Biochem 208, 80–87 (1993). [DOI] [PubMed] [Google Scholar]
- 31.Ismagilov RF, Stroock AD, Kenis PJA, Whitesides GM & Stone HA Experimental and theoretical scaling laws for transverse diffusive broadening in two-phase laminar flows in microchannels. Appl. Phys. Lett 76, 2376–2378 (2000). [Google Scholar]
- 32.Kamholz AE & Yager P Theoretical analysis of molecular diffusion in pressure-driven laminar flow in microfluidic channels. Biophys. J 80, 155–160 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kamholz AE, Schilling EA & Yager P Optical measurement of transverse molecular diffusion in a microchannel. Biophys. J 80, 1967–1972 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lima R, Wada S, Takeda M, Tsubota K & Yamaguchi T In vitro confocal micro-PIV measurements of blood flow in a square microchannel: the effect of the haematocrit on instantaneous velocity profiles. J. Biomech 40, 2752–2757 (2007). [DOI] [PubMed] [Google Scholar]
- 35.Hong JW & Quake SR Integrated nanoliter systems. Nat. Biotechnol 21, 1179–1183 (2003). [DOI] [PubMed] [Google Scholar]
- 36.Psaltis D, Quake SR & Yang C Developing optofluidic technology through the fusion of microfluidics and optics. Nature 442, 381–386 (2006). [DOI] [PubMed] [Google Scholar]
- 37.Weibel DB, Diluzio WR & Whitesides GM Microfabrication meets microbiology. Nat. Rev. Microbiol 5, 209–218 (2007). [DOI] [PubMed] [Google Scholar]
- 38.Wabuyele MB, Ford SM, Stryjewski W, Barrow J & Soper SA Single molecule detection of double-stranded DNA in poly(methylmethacrylate) and polycarbonate microfluidic devices. Electrophoresis 22, 3939–3948 (2001). [DOI] [PubMed] [Google Scholar]
- 39.McDonald JC & Whitesides GM Poly(dimethylsiloxane) as a material for fabricating microfluidic devices. Acc. Chem. Res 35, 491–499 (2002). [DOI] [PubMed] [Google Scholar]
- 40.Qi S et al. Microfluidic devices fabricated in poly(methyl methacrylate) using hot-embossing with integrated sampling capillary and fiber optics for fluorescence detection. Lab Chip 2, 88–95 (2002). [DOI] [PubMed] [Google Scholar]
- 41.Ladoux B, Quivy JP, Doyle PS, Almouzni G & Viovy JL Direct imaging of single-molecules: from dynamics of a single DNA chain to the study of complex DNA-protein interactions. Sci. Prog 84, 267–290 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Merenda F et al. Refractive multiple optical tweezers for parallel biochemical analysis in micro-fluidics in Proc. SPIE, 6483, (eds. Andrews DL, Galves EJ & Nienhuis G) 64830A (SPIE, Bellingham, Washington, USA, 2007). [Google Scholar]
- 43.Quake SR & Scherer A From micro- to nanofabrication with soft materials. Science 290, 1536–1540 (2000). [DOI] [PubMed] [Google Scholar]
- 44.De Jong J, Lammertink RG & Wessling M Membranes and microfluidics: a review. Lab Chip 6, 1125–1139 (2006). [DOI] [PubMed] [Google Scholar]
- 45.Ng JM, Gitlin I, Stroock AD & Whitesides GM Components for integrated poly(dimethylsiloxane) microfluidic systems. Electrophoresis 23, 3461–3473 (2002). [DOI] [PubMed] [Google Scholar]
- 46.Wuite GJL, Davenport RJ, Rappaport A & Bustamante C An integrated laser trap/flow control video microscope for the study of single biomolecules. Biophys. J 79, 1155–1167 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Noom MC, van den Broek B, van Mameren J & Wuite GJL Visualizing single DNA-bound proteins using DNA as a scanning probe. Nat. Methods 4, 1031–1036 (2007). [DOI] [PubMed] [Google Scholar]
- 48.Mijatovic D, Eijkel JC & van den Berg A Technologies for nanofluidic systems: top-down vs. bottom-up—a review. Lab Chip 5, 492–500 (2005). [DOI] [PubMed] [Google Scholar]
- 49.Rusu C et al. Direct integration of micromachined pipettes in a flow channel for single DNA molecule study by optical tweezers. J. Microelectromech. Syst 10, 238–244 (2001). [Google Scholar]
- 50.Takayama S et al. Patterning cells and their environments using multiple laminar fluid flows in capillary networks. Proc. Natl. Acad. Sci. USA 96, 5545–5548 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kenis PJ et al. Fabrication inside microchannels using fluid flow. Acc. Chem. Res 33, 841–847 (2000). [DOI] [PubMed] [Google Scholar]
- 52.Sia SK & Whitesides GM Microfluidic devices fabricated in poly(dimethylsiloxane) for biological studies. Electrophoresis 24, 3563–3576 (2003). [DOI] [PubMed] [Google Scholar]
- 53.Kim S, Blainey PC, Schroeder CM & Xie XS Multiplexed single-molecule assay for enzymatic activity on flow-stretched DNA. Nat. Methods 4, 397–399 (2007). [DOI] [PubMed] [Google Scholar]
- 54.Tanaka H, Ishijima A, Honda M, Saito K & Yanagida T Orientation dependence of displacements by a single one-headed myosin relative to the actin filament. Biophys. J 75, 1886–1894 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Leuba SH, Bennink ML & Zlatanova J Single-molecule analysis of chromatin. Methods Enzymol. 376, 73–105 (2004). [DOI] [PubMed] [Google Scholar]
- 56.Zheng H, Tomschik M, Zlatanova J & Leuba SH Evanescent field fluorescence microscopy for analysis of protein/DNA interactions at the single-molecule level in Protein Protein Interactions, a Molecular Cloning Manual. (eds. Golemis E & Adams P) 429–444 (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA, 2005). [Google Scholar]
- 57.Lee JB et al. DNA primase acts as a molecular brake in DNA replication. Nature 439, 621–624 (2006). [DOI] [PubMed] [Google Scholar]
- 58.Brewer L, Corzett M & Balhorn R Condensation of DNA by spermatid basic nuclear proteins. J. Biol. Chem 277, 38895–38900 (2002). [DOI] [PubMed] [Google Scholar]
- 59.Brewer L, Corzett M, Lau EY & Balhorn R Dynamics of protamine 1 binding to single DNA molecules. J. Biol. Chem 278, 42403–42408 (2003). [DOI] [PubMed] [Google Scholar]
- 60.Moffitt JR, Chemla YR, Izhaky D & Bustamante C Differential detection of dual traps improves the spatial resolution of optical tweezers. Proc. Natl. Acad. Sci. USA 103, 9006–9011 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.