

The blood vessels of the central nervous system tightly control the movement of ions, molecules, and cells between the blood and tissue. This “blood–brain barrier” is vital for neural homeostasis and protection. This review discusses current knowledge of the blood–brain barrier, emphasizing key unanswered questions.
Abstract
The blood vessels vascularizing the central nervous system exhibit a series of distinct properties that tightly control the movement of ions, molecules, and cells between the blood and the parenchyma. This “blood–brain barrier” is initiated during angiogenesis via signals from the surrounding neural environment, and its integrity remains vital for homeostasis and neural protection throughout life. Blood–brain barrier dysfunction contributes to pathology in a range of neurological conditions including multiple sclerosis, stroke, and epilepsy, and has also been implicated in neurodegenerative diseases such as Alzheimer’s disease. This review will discuss current knowledge and key unanswered questions regarding the blood–brain barrier in health and disease.
Blood vessels provide the vital infrastructure for delivery of oxygen and essential nutrients throughout the body, and the term “blood–brain barrier” (BBB) is used to describe the unique characteristics of the blood vessels that vascularize the central nervous system (CNS; Saunders et al., 2008; Zlokovic, 2008; Obermeier et al., 2013). The BBB is not a single physical entity but rather the combined function of a series of physiological properties possessed by endothelial cells (ECs) that limit vessel permeability. The BBB tightly regulates the movement of ions, molecules, and cells between the blood and the parenchyma and is thus critical for neuronal function and protection. The interaction of ECs with different neural and immune cells is commonly referred to as the neurovascular unit (NVU; Fig. 1 A). The complex properties that define the BBB are often altered in disease states, and BBB dysfunction has been identified as a critical component in several neurological conditions. This review will discuss BBB development, regulation, and dysfunction, emphasizing important unanswered questions.
Figure 1.

Cellular and molecular properties of the BBB. (A) A schematic comparison of the BBB capillaries with the continuous nonfenestrated, continuous fenestrated, and discontinuous capillaries found in peripheral organs. (B–F) Schematics of the molecular composition of junctional complexes of BBB ECs (B) and of ECs in peripheral organs (C), peripheral endothelial fenestra (D), and transport mechanisms in CNS ECs (E) and peripheral ECs (F). (G and H) Electron micrographs of a mouse brain EC (G) and a mouse muscle EC, which is densely packed with vesicles (arrows; H). BCRP, breast cancer resistance protein; GLUT1, glucose transporter 1; ICAM, intercellular adhesion molecule; JAM, junctional adhesion molecule; LAT1, L-type amino acid transporter 1; MRP, multidrug resistance-associated protein; PECAM-1, platelet and EC adhesion molecule 1; P-GP, p-glycoprotein; VE-cadherin, vascular endothelial cadherin; P-sel, P-selectin; E-sel, E-selectin.
The NVU
ECs
A cross-section of an artery or vein might contain dozens of ECs, while in the smallest capillaries, a single EC forms the vessel circumference (Aird, 2007). In all tissues, adherens junctions, composed of vascular endothelial cadherin and catenins, comprise the basic cellular adhesions between ECs, supporting the integrity of the vascular tube and regulating tensile forces. PECAM1 is a critical regulator of EC adhesion, promoting adherens junction formation (Biswas et al., 2006; Privratsky and Newman, 2014). CNS ECs are further specialized to restrict paracellular and transcellular movement of solutes.
Tight junctions (TJs)
TJs are cell adhesions consisting of multiple transmembrane proteins that directly interact via their extracellular components, linking two cells’ membranes together (Furuse, 2010; Fig. 1 B). CNS TJs are specialized in their molecular and structural P-face composition to form a high-resistance electrical barrier, and the specific combination of TJ proteins at the BBB determines its paracellular permeability.
The composition of claudins, a family of 27 four-pass transmembrane proteins, within a TJ is thought to determine the size and charge selectivity of paracellular permeability (Amasheh et al., 2005; Hou et al., 2006; Furuse et al., 1999). Claudin 5 (CLDN5) is the most abundant claudin at the BBB, and Cldn5 knockout mice exhibit size-selective leakage of the BBB and die at birth (Morita et al., 1999; Nitta et al., 2003). ECs in peripheral vascular beds also express CLDN5, and thus its expression alone is not sufficient for barrier formation. Other key components of TJs include claudin 12, occludin, and junctional adhesion molecules. Cytoplasmic proteins including ZO-1, ZO-2, ZO-3, cingulin, JACOP, MAG1, and MUPP1 aid TJ formation, binding TJs to the cytoskeleton, adherens junctions, and polarity complexes (Umeda et al., 2004; Tietz and Engelhardt, 2015; Sawada, 2013). It is still unknown why CLDN5 and ZO-1 expression does not confer the same low paracellular permeability in peripheral vessels as in the CNS. Expression data suggest that the answer might lie in the CNS-specific enrichment of certain cytoplasmic adaptors (e.g., JACOP, MPP7) and tricellular TJ molecules such as LSR and MARVELD2 (Daneman et al., 2010a; Sohet et al., 2015).
Transcellular permeability
Peripheral ECs possess properties that confer transcellular permeability, including high rates of caveolin-mediated transcytosis, diaphragm-containing pores termed fenestrae, or large discontinuities or gaps in the endothelial layer (Aird, 2007; Fig. 1 A). In contrast, CNS ECs form a continuous lining that lacks fenestrations and has low levels of transcytosis, properties that greatly limit transcellular permeability (Fig. 1, A–H). MFSD2A, enriched in CNS ECs, limits caveolin-dependent transcytosis by regulating EC lipid composition (Ben-Zvi et al., 2014; Nguyen et al., 2014; Andreone et al., 2017). Plasmalemma vesicle-associated protein (PLVAP) is important both for vesicle formation and fenestrations. Its down-regulation in CNS ECs, along with up-regulation of MFSD2A, coincides with BBB formation during embryogenesis (Hallmann et al., 1995; Hnasko et al., 2002; Chow and Gu, 2017).
Transporters
Numerous transporters are enriched in brain ECs, which generally fall into two categories: efflux and solute transporters (Miller, 2015; Nałęcz, 2017; Strazielle and Ghersi-Egea, 2015; Fig. 1 E).
Efflux transporters, concentrated on the luminal side of the membrane, use ATP hydrolysis to transport a wide range of small molecules up their concentration gradients back into the blood (Shen and Zhang, 2010). MDR1/P-glycoprotein (PGP) and breast cancer resistance protein are the most abundant BBB efflux proteins and limit entry of many xenobiotics and endogenous molecules, including steroids such as aldosterone (Hindle et al., 2017).
Solute transporters carry specific substrates down their concentration gradients, ensuring barrier passage to specific nutrients, such as glucose, that are vital for energy and homeostasis (Simpson et al., 2007). Transport of glucose, lactate, amino acids, and fatty acids occurs via GLUT1 (Slc2a1), MCT1 (Slc16a1), LAT1 (Slc7a5), and MFSD2A, respectively (Boado et al., 1999; Cornford et al., 1994; Kido et al., 2000; Nguyen et al., 2014). Other transporters provide receptor-mediated vesicular transport, including the transferrin receptor (TFR1) and low-density lipoprotein receptors (Jefferies et al., 1984; Méresse et al., 1989). Substrate-specific solute transporters can also be important for removing molecules from the CNS; lipoprotein receptor-related protein-1 (LRP1) is a critical transporter for eliminating β-amyloid (Shibata et al., 2000; Storck et al., 2016).
Leukocyte adhesion molecules
Leukocyte adhesion molecules on EC surfaces initiate binding of leukocytes, the first step of their entrance into tissues (Bevilacqua, 1993). Healthy CNS ECs exhibit lower leukocyte adhesion molecule expression compared with peripheral ECs (Daneman et al., 2010a), and thus there is minimal leukocyte crossing of the BBB in health (Fig. 1, E and F; and Fig. 2 A). Instead, CNS immune surveillance by lymphocytes in health occurs primarily at the blood–CSF interfaces of the meninges and choroid plexus (Ransohoff and Engelhardt, 2012; Kipnis et al., 2012; Shechter et al., 2013; Box 1).
Figure 2.

Dysfunction of the BBB in disease. (A and B) Schematic representation of the NVU in health and disease. (A) In health, CNS ECs exhibit TJs, low rates of transcytosis, and low expression of leukocyte adhesion molecules. Pericytes embedded in the BM help to maintain the barrier, and astrocyte endfeet contact the BM. (B) In disease, TJs are internalized or down-regulated, rates of transcytosis increase, increased leukocyte adhesion molecule expression leads to increased leukocyte extravasation, the BM degrades, and pericytes and astrocytes less tightly cover the ECs. Made with BioRender. (C–F) BBB disruption in models of MS, traumatic brain injury, and stroke. Sections showing BBB leakage to a sulfo-N-hydroxysulfosuccinimide-biotin tracer (green) in three disease models. (C and D) A section of spinal cord from a healthy mouse (C) and from the EAE model of MS (D). (E and F) The contralateral (E) and ipsilateral (F) hemispheres in a coronal section of the middle-cerebral artery occlusion model of ischemic stroke. (G–I) BBB leakage and edema in human cases of MS, stroke, and epilepsy. T1 weighted MRI images with gadolinium enhancement to show BBB leakage in (G) MS lesions and (H) stroke infarct. (I) T1 weighted and fluid attenuation inversion recovery (FLAIR) MRI images showing edema in epilepsy. Images courtesy of Dr. John Hesselink, University of California, San Diego, San Diego, CA.
Box 1. Barriers of the CNS
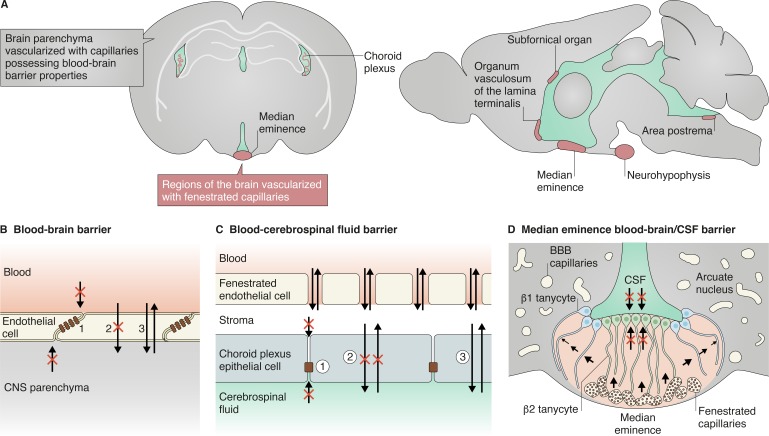
There are small regions of the brain that lack an endothelial BBB and are instead vascularized by permeable fenestrated capillaries. In these regions, a specialized glial barrier takes on the role of the endothelial BBB. (A) Among these regions is the choroid plexus, the structure that generates most of the cerebrospinal fluid. The specialized ependymal epithelial cells of the choroid plexus surround its fenestrated capillaries and filter the fluid that enters through fenestrae to generate the cerebrospinal fluid. The choroid plexus epithelial cells possess similar properties as the ECs of the BBB. (B and C) These properties include (compare panel B to C): (1) a dense formation of junctional complexes that restrict paracellular diffusion of hydrophilic solutes; (2) expression of efflux transporters and low rates of transcytosis that limit transcellular movement of molecules; and (3) expression of selective transporters that import necessary nutrients or export wastes (Marques et al., 2017). CVOs are vascularized by fenestrated capillaries and allow a small subset of neurons and glia to sense blood-derived signals or secrete hormones into the blood to regulate peripheral processes such as fluid homeostasis, osmoregulation, body temperature, energy balance, and inflammation. The subfornical organ, area postrema, and organum vasculosum of the lamina terminalis are the sensory CVOs, while the median eminence and neurohypophysis are the secretory CVOs. Each of these CVOs possess a glia-derived, cellular barrier generated by tanycytes or tanycyte-like cells that limit further diffusion of blood-derived solutes into neighboring regions or the cerebrospinal fluid (Ganong, 2000; Miyata, 2015). (D) In the median eminence, β1 tanycytes limit diffusion of solutes originating from the ventrally localized fenestrated capillaries into the arcuate nucleus while β2 tanycytes restrict chemical exchange between the median eminence and CSF (Miyata, 2015; Langlet et al., 2013).
The NVU
The luminal surface of the capillary endothelium is covered by the EC glycocalyx (Ausprunk et al., 1981a, b; Pillinger and Kam, 2017). Brain ECs have a denser glycocalyx than peripheral vasculature; average glycocalyx coverage is 40.1% in brain vessels compared with 15.1% and 3.2% in cardiac and pulmonary vessels, respectively (Ando et al., 2018). This dense network of luminal glycoproteins prevents larger molecules from interacting with the EC. While small dyes such as fluorescein (376 daltons) and Alexa Fluor (643 daltons) permeate the glycocalyx, dextrans (40–150 kD) penetrate <60% of its volume (Kutuzov et al., 2018). In disease, glycocalyx degradation is associated with more severe BBB leakage in models of multiple sclerosis (MS) and cardiac arrest (DellaValle et al., 2018; Zhu et al., 2018).
The abluminal surface of the EC is covered by the basal lamina (Fig. 1 A), a structural matrix of laminins, fibronectin, collagens, tenascin, and proteoglycans. This basement membrane (BM) surrounds ECs and pericytes, acting as an interface for the binding of molecules and migration of cells, while also limiting passage of macromolecules (Del Zoppo et al., 2006). The BM consists of two layers: the inner vascular BM secreted by ECs and pericytes, and the outer glial BM secreted by astrocytes (Sorokin, 2010). These BMs are merged surrounding capillaries but separate at post-capillary venules, creating a CSF-drained perivascular space for immune surveillance (Engelhardt and Ransohoff, 2012).
Mural cells—vascular smooth muscle cells (VSMCs) and pericytes—are found on the abluminal side of blood vessels in all tissues. VSMCs line all larger vessels but are more abundant on arteries and arterioles, forming a complete layer around them (Smyth et al., 2018; Vanlandewijck et al., 2018; Armulik et al., 2011). VSMC myosin fibers regulate blood flow via vasoconstriction and vasodilation (Aird, 2007). Pericytes are embedded in the BM and form an incomplete layer on the surface of CNS micro-vessels (Fig. 1 A). Pericytes play a key role in the regulation of angiogenesis, vascular remodeling, vascular tone, and BBB formation (Daneman et al., 2010a; Armulik et al., 2005, 2010; Winkler et al., 2011). Perivascular fibroblasts are found in the walls of large vessels (Vanlandewijck et al., 2018); however, their role in cerebrovascular function remains unexplored.
Astrocytes extend cellular processes terminating in endfeet that ensheath synapses, nodes of Ranvier, and ECs, contacting the BM around parenchymal vessels (Fig. 1 A). This astrocyte–endothelial interaction is critical in regulating blood flow (Mishra et al., 2016). Several groups have shown that CSF flows between the BM and astrocyte endfeet of arteries and capillaries, with arteriole pulsations driving bulk fluid flow through the parenchyma, although others have argued about the extent of bulk flow (Abbott et al., 2018; Hladky and Barrand, 2019). This “glymphatic” system helps to clear interstitial solutes such as amyloid via paravenous drainage pathways (Iliff et al., 2012; Xie et al., 2013; Mestre et al., 2018) and has been visualized in human patients via magnetic resonance imaging (MRI; Meng et al., 2019; Fultz et al., 2019). Expression of water channel aquaporin-4 in astrocyte endfeet has been reported to play a critical role in the movement of CSF into the parenchyma (Haj-Yasein et al., 2011; Iliff et al., 2012; Mestre et al., 2018).
CNS-associated macrophages, which express a gene signature of Mrc1 (CD206), Pf4, Cbr2, Ms4a7, and Stab1, include choroid plexus, dural, leptomeningeal, and perivascular macrophages (Kierdorf et al., 2019; Jordão et al., 2019). Perivascular macrophages are elongated cells residing between the astrocytic endfeet and parenchymal vessels (primarily arteries and veins). While nonmotile, they extend processes along the perivascular space, providing the first line of defense by collecting debris (Hickey and Kimura, 1988; Prinz et al., 2017). Microglia, derived from yolk-sac progenitor cells (Takahashi et al., 1989; Alliot et al., 1999), reside within the CNS parenchyma. They possess a highly ramified morphology and perform immune surveillance, phagocytosing infectious agents that evade the barrier (Streit et al., 2005; Prinz et al., 2011). Microglia have also been shown to regulate BBB resealing following vascular injury and disease (Fernández-López et al., 2016; Lou et al., 2016). In disease states, leukocytes such as neutrophils and T cells can interact with the BBB, increasing permeability via release of cytokines, reactive oxygen species, and other mediators of barrier dysfunction (Hudson et al., 2005; Persidsky et al., 1999).
Thus, the BBB is a series of structural, transport, and metabolic barriers that together limit CNS entry of nonspecific molecules while ensuring the delivery of specific nutrients, thereby controlling the extracellular environment. Several important questions remain. What exactly gets through the barrier, how much, and by which route(s)? The barrier is not absolute. Small, nonpolar molecules enter unrestricted through passive diffusion unless they are substrates of efflux transporters. In contrast, large or polar molecules are greatly restricted in access unless they are substrates of specific nutrient transporters. However, even large molecules enter the CNS parenchyma at 0.1% of their blood concentration through an unsaturable mechanism (Yu and Watts, 2013; Poduslo et al., 1994), likely via nonspecific transcytosis, which occurs at low rates. Future work fully characterizing the substrate specificity of BBB transporters and their dynamic response to various stimuli may enable manipulation of these transporters for CNS drug delivery.
There is heterogeneity of gene expression among different branches of the vascular tree (Macdonald et al., 2010; Vanlandewijck et al., 2018; Murugesan et al., 2011). It is thought that this heterogeneity enables capillaries, arterioles, and venules to be specialized for regulation of solute transport, blood flow, and inflammation, respectively. But what is the relevance of this arteriovenous zonation in terms of barrier function? How is this phenotypic continuum programmed during development?
It is also currently unknown whether there is regional heterogeneity of the BBB. Several regions of the CNS termed circumventricular organs (CVOs)—the area postrema, subfornical organ, pineal gland, and median eminence of the hypothalamus—have fenestrated capillaries that lack BBB properties (Box 1; Gross, 1992). This vascular permeability allows for the exchange of sensory or secretory signaling molecules between the brain and blood, enabling CVO-mediated regulation of body homeostasis. Much less is known about whether there are region-specific differences among areas with a functional BBB, including the cortex, hippocampus, cerebellum, and white matter tracks, and whether BBB heterogeneity might contribute to the specialized function of a particular brain region or render that region more vulnerable to disease.
BBB formation and regulation
How BBB properties are regulated in development and maintained in adulthood remains a fundamental field of study (Blanchette and Daneman, 2015). Transplanted CNS tissue is sufficient to induce BBB-like properties in the gut endothelium in vivo (Stewart and Wiley, 1981), suggesting a role for the neural microenvironment in BBB formation. Transplantation of astrocytes into nonneural tissues of adult rats induces barrier properties in local ECs (Janzer and Raff, 1987), and several astrocyte-secreted proteins are sufficient to induce EC barrier properties in vitro and in vivo, including Sonic hedgehog, angiotensin, and basic fibroblast growth factor (Alvarez et al., 2011; Sobue et al., 1999; Wosik et al., 2007). However, barrier properties arise during development before astrogliogenesis takes place (Ben-Zvi et al., 2014; Daneman et al., 2010a; Sohet et al., 2015; Sauvageot and Stiles, 2002), delaying astrocytic contact with ECs does not affect barrier formation (Saunders et al., 2016), and laser ablation of astrocyte endfeet in adult mice does not induce BBB leakage (Kubotera et al., 2019). These data suggest that astrocytes are not necessary for BBB formation, but perhaps provide dynamic BBB regulation in response to specific stimuli. For instance, reactive astrocytes have been shown to be critical for BBB repair following neurological disease (Bush et al., 1999).
Neural progenitor–derived Wnt signaling induces BBB properties during the angiogenic program (Daneman et al., 2009; Liebner et al., 2008; Stenman et al., 2008; Ye et al., 2009; Wang et al., 2012; Zhou and Nathans, 2014; Cho et al., 2017). Loss of Wnt signaling disrupts angiogenesis specifically in the CNS, reducing the expression of TJ proteins and solute transporters while increasing PLVAP (Daneman et al., 2009; Liebner et al., 2008; Stenman et al., 2008). Interestingly, β-catenin activation in the more permeable CVO vessels is sufficient to induce BBB properties (Benz et al., 2019; Wang et al., 2019). These data suggest that the same signal that drives angiogenic invasion of the CNS also induces initial BBB properties within the endothelium.
Pericytes are also essential in BBB development, and EC recruitment of pericytes is concomitant with development of barrier properties. The BBB fails to completely seal in mice lacking CNS pericytes, as they inhibit nonspecific transcytosis and leukocyte adhesion molecule expression (Daneman et al., 2010b; Armulik et al., 2010).
Thus, the BBB is regulated by a series of different cellular interactions: BBB “tight” properties are induced during the angiogenic program by Wnt signaling, “leaky” properties are inhibited by pericytes, and the overall phenotype of the BBB can be influenced by astrocytes, pericytes, and other cell types throughout life.
Important questions still remain. How is the induction of different BBB properties coordinated? Interestingly, Wnt signaling induces endothelial secretion of platelet-derived growth factor B, the key ligand for pericyte recruitment (Reis et al., 2012), suggesting that induction of different BBB properties is tightly coordinated via Wnt-mediated pericyte recruitment. Are the same signals required for induction also responsible for regulating BBB maintenance in adulthood? Although Wnt signaling decreases in ECs after angiogenesis, this pathway is critical for BBB maintenance; disruption of Wnt signaling in adulthood leads to cell-autonomous loss of TJ integrity and an increase in PLVAP in the retina and cerebellum (Wang et al., 2012). Additionally, pericytes are important for BBB function throughout life (Armulik et al., 2010), suggesting that similar signals are required for BBB formation and maintenance. Do region-specific differences in signaling influence BBB heterogeneity? Different Wnt ligands and receptor complexes have been shown to promote BBB formation in different regions of the CNS (Daneman et al., 2009; Wang et al., 2012, 2018; Zhou et al., 2014); however, it is not clear whether this induces regional heterogeneity or is merely a remnant of dorsal–ventral and rostral–caudal axis specification.
How dynamic is each BBB property in a healthy CNS? Are properties modulated by neural activity or environmental stimuli such as exercise and diet? Single-cell sequencing has revealed vascular changes in response to neural activity (Hrvatin et al., 2018), and neuronal activity has been shown to modulate BBB insulin-like growth factor 1 (Nishijima et al., 2010). However, whether neural activity dynamically regulates specific properties of the BBB to modulate circuit function remains unknown. While exercise might help to protect against BBB dysfunction in aging or disease, solid evidence is still forthcoming (Małkiewicz et al., 2019). A high-fat diet can increase BBB permeability (de Aquino et al., 2019; Salameh et al., 2019; Yamamoto et al., 2019), but the specific BBB properties affected have not been thoroughly characterized. Not only can diet affect the BBB, but the BBB can in turn dynamically regulate nutrient availability; animals entering hibernation up-regulate ketone transporters at the BBB to modulate energy utilization during inactivity (Andrews et al., 2009). How dynamic are BBB properties over the course of 24 h, and how might these fluctuations influence brain microenvironment and waste clearance? PGP expression levels follow a diurnal pattern (Savolainen et al., 2016; Kervezee et al., 2014), and a circadian clock in glial cells of the Drosophila melanogaster BBB regulates xenobiotic efflux (Cuddapah et al., 2019; Zhang et al., 2018), but the extent and functional implications of circadian oscillations at the BBB remain unclear.
Are there differences in the BBB across individuals? Are there sex differences in BBB properties? There is evidence for variation in male and female patient CSF/serum albumin ratio (Parrado-Fernández et al., 2018), and BBB sexual dimorphism has been proposed to underlie differences in response to traumatic brain injury and infection and in proclivity to autoimmune disease (Cruz-Orengo et al., 2014; Jullienne et al., 2018; Maggioli et al., 2016).
How do BBB properties change in age? Several studies have reported age-related decline in BBB function (Mooradian, 1988; Montagne et al., 2015; Erdő et al., 2017), and age-related pericyte dysfunction contributes to BBB permeability (Bell et al., 2010). VCAM1 up-regulation at the BBB is a crucial step in age-related cognitive deficits and increased inflammatory tone (Yousef et al., 2019), highlighting VCAM1 as a potential therapeutic target for age-related neurodegeneration.
BBB dysfunction
BBB dysfunction occurs in a number of diseases, including MS, epilepsy, and stroke. In these conditions, BBB dysfunction is a central element of the pathology, whereas in others, such as Alzheimer’s disease (AD), the incidence and extent of breakdown are more controversial and an area of burgeoning research. BBB disruption causes ion dysregulation, edema, and neuroinflammation, which can lead to neuronal dysfunction, increased intracranial pressure, and neuronal degeneration. However, the mechanisms underlying BBB dysfunction and its role in the onset and progression of disease or recovery are not fully understood.
The phrase “BBB breakdown” conjures images of the destruction of a physical wall, allowing an unabated flow of molecules from the blood into the brain. However, the BBB is not a wall but a series of physiological properties, and a change in just one property (transcytosis, transport) can significantly alter the neural environment (Fig. 2). For instance, dysfunction of GLUT1 glucose transport, LAT1 amino acid transport, and MCT8 thyroid hormone transport across the BBB leads to seizure, autism spectrum, and psychomotor retardation syndromes, respectively (Seidner et al., 1998; Tărlungeanu et al., 2016; Vatine et al., 2017).
Importantly, leakage of nonspecific molecules is distinct from leukocyte extravasation, which occurs via an active trafficking process. Single-cell sequencing has identified many subsets of immune cells with distinct roles in neuroinflammation that likely interact with the BBB in disease (Mrdjen et al., 2018; Jordão et al., 2019; Kierdorf et al., 2019; Masuda et al., 2019; Mundt et al., 2019). Parenchymal ECs up-regulate leukocyte adhesion molecules, thus increasing leukocyte trafficking. P-selectin and E-selectin mediate the rolling of leukocytes along the endothelium, ICAM1 and VCAM1 mediate firm adhesion, and proteins like PLVAP—also up-regulated in disease—aid in transmigration across ECs (Engelhardt and Ransohoff, 2012; Ioannidou et al., 2006). Leukocyte extravasation across the BBB can be either transcellular or paracellular (Carman et al., 2007; Winger et al., 2014). Levels of ICAM1 and PECAM1 can influence T cell diapedesis route (Abadier et al., 2015; Wimmer et al., 2019), and specific subsets of T cells prefer different routes (Lutz et al., 2017).
Thus, the BBB is not an on–off switch, and it is critical to understand the specificities and consequences underlying each instance of dysfunction.
BBB dysfunction in CNS disorders
MS
BBB dysfunction is a central feature of MS, and the time course of leakage has been studied with dynamic contrast-enhanced MRI (Bastianello et al., 1990; Harris et al., 1991; Guttmann et al., 2016; Gaitán et al., 2011; Fig. 2 G). While barrier leakage is almost always present in new lesions, it is rarely observed in older lesions (Bastianello et al., 1990, Harris et al., 1991). Interestingly, MRI evidence suggests that BBB permeability is the initial event in the formation of a subset of lesions, but in others, lesion formation occurs before barrier dysfunction (Guttmann et al., 2016).
CNS immune infiltration is a critical step in MS pathophysiology, and the dynamics of this process have been primarily studied in experimental autoimmune encephalomyelitis (EAE), a rodent model of MS. The primary sites of CNS immune surveillance in health are the blood–CSF barriers of the choroid plexus and meninges, and both are important sites of initial lymphocyte activation in EAE (Bartholomäus et al., 2009; Schläger et al., 2016; Mundt et al., 2019; Engelhardt et al., 2001, 2017; Carrithers et al., 2000; Reboldi et al., 2009). These immune cells first enter the perivascular space surrounding post-capillary venules (Greter et al., 2005) and gain parenchymal access after breaking down the BM (Song et al., 2017; Wu et al., 2009). Leukocyte-derived cytokines activate CNS ECs, inducing expression of leukocyte adhesion molecules (Carrithers et al., 2000; Barkalow et al., 1996; Lou et al., 1996), which leads to massive parenchymal infiltration of immune cells. Limiting immune cell trafficking across the BBB has proven effective in treating MS. Natalizumab, which targets the α4 integrin on immune cells, preventing their interaction with endothelial VCAM1, greatly reduces new lesion formation (Miller et al., 2003).
It is critical to note that while leukocyte invasion is often assumed to be detrimental, leukocyte trafficking is required at low levels in order to limit infections. Of great interest is the identification of leukocyte adhesion molecules that facilitate the extravasation of only certain subsets of immune cells (Steinman, 2015). This could enable targeting pathological inflammation without rendering patients more vulnerable to infection. Indeed, ninjurin1 (NINJ1; monocytes), activated leukocyte cell adhesion molecule (ALCAM; CD4+ T cells, monocytes), junction adhesion molecule–like (JAML; monocytes, CD8+ T cells), and melanoma cell adhesion molecule (MCAM; CD8, T helper cell 17) regulate the entry of specific immune cell populations into the CNS (Alvarez et al., 2015; Cayrol et al., 2008; Flanagan et al., 2012; Ifergan et al., 2011; Larochelle et al., 2015). It will be necessary to ensure that targeting these molecules does not produce secondary effects; Alcam knockout mice develop more severe EAE as ALCAM also enforces TJ integrity (Lécuyer et al., 2017).
Many questions remain unanswered. How much of MS pathophysiology directly results from BBB dysfunction? Is there a subset of lesions caused by leakage while others have a different etiology? If these lesion subsets exist, do they vary with respect to severity and repair processes? Does the BBB interact with the lymphatic system to regulate leukocyte efflux during remission?
Ischemia/stroke
BBB dysfunction during stroke follows a biphasic time course. Leakage is evident within hours of the primary insult, is subsequently reduced, and then reappears the day after (Huang et al., 1999; Kuroiwa et al., 1985; Fig. 2, E, F, and H). An increase in transcytosis of nonspecific molecules is the first stage of dysfunction, followed by structural alteration of TJs (Knowland et al., 2014). Questions still remain regarding the importance of leukocyte infiltration in pathogenesis. Several reports have shown that leukocyte adhesion molecule knockouts or antibodies directed against leukocyte adhesion molecules minimize infarct volume (Bowes et al., 1993; Connolly et al., 1996; Mayadas et al., 1993), whereas others have not been able to replicate this effect (Enzmann et al., 2018).
Much of the cell death that leads to neurological deficits occurs in the days following a stroke; thus, the second phase of BBB leakage may be an important therapeutic target. Major outstanding questions in stroke research surround the relevance of this biphasic BBB dysfunction. It is unknown whether the first and second openings are mechanistically different; perhaps the first opening is due to dynamic signaling while the second results from changes in BBB gene expression.
Epilepsy
There is a clear association between epilepsy and BBB dysfunction. BBB leakage in epilepsy patients is visible with contrast-enhanced MRI (Horowitz et al., 1992; Alvarez et al., 2010; Rüber et al., 2018; Fig. 2 I), and analysis of brain tissue from epileptic patients shows increased parenchymal albumin (Cornford et al., 1998a; Mihály and Bozóky, 1984), implicating blood-to-brain extravasation of large molecules. Furthermore, patient samples exhibit regional reduction in GLUT1 (Cornford et al., 1998b), and positron emission tomography scans demonstrate decreased uptake and metabolism in seizure foci (Cornford et al., 1998a; Janigro, 1999).
BBB dysfunction itself may be epileptogenic or may help propagate seizures. Experimental disruption of the BBB with osmotic shock leads to seizures in patients (Marchi et al., 2007), and diseases in which the BBB is compromised such as infection, inflammation, stroke, and traumatic brain injury can lead to seizures and epilepsy (Oby and Janigro, 2006; van Vliet et al., 2007). Furthermore, neuroinflammation has been hypothesized to be involved in seizure etiology; blockage of leukocyte–vascular interactions either pharmacologically or by genetic knockout inhibits both induction and recurrence of seizures (Fabene et al., 2008). Interestingly, patients with a BBB-GLUT1 deficiency develop epilepsy (De Vivo et al., 1991; De Vivo et al., 2002), demonstrating a critical role for BBB transport in normal brain function.
AD
The extent of BBB dysfunction in AD and its role in etiology are an important ongoing focus of research. Several techniques have been used to examine BBB function in AD patients, including staining postmortem brain tissue for serum components, measurement of blood/CSF albumin concentrations, and various imaging modalities. Histological analyses have shown increased albumin and immunoglobulins in areas of heavy plaque burden (Wisniewski et al., 1997) as well as increased levels of fibrinogen (Ryu and McLarnon, 2009). A three-dimensional in vitro AD model has shown evidence of BBB dysfunction, phenocopying vascular changes reported in patients (Shin et al., 2019). Additionally, several imaging studies have found evidence of a leakier BBB in AD patients and propose BBB dysfunction as an early biomarker of AD (Starr et al., 2009; Montagne et al., 2015; van de Haar et al., 2016; Nation et al., 2019). While many older reports found no change in CSF albumin levels or contrast-enhanced imaging (Alafuzoff et al., 1987; Frölich et al., 1991; Kay et al., 1987; Mecocci et al., 1991; Bronge and Wahlund, 2000; Dysken et al., 1990; Schlageter et al., 1987), several of these studies did find evidence of BBB leakage in patients with vascular disease, suggesting that even in the absence of widespread leakage, there is a crucial vascular component to pathology (Erickson and Banks, 2013; Farrall and Wardlaw, 2009; Mecocci et al., 1991; Alafuzoff et al., 1983). As new imaging technology with greater resolution has gained wider use, BBB dysfunction has been further implicated in the pathogenesis of AD (Montagne et al., 2015; van de Haar et al., 2016; Nation et al., 2019). With these new tools, it will be vital to perform a more detailed analysis to determine at what stage and in which brain regions BBB dysfunction occurs, whether leakage is transient or chronic, and which cellular BBB properties are affected.
Regardless of the extent of widespread BBB leakage, there are several links between BBB dysfunction and AD pathology (Petersen et al., 2018). Fibrin accumulates in amyloid-positive vessels in AD patients and mouse models, and fibrin depletion protects against cognitive deficits in mice (Paul et al., 2007; Cortes-Canteli et al., 2010). Perhaps small amounts of BBB leakage related to injury, infection, or aging increase fibrin deposition, setting in motion an inflammatory cascade that plays an important role in AD pathology (Petersen et al., 2018; Kumar et al., 2016; Kinney et al., 2018).
In addition to nonspecific leakage, dysfunction of BBB Aβ transport may drive AD pathology (Bell and Zlokovic, 2009; Erickson and Banks, 2013). LRP1, a cell-surface receptor expressed on ECs, regulates Aβ clearance from the parenchyma (Shibata et al., 2000). EC-specific Lrp1 knockout increases levels of soluble brain Aβ and the severity of learning and memory deficits in an AD mouse model (Storck et al., 2016). A phosphatidylinositol binding clathrin assembly protein (PICALM)/PGP-dependent mechanism also aids in the clearance of Aβ across the BBB. PICALM regulates clathrin-dependent internalization of Aβ, guiding receptor-mediated transcytosis and clearance of Aβ, potentially presenting Aβ to efflux transporters (Zhao et al., 2015). PGP deficiency in an AD mouse model cuts Aβ clearance rate in half and increases CNS Aβ deposition (Cirrito et al., 2005), and Aβ40 triggers ubiquitination and internalization of PGP (Hartz et al., 2016), suggesting a dangerous feedback cycle. Conversely, receptor for advanced glycation endproducts (RAGE) imports Aβ into the CNS (Deane et al., 2003), and alterations in LRP:RAGE activity are hypothesized to drive CNS amyloid deposition in AD patients (Jeynes and Provias, 2008).
Another factor that might contribute to BBB dysfunction in AD is apolipoprotein E (APOE) genotype. Transgenic mice expressing human APOE4, the AD risk allele, exhibit cerebral vasculature with a thinner BM and BBB dysfunction due to cyclophillin/MMP9 signaling in pericytes (Bell et al., 2012; Alata et al., 2015). Further, postmortem AD tissue has revealed decreased TJ proteins and MMP9 elevation along with pericyte degeneration in APOE4 carriers (Bell et al., 2012; Halliday et al., 2016; Nishitsuji et al., 2011). However, there are conflicting reports; others show no changes in BBB function in Apoe4 knockout or APOE4 transgenic mice (Bien-Ly et al., 2015). One possible explanation is that APOE4 might cause minor, highly localized BBB leakage while not disrupting global BBB integrity (Ulrich et al., 2015).
To address the outstanding questions in the field, a deeper understanding of the association between vascular damage and AD pathology is necessary. This will require a focus on finding causal rather than correlational information linking BBB leakage, inflammation, and AD pathology. For instance, a recent study found that BBB dysfunction is an early marker of cognitive decline independent of Aβ or tau accumulation (Nation et al., 2019), but more details are needed regarding the extent of BBB dysfunction at various points during the AD time course. Furthermore, it is critical to understand how the BBB, glymphatics, and lymphatics cooperate to remove Aβ and other waste products from the CNS parenchyma, and what role this plays in AD pathophysiology (Stower, 2018; Rasmussen et al., 2018; Sweeney and Zlokovic, 2018; Da Mesquita et al., 2018).
Looking forward
Several important questions remain regarding the BBB in the context of disease. How is each BBB property altered in neurological diseases, and how do these changes affect the extracellular environment of the CNS? One problem is that different studies in humans or mouse models often use a single modality to detect BBB breakdown, whether sampling postmortem tissue, measuring markers in the CSF or blood, quantifying leakage of an exogenous tracer, or performing live imaging with a contrast agent. The BBB is not a single entity that is “open” or “shut,” and moving forward, it is imperative to understand exactly how the complex physiology of the BBB changes in each disease. It is especially important to consider whether alterations are induced by the same or different signals across neurological conditions. If mechanistic similarities exist, it might be possible to design a therapeutic strategy applicable to a wide range of disorders (Munji et al., 2019). Indeed, several molecular factors regulate BBB dysfunction in multiple diseases, including vascular endothelial growth factor (Argaw et al., 2009, 2012), inflammatory cytokines (tumor necrosis factor α [Nishioku et al., 2010], interleukins 1 and 6 [Chiaretti et al., 2005; Paré et al., 2018; Wang et al., 2014]), reactive oxygen species (Maier et al., 2006; Pun et al., 2009; Relton et al., 1997), and matrix metalloproteinases (Gidday et al., 2005; Ugarte-Berzal et al., 2018). However, there is also evidence that barrier dysfunction is due not only to “breakdown signals” but also to disrupted maintenance signals. Disruption of Wnt signaling can lead to vascular permeability and worse disease outcomes (Wang et al., 2012; Chang et al., 2017); thus, increasing CNS EC Wnt signaling might have therapeutic potential.
Can subtle changes in different BBB properties cause specific neurological symptoms? Dysfunction in several BBB transporters causes specific developmental disorders (Seidner et al., 1998; Tărlungeanu et al., 2016; Vatine et al., 2017), and there may be more undiscovered instances of this pattern. It is possible that regional heterogeneity at the BBB renders particular brain regions vulnerable to certain disease pathologies. For instance, if the BBB is indeed specialized to cater to the distinct nutrient and signaling needs of individual brain regions, loss of one of those BBB specializations might lead to deficits in local circuit function.
It is also important to also think beyond ECs. Disruption of pericyte coverage leads to an increase in EC nonspecific transcytosis and leukocyte adhesion molecules expression, and it is unclear to what extent this drives neurological disease. Furthermore, disruption of astrocyte endfeet at the NVU would decrease glymphatic clearance, potentially contributing to pathological accumulation of proteins including Aβ. Future work analyzing how each cell type of the NVU, and the glycocalyx and BMs, is altered will be critical to understand the pathophysiology of different neurological diseases.
Another fundamental question is how the BBB is repaired. While the BBB becomes less permeable to molecular tracers at chronic phases of disease models, it is unclear whether there are functional or structural compromises made in the process of reversing leakiness. More work is needed to fully characterize the repaired BBB at the levels of physical integrity and transcriptomics. It is also unknown what endogenous signals induce BBB repair, and whether repair occurs cell-autonomously within ECs or with mediation from other cell types. Interestingly, both microglia and reactive astrocytes regulate repair of the BBB in response to injury, highlighting the importance of the interactions of cells within the NVU (Lou et al., 2016, Bush et al., 1999, Fernández-López et al., 2016).
Concluding remarks
The BBB is not a single entity, but rather a complex series of physiological properties allowing CNS ECs to tightly regulate the extracellular environment of the parenchyma. These properties are vital for proper neural function, and dysfunction of the BBB can lead to critical pathology in many neurological diseases. However, more work is needed in order to understand exactly what crosses the healthy BBB, the degree to which the BBB dynamically responds to environmental stimuli, the extent of its regional heterogeneity, and the signaling mechanisms underlying its maintenance, disruption, and repair (Box 2). As future research answers these questions and further reveals the cellular and molecular intricacies underlying the BBB, the clinical advantages will be twofold: a deeper knowledge of the BBB will provide therapeutic targets for BBB repair in a range of neurological conditions and will also enable more effective strategies for delivering drugs to the CNS.
Box 2. Important unanswered questions

Acknowledgments
We would like to thank Dr. John Hesselink (University of California, San Diego, San Diego, CA) for generously providing MRI and FLAIR images for Fig. 2.
Author contributions: C.P. Profaci, R.N. Munji, R.S. Pulido, and R. Daneman contributed to the conceptualization, research, and writing of the manuscript. C.P. Profaci and R. Daneman edited the manuscript. C.P. Profaci and R.N. Munji created the figures and boxes.
References
- Abadier M., Haghayegh Jahromi N., Cardoso Alves L., Boscacci R., Vestweber D., Barnum S., Deutsch U., Engelhardt B., and Lyck R.. 2015. Cell surface levels of endothelial ICAM-1 influence the transcellular or paracellular T-cell diapedesis across the blood-brain barrier. Eur. J. Immunol. 45:1043–1058. 10.1002/eji.201445125 [DOI] [PubMed] [Google Scholar]
- Abbott N.J., Pizzo M.E., Preston J.E., Janigro D., and Thorne R.G.. 2018. The role of brain barriers in fluid movement in the CNS: is there a ‘glymphatic’ system? Acta Neuropathol. 135:387–407. 10.1007/s00401-018-1812-4 [DOI] [PubMed] [Google Scholar]
- Aird W.C. 2007. Phenotypic heterogeneity of the endothelium: II. Representative vascular beds. Circ. Res. 100:174–190. 10.1161/01.RES.0000255690.03436.ae [DOI] [PubMed] [Google Scholar]
- Alafuzoff I., Adolfsson R., Bucht G., and Winblad B.. 1983. Albumin and immunoglobulin in plasma and cerebrospinal fluid, and blood-cerebrospinal fluid barrier function in patients with dementia of Alzheimer type and multi-infarct dementia. J. Neurol. Sci. 60:465–472. 10.1016/0022-510X(83)90157-0 [DOI] [PubMed] [Google Scholar]
- Alafuzoff I., Adolfsson R., Grundke-Iqbal I., and Winblad B.. 1987. Blood-brain barrier in Alzheimer dementia and in non-demented elderly. An immunocytochemical study. Acta Neuropathol. 73:160–166. 10.1007/BF00693782 [DOI] [PubMed] [Google Scholar]
- Alata W., Ye Y., St-Amour I., Vandal M., and Calon F.. 2015. Human apolipoprotein E ɛ4 expression impairs cerebral vascularization and blood-brain barrier function in mice. J. Cereb. Blood Flow Metab. 35:86–94. 10.1038/jcbfm.2014.172 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alliot F., Godin I., and Pessac B.. 1999. Microglia derive from progenitors, originating from the yolk sac, and which proliferate in the brain. Brain Res. Dev. Brain Res. 117:145–152. 10.1016/S0165-3806(99)00113-3 [DOI] [PubMed] [Google Scholar]
- Alvarez V., Maeder P., and Rossetti A.O.. 2010. Postictal blood-brain barrier breakdown on contrast-enhanced MRI. Epilepsy Behav. 17:302–303. 10.1016/j.yebeh.2009.12.025 [DOI] [PubMed] [Google Scholar]
- Alvarez J.I., Cayrol R., and Prat A.. 2011. Disruption of central nervous system barriers in multiple sclerosis. Biochim. Biophys. Acta. 1812:252–264. 10.1016/j.bbadis.2010.06.017 [DOI] [PubMed] [Google Scholar]
- Alvarez J.I., Kébir H., Cheslow L., Charabati M., Chabarati M., Larochelle C., and Prat A.. 2015. JAML mediates monocyte and CD8 T cell migration across the brain endothelium. Ann. Clin. Transl. Neurol. 2:1032–1037. 10.1002/acn3.255 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amasheh S., Schmidt T., Mahn M., Florian P., Mankertz J., Tavalali S., Gitter A.H., Schulzke J.D., and Fromm M.. 2005. Contribution of claudin-5 to barrier properties in tight junctions of epithelial cells. Cell Tissue Res. 321:89–96. 10.1007/s00441-005-1101-0 [DOI] [PubMed] [Google Scholar]
- Ando Y., Okada H., Takemura G., Suzuki K., Takada C., Tomita H., Zaikokuji R., Hotta Y., Miyazaki N., Yano H., et al. 2018. Brain-Specific Ultrastructure of Capillary Endothelial Glycocalyx and Its Possible Contribution for Blood Brain Barrier. Sci. Rep. 8:17523 10.1038/s41598-018-35976-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreone B.J., Chow B.W., Tata A., Lacoste B., Ben-Zvi A., Bullock K., Deik A.A., Ginty D.D., Clish C.B., and Gu C.. 2017. Blood-Brain Barrier Permeability Is Regulated by Lipid Transport-Dependent Suppression of Caveolae-Mediated Transcytosis. Neuron. 94:581–594.e5. 10.1016/j.neuron.2017.03.043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrews M.T., Russeth K.P., Drewes L.R., and Henry P.G.. 2009. Adaptive mechanisms regulate preferred utilization of ketones in the heart and brain of a hibernating mammal during arousal from torpor. Am. J. Physiol. Regul. Integr. Comp. Physiol. 296:R383–R393. 10.1152/ajpregu.90795.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Argaw A.T., Gurfein B.T., Zhang Y., Zameer A., and John G.R.. 2009. VEGF-mediated disruption of endothelial CLN-5 promotes blood-brain barrier breakdown. Proc. Natl. Acad. Sci. USA. 106:1977–1982. 10.1073/pnas.0808698106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Argaw A.T., Asp L., Zhang J., Navrazhina K., Pham T., Mariani J.N., Mahase S., Dutta D.J., Seto J., Kramer E.G., et al. 2012. Astrocyte-derived VEGF-A drives blood-brain barrier disruption in CNS inflammatory disease. J. Clin. Invest. 122:2454–2468. 10.1172/JCI60842 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armulik A., Abramsson A., and Betsholtz C.. 2005. Endothelial/pericyte interactions. Circ. Res. 97:512–523. 10.1161/01.RES.0000182903.16652.d7 [DOI] [PubMed] [Google Scholar]
- Armulik A., Genové G., Mäe M., Nisancioglu M.H., Wallgard E., Niaudet C., He L., Norlin J., Lindblom P., Strittmatter K., et al. 2010. Pericytes regulate the blood-brain barrier. Nature. 468:557–561. 10.1038/nature09522 [DOI] [PubMed] [Google Scholar]
- Armulik A., Genové G., and Betsholtz C.. 2011. Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev. Cell. 21:193–215. 10.1016/j.devcel.2011.07.001 [DOI] [PubMed] [Google Scholar]
- Ausprunk D.H., Boudreau C.L., and Nelson D.A.. 1981a Proteoglycans in the microvascular. II. Histochemical localization in proliferating capillaries of the rabbit cornea. Am. J. Pathol. 103:367–375. [PMC free article] [PubMed] [Google Scholar]
- Ausprunk D.H., Boudreau C.L., and Nelson D.A.. 1981b Proteoglycans in the microvasculature. I. Histochemical localization in microvessels of the rabbit eye. Am. J. Pathol. 103:353–366. [PMC free article] [PubMed] [Google Scholar]
- Barkalow F.J., Goodman M.J., Gerritsen M.E., and Mayadas T.N.. 1996. Brain endothelium lack one of two pathways of P-selectin-mediated neutrophil adhesion. Blood. 88:4585–4593. 10.1182/blood.V88.12.4585.bloodjournal88124585 [DOI] [PubMed] [Google Scholar]
- Bartholomäus I., Kawakami N., Odoardi F., Schläger C., Miljkovic D., Ellwart J.W., Klinkert W.E., Flügel-Koch C., Issekutz T.B., Wekerle H., and Flügel A.. 2009. Effector T cell interactions with meningeal vascular structures in nascent autoimmune CNS lesions. Nature. 462:94–98. 10.1038/nature08478 [DOI] [PubMed] [Google Scholar]
- Bastianello S., Pozzilli C., Bernardi S., Bozzao L., Fantozzi L.M., Buttinelli C., and Fieschi C.. 1990. Serial study of gadolinium-DTPA MRI enhancement in multiple sclerosis. Neurology. 40:591–595. 10.1212/WNL.40.4.591 [DOI] [PubMed] [Google Scholar]
- Bell R.D., and Zlokovic B.V.. 2009. Neurovascular mechanisms and blood-brain barrier disorder in Alzheimer’s disease. Acta Neuropathol. 118:103–113. 10.1007/s00401-009-0522-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell R.D., Winkler E.A., Sagare A.P., Singh I., LaRue B., Deane R., and Zlokovic B.V.. 2010. Pericytes control key neurovascular functions and neuronal phenotype in the adult brain and during brain aging. Neuron. 68:409–427. 10.1016/j.neuron.2010.09.043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell R.D., Winkler E.A., Singh I., Sagare A.P., Deane R., Wu Z., Holtzman D.M., Betsholtz C., Armulik A., Sallstrom J., et al. 2012. Apolipoprotein E controls cerebrovascular integrity via cyclophilin A. Nature. 485:512–516. 10.1038/nature11087 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben-Zvi A., Lacoste B., Kur E., Andreone B.J., Mayshar Y., Yan H., and Gu C.. 2014. Mfsd2a is critical for the formation and function of the blood-brain barrier. Nature. 509:507–511. 10.1038/nature13324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benz F., Wichitnaowarat V., Lehmann M., Germano R.F., Mihova D., Macas J., Adams R.H., Taketo M.M., Plate K.H., Guérit S., et al. 2019. Low wnt/β-catenin signaling determines leaky vessels in the subfornical organ and affects water homeostasis in mice. eLife. 8:e43818 10.7554/eLife.43818 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bevilacqua M.P. 1993. Endothelial-leukocyte adhesion molecules. Annu. Rev. Immunol. 11:767–804. 10.1146/annurev.iy.11.040193.004003 [DOI] [PubMed] [Google Scholar]
- Bien-Ly N., Boswell C.A., Jeet S., Beach T.G., Hoyte K., Luk W., Shihadeh V., Ulufatu S., Foreman O., Lu Y., et al. 2015. Lack of Widespread BBB Disruption in Alzheimer’s Disease Models: Focus on Therapeutic Antibodies. Neuron. 88:289–297. 10.1016/j.neuron.2015.09.036 [DOI] [PubMed] [Google Scholar]
- Biswas P., Canosa S., Schoenfeld D., Schoenfeld J., Li P., Cheas L.C., Zhang J., Cordova A., Sumpio B., and Madri J.A.. 2006. PECAM-1 affects GSK-3beta-mediated beta-catenin phosphorylation and degradation. Am. J. Pathol. 169:314–324. 10.2353/ajpath.2006.051112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanchette M., and Daneman R.. 2015. Formation and maintenance of the BBB. Mech. Dev. 138:8–16. 10.1016/j.mod.2015.07.007 [DOI] [PubMed] [Google Scholar]
- Boado R.J., Li J.Y., Nagaya M., Zhang C., and Pardridge W.M.. 1999. Selective expression of the large neutral amino acid transporter at the blood-brain barrier. Proc. Natl. Acad. Sci. USA. 96:12079–12084. 10.1073/pnas.96.21.12079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowes M.P., Zivin J.A., and Rothlein R.. 1993. Monoclonal antibody to the ICAM-1 adhesion site reduces neurological damage in a rabbit cerebral embolism stroke model. Exp. Neurol. 119:215–219. 10.1006/exnr.1993.1023 [DOI] [PubMed] [Google Scholar]
- Bronge L., and Wahlund L.O.. 2000. White matter lesions in dementia: an MRI study on blood-brain barrier dysfunction. Dement. Geriatr. Cogn. Disord. 11:263–267. 10.1159/000017248 [DOI] [PubMed] [Google Scholar]
- Bush T.G., Puvanachandra N., Horner C.H., Polito A., Ostenfeld T., Svendsen C.N., Mucke L., Johnson M.H., and Sofroniew M.V.. 1999. Leukocyte infiltration, neuronal degeneration, and neurite outgrowth after ablation of scar-forming, reactive astrocytes in adult transgenic mice. Neuron. 23:297–308. 10.1016/S0896-6273(00)80781-3 [DOI] [PubMed] [Google Scholar]
- Carman C.V., Sage P.T., Sciuto T.E., de la Fuente M.A., Geha R.S., Ochs H.D., Dvorak H.F., Dvorak A.M., and Springer T.A.. 2007. Transcellular diapedesis is initiated by invasive podosomes. Immunity. 26:784–797. 10.1016/j.immuni.2007.04.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrithers M.D., Visintin I., Kang S.J., and Janeway C.A. Jr. 2000. Differential adhesion molecule requirements for immune surveillance and inflammatory recruitment. Brain. 123:1092–1101. 10.1093/brain/123.6.1092 [DOI] [PubMed] [Google Scholar]
- Cayrol R., Wosik K., Berard J.L., Dodelet-Devillers A., Ifergan I., Kebir H., Haqqani A.S., Kreymborg K., Krug S., Moumdjian R., et al. 2008. Activated leukocyte cell adhesion molecule promotes leukocyte trafficking into the central nervous system. Nat. Immunol. 9:137–145. 10.1038/ni1551 [DOI] [PubMed] [Google Scholar]
- Chang J., Mancuso M.R., Maier C., Liang X., Yuki K., Yang L., Kwong J.W., Wang J., Rao V., Vallon M., et al. 2017. Gpr124 is essential for blood-brain barrier integrity in central nervous system disease. Nat. Med. 23:450–460. 10.1038/nm.4309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiaretti A., Genovese O., Aloe L., Antonelli A., Piastra M., Polidori G., and Di Rocco C.. 2005. Interleukin 1beta and interleukin 6 relationship with paediatric head trauma severity and outcome. Childs Nerv. Syst. 21:185–193, discussion: 194. 10.1007/s00381-004-1032-1 [DOI] [PubMed] [Google Scholar]
- Cho C., Smallwood P.M., and Nathans J.. 2017. Reck and Gpr124 Are Essential Receptor Cofactors for Wnt7a/Wnt7b-Specific Signaling in Mammalian CNS Angiogenesis and Blood-Brain Barrier Regulation. Neuron. 95:1056–1073.e5. 10.1016/j.neuron.2017.07.031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chow B.W., and Gu C.. 2017. Gradual Suppression of Transcytosis Governs Functional Blood-Retinal Barrier Formation. Neuron. 93:1325–1333.e3. 10.1016/j.neuron.2017.02.043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cirrito J.R., Deane R., Fagan A.M., Spinner M.L., Parsadanian M., Finn M.B., Jiang H., Prior J.L., Sagare A., Bales K.R., et al. 2005. P-glycoprotein deficiency at the blood-brain barrier increases amyloid-beta deposition in an Alzheimer disease mouse model. J. Clin. Invest. 115:3285–3290. 10.1172/JCI25247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Connolly E.S. Jr., Winfree C.J., Springer T.A., Naka Y., Liao H., Yan S.D., Stern D.M., Solomon R.A., Gutierrez-Ramos J.C., and Pinsky D.J.. 1996. Cerebral protection in homozygous null ICAM-1 mice after middle cerebral artery occlusion. Role of neutrophil adhesion in the pathogenesis of stroke. J. Clin. Invest. 97:209–216. 10.1172/JCI118392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornford E.M., Hyman S., and Swartz B.E.. 1994. The human brain GLUT1 glucose transporter: ultrastructural localization to the blood-brain barrier endothelia. J. Cereb. Blood Flow Metab. 14:106–112. 10.1038/jcbfm.1994.15 [DOI] [PubMed] [Google Scholar]
- Cornford E.M., Gee M.N., Swartz B.E., Mandelkern M.A., Blahd W.H., Landaw E.M., and Delgado-Escueta A.V.. 1998a Dynamic [18F]fluorodeoxyglucose positron emission tomography and hypometabolic zones in seizures: reduced capillary influx. Ann. Neurol. 43:801–808. 10.1002/ana.410430615 [DOI] [PubMed] [Google Scholar]
- Cornford E.M., Hyman S., Cornford M.E., Landaw E.M., and Delgado-Escueta A.V.. 1998b Interictal seizure resections show two configurations of endothelial Glut1 glucose transporter in the human blood-brain barrier. J. Cereb. Blood Flow Metab. 18:26–42. 10.1097/00004647-199801000-00003 [DOI] [PubMed] [Google Scholar]
- Cortes-Canteli M., Paul J., Norris E.H., Bronstein R., Ahn H.J., Zamolodchikov D., Bhuvanendran S., Fenz K.M., and Strickland S.. 2010. Fibrinogen and beta-amyloid association alters thrombosis and fibrinolysis: a possible contributing factor to Alzheimer’s disease. Neuron. 66:695–709. 10.1016/j.neuron.2010.05.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cruz-Orengo L., Daniels B.P., Dorsey D., Basak S.A., Grajales-Reyes J.G., McCandless E.E., Piccio L., Schmidt R.E., Cross A.H., Crosby S.D., and Klein R.S.. 2014. Enhanced sphingosine-1-phosphate receptor 2 expression underlies female CNS autoimmunity susceptibility. J. Clin. Invest. 124:2571–2584. 10.1172/JCI73408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuddapah V.A., Zhang S.L., and Sehgal A.. 2019. Regulation of the Blood-Brain Barrier by Circadian Rhythms and Sleep. Trends Neurosci. 42:500–510. 10.1016/j.tins.2019.05.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Da Mesquita S., Louveau A., Vaccari A., Smirnov I., Cornelison R.C., Kingsmore K.M., Contarino C., Onengut-Gumuscu S., Farber E., Raper D., et al. 2018. Functional aspects of meningeal lymphatics in ageing and Alzheimer’s disease. Nature. 560:185–191. 10.1038/s41586-018-0368-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daneman R., Agalliu D., Zhou L., Kuhnert F., Kuo C.J., and Barres B.A.. 2009. Wnt/beta-catenin signaling is required for CNS, but not non-CNS, angiogenesis. Proc. Natl. Acad. Sci. USA. 106:641–646. 10.1073/pnas.0805165106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daneman R., Zhou L., Agalliu D., Cahoy J.D., Kaushal A., and Barres B.A.. 2010a The mouse blood-brain barrier transcriptome: a new resource for understanding the development and function of brain endothelial cells. PLoS One. 5:e13741 10.1371/journal.pone.0013741 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daneman R., Zhou L., Kebede A.A., and Barres B.A.. 2010b Pericytes are required for blood-brain barrier integrity during embryogenesis. Nature. 468:562–566. 10.1038/nature09513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Aquino C.C., Leitão R.A., Oliveira Alves L.A., Coelho-Santos V., Guerrant R.L., Ribeiro C.F., Malva J.O., Silva A.P., and Oriá R.B.. 2019. Effect of Hypoproteic and High-Fat Diets on Hippocampal Blood-Brain Barrier Permeability and Oxidative Stress. Front. Nutr. 5:131 10.3389/fnut.2018.00131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Vivo D.C., Trifiletti R.R., Jacobson R.I., Ronen G.M., Behmand R.A., and Harik S.I.. 1991. Defective glucose transport across the blood-brain barrier as a cause of persistent hypoglycorrhachia, seizures, and developmental delay. N. Engl. J. Med. 325:703–709. 10.1056/NEJM199109053251006 [DOI] [PubMed] [Google Scholar]
- De Vivo D.C., Leary L., and Wang D.. 2002. Glucose transporter 1 deficiency syndrome and other glycolytic defects. J. Child Neurol. 17(Suppl 3):S15–S23, discussion: S24–S25. [PubMed] [Google Scholar]
- Deane R., Du Yan S., Submamaryan R.K., LaRue B., Jovanovic S., Hogg E., Welch D., Manness L., Lin C., Yu J., et al. 2003. RAGE mediates amyloid-beta peptide transport across the blood-brain barrier and accumulation in brain. Nat. Med. 9:907–913. 10.1038/nm890 [DOI] [PubMed] [Google Scholar]
- Del Zoppo G.J., Milner R., Mabuchi T., Hung S., Wang X., and Koziol J.A.. 2006. Vascular matrix adhesion and the blood-brain barrier. Biochem. Soc. Trans. 34:1261–1266. 10.1042/BST0341261 [DOI] [PubMed] [Google Scholar]
- DellaValle B., Manresa-Arraut A., Hasseldam H., Stensballe A., Rungby J., Larsen A., and Hempel C.. 2018. Detection of Glycan Shedding in the Blood: New Class of Multiple Sclerosis Biomarkers? Front. Immunol. 9:1254 10.3389/fimmu.2018.01254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dysken M.W., Nelson M.J., Hoover K.M., Kuskowski M., and McGeachie R.. 1990. Rapid dynamic CT scanning in primary degenerative dementia and age-matched controls. Biol. Psychiatry. 28:425–434. 10.1016/0006-3223(90)90410-4 [DOI] [PubMed] [Google Scholar]
- Engelhardt B., and Ransohoff R.M.. 2012. Capture, crawl, cross: the T cell code to breach the blood-brain barriers. Trends Immunol. 33:579–589. 10.1016/j.it.2012.07.004 [DOI] [PubMed] [Google Scholar]
- Engelhardt B., Wolburg-Buchholz K., and Wolburg H.. 2001. Involvement of the choroid plexus in central nervous system inflammation. Microsc. Res. Tech. 52:112–129. 10.1002/1097-0029(20010101)52:1<112::AID-JEMT13>3.0.CO;2-5 [DOI] [PubMed] [Google Scholar]
- Engelhardt B., Vajkoczy P., and Weller R.O.. 2017. The movers and shapers in immune privilege of the CNS. Nat. Immunol. 18:123–131. 10.1038/ni.3666 [DOI] [PubMed] [Google Scholar]
- Enzmann G.U., Pavlidou S., Vaas M., Klohs J., and Engelhardt B.. 2018. ICAM-1null C57BL/6 Mice Are Not Protected from Experimental Ischemic Stroke. Transl. Stroke Res. 9:608–621. 10.1007/s12975-018-0612-4 [DOI] [PubMed] [Google Scholar]
- Erdő F., Denes L., and de Lange E.. 2017. Age-associated physiological and pathological changes at the blood-brain barrier: A review. J. Cereb. Blood Flow Metab. 37:4–24. 10.1177/0271678X16679420 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erickson M.A., and Banks W.A.. 2013. Blood-brain barrier dysfunction as a cause and consequence of Alzheimer’s disease. J. Cereb. Blood Flow Metab. 33:1500–1513. 10.1038/jcbfm.2013.135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fabene P.F., Navarro Mora G., Martinello M., Rossi B., Merigo F., Ottoboni L., Bach S., Angiari S., Benati D., Chakir A., et al. 2008. A role for leukocyte-endothelial adhesion mechanisms in epilepsy. Nat. Med. 14:1377–1383. 10.1038/nm.1878 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farrall A.J., and Wardlaw J.M.. 2009. Blood-brain barrier: ageing and microvascular disease--systematic review and meta-analysis. Neurobiol. Aging. 30:337–352. 10.1016/j.neurobiolaging.2007.07.015 [DOI] [PubMed] [Google Scholar]
- Fernández-López D., Faustino J., Klibanov A.L., Derugin N., Blanchard E., Simon F., Leib S.L., and Vexler Z.S.. 2016. Microglial Cells Prevent Hemorrhage in Neonatal Focal Arterial Stroke. J. Neurosci. 36:2881–2893. 10.1523/JNEUROSCI.0140-15.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flanagan K., Fitzgerald K., Baker J., Regnstrom K., Gardai S., Bard F., Mocci S., Seto P., You M., Larochelle C., et al. 2012. Laminin-411 is a vascular ligand for MCAM and facilitates TH17 cell entry into the CNS. PLoS One. 7:e40443 10.1371/journal.pone.0040443 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frölich L., Kornhuber J., Ihl R., Fritze J., Maurer K., and Riederer P.. 1991. Integrity of the blood-CSF barrier in dementia of Alzheimer type: CSF/serum ratios of albumin and IgG. Eur. Arch. Psychiatry Clin. Neurosci. 240:363–366. 10.1007/BF02279767 [DOI] [PubMed] [Google Scholar]
- Fultz N.E., Bonmassar G., Setsompop K., Stickgold R.A., Rosen B.R., Polimeni J.R., and Lewis L.D.. 2019. Coupled electrophysiological, hemodynamic, and cerebrospinal fluid oscillations in human sleep. Science. 366:628–631. 10.1126/science.aax5440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furuse M. 2010. Molecular basis of the core structure of tight junctions. Cold Spring Harb. Perspect. Biol. 2:a002907 10.1101/cshperspect.a002907 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furuse M., Sasaki H., and Tsukita S.. 1999. Manner of interaction of heterogeneous claudin species within and between tight junction strands. J. Cell Biol. 147:891–903. 10.1083/jcb.147.4.891 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaitán M.I., Shea C.D., Evangelou I.E., Stone R.D., Fenton K.M., Bielekova B., Massacesi L., and Reich D.S.. 2011. Evolution of the blood-brain barrier in newly forming multiple sclerosis lesions. Ann. Neurol. 70:22–29. 10.1002/ana.22472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ganong W.F. 2000. Circumventricular organs: definition and role in the regulation of endocrine and autonomic function. Clin. Exp. Pharmacol. Physiol. 27:422–427. 10.1046/j.1440-1681.2000.03259.x [DOI] [PubMed] [Google Scholar]
- Gidday J.M., Gasche Y.G., Copin J.C., Shah A.R., Perez R.S., Shapiro S.D., Chan P.H., and Park T.S.. 2005. Leukocyte-derived matrix metalloproteinase-9 mediates blood-brain barrier breakdown and is proinflammatory after transient focal cerebral ischemia. Am. J. Physiol. Heart Circ. Physiol. 289:H558–H568. 10.1152/ajpheart.01275.2004 [DOI] [PubMed] [Google Scholar]
- Greter M., Heppner F.L., Lemos M.P., Odermatt B.M., Goebels N., Laufer T., Noelle R.J., and Becher B.. 2005. Dendritic cells permit immune invasion of the CNS in an animal model of multiple sclerosis. Nat. Med. 11:328–334. 10.1038/nm1197 [DOI] [PubMed] [Google Scholar]
- Gross P.M. 1992. Circumventricular organ capillaries. Prog. Brain Res. 91:219–233. 10.1016/S0079-6123(08)62338-9 [DOI] [PubMed] [Google Scholar]
- Guttmann C.R., Rousset M., Roch J.A., Hannoun S., Durand-Dubief F., Belaroussi B., Cavallari M., Rabilloud M., Sappey-Marinier D., Vukusic S., and Cotton F.. 2016. Multiple sclerosis lesion formation and early evolution revisited: A weekly high-resolution magnetic resonance imaging study. Mult. Scler. 22:761–769. 10.1177/1352458515600247 [DOI] [PubMed] [Google Scholar]
- Haj-Yasein N.N., Vindedal G.F., Eilert-Olsen M., Gundersen G.A., Skare Ø., Laake P., Klungland A., Thorén A.E., Burkhardt J.M., Ottersen O.P., and Nagelhus E.A.. 2011. Glial-conditional deletion of aquaporin-4 (Aqp4) reduces blood-brain water uptake and confers barrier function on perivascular astrocyte endfeet. Proc. Natl. Acad. Sci. USA. 108:17815–17820. 10.1073/pnas.1110655108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halliday M.R., Rege S.V., Ma Q., Zhao Z., Miller C.A., Winkler E.A., and Zlokovic B.V.. 2016. Accelerated pericyte degeneration and blood-brain barrier breakdown in apolipoprotein E4 carriers with Alzheimer’s disease. J. Cereb. Blood Flow Metab. 36:216–227. 10.1038/jcbfm.2015.44 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hallmann R., Mayer D.N., Berg E.L., Broermann R., and Butcher E.C.. 1995. Novel mouse endothelial cell surface marker is suppressed during differentiation of the blood brain barrier. Dev. Dyn. 202:325–332. 10.1002/aja.1002020402 [DOI] [PubMed] [Google Scholar]
- Harris J.O., Frank J.A., Patronas N., McFarlin D.E., and McFarland H.F.. 1991. Serial gadolinium-enhanced magnetic resonance imaging scans in patients with early, relapsing-remitting multiple sclerosis: implications for clinical trials and natural history. Ann. Neurol. 29:548–555. 10.1002/ana.410290515 [DOI] [PubMed] [Google Scholar]
- Hartz A.M., Zhong Y., Wolf A., LeVine H. III, Miller D.S., and Bauer B.. 2016. Aβ40 Reduces P-Glycoprotein at the Blood-Brain Barrier through the Ubiquitin-Proteasome Pathway. J. Neurosci. 36:1930–1941. 10.1523/JNEUROSCI.0350-15.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickey W.F., and Kimura H.. 1988. Perivascular microglial cells of the CNS are bone marrow-derived and present antigen in vivo. Science. 239:290–292. 10.1126/science.3276004 [DOI] [PubMed] [Google Scholar]
- Hindle S.J., Munji R.N., Dolghih E., Gaskins G., Orng S., Ishimoto H., Soung A., DeSalvo M., Kitamoto T., Keiser M.J., et al. 2017. Evolutionarily Conserved Roles for Blood-Brain Barrier Xenobiotic Transporters in Endogenous Steroid Partitioning and Behavior. Cell Rep. 21:1304–1316. 10.1016/j.celrep.2017.10.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hladky S.B., and Barrand M.A.. 2019. Is solute movement within the extracellular spaces of brain gray matter brought about primarily by diffusion or flow? A commentary on “Analysis of convective and diffusive transport in the brain interstitium” Fluids and Barriers of the CNS (2019) 16:6 by L. Ray, J.J. Iliff and J.J. Heys. Fluids Barriers CNS. 16:24 10.1186/s12987-019-0141-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hnasko R., McFarland M., and Ben-Jonathan N.. 2002. Distribution and characterization of plasmalemma vesicle protein-1 in rat endocrine glands. J. Endocrinol. 175:649–661. 10.1677/joe.0.1750649 [DOI] [PubMed] [Google Scholar]
- Horowitz S.W., Merchut M., Fine M., and Azar-Kia B.. 1992. Complex partial seizure-induced transient MR enhancement. J. Comput. Assist. Tomogr. 16:814–816. 10.1097/00004728-199209000-00025 [DOI] [PubMed] [Google Scholar]
- Hou J., Gomes A.S., Paul D.L., and Goodenough D.A.. 2006. Study of claudin function by RNA interference. J. Biol. Chem. 281:36117–36123. 10.1074/jbc.M608853200 [DOI] [PubMed] [Google Scholar]
- Hrvatin S., Hochbaum D.R., Nagy M.A., Cicconet M., Robertson K., Cheadle L., Zilionis R., Ratner A., Borges-Monroy R., Klein A.M., et al. 2018. Single-cell analysis of experience-dependent transcriptomic states in the mouse visual cortex. Nat. Neurosci. 21:120–129. 10.1038/s41593-017-0029-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang Z.G., Xue D., Preston E., Karbalai H., and Buchan A.M.. 1999. Biphasic opening of the blood-brain barrier following transient focal ischemia: effects of hypothermia. Can. J. Neurol. Sci. 26:298–304. 10.1017/S0317167100000421 [DOI] [PubMed] [Google Scholar]
- Hudson L.C., Bragg D.C., Tompkins M.B., and Meeker R.B.. 2005. Astrocytes and microglia differentially regulate trafficking of lymphocyte subsets across brain endothelial cells. Brain Res. 1058:148–160. 10.1016/j.brainres.2005.07.071 [DOI] [PubMed] [Google Scholar]
- Ifergan I., Kebir H., Terouz S., Alvarez J.I., Lécuyer M.A., Gendron S., Bourbonnière L., Dunay I.R., Bouthillier A., Moumdjian R., et al. 2011. Role of Ninjurin-1 in the migration of myeloid cells to central nervous system inflammatory lesions. Ann. Neurol. 70:751–763. 10.1002/ana.22519 [DOI] [PubMed] [Google Scholar]
- Iliff J.J., Wang M., Liao Y., Plogg B.A., Peng W., Gundersen G.A., Benveniste H., Vates G.E., Deane R., Goldman S.A., et al. 2012. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 4:147ra111 10.1126/scitranslmed.3003748 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ioannidou S., Deinhardt K., Miotla J., Bradley J., Cheung E., Samuelsson S., Ng Y.S., and Shima D.T.. 2006. An in vitro assay reveals a role for the diaphragm protein PV-1 in endothelial fenestra morphogenesis. Proc. Natl. Acad. Sci. USA. 103:16770–16775. 10.1073/pnas.0603501103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janigro D. 1999. Blood-brain barrier, ion homeostatis and epilepsy: possible implications towards the understanding of ketogenic diet mechanisms. Epilepsy Res. 37:223–232. 10.1016/S0920-1211(99)00074-1 [DOI] [PubMed] [Google Scholar]
- Janzer R.C., and Raff M.C.. 1987. Astrocytes induce blood-brain barrier properties in endothelial cells. Nature. 325:253–257. 10.1038/325253a0 [DOI] [PubMed] [Google Scholar]
- Jefferies W.A., Brandon M.R., Hunt S.V., Williams A.F., Gatter K.C., and Mason D.Y.. 1984. Transferrin receptor on endothelium of brain capillaries. Nature. 312:162–163. 10.1038/312162a0 [DOI] [PubMed] [Google Scholar]
- Jeynes B., and Provias J.. 2008. Evidence for altered LRP/RAGE expression in Alzheimer lesion pathogenesis. Curr. Alzheimer Res. 5:432–437. 10.2174/156720508785908937 [DOI] [PubMed] [Google Scholar]
- Jordão M.J.C., Sankowski R., Brendecke S.M., Sagar G., Locatelli G., Tai Y.H., Tay T.L., Schramm E., Armbruster S., Hagemeyer N., et al. 2019. Single-cell profiling identifies myeloid cell subsets with distinct fates during neuroinflammation. Science. 363:eaat7554 10.1126/science.aat7554 [DOI] [PubMed] [Google Scholar]
- Jullienne A., Salehi A., Affeldt B., Baghchechi M., Haddad E., Avitua A., Walsworth M., Enjalric I., Hamer M., Bhakta S., et al. 2018. Male and Female Mice Exhibit Divergent Responses of the Cortical Vasculature to Traumatic Brain Injury. J. Neurotrauma. 35:1646–1658. 10.1089/neu.2017.5547 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kay A.D., May C., Papadopoulos N.M., Costello R., Atack J.R., Luxenberg J.S., Cutler N.R., and Rapoport S.I.. 1987. CSF and serum concentrations of albumin and IgG in Alzheimer’s disease. Neurobiol. Aging. 8:21–25. 10.1016/0197-4580(87)90053-4 [DOI] [PubMed] [Google Scholar]
- Kervezee L., Hartman R., van den Berg D.J., Shimizu S., Emoto-Yamamoto Y., Meijer J.H., and de Lange E.C.. 2014. Diurnal variation in P-glycoprotein-mediated transport and cerebrospinal fluid turnover in the brain. AAPS J. 16:1029–1037. 10.1208/s12248-014-9625-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kido Y., Tamai I., Okamoto M., Suzuki F., and Tsuji A.. 2000. Functional clarification of MCT1-mediated transport of monocarboxylic acids at the blood-brain barrier using in vitro cultured cells and in vivo BUI studies. Pharm. Res. 17:55–62. 10.1023/A:1007518525161 [DOI] [PubMed] [Google Scholar]
- Kierdorf K., Masuda T., Jordão M.J.C., and Prinz M.. 2019. Macrophages at CNS interfaces: ontogeny and function in health and disease. Nat. Rev. Neurosci. 20:547–562. 10.1038/s41583-019-0201-x [DOI] [PubMed] [Google Scholar]
- Kinney J.W., Bemiller S.M., Murtishaw A.S., Leisgang A.M., Salazar A.M., and Lamb B.T.. 2018. Inflammation as a central mechanism in Alzheimer’s disease. Alzheimers Dement. (N. Y.). 4:575–590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kipnis J., Gadani S., and Derecki N.C.. 2012. Pro-cognitive properties of T cells. Nat. Rev. Immunol. 12:663–669. 10.1038/nri3280 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knowland D., Arac A., Sekiguchi K.J., Hsu M., Lutz S.E., Perrino J., Steinberg G.K., Barres B.A., Nimmerjahn A., and Agalliu D.. 2014. Stepwise recruitment of transcellular and paracellular pathways underlies blood-brain barrier breakdown in stroke. Neuron. 82:603–617. 10.1016/j.neuron.2014.03.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubotera H., Ikeshima-Kataoka H., Hatashita Y., Allegra Mascaro A.L., Pavone F.S., and Inoue T.. 2019. Astrocytic endfeet re-cover blood vessels after removal by laser ablation. Sci. Rep. 9:1263 10.1038/s41598-018-37419-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar D.K., Choi S.H., Washicosky K.J., Eimer W.A., Tucker S., Ghofrani J., Lefkowitz A., McColl G., Goldstein L.E., Tanzi R.E., and Moir R.D.. 2016. Amyloid-β peptide protects against microbial infection in mouse and worm models of Alzheimer’s disease. Sci. Transl. Med. 8:340ra72 10.1126/scitranslmed.aaf1059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuroiwa T., Ting P., Martinez H., and Klatzo I.. 1985. The biphasic opening of the blood-brain barrier to proteins following temporary middle cerebral artery occlusion. Acta Neuropathol. 68:122–129. 10.1007/BF00688633 [DOI] [PubMed] [Google Scholar]
- Kutuzov N., Flyvbjerg H., and Lauritzen M.. 2018. Contributions of the glycocalyx, endothelium, and extravascular compartment to the blood-brain barrier. Proc. Natl. Acad. Sci. USA. 115:E9429–E9438. 10.1073/pnas.1802155115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langlet F., Mullier A., Bouret S.G., Prevot V., and Dehouck B.. 2013. Tanycyte-like cells form a blood-cerebrospinal fluid barrier in the circumventricular organs of the mouse brain. J. Comp. Neurol. 521:3389–3405. 10.1002/cne.23355 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larochelle C., Lécuyer M.A., Alvarez J.I., Charabati M., Saint-Laurent O., Ghannam S., Kebir H., Flanagan K., Yednock T., Duquette P., et al. 2015. Melanoma cell adhesion molecule-positive CD8 T lymphocytes mediate central nervous system inflammation. Ann. Neurol. 78:39–53. 10.1002/ana.24415 [DOI] [PubMed] [Google Scholar]
- Lécuyer M.A., Saint-Laurent O., Bourbonnière L., Larouche S., Larochelle C., Michel L., Charabati M., Abadier M., Zandee S., Haghayegh Jahromi N., et al. 2017. Dual role of ALCAM in neuroinflammation and blood-brain barrier homeostasis. Proc. Natl. Acad. Sci. USA. 114:E524–E533. 10.1073/pnas.1614336114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liebner S., Corada M., Bangsow T., Babbage J., Taddei A., Czupalla C.J., Reis M., Felici A., Wolburg H., Fruttiger M., et al. 2008. Wnt/beta-catenin signaling controls development of the blood-brain barrier. J. Cell Biol. 183:409–417. 10.1083/jcb.200806024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lou J., Dayer J.M., Grau G.E., and Burger D.. 1996. Direct cell/cell contact with stimulated T lymphocytes induces the expression of cell adhesion molecules and cytokines by human brain microvascular endothelial cells. Eur. J. Immunol. 26:3107–3113. 10.1002/eji.1830261242 [DOI] [PubMed] [Google Scholar]
- Lou N., Takano T., Pei Y., Xavier A.L., Goldman S.A., and Nedergaard M.. 2016. Purinergic receptor P2RY12-dependent microglial closure of the injured blood-brain barrier. Proc. Natl. Acad. Sci. USA. 113:1074–1079. 10.1073/pnas.1520398113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lutz S.E., Smith J.R., Kim D.H., Olson C.V.L., Ellefsen K., Bates J.M., Gandhi S.P., and Agalliu D.. 2017. Caveolin1 Is Required for Th1 Cell Infiltration, but Not Tight Junction Remodeling, at the Blood-Brain Barrier in Autoimmune Neuroinflammation. Cell Rep. 21:2104–2117. 10.1016/j.celrep.2017.10.094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macdonald J.A., Murugesan N., and Pachter J.S.. 2010. Endothelial cell heterogeneity of blood-brain barrier gene expression along the cerebral microvasculature. J. Neurosci. Res. 88:1457–1474. [DOI] [PubMed] [Google Scholar]
- Maggioli E., McArthur S., Mauro C., Kieswich J., Kusters D.H., Reutelingsperger C.P., Yaqoob M., and Solito E.. 2016. Estrogen protects the blood-brain barrier from inflammation-induced disruption and increased lymphocyte trafficking. Brain Behav. Immun. 51:212–222. 10.1016/j.bbi.2015.08.020 [DOI] [PubMed] [Google Scholar]
- Maier C.M., Hsieh L., Crandall T., Narasimhan P., and Chan P.H.. 2006. Evaluating therapeutic targets for reperfusion-related brain hemorrhage. Ann. Neurol. 59:929–938. 10.1002/ana.20850 [DOI] [PubMed] [Google Scholar]
- Małkiewicz M.A., Szarmach A., Sabisz A., Cubała W.J., Szurowska E., and Winklewski P.J.. 2019. Blood-brain barrier permeability and physical exercise. J. Neuroinflammation. 16:15 10.1186/s12974-019-1403-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchi N., Angelov L., Masaryk T., Fazio V., Granata T., Hernandez N., Hallene K., Diglaw T., Franic L., Najm I., and Janigro D.. 2007. Seizure-promoting effect of blood-brain barrier disruption. Epilepsia. 48:732–742. 10.1111/j.1528-1167.2007.00988.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marques F., Sousa J.C., Brito M.A., Pahnke J., Santos C., Correia-Neves M., and Palha J.A.. 2017. The choroid plexus in health and in disease: dialogues into and out of the brain. Neurobiol. Dis. 107:32–40. 10.1016/j.nbd.2016.08.011 [DOI] [PubMed] [Google Scholar]
- Masuda T., Sankowski R., Staszewski O., Böttcher C., Amann L., Sagar C., Scheiwe C., Nessler S., Kunz P., van Loo G., et al. 2019. Spatial and temporal heterogeneity of mouse and human microglia at single-cell resolution. Nature. 566:388–392. 10.1038/s41586-019-0924-x [DOI] [PubMed] [Google Scholar]
- Mayadas T.N., Johnson R.C., Rayburn H., Hynes R.O., and Wagner D.D.. 1993. Leukocyte rolling and extravasation are severely compromised in P selectin-deficient mice. Cell. 74:541–554. 10.1016/0092-8674(93)80055-J [DOI] [PubMed] [Google Scholar]
- Mecocci P., Parnetti L., Reboldi G.P., Santucci C., Gaiti A., Ferri C., Gernini I., Romagnoli M., Cadini D., and Senin U.. 1991. Blood-brain-barrier in a geriatric population: barrier function in degenerative and vascular dementias. Acta Neurol. Scand. 84:210–213. 10.1111/j.1600-0404.1991.tb04940.x [DOI] [PubMed] [Google Scholar]
- Meng Y., Abrahao A., Heyn C.C., Bethune A.J., Huang Y., Pople C.B., Aubert I., Hamani C., Zinman L., Hynynen K., et al. 2019. Glymphatics visualization after focused ultrasound induced blood-brain barrier opening in humans. Ann. Neurol. 86:975–980. 10.1002/ana.25604 [DOI] [PubMed] [Google Scholar]
- Méresse S., Delbart C., Fruchart J.C., and Cecchelli R.. 1989. Low-density lipoprotein receptor on endothelium of brain capillaries. J. Neurochem. 53:340–345. 10.1111/j.1471-4159.1989.tb07340.x [DOI] [PubMed] [Google Scholar]
- Mestre H., Hablitz L.M., Xavier A.L., Feng W., Zou W., Pu T., Monai H., Murlidharan G., Castellanos Rivera R.M., Simon M.J., et al. 2018. Aquaporin-4-dependent glymphatic solute transport in the rodent brain. eLife. 7:e40070 10.7554/eLife.40070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mihály A., and Bozóky B.. 1984. Immunohistochemical localization of extravasated serum albumin in the hippocampus of human subjects with partial and generalized epilepsies and epileptiform convulsions. Acta Neuropathol. 65:25–34. 10.1007/BF00689824 [DOI] [PubMed] [Google Scholar]
- Miller D.S. 2015. Regulation of ABC transporters at the blood-brain barrier. Clin. Pharmacol. Ther. 97:395–403. 10.1002/cpt.64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller D.H., Khan O.A., Sheremata W.A., Blumhardt L.D., Rice G.P., Libonati M.A., Willmer-Hulme A.J., Dalton C.M., Miszkiel K.A., and O’Connor P.W.. International Natalizumab Multiple Sclerosis Trial Group . 2003. A controlled trial of natalizumab for relapsing multiple sclerosis. N. Engl. J. Med. 348:15–23. 10.1056/NEJMoa020696 [DOI] [PubMed] [Google Scholar]
- Mishra A., Reynolds J.P., Chen Y., Gourine A.V., Rusakov D.A., and Attwell D.. 2016. Astrocytes mediate neurovascular signaling to capillary pericytes but not to arterioles. Nat. Neurosci. 19:1619–1627. 10.1038/nn.4428 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyata S. 2015. New aspects in fenestrated capillary and tissue dynamics in the sensory circumventricular organs of adult brains. Front. Neurosci. 9:390 10.3389/fnins.2015.00390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montagne A., Barnes S.R., Sweeney M.D., Halliday M.R., Sagare A.P., Zhao Z., Toga A.W., Jacobs R.E., Liu C.Y., Amezcua L., et al. 2015. Blood-brain barrier breakdown in the aging human hippocampus. Neuron. 85:296–302. 10.1016/j.neuron.2014.12.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mooradian A.D. 1988. Effect of aging on the blood-brain barrier. Neurobiol. Aging. 9:31–39. 10.1016/S0197-4580(88)80013-7 [DOI] [PubMed] [Google Scholar]
- Morita K., Sasaki H., Furuse M., and Tsukita S.. 1999. Endothelial claudin: claudin-5/TMVCF constitutes tight junction strands in endothelial cells. J. Cell Biol. 147:185–194. 10.1083/jcb.147.1.185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mrdjen D., Pavlovic A., Hartmann F.J., Schreiner B., Utz S.G., Leung B.P., Lelios I., Heppner F.L., Kipnis J., Merkler D., et al. 2018. High-Dimensional Single-Cell Mapping of Central Nervous System Immune Cells Reveals Distinct Myeloid Subsets in Health, Aging, and Disease. Immunity. 48:599 10.1016/j.immuni.2018.02.014 [DOI] [PubMed] [Google Scholar]
- Mundt S., Mrdjen D., Utz S.G., Greter M., Schreiner B., and Becher B.. 2019. Conventional DCs sample and present myelin antigens in the healthy CNS and allow parenchymal T cell entry to initiate neuroinflammation. Sci. Immunol. 4:eaau8380 10.1126/sciimmunol.aau8380 [DOI] [PubMed] [Google Scholar]
- Munji R.N., Soung A.L., Weiner G.A., Sohet F., Semple B.D., Trivedi A., Gimlin K., Kotoda M., Korai M., Aydin S., et al. 2019. Profiling the mouse brain endothelial transcriptome in health and disease models reveals a core blood-brain barrier dysfunction module. Nat. Neurosci. 22:1892–1902. 10.1038/s41593-019-0497-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murugesan N., Macdonald J.A., Lu Q., Wu S.L., Hancock W.S., and Pachter J.S.. 2011. Analysis of mouse brain microvascular endothelium using laser capture microdissection coupled with proteomics. Methods Mol. Biol. 686:297–311. 10.1007/978-1-60761-938-3_14 [DOI] [PubMed] [Google Scholar]
- Nałęcz K.A. 2017. Solute Carriers in the Blood-Brain Barier: Safety in Abundance. Neurochem. Res. 42:795–809. 10.1007/s11064-016-2030-x [DOI] [PubMed] [Google Scholar]
- Nation D.A., Sweeney M.D., Montagne A., Sagare A.P., D’Orazio L.M., Pachicano M., Sepehrband F., Nelson A.R., Buennagel D.P., Harrington M.G., et al. 2019. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat. Med. 25:270–276. 10.1038/s41591-018-0297-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen L.N., Ma D., Shui G., Wong P., Cazenave-Gassiot A., Zhang X., Wenk M.R., Goh E.L., and Silver D.L.. 2014. Mfsd2a is a transporter for the essential omega-3 fatty acid docosahexaenoic acid. Nature. 509:503–506. 10.1038/nature13241 [DOI] [PubMed] [Google Scholar]
- Nishijima T., Piriz J., Duflot S., Fernandez A.M., Gaitan G., Gomez-Pinedo U., Verdugo J.M., Leroy F., Soya H., Nuñez A., and Torres-Aleman I.. 2010. Neuronal activity drives localized blood-brain-barrier transport of serum insulin-like growth factor-I into the CNS. Neuron. 67:834–846. 10.1016/j.neuron.2010.08.007 [DOI] [PubMed] [Google Scholar]
- Nishioku T., Matsumoto J., Dohgu S., Sumi N., Miyao K., Takata F., Shuto H., Yamauchi A., and Kataoka Y.. 2010. Tumor necrosis factor-alpha mediates the blood-brain barrier dysfunction induced by activated microglia in mouse brain microvascular endothelial cells. J. Pharmacol. Sci. 112:251–254. 10.1254/jphs.09292SC [DOI] [PubMed] [Google Scholar]
- Nishitsuji K., Hosono T., Nakamura T., Bu G., and Michikawa M.. 2011. Apolipoprotein E regulates the integrity of tight junctions in an isoform-dependent manner in an in vitro blood-brain barrier model. J. Biol. Chem. 286:17536–17542. 10.1074/jbc.M111.225532 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nitta T., Hata M., Gotoh S., Seo Y., Sasaki H., Hashimoto N., Furuse M., and Tsukita S.. 2003. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 161:653–660. 10.1083/jcb.200302070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obermeier B., Daneman R., and Ransohoff R.M.. 2013. Development, maintenance and disruption of the blood-brain barrier. Nat. Med. 19:1584–1596. 10.1038/nm.3407 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oby E., and Janigro D.. 2006. The blood-brain barrier and epilepsy. Epilepsia. 47:1761–1774. 10.1111/j.1528-1167.2006.00817.x [DOI] [PubMed] [Google Scholar]
- Paré A., Mailhot B., Lévesque S.A., Juzwik C., Ignatius Arokia Doss P.M., Lécuyer M.A., Prat A., Rangachari M., Fournier A., and Lacroix S.. 2018. IL-1β enables CNS access to CCR2hi monocytes and the generation of pathogenic cells through GM-CSF released by CNS endothelial cells. Proc. Natl. Acad. Sci. USA. 115:E1194–E1203. 10.1073/pnas.1714948115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parrado-Fernández C., Blennow K., Hansson M., Leoni V., Cedazo-Minguez A., and Björkhem I.. 2018. Evidence for sex difference in the CSF/plasma albumin ratio in ~20 000 patients and 335 healthy volunteers. J. Cell. Mol. Med. 22:5151–5154. 10.1111/jcmm.13767 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paul J., Strickland S., and Melchor J.P.. 2007. Fibrin deposition accelerates neurovascular damage and neuroinflammation in mouse models of Alzheimer’s disease. J. Exp. Med. 204:1999–2008. 10.1084/jem.20070304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Persidsky Y., Ghorpade A., Rasmussen J., Limoges J., Liu X.J., Stins M., Fiala M., Way D., Kim K.S., Witte M.H., et al. 1999. Microglial and astrocyte chemokines regulate monocyte migration through the blood-brain barrier in human immunodeficiency virus-1 encephalitis. Am. J. Pathol. 155:1599–1611. 10.1016/S0002-9440(10)65476-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petersen M.A., Ryu J.K., and Akassoglou K.. 2018. Fibrinogen in neurological diseases: mechanisms, imaging and therapeutics. Nat. Rev. Neurosci. 19:283–301. 10.1038/nrn.2018.13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pillinger N.L., and Kam P.. 2017. Endothelial glycocalyx: basic science and clinical implications. Anaesth. Intensive Care. 45:295–307. 10.1177/0310057X1704500305 [DOI] [PubMed] [Google Scholar]
- Poduslo J.F., Curran G.L., and Berg C.T.. 1994. Macromolecular permeability across the blood-nerve and blood-brain barriers. Proc. Natl. Acad. Sci. USA. 91:5705–5709. 10.1073/pnas.91.12.5705 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prinz M., Priller J., Sisodia S.S., and Ransohoff R.M.. 2011. Heterogeneity of CNS myeloid cells and their roles in neurodegeneration. Nat. Neurosci. 14:1227–1235. 10.1038/nn.2923 [DOI] [PubMed] [Google Scholar]
- Prinz M., Erny D., and Hagemeyer N.. 2017. Ontogeny and homeostasis of CNS myeloid cells. Nat. Immunol. 18:385–392. 10.1038/ni.3703 [DOI] [PubMed] [Google Scholar]
- Privratsky J.R., and Newman P.J.. 2014. PECAM-1: regulator of endothelial junctional integrity. Cell Tissue Res. 355:607–619. 10.1007/s00441-013-1779-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pun P.B., Lu J., and Moochhala S.. 2009. Involvement of ROS in BBB dysfunction. Free Radic. Res. 43:348–364. 10.1080/10715760902751902 [DOI] [PubMed] [Google Scholar]
- Ransohoff R.M., and Engelhardt B.. 2012. The anatomical and cellular basis of immune surveillance in the central nervous system. Nat. Rev. Immunol. 12:623–635. 10.1038/nri3265 [DOI] [PubMed] [Google Scholar]
- Rasmussen M.K., Mestre H., and Nedergaard M.. 2018. The glymphatic pathway in neurological disorders. Lancet Neurol. 17:1016–1024. 10.1016/S1474-4422(18)30318-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reboldi A., Coisne C., Baumjohann D., Benvenuto F., Bottinelli D., Lira S., Uccelli A., Lanzavecchia A., Engelhardt B., and Sallusto F.. 2009. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nat. Immunol. 10:514–523. 10.1038/ni.1716 [DOI] [PubMed] [Google Scholar]
- Reis M., Czupalla C.J., Ziegler N., Devraj K., Zinke J., Seidel S., Heck R., Thom S., Macas J., Bockamp E., et al. 2012. Endothelial Wnt/β-catenin signaling inhibits glioma angiogenesis and normalizes tumor blood vessels by inducing PDGF-B expression. J. Exp. Med. 209:1611–1627. 10.1084/jem.20111580 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Relton J.K., Beckey V.E., Hanson W.L., and Whalley E.T.. 1997. CP-0597, a selective bradykinin B2 receptor antagonist, inhibits brain injury in a rat model of reversible middle cerebral artery occlusion. Stroke. 28:1430–1436. 10.1161/01.STR.28.7.1430 [DOI] [PubMed] [Google Scholar]
- Rüber T., David B., Lüchters G., Nass R.D., Friedman A., Surges R., Stöcker T., Weber B., Deichmann R., Schlaug G., et al. 2018. Evidence for peri-ictal blood-brain barrier dysfunction in patients with epilepsy. Brain. 141:2952–2965. 10.1093/brain/awy242 [DOI] [PubMed] [Google Scholar]
- Ryu J.K., and McLarnon J.G.. 2009. A leaky blood-brain barrier, fibrinogen infiltration and microglial reactivity in inflamed Alzheimer’s disease brain. J. Cell. Mol. Med. 13(9a, 9A):2911–2925. 10.1111/j.1582-4934.2008.00434.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salameh T.S., Mortell W.G., Logsdon A.F., Butterfield D.A., and Banks W.A.. 2019. Disruption of the hippocampal and hypothalamic blood-brain barrier in a diet-induced obese model of type II diabetes: prevention and treatment by the mitochondrial carbonic anhydrase inhibitor, topiramate. Fluids Barriers CNS. 16:1 10.1186/s12987-018-0121-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saunders N.R., Ek C.J., Habgood M.D., and Dziegielewska K.M.. 2008. Barriers in the brain: a renaissance? Trends Neurosci. 31:279–286. 10.1016/j.tins.2008.03.003 [DOI] [PubMed] [Google Scholar]
- Saunders N.R., Dziegielewska K.M., Unsicker K., and Ek C.J.. 2016. Delayed astrocytic contact with cerebral blood vessels in FGF-2 deficient mice does not compromise permeability properties at the developing blood-brain barrier. Dev. Neurobiol. 76:1201–1212. 10.1002/dneu.22383 [DOI] [PubMed] [Google Scholar]
- Sauvageot C.M., and Stiles C.D.. 2002. Molecular mechanisms controlling cortical gliogenesis. Curr. Opin. Neurobiol. 12:244–249. 10.1016/S0959-4388(02)00322-7 [DOI] [PubMed] [Google Scholar]
- Savolainen H., Meerlo P., Elsinga P.H., Windhorst A.D., Dierckx R.A., Colabufo N.A., van Waarde A., and Luurtsema G.. 2016. P-glycoprotein Function in the Rodent Brain Displays a Daily Rhythm, a Quantitative In Vivo PET Study. AAPS J. 18:1524–1531. 10.1208/s12248-016-9973-3 [DOI] [PubMed] [Google Scholar]
- Sawada N. 2013. Tight junction-related human diseases. Pathol. Int. 63:1–12. 10.1111/pin.12021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schläger C., Körner H., Krueger M., Vidoli S., Haberl M., Mielke D., Brylla E., Issekutz T., Cabañas C., Nelson P.J., et al. 2016. Effector T-cell trafficking between the leptomeninges and the cerebrospinal fluid. Nature. 530:349–353. 10.1038/nature16939 [DOI] [PubMed] [Google Scholar]
- Schlageter N.L., Carson R.E., and Rapoport S.I.. 1987. Examination of blood-brain barrier permeability in dementia of the Alzheimer type with [68Ga]EDTA and positron emission tomography. J. Cereb. Blood Flow Metab. 7:1–8. 10.1038/jcbfm.1987.1 [DOI] [PubMed] [Google Scholar]
- Seidner G., Alvarez M.G., Yeh J.I., O’Driscoll K.R., Klepper J., Stump T.S., Wang D., Spinner N.B., Birnbaum M.J., and De Vivo D.C.. 1998. GLUT-1 deficiency syndrome caused by haploinsufficiency of the blood-brain barrier hexose carrier. Nat. Genet. 18:188–191. 10.1038/ng0298-188 [DOI] [PubMed] [Google Scholar]
- Shechter R., London A., and Schwartz M.. 2013. Orchestrated leukocyte recruitment to immune-privileged sites: absolute barriers versus educational gates. Nat. Rev. Immunol. 13:206–218. 10.1038/nri3391 [DOI] [PubMed] [Google Scholar]
- Shen S., and Zhang W.. 2010. ABC transporters and drug efflux at the blood-brain barrier. Rev. Neurosci. 21:29–53. 10.1515/REVNEURO.2010.21.1.29 [DOI] [PubMed] [Google Scholar]
- Shibata M., Yamada S., Kumar S.R., Calero M., Bading J., Frangione B., Holtzman D.M., Miller C.A., Strickland D.K., Ghiso J., and Zlokovic B.V.. 2000. Clearance of Alzheimer’s amyloid-ss(1-40) peptide from brain by LDL receptor-related protein-1 at the blood-brain barrier. J. Clin. Invest. 106:1489–1499. 10.1172/JCI10498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin Y., Choi S.H., Kim E., Bylykbashi E., Kim J.A., Chung S., Kim D.Y., Kamm R.D., and Tanzi R.E.. 2019. Blood-Brain Barrier Dysfunction in a 3D In Vitro Model of Alzheimer’s Disease. Adv. Sci. (Weinh.). 6:1900962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson I.A., Carruthers A., and Vannucci S.J.. 2007. Supply and demand in cerebral energy metabolism: the role of nutrient transporters. J. Cereb. Blood Flow Metab. 27:1766–1791. 10.1038/sj.jcbfm.9600521 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smyth L.C.D., Rustenhoven J., Scotter E.L., Schweder P., Faull R.L.M., Park T.I.H., and Dragunow M.. 2018. Markers for human brain pericytes and smooth muscle cells. J. Chem. Neuroanat. 92:48–60. 10.1016/j.jchemneu.2018.06.001 [DOI] [PubMed] [Google Scholar]
- Sobue K., Yamamoto N., Yoneda K., Hodgson M.E., Yamashiro K., Tsuruoka N., Tsuda T., Katsuya H., Miura Y., Asai K., and Kato T.. 1999. Induction of blood-brain barrier properties in immortalized bovine brain endothelial cells by astrocytic factors. Neurosci. Res. 35:155–164. 10.1016/S0168-0102(99)00079-6 [DOI] [PubMed] [Google Scholar]
- Sohet F., Lin C., Munji R.N., Lee S.Y., Ruderisch N., Soung A., Arnold T.D., Derugin N., Vexler Z.S., Yen F.T., and Daneman R.. 2015. LSR/angulin-1 is a tricellular tight junction protein involved in blood-brain barrier formation. J. Cell Biol. 208:703–711. 10.1083/jcb.201410131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song J., Zhang X., Buscher K., Wang Y., Wang H., Di Russo J., Li L., Lütke-Enking S., Zarbock A., Stadtmann A., et al. 2017. Endothelial Basement Membrane Laminin 511 Contributes to Endothelial Junctional Tightness and Thereby Inhibits Leukocyte Transmigration. Cell Rep. 18:1256–1269. 10.1016/j.celrep.2016.12.092 [DOI] [PubMed] [Google Scholar]
- Sorokin L. 2010. The impact of the extracellular matrix on inflammation. Nat. Rev. Immunol. 10:712–723. 10.1038/nri2852 [DOI] [PubMed] [Google Scholar]
- Starr J.M., Farrall A.J., Armitage P., McGurn B., and Wardlaw J.. 2009. Blood-brain barrier permeability in Alzheimer’s disease: a case-control MRI study. Psychiatry Res. 171:232–241. 10.1016/j.pscychresns.2008.04.003 [DOI] [PubMed] [Google Scholar]
- Steinman L. 2015. The re-emergence of antigen-specific tolerance as a potential therapy for MS. Mult. Scler. 21:1223–1238. 10.1177/1352458515581441 [DOI] [PubMed] [Google Scholar]
- Stenman J.M., Rajagopal J., Carroll T.J., Ishibashi M., McMahon J., and McMahon A.P.. 2008. Canonical Wnt signaling regulates organ-specific assembly and differentiation of CNS vasculature. Science. 322:1247–1250. 10.1126/science.1164594 [DOI] [PubMed] [Google Scholar]
- Stewart P.A., and Wiley M.J.. 1981. Developing nervous tissue induces formation of blood-brain barrier characteristics in invading endothelial cells: a study using quail--chick transplantation chimeras. Dev. Biol. 84:183–192. 10.1016/0012-1606(81)90382-1 [DOI] [PubMed] [Google Scholar]
- Storck S.E., Meister S., Nahrath J., Meißner J.N., Schubert N., Di Spiezio A., Baches S., Vandenbroucke R.E., Bouter Y., Prikulis I., et al. 2016. Endothelial LRP1 transports amyloid-β(1-42) across the blood-brain barrier. J. Clin. Invest. 126:123–136. 10.1172/JCI81108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stower H. 2018. Meningeal lymphatics in aging and Alzheimer’s disease. Nat. Med. 24:1781. [DOI] [PubMed] [Google Scholar]
- Strazielle N., and Ghersi-Egea J.F.. 2015. Efflux transporters in blood-brain interfaces of the developing brain. Front. Neurosci. 9:21 10.3389/fnins.2015.00021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Streit W.J., Conde J.R., Fendrick S.E., Flanary B.E., and Mariani C.L.. 2005. Role of microglia in the central nervous system’s immune response. Neurol. Res. 27:685–691. [DOI] [PubMed] [Google Scholar]
- Sweeney M.D., and Zlokovic B.V.. 2018. A lymphatic waste-disposal system implicated in Alzheimer’s disease. Nature. 560:172–174. 10.1038/d41586-018-05763-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi K., Yamamura F., and Naito M.. 1989. Differentiation, maturation, and proliferation of macrophages in the mouse yolk sac: a light-microscopic, enzyme-cytochemical, immunohistochemical, and ultrastructural study. J. Leukoc. Biol. 45:87–96. 10.1002/jlb.45.2.87 [DOI] [PubMed] [Google Scholar]
- Tărlungeanu D.C., Deliu E., Dotter C.P., Kara M., Janiesch P.C., Scalise M., Galluccio M., Tesulov M., Morelli E., Sonmez F.M., et al. 2016. Impaired Amino Acid Transport at the Blood Brain Barrier Is a Cause of Autism Spectrum Disorder. Cell. 167:1481–1494.e18. 10.1016/j.cell.2016.11.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tietz S., and Engelhardt B.. 2015. Brain barriers: Crosstalk between complex tight junctions and adherens junctions. J. Cell Biol. 209:493–506. 10.1083/jcb.201412147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ugarte-Berzal E., Berghmans N., Boon L., Martens E., Vandooren J., Cauwe B., Thijs G., Proost P., Van Damme J., and Opdenakker G.. 2018. Gelatinase B/matrix metalloproteinase-9 is a phase-specific effector molecule, independent from Fas, in experimental autoimmune encephalomyelitis. PLoS One. 13:e0197944 10.1371/journal.pone.0197944 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulrich J.D., Huynh T.P., and Holtzman D.M.. 2015. Re-evaluation of the Blood-Brain Barrier in the Presence of Alzheimer’s Disease Pathology. Neuron. 88:237–239. 10.1016/j.neuron.2015.10.008 [DOI] [PubMed] [Google Scholar]
- Umeda K., Matsui T., Nakayama M., Furuse K., Sasaki H., Furuse M., and Tsukita S.. 2004. Establishment and characterization of cultured epithelial cells lacking expression of ZO-1. J. Biol. Chem. 279:44785–44794. 10.1074/jbc.M406563200 [DOI] [PubMed] [Google Scholar]
- van de Haar H.J., Burgmans S., Jansen J.F., van Osch M.J., van Buchem M.A., Muller M., Hofman P.A., Verhey F.R., and Backes W.H.. 2016. Blood-Brain Barrier Leakage in Patients with Early Alzheimer Disease. Radiology. 281:527–535. 10.1148/radiol.2016152244 [DOI] [PubMed] [Google Scholar]
- van Vliet E.A., da Costa Araújo S., Redeker S., van Schaik R., Aronica E., and Gorter J.A.. 2007. Blood-brain barrier leakage may lead to progression of temporal lobe epilepsy. Brain. 130:521–534. 10.1093/brain/awl318 [DOI] [PubMed] [Google Scholar]
- Vanlandewijck M., He L., Mäe M.A., Andrae J., Ando K., Del Gaudio F., Nahar K., Lebouvier T., Laviña B., Gouveia L., et al. 2018. A molecular atlas of cell types and zonation in the brain vasculature. Nature. 554:475–480. 10.1038/nature25739 [DOI] [PubMed] [Google Scholar]
- Vatine G.D., Al-Ahmad A., Barriga B.K., Svendsen S., Salim A., Garcia L., Garcia V.J., Ho R., Yucer N., Qian T., et al. 2017. Modeling Psychomotor Retardation using iPSCs from MCT8-Deficient Patients Indicates a Prominent Role for the Blood-Brain Barrier. Cell Stem Cell. 20:831–843.e5. 10.1016/j.stem.2017.04.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y., Rattner A., Zhou Y., Williams J., Smallwood P.M., and Nathans J.. 2012. Norrin/Frizzled4 signaling in retinal vascular development and blood brain barrier plasticity. Cell. 151:1332–1344. 10.1016/j.cell.2012.10.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y., Jin S., Sonobe Y., Cheng Y., Horiuchi H., Parajuli B., Kawanokuchi J., Mizuno T., Takeuchi H., and Suzumura A.. 2014. Interleukin-1β induces blood-brain barrier disruption by downregulating Sonic hedgehog in astrocytes. PLoS One. 9:e110024 10.1371/journal.pone.0110024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y., Cho C., Williams J., Smallwood P.M., Zhang C., Junge H.J., and Nathans J.. 2018. Interplay of the Norrin and Wnt7a/Wnt7b signaling systems in blood-brain barrier and blood-retina barrier development and maintenance. Proc. Natl. Acad. Sci. USA. 115:E11827–E11836. 10.1073/pnas.1813217115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y., Sabbagh M.F., Gu X., Rattner A., Williams J., and Nathans J.. 2019. Beta-catenin signaling regulates barrier-specific gene expression in circumventricular organ and ocular vasculatures. eLife. 8:e43257 10.7554/eLife.43257 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wimmer I., Tietz S., Nishihara H., Deutsch U., Sallusto F., Gosselet F., Lyck R., Muller W.A., Lassmann H., and Engelhardt B.. 2019. PECAM-1 Stabilizes Blood-Brain Barrier Integrity and Favors Paracellular T-Cell Diapedesis Across the Blood-Brain Barrier During Neuroinflammation. Front. Immunol. 10:711 10.3389/fimmu.2019.00711 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winger R.C., Koblinski J.E., Kanda T., Ransohoff R.M., and Muller W.A.. 2014. Rapid remodeling of tight junctions during paracellular diapedesis in a human model of the blood-brain barrier. J. Immunol. 193:2427–2437. 10.4049/jimmunol.1400700 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winkler E.A., Bell R.D., and Zlokovic B.V.. 2011. Central nervous system pericytes in health and disease. Nat. Neurosci. 14:1398–1405. 10.1038/nn.2946 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wisniewski H.M., Vorbrodt A.W., and Wegiel J.. 1997. Amyloid angiopathy and blood-brain barrier changes in Alzheimer’s disease. Ann. N. Y. Acad. Sci. 826(1 Cerebrovascul):161–172. 10.1111/j.1749-6632.1997.tb48468.x [DOI] [PubMed] [Google Scholar]
- Wosik K., Cayrol R., Dodelet-Devillers A., Berthelet F., Bernard M., Moumdjian R., Bouthillier A., Reudelhuber T.L., and Prat A.. 2007. Angiotensin II controls occludin function and is required for blood brain barrier maintenance: relevance to multiple sclerosis. J. Neurosci. 27:9032–9042. 10.1523/JNEUROSCI.2088-07.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu C., Ivars F., Anderson P., Hallmann R., Vestweber D., Nilsson P., Robenek H., Tryggvason K., Song J., Korpos E., et al. 2009. Endothelial basement membrane laminin alpha5 selectively inhibits T lymphocyte extravasation into the brain. Nat. Med. 15:519–527. 10.1038/nm.1957 [DOI] [PubMed] [Google Scholar]
- Xie L., Kang H., Xu Q., Chen M.J., Liao Y., Thiyagarajan M., O’Donnell J., Christensen D.J., Nicholson C., Iliff J.J., et al. 2013. Sleep drives metabolite clearance from the adult brain. Science. 342:373–377. 10.1126/science.1241224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto M., Guo D.H., Hernandez C.M., and Stranahan A.M.. 2019. Endothelial Adora2a Activation Promotes Blood-Brain Barrier Breakdown and Cognitive Impairment in Mice with Diet-Induced Insulin Resistance. J. Neurosci. 39:4179–4192. 10.1523/JNEUROSCI.2506-18.2019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye X., Wang Y., Cahill H., Yu M., Badea T.C., Smallwood P.M., Peachey N.S., and Nathans J.. 2009. Norrin, frizzled-4, and Lrp5 signaling in endothelial cells controls a genetic program for retinal vascularization. Cell. 139:285–298. 10.1016/j.cell.2009.07.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yousef H., Czupalla C.J., Lee D., Chen M.B., Burke A.N., Zera K.A., Zandstra J., Berber E., Lehallier B., Mathur V., et al. 2019. Aged blood impairs hippocampal neural precursor activity and activates microglia via brain endothelial cell VCAM1. Nat. Med. 25:988–1000. 10.1038/s41591-019-0440-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu Y.J., and Watts R.J.. 2013. Developing therapeutic antibodies for neurodegenerative disease. Neurotherapeutics. 10:459–472. 10.1007/s13311-013-0187-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang S.L., Yue Z., Arnold D.M., Artiushin G., and Sehgal A.. 2018. A Circadian Clock in the Blood-Brain Barrier Regulates Xenobiotic Efflux. Cell. 173:130–139.e10. 10.1016/j.cell.2018.02.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Z., Sagare A.P., Ma Q., Halliday M.R., Kong P., Kisler K., Winkler E.A., Ramanathan A., Kanekiyo T., Bu G., et al. 2015. Central role for PICALM in amyloid-β blood-brain barrier transcytosis and clearance. Nat. Neurosci. 18:978–987. 10.1038/nn.4025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Y., and Nathans J.. 2014. Gpr124 controls CNS angiogenesis and blood-brain barrier integrity by promoting ligand-specific canonical wnt signaling. Dev. Cell. 31:248–256. 10.1016/j.devcel.2014.08.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Y., Wang Y., Tischfield M., Williams J., Smallwood P.M., Rattner A., Taketo M.M., and Nathans J.. 2014. Canonical WNT signaling components in vascular development and barrier formation. J. Clin. Invest. 124:3825–3846. 10.1172/JCI76431 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu J., Li X., Yin J., Hu Y., Gu Y., and Pan S.. 2018. Glycocalyx degradation leads to blood-brain barrier dysfunction and brain edema after asphyxia cardiac arrest in rats. J. Cereb. Blood Flow Metab. 38:1979–1992. 10.1177/0271678X17726062 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zlokovic B.V. 2008. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron. 57:178–201. 10.1016/j.neuron.2008.01.003 [DOI] [PubMed] [Google Scholar]