

Abstract
This chapter describes a variety of pathogens found in the environment that are capable of infecting humans and causing disease. Different classes of pathogens are discussed including bacteria, parasites and viruses. For each pathogen, data are provided on the incidence of the organism in a particular environmental matrix such as water, food or municipal waste. In addition, we discuss: the disease caused by the particular microbe; the mode of transmission; the incubation time needed within the host prior to the onset of disease; and the duration of illness. Fate and transport of pathogens in the environment are also described. A variety of bacterial pathogens are discussed including waterborne and waterbased bacteria. Parasites discussed include protozoa, nematodes, cestodes and trematodes. For viruses, both enteric and respiratory viruses are included.
Keywords: bacterial pathogens, cestodes, enteric viruses, human pathogens, nematodes, protozoan pathogens, respiratory viruses, trematodes, waterbased pathogens, waterborne pathogens
22.1. Environmentally Transmitted Pathogens
Although humans are continually exposed to a vast array of microorganisms in the environment, only a small proportion of these microbes are capable of interacting with the host in such a manner that infection and disease will result. Disease-causing microorganisms are called pathogens. Infection is the process in which the microorganism multiplies or grows in or on the host. Infection does not necessarily result in disease, since it is possible for the organism to grow in or on the host without producing an illness (see Chapter 23). In the case of enteric infections (i.e., diarrhea) caused by Salmonella, only half of the individuals infected develop clinical signs of illness. A frank pathogen is a microorganism capable of producing disease in both normal healthy and immunocompromised persons. Opportunistic pathogens are usually capable of causing infections only in immunocompromised individuals such as burn patients, patients taking antibiotics, those with impaired immune systems or elderly patients with diabetes. Opportunistic pathogens are common in the environment, and may be present in the human gut or skin without causing disease.
To cause illness, the pathogen must usually first grow within or on the host. The time between infection and the appearance of clinical signs and symptoms (diarrhea, fever, rash, etc.) is the incubation time (Table 22.1 ). This may range from as short as 6 to 12 hours in the case of norovirus diarrhea, or up to 30 to 60 days for hepatitis A virus, which causes liver disease. At any time during infection the pathogen may be released into the environment by the host in feces, urine or respiratory secretions. Although the maximum release may occur at the height of the disease, it can also precede the first signs of clinical illness. In the case of hepatitis A virus, the maximum excretion in the feces occurs before the onset of signs of clinical illness. The concentration of organisms released into the environment varies with the type of organism and the route of transmission (Table 22.2 ). The concentration of enteric viruses during gastroenteritis may be as high as 1010–1012 per gram of feces.
Table 22.1.
Incubation Time for Common Enteric Pathogens
| Agent | Incubation Period | Modes of Transmission | Duration of Illness |
|---|---|---|---|
| Adenovirus | 8–10 days | Fecal–oral–respiratory | 8 days |
| Campylobader jejuni | 3–5 days | Food ingestion, direct contact | 2–10 days |
| Cryptosporidium | 2–14 days | Food or water ingestion, direct and indirect contact | Weeks to months |
| Escherichia coli ETEC | 16–72 h | Food or water ingestion | 3–5 days |
| EPEC | 16–48 h | Food or water ingestion, direct and indirect contact | 5–12 days |
| EHEC | 72–120 h | Food/ingestion, direct or indirect contact | 2–15 days |
| Giardia lamblia | 7–14 days | Food or water ingestion, direct and indirect contact | Weeks to months |
| Norovirus | 24–48 h | Food or water ingestion, direct and indirect contact, aerosol? | 1–2 days |
| Rotavirus | 24–72 h | Direct and indirect contact | 4–6 days |
| Hepatitis A | 30–60 days | Fecal–oral, fomites | 2–4 weeks |
| Salmonella | 16–72 h | Food ingestion, direct and indirect contact | 2–7 days |
| Shigella | 16–72 h | Food or water ingestion, direct and indirect contact | 2–7 days |
| Yersinia enterocolitica | 3–7 days | Food ingestion, direct contact | 1–3 weeks |
Table 22.2.
Concentration of Enteric Pathogens in Feces
| Organism | Per Gram of Feces |
|---|---|
| Protozoan parasites | 106–107 |
| Helminths | |
| Ascaris | 104–105 |
| Enteric viruses | |
| Enteroviruses | 103–107 |
| Rotavirus | 1010 |
| Adenovirus/Norovirus | 1011 |
| Enteric bacteria | |
| Salmonella spp. | 104–1010 |
| Shigella | 105–109 |
| Indicator bacteria | |
| Coliforms | 107–109 |
| Fecal coliforms | 106–109 |
Pathogenic microorganisms usually originate from an infected host (either human or other animal), or directly from the environment. Many human pathogens can only be transmitted by direct or close contact with an infected person or animal. Examples include the herpes virus, Neisseria gonorrhoeae (gonorrhea) and Treponema pallidum (syphilis). This is because their survival time outside the host is very brief. Pathogens transmitted through the environment may survive from hours to years outside the host, depending on the organism and the environment. Pathogens may exit a host in respiratory secretions from the nose and mouth, or be shed on dead skin or in feces, urine, saliva or tears. Thus, they may contaminate the air, water, food, or inanimate objects (fomites) (see Chapter 30). When contaminated air is inhaled or food consumed, the organisms are effectively transmitted to another host, where the infection process begins again. Airborne transmission can occur via release from the host in droplets (i.e., coughing) or through natural (surf at a beach) or human activities (cooling towers, showers) (see Chapter 5). Some organisms may be carried great distances, hundreds of meters or miles (e.g., Legionnaires’ disease and foot-and-mouth disease). Virus transmission by the airborne route may be both direct and indirect. Infection of a host may be by direct inhalation of infectious droplets, or through contact with fomites on which the airborne droplets have settled. Hand or mouth contact with the organism on the surface of a fomite results in the transfer of the organism to the portal of entry, i.e., nose, mouth or eye.
Microorganisms transmitted by the fecal–oral route are usually referred to as enteric pathogens because they infect the gastrointestinal tract. They are characteristically stable in water and food, and, in the case of enteric bacteria, are capable of growth outside the host under the right environmental conditions.
Waterborne diseases (Table 22.3 ) are those transmitted through the ingestion of contaminated water that serves as the passive carrier of the infectious agent. The classic waterborne diseases, cholera and typhoid fever, which have frequently ravaged densely populated areas throughout human history, have been effectively controlled by the protection of water sources and by treatment of contaminated water supplies. In fact, the control of these classic diseases illustrates the importance of water supply treatment which played an important role in the reduction of infectious diseases. Other diseases caused by bacteria, viruses, protozoa and helminths may also be transmitted by contaminated drinking water. However, it is important to remember that waterborne diseases are transmitted by the fecal–oral route, from human to human or animal to human, so that drinking water is only one of several possible sources of infection.
Table 22.3.
Classification of Water-related Illnesses Associated with Microorganisms
| Class | Cause | Example |
|---|---|---|
| Waterborne | Pathogens that originate in fecal material and are transmitted by ingestion | Cholera, typhoid fever |
| Water-washed | Organisms that originate in feces are transmitted through contact because of inadequate sanitation or hygiene | Trachoma |
| Water-based | Organisms that originate in the water or spend part of their life cycle in aquatic animals and come in direct contact with humans in water or by inhalation | Schistosomiasis, Legionellosis |
| Water-related | Microorganisms with life cycles associated with insects that live or breed in water | Yellow fever |
Water-washed diseases are those closely related to poor hygiene and improper sanitation. In this case, the availability of a sufficient quantity of water is generally considered more important than the quality of the water. The lack of water for washing and bathing contributes to diseases that affect the eye and skin, including infectious conjunctivitis and trachoma, as well as to diarrhea illnesses, which are a major cause of infant mortality and morbidity in developing countries. Diarrheal diseases may be directly transmitted through person-to-person contact, or indirectly through contact with contaminated foods and utensils used by persons whose hands are fecally contaminated. When enough water is available for hand washing, the incidence of diarrheal diseases has been shown to decrease dramatically, as has the prevalence of enteric pathogens such as Shigella.
Water-based diseases are caused by pathogens that either spend all (or essential parts) of their lives in water or depend on aquatic organisms for the completion of their life cycles. Examples of such organisms are the parasitic helminth Schistosoma and the bacterium Legionella, which cause schistosomiasis and Legionnaires’ disease, respectively.
Water-related diseases, such as yellow fever, dengue, filariasis, malaria, onchocerciasis and sleeping sickness, are transmitted by insects that breed in water (e.g., mosquitoes that carry malaria) or live near water (e.g., the flies that transmit the filarial infection onchocerciasis). Such insects are known as vectors.
22.2. Bacteria
22.2.1. Salmonella
In the United States, the concept of waterborne and foodborne disease was poorly understood until the late 19th century. During the Civil War (1861–1865), encamped soldiers often disposed of the waste upriver, but drew drinking water from downriver. This practice resulted in widespread dysentery. In fact, dysentery, together with its sister disease, typhoid fever (Salmonella typhi), was the leading cause of death among soldiers of all armies until the 20th century. It was not until the end of the 19th century that this state of affairs began to change. At that time, the germ theory was generally accepted, and steps were taken to treat wastes properly, and protect drinking water and food supplies.
In 1890, more than 30 people out of every 100,000 in the United States died of typhoid. But by 1907, water filtration was becoming common in most U.S. cities, and in 1914 chlorination was introduced. Because of these new practices, the national typhoid death rate in the United States between 1900 and 1928 dropped from 36 to five cases per 100,000 people. The lower death toll was largely the result of a reduced number of outbreaks of waterborne diseases. In Cincinnati, for instance, the yearly typhoid rate of 379 per 100,000 people in the years 1905–1907 decreased to 60 per 100,000 people between 1908 and 1910, following the inception of sedimentation and filtration treatment of drinking water. The introduction of chlorination after 1910 further decreased this rate.
Salmonella is a very large group of rod-shaped, Gram-negative, bacteria comprising more than 2000 known serotypes that are members of the family Enterobacteriaceae. All these serotypes are pathogenic to humans, and can cause a range of symptoms from mild gastroenteritis to severe illness or death. Salmonella are capable of infecting a large variety of both cold- and warm-blooded animals. Typhoid fever, caused by S. typhi, and paratyphoid fever, caused by S. paratyphi, are normally found only in humans, although S. paratyphi is found in domestic animals on rare occasions. In the United States, salmonellosis is due primarily to foodborne transmission, due to bacteria that infect beef and poultry and are subsequently capable of growing in these foods. Salmonellosis is the leading cause of foodborne illness in the United States. Since the route of transmission is fecal–oral, any food or water contaminated with feces may transmit the organism to a new host.
An estimated 94 million cases of gastroenteritis caused by Salmonella species occur globally each year; and of these, nearly 80 million cases are foodborne (Majowicz et al., 2010). In the United States, a little over 1 million cases are estimated to occur annually. All age groups are susceptible, but symptoms are most severe in the elderly, infants and the infirm. The onset time is 12 and 36 hours after ingestion of contaminated food or water. Intestinal disease occurs with the penetration of Salmonella organisms from the gut lumen into the lining of the intestines, where inflammation occurs and an enterotoxin is produced. The immediate symptoms include nausea, vomiting, abdominal cramps, diarrhea, fever and headache. Acute symptoms may last for 1 to 2 days or may be prolonged, again depending on host factors and individual strain characteristics.
S. typhi and S. paratyphi A, B and C produce typhoid and typhoid-like (paratyphoid) fever in humans. Any of the internal organs may be infected. The fatality rate of untreated cases of typhoid fever is 10%, compared to less than 1% for most forms of salmonellosis. Salmonella septicemia (bacteria multiplying in the blood) has been associated with the subsequent infection of virtually every organ system.
Typhoid fever presents a very different clinical picture than salmonellosis. The onset of typhoid fever (1 to 3 weeks) is usually insidious, with fever, headache, anorexia, enlarged spleen, and coughing, and with constipation being more common than diarrhea in adults. Paratyphoid presents a similar clinical picture but tends to be milder. A carrier state may follow infection (<1.0 to 3.9% of the population). A carrier is a person who is permanently infected and may transmit the organism, but does not demonstrate any signs or symptoms of disease. The organism is usually carried in the gallbladder and secreted in the bile.
22.2.2. Escherichia coli and Shigella
Escherichia coli is a Gram-negative rod found in the gastrointestinal tract of all warm-blooded animals, and is usually considered a harmless organism. However, several strains are capable of causing gastroenteritis; among these are the enterotoxigenic (ETEC), enteropathogenic (EPEC), enteroaggregative (EAEC), enteroinvasive (EIEC) and enterohemorrhagic (EHEC) E. coli. They are grouped by their mechanisms of pathogenesis (Figure 22.1 ). All of the E. coli are spread by the fecal–oral route of transmission.
Figure 22.1.
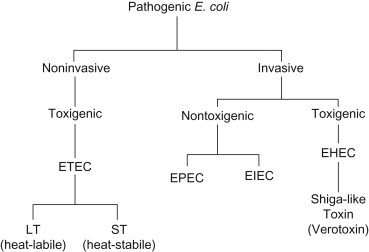
Mechanisms of pathogenic Escherichia coli strains.
The enterotoxigenic E. coli are a major cause of traveler’s diarrhea in persons from industrialized countries who visit less developed countries, and it is also an important cause of diarrhea in infants and children in less developed countries (Table 22.4 ). Following an incubation period of 10–72 hours, symptoms including cramping, vomiting, diarrhea (may be profuse), prostration and dehydration occur. The illness usually lasts less than 3 to 5 days. Disease is caused by two toxins, one the heat-labile toxin and the other the heat-stable toxin. The ETEC E. coli are usually species specific; that is, humans are the reservoir for the strains causing diarrhea in humans. Only a few outbreaks have been documented in the United States. Conservative estimates suggest that each year, approximately 210 million cases and 380,000 deaths occur (Wenneras and Erling, 2004). EPEC is the oldest recognized category of diarrhea-causing E. coli. Diarrheal disease caused by this group of E. coli is virtually confined to infants less than 1 year of age. It is associated with infant summer diarrhea, outbreaks of diarrhea in nurseries and community epidemics of infant diarrhea. Symptoms include watery diarrhea with mucus, fever and dehydration. The diarrhea can be severe and prolonged with a high fatality rate (as high as 50%). Since the 1960s, EPEC has largely disappeared as an important cause of infant diarrhea in North America and Europe. However, it remains a major agent of infant diarrhea in South America, Africa and Asia. Humans, cattle and pigs can be infected by this organism. Thus, foods implicated in outbreaks are raw beef and chicken.
Table 22.4.
Summary of Pathogenic E. coli Incidence and Epidemiology
| Pathogenic E. Coli | Site of Infection | Associated Disease | Incidence | Target Population | Significant Transmission Route |
|---|---|---|---|---|---|
| ETEC | Small intestine | Travelers’ diarrhea, chronic childhood diarrhea (in developing countries) | 16 U.S. outbreaks (1996–2003); prevalence 1.4% in patients with diarrhea; 79,420 cases of travelers’ diarrhea each year (in the U.S.) | International travelers and children in developing countries | Food (raw produce, street vendors) and water |
| EPEC | Small intestine | Infant diarrhea | Hundreds of thousands of deaths worldwide | Children in developing countries | Water, infant formula |
| EHEC | Large intestine | Hemorrhagic colitis, HUS | 110,000 cases and 61 deaths annually in the U.S. | All ages | Food (beef, produce), person-to-person, water, animals |
| EIEC | Large intestine | Dysentery | Low in developed countries | Children in developing countries | Water (rare), person-to-person |
| EAEC | Intestine | Watery diarrhea with or without blood in the stool, acute and chronic | Developed and developing countries | Children and adults, travelers | Food, water, person-to-person |
The disease caused by EIEC closely resembles that caused by Shigella. Illness begins with severe abdominal cramps, watery stools and fever. Disease is usually self-limiting with no known complications. Any age group is susceptible. This type of E. coli carries a plasmid that enables it to invade cells lining the gastrointestinal tract, resulting in a mild form of dysentery. EIEC infections are endemic in less developed countries, although occasional cases and outbreaks are reported in industrialized countries. Outbreaks also have been associated with hamburgers and unpasteurized milk.
The EHEC E. coli (also referred to as Shigella toxin producing E. coli or STEC) were described in 1982 when a multistate epidemic of hemorrhagic colitis occurred in the United States, and was shown to be due to a specific serotype known as E. coli O157: H7. This organism produces two toxins called verotoxins I and II. These toxins are closely related to or identical to the toxin produced by Shigella dysenteriae. The toxin production depends on the presence of a prophage. A prophage is a bacterial virus that inserts its own DNA into a bacterial chromosome. A plasmid codes for a novel type of fimbria (hairlike projection) that enabled the organism to adhere to the intestinal lining and initiate disease.
Enteroaggregative Escherichia coli (EAEC), an increasingly recognized cause of diarrhea in children in developing countries, has been particularly associated with persistent diarrhea (more than 14 days), a major cause of illness and death. Recent outbreaks implicate EAEC as a cause of foodborne illness in industrialized countries.
EHEC infections are now recognized to be an important problem in North America, Europe and some areas of South America. The illness usually includes severe cramping and diarrhea, which is initially watery but becomes grossly bloody. The illness is usually self-limiting and lasts for an average of 8 days. However, some victims, particularly the very young, develop hemolytic–uremic syndrome (HUS) resulting in renal failure and hemolytic anemia (Figure 22.2 ). This disease can result in permanent loss of kidney function. In older individuals, HUS plus two other symptoms, fever and neurologic symptoms, constitutes thrombotic thrombocytopenic purpura (TTP). This illness can have a mortality rate in the elderly as high as 50%. Most outbreaks are associated with undercooked or raw hamburger. Raw milk, unpasteurized fruit juices and vegetables contaminated with cow dung have also been implicated. Infected cattle are believed to be a major environmental source of this organism. Waterborne outbreaks involving both nondisinfected groundwater and recreational waters have also occurred. Various studies have shown that E. coli O157:H7 is commonly present in domestic sewage at concentrations from 10 to 100 colony forming units (CFU) per 100 ml, and in wastewater from slaughterhouses from 100 to 1000 CFU/100 ml (Nwachuku and Gerba, 2008).
Figure 22.2.
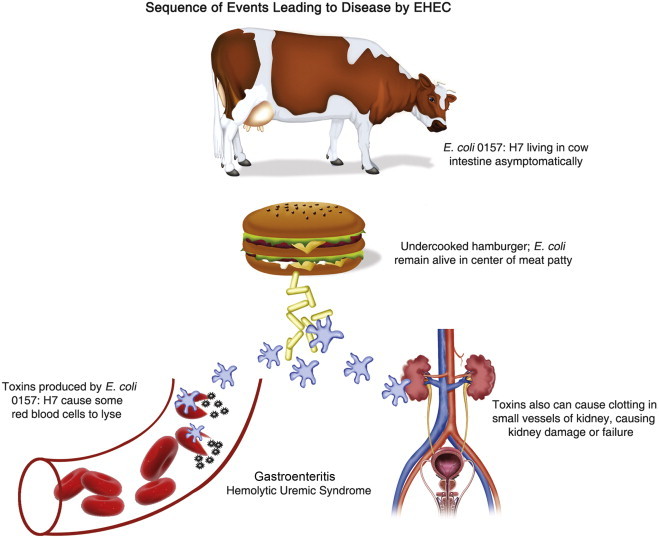
Sequence of events leading to disease by EHEC.
Shigella is closely related to E. coli. Four species have been described: S. dysenteriae, S. flexneri, S. boydii and S. sonnei. S. dysenteriae causes the most severe disease, and S. sonnei causes the mildest symptoms. S. sonnei is the serotype most often found in the United States (Lee et al., 1991). Shigella very rarely occurs in animals. It is principally a disease of humans and occasionally of other primates such as monkeys and chimpanzees. The organism is often found in water polluted with human sewage, and is transmitted by the fecal–oral route. Shigella has been a common cause of waterborne recreational outbreaks in the United States. However, most cases of shigellosis are the result of person-to-erson transmission through the fecal–oral route. Secondary attack rates are 20–40% of household contacts. An estimated 440,000 cases of shigellosis occur annually in the United States (Gupta et al., 2004). Shigella is associated with certain foods such as salads, raw vegetables, milk and dairy products, and poultry. After an onset time of 12 hours to 1 week, symptoms of abdominal pain, cramps, diarrhea and vomiting may occur. The organisms multiply in the cells of the gastrointestinal tract and spread to neighboring cells, resulting in tissue destruction. Some infections are associated with ulceration, rectal bleeding and drastic dehydration, with fatality rates as high as 10–15%. Infants, the elderly and the infirm are most susceptible.
22.2.3. Campylobacter
Campylobacter jejuni is a Gram-negative curved rod. It is relatively fragile, and sensitive to environmental stress (including an oxygen content of 21%, drying, heating, contact with disinfectants or acidic conditions). Before 1972, when methods were developed for its isolation from feces, it was believed to be primarily an animal pathogen causing abortion and enteritis in sheep and cattle. Recent surveys have shown that C. jejuni is a leading cause of bacterial diarrheal illness in the United States, with an estimated 2 million cases per year (Samuel et al., 2004). Although C. jejuni is not carried by healthy individuals in the United States or Europe, it is often isolated from healthy cattle, chickens, birds and even flies. It is sometimes present in nonchlorinated water sources such as streams and ponds.
C. jejuni infections cause diarrhea with fever, abdominal pain, nausea, headache and muscle pain. The illness usually occurs 2–5 days after the ingestion of the contaminated food or water. Illness generally lasts 7–10 days, and relapses are not uncommon. Although anyone can be infected with C. jejuni, children under 5 years of age and young adults (15–29 years old) are more frequently afflicted than other age groups.
Surveys show that 20 to 100% of retail chickens are contaminated with C. jejuni. This is not surprising because many healthy chickens carry these bacteria in their intestinal tracts. Nonchlorinated water may also be a source of infection. However, properly cooking chicken, pasteurizing milk and chlorinating water will kill the bacteria (Blaser et al., 1986).
C. jejuni has been isolated from 22% of coastal and estuary water samples in concentrations ranging from 10 to 230 per 100 ml, and from 28% of river samples in concentrations of 10 to 36 cells per 100 ml. Carter et al. (1987) found Campylobacter in 10–44% of pond water samples. However, it is thought that virtually all surface waters contain Campylobacter. Recovery rates from surface waters are highest in the fall and winter months, and lowest during the spring and summer months. Campylobacter density does not show a significant correlation with the isolation of indicator bacteria such as fecal or total coliforms (Carter et al., 1987).
This bacterium dies off rapidly in stream water at 37°C, showing a 9-log decrease within 3–12 days (Rollins and Colwell, 1986). However, the organism can remain viable for extended periods at cooler temperatures, surviving over 120 days at 4°C in stream water.
22.2.4. Yersinia
Yersinia enterocolitica and Y. pseudotuberculosis are small, rod-shaped, Gram-negative bacteria. Y. enterocolitica causes diarrhea often with a bloody stool, while Y. pseudotuberculosis causes fever and abdominal pain, usually on the right side. Yersinia enterocolitica is a ubiquitous microorganism, with the majority of isolates being recovered from asymptomatic carriers, infected animals, contaminated food and untreated water. Most are nonpathogenic having no clinical importance (Rahman et al., 2011). Only a few serotypes have been associated with illness. Pigs have been shown to be a major reservoir of pathogenic Y. enterocolitica involved in human infections, particularly for strains of serotype 4/O:3. Y. enterocoltica has been largely associated with foodborne outbreaks, but waterborne outbreaks have also occurred. Yersinia pseudotuberculosis also appears to occur in water; animal- and food-associated infections occur primarily in the northern hemisphere, and carrots and iceberg lettuce have been implicated in outbreaks.
Symptoms usually begin 24 to 48 hours after ingestion of contaminated food or drink. Yersiniosis is frequently characterized by such symptoms as gastroenteritis with diarrhea and/or vomiting. However, fever and abdominal pain are the hallmark symptoms. Yersinia infections mimic appendicitis, but the bacteria may also cause infections of other sites such as wounds, joints and the urinary tract. Overall, yersiniosis is more common in Northern Europe, Scandinavia and Japan than in the United States. The greatest incidence of disease is seen during cold seasons.
22.2.5. Vibrio
Dr. John Snow (1813–1858) of London was one of the first to make a connection between certain infectious diseases and drinking water contaminated with sewage. In his famous study of London’s Broad Street pump, published in 1854, he noted that people afflicted with cholera were clustered in a single area around the Broad Street pump, which he determined was the source of the infection. When, at his insistence, city officials removed the handle of the pump, Broad Street residents were forced to obtain their water elsewhere. Subsequently, the cholera epidemic in that area subsided.
The Gram-negative genus Vibrio contains more than one member that is pathogenic to humans. The most famous member of the genus is still V. cholerae. Cholera is transmitted through the ingestion of fecally contaminated food and water. Cholera remains prevalent in many parts of Central America, South America, Asia and Africa.
V. cholerae serogroup Ol includes two biovars, cholerae (classical) and El Tor, each of which includes organisms of the Inaba and Ogawa serotypes. A similar enterotoxin is elaborated by each of these organisms, so the clinical pictures are similar. Asymptomatic infection is much more common than disease, but mild cases of diarrhea are also common. In severe untreated cases, death may occur within a few hours and the fatality rate without treatment may exceed 50%. This is due to a profuse watery diarrhea referred to as rice-water stools. The rice-water appearance is due to the shedding of intestinal mucosa and epithelial cells. With proper treatment, the fatality rate is below 1%. Humans are the only known natural host. Thus, the reservoir for V. cholerae is human, although environmental reservoirs may exist, apparently in association with copepods or other phytoplankton.
Vibrios that are biochemically indistinguishable, but do not agglutinate in V. cholerae serogroup Ol antiserum, were formerly known as nonagglutinable vibrios (NAGs) or noncholera vibrios (NCVs). They are now included in the species V. cholerae. Some of these strains produce enterotoxin but most do not. Thus reporting of non-Ol V. cholerae infections as cholera is inaccurate and leads to confusion. V. mimicus is a closely related species that can cause diarrhea, some strains of which produce an enterotoxin indistinguishable from that produced by V. cholerae.
The following example illustrates the possible devastating impacts of this disease. A cholera epidemic caused by V. cholerae Ol began in January 1991 in Peru, and spread to Central America and South America. A total of 1,041,422 cases occurred with 9642 deaths (MMWR, 1995). The epidemic was believed to have been initiated by inadequate chlorination of drinking water. A second example is an outbreak in southern Asia, an epidemic caused by the newly recognized strain V. cholerae O139 which began in late 1992 and continued to spread to at least 11 countries. This latter strain of V. cholerae also produces severe watery diarrhea and dehydration that is indistinguishable from the illness caused by V. cholerae Ol.
Vibrio cholerae is a native marine organism, and its potential for transmission to humans is related to a complex ecology that controls its occurrence and concentration in the marine food chain (Lipp et al., 2002). Warmer temperatures in combination with elevated pH and plankton blooms can influence V. cholerae attachment, growth and multiplication in the aquatic environment, particularly in association with copepods (Figure 22.3 ). Thus, factors such as climate change, climate variability (El Niño-Southern Oscillation) and monsoons can influence the V. cholerae concentration in marine waters and exposure via the marine food chain to humans (see also Chapter 31).
Figure 22.3.
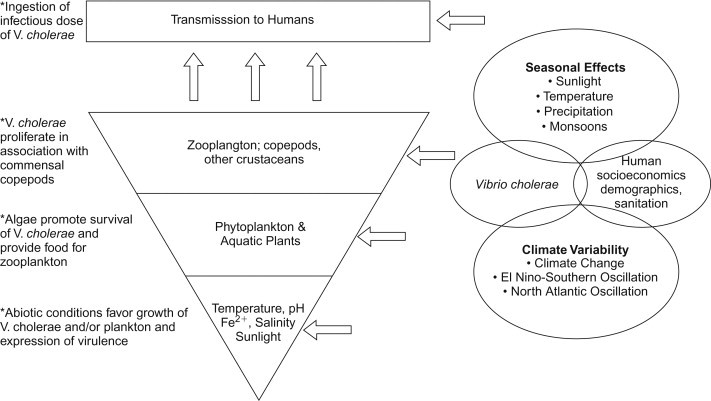
Hierarchical model for environmental cholera transmission (Lipp et al., 2002).
Another species, V. parahaemolyticus, is usually transmitted by contaminated food. The ingestion of inadequately cooked seafood, any food cross-contaminated by handling raw seafood or food rinsed with contaminated seawater may transmit this disease. A period of time at room temperature is generally necessary to allow multiplication of the organisms. Disease symptoms include watery diarrhea and abdominal cramps in the majority of cases, sometimes with nausea, vomiting, fever and headache. Usually, the disease is self-limiting and lasts 1 to 7 days. Most cases are reported during the warmer months, and marine coastal environments are a natural habitat. The organisms have been found in marine silt, in coastal waters and in fish and shellfish.
Of all foodborne infectious diseases, infection with V. vulnificus is one of the most severe; the fatality rate for V. vulnificus septicemia exceeds 50% (Tacket et al., 1984). Cases are most commonly reported during warm-weather months (April–November), and are often associated with eating raw oysters. Many patients are found to have had a preexisting liver disease, usually associated with alcohol use or viral hepatitis. All of these latter patients had eaten raw oysters 1–2 days before the onset of symptoms. These symptoms include thrombocytopenia, bulbous skin lesions, hypotension and shock. Cases have also been reported that arose through the contamination of a wound by seawater or seafood drippings.
22.2.6. Helicobacter
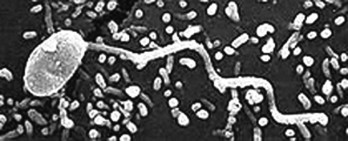
In 1982, a physician in Australia cultured a Gram-negative, spiral-shaped bacterium observed in biopsied tissue of a stomach ulcer. Initially called Campylobacter pylori based on biochemical and morphological characteristics, the organism is now named Helicobacter pylori (Figure 22.4 ). The stomach mucosa contains cells that secrete proteolytic enzymes and hydrochloric acid. Other specialized cells produce a layer of mucus that protects the stomach itself from digestion. If this mucous layer is disrupted, the ensuing inflammation leads to an ulcer. H. pylori has been shown to bind to O− blood group antigens on the gastric epithelial cells. People of this blood group are twice as likely to develop gastric ulcers. H. pylori produces large amounts of urease, an enzyme that converts urea to the alkaline product ammonia. This results in a high pH in the local region.
Figure 22.4.
Helicobacter pylori from Goai Nobel, 2005.
Humans are thought to be a natural host for H. pylori. Person-to-person spread is probably the dominant source of transmission, although controversy exists over whether the fecal–oral or oral–oral route of transmission predominates. H. pylori has been isolated from feces (Thomas et al., 1992) and detected in saliva (Nguyen et al., 1993). Infection may also be spread by ingesting contaminated food or water. Several studies have shown a greater risk of acquiring H. pylori infections from drinking untreated well water (Klein et al., 1991, Rolle-Kampczyk et al., 2004, Reavis, 2005). A recent study found a possible outbreak associated with using a public drinking water supply (Melius et al., 2012). Nonculturable H. pylori organisms have been detected in river water and groundwater in the United States (Hegarty et al., 1999).
22.2.7. Legionella
Legionella is the causative agent of Legionnaires’ disease and Pontiac fever. Both syndromes are characterized initially by anorexia, malaise, myalgia and headache. Within 24 hours, a fever ensues with chills. A nonproductive cough may occur and abdominal pain and diarrhea are seen in many patients. In Legionnaires’ disease, chest radiographs show areas of consolidation indicative of pneumonia and, indeed, respiratory failure may occur. This disease has a 15% fatality rate in hospitalized cases. Pontiac fever is not associated with pneumonia or death. Patients recover spontaneously in 2–5 days.
Legionella are poorly staining Gram-negative bacilli that require cysteine and other specific nutrients when grown on artificial laboratory media. At least 40 species of Legionella have been shown to cause disease, but the most prominent pathogenic species is L. pneumophila, which first received extensive attention after an outbreak in 1976 in Philadelphia. The disease is found worldwide, with most sporadic cases occurring during the summer and fall months. The reservoir is primarily aquatic. Hot water systems, air-conditioning cooling towers and evaporative condensers have all been implicated in outbreaks, as have decorative fountain and retail store misters (Figure 22.5 ). The organism has also been found in creeks and ponds, and the soil from their banks. The bacterium survives for months in tap and distilled water. The primary route of transmission is thought to be through the inhalation of aerosols. Exposure is not uncommon, as reflected in serologic assays that show that 1–20% of the general population have antibodies to L. pneumophila.
Figure 22.5.
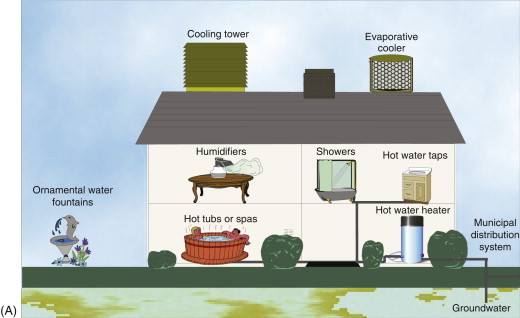

(A) Sources of Legionella in the environment. (B) Cooling towers. Outbreaks of Legionnaires’ disease have been commonly traced to cooling towers.
Concentrations of 1.4×104 to 1.7×105 cells per liter of Legionella spp. have been detected in raw drinking water sources using direct fluorescent antibody (DFA) techniques (Tison and Seidler, 1983). The concentrations of Legionella found in distribution water samples by DFA were as follows (CFU/L): chlorinated water, <8×103 to 1.4×104; water treated by slow sand filtration and chlorination, <5.4×103 to 4.6×104; water treated by flocculation, filtration and chlorination, <8×103 to 2.2×104. Zacheus and Martikainen (1994) found that 30% of hot water systems in apartment buildings contained Legionella. The mean number of L. pneumophila was 2.7×103 CFU/L with a range of <50 to 3.2×105 CFU/L. For all positive hot water systems, Legionella was also isolated from the hot water tap and shower head. Legionella pneumophila was isolated from 12.5, 29.0 and 37.5% of the hot water distribution systems receiving chlorinated groundwater, unchlorinated groundwater and chlorinated surface water, respectively.
A great deal of work has been carried out on the survival and growth of legionellae in potable water distribution systems and plumbing in hospitals and homes. Legionellae appear to be more resistant to chlorine than E. coli. For example, Legionella has been shown to exist in potable water systems even when exposed to 0.75 to 1.5 ppm free chlorine residual. Such protection is afforded by intracellular growth in protozoa such as Tetrahymena pyriformis and Acanthamoeba castellani (Fields et al., 1984, Moffat and Tompkins, 1992).
Legionella survives well at 50°C, and environmental isolates are able to grow in tap water at temperatures as high as 42°C (Figure 22.6 ). The enhanced survival and growth in these systems have been linked to stagnation stimulated by rubber fittings in the plumbing system (Colbourne et al., 1988), and trace concentrations of metals such as iron, zinc and potassium (States et al., 1989). Sediment promotes the growth of Legionella, as does stagnation in the hot water tank. Legionella can be removed from hot water heaters by raising the temperature to over 60°C near the heating element and to over 50°C at outlets, combined with regular flushing (Meenhorst et al., 1985).
Figure 22.6.
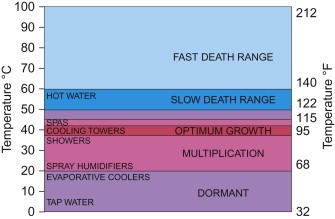
The effect of temperature on the growth of Legionella pneumophila.
The overall attack rate of pneumonia in the United States is 12–15 cases per 1000 persons per year, resulting in approximately 3,957,000 cases annually. Pneumonia is the sixth leading cause of death in the United States, with an estimated annual cost of $23 billion. L. pneumophila causes 4.1 to 20.1% of community-acquired cases, many of which result in hospitalization (Marrie, 1994). These data suggest that L. pneumophila is a major cause of serious cases of pneumonia.
22.2.8. Opportunistic Bacterial Pathogens
An opportunistic pathogen is one that usually causes disease only in those whose immune system is compromised. The weakened immune system may be due to very young or old age, pregnancy, cancer therapy, immunosuppressive drugs, human immunodeficiency virus (HIV) and other causes. Opportunistic pathogens are numerous in the environment. Many opportunistic bacterial pathogens are found in surface and drinking waters.
Concern has been generated in the drinking water industry regarding the health effects of heterotrophic bacteria that are found in tap water, bottled water and other sources of potable water. Heterotrophic bacteria are those that require organic carbon rather than carbon dioxide as a carbon source. All human bacterial pathogens are heterotrophic.
Most of the heterotrophic bacteria in drinking water are not human pathogens. However, some of the genera, including Legionella, Mycobacterium, Pseudomonas, Acinetobacter, Stenotrophomonas and Aeromonas, include species that are opportunistic pathogens. See Section 22.2.7 for a discussion of the genus Legionella.
The most important opportunistic pathogen in the genus Pseudomonas is P. aeruginosa, which is primarily a nosocomial (hospital-acquired) pathogen responsible for 10% of nosocomial infections. Pseudomonas spp. infects the lungs, urinary tract, eye, burn patients and wounds. Human disease is often associated with water-related reservoirs such as swimming pools, whirl-pools, hot tubs, taps (faucets) and contact lens solutions. In hospitals, growth of P. aeruginosa in the drinking water taps appears to be the major source infecting patients (Asghari et al., 2013). The source of community-acquired infections of cystic fibrosis patients appears to be largely from environmental exposure (Ranganathan et al., 2013). It is important as the ultimate cause of death in these patients is often lung infections by P. aeruginosa. Although P. aeruginosa is found in drinking water, it is not ubiquitous. Results of surveys showed that 2% of bottled water and 2–3% of tap water samples contain P. aeruginosa at concentrations between 1 and 2300 organisms/ml (Allen and Geldreich, 1975).
Acinetobacter is the second most frequently isolated, nonfermentative, Gram-negative rod in the clinical laboratory. However, it is generally considered to be of low virulence. Up to 25% of healthy adults carry this organism in the respiratory tract. The human skin is the likely source for most outbreaks of hospital infections, but sinks and taps have also been implicated in some outbreaks. As an opportunistic pathogen, Acinetobacter is involved in nosocomial urinary tract infections, bacteremia, wound infections and pneumonia. Acinetobacter can also be a cause of community-acquired pneumonia and urinary tract infections. Acinetobacter has been isolated from 97% of natural surface water samples with a concentration of 0.1–100 cells/ml (Baumann, 1968). Bifulco et al. (1989) isolated Acinetobacter from 38% of groundwater supplies at an arithmetic mean density of 8 CFU/100 ml in the positive samples.
Acinetobacter has been isolated from 5 to 92% of distribution water samples (LeChevallier et al., 1980, Bifulco et al., 1989). It comprised 1.0 to 5.5% of the heterotrophic plate count (HPC) flora in drinking water samples (Payment et al., 1988) with concentrations of 6–21 CFU/ml. It has also been found in 5 to 35% of bottled water samples, at concentrations of 2–30 CFU/ml.
Pseudomonas maltophilia has been reclassified as Stenotrophomonas maltophilia and is the third most commonly isolated nonfermentative Gram-negative rod in clinical laboratories. This organism can colonize the body and cause disease. Risk factors for infection include stays in intensive care units, mechanical ventilation, antibiotic treatment and cancer. Diseases it can cause include septicemia, pneumonia, wound infections and more rarely meningitis and endocarditis.
S. maltophilia constituted 5.7% of the HPC found in raw surface water samples and 0–1.2% of the flora in distribution water samples (LeChevallier et al., 1980). Using PCR, S. maltophilia was detected in 3.7 to 36.4% of unchlorinated drinking water samples, derived from surface waters in the Netherlands (Van der Wielen and van der Kooij, 2013). The bacterium has also been found in 2% of bottled water samples at concentrations of 2–22 CFU/ml and in <5% of cooler water samples at 34 CFU/ml.
Aeromonas hydrophila, A. sobria and A. caviae are very similar and were all referred to as A. hydrophila until 1976, when they were first split into separate species. Papers written before 1985 may use the term A. hydrophila including all three species and biochemical variants. The three Gram-negative species are biochemically very similar, and have all been implicated as diarrheal agents in humans. The exact mechanism for diarrhea has not been elucidated.
A strong association has been found between drinking untreated water and the occurrence of diarrhea with the isolation of Aeromonas spp. Higher counts of A. hydrophila in distribution water correlate with greater frequency of diarrheal isolates; however, no waterborne outbreaks have been documented.
Children and the elderly tend to be affected most often. As with many diarrheal agents, outbreaks of diarrhea, in which Aeromonas has been implicated, have been associated with day care centers. Aeromonas is frequently found in environmental water samples. It has been recovered from 0.6 to 18.2% of natural freshwater samples at a concentration range of 0.1–3600 CFU/ml. It has also been recovered from 0.9 to 27% of distribution water samples at an average concentration of 0.022 CFU/ml. However, even at large oral doses of up to 1010 CFU, A. hydrophila failed to produce diarrhea in human volunteers (Morgan et al., 1985).
Mycobacteria are rod-shaped bacteria that contain high levels of lipid (waxy) material. They are among the most resistant nonspore-forming bacteria to chlorine and other common disinfectants. The Mycobacterium avium complex (MAC) consists of at least 28 serovars of two distinct species, M. avium and M. intracellulare. It also included three serovars of M. scrofulaceum in the past, but the inclusion is no longer appropriate due to recent advances in mycobacterial systematics. Pulmonary disease caused by MAC has dramatically increased in the United States over the last three decades (Kendall and Winthrop, 2013). The disease is most common among individuals over 60 years of age. Predisposing factors include age, chronic lung disease, bronchogenic carcinoma, previous gastrectomy and AIDS. MAC can also cause pulmonary disease, osteomyelitis and septic arthritis in people with no known predisposing factors (Jones et al., 1995). However, disease by MAC can be lethal and is difficult to treat because of resistance to many antimycobacterial agents.
It is believed that municipal drinking water systems are an important reservoir for MAC since it is among the most common bacteria identified in biofilms in chlorinated drinking water distribution systems and fixtures (i.e., showerheads). Epidemiological investigations have associated water sources with infections by atypical mycobacteria (Burns et al., 1991). These bacteria can multiply in water that is oligotrophic (Falcao et al., 1993) or essentially free of nutrients, including dialysis water, and they are relatively resistant to disinfection by chlorination and chloramines (Collins et al., 1984). Atypical mycobacteria are widespread in water environments. Mycobacteria have been isolated from 11 to 38% of raw water samples at concentrations of <0.1–48 CFU/ml. MAC has also been found in up to 50% of municipal and private drinking water samples at concentrations of 0.01–5.2 CFU/ml.
Hospital water systems often harbor MAC and may be a source of nosocomial infection. M. avium has primarily been associated with hot water systems, and in some cases, a hot water system may be persistently colonized by the same strain of M. avium (Whiley et al., 2012).
22.2.9. Blue–Green Algae
Blue–green algae or cyanobacteria occur in an enormous diversity of habitats, freshwater and marine, as plankton (free floating), mats and periphyton (attached to surfaces). Hot spring mats of some Oscillatoria develop up to temperatures of 62°C (Figure 22.7 ). They have many beneficial functions such as nitrogen fixation and cycling of nutrients in the food chain.
Figure 22.7.
Oscillatoria—a blue–green algae.
Courtesy Michael Clayton (2006).
Despite their beneficial roles in the environment, cyanobacteria sometimes become problematic. Occasionally, they increase rapidly resulting in cyanobacterial blooms (Figure 22.8 ). Blooms are associated with eutrophic water, especially with levels of total phosphorus >0.01 mg/L and levels of ammonia- or nitrate-nitrogen >0.1 mg/L. Optimal temperatures for blooms are 15–30°C, and optimal pH is 6–9. Calm or mild wind conditions sometimes allow blooms to cover the water surface, but the highest concentrations of cyanobacteria may occur at depths ranging from 2 to 9 m, which will not be visible from the shore. The offending bacteria may also grow in the sediment. These blooms can impart an off-taste and odor to the water, and/or result in the production of toxins.
Figure 22.8.

Cyanobacterial bloom.
Courtesy C.P. Gerba.
The most common complaints related to such blooms are of taste and odor. Geosmin and 2-methylisoborneol (MIB) can produce odors at levels as low as 1.3–10 and 6.3–29 ng/L, respectively (Young et al., 1996). The odor produced by geosmin is described as earthy, and that of MIB as musty or camphorous smelling. Concentrations of MIB and geosmin are usually highest in summer and fall. Several compounds produced by cyanobacteria can cause off-tastes and odors as shown in Table 22.5 .
Table 22.5.
Cyanobacterial Compounds Producing Off-tastes and Odors
| Compound | Odor | Taste |
|---|---|---|
| Geosmin | Earthy, musty, grassy | Musty, earthy, stale |
| MIB | Musty, earthy, peaty | Musty, earthy, stale |
| Isobutylmethoxypyrazine | Woody, stale, musty | Creosote, stale, dusty |
| Isopropylmethoxypyrazine | Sooty, dusty, cabbage | Musty, vegetable water |
| Octa-1,3-diene | Musty | |
| Hexanal | Green apple-like | |
| Octan-1-ol | Rancid | |
| β-Cyclocitral | Tobacco |
Geosmin is produced by several cyanobacteria including Oscillatoria, Anabaena, Lyngbya, Phormidium, Symploca (Narayan and Nunez, 1974), Aphanizominon and Fischerella (Wu and Juttner, 1988). Lyngbya, Oscillatoria and Phormidium (Izaguirre, 1992) are the most common genera producing MIB. Some strains of Diplocystis and Schizothrix can also cause off-tastes and odors. Microcystis release some odorous sulfur compounds, especially when they decay.
Many cyanobacteria found in algal blooms can produce toxins that cause liver damage, neural damage and gastrointestinal (GI) disturbances. This has been well documented in many wild animal and livestock cases and implicated in human cases as well. Microcystis is the number one offender worldwide. Other toxin-producing genera include Anabaena, Aphanizomenon, Alexandrium, Cylindrospermopsis, Nodularia, Nostoc and Oscillatoria (Turner et al., 1990). Different types of toxins produced are shown in Table 22.6, Table 22.7 . Most toxic species are associated with temperate rather than tropical climates.
Table 22.6.
Cyanobacterial and Types of Toxins Produced
| Genus | Toxins Produced |
|---|---|
| Anabaena | Anatoxin-a, hepatotoxins |
| Aphanizomenon | Saxitoxin, neosaxitoxin, hepatotoxins |
| Alexandrium | Saxitoxin |
| Cylindrospermopsis | Hepatotoxin |
| Nodularia | Nodularins |
| Oscillatoria | Neurotoxins, hepatotoxins |
| Microcystis | Microcystins |
Table 22.7.
Characterization of Cyanobacterial Toxins
| Toxin | Characterization of Toxin |
|---|---|
| Anatoxins | Neurotoxins |
| Microcystins | Hepatotoxins |
| Nodularins | Hepatotoxins |
| Saxitoxins | Neurotoxins |
In livestock and wild animals, the hepatotoxins cause weakness, anorexia and liver damage. They can be lethal within minutes to a few days. Neurotoxins can cause twitching, muscle contraction, convulsions and death. Signs and symptoms in humans associated with the ingestion of water with algal blooms are dizziness, headaches, muscle cramps, nausea, vomiting, gastroenteritis and pneumonia (Phillip et al., 1992). Long-term exposure to toxins is associated with liver cancer as well (Carmichael, 1994).
22.3. Parasitology
The study of parasitology embodies a large diversity of eukaryotic organisms. This group includes organisms that are unicellular, multicellular and multinucleate; aerobic and anaerobic; motile and nonmotile; sexual and asexual. For this chapter parasites will be grouped into two categories: protozoa and helminthes (Table 22.8 ). Protozoa are unicellular microorganisms in the Kingdom Protista, and are classified according to their means of locomotion: flagella, cilia, pseudopodia or no locomotion. Some are parasitic, although the majority are nonparasitic. Some undergo a sexual stage, whereas others reproduce by asexual means: fission, budding or schizogony. Helminths belong to Kingdom Animalia, and include roundworms, flatworms, tapeworms and flukes. They are multicellular complex organisms containing organs and tissue. They usually develop in soil or in intermediate hosts to complete their life cycle and have elaborate life cycles, including larva, eggs and adult stages. Parasitic protozoa and helminths can be acquired from the environment—water, soil and contaminated food—and have had great health and economic impacts in many developing and developed countries. In the United States, there are two protozoan genera that are especially of concern: Giardia and Cryptosporidium. They form hardy cysts and oocysts that can survive water treatment disinfection and are one of the biggest concerns of water utilities today. Some of the characteristics of environmentally transmitted parasites are summarized in Table 22.9 .
Table 22.8.
Classification of some Environmentally Transmitted Protozoa and Helminthes
| Protozoa |
| Phylum Apicomplexa |
| Cyclospora cayetanensis |
| Cryptosporidium parvum |
| Toxoplasma gondii |
| Phylum Microspora |
| Enterocytozoon bieneusi |
| Encephalitozoon cuniculi |
| Encephalitozoon hellem |
| Encephalitozoon intestinalis |
| Phylum Sarcomastigophora |
| Entamoeba histolytica |
| Giardia lamblia |
| Naegleria fowleri |
| Helminthes |
| Phylum Nematoda |
| Ascaris lumbricoides |
| Necator americanus |
| Trichuris trichiura |
| Phylum Platyhelminthes |
| Class Cestoidea |
| Taenia saginata |
| Class Trematoda |
| Schistosoma mansoni |
Table 22.9.
Some Characteristics of Environmentally Transmitted Parasites
| Organism | Infective Form (Size) | Mechanism of Transmission | Distribution | Reservoirs |
|---|---|---|---|---|
| Giardia lamblia | Cyst (8–16 μm) | Person–person waterborne, foodborne | Worldwide | Humans, beavers, muskrats, voles |
| Cryptosporidium parvum | Oocyst (3–6 μm) | Person–person waterborne, foodborne | Worldwide | Many vertebrates, especially cattle |
| Entamoeba histolytica | Cyst (10–16 m) | Person–person waterborne, foodborne | Areas of poor sanitation | Usually humans (potentially pigs, primates, and dogs) |
| Naegleria fowleri | Trophozoite (10–15 μm) | Trophozoite swims up nasal cavity | Worldwide | None: free-living in aquatic or soil environment |
| Cylospora cayetanensis | Sporulated oocyst (8–10 μm) | Waterborne, foodborne | Asia, Caribbean, Mexico, and Peru | None known |
| Enterocytozoon bieneusi | Spore 1–1.6 μm | Fecal–oral | Not known | None known |
| Encephalitozoon hellem | Spore 1–2.5 μm | Urine–oral | Not known | None known |
| Encephalitozoon cuniculi | Spore 1–2.5 μm | Fecal/urine–oral | Not known | Laboratory rabbits, rodents, dogs |
| Encephalitozoon intestinalis | Spore 1–2 µm | Fecal/urine–oral | Not known | None known |
| Toxoplasma gondii | Sporulated (11–12 μm) | Oral ingestion from soil or litterbox (oocyst), undercooked meats (oocyst), undercooked meats (tissue cysts), Waterborne | Worldwide | Cats (definitive host), humans, sheep, goats, pigs, cattle, birds (intermediate hosts) |
| Ascaris lumbricoides | Embryonated egg | Oral from soil contact | Worldwide | Humans |
| Trichuris trichiura | Embryonated egg | Waterborne, foodborne | Worldwide especially tropics | Humans |
| Necator americanus | Filariform larva | Skin penetration | Tropical Africa, Asia, Central and South America and Caribbean | Humans |
| Ancylostoma duodenale | Filariform larva | Skin penetration, oral from soil contact | Tropical Africa, Asia, Central and South America and Caribbean | Humans |
| Taenia saginata | Cysticercus | Ingestion of undercooked beef containing cysticerci, waterborne | Worldwide | Cattle (intermediate host) |
| Schistosoma mansoni | Cercaria | Penetrate skin | Arabia, Africa, South America, and Caribbean | Humans, snail (intermediate host) |
22.3.1. Protozoa
22.3.1.1. Giardia lamblia
Giardia lamblia was first described in 1681 by Antonie van Leeuwenhoek, who found them in his own feces. He called the trophozoites “animalcules.” In 1859, Vilern Lambl rediscovered Giardia by finding the trophozoites in stools of young children with diarrhea. It was not until the early 20th century that physicians began associating diarrhea with the presence of Giardia in stools. In 1954, Robert Rendtorff confirmed infectivity in human volunteers with oral administration of Giardia cysts.
Giardia is the most frequently identified intestinal parasite in the United States (Schaefer, 2006). Worldwide it is estimated that 13% of adults and up to 50% of children are infected with Giardia (Schaefer, 2006). In the United States, it is one of the most frequent identified causes of waterborne disease (Craun et al., 2010).
Humans become infected with G. lamblia by ingesting the environmentally resistant stage, the cyst (Figure 22.9 ). Once ingested, it passes through the stomach and into the upper intestine. The increase in acidity via passage through the stomach stimulates the cyst to excyst, which releases two trophozoites into the upper intestine. The trophozoites (Figure 22.10 ) attach to the epithelial cells of the small intestine. It is believed that the trophozoites use their sucking disks to adhere to epithelial cells. The adherence to the cells flattens the villi, causing malabsorption and diarrhea by not allowing adsorption of water and nutrients across the intestine. This can cause both acute and chronic diarrhea within 1–4 weeks of ingestion of cysts resulting in foul-smelling, loose and greasy stools. Once the trophozoites detach from the epithelial cells and travel down the intestine, cholesterol starvation is believed to stimulate the trophozoites to encyst and pass back into the environment as a cyst. Giardiasis can be treated with metronidazole (Flagyl) or nitazoxanide. Although in some cases the symptoms will spontaneously disappear without treatment, in most cases without treatment the symptoms will wax and wane for many months. In symptomatic patients, more trophozoites than cysts are excreted into the feces and cannot withstand the harshness outside the human body. In asymptomatic humans, mostly cysts are passed in stools; therefore Giardia carriers can serve as a source of cysts in the environment.
Figure 22.9.
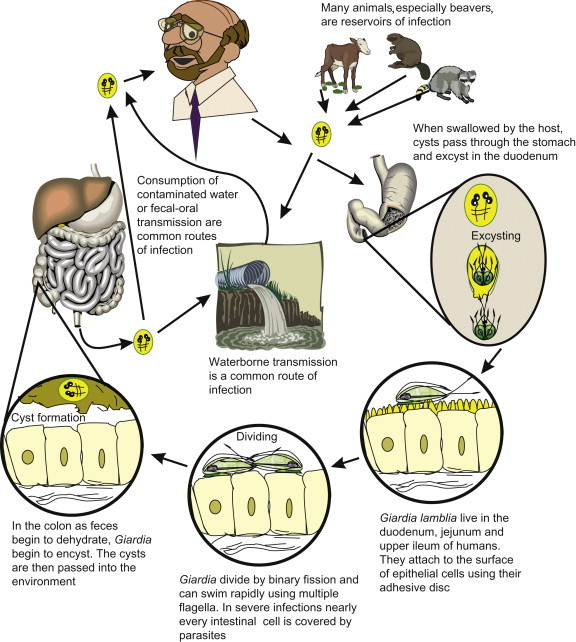
Life cycle of Giardia lamblia.
Figure 22.10.
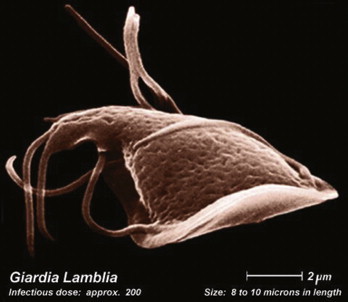
Trophozite of Giardia lamblia, the reproductive stage of the waterborne parasite.
Photo courtesy E.A. Myer.
When Giardia cysts enter the environment they can survive for prolonged periods. G. lamblia cysts have been documented to survive for up to 77 days at 8°C and 4 days at 37°C in distilled water (Bingham et al., 1979). In another study, cysts of Giardia muris (a species that infects mice but is often used as a model for G. lamblia) were suspended in lake and river water, and found to survive 28 days in lake water at a depth of 15 feet at 19.2°C. At a 30-foot depth at a temperature of 6.6°C, the cysts remained viable for 56 days.
It is still controversial whether animals such as beavers produce a G. lamblia strain that can infect humans. Studies have shown that beavers can be infected with G. lamblia from humans, but the reverse has not been demonstrated (Erlandsen et al., 1988). In one study, 40 to 45% of beavers in Colorado were found to be infected with Giardia and shedding up to 1×108 cysts/animal/day, making them a major source of Giardia in the environment (Hibler and Hancock, 1990). Other animals that may contribute to Giardia numbers in the environment are muskrats, where 95% of the population is infected. Various other animals found to be infected are cattle, goats, sheep, pigs, cats and dogs (Erlandsen, 1995). To date, no infections with G. lamblia in humans have been directly linked to an animal host, but there is evidence that suggests that animal-source Giardia could potentially infect humans. Studies based upon isoenzyme analysis and pulsed field gel electrophoresis (PFGE) banding patterns did not find a difference between cysts from beaver hosts and human hosts (Isaac-Renton et al., 1993).
22.3.1.2. Cryptosporidium
Cryptosporidium was first described by Tyzzer in 1907, when he identified the organism in the intestinal epithelium of a mouse. It was not identified as a human pathogen until 1976, when it was described in the stools of immunocompromised hosts. Since that time, there have been several waterborne outbreaks, the most notable being the Milwaukee outbreak in April 1993, which infected over 400,000 people (MacKenzie et al., 1994) and killed more than 50 (Case Study 22.1 ).
Case Study 22.1. Cryptosporidiosis in Milwaukee.
Early in the spring of 1993, heavy rains flooded the rich agricultural plains of Wisconsin. These rains produced an abnormal runoff into a river that drains into Lake Michigan, from which the city of Milwaukee obtains its drinking water. The city’s water treatment plant seemed able to handle the extra load: it had never failed before, and all existing water quality standards for drinking water were properly met. Nevertheless, by April 1, thousands of Milwaukee residents came down with acute watery diarrhea, often accompanied by abdominal cramping, nausea, vomiting, and fever. In a short period of time, more than 400,000 people developed gastroenteritis, and more than 100—mostly elderly and infirm individuals—ultimately died, despite the best efforts of modern medical care. Finally, after much testing, it was discovered that Cryptosporidium oocysts were present in the finished drinking water after treatment. These findings pointed to the water supply as the likely source of infection; and on the evening of April 7, the city put out an urgent advisory for residents to boil their water. This measure effectively ended the outbreak. All told, direct costs and costs associated with loss of life are believed to have exceeded $150 million.
The Milwaukee episode was the largest waterborne outbreak of disease ever documented in the United States. But what happened? How could such a massive outbreak occur in a modern U.S. city in the 1990s? And how could so many people die? Apparently, high concentrations of suspended matter and oocysts in the raw water resulted in failure of the water treatment process—a failure in which Cryptosporidium oocysts passed right through the filtration system in one of the city’s water treatment plants, thereby affecting a large segment of the population. And among this general population were some whose systems could not withstand the resulting illness. In immunocompetent people, Cryptosporidiosis is a self-limiting illness; it is very uncomfortable, but it goes away of its own accord. However, in the immunocompromised, Cryptosporidiosis can be unrelenting and fatal.
It is now recognized that there are several species of Cryptosporidium capable of infecting humans. The most common species infecting humans is C. hominis, which infects only humans, and C. parvum, which primarily infects cattle, but also humans. Cryptosporidium hominis has a complex life cycle involving both sexual and asexual stages (Figure 22.11 ). The host ingests sporulated oocysts (ranging from 3 to 6 μm in diameter) from contaminated water, food or direct contact (Figure 22.12 ). In the small intestine, the oocyst excysts, releasing four sporozoites, which attach to the epithelial cells of the mucosa. The sporozoite becomes enveloped by the microvilli, which fuse and elongate to cover the sporozoite. The sporozoite then matures into a trophozoite and into a schizont. The schizont, an asexual reproductive form in which multiple mitosis occurs followed by cytokinesis, results in eight first-generation merozoites. The cell ruptures, releasing the merozoites, which then infect neighboring epithelial cells, and schizogony occurs again, but forming only four second-generation merozoites. When the cell ruptures, the merozoites attach to uninfected epithelial cells and form either macrogametocytes or microgametocytes. The macrogametocytes and microgametocytes further divide, and form macrogametes and microgametes, respectively. They join, forming a zygote, which differentiates to form the unsporulated oocyst, which is then expelled from the cell surface, sporulates, and is shed in the host’s feces.
Figure 22.11.
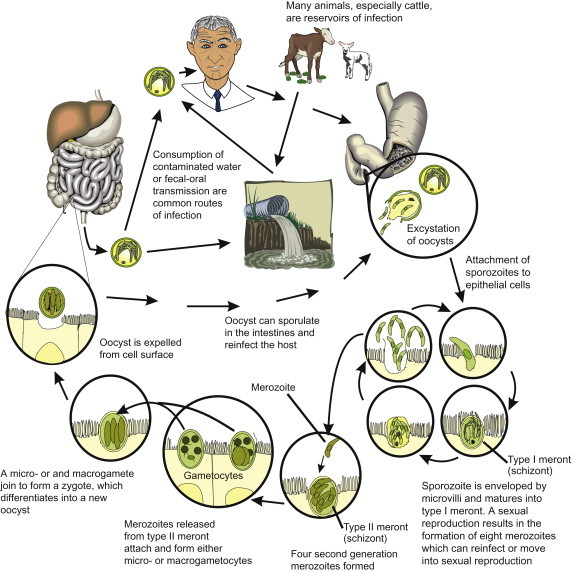
Life cycle of Cryptosporidium hominis.
Figure 22.12.
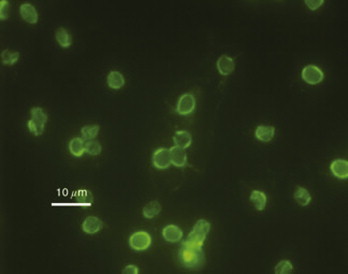
Cryptosporidium parvum oocysts stained with fluorescein isothiocyanate-labeled antibodies.
Photo courtesy H.D.A. Lindquest, U.S. Environmental Protection Agency.
Within 3–10 days after ingestion of oocysts, nonbloody, voluminous, watery diarrhea begins and lasts for 10–14 days in most immunocompetent hosts. In normal healthy individuals the disease is usually self-limiting. Nitazoxanide has been approved for treatment of diarrhea caused by Cryptosporidium in people with healthy immune systems. In the immunocompromised host (e.g., AIDS patients), the disease can be life threatening. The prevalence of cryptosporidiosis in the United States and Europe is 1 to 3% (Sterling and Marshall, 2006). The oocysts are very infectious and the presence of low numbers of oocysts in water or food poses a health threat (Messner et al., 2001).
Cryptosporidium oocysts can enter the environment via human and animal wastes. Cryptosporidiosis has been reported in many domestic animals, especially cattle. An infected calf can excrete 1010 oocysts per day. In a study by Kemp et al. (1995), farm drains were found to contain 0.06 to 19.4 oocysts per liter. This can result in agricultural land runoff that can contaminate surface water.
Cryptosporidium forms an extremely hardy oocyst that survives chlorine disinfection as commonly practiced at conventional water treatment plants. It is now the most commonly identified cause of recreational outbreaks in the United States, largely associated with chlorinated swimming pools (Hlavsa et al., 2011). It has also been found to survive for weeks in surface waters (Johnson et al., 1997).
22.3.1.3. Entamoeba histolytica
Entamoeba histolytica was discovered in 1873 by D.F. Lösch in St. Petersburg, Russia, although its life cycle was not determined until Dobell did so in 1928. It causes amoebic dysentery (bloody diarrhea), and is the third most common cause of parasitic death in the world. The world prevalence exceeds 500 million infections with more than 40,000 deaths each year. The protozoan only infects humans. There are two sizes of cysts, small (5–9 μm) and large (10–20 μm), with each cyst producing eight trophozoites in the host. Only the larger cyst has been associated with disease; the smaller cyst tends to be associated with a commensal lifestyle (the organism benefits from the host, while the host is unaffected). About 2–8% of people infected develop invasive amoebic dysentery in which the trophozoites actively invade the intestinal wall, bloodstream and liver. It is unknown why this occurs. This organism is generally a problem in developing countries where sanitation is substandard, and is transmitted via contaminated food and water. Humans are the main reservoir, although pigs, monkeys and dogs have also been found to serve as reservoirs. No waterborne outbreaks have occurred in the United States since 1984. Entamoeba is not as resistant to disinfectants as Giardia and Cryptosporidium, and is easily removed during water treatment.
22.3.1.4. Naegleria fowleri
Naegleria fowleri is an amoeboflagellate, changing between a cyst, amoeba and flagellate with the amoeba stage dominant. The free-living protozoa are ubiquitous and found throughout the world in fresh waters (John, 1982). Cysts are usually present in low numbers, but when the water temperature exceeds 35°C (hot springs and warm stagnant waters), the amoeba transforms to the flagellated form quite rapidly, which enables the microorganism to swim. Infections are usually associated with children swimming in natural springs or warm waters, although they are rare in the United States. There have been four cases associated with drinking water in the United States: in Arizona and Louisiana. The flagellate swims into the nose of a host and sheds its flagella (or it may be forced into the nose via diving). The amoeba then follows the nerves to the brain, producing a toxin that liquefies the brain. The organisms do not form cysts in the host. Primary amoebic meningoencephalitis (PAM) develops, causing severe, massive headaches. Death usually follows 4–6 days later. Diagnosis is most frequently postmortem upon brain examination. Treatment (amphotericin B) is available if the diagnosis is made quickly enough, although permanent brain damage may already have occurred.
22.3.1.5. Cyclospora sp.
Cyclospora cayetanensis is an emerging waterborne and foodborne protozoan pathogen. It was first identified in the intestine of a mole in 1870 by Eimer. It was not recognized as a human pathogen until the early 1980s, and then it was believed to be a cyanobacterium (Soave and Johnson, 1995). It is a coccidian protozoan in the phylum Apicomplexa. It produces a round spherical oocyst (Figure 22.13 ) measuring 8×10 μm which contains two sporocysts, each containing two sporozoites. Its sporulation life cycle is typical of a coccidian protozoan parasite. Desiccation of the organism kills it, and therefore it must be in an aquatic environment during maturation.
Figure 22.13.
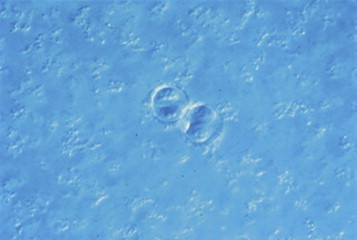
Oocyst of Cyclospora cayotanensis; 100×under DIC.
Photo courtesy H. Smith.
Cyclosporiasis causes voluminous, explosive, watery, nonbloody diarrhea in addition to abdominal cramps, nausea and fatigue. Illness associated with Cyclospora averages 43 days in the immunocompetent host, which can be compared to cryptosporidiosis, which has a duration of only 10 to 14 days (Ortega et al., 1993). It can be treated with Bactrim (trimethoprim–sulfamethoxazole), which appears to eliminate the parasite (Knight, 1995).
Acquisition of Cyclospora is not completely understood, as the host range and reservoirs are not known. However, water and food are believed to be a major route of infection, because for the organism to be infectious it must mature (sporulate) in the environment for 2 weeks (Ortega et al., 1993). Water can potentially become contaminated with sporulated oocysts, but most infections have been associated with contaminated produce. Outbreaks related to imported contaminated raspberries, snow peas, basil and mesclun lettuce have been implicated in various outbreaks of cyclosporiasis in the United States (CDC, 2004). Several suspected waterborne outbreaks have been documented (Rebold et al., 1994, Huang et al., 1995, Baldursson and Karanis, 2011). Infections have also been associated with individuals who live in or visit the Caribbean Islands, Central America and South America, Southeast Asia and Eastern Europe (Knight, 1995).
22.3.1.6. Microsporidia
Microsporidia, the nontaxonomic name to describe organisms belonging to the phylum Microspora, were first described in 1857, when Nägeli identified Nosema bombycis, a microsporidian responsible for destruction of the silkworm industry. To date, over 1000 species of microsporidia infecting insects, invertebrates and all five phyla of vertebrate hosts have been described. Microsporidia are for the most part considered to be opportunistic pathogens in humans. There were only a handful of documented cases before the advent of the AIDS epidemic. Since then there have been hundreds of documented cases in immunocompromised patients. However, there have also been cases documented among the immunocompetent. Five genera have been associated with the majority of human infections: Enterocytozoon bieneusi, Encephalitozoon hellem, Encephalitozoon cuniculi, Encephalitozoon intestinalis, Pleistophora spp. and Nosema corneum. The first four have the potential to be waterborne because they are shed in feces and urine. E. bieneusi, E. hellem and E. intestinalis are the most common cause of microsporidian infections in patients with AIDS (Curry and Canning, 1993). In addition, they are much smaller (0.8×1.5 μm depending on species) than other parasites, and potentially more difficult to remove by water treatment filtration.
The microsporidian spore has the potential of being transmitted by water. The life cycle of microsporidia contains three stages: the environmentally resistant spore, merogony and sporogony. The spore is ingested by a host or possibly inhaled in some cases. Once in the body, it infects cells and goes through merogony followed by sporogony, which results in production of resistant infective spores (Figure 22.14 ). The spores are then shed via bodily fluids such as urine and excreta. Once in the environment they have a strong potential to enter water sources. E. intestinalis spores have been identified in sewage, surface and ground waters, supporting the notion of environmental transmission. The spores are highly resistant to heat inactivation and drying. Waller (1979) found that E. cuniculi survived 98 days at 4°C and 6 days at 22°C.
Figure 22.14.
Microsporidium spore with tube by which it penetrates host cells.
Courtesy Centers for Disease Control.
Enterocytozoon bieneusi, Encephalitozoon hellem, Encephalitozoon cuniculi and Encephalitozoon intestinalis cause a variety of illnesses. E. bieneusi causes diarrhea and wasting disease. It is the most important cause of microsporidiosis in AIDS patients. Several surveys have determined that 7 to 30% of AIDS patients who have unexplained chronic diarrhea are infected with E. bieneusi (Weber et al., 1994). E. intestinalis is similar to E. bieneusi in that it infects the intestines and causes diarrhea, but it can also infect kidneys and bronchial and nasal cells. It can infect macrophages, which allows it to disseminate throughout the body. It is secreted in feces and urine, which supports the notion of water transmission. E. cuniculi is not an intestinal parasite but it can be shed in urine (Zeman and Baskin, 1985), and therefore environmental transmission is a possibility. It has also been described infecting many different mammals, which means that there could be many animal reservoirs that can contaminate the environment. E. hellem has been recognized and shown to cause eye infections (keratoconjunctivitis) and disseminated infections such as ureteritis and pneumonia. It does not invade the intestine, but it can be shed in the urine (Schwartz et al., 1994). Both water- and foodborne outbreaks have been documented (Cotte et al., 1999, Decraene et al., 2012).
22.3.1.7. Toxoplasma gondii
Toxoplasma gondii, the causative agent of toxoplasmosis, is an intestinal coccidium of felines with a very wide range of intermediate hosts. T. gondii causes a wide range of clinical conditions including brain damage in children, lymphadenopathy, ocular disease and encephalitis. In the immunocompetent adult it causes symptoms that could be mistaken for influenza. It is estimated that 13% of the world’s population is infected with T. gondii (Hughes, 1985). Acquisition of the organism can be through contact with infected undercooked meat containing a bradyzoite (tissue cyst), and through contact with the environmental stage, the oocysts. The oocyst is excreted only in cat feces (domestic and wild) (Figure 22.15 ). Congenital infection occurs when a pregnant woman becomes infected for the first time. Once infected, tachyzoites can cross the placenta and infect the fetus. Of the fetuses that become infected, 15% have severe complications. The oocysts can persist in soil and water, which can serve as a route of transmission. One study in England found that 100% of wild cats had antibodies to T. gondii (McOrist et al., 1991). Various studies of cats in the United States have found a seropositive prevalence of approximately 40% (Dubey and Beattie, 1988). The high prevalence of T. gondii in the cat population demonstrates that felines can be a significant source of oocysts in the environment. The oocyst can survive 18 months in the soil (−20 to 33°C) in Kansas (Frenkel et al., 1975) and over 410 days in water (Yilmaz and Hopkins, 1972).
Figure 22.15.
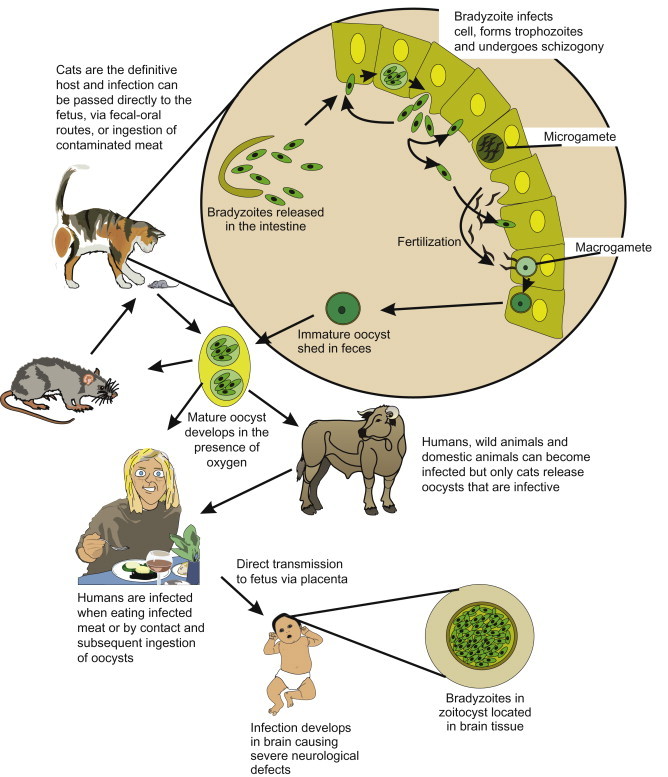
Life cycle of Toxoplasma gondii.
There have been several drinking water outbreaks of toxoplasmosis, and water may be a more important route than previously thought (Baldursson and Karanis, 2011, Karanis et al., 2013). Several epidemiological studies have associated increase risk of infection with the consumption of unfiltered drinking water (Jones et al., 1995). T. gondii caused a waterborne outbreak in British Colombia, Canada, where more than 110 people including 12 newborns were infected after exposure to an unfiltered water supply (Mullens, 1996). In a later study of the area, four of seven domestic cats found in the watershed had antibodies to T. gondii, potentially being the initial source of contamination (Stephen et al., 1996).
The oocysts are very resistant to inactivation by iodine, chlorine and ozone (Wainwright et al., 2007). The cysts can be inactivated in meat by either freezing at −12°C or cooking to an internal temperature of 67°C (Dubey, 1996).
22.3.2. Nematodes
Nematodes are nonsegmented roundworms belonging to the phylum Nematoda. The majority of roundworms are free living in soil and fresh and salt water. The typical nematode has a flexible outer cuticle that protects the worm. Nematodes move via a muscular system and most lay eggs.
22.3.2.1. Ascaris lumbricoides
Ascaris lumbricoides is probably the most prevalent parasitic infection, with over 1 billion humans affected or about 22% of the world’s population (CDC, 2013). The prevalence is quite high in some regions of the world. Infection percentages range from 40 to 98% in Africa, to 73% in Southeast Asia, and to 45% in Central America and from South America, and 1 to 2% in the United States (Freedman, 1992).
Infection in humans occurs by ingestion of embryonated eggs (Figure 22.16, Figure 22.17 ). There are no known animal reservoirs. The eggs are swallowed and the larvae hatch in the small intestine. The larvae develop into second-stage larvae, which penetrate the lumen and enter into the bloodstream and capillaries. They travel via pulmonary circulation to the liver and heart, where the larvae develop into third-stage organisms that can lodge in the alveolar space (Figure 22.18 ). This migration through the lungs can cause pneumonitis (Loeffler’s syndrome). The immature worms leave the lungs and travel through the bronchi, trachea and epiglottis. They are then swallowed and arrive in the small intestine. There they undergo two molts, mate and produce eggs. There is medical treatment against the adult intestinal worm: mebendazole or pyrantel pamoate. Symptoms usually correspond to the worm load, and a heavy worm load can lead to intestinal blockage. The adult worm can reach more than 30 cm in size. Although most infections are mild, more than 20,000 people die annually with complications caused by intestinal blockage (Freedman, 1992).
Figure 22.16.
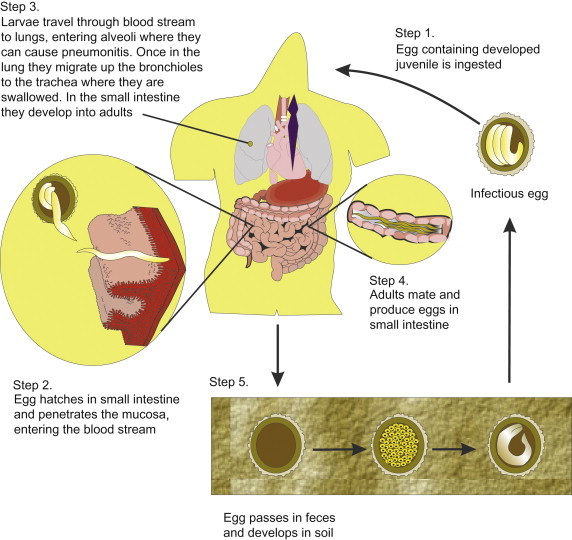
Life cycle of Ascaris lumbricoides.
Figure 22.17.
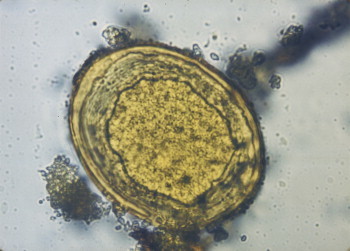
Ascaris lumbricoides ovum.
Photo courtesy P. Watt.
Figure 22.18.

Ascaris lumbricoides adult.
Photo courtesy Centers for Disease Control and Prevention.
An adult worm can produce more than 200,000 eggs per day. In 2–4 weeks after deposition in soil, they embryonate if the soil conditions are suitable (humid and warm) and are infectious. The eggs can survive months before embryonation if soil conditions are not appropriate. The eggs can survive freezing, chemicals, disinfectants and sewage treatment. Because of their large size (35×55 μm), they accumulate in sewage sludge. This is especially a concern with sewage sludge-amended soils. In one study, 31–53% of eggs that had been deposited in soil were still viable 10 years later (Brudastov et al., 1971).
22.3.2.2. Trichuris trichiura
Trichuris trichiura is a worm that measures about 30–50 mm in length and is referred to as a whipworm, as the worm’s shape resembles a whip. It has been estimated that there are over 1 billion infections worldwide (de Silva et al., 2003), and it is the third most common nematode infection. It is common in the southeastern United States as the weather conditions are ideal for egg survival in the soil (Figure 22.19 ). The egg must be deposited in the soil and requires 21 days in moist, shady, warm soil to embryonate. In one study, 20% of ova deposited in soil were viable after 18 months (Burden et al., 1976). Infection occurs in humans via ingestion of contaminated water or soil. The worms can survive for years in a host, causing disease symptoms of diarrhea or constipation, anemia, inflamed appendix, vomiting, flatulence and insomnia. The infection is diagnosed by identification of worms or eggs in the stool. The infection can be treated successfully with mebendazole. To prevent transmission, education on hand washing and sanitary feces disposal is necessary.
Figure 22.19.
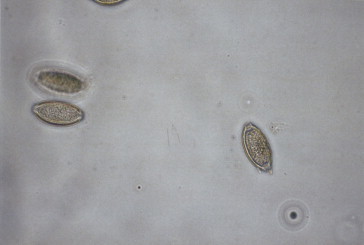
Trichuris trichiura eggs.
22.3.2.3. Necator americanus and Ancylostoma duodenale
There are two major hookworm species that infect humans: Necator americanus (New World hookworm) and Ancylostoma duodenale (Old World hookworm). The species are differentiated by mouth parts of adults and body size. There are no known reservoirs. They inhabit the small intestine and feed on intestinal mucosa and blood. They secrete an anticoagulant, causing great blood loss and anemia. They are the leading cause of iron deficiency in the tropics.
N. americanus have round cutting teeth and are 7–10 mm in length. They lay an average of 10,000 eggs a day, and enter humans via skin penetration. Each worm consumes 0.03 ml of blood per day. A. duodenale are larger (10–12 mm in length) and cause even greater blood loss. Each worm consumes 0.26 ml of blood per day. A. duodenale also lays more eggs (28,000), and are orally infective as well as able to penetrate the skin. In addition, A. duodenale also have sharp cutting teeth.
The eggs embryonate once they are passed into the small intestine (Figure 22.20 ). The eggs further develop into the rhabditiform larvae within 48 hours in warm, moist sandy or loamy soil. The larvae feed, grow and molt twice and then transform to filariform larvae. The filariforms do not eat. They seek out the highest point in the surroundings (e.g., top of grass blade) waiting for a host. Upon contact with skin, they penetrate the tissue and pass through a hair follicle or cut. They burrow through the subcutaneous tissue, and then through the capillaries to the lungs. In the lungs, they break out of the alveolar capillaries, and migrate up the bronchi and trachea, where they are swallowed and enter the stomach and small intestine. They can live an average of 5 years but have been found to survive up to 15 years. The larvae can survive up to 6 weeks in moist, shady sandy or loamy soil. They do not survive well in clay soil, dry conditions or at temperatures below freezing or greater than 45°C.
Figure 22.20.
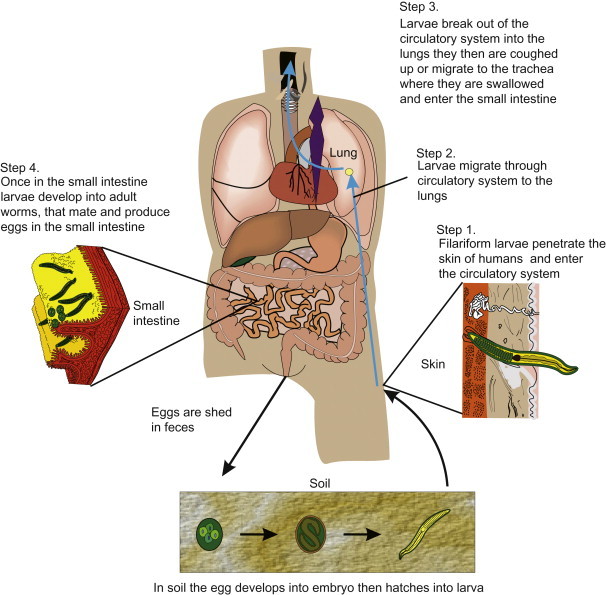
Life cycle of Necator americanus.
22.3.3. Cestodes (Taenia saginata)
Cestodes are tapeworms consisting of a flat segmented body and a scolex (head) containing hooks and/or suckers and grooves for attachment. The segments are called proglottids and pregnant segments are called gravids. The adults are parasitic and live in the intestinal lumen of many vertebrates.
Taenia saginata is transmitted by infected beef products and is the most common tapeworm found in humans. It is present in every country where beef is consumed (Figure 22.21 ). Cattle become infected from eating grass or soil contaminated with human waste containing gravid proglottids. This occurs in areas where night soil (human waste) is used as fertilizer. The proglottids can survive in the environment for weeks. One study found that they could survive 71 days in liquid manure, 16 days in untreated sewage and up to 159 days on grass (Jepsen and Roth, 1952). The eggs hatch in the duodenum, and hexacanths (tapeworm embryos containing six pairs of hooklets) are released. The hexacanths penetrate the mucosa, enter the intestine and travel throughout the body. They then enter muscle and form a cysticercus (larval tapeworm enclosed in a cyst), which becomes infective in a few months. Humans become infected when they eat undercooked beef containing such cysticerci. The cysticerci can be inactivated at 56°C or by freezing at −5°C for 1 week (Schmidt and Roberts, 1989). Within 2–12 weeks the worm begins shedding gravid proglottids. The average worm length is 10–15 feet. Symptoms of infection are abdominal pain, headache, nausea, diarrhea, intestinal blockage and loss of appetite (contrary to the belief that tape worms infections cause an increase in appetite). The infection can be diagnosed by examining a gravid proglottid or scolex. The best methods for prevention are sanitation (proper disposal of human wastes) and thorough cooking of beef.
Figure 22.21.
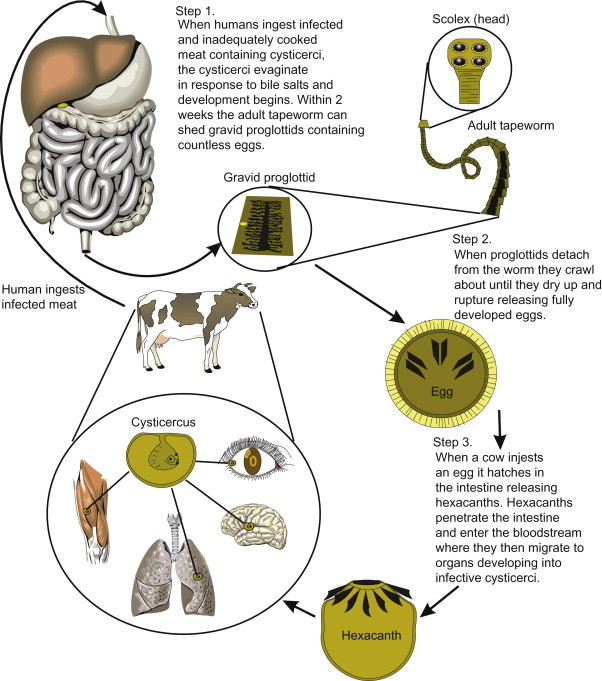
Life cycle of Taenia saginata.
22.3.4. Trematodes (Schistosoma mansoni)
Trematodes, or flukes, are bilaterally symmetric worms that have two deep suckers and flame cells (for excretion). The suckers are used for both attachment and locomotion. The life cycles are complex, with trematodes being either hermaphrodites (adults have both female and male gonads) or schistosomes (separate sexes). Trematodes require an intermediate host (snail) to complete their life cycle; the human is the definitive host, excreting eggs in the feces.
Three species of Schistosoma are medically important. In the past, S. japonicum and S. haematobium were the main causes of schistosomiasis, but now S. mansoni is recognized to be the most widespread of the three. The genus Schistosoma is responsible for more than 200 million infections worldwide and causes up to 200,000 deaths annually (Hopkins, 1992). More than 400,000 of those infected live in the United States (West and Olds, 1992). However, all these cases are imported as the intermediate host, the freshwater snail (Biomphalaria sp.), is not present in the United States. S. mansoni is distributed through most of Africa and the Middle East but is also found in parts of Central America and South America as well as some Caribbean Islands (Savioli et al., 1997).
The larvae of schistosomes of bird and mammals can penetrate human skin, causing what is known as “swimmer’s itch.” These schistosomes do not mature in humans. This occurs in the Great Lakes region and along the coast of California.
The life cycle of S. mansoni is complicated, and human infection begins with the cercariae penetrating human skin. In addition, each parasitic stage involves different organs and thus different medical symptoms. The cercariae from infected snails are found in freshwater bodies. Once the cercariae penetrate the skin, they transform into the schistosomula, the first parasitic stage. This causes the disease cercarial dermatitis, which consists in immediate or delayed hypersensitivity to the penetration. After 1–2 days of living in the subcutaneous tissues, the schistomsomula migrate through the lungs to the liver. The organisms mature through this migration, developing into male or female adult worms. The adult worms then migrate to the circulatory system and to other organs. Each pair of adult worms live an average of 3–10 years and potentially up to 30 years (Lucey and Maguire, 1993). They also produce 300 eggs a day, only a percentage of which reach the lumen to be passed in the feces. Many of the eggs become trapped in the small blood vessels and tissues, causing granulomas and intestinal, visceral, liver and fibroobstructive diseases. The acute form of disease, Katayama fever, can occur in previously uninfected persons 2–6 weeks after exposure to cercariae-contaminated water. Chemotherapy is available using praziquantel, although no vaccine is available.
No decrease of infection has been noted, and in fact the number of cases has increased over the past 50 years. The spread of this parasite has occurred as water development projects and population movements have introduced it into previously uninfected regions. However, it has been controlled in some areas: the Caribbean, excluding Puerto Rico, and Brazil. Strategies that have been successful are chemotherapy, health education, water supply treatment and sanitation. Very little success has been achieved by eradicating the intermediate host, the snail, with either mulluscicides or predator fish.
22.4. Viruses
22.4.1. Enteric Viruses
Viruses are a leading cause of gastroenteritis, in particular in infants and young children, in which they are a major cause of mortality worldwide. Four major groups of human gastroenteritis viruses have been identified: rotavirus; enteric adenovirus; caliciviruses (norovirus and sapporovirus); and astrovirus (Table 22.10 ). Of these, norovirus is of note because it has become the enteric virus most commonly associated with water- and foodborne illness worldwide. Although endemic viral gastroenteritis can be transmitted person to person by the oral–fecal route, outbreaks of viral gastroenteritis may be triggered by contamination of a common water or food source.
Table 22.10.
Human Enteric Viruses
| Enterovirus |
| Hepatitis A virus |
| Reoviruses |
| Rotaviruses |
| Adenoviruses |
| Astroviruses |
| Torovirus |
| Human caliciviruses (norovirus, sapporovirus) |
| Hepatitis E virus |
| Picobirnaviruses |
| Bocaviruses |
| Coronaviruses |
Many human viruses can infect the gastrointestinal tract and be excreted in the feces into the environment. It has been estimated that an individual with an enteric viral infection may excrete 1011 viral particles per gram of feces. Once in the environment, viruses can reach water supplies, recreational waters, crops and shellfish, through contact with sewage, land runoff, solid waste landfills and septic tanks.
Diseases caused by enteric viruses range from trivial to severe or even fatal. Waterborne outbreaks caused by enteric viruses are difficult to document because many infections by these agents are subclinical; i.e., the virus may replicate in an individual, resulting in virus shedding but without signs of overt disease. Therefore, an individual with waterborne infection but without overt disease may infect others, who in turn may become ill, spreading the infection throughout the community. In addition, epidemiological techniques lack the sensitivity to detect low-level transmission of viruses through water. Recreational activities in swimming pools have sometimes resulted in waterborne outbreaks caused by norovirus, hepatitis A virus, coxsackievirus, echovirus and adenoviruses. Enteric viruses from infected individuals may contaminate recreational waters by direct contact or by fecal release.
22.4.1.1. Astroviruses
Astroviruses were first observed by electron microscopy in diarrheal stools in 1975. These agents are icosahedral viruses with a starlike appearance and with a diameter of approximately 28 nm. Astroviruses have a single-stranded RNA (ssRNA) genome. The sequencing of the astrovirus genome allowed the establishment of its own virus family the Astroviridae, with Mamastrovirus which infects mammals and Avastrovirus which infects birds. Serological assays and sequence analysis have resulted in the identification of eight distinct serotypes which infect humans.
Astrovirus type 1 seems to be the most prevalent strain in children. Type 4 has been associated with severe gastroenteritis in young adults. Astrovirus-like particles have been found in the feces of a number of animals suffering from a mild self-limiting diarrheal infection, but no antigenic cross-reactivity has been found between these agents and human astroviruses. Astrovirus infections occur throughout the year, with a peak during the winter–spring seasons in temperate zones.
Astroviruses cause a mild gastroenteritis after an incubation period of 3 to 4 days. Overt disease is common in 1- to 3-year-old children. However, adults and young children are also affected. Outbreaks of astrovirus infection have been associated with oysters and drinking water (Kurtz and Lee, 1987, Maunula et al., 2004).
22.4.1.2. Adenoviruses
Adenoviruses are double-stranded DNA (dsDNA) icosahedral viruses approximately 70 nm in diameter with protruding spikes called pentons (Figure 22.22 ). At least 57 human adenovirus types have been identified. Although there are many avian and mammalian adenovirus types, they are species specific. Adenoviruses can replicate in the respiratory tract, the eye mucosa, the intestinal tract, the urinary bladder and the liver.
Figure 22.22.
Adenovirus.
Most adenovirus human illness is associated only with one-third of adenovirus types. Although many adenovirus infections are subclinical, these viruses may cause: acute respiratory disease (types 1–7, 14 and 21); conjunctivitis (types 3, 7, 8, 11, 14, 19 and 37); acute hemorrhagic cystitis (11 and 21); acute respiratory disease (ARD) of military recruits (types 3, 4, 7, 14 and 21); and gastroenteritis (types 31, 40, 41 and 52) (Table 22.11 ). Adenovirus type 36 has been associated with obesity in humans and animals (Atkinson et al., 2005).
Table 22.11.
Classification of Human Adenoviruses
| Subgenus | Serotype | Human Illness |
|---|---|---|
| A | 12 | Meningoencephalitis |
| 18, 31 | Diarrhea | |
| B | 3 | Acute febrile pharyngitis; adenopharyngo-conjunctival fever; pneumonia; follicular conjunctivitis; fatal infection in neonates |
| 7 | Acute febrile pharyngitis; adenopharyngo-conjunctival fever; acute respiratory disease with pneumonia; fatal infection in neonates; meningoencephalitis | |
| 11 | Follicular conjunctivitis; hemorrhagic cystitis in children | |
| 21 | Hemorrhagic cystitis in children; fatal infection in neonates | |
| 14, 16, 50, 55 | Acute respiratory disease with pneumonia | |
| 34, 35 | Acute and chronic infection in patients with immunosuppression and AIDS | |
| C | 1, 2, 6 | Acute febrile pharyngitis; pneumonia in children |
| 5 | Acute febrile pharyngitis; pertussis-like syndrome; acute and chronic infection in patients with immunosuppression and AIDS | |
| D | 8, 19, 37 | Epidemic keratoconjunctivitis |
| 9, 10, 13, 15, 17, 42 | ||
| 19, 20, 22–29 | ||
| 30 | Fatal infection in neonates | |
| 32, 33, 36, 38 | Asymptomatic (36 obesity) | |
| 39, 42–51, 53, 54, 56 | Acute and chronic infection in patients with immunosuppression and AIDS | |
| E | 4 | Respiratory infection |
| F | 40, 41 | Diarrhea |
Adenoviruses gain access to susceptible individuals through the mouth, the nasopharynx or the conjunctiva. Although initial infection may occur via the respiratory route, fecal–oral transmission accounts for most adenovirus infections in young children because of the prolonged shedding of viruses in feces (Horwitz, 1996).
Nose, throat and eye infections caused by adenoviruses have been associated with improperly disinfected swimming pool water. The enteric adenoviruses 40 and 41 have been recognized as the second most important etiological agents of viral gastroenteritis in children. These viruses, in contrast to other adenoviruses, are not shed in respiratory secretions; thus, their transmission is limited to the oral–fecal route. They have been associated with several outbreaks associated with drinking water (Divizia et al., 2004).
Adenoviruses are among the most common viruses in untreated sewage, where they have been found in concentrations 10 times greater than that of the enteroviruses. Contaminated inanimate surfaces may play a significant role in adenovirus transmission because of its stability to drying. At room temperature, adenovirus 2 survives for 8 and 12 weeks at low (7%) and high (96%) relative humidity, respectively, and is more resistant than poliovirus 2, vaccinia virus, coxsackievirus B3 and herpes virus (Boone and Gerba, 2007). Longer survival of adenoviruses in water has also been observed. Adenovirus type 5 survives longer in tap water, at either 4 or 18°C, than either poliovirus 1 or echovirus 7 (Bagdasar’yan and Abieva, 1971), and enteric adenoviruses 40 and 41 survive longer than poliovirus 1 and HAV in tapwater and seawater (Enriquez et al., 1995b). The increased survival of the enteric adenoviruses in tapwater and seawater, and the faster inactivation in sewage, may indicate that these viruses are inactivated by different mechanisms than those affecting the enteroviruses. The enteric adenoviruses are more thermally stable than poliovirus 1, which is inactivated faster at temperatures above 50°C (Enriquez et al., 1995a). In addition, enteric adenoviruses 40 and 41 are more resistant to ultraviolet (UV) light disinfection than poliovirus type 1 and coliphage MS-2, which has been suggested as a model for enteric virus disinfection (Meng and Gerba, 1996).
The increased resistance showed by enteric adenoviruses, compared with other enteric viruses, may be associated with the double-stranded nature of their DNA, which, if damaged, may be repaired by the host cell DNA repair mechanisms. This mechanism would not be effective with ssRNA genome viruses such as poliovirus 1 or HAV. It has been suggested that the longer survival of the enteric adenoviruses in tapwater and seawater may be associated with DNA damage, and the faster inactivation in sewage may result from protein capsid damage.
22.4.1.3. Enteroviruses and Paraechoviruses
The enteroviruses are members of the family Picornaviridae, which are among the smallest ribonucleic acid (RNA) viruses. “Picornavirus” means small RNA virus. Enteroviruses and Paraechoviruses are icosahedral viruses approximately 27 to 32 nm in diameter. Enteroviruses are divided into five groups (Table 22.12 ). The nucleic acid of enteroviruses consists of ssRNA. These are the viruses most often detected in sewage polluted water. However, their apparent higher prevalence may be associated, in part, with available cell lines for their propagation, because many pathogenic enteric viruses such as HAV, enteric adenoviruses, rotavirus, norovirus and other small round viruses are difficult to grow in conventional cell lines.
Table 22.12.
Some Human Enteroviruses and Paraechoviruses and Clinical Illness
| Virus | Serotypes | Clinical Illness |
|---|---|---|
| Poliovirus | 3 types | Paralysis, aseptic meningitis, febrile illness |
| Enterovirus A | 12 types | Paralysis, aseptic meningitis, hand, foot, and mouth disease, encephalitis |
| Coxsackievirus | A 2–7 | |
| A 8–16 | ||
| Enterovirus | 71 | Herpangina, exanthema, diarrhea |
| 76 | ||
| Enterovirus B | 37 types | Aseptic meningitis, paralysis, exanthema, respiratory diseases |
| Coxsackievirus | B1-B6, A9 | |
| Echovirus | 1–9, 11–21, 24–33 | Diarrhea |
| Enterovirus | 69, 73–91 | Pericarditis, myocarditis, febrile illness |
| Enterovirus C | 11 types | Paralysis, aseptic meningitis |
| Coxsackievirus A | 1, 11, 13, 15, 17–22, 24 | Myocarditis, encephalitis |
| Enterovirus D | 68, 70 | Pneumonia, acute hemorrhagic conjunctivitis |
| Enterovirus | ||
| Paraechoviruses | 1–6 | Pericarditis, herpangina, respiratory disease |
Although viruses belonging to the Enterovirus and Paraechovirus genera are capable of causing a wide variety of clinical conditions, from asymptomatic to disabling, or even fatal infections, they often do not cause overt disease (Table 22.12).
Despite the fact that there are bovine, porcine, simian and murine enteroviruses, it is believed that humans are the only natural host of human enteroviruses. There are currently more than 100 human enterovirus types known (Piralla et al., 2013). Enteroviruses replicate primarily in the gastrointestinal tract, and may be shed in large numbers (approximately 1010/g feces). The most common forms of transmission include the fecal–oral and respiratory routes (Figure 22.23 ). Waterborne transmission may be considered a form of fecal–oral transmission in which the responsible vehicle is water instead of hands or fomites. Although enteroviruses are readily found in fecally contaminated drinking or recreational waters, waterborne enterovirus infection has been only occasionally documented. Waterborne outbreaks related to enteroviruses are difficult to document because many infections by these agents are subclinical. Therefore, an individual with waterborne infection without overt disease may infect others, who in turn may become ill, spreading the infection further. Attack rates of enteroviruses vary depending on the virus and the host age. Asymptomatic infections by poliovirus outnumber symptomatic disease (10:1), whereas symptomatic infection by echovirus 9 is relatively high (10:7) (Morens and Pallansch, 1995).
Figure 22.23.
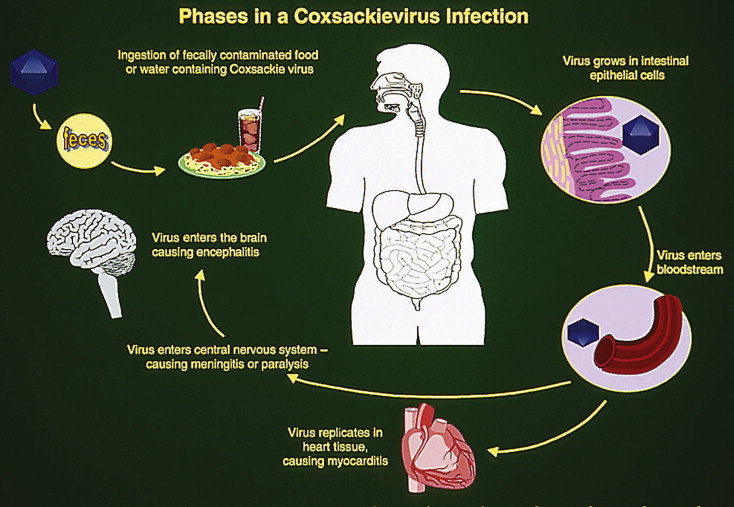
Infection by Coxsackieviruses.
Both disease and infection caused by polioviruses are age related. Generally, infection is more common in infants, but adults and older children are more severely affected. However, some exceptions exist. Coxsackievirus B virus infection is usually more severe in newborns than in older children and adults, often causing fulminant myocarditis, encephalitis, hepatitis and death. Coxsackieviruses are the most prevalent nonpolio enteroviruses (Morens and Pallansch, 1995). Coxsackieviruses are also the most common nonpolio enteroviruses isolated from water and wastewater. These viruses have been associated with several serious illnesses (Table 22.12). Coxsackievirus B5 infection has been associated with recreational water. In an outbreak at a boys’ summer camp, this virus was isolated from lake water; however, person-to-person contact appeared to be the main form of transmission (Hawley et al., 1973).
It is believed that almost all enteroviruses can be transmitted by the fecal–oral route; however, it is not clear if all of them can additionally be transmitted by the respiratory route. Airborne transmission of enterovirus might include aerosol spread or direct exposure to respiratory secretions. Fecal–oral transmission may predominate in areas with poor sanitary conditions, whereas respiratory transmission may occur more often with better sanitation. In temperate climates, enteroviruses are more common during the summer season. In the United States, most enterovirus isolations (82%) occur from June to October. However, vaccine strains of poliovirus are isolated year-round because of routine vaccination of children. In contrast, in tropical and semitropical areas, enteroviruses do not show seasonality. Transmission of enteroviruses within a household is usually started by young children; then the infection spreads quickly to other family members, especially in larger families living under crowded conditions with poor hygiene. Paralytic poliomyelitis and perhaps some nonpolio enteroviral diseases are more often observed in developing countries, where sanitary conditions are improving. With poor hygiene, most individuals are infected at a very early age, when infection rarely results in overt disease and maternal immunity limits infection. This early exposure to the virus elicits a protective immune response on reexposure to these viruses later in life. In contrast, when early exposure is prevented or delayed as a result of better sanitation, an initial poliovirus infection is likely to occur at an older age, when maternal immunity has waned and the possibility of developing a more severe clinical condition is greater.
22.4.1.4. Hepatitis A Virus (HAV)
HAV is a picornavirus morphologically indistinguishable from other members of the same family. This agent was formerly classified as a member of the Enterovirus genus (enterovirus 72). However, differences in nucleotide and amino acid sequences resulted in its classification as the only member of the hepatovirus group.
The average HAV incubation is approximately 30 days, but it may vary from 10 to 50 days, a variation associated with the dose. Infection with very few particles results in longer incubation periods and vice versa. The period of communicability extends from early in the incubation period to about a week after the development of jaundice. The greatest danger of spreading the disease to others occurs during the middle of the incubation period, well before the first presentation of symptoms. During this period the patient remains asymptomatic; however, active shedding of the virus is the norm. Therefore, it is during the incubation time that the infected individual has the highest potential for spreading HAV.
Hepatitis A is usually a mild illness, which almost always results in complete recovery. Severity and disease manifestation are age related. An estimated 80 to 95% of infected children younger than 5 years of age do not develop overt disease, whereas clinical manifestations are observed in approximately 75 to 90% of infected adults. The mortality rate in children 14 years of age or younger is 0.1%; this rate rises to 0.3% in individuals between the ages of 15 and 39 years and 2.1% in those older than 40 years.
Hepatitis A is characterized by sudden onset of fever, malaise, nausea, anorexia and abdominal discomfort, followed in several days by jaundice. In contrast to hepatitis B, HAV infection is not chronic. HAV is excreted in feces of infected people, and can produce clinical disease when susceptible individuals consume contaminated water or foods. Water, shellfish and produce are the most frequent sources. Contamination of foods by infected workers in food processing plants and restaurants is not uncommon.
HAV is very stable in the environment. It is more resistant to high temperatures than poliovirus. HAV can survive at 60°C for 1 hour, and temperatures up to 85 to 95°C are needed to inactivate it in shellfish. Poor sanitation and crowding facilitate HAV transmission. Therefore, outbreaks of hepatitis A are common in institutions, crowded house projects, prisons and military forces. In developing countries, the incidence of disease in adults is relatively low because of exposure to the virus in childhood. Most individuals 18 years and older possess an immunity that provides lifelong protection against reinfection.
The survival of HAV on hands and its transfer to hands or inanimate surfaces were studied by Mbithi et al. (1992). They found that approximately 20% of the initial HAV inoculated on hands remained infectious for at least 4 hours, and that HAV inoculated onto a stainless steel surface survived for 2 hours. They also determined that exerting higher pressure and friction between HAV-contaminated hands or fomites resulted in more efficient transfer of this virus to clean hands.
With better sanitation, HAV infection shifts to older individuals, and the incidence of overt disease increases. In more developed countries, low levels of HAV transmission occur. However, disease outbreaks are relatively common in most of these countries as a significant segment of the population is susceptible to HAV infection. The introduction of a vaccine for HAV has resulted in a steady decline in cases in the United States (see Case Study 22.2 ).
Case Study 22.2. A Massive Waterborne Outbreak of Infectious Hepatitis.
Approximately 35,000 young adults in New Delhi, India came down with hepatitis in the fall of 1955. There were 73 deaths recorded among those affected, one of the largest viral water-borne outbreaks ever documented. New Delhi obtains its drinking water from the Jamuna River. Because of flooding following the monsoon season, water drawn from the river for treatment and distribution became heavily contaminated with sewage from a nearby creek. From November 11 to November 16, raw wastewater freely entered the drinking water treatment plant. The staff at the plant, aware of the contamination, increased the dose of alum coagulant and chlorine in an effort to eliminate contaminating pathogens. About three weeks later, the first hepatitis cases were identified. The observed incubation period ranged from 22 to 60 days, with an average of 40 days. Surprisingly, no increase in other types of viral, bacterial, or parasitic infections was observed during the hepatitis epidemic, possibly because of the partial success of emergency disinfection using high levels of chlorine.
In an attempt to isolate the epidemic’s causative agent, researchers at the Virus Research Center in Poona inoculated eight different cell lines with a variety of clinical specimens obtained from hepatitis patients. Unfortunately, no viruses were isolated from any of the samples tested. Although the etiologic agent was not identified, it was believed that the epidemic was caused by the hepatitis A virus (HAV). A retrospective study conducted in 1980 showed that the outbreak in New Delhi was not caused by HAV or hepatitis B virus (HBV) but by the so-called enterically transmitted non-A, non-B (ENANB) hepatitis virus. This agent was later renamed hepatitis E virus (HEV). Although waterborne outbreaks of hepatitis E have occurred in many regions of the world, outbreaks have almost exclusively been in developing countries. HEV waterborne outbreaks primarily affect young adults, and fatalities among pregnant women are not uncommon.
The raw sewage contamination of drinking water that led to the large HEV outbreak in New Delhi was thought to be controlled by the use of high doses of alum coagulant and chlorine. Although this measure prevented an outbreak of bacterial, parasitic, and other enteric viral diseases, it failed to eliminate HEV. Therefore, prevention of contamination, rather than extensive treatment, offers the best alternative for safe drinking water.
22.4.1.5. Hepatitis E Virus (HEV)
The hepatitis E virus is the leading cause of acute viral hepatitis among young and middle-aged adults in developing countries. HEV has a diameter of 32–34 nm and an ssRNA genome. Currently, HEV is placed in a sole genus Hepevirus within a new family Hepeviridae. It has been suggested that the higher prevalence of HEV in adults may be due to a silent infection early in life, with subsequent waning of immunity after 10 to 20 years, when they again become susceptible to infection by HEV (Bradley, 1992). Important epidemiological features of HEV infection are the frequent occurrence of outbreaks associated with consumption of sewage-polluted water and its severity, particularly among pregnant women, in whom the case-fatality rate may be as high as 25% (Balayan, 1993). HEV virus also infects swine and other animals, and has been transmitted by the consumption of undercooked meat (Tamada et al., 2004). To date, no outbreak has occurred in the United States.
Hepatitis E is clinically indistinguishable from hepatitis A. It is characterized by jaundice, malaise, anorexia, abdominal pain, arthralgia and fever. The incubation period for hepatitis E varies from 2 to 9 weeks. The disease is most often seen in young to middle-aged adults (15–40 years old). The disease is usually mild and resolves in 2 weeks, leaving no long-term effects, with a relatively low fatality rate (0.1–1%). There is no evidence of immunity against HEV in a population that has been exposed to this virus (Margolis et al., 1997).
22.4.1.6. Rotavirus
Rotaviruses are classified within the family Reoviridae. These viruses have a characteristic genome consisting of 11 dsRNA segments surrounded by a distinctive two-layered protein capsid. Particles are approximately 70 nm in diameter (Figure 22.24 ). Six serological groups (A–F) have been identified; groups A, B and C infect both humans and animals, and D, E and F have been detected only in animals. Group A rotaviruses have been associated with the majority of infantile acute gastroenteritis cases, group B with severe diarrhea epidemics in adults in China and group C with sporadic cases of diarrhea in children, but their clinical importance has not been determined. Within each group, rotaviruses are classified into serotypes.
Figure 22.24.
Electron micrograph of human rotavirus.
Courtesy Centers for Disease Control.
Rotaviruses are the most important agents of infantile gastroenteritis around the world. Group A rotavirus is endemic worldwide. It is the leading cause of severe diarrhea among infants and children, and accounts for about half of the cases requiring hospitalization. In temperate areas, it occurs primarily in the winter, but in the tropics it occurs throughout the year. Group B rotavirus, also called adult diarrhea rotavirus, has caused major epidemics of severe diarrhea affecting thousands of persons of all ages in China. Group C rotavirus has been associated with rare and sporadic cases of diarrhea in children in many countries.
Rotaviruses are shed in large numbers, up to 1010 viral particles per gram of feces (White and Fenner, 1994). Names applied to the infection caused by the most common and widespread group A rotavirus include: acute gastroenteritis; infantile diarrhea; winter diarrhea; acute nonbacterial infectious gastroenteritis; and acute viral gastroenteritis.
Rotaviruses are transmitted by the fecal–oral route. Exposure by person-to-person spread via contaminated hands is probably one of the most important routes by which rotaviruses are transmitted. Institutions or close communities such as pediatric wards, day care centers and family homes are usually most affected by outbreaks of gastroenteritis caused by rotaviruses. Because of high infectivity, rotavirus-infected food handlers may contaminate foods that require handling but do not require further cooking, such as cakes, salads and fruits. Among adults, multiple foods served at banquets were implicated in two outbreaks, and an outbreak related to contaminated municipal water occurred in Colorado in 1981. Several large outbreaks of group B rotavirus involving millions of persons as a result of sewage contamination of drinking water supplies have occurred in China since 1982. Although to date, outbreaks caused by group B rotavirus have been confined to mainland China, seroepidemiological surveys have indicated a lack of immunity in the United States to this group of viruses. Rotaviruses are quite stable in the environment and have been found in estuary samples at levels as high as one to five infectious particles per gallon (Anonymous, 1992). Sanitary measures adequate for bacteria and parasites seem to be ineffective in endemic control of rotavirus, as similar incidences of rotavirus infection are observed in countries with both high and low health standards. Rotaviruses have caused numerous water and foodborne outbreaks in both adults and children (Gerba et al., 1996, Greening, 2006).
Rotavirus gastroenteritis is a self-limiting disease, which can be mild to severe. It is characterized by vomiting, watery diarrhea and mild fever. Asymptomatic rotavirus excretion has been well documented and may play a role in perpetuating endemic disease. The incubation period ranges from 1 to 3 days. Symptoms often start with vomiting followed by 4–8 days of diarrhea. As with other viral gastroenteritis, rotavirus gastroenteritis treatment consists of fluid and electrolyte replacement; if it is untreated, severe diarrhea with dehydration and death may occur. Individuals of all ages are susceptible to rotavirus infection. Premature infants, children 6 months to 2 years of age, the elderly and the immunocompromised are particularly prone to develop more severe symptoms. In adults, rotaviral infection is usually subclinical. Each year rotavirus causes millions of cases of diarrhea in developing countries, almost 2 million resulting in hospitalization, and an estimated 453,000 resulting in the death of a child younger than 5 years, 85% of whom live in developing countries (Tate et al., 2012). The introduction of a vaccine for children has been reducing the number of serious cases of rotavirus in the United States. Rotavirus infection does not result in an efficient or long-lasting immunity. Therefore, rotavirus infection in the same child often occurs up to six times during childhood.
Some animal rotaviruses, such as SA-11, are readily propagated in cell culture; the human rotaviruses, however, are rather fastidious. These viruses have not been grown efficiently in any conventional tissue culture system.
22.4.1.7. Human Caliciviruses
The use of electron microscopy (EM) since the early 1970s for the examination of fecal specimens from individuals suffering from nonbacterial gastroenteritis has shown many previously unknown viruses. Among these, viruses such as adenoviruses and rotaviruses are larger and well defined; thus, they are relatively easy to identify by EM. However, many smaller (20–40 nm) viruses without a distinctive morphology are often present in fecal samples of patients suffering from gastroenteritis (Figure 22.25 ). Norwalk virus was the first to be described (Kapikian et al., 1972). Subsequently, other small round viruses were observed in diarrheal stools, namely Montgomery County, Hawaii, Wollan, Ditchling, the Parramatta, the Cockle agents and minirotavirus. Norwalk-like viruses are in the calicivirus family, and are now divided into two genera which infect humans; Norovirus and Sapovirus. There are also several genera of caliciviruses that only infect animals. Norovirus are major causes of both food- and waterborne disease. It is estimated that 67% of the foodborne illness in the United States are caused by noroviruses (Gerba and Kayed, 2003). Caliciviruses are nonenveloped viruses with a diameter of approximately 26 to 35 nm and a positive-sense ssRNA genome.
Figure 22.25.
Norwalk virus (norovirus) capsid.
Infectivity studies with volunteers have shown that individual susceptibility is more important than acquired immunity. It has been suggested that genetically determined factors are the primary determinants of resistance to Norovirus infection, perhaps at the level of cellular receptor sites (i.e., blood group antigens expressed in the gut). In developing countries, antibodies to Norovirus are acquired early in life, and the peak incidence of illness may also occur among younger age groups than in developed nations. Noroviruses and related viruses usually produce a mild and brief illness, lasting 1 to 2 days. It is characterized by nausea and abdominal cramps, followed commonly by vomiting in children and diarrhea in adults. Human noroviruses cannot be grown in cell culture on a continuous basis, and the murine Norovirus has been often used as a model for environmental studies.
22.4.2. Respiratory Viruses
Worldwide, respiratory illnesses are the most common illnesses in humans and most have a viral etiology. Respiratory disease is associated with a large number of viruses, including: rhinoviruses; coronaviruses; parainfluenza viruses; respiratory syncytial virus (RSV); influenza virus; and adenovirus. These viruses, when they infect the upper respiratory tract, can cause acute viral rhinitis or pharyngitis (common cold); when the primary site of infection is the lower respiratory tract, they can cause laryngotracheitis (croup), bronchitis or pneumonia.
Mortality related to acute respiratory disease may be especially significant in children and in the elderly. In adults, temporary disability results in important economic loss. Respiratory infection often results from self-inoculation, when virus-contaminated hands or fingers rub the eyes or when viruses are introduced into the mouth or nose. Another important route of transmission of respiratory viruses is inhalation of contaminated aerosols.
22.4.2.1. Rhinoviruses
Rhinoviruses (Latin rhino, nose) belong to the family Picornaviridae. Two important characteristics differentiate rhinoviruses from enteroviruses: stability at low pH values and temperature. Whereas the rhinoviruses are inactivated at pH values below 6, the enteroviruses are stable at low pH values. In contrast, rhinoviruses are stable at temperatures (50°C) that would inactivate most enteroviruses (Couch, 1996). Human rhinoviruses have an icosahedral morphology, containing a single-stranded RNA genome. The diameter of these viruses is approximately 25 to 30 nm. Although no etiologic agent is identified in half of the acute upper respiratory illnesses, it has been estimated that 30 to 50% are caused by rhinoviruses. In fact, rhinovirus infection is probably the most common type of human acute infection (Gwaltney, 1997). The optimal temperature for rhinovirus growth is 33 to 35°C, which corresponds to the normal temperature of the nasal mucosa. This may explain why these viruses propagate most efficiently in the upper respiratory tract. These viruses can be propagated in monkey and human cell lines. There are more than 100 different human rhinovirus serotypes. Although these viruses are species specific, an equine rhinovirus can infect other species, including humans. Experimentally, chimpanzees and gibbons have been infected with human rhinoviruses, but these animals do not develop disease. Volunteer studies have shown that infection can be started with less than one tissue culture median infectious dose (TCID50).
Rhinovirus infection, after 1 to 4 days of incubation, is characterized by nasal obstruction and discharge, sneezing, scratchy throat, mild cough and malaise. The relatively high rate of rhinovirus infection may be associated with the large number of rhinovirus serotypes, and the fact that the same serotype can infect an individual more than once. In addition, immunity to rhinovirus infection is short lived.
Although rhinovirus infection occurs throughout the year, in temperate climates its frequency increases during colder months, and in the tropics the peak incidence occurs during the rainy season. Experiments with volunteers have failed to associate exposure to low temperatures with an increased susceptibility to rhinovirus infection. Therefore, the cause of this seasonality remains unclear. Nevertheless, it has been suggested that colder temperatures or rain may increase the survivability of these viruses and/or promote crowding conditions, in which the virus may propagate more efficiently.
During rhinovirus infection, the highest concentration of viruses is found in the nasal and pharyngeal mucosa. In contrast, in saliva, rhinoviruses are found in low numbers, and inconsistently. This suggests that aerosols may not play an important role in the transmission of these agents, as sneezing and coughing generate aerosols composed mainly of saliva (Figure 22.26 ). In fact, studies with volunteers have shown that rhinovirus transmission by hand contact and self-inoculation of the eye or nasal mucosa is much more efficient than through aerosols (Hendley et al., 1973).
Figure 22.26.
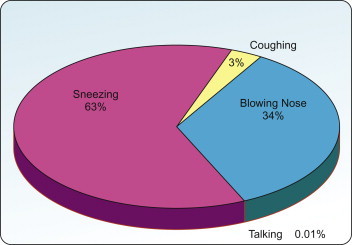
Source of rhinovirus dispersion.
Data from Buckland and Tyrrell (1964).
The high concentration of rhinoviruses in nasal secretions easily leads to hand contamination. Rhinoviruses have been isolated from 40 to 90% of hands of ill persons, and from 6 to 15% of environmental objects such as doorknobs, dolls, coffee cups and glasses. It is believed that contamination of objects in the environment may play a significant role in the transmission of rhinoviruses. This, and the ability of rhinoviruses to survive on human skin and on inanimate objects, may explain the more prevalent self-inoculation form of transmission. Both finger-to-eye and finger-to-nose contact are part of normal human behavior. Therefore, experimental rhinovirus transmission has been effectively interrupted by hand disinfection. Rhinoviruses are relatively stable on inanimate surfaces. They can survive at least 3 hours on human skin, nonporous materials such as plastic surfaces, Formica, stainless steel and hard synthetic fabrics such as nylon and dacron. In porous materials such as paper tissue and cotton fabric, rhinoviruses can survive for 1 hour (Hendley et al., 1973).
22.4.2.2. Paramyxovirus
Parainfluenza viruses and respiratory syncytial viruses (RSV) belong to the family Paramyxoviridae. These viruses are spherical enveloped particles of heterogeneous size, ranging from 125 to 250 nm. The paramyxovirus viral particle has a lipid envelope covered with spikes of about 10 nm. The nucleic acid of these agents consists of a single-stranded RNA molecule (Ginsberg, 1990).
RSV and parainfluenza viruses do not survive well in the environment. If suspended in a protein-free medium at 4°C, 90 to 99% of their infectivity is lost within 4 hours. Organic solvents and detergents rapidly inactivate these viruses by dissolving their lipid envelopes. Both RSV and parainfluenza viruses cause a variety of illnesses, primarily in infants and young children. The most important are common cold, bronchitis, bronchiolitis, laryngotracheobronchitis (croup) and pneumonia.
22.4.2.3. Parainfluenza Viruses
The parainfluenza viruses constitute two of the four genera of the family Paramyxoviridae: Paramyxovirus (parainfluenza virus types 1 and 3) and Rubulavirus (parainfluenza virus types 2 and 4). Parainfluenza viruses infect most people during childhood. Types 1 and 2 are often associated with croup in infants and type 3 with bronchiolitis and pneumonia; type 4 seldom causes illness. Although parainfluenza infections in children and infants can result in serious disease, most infections are subclinical. In adults, these viruses can cause a mild cold, but, as in children, most infections are also subclinical. Survival studies have shown that parainfluenza virus type 3 remains infectious in aerosol particles for at least 1 hour (Miller and Artenstein, 1967).
Disease caused by parainfluenza viruses is observed throughout the year. Infection by parainfluenza virus types 1 and 2 occurs endemically, but small epidemics caused by these agents are observed every 2 years. Parainfluenza virus type 3 infects approximately 60% of infants during the first 2 years of life, reaching 80% by 4 years of age, whereas infection by types 1 and 2 does not reach 80% until 10 years of age. In children, an incubation period of 2 to 4 days has been estimated for parainfluenza virus illness. Studies have shown that a child infected with parainfluenza type 3 may shed viruses over an average of 8 days, but shedding viruses for up to 4 weeks has been documented (Frank et al., 1981). During the fall, parainfluenza has been found on 30% of common fomites in office buildings in the United States (Boone and Gerba, 2010).
22.4.2.4. Respiratory Syncytial Virus
RSV belongs to the family Paramyxoviridae, as the only member of the genus Pneumovirus. The characteristics of RSV have already been described. This virus is the most important respiratory pathogen during infancy and early childhood. It causes approximately half of the cases of bronchiolitis and 25% of cases of pneumonia in infants. Approximately 90,000 hospital admissions and 4500 deaths are associated each year with RSV in the United States in both infants and young children. In approximately half of infants younger than 8 months, the infection spreads to the lower respiratory tract, resulting in life-threatening bronchitis, bronchiolitis, bronchopneumonia and croup (Ginsberg, 1990). Asymptomatic first infection with RSV is rare; almost 100% of infected children develop disease. Because of poor protective immunity, reinfection is very common, but the disease is not as severe. Inoculation of adult volunteers with RSV has shown that the nose and eye mucosae are the most important portals of entry of this virus. Efficient transmission of RSV by large droplets or by touching occurs when susceptible individuals are in close contact with children shedding the virus, but not when they are exposed to small-particle aerosol (McIntosh, 1997).
The incubation period of RSV respiratory disease is 4 to 5 days. Replication of RSV in the upper respiratory tract can reach concentrations of 104 to 106 TCID50/ml of secretion, with higher titers in infants. Infected individuals may shed RSV for up to 3 weeks. In temperate climates, RSV infection may occur throughout the year, but it peaks during winter months with few occurrences in the summer; in tropical areas, outbreaks often occur during the rainy season.
22.4.2.5. Influenza Viruses
The influenza viruses belong to three genera of the family Orthomyxoviridae: influenza virus A, influenza virus B and influenza virus C. These viruses, approximately 80 to 120 nm in size, possess a lipid envelope with a genome consisting of eight segments of single-stranded RNA. The influenza virus type A is further classified into many subtypes according to host of origin, year and geographic location of first isolation.
It is not known why several recent influenza pandemics have started in China. It has been suggested that the large pig, duck and human population in the Canton area may facilitate coinfection of animals with influenza viruses that originated from different species, leading to genetic reassortment and to the generation of viruses with novel antigenic and virulence characteristics. There is strong evidence that aquatic birds are the main reservoir of all influenza viruses in other species. For example, the catastrophic influenza pandemic of 1918, in which over 20 million people died, is believed to have been caused by an influenza A virus derived from a bird.
Influenza A virus is a highly contagious agent, causing epidemics of an acute respiratory infection known as influenza, with high mortality in the elderly. Usually, about 75% of all influenza deaths occur in individuals over 55 years of age. Influenza A virus infection may be asymptomatic, but more often it may be manifested by a wide variety of clinical conditions, ranging from an annoying flu to a fatal pneumonia. For instance, a subtype of influenza A virus that was responsible for epidemics of severe disease at the beginning of the century caused minor illness in older people during widespread epidemics in the 1980s. The reason for this wide variation is poorly understood, but it has been speculated that age, underlying illnesses, previous exposure to a similar influenza A virus subtype and virus virulence may be associated with the type of disease presentation. Mortality due to influenza is significant and varies from season to season. During nine of 20 influenza seasons in the United States (1972–1992), more than 20,000 people died each season; during four seasons, more than 40,000 deaths were recorded (Centers for Disease Control, 1997).
Influenza A virus can be transmitted most efficiently through aerosols. It has been demonstrated that influenza A virus remains infectious for at least 1 hour in aerosols at room temperature (Murphy and Webster, 1996). Clinical manifestations of influenza start rather suddenly, about 1 to 4 days after infection. A disabling syndrome characterized by high fever, together with muscle pain, sore throat, nasal congestion, conjunctivitis, cough and headache, is usually the norm.
Influenza epidemics occur intermittently. In temperate climates they usually occur from early fall to late spring, but in tropical regions epidemics are observed throughout the year. Influenza epidemics spread rapidly and tend to occur worldwide. Although variable, epidemics caused by influenza A virus are observed every 2 to 4 years, whereas influenza B virus epidemics normally occur every 3 to 6 years. Immunity to influenza is long lived; however, it is virus subtype specific. Epidemiological studies have shown that individuals previously infected with influenza A subtype H1N1 during the 1957 pandemic were resistant to infection when the same subtype reappeared in the 1977 pandemic, but they were fully susceptible when exposed to other influenza A subtypes.
22.4.2.6. SARS
Severe acute respiratory virus, or SARS, is a serious respiratory virus that resulted in a worldwide outbreak in 2003 resulting in about 8000 cases and nearly 800 deaths (Figure 22.27 ). It is caused by a coronavirus believed to have originated from bats in Southeast Asia, and later transmitted to other animals. It causes a fever with chills and coughing, with 10 to 20% of persons infected developing diarrhea. It is believed to be spread by contact droplets or from fomite contact. The airborne route does not appear to be the major route of spread as close contact appears necessary for effective spread under most conditions. It is present in both the respiratory droplets and feces. It appears capable of surviving for a few days in liquids and for 24 hours on fomites (Rabenau et al., 2005). Current evidence suggests that the SARS coronavirus originated in bats, and spread to humans either directly or through animals held in Chinese markets.
Figure 22.27.
SARS virus.
Courtesy Centers for Disease Control.
22.5. Fate and Transport of Enteric Pathogens in the Environment
There are many potential routes for the transmission of excreted enteric pathogens. The ability of an enteric pathogen to be transmitted by any of these routes depends largely on its resistance to environmental factors, which control its survival and its capacity to be carried by water or air, as it moves through the environment. Some routes can be considered “natural” routes for the transmission of waterborne disease, but others—such as the use of domestic wastewater for groundwater recharge, large-scale aquaculture projects or land disposal of disposable diapers—are actually new routes created by modern human activities.
Human and animal excreta are sources of pathogens. Humans become infected by pathogens through consumption of contaminated foods, such as shellfish from contaminated waters or crops irrigated with wastewater; from drinking contaminated water; and through exposure to contaminated surface waters, as may occur during bathing or at recreational sites. Furthermore, those individuals infected by the above processes become sources of infection through their excrement, thereby completing the cycle.
In general, viral and protozoan pathogens survive longer in the environment than enteric bacterial pathogens (Figure 22.28 ). How long a pathogen survives in a particular environment depends on a number of complex factors, which are listed in Table 22.13 . Of all of the factors, temperature is probably the most important. Temperature is a well-defined factor with a consistently predictable effect on enteric pathogen survival in the environment. Usually, the lower the temperature, the longer the survival time. However, freezing temperatures generally result in the death of enteric bacteria and protozoan parasites. Enteric viruses, however, can remain infectious for months or years at freezing temperatures. Moisture—or lack thereof—can cause decreased survival, and UV light from the sun is a major factor in the inactivation of indicator bacteria in surface waters; thus, die-off in marine waters can be predicted by amount of exposure to daylight. Viruses are much more resistant to inactivation by UV light.
Figure 22.28.
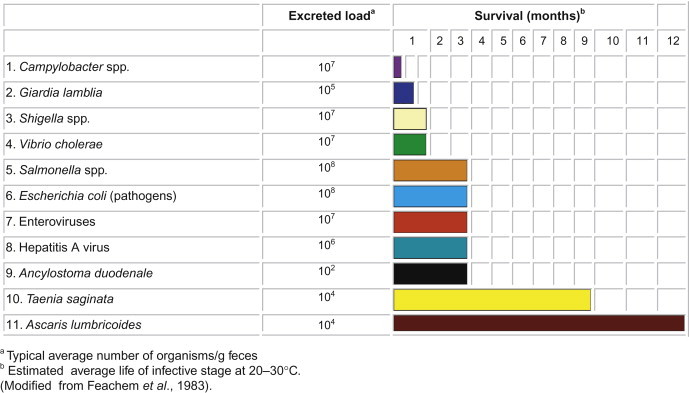
Survival times of enteric pathogens in water, wastewater, and soil and on crops.
Table 22.13.
Environmental Factors Affecting Enteric Pathogen Survival in Natural Waters
| Factor | Remarks |
|---|---|
| Temperature | Probably the most important factor; longer survival at lower temperatures; freezing kills bacteria and protozoan parasites, but prolongs virus survival. |
| Moisture | Low moisture content in soil can reduce bacterial populations. |
| Light | UV in sunlight is harmful. |
| pH | Most are stable at pH values of natural waters. Enteric bacteria are less stable at pH >9 and pH <6 |
| Salts | Some viruses are protected against heat inactivation by the presence of certain cations. |
| Organic matter | The presence of sewage usually results in longer survival. |
| Suspended solids or sediments | Association with solids prolongs survival of enteric bacteria and virus. |
| Biological factors | Native microflora is usually antagonistic. |
Many laboratory studies have shown that the microflorae of natural waters and sewage are antagonistic to the survival of enteric pathogens. It has been shown, for example, that enteric pathogens survive longer in sterile water than in water from lakes, rivers and oceans. Bacteria in natural waters can feed upon indicator bacteria. Suspended matter (clays, organic debris and the like) and fresh or marine sediments have been shown to prolong their survival time (Figure 22.29 ).
Figure 22.29.
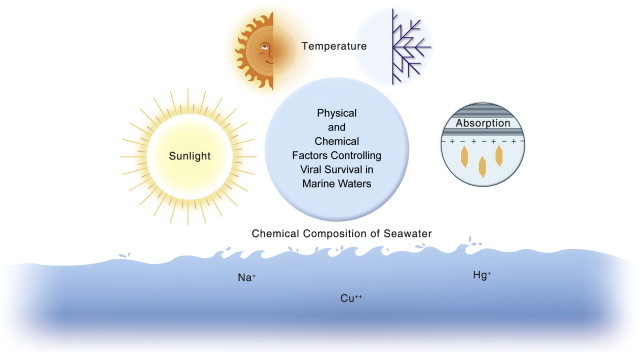
Factors affecting the survival of enteric bacteria and viral pathogens in seawater.
Questions and Problems
-
1.
What are pathogens? What is an enteric pathogen?
-
2.
What is the difference between a waterborne and a water-based pathogen? List three of each.
-
3.
Which group of enteric pathogens survives longest in the environment and why?
-
4.
What are some of the niches in which Legionella can grow to high numbers?
-
5.
Why are Cryptosporidium and Giardia major causes of waterborne disease in the United States today?
-
6.
What are the names of the environmentally resistant forms of waterborne protozoan parasites?
-
7.
What group of animals are the reservoirs of Toxoplasma gondii?
-
8.
What is the difference between a frank pathogen and an opportunistic pathogen? Give examples of each.
-
9.
Which groups of pathogens cannot grow in the environment outside a host? Which ones can?
-
10.
Which virus is the leading cause of childhood gastroenteritis worldwide?
-
11.
Which virus is most commonly associated with waterborne disease outbreaks?
-
12.
Which bacterium is the leading cause of gastroenteritis in the United States?
-
13.
Fomites are important in the spread of what respiratory viruses?
-
14.
Why are respiratory infections more common at certain times of the year?
-
15.
What virus can be transmitted by contact with the eyes?
-
16.
What type of hepatitis has a high mortality for pregnant women?
-
17.
What type of virus causes eye infections in persons swimming in contaminated waters?
-
18.
What are nosocomial infections?
-
19.
What organism is responsible for typhoid? Cholera? Winter diarrhea in infants?
-
20.
Why are only certain strains of E. coli capable of causing disease in humans?
-
21.
What type of pathogenic E. coli is transmitted primarily by cattle feces?
-
22.
What are the factors determining the survival of enteric pathogens in the environment? Which is the most important factor?
-
23.
The SARS virus is thought to originate from what group of animals?
-
24.
What group of animals is a major source of influenza in humans?
References and Recommended Reading
- Allen M.J., Geldreich E.E. Bacteriological criteria for groundwater quality. Groundwater. 1975;13:45–52. [Google Scholar]
- Anonymous . Foodborne Pathogenic Microorganisms and Natural Toxins. Federal Drug Administration; Washington, DC: 1992. [Google Scholar]
- Asghari F.B., Nikaeen M., Mirhendi H. Rapid monitoring of Pseudomonas in hospital water systems: a key priority in prevention of nosocomial infection. FEMS Microbiol. Lett. 2013;343:77–81. doi: 10.1111/1574-6968.12132. [DOI] [PubMed] [Google Scholar]
- Atkinson R.L., Dhurandhar N.V., Allison D.B., Bowen R.L., Israel B.A., Albu J.B. Human adenovirus-36 is associated with increased body weight and paradoxical reduction of serum lipids. Int. J. Obes. 2005;29:281–286. doi: 10.1038/sj.ijo.0802830. [DOI] [PubMed] [Google Scholar]
- Bagdasar’yan G.A., Abieva R.M. Survival of enteroviruses and adenoviruses in water. Hyg. Sanit. 1971;36:333–337. [PubMed] [Google Scholar]
- Balayan M.S. Hepatitis E virus infection in Europe: regional situation regarding laboratory diagnosis and epidemiology. Clin. Diagn. Virol. 1993;1:1–9. doi: 10.1016/0928-0197(93)90027-3. [DOI] [PubMed] [Google Scholar]
- Baldursson S., Karanis P. Waterborne transmission of protozoan parasites: review of worldwide outbreak—an update 2004–2010. Wat. Res. 2011;45:6603–6614. doi: 10.1016/j.watres.2011.10.013. [DOI] [PubMed] [Google Scholar]
- Baumann P. Isolation of Acinetobacter from soil and water. J. Bacteriol. 1968;96:39–42. doi: 10.1128/jb.96.1.39-42.1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bifulco J.M., Shirey J.J., Bissonnette G.K. Detection of Acinetobacter spp. in rural drinking water supplies. Appl. Environ. Microbiol. 1989;55:2214–2219. doi: 10.1128/aem.55.9.2214-2219.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bingham A.K., Jarroll E., Meyer E. Giardia spp.: physical factors of excystation in vitro, and excystation vs. eosin exclusion as determinants of viability. Exp. Parasitol. 1979;47:284–291. doi: 10.1016/0014-4894(79)90080-8. [DOI] [PubMed] [Google Scholar]
- Blaser M.J., Smith P.F., Wang W.L.L., Hoff J.C. Inactivation of Campylobacter by chlorine and monochloramine. Appl. Environ. Microbiol. 1986;51:307–311. doi: 10.1128/aem.51.2.307-311.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boone A.A., Gerba C.P. The prevalence of human parainfluenza virus 1 on indoor office fomites. Food Environ. Virol. 2010;2:41–46. [Google Scholar]
- Boone S.A., Gerba C.P. Significance of fomites in the spread of respiratory and enteric viral disease. Appl. Environ. Microbiol. 2007;73:1687–1696. doi: 10.1128/AEM.02051-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradley D.W. Hepatitis E: epidemiology, aetiology and molecular biology. Rev. Med. Virol. 1992;2:19–28. [Google Scholar]
- Brudastov A.N., Lemelev V.R., Kholnukhanedov S.K., Krasnos L.N. The clinical picture of the migration phase of ascaris in self-infection. Medskaya Parazitol. 1971;40:165–168. [PubMed] [Google Scholar]
- Buckland F.E., Tyrrell D.A. Experiments on the spread of colds. 1. Laboratory studies on the dispersal of nasal secretion. J. Hyg. 1964;62:365–377. doi: 10.1017/s0022172400040080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burden D.J., Whitehead A., Green E.A., McFadzean J.A., Beer R. The treatment of soil infected with human whipworm. Trichuris trichiura. J. Hyg. 1976;77:377–382. doi: 10.1017/s0022172400055753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burns D., Wallace R.J., Jr., Schultz M.E., Zhang Y., Zubairi S.Q., Pang Y. Nosocomial outbreak of respiratory tract colonization with Mycobacterium fortuitum: demonstration of the usefulness of pulsed-field gel electrophoresis in an epidemiological investigation. Am. Rev. Respir. Dis. 1991;144:1153–1159. doi: 10.1164/ajrccm/144.5.1153. [DOI] [PubMed] [Google Scholar]
- Carmichael W.W. The toxins of cyanobacteria. Sci. Am. 1994;273:64–72. doi: 10.1038/scientificamerican0194-78. [DOI] [PubMed] [Google Scholar]
- Carter A.M., Pacha R.E., Clark G.W., Williams E.A. Seasonal occurrence of Campylobacter spp. in surface waters and their correlation with standard indicator bacteria. Appl. Environ. Microbiol. 1987;53:523–526. doi: 10.1128/aem.53.3.523-526.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Centers for Disease Control Outbreak of cyclosporiasis associated with snow peas—Pennsylvania. MMWR. 2004;53:876–878. [PubMed] [Google Scholar]
- Centers for Disease Control (CDC) Influenza surveillance—United States, 1992–93 and 1993–94. CDC Surveill. Summ. 1997;46:SS-1. [PubMed] [Google Scholar]
- Centers for Disease Control (CDC) (2013) Ascariasis. http://www.cdc.gov/parasites/ascariasis, accessed July 10, 2013.
- Colbourne, J. S., Dennis, P. J., Trew, R. M., Beery, C., and Vesey, G. (1988) Legionella and public water supplies. Proceedings for the International Conference on Water and Wastewater Microbiology, Newport Beach, CA, February 8–11, 1988, Vol. 1.
- Collins C.H., Grange J.M., Yates M.D. Mycobacteria in water. J. Bacteriol. 1984;57:193–211. doi: 10.1111/j.1365-2672.1984.tb01384.x. [DOI] [PubMed] [Google Scholar]
- Cotte L., Rabodonirina M., Chapuis F., Bailly F., Bissuel F., Raynal C. Waterborne outbreak of intestinal microsporidiosis in persons with and without human immunodeficiency virus infection. J. Infect. Dis. 1999;180:2003–2008. doi: 10.1086/315112. [DOI] [PubMed] [Google Scholar]
- Couch R.B. Rhinoviruses. In: Fields B.N., Knipe D.M., Howley P.M., editors. Virology. Raven Press; New York: 1996. pp. 713–734. [Google Scholar]
- Craun G.F., Brunkard J.M., Yoder J.S., Roberts V.A., Carpenter J., Wade T. Causes of outbreaks associated with drinking water in the United States from 1971 to 2006. Clin. Microbiol. Rev. 2010;23:507–528. doi: 10.1128/CMR.00077-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curry A., Canning E. Human microsporidiosis. J. Infect. 1993;27:229–236. doi: 10.1016/0163-4453(93)91923-d. [DOI] [PubMed] [Google Scholar]
- de Silva D.R., Booker S., Hotez P.J., Montresor A., Engles D., Savioli L. Soil-transmitted helminth infections: updating the global picture. Trends Parasitol. 2003;19:547–551. doi: 10.1016/j.pt.2003.10.002. [DOI] [PubMed] [Google Scholar]
- Decraene V., Lebbad M., Botero-Kleiven S., Gustavsson A.M., Lofdahl M. First reported foodborne outbreak associated with microsporidia, Sweden, October 2009. Epidemiol. Infect. 2012;140:519–527. doi: 10.1017/S095026881100077X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Divizia M., Gabrieli R., Donia D., Macaluso A., Bosch A., Guix S. Waterborne gastroenteritis outbreak in Albania. Wat. Sci. Technol. 2004;50:57–61. [PubMed] [Google Scholar]
- Dubey J.P. Strategies to reduce transmission of Toxoplasma gondii to animals and humans. Vet. Parasitol. 1996;64:65–70. doi: 10.1016/0304-4017(96)00961-2. [DOI] [PubMed] [Google Scholar]
- Dubey J.P., Beattie C.P. Toxoplasmosis of Animals and Man. CRC Press; Boca Raton, FL: 1988. [Google Scholar]
- Enriquez, C. E., Garzon-Sandoval, J., and Gerba, C. P. (1995a) Survival, detection and resistance to disinfection of enteric adenoviruses. Proceedings of the 1995 Water Quality Technology Conference, American Water Works Association, New Orleans, pp. 2059–2086.
- Enriquez C.E., Hurst C.J., Gerba C.P. Survival of the enteric adenoviruses 40 and 41 in tap, sea, and wastewater. Wat. Res. 1995;29:2548–2553. [Google Scholar]
- Erlandsen S., Sherlock L., Januschka M., Schupp D., Schaefer F., Jakubowski W. Cross-species transmission of Giardia spp.: inoculation of beavers and muskrats with cysts of human, beaver, mouse, and muskrat origin. Appl. Environ. Microbiol. 1988;54:2777–2785. doi: 10.1128/aem.54.11.2777-2785.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erlandsen S.L. Biotic transmission—giardiasis a zoonosis? In: Thompson R., Reynoldson J., Lymbery A., editors. Giardia: From Molecules to Disease. University Press; Cambridge: 1995. pp. 83–97. [Google Scholar]
- Falcao D.P., Valentini S.R., Leite C.Q.F. Pathogenic or potentially pathogenic bacteria as contaminants of fresh water from different sources in Araraquara, Brazil. Wat. Res. 1993;27:1737–1741. [Google Scholar]
- Feachem R.G., Bradley D.J., Garelick H., Mara D.D. Sanitation and Disease: Health Aspects of Excreta and Wastewater Management. John Wiley and Sons; Chichester: 1983. [Google Scholar]
- Fields B.S., Shotts E.B., Jr., Feeley J.C., Gorman G.W, Martin W.T. Proliferation of Legionella pneumophila as an intracellular parasite of the ciliated protozoan Tetrahymena pyriformis. Appl. Environ. Microbiol. 1984;47:467–471. doi: 10.1128/aem.47.3.467-471.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank A.L., Taber L.H., Wells C.R., Wells J.M., Glezen W.P., Paredes A. Patterns of shedding of myxoviruses and paramyxoviruses in children. J. Infect. Dis. 1981;144:433–441. doi: 10.1093/infdis/144.5.433. [DOI] [PubMed] [Google Scholar]
- Freedman D.O. Intestinal nematodes. In: Gorbach S.L., Bartlett J.G., Blacklow N., editors. Infectious Diseases. W. B. Saunders; Philadelphia: 1992. pp. 2003–2008. [Google Scholar]
- Frenkel J.K., Ruiz A., Chinchilla M. Soil survival of Toxoplasma oocysts in KS and Costa Rica. Am. J. Trop. Med. Hyg. 1975;24:439–443. doi: 10.4269/ajtmh.1975.24.439. [DOI] [PubMed] [Google Scholar]
- Gerba C.P., Kayed D. A major cause of foodborne illness. J. Food Sci. 2003;68:1136–1142. [Google Scholar]
- Gerba C.P., Rose J.B., Haas C.N., Crabtree K.D. Waterborne rotavirus: a risk assessment. Wat. Res. 1996;30:2929–2940. [Google Scholar]
- Ginsberg H.S. Paramyxoviruses. In: Davis B.D., Dulbecco R., Eisen H.N., Ginsberg H.S., editors. Microbiology. 4th ed. J. B. Lippincott; Philadelphia: 1990. pp. 947–959. [Google Scholar]
- Greening G.E. Human and animal viruses in food. In: Goyal S., editor. Viruses in Foods. Springer; New York: 2006. pp. 5–42. [Google Scholar]
- Gupta A., Polyak C.S., Bishop R.D., Sobel J., Mintz E.D. Laboratory-confirmed shigellosis in the United States, 1989–2002: epidemiologic trends and patterns. Clin. Infect. Dis. 2004;38:1372–1377. doi: 10.1086/386326. [DOI] [PubMed] [Google Scholar]
- Gwaltney J.M. Rhinoviruses. In: Evans A.S., Kaslow R.A., editors. Viral Infections of Humans, Epidemiology and Control. 4th ed. Plenum; New York: 1997. pp. 815–838. [Google Scholar]
- Hawley H.B., Morin D.P., Geraghty M.E., Tomkow J., Phillips A. Coxsackievirus B epidemic at a boys’ summer camp. JAMA. 1973;226:33–36. [PubMed] [Google Scholar]
- Hegarty J.P., Dowd M.T., Baker K.H. Occurrence of Helicobacter pylori in surface water in the United States. J. Appl. Microbiol. 1999;87:697–701. doi: 10.1046/j.1365-2672.1999.00912.x. [DOI] [PubMed] [Google Scholar]
- Hendley J.O., Wenzel R.P., Gwaltney J.M. Transmission of rhinovirus colds by self-inoculation. N. Engl. J. Med. 1973;288:1361–1363. doi: 10.1056/NEJM197306282882601. [DOI] [PubMed] [Google Scholar]
- Herwaldt B.L., Craun G.F, Stokes S.L., Juranek D.D. Outbreaks of waterborne disease in the United States: 1989–90. J. Am. Wat. Works Assoc. 1992;84:129–135. [Google Scholar]
- Hibler C., Hancock C. Waterborne giardiasis. In: McFeters G., editor. Drinking Water Microbiology. Springer-Verlag; New York: 1990. pp. 271–293. [Google Scholar]
- Hlavsa M.C., Roberts V.A., Hill V.R., Kahler A.M., Orr M., Garrison L.E. Surveillance for waterborne disease outbreaks and other health events associated with recreational water—United States, 2007–2008. MMWR Surveill. Summ. 2011;23:1–32. [PubMed] [Google Scholar]
- Hopkins D.R. Homing in on helminths. Am. J. Trop. Med. Hyg. 1992;46:626. doi: 10.4269/ajtmh.1992.46.626. [DOI] [PubMed] [Google Scholar]
- Horwitz M.S. Adenoviruses. In: Fields B.N., Knipe D.M., Howley P.M., editors. Fields Virology. 3rd ed. Lippincott-Raven; Philadelphia, PA: 1996. pp. 2149–2171. [Google Scholar]
- Huang P., Wever W., Sosin D., Griffin P., Long E., Murphy J. The first reported outbreak of diarrheal illness associated with Cyclospora in the United States. Ann. Intern. Med. 1995;123:409–414. doi: 10.7326/0003-4819-123-6-199509150-00002. [DOI] [PubMed] [Google Scholar]
- Hughes H.P.A. How important is toxoplasmosis? Parasitol. Today. 1985;1:41–44. doi: 10.1016/0169-4758(85)90112-7. [DOI] [PubMed] [Google Scholar]
- Isaac-Renton J., Corderio C., Sarafis K., Shahriar H. Characterization of Giardia duodenalis isolates from a waterborne outbreak. J. Infect. Dis. 1993;167:431–440. doi: 10.1093/infdis/167.2.431. [DOI] [PubMed] [Google Scholar]
- Izaguirre G. A copper-tolerant Phormidium species from Lake Mathews, CA, that produces 2-methylisoborneol and geosmin. Wat. Sci. Technol. 1992;25:217–223. [Google Scholar]
- Jepsen A., Roth H. Epizootiology of Cysticercus bovis-resistance of the eggs of Taenia saginata. Report 14. Vet. Cong. 1952;22:43–50. [Google Scholar]
- John D.T. Primary amebic meningoencephalitis and the biology of Naegleria fowleri. Annu. Rev. Microbiol. 1982;36:101–103. doi: 10.1146/annurev.mi.36.100182.000533. [DOI] [PubMed] [Google Scholar]
- Johnson D.C., Enriquez C.E., Pepper I.L., Davis T.L., Gerba C.P., Rose J.B. Survival of Giardia, Cryptosporidium, poliovirus and Salmonella in marine waters. Wat. Sci. Technol. 1997;35:261–268. [Google Scholar]
- Jones A.R., Bartlett J., McCormack J.G. Mycobacterium avium complex (MAC) osteomyelitis and septic arthritis in an immunocompetent host. J. Infect. 1995;30:59–62. doi: 10.1016/s0163-4453(95)92925-8. [DOI] [PubMed] [Google Scholar]
- Kapikian A.Z., Wyatt R.G., Dolin R., Thornhill T.S., Kalica A.R., Chanock R.M. Visualization by immune electron microscopy of 27 nm particle associated with acute infectious non-bacterial gastroenteritis. J. Virol. 1972;10:1075–1081. doi: 10.1128/jvi.10.5.1075-1081.1972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karanis P., Aldeyarbi H.M., Mirhashemi M.E., Khalil K.M. The impact of the waterborne transmission of Toxoplasma gondii and analysis for water detection: an overview and update. Environ. Sci. Pollut. Res. Int. 2013;20:86–99. doi: 10.1007/s11356-012-1177-5. [DOI] [PubMed] [Google Scholar]
- Kemp J.S., Wright S.E., Bukhari Z. On farm detection of Cryptosporidium parvum in cattle, calves and environmental samples. In: Betts W.B., Casemore D., Fricker C., Smith H., Watkins J., editors. Protozoan Parasites and Water. The Royal Society of Chemistry; Cambridge, UK: 1995. [Google Scholar]
- Kendall B.A., Winthrop K.L. Update on the epidemiology of pulmonary nontuberculus mycobacteria infections. Semin. Res. Crit. Care Med. 2013;34:87–94. doi: 10.1055/s-0033-1333567. [DOI] [PubMed] [Google Scholar]
- Klein P.D., Graham D.Y, Gaillour A., Opekun A.R., Smith E.O. Water source as risk factor for Helicobacter pylori infection in Peruvian children. Lancet. 1991;337:1503–1506. doi: 10.1016/0140-6736(91)93196-g. [DOI] [PubMed] [Google Scholar]
- Knight P. Once misidentified human parasite is a cyclosporan. ASM News. 1995;61:520–522. [Google Scholar]
- Kurtz J.B., Lee T.W. Astroviruses: human and animal. In: Bock G., Whelan J., editors. Novel Diarrhea Viruses. John Wiley & Sons; Chichester, UK: 1987. pp. 92–107. [Google Scholar]
- LeChevallier M.W, Seidler R.J., Evans T.M. Enumeration and characterization of standard plate count bacteria in chlorinated and raw water supplies. Appl. Environ. Microbiol. 1980;40:922–930. doi: 10.1128/aem.40.5.922-930.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee L.A., Shapiro C.N., Hargrett-Bean N., Tauxe R.V. Hyperendemic shigellosis in the United States: a review of surveillance data for 1967–1988. J. Infect. Dis. 1991;164:894–900. doi: 10.1093/infdis/164.5.894. [DOI] [PubMed] [Google Scholar]
- Lipp E.K., Hug A., Colwell R.R. Effects of global climate on infectious disease: the cholera model. Clin. Microbiol. Rev. 2002;15:757–770. doi: 10.1128/CMR.15.4.757-770.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucey D.R., Maguire J.H. Schistosomiasis. Infect. Dis. Clin. North Am. 1993;7:635–653. [PubMed] [Google Scholar]
- MMWR Update, Vibrio cholerae O1 Western hemisphere, 1991–1994, and V. cholerae O139—Asia, 1994. MMWR. 1995;44:215–219. [PubMed] [Google Scholar]
- MacKenzie W, Hoxie N., Proctor M., Gradus M., Blair K., Peterson D. A massive outbreak in Milwaukee of Cryptosporidium infection transmitted through the public water supply. N. Engl. J. Med. 1994;331:161–167. doi: 10.1056/NEJM199407213310304. [DOI] [PubMed] [Google Scholar]
- Majowicz S.E., Musto J., Scallan E., Angulo F.J., O’Brien S.J., Jones T.F. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010;15:882–889. doi: 10.1086/650733. [DOI] [PubMed] [Google Scholar]
- Margolis H.S., Alter M.J., Hadler S.C. Viral hepatitis. In: Evans A.S., Kaslow R.A., editors. Viral Infections of Humans, Epidemiology and Control. 4th ed. Plenum; New York: 1997. pp. 363–418. [Google Scholar]
- Marrie T.J. Community-acquired pneumonia. Clin. Infect. Dis. 1994;18:501–515. doi: 10.1093/clinids/18.4.501. [DOI] [PubMed] [Google Scholar]
- Maunula L., Kalso S., Von Bonsdorf C.H., Ponka A. Wading pool water contaminated with both noroviruses and astroviruses as the source of gastroenteritis. Epidemiol. Infect. 2004;132:737–743. doi: 10.1017/s0950268804002249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mbithi J.N., Springthorpe V.S., Sattar S.A. Survival of hepatitis A virus on human hands and its transfer on contact with animate and inanimate surfaces. J. Clin. Microbiol. 1992;30:757–763. doi: 10.1128/jcm.30.4.757-763.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McIntosh K. Respiratory syncytial virus. In: Robart H.A., editor. Human Enterovirus Infections. ASM Press; Washington, DC: 1997. [Google Scholar]
- McOrist S., Boid R., Jones T, Easterbee N., Hubbard A., Jarrett O. Some viral and protozool diseases in the European wildcat (Felis silvestris) J. Wildlife Dis. 1991;27:693–696. doi: 10.7589/0090-3558-27.4.693. [DOI] [PubMed] [Google Scholar]
- Meenhorst P.L., Reingold A.L., Groothuis D.G., Gorman G.W, Wilkinson H.W, McKinney R.M. Water-related nosocomial pneumonia caused by Legionella pneumophila serogroup 1 and 10. J. Infect. Dis. 1985;152:356–363. doi: 10.1093/infdis/152.2.356. [DOI] [PubMed] [Google Scholar]
- Melius E.J., Davis S.I., Redd J.T., Lewin M., Herlihy R., Henderson A. Estimating the prevalence of active Helicobacter pylori infection in a rural community with global positioning system technology-assisted sampling. Epidemiol. Infect. 2012;17:1–9. doi: 10.1017/S0950268812000714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng Q.S., Gerba C.P. Comparative inactivation of enteric adenovirus, polio virus, and coliphages by ultraviolet irradiation. Wat. Res. 1996;30:2665–2668. [Google Scholar]
- Messner M.J., Chappell C.L., Okhuysen P.C. Risk assessment for Cryptosporidium: a hierarchical Bayesian analysis of human dose response data. Wat. Res. 2001;35:3934–3940. doi: 10.1016/s0043-1354(01)00119-1. [DOI] [PubMed] [Google Scholar]
- Miller W.S., Artenstein M.S. Aerosol stability of three acute disease viruses. Proc. Soc. Exp. Biol. Med. 1967;125:222–227. doi: 10.3181/00379727-125-32054. [DOI] [PubMed] [Google Scholar]
- Moffat J.E., Tompkins L.S. A quantitative model of intracellular growth of Legionella pneumophila in Acanthamoeba castellanii. Infect. Immun. 1992;60:292–301. doi: 10.1128/iai.60.1.296-301.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morens D.M., Pallansch M.A. Epidemiology. In: Rotbart H.A., editor. Human Enterovirus Infections. ASM Press; Washington, DC: 1995. pp. 3–24. [Google Scholar]
- Morgan D.R., Johnson P.C., DuPont H.L., Satterwhite T.K., Wood L.V. Lack of correlation between known virulence properties of Aeromonas hydrophila and enteropathogenicity for humans. Infect. Immun. 1985;50:62–65. doi: 10.1128/iai.50.1.62-65.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mullens A. I think we have a problem in Victoria: MD’s respond quickly to toxoplasmosis outbreak in BC. Can. Med. Assoc. J. 1996;154:1721–1724. [PMC free article] [PubMed] [Google Scholar]
- Murphy B.R., Webster R.G. Orthomyxoviruses. In: Fields B.N., Knipe D.M., Howley P.M., editors. Fields Virology. 3rd ed. Lippincott-Raven; Philadelphia: 1996. pp. 1397–1445. [Google Scholar]
- Narayan L.V, Nunez W.J., III Biological control: isolation and bacterial oxidation of the taste-and-odor compound geosmin. J. Am. Wat. Works Assoc. 1974;66:532–536. [Google Scholar]
- Nguyen A.M., Engstrand L., Genta R.M., Graham D.Y., el-Zaatari F.A. Detection of Helicobacter pylori in dental plaque by reverse-transcriptase–polymerase chain reaction. J. Clin. Microbiol. 1993;31:783–787. doi: 10.1128/jcm.31.4.783-787.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nwachuku N., Gerba C.P. Occurrence and persistence of Escherichia coli O157: H7 in water. Rev. Environ. Sci. Biotechnol. 2008;7:267–273. [Google Scholar]
- Ortega Y., Sterling C., Gilman R., Cama V, Diaz F. Cyclospora species—a new protozoan pathogen of humans. N. Engl. J. Med. 1993;328:1308–1312. doi: 10.1056/NEJM199305063281804. [DOI] [PubMed] [Google Scholar]
- Payment P., Gamache F., Paquette G. Microbiological and virological analysis of water from two water filtration plants and their distribution systems. Can. J. Microbiol. 1988;34:1304–1309. doi: 10.1139/m88-228. [DOI] [PubMed] [Google Scholar]
- Phillip R., Brown M., Bell R., Francis F. Health risks associated with recreational exposure to blue–green algae (cyanobacteria) when windsurfing and fishing. Health Hyg. 1992;13:115–119. [Google Scholar]
- Piralla A., Daleno C., Scala A., Greenberg D., Usonis V., Principi N. Genome characterization of enteroviruses 117 and 118: a new group with human enterovirus species C. PLoS One. 2013;8:e60641. doi: 10.1371/journal.pone.0060641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rabenau H.F., Cinatl J., