

Abstract
Fucoidans are multifunctional marine macromolecules that are subjected to numerous and various downstream processes during their production. These processes were considered the most important abiotic factors affecting fucoidan chemical skeletons, quality, physicochemical properties, biological properties and industrial applications. Since a universal protocol for fucoidans production has not been established yet, all the currently used processes were presented and justified. The current article complements our previous articles in the fucoidans field, provides an updated overview regarding the different downstream processes, including pre-treatment, extraction, purification and enzymatic modification processes, and shows the recent non-traditional applications of fucoidans in relation to their characters.
Keywords: fucoidans, extraction, brown algae, production, bioactivities
1. Introduction
Polysaccharides, nucleic acids, and peptides are considered the main three types of bioactive polymeric macromolecules [1]. Among these, polysaccharides serve various roles in living cells including structural functions, where cellulose and chitin represent the major components of the different cell wall matrices [2,3], energy storage (e.g., starch and glycogen) [4,5], and hydration and signaling functions (e.g., mucilage and alginic acid) [6,7].
Particularly, marine homo- and heteropolysaccharides are derived from marine organisms, which represent a large part of global biodiversity [8]. Among these are the algal polysaccharides, such as fucoidan and alginate in brown seaweeds, carrageenan in red seaweeds and ulvan in green seaweeds. These were reported to have interesting nutraceutical, biomedical, pharmaceutical and cosmeceutical applications, including dietary fibers; anti-inflammatory, anti-tumor, anti-oxidant, hepatoprotective and anti-coagulant properties; and drug carrier functionality. Therefore, they have been extensively investigated during the last few decades [9,10,11,12,13], especially after the emergence of glycobiology and glycomics [14,15,16,17].
Polysaccharides such as dietary fibers of brown algae are abundant and diverse (e.g., alginates, cellulose, fucoidans and laminarins) constituting the major components (up to 75%) of the dried thallus weight (% DW) [18,19,20]. Previous work investigated their abundance in different species, reporting Fucus, Ascophyllum, Saccharina, and Sargassum to contain 65.7, 69.6, 57.8 and 67.8 % DW, respectively [21,22]. Specifically, fucoidans are found in the cell walls and extracellular matrices of brown algae in addition to more than 265 genera and 2040 species of marine invertebrates (e.g., sea cucumbers), where they perform vital structural functions [23,24,25,26]. Fucoidans are assumed to act as cross-linkers between the major threads of cellulose and hemicellulose, promoting cellular integrity and maintaining cellular hydration (especially during drought seasons) [27]. They also act in other reproductive, immune and cell-to-cell communicative roles [23]. As recommended by the International Union of Pure and Applied Chemistry (IUPAC), fucoidans is a general term used to describe sulfated L-fucose-based polymers including sulfated fucans cited by the Swedish scholar Kylin, as well as other fucose-rich sulfated heteropolysaccharides [23,28]. Their chemical structures, in terms of monomeric composition and branching, are quite simple in marine invertebrates compared to their analogues in brown algae [13,29].
Hundreds of articles have thoroughly discussed and reviewed the biological, pharmacological and pharmaceutical applications of fucoidans [30,31,32,33], including nanomedicine, [34] which has made it a hot topic in the last few decades [35,36,37]. All these studies tried to investigate fucoidans molecular mechanisms in relation to their chemical structure and physicochemical properties. Therefore, different hypotheses were suggested for each activity, such as anti-tumor [31,38,39,40], anti-coagulant [41,42], anti-viral [43,44] and anti-inflammatory activity[45,46]. These investigations revealed that various factors are relevant, such as molecular weight, sulfation pattern, sulfate content and monomeric composition [47,48,49]. For example, different fractions were produced with different physicochemical properties in our previous experiments; sulfation pattern and sulfate content were highly related to anti-viral and cytotoxic activities against HSV-1 and Caco-2 cell lines, respectively, while molecular weight and sugar composition were potential factors in anti-coagulation activity [41,50]. In addition, degree of purity was reported as an influential factor [32], where co-extracted contaminants (e.g., phlorotannins or polyphenols) could lead to significant interference in anti-oxidant activity and, consequently, cosmetic applications [51,52].
Therefore, several key production challenges regarding fucoidans were discussed in our last review article in order to obtain a product that follows the universal good manufactured practice (GMP) guidelines. The article discussed sources of heterogeneity in extracted fucoidans, including the different biotic (e.g., biogenic, geographical and seasonal factors) and abiotic (e.g., downstream processes) factors affecting the fucoidans physicochemical and chemical properties [53]. Others patented production techniques that have assisted in the marketing of several commercial fucoidans by well-known companies (e.g., Sigma-Aldrich®, Algues and Mer and Marinova®) derived from Fucus vesiculosus and other brown algae species [54,55,56].
Furthermore, the improvement of fucoidans activity was investigated, targeting several points. Among these was the modification of the chemical structure of the native fucoidans scaffolding, including depolymerization [57,58] and over-sulfation [59]. These modifications could be attempted chemically [60], enzymatically [35,61] or physically [62]. Predetermined synthesis of oligomers [63,64] and low molecular weight polymers with defined monomeric units [65] is also involved. Additionally, fractionation of fucoidans is a common approach during extraction and purifications steps by applying different extraction and purification conditions (e.g., pH, time, molarity of NaCl) [49,55].
The current article aimed at complementing our previously published article discussing the reasons for heterogeneity of fucoidans [53]. It reviewed and evaluated the different downstream processes used in production as the most important abiotic factors affecting the fucoidans quality and structural features; it then addressed recent uncommon applications and prospective bioproduction of fucoidans. In addition, the updated status of enzymatic structural modifications of fucoidans, especially by fucoidanases, were presented.
2. Global Market and Cultivation of Brown Algae
Marine hydrocolloids (e.g., agar, carrageenan and alginate) are of particular industrial interest, with worldwide annual production of approx. 100,000 tons and a value above US $1.1 billion [66]. Based on FAO periodical reports (FAO, 2014, 2016), among the top seven most-cultivated seaweeds, three taxa are mainly used for hydrocolloids production; these include Rhodophyta (e.g., Eucheuma sp. and Kappaphycus alvarezii) for carrageenan production and Gracilaria sp. for agar production [67]. These data encouraged the global marine market to escalate the production yield by finding alternative, eco-friendly seaweed cultivation techniques, such as sea farming or aquaculture and biotechnology [53]. In 2014, the annual production of cultivated seaweeds reached 27.3 million tons [68], representing 27% of the total marine aquaculture production, while the global market of marine biotechnology (blue biotechnology) for industrial applications has been expected to achieve US $4.8 billion in 2020 and grow to US $6.4 billion by 2025 [69].
Species of brown macroalgae (Phaeophyceae) are distributed among the orders Fucales and Laminariales, which are the major commercial sources of the algal sulfated polysaccharides, in addition to Chordariales, Dictyotales, Dictyosiphonales, Ectocarpales, and Scytosiphonales. Moreover, phylogenetic analysis showed that Fucales are one of the largest and most diversified orders within Phaeophyceae, having eight families (41 genera and 485 species), named Ascoseiraceae, Cystoseiraceae, Durvillaeaceae, Fucaceae, Hormosiraceae, Himanthaliaceae, Sargassaceae, and Seirococcaceae [70]. Figure 1 illustrates the distribution of several examples of well-known brown algae species which are considered potential sources of sulfated polysaccharides dominating tropical to temperate marine forests and intertidal regions. The data were based on Wahl, et al. [71].
Figure 1.

Global distribution of the major brown seaweeds’ species. They dominate tropical to temperate marine forests and intertidal regions.
Furthermore, like terrestrial plant tissue culture (PTC), several biotechnological attempts were performed to cultivate and/or regenerate thallus from different species of brown seaweeds using seaweeds tissue culture [72]. They include micropropagation, callus induction and protoplast isolation [69,73,74,75]. They are very promising techniques as it may not only help to overcome the previously mentioned fucoidans production heterogeneity challenges [53] but also provide a sustainable supply [76]. However, compared to PTC, STC is still not well-enough established to be used for production of hydrocolloids and fucoidans [77] or cultivation in closed, well-controlled bioreactors, as in case of the red algae organism Agardhiella subulata [78].
3. Downstream Processes
Fucoidans are anionic polymers occurring in highly complicated matrices in cell walls and intercellular spaces along with other carbohydrate polymers (e.g., alginate, cellulose and laminarin), polyphenols and proteins [79]. Additionally, due to the sulfate ester groups, fucoidans are water soluble polysaccharide polymers [80] exhibiting high affinity to other cell wall components, especially polyphenols [81]. Therefore, various and complicated downstream processes are required to remove such extraneous substances before and after precipitation with ethanol or cationic surfactants to obtain high-purity fucoidans [82,83]. The processes always include pre-treatment, extraction and purification stages as shown in Figure 2.
Figure 2.

Required downstream processes including steps in each process for fucoidans production.
3.1. Pre-Treatment
After harvesting algal biomass from beaches, the biomass should be washed thoroughly with tap water to remove sands and epiphytes, then dried and milled to increase the area-to-mass ratio. Several pre-treatment steps are performed before the extraction step to release fucoidans from intercalating components, ease the following extraction process, improve the extraction yield, and decrease the possible interferences from co-extracted components in purification and biological investigations.
Previous experiments tried to remove pigments (e.g., chlorophyll, flavins and carotenoids) and lipids in specific bleaching and defatting steps with acetone, toluene, charcoal or 80%–85% (v/v) ethanol [34,84,85]. Since fucoidans are negatively charged molecules, they remained unaffected by incubation with organic solvents (e.g., acetone, toluene or hexane:isopropanol (3:2) mixture) during pre-treatment of the dried algal biomass. Such extracts were further treated to obtain carotenoids, represented by fucoxanthin in brown algae [86], lipids and fatty acid metabolites (especially essential polyunsaturated fatty acids (PUFA) and fucosterol), adding to nutraceutical applications of brown algae [87,88]. In contrast, activated carbon materials, such as charcoal, adsorb the target fucoidans molecules, adversely affecting the final production yield [79].
Other studies tried to exclude the tightly non-covalently bound polyphenolic compounds represented by phloroglucinol-type phlorotannins [89], which contribute to the light to dark brown color of the crude fucoidans extract (along with fucoxanthin) [41,81]. They reported comparatively high phlorotannins content, reaching approximately one fifth of the brown algae dry weight [25]. Phlorotannins perform major structural and physiological functions, like tannins found in plants, including defense against biotic and abiotic stresses [90,91]. Despite of the great pharmacological importance of phlorotannins [92,93], their presence in high-quality fucoidans is not acceptable because of the possibility of interference with the anti-oxidant [25,52,94] and anti-tumor activities of fucoidans [95]. Therefore, the natural phenolics content of fucoidans should be determined before the measurement of their biological activities [96]. Therefore, nearly all pre-extraction protocols for fucoidans involved strategies to remove such contaminants, e.g., incubation with EtOH:H2O:HCHO (16:3:1) (v/v/v) at pH 2. Under such conditions, formaldehyde enhances the crosslinking and polymerization of such polyphenolic contaminants and the high volume of ethanol results in protein denaturation [41,60,97,98]. However, the toxicity of formaldehyde limits its utilization in pre-treatment protocols [51].
Furthermore, pre-treatment steps are performed to remove other carbohydrates such as alginate, the major hydrocolloids in brown algae [99]. This is commonly removed by formation of water-insoluble calcium complex either before [60] or during the extraction procedure using 1%–4% (w/v) CaCl2 followed by a filtration or centrifugation step to remove the formed precipitate [58,98,100,101]. These previously mentioned procedures were optimized using successive incubation, centrifugation or filtration, washing and drying for the main extraction step of the dried, milled algal biomass, as described in Figure 3. The application of such an optimized protocol resulted in a dried, pre-treated powder representing 71% (w/w) of the starting material [98]. Despite these results, downstream processing of fucoidans, except with enzymatic modification, starts with a small scale (e.g., 5–10 g of the dried algal biomass) to optimize parameters like dried biomass to solvent ratio, temperatures, pH, and incubation time, based on preliminary quality and yield of crude fucoidans measured by infra-red spectroscopy (IR), simple sugar tests and elemental analysis. After this, transfer to large scale production could be accomplished using larger biomass quantities (e.g., 500–1000 g).
Figure 3.
