

Abstract
The homeostasis of the gut-brain axis has been shown to exert several effects on physiological and psychological health. The gut hormones released by enteroendocrine cells scattered throughout the gastrointestinal tract are important signaling molecules within the gut-brain axis. The interaction between gut microbiota and gut hormones has been greatly appreciated in gut-brain cross-talk. The microbiota plays an essential role in modulating many gut-brain axis-related diseases, ranging from gastrointestinal disorders to psychiatric diseases. Similarly, gut hormones also play pleiotropic and important roles in maintaining health, and are key signals involved in gut-brain axis. More importantly, gut microbiota can affect the release and functions of gut hormones. This review highlights the role of gut microbiota in the gut-brain axis and focuses on how microbiota-related gut hormones modulate various physiological functions. Future studies could target the microbiota-hormones-gut brain axis to develop novel therapeutics for different psychiatric and gastrointestinal disorders, such as obesity, anxiety, and depression.
Keywords: Microbiota, Gut hormones, Gut-brain axis, Appetite, Anxiety, Depression
Introduction
Recent preclinical and clinical studies have shown bidirectional interactions within the gut-brain axis. The brain exerts a far-reaching influence on the gastrointestinal tract and vice versa. An aberrant reciprocal gut-brain interaction may influence several conditions, such as inflammation disorders, abnormal responses to stress, altered behaviors, and metabolic disturbances. However, the mechanisms underlying these conditions have not yet been fully understood.
With the development of sequencing technology and bioinformatics analysis, the fields of microbiology and neuroscience have become ever more attractive. During the past decades, increasing evidence shows that the gut microbiota is essential for the development of a large number of diseases, ranging from gastrointestinal disorders to psychiatric diseases. Although the mechanisms underlying the gut and brain interactions are yet to be fully resolved, the concept of the gut-brain axis is becoming more relevant as the gut microbiota can exert considerable influence on the central nervous system (CNS).[1]
Similarly, the gut hormones produced and secreted by the enteroendocrine cells (EECs) also have a wide range of targets and undoubtedly play pleiotropic and important roles in maintaining health. Owing to the complexity in the types and functions, most of these hormones play more than one physiological role, and most of the physiological roles are played by more than one hormone. The EECs coordinate with the nutrients-related signals in the gut and release different gut hormones and then signals to the CNS. Previous studies have shown that a majority of the gut hormones mainly play roles in the central regulation of appetite and food intake; however, recent studies suggest that they are also closely related to other physiological processes, for example, inflammation, that can be linked to different brain disorders, such as anxiety and depression.
Interestingly, studies have documented that the functions of the EECs are modulated by the gut microbiota, whose diversity and composition greatly influence the release of variable gut hormones, including cholecystokinin (CCK), peptide YY (PYY), glucagon-like peptide 1 (GLP-1), and gastric inhibitory polypeptide.[2] It has been proposed that the interactions between the microbiota and EECs may help in explaining the complicated communication between the gut and the brain.[3] New therapeutic strategies may be developed to prevent or alleviate gut-brain axis-related disorders. We retrieved the published literature referring to gut hormones, gut microbiota, and gut-brain axis, then selected the original articles and reviews to summarize. In this review, we provide an overview of gut hormones in the gut-brain axis and focus on how gut microbiota interact with gut hormones.
Microbiota-gut-brain axis
The gut and the brain communicate to regulate health and disease through the brain-gut axis.[4] The CNS modulates the intestinal function via the hypothalamic-pituitary-adrenal axis (HPA) axis, as well as via sympathetic and parasympathetic branches of the autonomic nervous system (ANS). Stressful experience dysregulates the HPA axis significantly and then stimulates the release of neuronal and neuroendocrine signaling molecules, such as norepinephrine, catecholamines, serotonin (5-HT), and cytokines. These molecules are released by the neurons, enterochromaffin cells, and immune cells into the gut lumen, and subsequently, affect the composition and function of the gut microbiota.[5] It has been verified that norepinephrine, whose level increases after stress, can stimulate the proliferation of enteric pathogens.[6] Additionally, ANS is another pathway through which the CNS influences the enteric microbiota. Parasympathetic and vagal outputs to the intestine and stomach are altered after acute stress stimulation,[7] which participate in the modulation of gut functions, including gut motility, permeability, acid secretion, and immune response.[8] All these changes are involved in the modulation of the enteric environment which is associated with microbial colonization in the small intestine and colon.
The CNS receives constant neural and chemical signals from the gut and is responsible for integrating this information and generating appropriate responses to maintain homeostasis. Such effects play important roles in mediating physiological functions, ranging from appetite and food reward to mood response and neurodevelopment. Current evidence indicates that the gut modulates CNS functions primarily through the immune system and neuroimmune mechanisms, neurotransmitters, and ANS, which usually involves the vagus nerve, enteric nervous system, enteroendocrine signaling, and metabolites originating from the gut microbiota.[9] For example, the vagus nerve is the most direct route connecting the gut and the brain. It can detect specific stimuli from the gut depending on the variety of receptors expressed on the vagal afferents and then transmit the gut signals to the brain.
Growing evidence now suggests that microbiota exerts important impact on CNS. Germ-free (GF) mice or mice treated with broad-spectrum antibiotics exhibit significant alteration in neurophysiology and behaviors compared to conventional mice, which suggests the critical roles of gut microbiota in gut-brain axis. Moreover, neurological diseases are associated with dysbiosis of gut microbiota, including neurodegenerative disorders, epilepsy, autism and Parkinson's disease.[10] GF mice received fecal microbiota transplantation (FMT) showed the similar phenotypes as the “donors,” for examples, FMT of GF mice with microbiota from major depressive disorders patients results in increased depression-like behaviors.[11] Colonization with the microbiota from patients with schizophrenia induces schizophrenia-relevant behaviors in mice.[4] All these studies support the links from microbiota to brain.
The gut microbiota affects the brain through several molecules, including neurotransmitter homologs and other metabolites. While on one hand, these molecules are recognized by the receptors located on the host cells and then affect the nerve endings, immune cells, or EECs, which is referred to as microbiota-gut-brain axis. On the other hand, some molecules can cross the intestinal barrier, enter the circulation, cross the blood-brain barrier, and deliver into the brain, which is well known as gut microbiota-brain axis, as shown in Figure 1.
Figure 1.
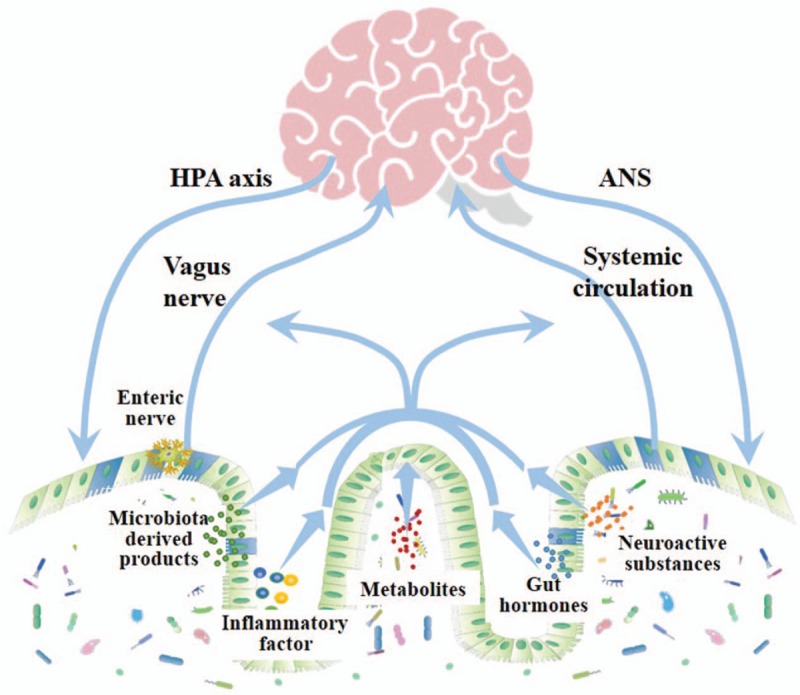
The known bidirectional pathways of interaction between the gut microbiota and brain. The pathways of gut microbiota interact with brain include HPA axis and ANS through which the brain regulates gut microbiota. Through vagus nerve and systemic circulation microbiota-derived products, metabolites, neuroactive substances, gut hormones, and inflammatory factors modulate the function of CNS. HPA: Hypothalamic-pituitary-adrenal axis; ANS: Autonomic nervous system; CNS: Central nervous system.
Neuroactive substances
Neuroactive substances, such as 5-HT, γ-aminobutyric acid (GABA), and tryptophan metabolites play important roles in the CNS modulation. Intriguingly, these molecules could be synthesized and released by the gut microbes. For example, Candida and Escherichia can utilize tryptophan in food and produce 5-HT, and Bacillus can produce dopamine. These microorganisms are considered to influence the CNS by a specific mechanism in which they can produce neurochemicals that are very similar in structure to the neurotransmitters produced by the neuronal cells. However, they are incapable of influencing the brain functions directly as they are unlikely to cross the blood-brain barrier. The pathway through which they affect the brain functions is not known, and it is speculated that the neuroactive substances synthesized by the gut microbes can cross the gut mucosal layer and act on the enteric nervous system.
Microbiota-derived metabolites
A recent study using single-cell sequencing in the brain revealed great alteration in gene expression in the prefrontal cortex of the brain, which suggests that microbes related metabolites may affect CNS, such as neuronal function and fear extinction learning.[12] Microbiota-derived metabolites, such as short-chain fatty acids (SCFAs), tryptophan precursors, and metabolites may exert their central effects through interactions with host cells that express the receptors located in various host tissues (including the gut, muscle, liver, pancreas, and adipose tissues) or immune cells. The fermentable carbohydrates could be decomposed by the gut microbes and converted into SCFAs that include acetate, propionate, and butyrate. They are essential metabolic products of gut microbes and the most well-studied microbe derived metabolites. It has been reported that SCFAs could play roles in glucose homeostasis, reduction of food intake, and modulation of lymphocyte function[13,14] either through G-protein-coupled receptors (GPCRs) or acting as epigenetic modulators of histone deacetylases.[15] For example, GPCRs are widely distributed in various cells and G-protein-coupled receptor (GPR) 41 is expressed in the enteric nerves allowing the signals from SCFAs to reach the nervous system directly.[16] GPR43 was expressed in white adipose tissue, which allows SCFAs act as hormonal molecules and stimulate energy expenditure in the skeletal muscles and liver.[17] More importantly, the effects of different SCFAs on the host physiology are distinct and the results are inconsistent. Acetate has been implied to play a direct role in the central appetite regulation,[18] however, a study has shown that acetate produced by gut microbes could stimulate the parasympathetic nervous system and secretion of ghrelin, thereby increasing food intake.[19]
Endogenous tryptophan is another well-studied microbiota-related metabolite. The microbes in the gastrointestinal tract contribute significantly to the metabolism of dietary tryptophan converting it to indole-3-acetic by the enzymes involved in the indole-3-acetamide pathway, such as tryptophan monooxygenase and indole-3-acetamide hydrolase, and then convert it to 3-methyl indole by decarboxylation of indole acetic acid. These indole compounds have been found to activate the aryl hydrocarbon receptor (AHR), and play extensive roles in the cell cycle, mucosal barrier, and immune regulation.[20,21] Some species (Lactobacillus reuteri, Lactobacillus johnsonii, and Lactobacillus murinus) have been identified to provide indole derivatives from dietary tryptophan, then activate AHR, and participate in the differentiation of T cell.[22] Lamas et al[23] reported that mice lacking bacteria capable of catabolizing tryptophan were much more susceptible to colitis. As it is well known that inflammation plays a critical role in various CNS diseases, including Parkinson's disease, multiple sclerosis, amyotrophic lateral sclerosis, and Alzheimer's disease,[24] these studies make substantial contributions to provide novel strategies for the treatment of these diseases.
Microbiota-derived products
Microbiota-derived products, such as lipopolysaccharide (LPS), LPS binding protein (LBP), peptidoglycan, and flagellin are also the key molecules that deliver messages to the gut-brain axis. LPS is a specific component of the cellular wall of gram-negative (G−) bacteria and is released once the G− bacteria die and the cell walls are destroyed. Toll-like receptors 4, which are widely expressed in the immune cells, such as B cells and dendritic cells, can recognize LPS and induce cytokine production, including tumor necrosis factor-α, interleukin (IL)-6, and IL-1. Furthermore, this immune response has been reported to be associated with anxiety, depression, or memory impairment.[25] Polysaccharide A is a kind of product secreted from the microbiota. It has been reported that PSA from B. fragilis can protect against CNS inflammation through a toll-like receptors 2 dependent pathway.[26]
Gut hormones in the gut-brain crosstalk
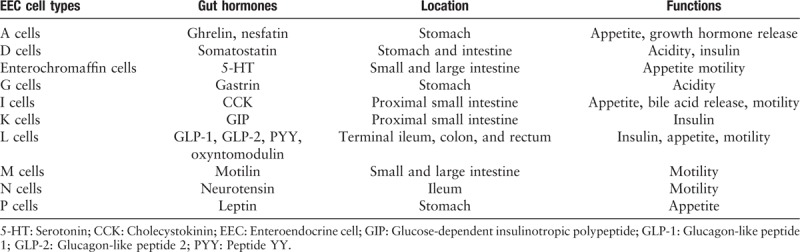
EECs are found to scatter throughout the gastrointestinal tract although they represent only 1% of the epithelial cells in the gastrointestinal tract. They can release a variety of gut hormones in response to diet-related stimuli and play key roles in the control of gut motility, appetite, and hormone release. EECs are generally classified into ten different types according to the primary hormone produced by them[27] [Table 1]. All these EECs are sensory cells that coordinate signal changes between the gut content the host responses, such as regulation of food intake, insulin secretion, and behavioral adaption.
Table 1:
Main enteroendocrine cell types, gut hormones, and functions.
Gut hormones exert a wide variety of functions involved in diverse tissues, ranging from the gastrointestinal tract to the CNS. There are more than 20 active hormones found so far, with overlapping actions and targets. Gut hormones are usually studied for their functions in the detection of nutrients, mediation of digestion, and modulation of insulin release. Recently, it has been reported that gut hormones are also key regulators of anxiety and depression.[28]
Gut hormones and metabolic control
CNS has always been considered as the most important organ to govern metabolic control. The hypothalamus expresses a multitude of nutrient sensors and hormone receptors and directly receive circulating nutrient and hormonal signals. Several gut hormones are found to be the key signals involved in the gut-brain crosstalk and human energy metabolism, such as ghrelin, PYY, GLP-1, glucose-dependent insulinotropic polypeptide (GIP), and CCK,[29] all of which are released in response to food ingestion and act to modulate different functions of the CNS.
Ghrelin is primarily produced by the proximal X/A-like cells in the stomach and is released during food restriction. Its level was identified to decline sharply following gastrectomy.[30] Ghrelin is now the only peripherally-derived hormone to increase appetite and food intake. It can play its role through ghrelin receptors expressed on the vagal afferents and neurons in the nodose ganglion which signals to the brain. Additionally, ghrelin can cross the blood-brain barrier and reach the hypothalamus where the food intake regulatory center is present.[31] The ghrelin receptors are widely expressed in various areas of the brain, including the hippocampus, arcuate nucleus, ventromedial nucleus, substantia nigra, dentate gyrus, ventral tegmental area, and pituitary.[32] Ghrelin binds to the growth hormone secretagogue receptor 1a isoform (GHS-R1a) and plays various roles in food intake and memory regulation.[33] Our previous study in obese patients included 30 individuals who underwent laparoscopic sleeve gastrectomy (LSG). There was a significant reduction in the plasma ghrelin and amplitude of low frequency fluctuation (ALFF) value in the hippocampus one-month post-surgery, and a correlation analysis showed that the reduction in the ALFF values of the hippocampus positively correlated with that of ghrelin. This suggests that ghrelin plays an important role in the normalization of hippocampal activity post-LSG.[34] Another study showed that LSG significantly reduced the craving for high-calorie food and activation of the right dorsolateral prefrontal cortex following high-calorie food stimulation. In addition, a reduction in ghrelin was positively correlated with a reduction in the activity of the right dorsolateral prefrontal cortex, suggesting that ghrelin might be an important factor leading to the reduction in appetite post-LSG.[35] These findings in subjects undergoing LSG demonstrate that ghrelin could be a key signaling molecule governing the communication between the gastrointestinal tract and the CNS.
CCK is a gut-derived peptide hormone that is produced and released by the enteroendocrine I cells located in the mucosal epithelium of the small intestine. It can stimulate the digestion of some nutrients, including fats and proteins. CCK binds to the GPCRs, often known as CCK-A and CCK-B, which are mainly expressed in the gut and brain, respectively.[36] CCK then inhibits food intake via interaction with leptin in the vagal afferents and the brain. Additionally, CCK can cross the blood-brain barrier and directly bind to CCK-A receptors within the hypothalamus and hindbrain, which trigger its action to regulate appetite.[37]
GLP-1 and PYY are secreted by the enteroendocrine L cells. They promote satiety and have an inhibitory effect on energy intake. GLP-1 is released mainly in response to carbohydrates and fats. GLP-1 receptors are widely distributed in different tissues, such as the gut, kidney, pancreatic β-cells, vagus nerve, and hypothalamus. Recently, a large number of GLP-1 receptor agonists are used to treat type 2 diabetes mellitus for their ability to reduce blood glucose level, which is related to its role in the pancreatic β-cells. As to its effect on food intake in CNS, GLP-1 is rapidly degraded in a very short time after its release, and hence, it is almost impossible that it is capable of reaching to the hypothalamus and brainstem. It is more likely to activate the receptors located on enteric nervous system (ENS) or vagal afferent terminals close to the site of secretion.[38] PYY is primarily produced in the ileum and colon in response to lipids and other nutrients, such as proteins and carbohydrates. It is a well-studied member of the neuropeptide hormone group which takes part in the central and peripheral control of food intake within the brain-gut axis. There are five different types of receptors (Y1, Y2, Y3, Y4, and Y5) expressed in different areas of the CNS, such as the hypothalamus, NTS, spinal cord, and pons. PYY binds to and activates the Y2 receptors located in the hypothalamus and then participates in reducing food intake.[39]
GIP is mainly produced by the K cells scattered throughout the duodenum and the jejunum. Initial studies have shown that GIP could bind to its receptors and stimulate the release of insulin in the pancreatic β-cells, which play an important role in glucose homeostasis.[40] However, its role in food intake is debated as there was evidence suggesting it might rather promote adiposity.[41] A recent study identified that the neurons expressing the GIP receptor play a key role in food intake control in mouse and human brain,[42] which provide novel insights into the functions of GIP.
Gut hormones and mood disorders
It is now well accepted that a majority of the gut hormones play an important role in the regulation of food intake in the CNS. Most intriguingly, obesity and mood disorders often tend to co-exist.[43] There are several gut hormones, including 5-HT, NPY, GLP-1, CCK, and ghrelin, that are identified with known roles in mood disorders, such as anxiety and depression.
5-HT is mainly produced by the enterochromaffin cells distributed throughout the gastrointestinal tract, including the stomach, small, and large intestine. Peripheral 5-HT participates in the modulation of intestinal motility, pain perception, electrolyte secretion, cardiac functions, vascular tone, organ development, and inflammation. Additionally, in CNS it can also act as a neurotransmitter to regulate mood, sleep, and appetite.[44] Although the peripheral and central 5-HT are synthesized in different ways and separated by the blood-brain barrier, they are observed closely linked to CNS functions. First, tryptophan, an essential amino acid and the precursor of 5-HT, has many important implications on the CNS and ENS functions, and thus, the brain-gut axis signaling. The availability of tryptophan in the CNS is largely affected by its metabolism via the kynurenine pathway in the peripheral tissues.[45] Second, 5-HT derived from enterochromaffin cells exerts critical effects on immune regulation. A previous study observed that the injection of endotoxin could stimulate the release of 5-HT from the platelets to the plasma. More importantly, 5-HT was thought to exert key effects on innate and adaptive immunity. It can promote the secretion of cytokines from lymphocytes and monocytes[46,47] and participate in the modulation of CNS functions. Third, 5-HT released from the enterochromaffin cells leads to alteration of the vagal afferent activity and then potentially affects the gut-brain signaling. A characterized example is that a rapid release of 5-HT during chemotherapy induces nausea and emesis, which is largely dependent on the activation of the vagal afferents in the gut.[48] All this evidence suggests that 5-HT is largely involved in the modulation of the gut-brain axis.
The neuropeptide Y family (NPY), including NPY, PYY, and pancreatic polypeptide (PP), affects stress-related disorders, neuroprotection, neuroinflammation, and neurogenesis. The Y4 receptors can be activated by NPY and PP, and then they participate in the modulation of anxiety and depression.[49]
GLP-1, best known as a hormone stimulating glucose-dependent insulin secretion, also responses to stress through the activation of the GLP-1 receptor, for example, endogenous or exogenous glucocorticoids reduce the bioavailability of GLP-1.[50]
CCK, as mentioned above, is abundantly produced in the peripheral nervous system and CNS, and mainly participates in the regulation of food intake. It has also been reported that the CCK levels were positively correlated with increased anxiety-like behaviors in both humans and mice.[51] It seems that CCK modulates mood disorders through other neurotransmitters, including glutamate, dopamine, acetylcholine, and GABA, all of which play key roles in emotional behaviors.[52]
Ghrelin, best known for its adipogenic and orexigenic effects, was also identified as a regulator of stress response, anxiety, and depression. Evidence from rodents has shown that various stress factors, such as restraint stress and social defeat, could increase ghrelin levels. More interestingly, a recent study has demonstrated that an elevated ghrelin level following hunger instigates adaptation to stress.[53] Repeated injection of ghrelin receptor agonists in animals increased the fear memory induced by stress, whereas its antagonists inhibited fear memory, implying that ghrelin promotes anxiety and depression-like behaviors in rodents.[54] In our previous study, the anxiety scores were decreased post-LSG accompanied by a sharp reduction in plasma ghrelin, which also provides evidence to support this view.[34]
Microbiota and the gut hormones
Although the precise pathways through which the gut microbiota communicates with the hormones have not yet been deciphered, growing pieces of evidence now suggest that microbiota is a key factor involved in the metabolism of hormones, and host hormones greatly influence the microbiota [Figure 2]. Homeostatic mechanisms influence several gut-brain axis-related host physiological processes, such as appetite, immune response, stress response, and metabolism.
Figure 2.
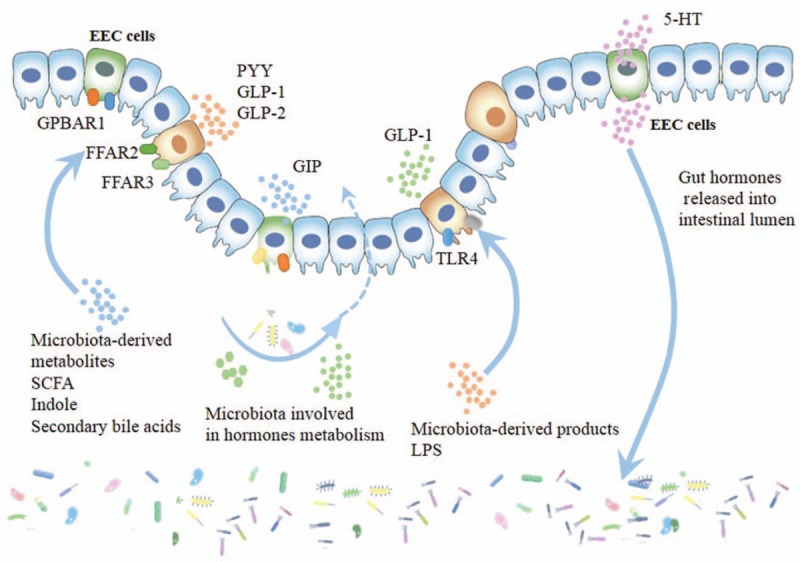
Interactions between gut microbiota and EEC cells. The gut microbiota affects EEC cells through microbiota-derived products (eg, LPS), microbiota-derived metabolites, including SCFA, indole and secondary bile acids, in addition, some microbiota is involved in hormones metabolism. EEC cells can also release gut hormones, and part of hormones released into gastrointestinal lumen can influence gut microbiota. EEC: Enteroendocrine cells; GPBAR1: G protein-coupled bile acid receptors 1; FFAR2: Free fatty acid receptor 2; FFAR3: Free fatty acid receptor 3; PYY: Peptide YY; GLP-1: Glucagon-like peptide 1; GLP-2: Glucagon-like peptide 2; GIP: Glucose-dependent insulinotropic polypeptide; TLR4: Toll-like receptor 4; 5-HT: Serotonin; LPS: Lipopolysaccharide; SCFA: Short chain fatty acid; GIP: Glucose-dependent insulinotropic polypeptide.
The gut microbiota may affect the host hormones directly since some bacteria can produce or metabolize these hormones. Gut bacteria were found to produce neurohormones; for example, dopamine can be produced by some bacteria, such as Bacillus and Serratia. It has been reported that many hormones were altered significantly in GF mice as compared to those in the conventionally raised mice. The levels of norepinephrine, 5-HT, and dopamine were decreased, while those of GLP-1, corticosterone, and adrenocorticosterone were increased.[3] The level of plasma leptin decreased after treatment with antibiotics.[55] A series of studies have provided integral evidence about the gut microbiota, host hormones, and gut-brain axis. Children with obesity or overweight were administered with prebiotics for 16 weeks. Bifidobacterium in the gut microbiota increased and Bacteroides vulgatus decreased significantly. Fasting ghrelin level increased and behavioral tests showed that prebiotic supplementation resulted in lower prospective food consumption and more feeling of fullness. Anthropometric results showed that the body weight z-score and percent body fat were reduced by 2.4%.[56,57] An evolutionary-oriented study hypothesized that a variety of hormone-metabolism-related enzymes might have evolved from the bacterial genes.[56,57] In addition, microbes may regulate the hormones indirectly since they may modulate the functions of the adrenal cortex and inflammatory response, all of which are closely related to the CNS and the gastrointestinal tract.
Apart from that bacteria directly or indirectly affect the host hormones, the gut microbiota can also promote the release of gut hormones from the EECs through metabolites or bacterial components. LPS is identified to bind to Toll-like receptor 4 expressed in the L cells. They promote the secretion of GLP-1 in mice[58] and trigger the release of CCK in vitro.[59] SCFA produced by the bacteria can activate G-coupled receptor FFAR2 and FFAR3 in the L-cells, elevate intracellular Ca2+ and stimulate GLP-1 secretion.[60] Indole is a product of tryptophan produced by bacteria, and it has been observed to elicit a rapid stimulation of GLP-1 release in vitro by inhibiting voltage-gated K+ channels and increasing Ca2+ influx.[61]
The gut hormones also greatly affect the microbiota. For example, 5-HT released by the enterochromaffin cells is not only secreted towards the intestinal submucosa, but also towards the gut lumen,[62] which may lead to alterations in the gut microbiota. Our previous study showed that in a depression model (generally accompanied by high cortisone), the gut microbiota exhibited a specific signature, including low bacterial diversity, simple bacterial network, and high abundance of pathogens. Interestingly, this alteration in the microbiota was largely ameliorated by the classical antidepressant fluoxetine.[63] It is well known that fluoxetine inhibits the reuptake of 5-HT and increases serotonergic neurotransmission. Although the alteration of 5-HT is not derived from the EECs, these findings provide clues to link the gut hormones and the microbiota.
Although it is presently unclear how the gut interacts with the brain, this field still receives great attention because the roles of gut microbiota in several diseases have been identified. Given the roles of gut microbiota in modulating gut hormones and thus gut hormones in the gut-brain axis, we may focus on a microbiota-gut hormones-gut brain axis mediating different diseases.
Funding
This work was supported by grants from the National Natural Science Foundation of China (Nos. 81730016 and 81900483).
Conflicts of interest
None.
Footnotes
How to cite this article: Sun LJ, Li JN, Nie YZ. Gut hormones in microbiota-gut-brain cross-talk. Chin Med J 2020;133:826–833. doi: 10.1097/CM9.0000000000000706
References
- 1.Wang HX, Wang YP. Gut Microbiota-brain Axis. Chin Med J - 2016; 129:2373–2380. doi: 10.4103/0366-6999.190667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Olivares M, Schuppel V, Hassan AM, Beaumont M, Neyrinck AM, Bindels LB, et al. The potential role of the dipeptidyl Peptidase-4-like activity from the gut microbiota on the host health. Front Microbiol 2018; 9:1900.doi: 10.3389/fmicb.2018.01900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Neuman H, Debelius JW, Knight R, Koren O. Microbial endocrinology: the interplay between the microbiota and the endocrine system. FEMS Microbiol Rev 2015; 39:509–521. doi: 10.1093/femsre/fuu010. [DOI] [PubMed] [Google Scholar]
- 4.Zheng P, Zeng B, Liu M, Chen J, Pan J, Han Y, et al. The gut microbiome from patients with schizophrenia modulates the glutamate-glutamine-GABA cycle and schizophrenia-relevant behaviors in mice. Sci Adv 2019; 5:u8317.doi: 10.1126/sciadv.aau8317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lyte M. Microbial endocrinology in the microbiome-gut-brain axis: how bacterial production and utilization of neurochemicals influence behavior. PLoS Pathog 2013; 9:e1003726.doi: 10.1371/journal.ppat.1003726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hughes DT, Sperandio V. Inter-kingdom signalling: communication between bacteria and their hosts. Nat Rev Microbiol 2008; 6:111–120. doi: 10.1038/nrmicro1836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mayer EA. The neurobiology of stress and gastrointestinal disease. Gut 2000; 47:861–869. doi: 10.1136/gut.47.6.861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mayer EA, Tillisch K, Gupta A. Gut/brain axis and the microbiota. J Clin Invest 2015; 125:926–938. doi: 10.1172/JCI76304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cryan JF, O’Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, Boehme M, et al. The microbiota-gut-brain axis. Physiol Rev 2019; 99:1877–2013. doi: 10.1152/physrev. 00018.2018. [DOI] [PubMed] [Google Scholar]
- 10.Fung TC, Olson CA, Hsiao EY. Interactions between the microbiota, immune and nervous systems in health and disease. Nat Neurosci 2017; 20:145–155. doi: 10.1038/nn.4476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zheng P, Zeng B, Zhou C, Liu M, Fang Z, Xu X, et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host's metabolism. Mol Psychiatry 2016; 21:786–796. doi: 10.1038/mp.2016.44. [DOI] [PubMed] [Google Scholar]
- 12.Chu C, Murdock MH, Jing D, Won TH, Chung H, Kressel AM, et al. The microbiota regulate neuronal function and fear extinction learning. Nature 2019; 574:543–548. doi: 10.1038/s41586-019-1644-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Smith PM, Howitt MR, Panikov N, Michaud M, Gallini CA, Bohlooly YM, et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013; 341:569–573. doi: 10.1126/science.1241165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Layden BT, Angueira AR, Brodsky M, Durai V, Lowe WJ. Short chain fatty acids and their receptors: new metabolic targets. Transl Res 2013; 161:131–140. doi: 10.1016/j.trsl.2012.10.007. [DOI] [PubMed] [Google Scholar]
- 15.Stilling RM, Dinan TG, Cryan JF. Microbial genes, brain & behaviour - epigenetic regulation of the gut-brain axis. Genes Brain Behav 2014; 13:69–86. doi: 10.1111/gbb.12109. [DOI] [PubMed] [Google Scholar]
- 16.Nohr MK, Pedersen MH, Gille A, Egerod KL, Engelstoft MS, Husted AS, et al. GPR41/FFAR3 and GPR43/FFAR2 as cosensors for short-chain fatty acids in enteroendocrine cells vs. FFAR3 in enteric neurons and FFAR2 in enteric leukocytes. Endocrinology 2013; 154:3552–3564. doi: 10.1210/en.2013-1142. [DOI] [PubMed] [Google Scholar]
- 17.Kimura I, Ozawa K, Inoue D, Imamura T, Kimura K, Maeda T, et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat Commun 2013; 4:1829.doi: 10.1038/ncomms2852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Frost G, Sleeth ML, Sahuri-Arisoylu M, Lizarbe B, Cerdan S, Brody L, et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat Commun 2014; 5:3611.doi: 10.1038/ncomms4611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Perry RJ, Peng L, Barry NA, Cline GW, Zhang D, Cardone RL, et al. Acetate mediates a microbiome-brain-beta-cell axis to promote metabolic syndrome. Nature 2016; 534:213–217. doi: 10.1038/nature18309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zelante T, Iannitti RG, Cunha C, De Luca A, Giovannini G, Pieraccini G, et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013; 39:372–385. doi: 10.1016/j.immuni. 2013.08.003. [DOI] [PubMed] [Google Scholar]
- 21.Hubbard TD, Murray IA, Perdew GH. Indole and tryptophan metabolism: endogenous and dietary routes to Ah receptor activation. Drug Metab Dispos 2015; 43:1522–1535. doi: 10.1124/dmd.115.064246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cervantes-Barragan L, Chai JN, Tianero MD, Di Luccia B, Ahern PP, Merriman J, et al. Lactobacillus reuteri induces gut intraepithelial CD4(+)CD8alphaalpha(+) T cells. Science 2017; 357:806–810. doi: 10.1126/science.aah5825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lamas B, Richard ML, Leducq V, Pham HP, Michel ML, Da CG, et al. CARD9 impacts colitis by altering gut microbiota metabolism of tryptophan into aryl hydrocarbon receptor ligands. Nat Med 2016; 22:598–605. doi: 10.1038/nm.4102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chitnis T, Weiner HL. CNS inflammation and neurodegeneration. J Clin Invest 2017; 127:3577–3587. doi: 10.1172/JCI90609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wright CE, Strike PC, Brydon L, Steptoe A. Acute inflammation and negative mood: mediation by cytokine activation. Brain Behav Immun 2005; 19:345–350. doi: 10.1016/j.bbi.2004.10.003. [DOI] [PubMed] [Google Scholar]
- 26.Wang Y, Telesford KM, Ochoa-Reparaz J, Haque-Begum S, Christy M, Kasper EJ, et al. An intestinal commensal symbiosis factor controls neuroinflammation via TLR2-mediated CD39 signalling. Nat Commun 2014; 5:4432.doi: 10.1038/ncomms5432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gribble FM, Reimann F. Enteroendocrine cells: chemosensors in the intestinal epithelium. Annu Rev Physiol 2016; 78:277–299. doi: 10.1146/annurev-physiol-021115-105439. [DOI] [PubMed] [Google Scholar]
- 28.Lach G, Schellekens H, Dinan TG, Cryan JF. Anxiety, depression, and the microbiome: a role for gut peptides. Neurotherapeutics 2018; 15:36–59. doi: 10.1007/s13311-017-0585-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Clemmensen C, Muller TD, Woods SC, Berthoud HR, Seeley RJ, Tschop MH. Gut-brain cross-talk in metabolic control. Cell 2017; 168:758–774. doi: 10.1016/j.cell.2017.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jeon TY, Lee S, Kim HH, Kim YJ, Son HC, Kim DH, et al. Changes in plasma ghrelin concentration immediately after gastrectomy in patients with early gastric cancer. J Clin Endocrinol Metab 2004; 89:5392–5396. doi: 10.1210/jc.2004-0872. [DOI] [PubMed] [Google Scholar]
- 31.Stengel A, Goebel M, Wang L, Tache Y. Ghrelin, des-acyl ghrelin and nesfatin-1 in gastric X/A-like cells: role as regulators of food intake and body weight. Peptides 2010; 31:357–369. doi: 10.1016/j.peptides.2009.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zigman JM, Jones JE, Lee CE, Saper CB, Elmquist JK. Expression of ghrelin receptor mRNA in the rat and the mouse brain. J Comp Neurol 2006; 494:528–548. doi: 10.1002/cne.20823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Nakazato M, Murakami N, Date Y, Kojima M, Matsuo H, Kangawa K, et al. A role for ghrelin in the central regulation of feeding. Nature 2001; 409:194–198. doi: 10.1038/35051587. [DOI] [PubMed] [Google Scholar]
- 34.Zhang Y, Ji G, Li G, Hu Y, Liu L, Jin Q, et al. Ghrelin reductions following bariatric surgery were associated with decreased resting state activity in the hippocampus. Int J Obes (Lond) 2019; 43:842–851. doi: 10.1038/s41366-018-0126-x. [DOI] [PubMed] [Google Scholar]
- 35.Li G, Ji G, Hu Y, Liu L, Jin Q, Zhang W, et al. Reduced plasma ghrelin concentrations are associated with decreased brain reactivity to food cues after laparoscopic sleeve gastrectomy. Psychoneuroendocrinology 2019; 100:229–236. doi: 10.1016/j.psyneuen.2018.10.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rasmussen BA, Breen DM, Lam TK. Lipid sensing in the gut, brain and liver. Trends Endocrinol Metab 2012; 23:49–55. doi: 10.1016/j.tem.2011.11.001. [DOI] [PubMed] [Google Scholar]
- 37.Blevins JE, Stanley BG, Reidelberger RD. Brain regions where cholecystokinin suppresses feeding in rats. Brain Res 2000; 860:1–10. doi: 10.1016/s0006-8993(99)02477-4. [DOI] [PubMed] [Google Scholar]
- 38.Abbott CR, Monteiro M, Small CJ, Sajedi A, Smith KL, Parkinson JR, et al. The inhibitory effects of peripheral administration of peptide YY(3-36) and glucagon-like peptide-1 on food intake are attenuated by ablation of the vagal-brainstem-hypothalamic pathway. Brain Res 2005; 1044:127–131. doi: 10.1016/j.brainres.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 39.Batterham RL, Cowley MA, Small CJ, Herzog H, Cohen MA, Dakin CL, et al. Gut hormone PYY(3-36) physiologically inhibits food intake. Nature 2002; 418:650–654. doi: 10.1038/nature00887. [DOI] [PubMed] [Google Scholar]
- 40.Dupre J, Ross SA, Watson D, Brown JC. Stimulation of insulin secretion by gastric inhibitory polypeptide in man. J Clin Endocrinol Metab 1973; 37:826–828. doi: 10.1210/jcem-37-5-826. [DOI] [PubMed] [Google Scholar]
- 41.Miyawaki K, Yamada Y, Ban N, Ihara Y, Tsukiyama K, Zhou H, et al. Inhibition of gastric inhibitory polypeptide signaling prevents obesity. Nat Med 2002; 8:738–742. doi: 10.1038/nm727. [DOI] [PubMed] [Google Scholar]
- 42.Adriaenssens AE, Biggs EK, Darwish T, Tadross J, Sukthankar T, Girish M, et al. Glucose-dependent insulinotropic polypeptide receptor-expressing cells in the hypothalamus regulate food intake. Cell Metab 2019; 30:987–996. doi: 10.1016/j.cmet.2019.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Milaneschi Y, Simmons WK, van Rossum E, Penninx BW. Depression and obesity: evidence of shared biological mechanisms. Mol Psychiatry 2019; 24:18–33. doi: 10.1038/s41380-018-0017-5. [DOI] [PubMed] [Google Scholar]
- 44.Herr N, Bode C, Duerschmied D. The Effects of serotonin in immune cells. Front Cardiovasc Med 2017; 4:48.doi: 10.3389/fcvm.2017.00048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kennedy PJ, Cryan JF, Dinan TG, Clarke G. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology 2017; 112:399–412. doi: 10.1016/j.neuropharm.2016.07.002. [DOI] [PubMed] [Google Scholar]
- 46.Iken K, Chheng S, Fargin A, Goulet AC, Kouassi E. Serotonin upregulates mitogen-stimulated B lymphocyte proliferation through 5-HT1A receptors. Cell Immunol 1995; 163:1–9. doi: 10.1006/cimm.1995.1092. [DOI] [PubMed] [Google Scholar]
- 47.Durk T, Panther E, Muller T, Sorichter S, Ferrari D, Pizzirani C, et al. 5-Hydroxytryptamine modulates cytokine and chemokine production in LPS-primed human monocytes via stimulation of different 5-HTR subtypes. Int Immunol 2005; 17:599–606. doi: 10.1093/intimm/dxh242. [DOI] [PubMed] [Google Scholar]
- 48.Gale JD. Serotonergic mediation of vomiting. J Pediatr Gastroenterol Nutr 1995; 21: Suppl 1: S22–S28. doi: 10.1097/00005176-199501001-00008. [DOI] [PubMed] [Google Scholar]
- 49.Verma D, Wood J, Lach G, Herzog H, Sperk G, Tasan R. Hunger promotes fear extinction by activation of an amygdala microcircuit. Neuropsychopharmacology 2016; 41:431–439. doi: 10.1038/npp.2015.163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ghosal S, Myers B, Herman JP. Role of central glucagon-like peptide-1 in stress regulation. Physiol Behav 2013; 122:201–207. doi: 10.1016/j.physbeh.2013.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Del BC, Lutz PE, Le Merrer J, Koebel P, Kieffer BL. Cholecystokinin knock-down in the basolateral amygdala has anxiolytic and antidepressant-like effects in mice. Neuroscience 2012; 218:185–195. doi: 10.1016/j.neuroscience.2012.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Rezayat M, Roohbakhsh A, Zarrindast MR, Massoudi R, Djahanguiri B. Cholecystokinin and GABA interaction in the dorsal hippocampus of rats in the elevated plus-maze test of anxiety. Physiol Behav 2005; 84:775–782. doi: 10.1016/j.physbeh.2005.03.002. [DOI] [PubMed] [Google Scholar]
- 53.Hirano Y, Masuda T, Naganos S, Matsuno M, Ueno K, Miyashita T, et al. Fasting launches CRTC to facilitate long-term memory formation in Drosophila. Science 2013; 339:443–446. doi: 10.1126/science.1227170. [DOI] [PubMed] [Google Scholar]
- 54.Meyer RM, Burgos-Robles A, Liu E, Correia SS, Goosens KA. A ghrelin-growth hormone axis drives stress-induced vulnerability to enhanced fear. Mol Psychiatry 2014; 19:1284–1294. doi: 10.1038/mp.2013.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lam V, Su J, Koprowski S, Hsu A, Tweddell JS, Rafiee P, et al. Intestinal microbiota determine severity of myocardial infarction in rats. FASEB J 2012; 26:1727–1735. doi: 10.1096/fj.11-197921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Nicolucci AC, Hume MP, Martinez I, Mayengbam S, Walter J, Reimer RA. Prebiotics reduce body fat and alter intestinal microbiota in children who are overweight or with obesity. Gastroenterology 2017; 153:711–722. doi: 10.1053/j.gastro.2017.05.055. [DOI] [PubMed] [Google Scholar]
- 57.Hume MP, Nicolucci AC, Reimer RA. Prebiotic supplementation improves appetite control in children with overweight and obesity: a randomized controlled trial. Am J Clin Nutr 2017; 105:790–799. doi: 10.3945/ajcn.116.140947. [DOI] [PubMed] [Google Scholar]
- 58.Iyer LM, Aravind L, Coon SL, Klein DC, Koonin EV. Evolution of cell-cell signaling in animals: did late horizontal gene transfer from bacteria have a role? Trends Genet 2004; 20:292–299. doi: 10.1016/j.tig.2004.05.007. [DOI] [PubMed] [Google Scholar]
- 59.Bogunovic M, Dave SH, Tilstra JS, Chang DT, Harpaz N, Xiong H, et al. Enteroendocrine cells express functional Toll-like receptors. Am J Physiol Gastrointest Liver Physiol 2007; 292:G1770–G1783. doi: 10.1152/ajpgi.00249.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tolhurst G, Heffron H, Lam YS, Parker HE, Habib AM, Diakogiannaki E, et al. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012; 61:364–371. doi: 10.2337/db11-1019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Chimerel C, Emery E, Summers DK, Keyser U, Gribble FM, Reimann F. Bacterial metabolite indole modulates incretin secretion from intestinal enteroendocrine L cells. Cell Rep 2014; 9:1202–1208. doi: 10.1016/j.celrep.2014.10.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Patel BA. Electroanalytical approaches to study signaling mechanisms in the gastrointestinal tract. Neurogastroenterol Motil 2011; 23:595–605. doi: 10.1111/j.1365-2982.2011.01708.x. [DOI] [PubMed] [Google Scholar]
- 63.Sun L, Zhang H, Cao Y, Wang C, Zhao C, Wang H, et al. Fluoxetine ameliorates dysbiosis in a depression model induced by chronic unpredicted mild stress in mice. Int J Med Sci 2019; 16:1260–1270. doi: 10.7150/ijms.37322. eCollection 2019. [DOI] [PMC free article] [PubMed] [Google Scholar]