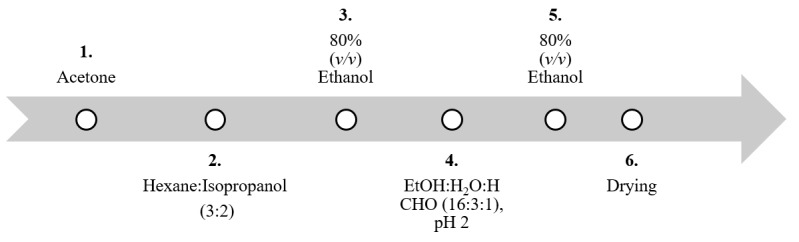
Overview of optimized pre-treatment steps of the dried algae biomass before fucoidans extraction. All steps were performed at 25 °C overnight and the ratio between dried algal biomass to solvent was 1:10, except for the acetone step, which was 1:20 (modified after [98,102]).
Due to several complicated pre-treatment steps, general protocols always employ a single incubation step using the ternary mixture composed of CH3OH:CH3Cl:H2O (4:2:1) (v/v/v) [103], binary mixture of CH2Cl2:EtOH (94.2:5.8, v/v) [104], or aqueous ethanol (e.g., 95% v/v) [105,106] to remove pigments [107], polyphenols [51,103] and lipids [108]. Nevertheless, pre-treatment steps may be insufficient for complete removal or prevention of some residual co-extraction.
Notably, all these procedures were carried out at room temperature in organic solvents and high volumes of ethanol, in which fucoidans are insoluble. Theoretically, the native structural backbone should not be affected. However, similar polymeric carbohydrates such as laminarin may still be present, contaminating the extract after these steps.
Recently, in order to decrease pollution of organic toxic solvents, compressional-puffing pre-treatment was applied for Sargassum hemiphyllum and S. glaucescens fucoidans. The pre-treatment method was based on mechanical pressure at higher temperatures that loosen the cell wall matrix before the step of extraction. Such methods succeeded in increasing the production yield, but they affected the molecular features of the fucoidans, including molecular weight [109,110].
3.2. Extraction
As previously mentioned, fucoidans are principally anionic water-soluble macromolecules. Therefore, they can be extracted from the pre-treated biomass using a simple hot- or cold-water incubation. Afterwards, the extracted fucoidans can be precipitated by high volumes of solvents with a low dielectric constant (e.g., >70% (v/v), > 2.5 volume ethanol [111,112], <2 volume acetone [113]) or cationic surfactants (e.g., hexadecyltrimethylammonium bromide (Cetavlon®) 10% (v/v)) [55] via an affinity complex formation at low temperatures (4 °C) to remove the undesired salts from the sulfated polysaccharides [52]. This specific precipitation reaction between fucoidans and Cetavlon® is applied in screening tests of microorganisms for putative fucoidanase activity [114].
Ale et al. published comprehensive articles discussing the history of extraction, including the different classical extraction methods of fucoidans, and reported that extraction procedures significantly affect the polymers monomeric composition, even for the same organism [60,115]. Beyond simple hot water extraction [58,116], attempts were made to increase the selectivity and extraction yields, including extraction in acidic [117], alkaline [118], and buffered [41,119] aqueous solutions. However, a neutralization step is required, using Na2CO3 or (NH4)2CO3, directly after extraction to guard against the non-specific acidic hydrolysis of the polymer [101,115]. Such drastic pH changes affect the chemical and physicochemical properties of fucoidans during the extraction step.
Currently, besides the previously discussed classical extraction methods based on thermal energy, extraction protocols based on vibrational energy have been developed. These protocols are based on microwave-assisted (MAE) [120,121] or ultrasound-assisted (UAE) [94,122] extraction steps to elicit cell wall degradation which improves the polymer release into aqueous solvent. These protocols were optimized either using an approach that modified one factor at a time or a multiple factorial design, setting the polymers production yield, monomeric composition and biological activities as the measured responses.
Recently, combined sulfated polysaccharides extraction protocols were optimized from different brown algae species using hydrothermal-assisted extraction (HAE) followed by sequential ultrasound and thermal technologies [123]. Similarly, subcritical water extraction was applied to increase the production yield of fucoidans from Nizamuddinia zanardinii [124]; such mild conditions may be advantageous to preserve the native chemical backbone and physicochemical characters of fucoidans.
Recently, as a trial to reduce such undesirable effects, enzyme-aided or assisted extraction (EAE) protocols are being developed using enzymes instead of harsh chemicals and high extraction temperatures during extraction. These include cellulase, papain, laminarinase, alginate lyase, and protease, which are present in products of Novozymes [79,125,126,127,128]. In addition, other cost-effective and time-saving techniques are reported, like those for terrestrial plant polysaccharides, such as extraction under vacuum to lower the boiling point of water and hence avoid possible heat-induced fucoidans degradation [129]. Alternatively, 0.5% (w/v) ethylenediaminetetraacetic acid (EDTA) was applied at 70 °C for simultaneous extraction of Laminaria japonica fucoidans and removal of pigments [130].
3.3. Separation Physical Methods
Filtration, dialysis and centrifugation, either for the algal biomass or precipitates, are also among the downstream processes after pre-treatment and extraction steps [131,132,133]. Cross-flow filtration and dialysis against water are usually performed using different molecular weight cut-off (MWCO) membranes for isolation of fucoidans from smaller compounds depending on the high molecular weight of fucoidans [134] and also for fractionation purposes, where low molecular weight fucoidans (LMWF) can be separated from high molecular weight analogues (HMWF) [49].
In addition, filtration, concentration, and fractionation are simultaneously performed using centrifugal concentrators (Vivaspin®) equipped with membranes with certain MWCO, like in protein purification. However, in some cases, especially in the presence of bulk masses or high concentrations of salts and small contaminants, the use of centrifugal concentrators becomes practically and economically unsuitable for fucoidans purification. In such cases, bulky contaminants result in membrane clogging leading to its deterioration and increasing the production cost.
3.4. Purification
Despite the previously mentioned purification steps, residuals of co-extracted contaminants are still present, and resulting fucoidans are still crude-type. [27]. Therefore, further selective purification steps are needed to obtain a high-quality product for reproducible and accurate biological investigations. Some researches adopted simple, non-chromatographic steps, such as bleaching of the crude fucoidans (NaClO2 in dilute HCl) followed by precipitation with cetyltrimethylammonium bromide [135] or by cold overnight incubation in aqueous buffered solution of calcium acetate (20 mM, pH = 6.5 -7.5) followed by dialysis [136]. In addition, membrane filtration was reported to produce fucoidans fractions of different molecular weight [137].
However, other chromatographic purification techniques were discussed in our previous publications [41,53,98,102]. Almost all the chromatographic techniques are based on the permanent negatively charged sulfate ester groups distributed on the polymer backbone which allow selective fucoidans capture. However, carboxylated (e.g., alginate) and phosphorylated (e.g., nucleic acids) compounds might interfere [138,139]. Therefore, the pH value of the applied solvents is critical during chromatographic purification. One option for this uses anionic exchange resins (e.g., diethylaminoethyl cellulose or DEAE-cellulose), which was performed at pH 7.2 using 0.1 M sodium phosphate buffer [140]. An alternative is cationic dyes (e.g., toluidine blue- or perylene diimide derivative), modified resins or chitosan functioning in buffered solutions [27,102]. Both anion exchange and dye affinity chromatography involve the use of highly concentrated NaCl elution solvents. As a result, a subsequent purification step using chromatographic gel permeation [141] or dialysis [140] is required to remove salts, increasing the production costs. Other methods based on the use of biological macromolecules, such as lectins and anti-thrombin III, were also reported [53].
Novel innovative purification techniques were recently developed, such as selective solid phase extraction for purifying fucoidans and other complex seaweeds polymers by molecularly imprinted polymers (MIP) [142,143] or MIP modified by deep eutectic solvents [142,143]. Abdella et al., developed a green and time-saving purification protocol using genipin cross-linked toluidine blue immobilized-chitosan beads employing fucoidans affinity to cationic thiazine dyes [102].
4. Recent Uncommon Applications
In addition to the classical therapeutic applications of fucoidans, including anti-coagulant [41,144], anti-viral [145,146], anti-inflammatory [46,147] and selective cytotoxic and anti-tumor uses [39,50], uncommon bioactivities, including cosmeceutical, pharmaceutical, diagnostic, and synergistic therapeutic applications were recently reported [32]. Recent fucoidans uses included therapeutic treatment of major blindness diseases [148]. It has also been used as a drug carrier, especially for anti-cancer treatments and anti-biotics. Additionally, fucoidans have been shown to improve drug bioavailability and efficacy in pharmaceutical formulations, including in nanoparticles, liposomes, microparticles, and semisolid formulations [28,149,150]. Table 1 summarizes some of the recent and uncommon fucoidans applications based on in-vitro or in-vivo studies, in addition to biogenic resources and physicochemical features.
Table 1.
Some selected recent therapeutic, diagnostic and pharmaceutical applications of fucoidans including the biogenic sources.
| Application | Biogenic Source | Quality Grade/Purification Method | Structural Features | Involved Mechanism | Ref. |
|---|---|---|---|---|---|
| Therapeutic | |||||
| Anti-viral (IAV) |
Kjellmaniella crassifolia (Laminariales) | * | Inhibition of the viral neuraminidase (NA) Interference with the cellular EGFR pathway |
[43] | |
| Anti-metabolic syndrome | Fucus vesiculosus (Fucales) | Dialysis of crude alginate-free fucoidans | Alternating α(1→3)/α(1→4)linked fucose, Mw > 7.0 × 103 g/mol |
Regulation of jnk, akt, and ampk signaling Alleviation of insulin resistance Regulation of lipid metabolism |
[151] |
| Anti-leishmaniasis | Commercial product purchased from Sigma-Aldrich® | Polymer of α-(1→3) linked fucose | Activation of the mitogen-activated protein kinase (MAPK)/NF-κB pathway against Leishmania donovani-infected macrophages | [152] | |
| Enhancement of dendritic cells maturation, production of pro-inflammatory cytokines, and down-regulation of anti-inflammatory cytokines |
[153,154] | ||||
| Immunostimulant | Nizamuddinia zanardinii (Fucales) | A fraction of DEAE Sepharose Fast Flow column | Highly branched polymer Mw: 953.6 × 103 g/mol |
Stimulation of RAW264.7 murine macrophage and NK cells | [155] |
| Anti-metastasis | Undaria pinnatifida (Laminariales) | DEAE-cellulose, and Sephadex G-100 column chromatography (purity>90%) | Mw: of 10.4356 × 104 g/mol | - Suppression of Hca-F cell growth, adhesion, invasion, and metastasis capabilities, - Inactivation of the NF-κB pathway |
[156] |
| Gastrointestinal tract protective | Purity ≥ 95% (Commercial product purchased from Sigma-Aldrich®) | Protection against H2O2-induced damage via activation of the NRF2 signaling pathway | [157] | ||
| Anti-malaria | - Partial purification by cetylpyridinum chloride Fractionation by DEAE-Sephadex A-25 column |
Sugar monomers, and uronic acid, M.wt: approx. 15 × 103 g/mol |
In-vitro and in-vivo inhibition of erythrocytes invasion by P. falciparum merozoites | [158] | |
| Renal protective |
Laminaria japonica (Laminariales) |
LMWF (Mw: 7 x 103 g/mol) |
Inhibition of overexpression of pro-inflammatory and pro-fibrotic factors, oxidative stress and apoptosis | [159,160] | |
| Cardio-, hepatic- and renal protective | Commercial product purchased from Absunutrix Lyfetrition® | Reduction of oxidative stress, pro-inflammatory effects and injuries to the cardiac, hepatic, and renal tissues | [161] | ||
| Inhibition of tumor angiogenesis | Sargassum hemiphyllum (Fucales) | Hydrolyzed crude extract | LMWF; 760 g/mol |
Suppression of HIF-1/VEGF-regulated signaling pathway | [162] |
| Pro-angiogenic |
Ascophyllum nodosum (Fucales) |
Fractionated with dialysis commercial crude fucoidan (ASPHY) | LMWF (<4.9 x 103 g/mol) |
Increase of the vascular network formation regulated via Erk1/2 and PI3K/AKT cell signaling pathways | [163] |
| Alleviation of diabetic complications |
S. Fusiforme (Fucales) |
Crude extract | Mw: 205.89x103 g/mol, high sulfate content |
- Suppression of oxidative stress - Alteration of the gut microbiota - Attenuation of the pathological changes in heart and liver |
[164] |
| Diagnostic | |||||
| Imaging of cardiovascular diseases |
Ascophyllum nodosum (Fucales) |
An oxidative-reductive degraded crude extract (purchased from Algues and Mer, Ascophyscient®) | GMP-grade LMWF (7.1x103 g/mol) | Synthesis of technetium-99m-fucoidan radiotracer for detection of P-selectin | [56] |
| Commercial product from Algues and Mer | Synthesis of polycyanoacrylate-fucoidan microcapsules (Fuco-MCs) for detection of P-selectin | [165] | |||
| Cosmeceutical | |||||
| Anti-Photoaging |
Ecklonia cava (Laminariales) |
Enzymatic degradation of a commercial HMWF | LMWF (Mw: 8 × 103 g/mol) | Anti-oxidant, anti-apoptotic, and MMP-9-inhibiting effects | [166] |
| Skin brightening and age spot reduction | F. vesiculosus (Fucales) | Crude extracts purchased from Marinova® Pty Ltd. | 58.6% fucoidans, 33.7% polyphenol |
Increase of Sirtuin 1 (SIRT1) expression in vitro | [167,168] |
| Skin immunity, soothing and protection | U. pinnatifida (Laminariales) | 89.6% fucoidans, <2% polyphenol |
|||
| Reconstruction of skin | F. vesiculosus (Fucales) | Commercial product from Sigma-Aldrich® (not determined the degree of purity) |
Increase of proliferating cell nuclear antigen (PCNA) p63 and α6-integrin expression | [169] | |
| Pharmaceutical technology | |||||
| As vehicle for drug delivery |
F. vesiculosus (Fucales) |
Commercial product purchased from Sigma-Aldrich® | Mw: 57.26 ×103 g/mol | - Chitosan-fucoidans-based nanoparticles for delivery of anti-cancers (e.g., curcumin-loaded NPs) - Nanoencapsulation of poly L-lysine |
[170,171] |
| Piperlongumine (PL)-loaded chitosan-fucoidan nanoparticles (PL-CS-F NPs) | [172] | ||||
| Synthesis of fucoidan/trimethylchitosan nanoparticles (FUC-TMC-NPs) as adjuvant in anthrax vaccine adsorbed | [173] | ||||
| Green synthesis of silver nanoparticles | Synthesis of chitosan-fucoidan complex-coated AgNPs | [174] | |||
*: Not specified.
5. Enzymatic Modification of Native Fucoidans
Owing to their high molecular weight, therapeutic applications of native fucoidans face many challenges including structure elucidation, solubility, manufacturing, and handling [63,116], in addition to safety as a food supplement [175]. Structure elucidation and quantitation of native fucoidans is highly complicated and requires advanced or hyphenated spectroscopic techniques such asHPLC-MS/MS as it applied in Sea Cucumbers fucoidans [176,177]. Also, these techniques must be applied after a step of enzymatic or acid hydrolysis to transform the fucoidans polymers to oligomers. According to their molecular weight, fucoidans are classified into three classes: LMWF (<10 kDa), medium molecular weight fucoidan (MMWF) (10–10000 kDa), and HMWF (>10000 kDa) [31]. LMWF demonstrated better bioavailability and bioactivities than HMWF [178,179]. As a consequence, several articles reported physical, chemical and enzymatic modification of the native HMWF to get LMWF of higher biological activity [62]. Specifically, enzymatic modification of macroalgal polysaccharides, including fucoidans by either fucoidanases or sulfatases, is characterized by regioselectivity and stereospecificity. This new trend is considered crucial and highly promising for current and future applications of polysaccharides [180].
Nevertheless, our publications in 2009 particularly reviewed the specific enzymatic degradation of fucoidans induced by fucoidanases (EC 3.2.1.44) and α-L-fucosidases (EC 3.2.1.51), mainly those isolated from marine bacteria [35]. Cumashi, et al. studied the chemical structures of different fucoidans isolated from a number of brown algal species [181]. Their proposed models, which were highly appreciated and recommended by many researchers [60], showed the backbone of fucoidans to be mainly an alternating α-(1-4) and α-(1-3) linked L-fucopyranoside. Regarding the sulfation pattern, C-2 is usually substituted with sulfate ester groups in addition to alternating C-3 or C-4 in L-fucopyranose residue, according to the glycosidic linkages. In addition, branched chain polymers were also found as in F. serratus. Other minor sugar units (e.g., mannose, galactose, glucose and xylose) occur as well in fucoidans structure; however, their distribution pattern and positions are still unknown [60,181]. Now, the mechanism of enzymatic degradation can be described in relation to fucoidans chemical structures.
Despite the increasing number of publications investigating fucoidanase activity of different marine species cell extracts, few of these enzymes have been isolated and characterized. Moreover, genome sequences encoding few fucoidanases have been published, including Ffa2 and FFA1 from Formosa algae KMM 3553T [182,183], FcnA from Mariniflexili fucanivorans SW5T [184]. Therefore, specificity of fucoidanases, type of cleaved glycoside bond, structure-activity relationship studies and enzyme stability are still poorly described. It was only observed that identified microbial fucoidanses act only on fucoidans isolated from their respective symbionts [185]. In fact, fucoidanases have not actually been fully utilized yet as a powerful tool either for the structural studies of fucoidans or production of defined and well-characterized bioactive fragments of extracted fucoidans, as shown in Table 2.
Table 2.
Source of fucoidans as a substrate and mode of action of some fucoidanases.
| Biogenic Source of Fucoidans | Fucoidanase Source | Mode of Action | Ref. |
|---|---|---|---|
| F. evanescens | Formosa algae KMM 3553 | Endo α-1→4 |
[61,182] |
|
Pseudoalteromonas citrea strains KMM 3296, KMM 3297, KMM 3298 |
Endo α-1→3 |
[186] | |
| F. vesiculosus | Dendryphiella. arenaria TM94 | Endo n.d. * |
[187] |
| Kjellmaniella crassifolia | Fucobacter marina SA-0082 | Endo β-1→4 |
[188] |
| Cladosiphon okamuranus | Fucophilus fucoidanolyticus SI-1234 | Endo α-1→3 |
[189] |
| Flavobacterium sp. F-31 | Endo n.d. |
[190] | |
| F. distichus | Littorina kurila | Endo α-1→3 |
[191] |
| Pelvetia canaliculata | Mariniflexile fucanivorans SW5T | Endo α-1→4 |
[184] |
| Undara pinnatifida | Sphingomonas paucimobilis PF-1 | Endo n.d. |
[192,193] |
| Saccharina cichorioides |
Pseudoalteromonas citrea strains KMM 3296, KMM 3297, KMM 3298 |
Endo α-1→3 |
[186] |
| Nemacystus decipiens | Mizuhopecten yessoensis | Endo n.d. |
[194] |
| Ascophyllum nodosum | Pecten maximus | Exo n.d. |
[195,196] |
| Thelenota ananas (Wild sea cucumber) | Wenyingzhuangia Fucanilytica | Endo n.d. |
[197] |
* n.d.: not determined.
Similarly, recent advances in bioinformatics and genome sequencing of microbial species have resulted in a continual increase of novel genome sequences. These genomes demonstrated various potential genes encoding for enzymes with biopolymer-degrading capabilities, such as Shewanella violacea DSS12 (NC_014012.1), Formosa algae KMM 3553 (NZ_LMAK01000014.1) [182], Formosa haliotis MA1 (NZ_BDEL01000001.1) [198], Wenyingzhuangia fucanilytica CZ1127 (NZ_CP014224.1) [199] and Pseudoalteromonas sp. strain A601 (MXQF01000000) [200]. Moreover, production of stabilized fucoidanases has been achieved by targeted truncation of the C-terminal of FcnA2, Fda1 and Fda2. This recently developed method may help with enzymatic production of defined degrees of polymerization and more bioactive products from native fucoidan substrates [201].
6. Conclusion and Future Prospective
As multifunctional molecules, fucoidans have received special interest based on their proven efficacy in different fields. The current article reviewed many aspects related to fucoidans’ production, mainly from brown algae. Biogenic source and downstream processes were shown as major factors determining their application, which is affected by molecular weight and quality grade of fucoidans. Therefore, the alteration of fucoidans’ native structure was recommended, especially as performed by fucoidanases. Their production in nanoform or in combination with other polymers can improve or modify their potential uses, allowing their expanded potential as therapeutic agents, e.g., in anti-cancer applications [202].
Production of high-quality purified fucoidans is urgently required to clarify the relationships between chemical structure and the various bioactivities attributed to fucoidans, eliminating any interference from contaminants. However, it was observed in some cases that crude extracts and presence of co-extracted contaminants, especially polyphenolic phlorotannins, have advantageous cosmeceutical effects due to their powerful anti-oxidant activity [203,204].
Novel techniques, either in cultivation or downstream processes, have been established, increasing the global production yields and reducing ecological and economic problems. A new advance toward achieving such goals was established by optimization of water extraction via measurement of kinetic parameters [205]. In addition to this, it is expected that most future trends in marine biotechnology research will focus on the cell wall and extracellular matrix components of brown algae, including fucoidans’ biosynthetic genes and production regulators [23,53,63,206,207,208]. Such trials may enable the scientific community to produce more bioactive molecules of fucoidans with defined characteristics, including degree of polymerization, sulfate content and pattern, in reproducible manners.
Acknowledgments
The research is funded by the „Deutsche Forschungsgemeinschaft (DFG, German Research Foundation)-Project-ID 172116086-SFB 926“. The authors would like also to thank Mrs. Aya Abdella and Ms. Gabrielle Phillips for helpful comments and English editing of the manuscript.
Author Contributions
R.U. planned the manuscript’s topics and is the corresponding author, while A.Z. collected the data and wrote the article. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interests.
References
- 1.Munoz-Bonilla A., Echeverria C., Sonseca A., Arrieta M.P., Fernandez-Garcia M. Bio-based polymers with antimicrobial properties towards sustainable development. Materials. 2019;12:641. doi: 10.3390/ma12040641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Amos R.A., Mohnen D. Critical review of plant cell wall matrix polysaccharide glycosyltransferase activities verified by heterologous protein expression. Front. Plant Sci. 2019;10:915. doi: 10.3389/fpls.2019.00915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lampugnani E.R., Flores-Sandoval E., Tan Q.W., Mutwil M., Bowman J.L., Persson S. Cellulose synthesis—central components and their evolutionary relationships. Trends Plant Sci. 2019;24:402–412. doi: 10.1016/j.tplants.2019.02.011. [DOI] [PubMed] [Google Scholar]
- 4.Helle S., Bray F., Verbeke J., Devassine S., Courseaux A., Facon M., Tokarski C., Rolando C., Szydlowski N. Proteome analysis of potato starch reveals the presence of new starch metabolic proteins as well as multiple protease inhibitors. Front. Plant Sci. 2018;9:746. doi: 10.3389/fpls.2018.00746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ball S., Colleoni C., Cenci U., Raj J.N., Tirtiaux C. The evolution of glycogen and starch metabolism in eukaryotes gives molecular clues to understand the establishment of plastid endosymbiosis. J. Exp. Bot. 2011;62:1775–1801. doi: 10.1093/jxb/erq411. [DOI] [PubMed] [Google Scholar]
- 6.Edmond Ghanem M., Han R.-M., Classen B., Quetin-Leclerq J., Mahy G., Ruan C.-J., Qin P., Pérez-Alfocea F., Lutts S. Mucilage and Polysaccharides in the Halophyte plant species Kosteletzkya virginica: Localization and composition in relation to salt stress. J. Plant Physiol. 2010;167:382–392. doi: 10.1016/j.jplph.2009.10.012. [DOI] [PubMed] [Google Scholar]
- 7.Shukla P.S., Mantin E.G., Adil M., Bajpai S., Critchley A.T., Prithiviraj B. Ascophyllum nodosum-based biostimulants: sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 2019;10:655. doi: 10.3389/fpls.2019.00655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hamed I., Özogul F., Özogul Y., Regenstein J.M. Marine bioactive compounds and their health benefits: A review. Compr. Rev. Food Sci. Food Saf. 2015;14:446–465. doi: 10.1111/1541-4337.12136. [DOI] [Google Scholar]
- 9.Lee Y.-E., Kim H., Seo C., Park T., Lee K.B., Yoo S.Y., Hong S.C., Kim J.T., Lee J. Marine polysaccharides: therapeutic efficacy and biomedical applications. Arch. Pharmacal Res. 2017;40:1006–1020. doi: 10.1007/s12272-017-0958-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ruocco N., Costantini S., Guariniello S., Costantini M. Polysaccharides from the marine environment with pharmacological, cosmeceutical and nutraceutical potential. Molecules. 2016;21:551. doi: 10.3390/molecules21050551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Laurienzo P. Marine Polysaccharides in Pharmaceutical Applications: An Overview. Mar. Drugs. 2010;8:2435–2465. doi: 10.3390/md8092435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Meenakshi S., Umayaparvathi S., Saravanan R., Manivasagam T., Balasubramanian T. Hepatoprotective effect of fucoidan isolated from the seaweed turbinaria decurrens in ethanol intoxicated rats. Int. J. Biol. Macromol. 2014;67:367–372. doi: 10.1016/j.ijbiomac.2014.03.042. [DOI] [PubMed] [Google Scholar]
- 13.Cunha L., Grenha A. Sulfated seaweed polysaccharides as multifunctional materials in drug delivery applications. Mar. Drugs. 2016;14:42. doi: 10.3390/md14030042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hudak J.E., Bertozzi C.R. Glycotherapy: New advances inspire a reemergence of glycans in medicine. Chem. Biol. 2014;21:16–37. doi: 10.1016/j.chembiol.2013.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Novotny M.V., Alley W.R., Jr. Recent trends in analytical and structural glycobiology. Curr. Opin. Chem. Biol. 2013;17:832–840. doi: 10.1016/j.cbpa.2013.05.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Caldwell G.S., Pagett H.E. Marine glycobiology: Current status and future perspectives. Mar. Biotechnol. 2010;12:241–252. doi: 10.1007/s10126-010-9263-5. [DOI] [PubMed] [Google Scholar]
- 17.Pomin V.H. Marine medicinal Gglycomics. Front. Cell. Infect. Microbiol. 2014;4:5. doi: 10.3389/fcimb.2014.00005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.De Jesus Raposo M.F., de Morais A.M.B., de Morais R.M.S.C. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs. 2015;13:2967–3028. doi: 10.3390/md13052967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gobet A., Barbeyron T., Matard-Mann M., Magdelenat G., Vallenet D., Duchaud E., Michel G. Evolutionary evidence of algal polysaccharide degradation acquisition by Pseudoalteromonas carrageenovora 9T to adapt to macroalgal niches. Front. Microbiol. 2018;9:2740. doi: 10.3389/fmicb.2018.02740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Maneein S., Milledge J.J., Nielsen B.V., Harvey P.J. A Review of seaweed pre-treatment methods for enhanced biofuel production by anaerobic digestion or fermentation. Fermentation. 2018;4:100. doi: 10.3390/fermentation4040100. [DOI] [Google Scholar]
- 21.Afonso N.C., Catarino M.D., Silva A.M.S., Cardoso S.M. Brown macroalgae as valuable food ingredients. Antioxidants. 2019;8:365. doi: 10.3390/antiox8090365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Catarino M.D., Silva A.M.S., Cardoso S.M. Phycochemical constituents and biological activities of Fucus spp. Mar. Drugs. 2018;16:249. doi: 10.3390/md16080249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Deniaud-Bouët E., Hardouin K., Potin P., Kloareg B., Hervé C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr. Polym. 2017;175:395–408. doi: 10.1016/j.carbpol.2017.07.082. [DOI] [PubMed] [Google Scholar]
- 24.Deniaud-Bouët E., Kervarec N., Michel G., Tonon T., Kloareg B., Hervé C. Chemical and enzymatic fractionation of cell walls from Fucales: Insights into the structure of the extracellular matrix of brown algae. Ann. Bot. 2014;114:1203–1216. doi: 10.1093/aob/mcu096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Generalić Mekinić I., Skroza D., Šimat V., Hamed I., Čagalj M., Popović Perković Z. Phenolic content of brown algae (Pheophyceae) species: Extraction, identification, and quantification. Biomolecules. 2019;9:244. doi: 10.3390/biom9060244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chang Y., Xue C., Tang Q., Li D., Wu X., Wang J. Isolation and characterization of a sea cucumber fucoidan-utilizing marine bacterium. Lett. Appl. Microbiol. 2010;50:301–307. doi: 10.1111/j.1472-765X.2009.02792.x. [DOI] [PubMed] [Google Scholar]
- 27.Zayed A., Dienemann C., Giese C., Krämer R., Ulber R. An immobilized perylene diimide derivative for fucoidan purification from a crude brown algae extract. Process Biochem. 2018;65:233–238. doi: 10.1016/j.procbio.2017.10.012. [DOI] [Google Scholar]
- 28.Citkowska A., Szekalska M., Winnicka K. Possibilities of fucoidan utilization in the development of pharmaceutical dosage forms. Mar. Drugs. 2019;17:458. doi: 10.3390/md17080458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li S., Li J., Zhi Z., Hu Y., Ge J., Ye X., Tian D., Linhardt R.J., Chen S. 4-O-sulfation in sea cucumber fucodians contribute to reversing dyslipidiaemia caused by HFD. Int. J. Biol. Macromol. 2017;99:96–104. doi: 10.1016/j.ijbiomac.2017.01.145. [DOI] [PubMed] [Google Scholar]
- 30.Fitton J.H., Stringer D.S., Park A.Y., Karpiniec S.N. Therapies from fucoidan: new developments. Mar. Drugs. 2019;17:571. doi: 10.3390/md17100571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Van Weelden G., Bobinski M., Okla K., van Weelden W.J., Romano A., Pijnenborg J.M.A. Fucoidan structure and activity in relation to anti-cancer mechanisms. Mar. Drugs. 2019;17:32. doi: 10.3390/md17010032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wang Y., Xing M., Cao Q., Ji A., Liang H., Song S. Biological activities of fucoidan and the factors mediating its therapeutic effects: A review of recent studies. Mar. Drugs. 2019;17:183. doi: 10.3390/md17030183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Luthuli S., Wu S., Cheng Y., Zheng X., Wu M., Tong H. Therapeutic effects of fucoidan: A review on recent studies. Mar. Drugs. 2019;17:487. doi: 10.3390/md17090487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chollet L., Saboural P., Chauvierre C., Villemin J.N., Letourneur D., Chaubet F. Fucoidans in nanomedicine. Mar. Drugs. 2016;14:145. doi: 10.3390/md14080145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Holtkamp A.D., Kelly S., Ulber R., Lang S. Fucoidans and fucoidanases-focus on techniques for molecular structure elucidation and modification of marine polysaccharides. Appl. Microbiol. Biotechnol. 2009;82:1–11. doi: 10.1007/s00253-008-1790-x. [DOI] [PubMed] [Google Scholar]
- 36.Ustyuzhanina N.E., Bilan M.I., Ushakova N.A., Usov A.I., Kiselevskiy M.V., Nifantiev N.E. Fucoidans: Pro- or antiangiogenic agents? Glycobiology. 2014;24:1265–1274. doi: 10.1093/glycob/cwu063. [DOI] [PubMed] [Google Scholar]
- 37.Zhao Y., Zheng Y., Wang J., Ma S., Yu Y., White W.L., Yang S., Yang F., Lu J. Fucoidan extracted from Undaria pinnatifida: Source for nutraceuticals/functional foods. Mar. Drugs. 2018;16:321. doi: 10.3390/md16090321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Arumugam P., Arunkumar K., Sivakumar L., Murugan M., Murugan K. Anticancer effect of fucoidan on cell proliferation, cell cycle progression, genetic damage and apoptotic cell death in HepG2 cancer cells. Toxicol. Rep. 2019;6:556–563. doi: 10.1016/j.toxrep.2019.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Atashrazm F., Lowenthal R.M., Woods G.M., Holloway A.F., Dickinson J.L. Fucoidan and cancer: A multifunctional molecule with anti-tumor potential. Mar. Drugs. 2015;13:2327–2346. doi: 10.3390/md13042327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chen L.-M., Liu P.-Y., Chen Y.-A., Tseng H.-Y., Shen P.-C., Hwang P.-A., Hsu H.-L. Oligo-fucoidan prevents IL-6 and CCL2 production and cooperates with p53 to suppress ATM signaling and tumor progression. Sci. Rep. 2017;7:11864. doi: 10.1038/s41598-017-12111-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zayed A., Muffler K., Hahn T., Rupp S., Finkelmeier D., Burger-Kentischer A., Ulber R. Physicochemical and biological characterization of fucoidan from Fucus vesiculosus purified by dye affinity chromatography. Mar. Drugs. 2016;14:79. doi: 10.3390/md14040079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zhao X., Dong S., Wang J., Li F., Chen A., Li B. A comparative study of antithrombotic and antiplatelet activities of different fucoidans from Laminaria japonica. Thromb. Res. 2012;129:771–778. doi: 10.1016/j.thromres.2011.07.041. [DOI] [PubMed] [Google Scholar]
- 43.Wang W., Wu J., Zhang X., Hao C., Zhao X., Jiao G., Shan X., Tai W., Yu G. Inhibition of influenza A virus infection by fucoidan targeting viral neuraminidase and cellular EGFR pathway. Sci. Rep. 2017;7:40760. doi: 10.1038/srep40760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Prokofjeva M.M., Imbs T.I., Shevchenko N.M., Spirin P.V., Horn S., Fehse B., Zvyagintseva T.N., Prassolov V.S. Fucoidans as potential inhibitors of HIV-1. Mar. Drugs. 2013;11:3000–3014. doi: 10.3390/md11083000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Takahashi H., Kawaguchi M., Kitamura K., Narumiya S., Kawamura M., Tengan I., Nishimoto S., Hanamure Y., Majima Y., Tsubura S., et al. An exploratory study on the anti-inflammatory effects of fucoidan in relation to quality of life in advanced cancer patients. Integr. Cancer Ther. 2018;17:282–291. doi: 10.1177/1534735417692097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Park H.Y., Han M.H., Park C., Jin C.-Y., Kim G.-Y., Choi I.-W., Kim N.D., Nam T.-J., Kwon T.K., Choi Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011;49:1745–1752. doi: 10.1016/j.fct.2011.04.020. [DOI] [PubMed] [Google Scholar]
- 47.Li B., Lu F., Wei X., Zhao R. Fucoidan: Structure and bioactivity. Molecules. 2008;13:1671–1695. doi: 10.3390/molecules13081671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lu J., Shi K.K., Chen S., Wang J., Hassouna A., White L.N., Merien F., Xie M., Kong Q., Li J., et al. Fucoidan extracted from the New Zealand Undaria pinnatifida-Physicochemical comparison against five other fucoidans: Unique low molecular weight fraction bioactivity in breast cancer cell lines. Mar. Drugs. 2018;16:461. doi: 10.3390/md16120461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yoo H.J., You D.-J., Lee K.-W. Characterization and immunomodulatory effects of high molecular weight fucoidan fraction from the sporophyll of Undaria pinnatifida in cyclophosphamide-induced immunosuppressed mice. Mar. Drugs. 2019;17:447. doi: 10.3390/md17080447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zayed A., Hahn T., Finkelmeier D., Burger-Kentischer A., Rupp S., Krämer R., Ulber R. Phenomenological investigation of the cytotoxic activity of fucoidan isolated from Fucus vesiculosus. Process Biochem. 2019;81:182–187. doi: 10.1016/j.procbio.2019.03.026. [DOI] [Google Scholar]
- 51.Imbs T.I., Skriptsova A.V., Zvyagintseva T.N. Antioxidant activity of fucose-containing sulfated polysaccharides obtained from Fucus evanescens by different extraction methods. J. Appl. Phycol. 2015;27:545–553. doi: 10.1007/s10811-014-0293-7. [DOI] [Google Scholar]
- 52.Hifney A.F., Fawzy M.A., Abdel-Gawad K.M., Gomaa M. Industrial optimization of fucoidan extraction from Sargassum sp. and its potential antioxidant and emulsifying activities. Food Hydrocoll. 2016;54:77–88. doi: 10.1016/j.foodhyd.2015.09.022. [DOI] [Google Scholar]
- 53.Zayed A., Ulber R. Fucoidan Production: Approval Key Challenges and Opportunities. Carbohydr. Polym. 2019;211:289–297. doi: 10.1016/j.carbpol.2019.01.105. [DOI] [PubMed] [Google Scholar]
- 54.Fitton J.H., Stringer D.N., Karpiniec S.S. Therapies from fucoidan: An update. Mar. Drugs. 2015;13:5920–5946. doi: 10.3390/md13095920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mak W., Wang S.K., Liu T., Hamid N., Li Y., Lu J., White W.L. Anti-proliferation potential and content of fucoidan extracted from sporophyll of New Zealand Undaria pinnatifida. Front. Nutr. 2014;1:9. doi: 10.3389/fnut.2014.00009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chauvierre C., Aid-Launais R., Aerts J., Chaubet F., Maire M., Chollet L., Rolland L., Bonafe R., Rossi S., Bussi S., et al. Pharmaceutical development and safety evaluation of a GMP-grade fucoidan for molecular diagnosis of cardiovascular diseases. Mar. Drugs. 2019;17:699. doi: 10.3390/md17120699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Torres M.D., Flórez-Fernández N., Simón-Vázquez R., Giménez-Abián J.F., Díaz J.F., González-Fernández Á., Domínguez H. Fucoidans: The importance of processing on their anti-tumoral properties. Algal Res. 2020;45:101748. doi: 10.1016/j.algal.2019.101748. [DOI] [Google Scholar]
- 58.Borazjani N.J., Tabarsa M., You S., Rezaei M. Improved immunomodulatory and antioxidant properties of unrefined fucoidans from Sargassum angustifolium by hydrolysis. J. Food Sci. Technol. 2017;54:4016–4025. doi: 10.1007/s13197-017-2867-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Koyanagi S., Tanigawa N., Nakagawa H., Soeda S., Shimeno H. Oversulfation of fucoidan enhances its anti-angiogenic and antitumor activities. Biochem. Pharmacol. 2003;65:173–179. doi: 10.1016/S0006-2952(02)01478-8. [DOI] [PubMed] [Google Scholar]
- 60.Ale M.T., Mikkelsen J.D., Meyer A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs. 2011;9:2106–2130. doi: 10.3390/md9102106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Silchenko A.S., Rasin A.B., Kusaykin M.I., Malyarenko O.S., Shevchenko N.M., Zueva A.O., Kalinovsky A.I., Zvyagintseva T.N., Ermakova S.P. Modification of native fucoidan from fucus evanescens by recombinant fucoidanase from marine bacteria Formosa algae. Carbohydr. Polym. 2018;193:189–195. doi: 10.1016/j.carbpol.2018.03.094. [DOI] [PubMed] [Google Scholar]
- 62.Jiao G., Yu G., Zhang J., Ewart H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs. 2011;9:196–223. doi: 10.3390/md9020196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Suprunchuk V.E. Low-molecular-weight fucoidan: Chemical modification, synthesis of its oligomeric fragments and mimetics. Carbohydr. Res. 2019;485:107806. doi: 10.1016/j.carres.2019.107806. [DOI] [PubMed] [Google Scholar]
- 64.Kardeby C., Fälker K., Haining E.J., Criel M., Lindkvist M., Barroso R., Påhlsson P., Ljungberg L.U., Tengdelius M., Rainger G.E., et al. Synthetic glycopolymers and natural fucoidans cause human platelet aggregation via PEAR1 and GPIbα. Blood Adv. 2019;3:275–287. doi: 10.1182/bloodadvances.2018024950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kasai A., Arafuka S., Koshiba N., Takahashi D., Toshima K. Systematic synthesis of low-molecular weight fucoidan derivatives and their effect on cancer cells. Org. Biomol. Chem. 2015;13:10556–10568. doi: 10.1039/C5OB01634G. [DOI] [PubMed] [Google Scholar]
- 66.Rhein-Knudsen N., Ale M.T., Meyer A.S. Seaweed hydrocolloid production: An update on enzyme assisted extraction and modification technologies. Mar. Drugs. 2015;13:3340–3359. doi: 10.3390/md13063340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Buschmann A.H., Camus C., Infante J., Neori A., Israel Á., Hernández-González M.C., Pereda S.V., Gomez-Pinchetti J.L., Golberg A., Tadmor-Shalev N., et al. Seaweed production: Overview of the global state of exploitation, farming and emerging research activity. Eur. J. Phycol. 2017;52:391–406. doi: 10.1080/09670262.2017.1365175. [DOI] [Google Scholar]
- 68.Duarte C.M., Wu J., Xiao X., Bruhn A., Krause-Jensen D. Can seaweed farming play a role in climate change mitigation and adaptation? Front. Mar. Sci. 2017;4:100. [Google Scholar]
- 69.Zayed A., Kovacheva M., Muffler K., Breiner H.-W., Stoeck T., Ulber R. Induction and genetic identification of a callus-like growth developed in the brown alga Fucus vesiculosus. Eng. Life Sci. 2019;19:363–369. doi: 10.1002/elsc.201800211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Cho G.Y., Rousseau F., de Reviers B., Boo S.M. Phylogenetic relationships within the fucales (Phaeophyceae) assessed by the photosystem I coding psaa sequences. Phycologia. 2006;45:512–519. doi: 10.2216/05-48.1. [DOI] [Google Scholar]
- 71.Wahl M., Molis M., Hobday A.J., Dudgeon S., Neumann R., Steinberg P., Campbell A.H., Marzinelli E., Connell S. The responses of brown macroalgae to environmental change from local to global scales: Direct versus ecologically mediated effects. Perspect. Phycol. 2015;2:11–29. doi: 10.1127/pip/2015/0019. [DOI] [Google Scholar]
- 72.Crous P.W., Wingfield M.J., Burgess T.I., Hardy G.S.J., Gené J., Guarro J., Baseia I.G., García D., Gusmão L.F.P., Souza-Motta C.M., et al. Fungal planet description sheets: 716–784. Persoonia. 2018;40:240–393. doi: 10.3767/persoonia.2018.40.10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kumar G.R., Reddy C.R.K., Jha B. Callus induction and thallus regeneration from callus of phycocolloid yielding seaweeds from the Indian coast. J. Appl. Phycol. 2007;19:15–25. doi: 10.1007/s10811-006-9104-0. [DOI] [Google Scholar]
- 74.Muhamad S.N.S., Ling A.P.-K., Wong C.-L. Effect of plant growth regulators on direct regeneration and callus induction from Sargassum polycystum C. Agardh. J. Appl. Phycol. 2018;30:3299–3310. doi: 10.1007/s10811-018-1649-1. [DOI] [Google Scholar]
- 75.Avila-Peltroche J., Won B.Y., Cho T.O. Protoplast isolation and regeneration from Hecatonema terminale (Ectocarpales, Phaeophyceae) using a simple mixture of commercial enzymes. J. Appl. Phycol. 2019;31:1873–1881. doi: 10.1007/s10811-018-1660-6. [DOI] [Google Scholar]
- 76.Luiten E.E., Akkerman I., Koulman A., Kamermans P., Reith H., Barbosa M.J., Sipkema D., Wijffels R.H. Realizing the promises of marine biotechnology. Biomol. Eng. 2003;20:429–439. doi: 10.1016/S1389-0344(03)00074-1. [DOI] [PubMed] [Google Scholar]
- 77.Baweja P., Sahoo D., García-Jiménez P., Robaina R.R. Review: Seaweed tissue culture as applied to biotechnology: Problems, achievements and prospects. Phycol. Res. 2009;57:45–58. doi: 10.1111/j.1440-1835.2008.00520.x. [DOI] [Google Scholar]
- 78.Huang Y.M., Rorrer G.L. Cultivation of microplantlets derived from the marine red alga Agardhiella subulata in a stirred tank photobioreactor. Biotechnol. Prog. 2003;19:418–427. doi: 10.1021/bp020123i. [DOI] [PubMed] [Google Scholar]
- 79.Hahn T., Lang S., Ulber R., Muffler K. Novel procedures for the extraction of fucoidan from brown algae. Process Biochem. 2012;47:1691–1698. doi: 10.1016/j.procbio.2012.06.016. [DOI] [Google Scholar]
- 80.Thinh P.D., Menshova R.V., Ermakova S.P., Anastyuk S.D., Ly B.M., Zvyagintseva T.N. Structural characteristics and anticancer activity of fucoidan from the brown alga Sargassum mcclurei. Mar. Drugs. 2013;11:1456–1476. doi: 10.3390/md11051456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.De Reviers B. Fucans and alginates without phenolic compounds. J. Appl. Phycol. 1989;1:75–76. doi: 10.1007/BF00003538. [DOI] [Google Scholar]
- 82.Yang W.N., Chen P.W., Huang C.Y. Compositional characteristics and in vitro evaluations of antioxidant and neuroprotective properties of crude extracts of fucoidan prepared from compressional puffing-pretreated Sargassum crassifolium. Mar. Drugs. 2017;15:183. doi: 10.3390/md15060183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zhang R., Zhang X., Tang Y., Mao J. Composition, isolation, purification and biological activities of Sargassum fusiforme polysaccharides: A review. Carbohydr. Polym. 2020;228:115381. doi: 10.1016/j.carbpol.2019.115381. [DOI] [PubMed] [Google Scholar]
- 84.Spicer S.E., Adams J.M.M., Thomas D.S., Gallagher J.A., Winters A.L. Novel rapid method for the characterisation of polymeric sugars from macroalgae. J. Appl. Phycol. 2017;29:1507–1513. doi: 10.1007/s10811-016-0995-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zou P., Lu X., Zhao H., Yuan Y., Meng L., Zhang C., Li Y. Polysaccharides derived from the brown algae Lessonia nigrescens enhance salt stress tolerance to wheat seedlings by enhancing the antioxidant system and modulating intracellular ion concentration. Front. Plant Sci. 2019;10:48. doi: 10.3389/fpls.2019.00048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Peng J., Yuan J.-P., Wu C.-F., Wang J.-H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs. 2011;9:1806–1828. doi: 10.3390/md9101806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Da Costa E., Domingues P., Melo T., Coelho E., Pereira R., Calado R., Abreu M.H., Domingues M.R. Lipidomic signatures reveal seasonal shifts on the relative abundance of high-valued lipids from the brown algae Fucus vesiculosus. Mar. Drugs. 2019;17:335. doi: 10.3390/md17060335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Terasaki M., Hirose A., Narayan B., Baba Y., Kawagoe C., Yasui H., Saga N., Hosokawa M., Miyashita K. Evaluation of recoverable functional lipid components of several brown seaweeds (Phaeophyta) from Japan with special reference to fucoxanthin and fucosterol contents. J. Phycol. 2009;45:974–980. doi: 10.1111/j.1529-8817.2009.00706.x. [DOI] [PubMed] [Google Scholar]
- 89.Zhu F. Interactions between cell wall polysaccharides and polyphenols. Crit. Rev. Food Sci. Nutr. 2018;58:1808–1831. doi: 10.1080/10408398.2017.1287659. [DOI] [PubMed] [Google Scholar]
- 90.Li Y., Fu X., Duan D., Liu X., Xu J., Gao X. Extraction and Identification of Phlorotannins from the Brown Alga, Sargassum fusiforme (Harvey) Setchell. Mar. Drugs. 2017;15:49. doi: 10.3390/md15020049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Bertoni G. A key step in phlorotannin biosynthesis revealed. Plant Cell. 2013;25:2770. doi: 10.1105/tpc.113.250813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Catarino M.D., Silva A.M.S., Cardoso S.M. Fucaceae: A source of bioactive phlorotannins. Int. J. Mol. Sci. 2017;18:1327. doi: 10.3390/ijms18061327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Thomas N.V., Kim S.K. Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Environ. Toxicol. Pharmacol. 2011;32:325–335. doi: 10.1016/j.etap.2011.09.004. [DOI] [PubMed] [Google Scholar]
- 94.Agregán R., Munekata P.E.S., Franco D., Carballo J., Barba F.J., Lorenzo J.M. Antioxidant potential of extracts obtained from macro- (Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata) and micro-Algae (Chlorella vulgaris and Spirulina platensis) assisted by ultrasound. Medicines. 2018;5:33. doi: 10.3390/medicines5020033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Zhang M.Y., Guo J., Hu X.M., Zhao S.Q., Li S.L., Wang J. An in vivo anti-tumor effect of eckol from marine brown algae by improving the immune response. Food Funct. 2019;10:4361–4371. doi: 10.1039/C9FO00865A. [DOI] [PubMed] [Google Scholar]
- 96.Gall E.A., Lelchat F., Hupel M., Jégou C., Stiger-Pouvreau V. Extraction and purification of phlorotannins from brown algae. In: Stengel D.B., Connan S., editors. Natural Products from Marine Algae: Methods and Protocols. Springer; New York, NY, USA: 2015. pp. 131–143. [DOI] [PubMed] [Google Scholar]
- 97.Brzonova I., Kozliak E.I., Andrianova A.A., LaVallie A., Kubátová A., Ji Y. Production of lignin based insoluble polymers (anionic hydrogels) by C. versicolor. Sci. Rep. 2017;7:17507. doi: 10.1038/s41598-017-17696-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Hahn T., Zayed A., Kovacheva M., Stadtmüller R., Lang S., Muffler K., Ulber R. Dye affinity chromatography for fast and simple purification of fucoidan from marine brown algae. Eng. Life Sci. 2016;16:78–87. doi: 10.1002/elsc.201500044. [DOI] [Google Scholar]
- 99.Chades T., Scully S.M., Ingvadottir E.M., Orlygsson J. Fermentation of mannitol extracts from brown macro algae by Thermophilic clostridia. Front. Microbiol. 2018;9:1931. doi: 10.3389/fmicb.2018.01931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Balboa E.M., Rivas S., Moure A., Dominguez H., Parajo J.C. Simultaneous extraction and depolymerization of fucoidan from Sargassum muticum in aqueous media. Mar. Drugs. 2013;11:4612–4627. doi: 10.3390/md11114612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Descamps V., Colin S., Lahaye M., Jam M., Richard C., Potin P., Barbeyron T., Yvin J.-C., Kloareg B. Isolation and culture of a marine bacterium degrading the sulfated fucans from marine brown algae. Mar. Biotechnol. 2006;8:27–39. doi: 10.1007/s10126-005-5107-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Abdella A.A., Ulber R., Zayed A. Chitosan-toluidine blue beads for purification of fucoidans. Carbohydr. Polym. 2020;231:115686. doi: 10.1016/j.carbpol.2019.115686. [DOI] [PubMed] [Google Scholar]
- 103.Kadam S.U., Tiwari B.K., O’Donnell C.P. Application of novel extraction technologies for bioactives from marine algae. J. Agric. Food Chem. 2013;61:4667–4675. doi: 10.1021/jf400819p. [DOI] [PubMed] [Google Scholar]
- 104.Pozharitskaya O.N., Shikov A.N., Faustova N.M., Obluchinskaya E.D., Kosman V.M., Vuorela H., Makarov V.G. Pharmacokinetic and tissue distribution of fucoidan from Fucus vesiculosus after oral administration to rats. Mar. Drugs. 2018;16:132. doi: 10.3390/md16040132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Hadj Ammar H., Lajili S., Ben Said R., Le Cerf D., Bouraoui A., Majdoub H. Physico-chemical characterization and pharmacological evaluation of sulfated polysaccharides from three species of Mediterranean brown algae of the genus Cystoseira. DARU J. Pharm. Sci. 2015;23:1. doi: 10.1186/s40199-015-0089-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Zhang Y., Xu M., Hu C., Liu A., Chen J., Gu C., Zhang X., You C., Tong H., Wu M., et al. Sargassum fusiforme fucoidan SP2 extends the lifespan of Drosophila melanogaster by upregulating the Nrf2-mediated antioxidant signaling pathway. Oxidative Med. Cell. Longev. 2019;2019:8918914. doi: 10.1155/2019/8918914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Saepudin E., Sinurat E., Suryabrata I.A. Depigmentation and characterization of fucoidan from brown seaweed Sargassum binderi Sonder. IOP Conference Series: Mater. Sci. Eng. 2018;299:012027. doi: 10.1088/1757-899X/299/1/012027. [DOI] [Google Scholar]
- 108.Patel A., Mikes F., Matsakas L. An overview of current pretreatment methods used to improve lipid extraction from Oleaginous micro-organisms. Molecules. 2018;23:1562. doi: 10.3390/molecules23071562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Huang C.-Y., Kuo C.-H., Chen P.-W. Compressional-puffing pretreatment enhances neuroprotective effects of fucoidans from the brown seaweed Sargassum hemiphyllum on 6-hydroxydopamine-induced apoptosis in SH-SY5Y cells. Molecules. 2017;23:78. doi: 10.3390/molecules23010078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Huang C.-Y., Wu S.-J., Yang W.-N., Kuan A.-W., Chen C.-Y. Antioxidant activities of crude extracts of fucoidan extracted from Sargassum glaucescens by a compressional-puffing-hydrothermal extraction process. Food Chem. 2016;197:1121–1129. doi: 10.1016/j.foodchem.2015.11.100. [DOI] [PubMed] [Google Scholar]
- 111.Kordjazi M., Shabanpour B., Zabihi E., Faramarzi M.A., Feizi F., Ahmadi Gavlighi H., Feghhi M.A., Hosseini S.A. Sulfated polysaccharides purified from two species of Padina improve collagen and epidermis formation in the rat. Int. J. Mol. Cell. Med. 2013;2:156–163. [PMC free article] [PubMed] [Google Scholar]
- 112.Cho M.L., Lee B.-Y., You S.G. Relationship between oversulfation and conformation of low and high molecular weight fucoidans and evaluation of their in vitro anticancer activity. Molecules. 2010;16:291–297. doi: 10.3390/molecules16010291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Oliveira R.M., Câmara R.B.G., Monte J.F.S., Viana R.L.S., Melo K.R.T., Queiroz M.F., Filgueira L.G.A., Oyama L.M., Rocha H.A.O. Commercial fucoidans from Fucus vesiculosus can be grouped into antiadipogenic and adipogenic agents. Mar. Drugs. 2018;16:193. doi: 10.3390/md16060193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Kusaykin M.I., Silchenko A.S., Zakharenko A.M., Zvyagintseva T.N. Fucoidanases. Glycobiology. 2015;26:3–12. doi: 10.1093/glycob/cwv072. [DOI] [PubMed] [Google Scholar]
- 115.Ale M.T., Meyer A.S. Fucoidans from brown seaweeds: An update on structures, extraction techniques and use of enzymes as tools for structural elucidation. RSC Adv. 2013;3:8131–8141. doi: 10.1039/C3RA23373A. [DOI] [Google Scholar]
- 116.Huang C.-Y., Kuo C.-H., Lee C.-H. Antibacterial and antioxidant capacities and attenuation of lipid accumulation in 3T3-L1 adipocytes by low-molecular-weight fucoidans prepared from compressional-puffing-pretreated Sargassum crassifolium. Mar. Drugs. 2018;16:24. doi: 10.3390/md16010024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Imbs T.I., Shevchenko N.M., Sukhoverkhov S.V., Semenova T.L., Skriptsova A.V., Zvyagintseva T.N. Seasonal variations of the composition and structural characteristics of polysaccharides from the brown alga Costaria costata. Chem. Nat. Compd. 2009;45:786–791. doi: 10.1007/s10600-010-9507-7. [DOI] [Google Scholar]
- 118.Fidelis G.P., Silva C.H.F., Nobre L., Medeiros V.P., Rocha H.A.O., Costa L.S. Antioxidant fucoidans obtained from tropical seaweed protect pre-osteoblastic cells from hydrogen peroxide-induced damage. Mar. Drugs. 2019;17:506. doi: 10.3390/md17090506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Rohwer K., Neupane S., Bittkau K.S., Galarza Perez M., Dorschmann P., Roider J., Alban S., Klettner A. Effects of Crude Fucus distichus Subspecies evanescens Fucoidan Extract on Retinal Pigment Epithelium Cells-Implications for Use in Age-Related Macular Degeneration. Mar. Drugs. 2019;17:538. doi: 10.3390/md17090538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Rodriguez-Jasso R.M., Mussatto S.I., Pastrana L., Aguilar C.N., Teixeira J.A. Microwave-assisted extraction of sulfated polysaccharides (fucoidan) from brown seaweed. Carbohydr. Polym. 2011;86:1137–1144. doi: 10.1016/j.carbpol.2011.06.006. [DOI] [Google Scholar]
- 121.Mussatto S.I. Microwave-assisted extraction of fucoidan from marine algae. In: Stengel D.B., Connan S., editors. Natural Products from Marine Algae: Methods and Protocols. Springer; New York, NY, USA: 2015. pp. 151–157. [DOI] [PubMed] [Google Scholar]
- 122.Alboofetileh M., Rezaei M., Tabarsa M., You S. Ultrasound-assisted extraction of sulfated polysaccharide from Nizamuddinia zanardinii: Process optimization, structural characterization, and biological properties. J. Food Process Eng. 2019;42:e12979. doi: 10.1111/jfpe.12979. [DOI] [Google Scholar]
- 123.Garcia-Vaquero M., O’Doherty J.V., Tiwari B.K., Sweeney T., Rajauria G. Enhancing the extraction of polysaccharides and antioxidants from macroalgae using sequential hydrothermal-assisted extraction followed by ultrasound and thermal technologies. Mar. Drugs. 2019;17:457. doi: 10.3390/md17080457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Alboofetileh M., Rezaei M., Tabarsa M., You S., Mariatti F., Cravotto G. Subcritical water extraction as an efficient technique to isolate biologically-active fucoidans from Nizamuddinia zanardinii. Int. J. Biol. Macromol. 2019;128:244–253. doi: 10.1016/j.ijbiomac.2019.01.119. [DOI] [PubMed] [Google Scholar]
- 125.Qin Y., Yuan Q., Zhang Y., Li J., Zhu X., Zhao L., Wen J., Liu J., Zhao L., Zhao J. Enzyme-assisted extraction optimization, characterization and antioxidant activity of polysaccharides from sea cucumber Phyllophorus proteus. Molecules. 2018;23:590. doi: 10.3390/molecules23030590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Wijesinghe W.A., Jeon Y.J. Enzyme-assistant extraction (EAE) of bioactive components: A useful approach for recovery of industrially important metabolites from seaweeds: A review. Fitoterapia. 2012;83:6–12. doi: 10.1016/j.fitote.2011.10.016. [DOI] [PubMed] [Google Scholar]
- 127.Ahn G., Lee W., Kim K.-N., Lee J.-H., Heo S.-J., Kang N., Lee S.-H., Ahn C.-B., Jeon Y.-J. A sulfated polysaccharide of Ecklonia cava inhibits the growth of colon cancer cells by inducing apoptosis. EXCLI J. 2015;14:294–306. doi: 10.17179/excli2014-676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Badrinathan S., Shiju T.M., Sharon Christa A.S., Arya R., Pragasam V. Purification and structural characterization of sulfated polysaccharide from Sargassum myriocystum and its efficacy in scavenging free radicals. Indian J. Pharm. Sci. 2012;74:549–555. doi: 10.4103/0250-474X.110600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Liu Y., Huang G. Extraction and derivatisation of active polysaccharides. J. Enzym. Inhib. Med. Chem. 2019;34:1690–1696. doi: 10.1080/14756366.2019.1660654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Zhao D., Xu J., Xu X. Bioactivity of fucoidan extracted from Laminaria japonica using a novel procedure with high yield. Food Chem. 2018;245:911–918. doi: 10.1016/j.foodchem.2017.11.083. [DOI] [PubMed] [Google Scholar]
- 131.Xing R., Liu S., Yu H., Chen X., Qin Y., Li K., Li P. Extraction and separation of fucoidan from Laminaria japonica with chitosan as extractant. BioMed Res. Int. 2013;2013:193689. doi: 10.1155/2013/193689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Ertani A., Francioso O., Tinti A., Schiavon M., Pizzeghello D., Nardi S. Evaluation of seaweed extracts from Laminaria and Ascophyllum nodosum spp. as biostimulants in Zea mays L. using a combination of chemical, biochemical and morphological approaches. Front. Plant Sci. 2018;9:428. doi: 10.3389/fpls.2018.00428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Lee J.M., Oh S.Y., Johnston T.V., Ku S., Ji G.E. Biocatalysis of fucodian in Undaria pinnatifida sporophyll using Bifidobacterium longum RD47 for production of prebiotic fucosylated oligosaccharide. Mar. Drugs. 2019;17:117. doi: 10.3390/md17020117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Somasundaram S.N., Shanmugam S., Subramanian B., Jaganathan R. Cytotoxic effect of fucoidan extracted from Sargassum cinereum on colon cancer cell line HCT-15. Int. J. Biol. Macromol. 2016;91 doi: 10.1016/j.ijbiomac.2016.06.084. [DOI] [PubMed] [Google Scholar]
- 135.Ustyuzhanina N.E., Ushakova N.A., Zyuzina K.A., Bilan M.I., Elizarova A.L., Somonova O.V., Madzhuga A.V., Krylov V.B., Preobrazhenskaya M.E., Usov A.I., et al. Influence of fucoidans on hemostatic system. Mar. Drugs. 2013;11:2444–2458. doi: 10.3390/md11072444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Saboural P., Chaubet F., Rouzet F., Al-Shoukr F., Azzouna R.B., Bouchemal N., Picton L., Louedec L., Maire M., Rolland L., et al. Purification of a low molecular weight fucoidan for SPECT molecular imaging of myocardial infarction. Mar. Drugs. 2014;12:4851–4867. doi: 10.3390/md12094851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Garcia-Vaquero M., Rajauria G., O’Doherty J.V., Sweeney T. Polysaccharides from macroalgae: Recent advances, innovative technologies and challenges in extraction and purification. Food Res. Int. 2017;99:1011–1020. doi: 10.1016/j.foodres.2016.11.016. [DOI] [PubMed] [Google Scholar]
- 138.Hahn T., Schulz M., Stadtmüller R., Zayed A., Muffler K., Lang S., Ulber R. Cationic dye for the specific determination of sulfated polysaccharides. Anal. Lett. 2016;49:1948–1962. doi: 10.1080/00032719.2015.1126839. [DOI] [Google Scholar]
- 139.Lee J.M., Shin Z.U., Mavlonov G.T., Abdurakhmonov I.Y., Yi T.-H. Solid-phase colorimetric method for the quantification of fucoidan. Appl. Biochem. Biotechnol. 2012;168:1019–1024. doi: 10.1007/s12010-012-9837-y. [DOI] [PubMed] [Google Scholar]
- 140.Palanisamy S., Vinosha M., Manikandakrishnan M., Anjali R., Rajasekar P., Marudhupandi T., Manikandan R., Vaseeharan B., Prabhu N.M. Investigation of antioxidant and anticancer potential of fucoidan from Sargassum polycystum. Int. J. Biol. Macromol. 2018;116:151–161. doi: 10.1016/j.ijbiomac.2018.04.163. [DOI] [PubMed] [Google Scholar]
- 141.Cong Q., Chen H., Liao W., Xiao F., Wang P., Qin Y., Dong Q., Ding K. Structural characterization and effect on anti-angiogenic activity of a fucoidan from Sargassum fusiforme. Carbohydr. Polym. 2016;136:899–907. doi: 10.1016/j.carbpol.2015.09.087. [DOI] [PubMed] [Google Scholar]
- 142.Li G., Row K.H. Magnetic molecularly imprinted polymers for recognition and enrichment of polysaccharides from seaweed. J. Sep. Sci. 2017;40:4765–4772. doi: 10.1002/jssc.201700947. [DOI] [PubMed] [Google Scholar]
- 143.Guthrie L., Wolfson S., Kelly L. The human gut chemical landscape predicts microbe-mediated biotransformation of foods and drugs. eLife. 2019;8:e42866. doi: 10.7554/eLife.42866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Wang J., Zhang Q., Zhang Z., Song H., Li P. Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2010;46:6–12. doi: 10.1016/j.ijbiomac.2009.10.015. [DOI] [PubMed] [Google Scholar]
- 145.Lee J.B., Hayashi K., Hashimoto M., Nakano T., Hayashi T. Novel antiviral fucoidan from sporophyll of Undaria pinnatifida (Mekabu) Chem. Pharm. Bull. 2004;52:1091–1094. doi: 10.1248/cpb.52.1091. [DOI] [PubMed] [Google Scholar]
- 146.Elizondo-Gonzalez R., Cruz-Suarez L.E., Ricque-Marie D., Mendoza-Gamboa E., Rodriguez-Padilla C., Trejo-Avila L.M. In vitro characterization of the antiviral activity of fucoidan from Cladosiphon okamuranus against Newcastle disease virus. Virol. J. 2012;9:307. doi: 10.1186/1743-422X-9-307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Jeong J.-W., Hwang S.J., Han M.H., Lee D.-S., Yoo J.S., Choi I.-W., Cha H.-J., Kim S., Kim H.-S., Kim G.-Y., et al. Fucoidan inhibits lipopolysaccharide-induced inflammatory responses in RAW 264.7 macrophages and zebrafish larvae. Mol. Cell. Toxicol. 2017;13:405–417. doi: 10.1007/s13273-017-0045-2. [DOI] [Google Scholar]
- 148.Klettner A. Fucoidan as a potential therapeutic for major blinding diseases - a hypothesis. Mar. Drugs. 2016;14:31. doi: 10.3390/md14020031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Barbosa A.I., Coutinho A.J., Costa Lima S.A., Reis S. Marine polysaccharides in pharmaceutical applications: Fucoidan and chitosan as key players in the drug delivery match field. Mar. Drugs. 2019;17:654. doi: 10.3390/md17120654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Wang P., Kankala R.K., Fan J., Long R., Liu Y., Wang S. Poly-L-ornithine/fucoidan-coated calcium carbonate microparticles by layer-by-layer self-assembly technique for cancer theranostics. J. Mater. Sci. Mater. Med. 2018;29:68. doi: 10.1007/s10856-018-6075-z. [DOI] [PubMed] [Google Scholar]
- 151.Wang X., Shan X., Dun Y., Cai C., Hao J., Li G., Cui K., Yu G. Anti-metabolic syndrome effects of fucoidan from Fucus vesiculosus via reactive oxygen species-mediated regulation of JNK, Akt, and AMPK signaling. Molecules. 2019;24:3319. doi: 10.3390/molecules24183319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Sharma G., Kar S., Basu Ball W., Ghosh K., Das P.K. The curative effect of fucoidan on visceral leishmaniasis is mediated by activation of MAP kinases through specific protein kinase C isoforms. Cell. Mol. Immunol. 2014;11:263–274. doi: 10.1038/cmi.2013.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Varikuti S., Jha B.K., Volpedo G., Ryan N.M., Halsey G., Hamza O.M., McGwire B.S., Satoskar A.R. Host-directed drug therapies for neglected tropical diseases caused by protozoan parasites. Front. Microbiol. 2018;9:2655. doi: 10.3389/fmicb.2018.02655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Jin J.O., Zhang W., Du J.Y., Wong K.W., Oda T., Yu Q. Fucoidan can function as an adjuvant in vivo to enhance dendritic cell maturation and function and promote antigen-specific T cell immune responses. PLoS ONE. 2014;9:e99396. doi: 10.1371/journal.pone.0099396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Tabarsa M., Dabaghian E.H., You S., Yelithao K., Cao R., Rezaei M., Alboofetileh M., Bita S. The activation of NF-κB and MAPKs signaling pathways of RAW264.7 murine macrophages and natural killer cells by fucoidan from Nizamuddinia zanardinii. Int. J. Biol. Macromol. 2020;148:56–67. doi: 10.1016/j.ijbiomac.2020.01.125. [DOI] [PubMed] [Google Scholar]
- 156.Wang P., Liu Z., Liu X., Teng H., Zhang C., Hou L., Zou X. Anti-metastasis effect of fucoidan from Undaria pinnatifida sporophylls in mouse hepatocarcinoma Hca-F cells. PLoS ONE. 2014;9:e106071. doi: 10.1371/journal.pone.0106071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Li Y., Zhao W., Wang L., Chen Y., Zhang H., Wang T., Yang X., Xing F., Yan J., Fang X. Protective effects of fucoidan against hydrogen peroxide-induced oxidative damage in porcine intestinal epithelial cells. Animals. 2019;9:1108. doi: 10.3390/ani9121108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Chen J.H., Lim J.D., Sohn E.H., Choi Y.S., Han E.T. Growth-inhibitory effect of a fucoidan from brown seaweed Undaria pinnatifida on Plasmodium parasites. Parasitol. Res. 2009;104:245–250. doi: 10.1007/s00436-008-1182-2. [DOI] [PubMed] [Google Scholar]
- 159.Jia Y., Sun Y., Weng L., Li Y., Zhang Q., Zhou H., Yang B. Low molecular weight fucoidan protects renal tubular cells from injury induced by albumin overload. Sci. Rep. 2016;6:31759. doi: 10.1038/srep31759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Chen C.-H., Sue Y.-M., Cheng C.-Y., Chen Y.-C., Liu C.-T., Hsu Y.-H., Hwang P.-A., Huang N.-J., Chen T.-H. Oligo-fucoidan prevents renal tubulointerstitial fibrosis by inhibiting the CD44 signal pathway. Sci. Rep. 2017;7:40183. doi: 10.1038/srep40183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Abdel-Daim M.M., Abushouk A.I., Bahbah E.I., Bungău S.G., Alyousif M.S., Aleya L., Alkahtani S. Fucoidan protects against subacute diazinon-induced oxidative damage in cardiac, hepatic, and renal tissues. Environ. Sci. Pollut. Res. 2020 doi: 10.1007/s11356-020-07711-w. Online ahead of print. [DOI] [PubMed] [Google Scholar]
- 162.Chen M.-C., Hsu W.-L., Hwang P.-A., Chou T.-C. Low molecular weight fucoidan inhibits tumor angiogenesis through downregulation of HIF-1/VEGF signaling under hypoxia. Mar. Drugs. 2015;13:4436–4451. doi: 10.3390/md13074436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Marinval N., Saboural P., Haddad O., Maire M., Bassand K., Geinguenaud F., Djaker N., Ben Akrout K., Lamy de la Chapelle M., Robert R., et al. Identification of a pro-angiogenic potential and cellular uptake mechanism of a LMW highly sulfated fraction of fucoidan from Ascophyllum nodosum. Mar. Drugs. 2016;14:185. doi: 10.3390/md14100185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Cheng Y., Sibusiso L., Hou L., Jiang H., Chen P., Zhang X., Wu M., Tong H. Sargassum fusiforme fucoidan modifies the gut microbiota during alleviation of streptozotocin-induced hyperglycemia in mice. Int. J. Biol. Macromol. 2019;131:1162–1170. doi: 10.1016/j.ijbiomac.2019.04.040. [DOI] [PubMed] [Google Scholar]
- 165.Li B., Juenet M., Aid-Launais R., Maire M., Ollivier V., Letourneur D., Chauvierre C. Development of polymer microcapsules functionalized with fucoidan to target p-selectin overexpressed in cardiovascular diseases. Adv. Healthc. Mater. 2017;6:1601200. doi: 10.1002/adhm.201601200. [DOI] [PubMed] [Google Scholar]
- 166.Kim Y.-I., Oh W.-S., Song P.H., Yun S., Kwon Y.-S., Lee Y.J., Ku S.-K., Song C.-H., Oh T.-H. Anti-photoaging effects of low molecular-weight fucoidan on ultraviolet B-irradiated mice. Mar. Drugs. 2018;16:286. doi: 10.3390/md16080286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Kim J.H., Lee J.-E., Kim K.H., Kang N.J. Beneficial effects of marine algae-derived carbohydrates for skin health. Mar. Drugs. 2018;16:459. doi: 10.3390/md16110459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Fitton J.H., Dell’Acqua G., Gardiner V.-A., Karpiniec S.S., Stringer D.N., Davis E. Topical benefits of two fucoidan-rich extracts from marine macroalgae. Cosmetics. 2015;2:66–81. doi: 10.3390/cosmetics2020066. [DOI] [Google Scholar]
- 169.Song Y.S., Li H., Balcos M.C., Yun H.-Y., Baek K.J., Kwon N.S., Choi H.-R., Park K.-C., Kim D.-S. Fucoidan promotes the reconstruction of skin equivalents. Korean J. Physiol. Pharmacol. 2014;18:327–331. doi: 10.4196/kjpp.2014.18.4.327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Venkatesan J., Anil S., Kim S.-K., Shim M.S. Seaweed polysaccharide-based nanoparticles: Preparation and applications for drug delivery. Polymers. 2016;8:30. doi: 10.3390/polym8020030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Pinheiro A.C., Bourbon A.I., Cerqueira M.A., Maricato É., Nunes C., Coimbra M.A., Vicente A.A. Chitosan/fucoidan multilayer nanocapsules as a vehicle for controlled release of bioactive compounds. Carbohydr. Polym. 2015;115:1–9. doi: 10.1016/j.carbpol.2014.07.016. [DOI] [PubMed] [Google Scholar]
- 172.Choi D.G., Venkatesan J., Shim M.S. Selective anticancer therapy using pro-oxidant drug-loaded chitosan-fucoidan nanoparticles. Int. J. Mol. Sci. 2019;20:3220. doi: 10.3390/ijms20133220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Tsai M.-h., Chuang C.-c., Chen C.-c., Yen H.-j., Cheng K.-m., Chen X.-a., Shyu H.-f., Lee C.-y., Young J.-j., Kau J.-h. Nanoparticles assembled from fucoidan and trimethylchitosan as anthrax vaccine adjuvant: In vitro and in vivo efficacy in comparison to CpG. Carbohydr. Polym. 2020;236:116041. doi: 10.1016/j.carbpol.2020.116041. [DOI] [PubMed] [Google Scholar]
- 174.Venkatesan J., Singh S.K., Anil S., Kim S.-K., Shim M.S. Preparation, characterization and biological applications of biosynthesized silver nanoparticles with chitosan-fucoidan coating. Molecules. 2018;23:1429. doi: 10.3390/molecules23061429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Hwang P.-A., Yan M.-D., Lin H.-T.V., Li K.-L., Lin Y.-C. Toxicological evaluation of low molecular weight fucoidan in vitro and in vivo. Mar. Drugs. 2016;14:121. doi: 10.3390/md14070121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Zhu Z., Zhu B., Ai C., Lu J., Wu S., Liu Y., Wang L., Yang J., Song S., Liu X. Development and application of a HPLC-MS/MS method for quantitation of fucosylated chondroitin sulfate and fucoidan in sea cucumbers. Carbohydr. Res. 2018;466:11–17. doi: 10.1016/j.carres.2018.07.001. [DOI] [PubMed] [Google Scholar]
- 177.Yu L., Xue C., Chang Y., Xu X., Ge L., Liu G., Wang Y. Structure elucidation of fucoidan composed of a novel tetrafucose repeating unit from sea cucumber Thelenota ananas. Food Chem. 2014;146:113–119. doi: 10.1016/j.foodchem.2013.09.033. [DOI] [PubMed] [Google Scholar]
- 178.Zhao X., Guo F., Hu J., Zhang L., Xue C., Zhang Z., Li B. Antithrombotic activity of oral administered low molecular weight fucoidan from Laminaria Japonica. Thromb. Res. 2016;144:46–52. doi: 10.1016/j.thromres.2016.03.008. [DOI] [PubMed] [Google Scholar]
- 179.Tsai H.L., Tai C.J., Huang C.W., Chang F.R., Wang J.Y. Efficacy of low-molecular-weight fucoidan as a supplemental therapy in metastatic colorectal cancer patients: A double-blind randomized controlled trial. Mar. Drugs. 2017;15:122. doi: 10.3390/md15040122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Jonsson M., Allahgholi L., Sardari R.R.R., Hreggviethsson G.O., Nordberg Karlsson E. Extraction and modification of macroalgal polysaccharides for current and next-generation applications. Molecules. 2020;25:930. doi: 10.3390/molecules25040930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Cumashi A., Ushakova N.A., Preobrazhenskaya M.E., D’Incecco A., Piccoli A., Totani L., Tinari N., Morozevich G.E., Berman A.E., Bilan M.I., et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology. 2007;17:541–552. doi: 10.1093/glycob/cwm014. [DOI] [PubMed] [Google Scholar]
- 182.Silchenko A.S., Ustyuzhanina N.E., Kusaykin M.I., Krylov V.B., Shashkov A.S., Dmitrenok A.S., Usoltseva R.V., Zueva A.O., Nifantiev N.E., Zvyagintseva T.N. Expression and biochemical characterization and substrate specificity of the fucoidanase from Formosa algae. Glycobiology. 2017;27:254–263. doi: 10.1093/glycob/cww138. [DOI] [PubMed] [Google Scholar]
- 183.Silchenko A.S., Rasin A.B., Kusaykin M.I., Kalinovsky A.I., Miansong Z., Changheng L., Malyarenko O., Zueva A.O., Zvyagintseva T.N., Ermakova S.P. Structure, enzymatic transformation, anticancer activity of fucoidan and sulphated fucooligosaccharides from Sargassum horneri. Carbohydr. Polym. 2017;175:654–660. doi: 10.1016/j.carbpol.2017.08.043. [DOI] [PubMed] [Google Scholar]
- 184.Colin S., Deniaud E., Jam M., Descamps V., Chevolot Y., Kervarec N., Yvin J.C., Barbeyron T., Michel G., Kloareg B. Cloning and biochemical characterization of the fucanase FcnA: Definition of a novel glycoside hydrolase family specific for sulfated fucans. Glycobiology. 2006;16:1021–1032. doi: 10.1093/glycob/cwl029. [DOI] [PubMed] [Google Scholar]
- 185.Nagao T., Arai Y., Yamaoka M., Komatsu F., Yagi H., Suzuki H., Ohshiro T. Identification and characterization of the fucoidanase gene from Luteolibacter algae H18. J. Biosci. Bioeng. 2018;126:567–572. doi: 10.1016/j.jbiosc.2018.05.016. [DOI] [PubMed] [Google Scholar]
- 186.Bakunina I., Nedashkovskaia O.I., Alekseeva S.A., Ivanova E.P., Romanenko L.A., Gorshkova N.M., Isakov V.V., Zviagintseva T.N., Mikhailov V.V. Degradation of fucoidan by the marine proteobacterium Pseudoalteromonas citrea. Mikrobiologiia. 2002;71:49–55. [PubMed] [Google Scholar]
- 187.Wu Q., Zhang M., Wu K., Liu B., Cai J., Pan R. Purification and characteristics of fucoidanase obtained from Dendryphiella arenaria TM94. J. Appl. Phycol. 2011;23:197–203. doi: 10.1007/s10811-010-9588-5. [DOI] [Google Scholar]
- 188.Sakai T., Kimura H., Kojima K., Shimanaka K., Ikai K., Kato I. Marine bacterial sulfated fucoglucuronomannan (SFGM) lyase digests brown algal SFGM into trisaccharides. Mar. Biotechnol. 2003;5:70–78. doi: 10.1007/s10126-002-0056-3. [DOI] [PubMed] [Google Scholar]
- 189.Sakai T., Ishizuka K., Shimanaka K., Ikai K., Kato I. Structures of oligosaccharides derived from Cladosiphon okamuranus fucoidan by digestion with marine bacterial enzymes. Mar. Biotechnol. 2003;5:536–544. doi: 10.1007/s10126-002-0107-9. [DOI] [PubMed] [Google Scholar]
- 190.Ohshiro T., Ohmoto Y., Ono Y., Ohkita R., Miki Y., Kawamoto H., Izumi Y. Isolation and characterization of a novel fucoidan-degrading microorganism. Biosci. Biotechnol. Biochem. 2010;74:1729–1732. doi: 10.1271/bbb.100327. [DOI] [PubMed] [Google Scholar]
- 191.Bilan M.I., Kusaykin M.I., Grachev A.A., Tsvetkova E.A., Zvyagintseva T.N., Nifantiev N.E., Usov A.I. Effect of enzyme preparation from the marine mollusk Littorina kurila on fucoidan from the brown alga Fucus distichus. Biochemistry. 2005;70:1321–1326. doi: 10.1007/s10541-005-0264-3. [DOI] [PubMed] [Google Scholar]
- 192.Kim W.J., Kim S.M., Lee Y.H., Kim H.G., Kim H.K., Moon S.H., Suh H.H., Jang K.H., Park Y.I. Isolation and characterization of marine bacterial strain degrading fucoidan from korean Undaria pinnatifida Sporophylls. J. Microbiol. Biotechnol. 2008;18:616–623. [PubMed] [Google Scholar]
- 193.Kim W.J., Park J.W., Park J.K., Choi D.J., Park Y.I. Purification and characterization of a fucoidanase (FNase S) from a marine bacterium Sphingomonas paucimobilis PF-1. Mar. Drugs. 2015;13:4398–4417. doi: 10.3390/md13074398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Kitamura K., Matsuo M., Tsuneo Y. Enzymic degradation of fucoidan by fucoidanase from the hepatopancreas of Patinopecten yessoensis. Biosci. Biotechnol. Biochem. 1992;56:490–494. doi: 10.1271/bbb.56.490. [DOI] [PubMed] [Google Scholar]
- 195.Daniel R., Berteau O., Chevolot L., Varenne A., Gareil P., Goasdoue N. Regioselective desulfation of sulfated l-fucopyranoside by a new sulfoesterase from the marine mollusk Pecten maximus. Eur. J. Biochem. 2001;268:5617–5626. doi: 10.1046/j.1432-1033.2001.02497.x. [DOI] [PubMed] [Google Scholar]
- 196.Berteau O., McCort I., Goasdoué N., Tissot B., Daniel R. Characterization of a new alpha-L-fucosidase isolated from the marine mollusk Pecten maximus that catalyzes the hydrolysis of alpha-L-fucose from algal fucoidan (Ascophyllum nodosum) Glycobiology. 2002;12:273–282. doi: 10.1093/glycob/12.4.273. [DOI] [PubMed] [Google Scholar]
- 197.Dong S., Chang Y., Shen J., Xue C., Chen F. Purification, expression and characterization of a novel α-l-fucosidase from a marine bacteria Wenyingzhuangia fucanilytica. Protein Expr. Purif. 2017;129:9–17. doi: 10.1016/j.pep.2016.08.016. [DOI] [PubMed] [Google Scholar]
- 198.Tanaka R., Mizutani Y., Shibata T., Miyake H., Iehata S., Mori T., Kuroda K., Ueda M. Genome sequence of Formosa haliotis strain MA1, a brown alga-degrading bacterium isolated from the gut of Abalone Haliotis gigantea. Genome Announc. 2016;4:e01312-16. doi: 10.1128/genomeA.01312-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Chen F., Chang Y., Dong S., Xue C. Wenyingzhuangia fucanilytica sp. nov., a sulfated fucan utilizing bacterium isolated from shallow coastal seawater. Int. J. Syst. Evol. Microbiol. 2016;66:3270–3275. doi: 10.1099/ijsem.0.001184. [DOI] [PubMed] [Google Scholar]
- 200.Li J., Cheng Y., Wang D., Li J., Wang Y., Han W., Li F. Draft genome sequence of the polysaccharide-degrading marine bacterium Pseudoalteromonas sp. Strain A601. Genome Announc. 2017;5:e00590-17. doi: 10.1128/genomeA.00590-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Cao H.T.T., Mikkelsen M.D., Lezyk M.J., Bui L.M., Tran V.T.T., Silchenko A.S., Kusaykin M.I., Pham T.D., Truong B.H., Holck J., et al. Novel enzyme actions for sulphated galactofucan depolymerisation and a new engineering strategy for molecular stabilisation of fucoidan degrading enzymes. Mar. Drugs. 2018;16:422. doi: 10.3390/md16110422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Tran P.H.L., Duan W., Tran T.T.D. Fucoidan-based nanostructures: A focus on its combination with chitosan and the surface functionalization of metallic nanoparticles for drug delivery. Int. J. Pharm. 2019;575:118956. doi: 10.1016/j.ijpharm.2019.118956. [DOI] [PubMed] [Google Scholar]
- 203.Jesumani V., Du H., Pei P., Aslam M., Huang N. Comparative study on skin protection activity of polyphenol-rich extract and polysaccharide-rich extract from Sargassum vachellianum. PLoS ONE. 2020;15:e0227308. doi: 10.1371/journal.pone.0227308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Brunt E.G., Burgess J.G. The promise of marine molecules as cosmetic active ingredients. Int. J. Cosmet. Sci. 2018;40:1–15. doi: 10.1111/ics.12435. [DOI] [PubMed] [Google Scholar]
- 205.Ferreira R.M., Ramalho Ribeiro A., Patinha C., Silva A.M.S., Cardoso S.M., Costa R. Water extraction kinetics of bioactive compounds of Fucus vesiculosus. Molecules. 2019;24:3408. doi: 10.3390/molecules24183408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Michel G., Tonon T., Scornet D., Cock J.M., Kloareg B. The cell wall polysaccharide metabolism of the brown alga Ectocarpus siliculosus. Insights into the evolution of extracellular matrix polysaccharides in eukaryotes. New Phytol. 2010;188:82–97. doi: 10.1111/j.1469-8137.2010.03374.x. [DOI] [PubMed] [Google Scholar]
- 207.Chi S., Liu T., Wang X., Wang R., Wang S., Wang G., Shan G., Liu C. Functional genomics analysis reveals the biosynthesis pathways of important cellular components (alginate and fucoidan) of Saccharina. Curr. Genet. 2018;64:259–273. doi: 10.1007/s00294-017-0733-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Shao Z., Zhang P., Lu C., Li S., Chen Z., Wang X., Duan D. Transcriptome sequencing of Saccharina Japonica Sporophytes during whole developmental periods reveals regulatory networks underlying alginate and mannitol biosynthesis. BMC Genom. 2019;20:975. doi: 10.1186/s12864-019-6366-x. [DOI] [PMC free article] [PubMed] [Google Scholar]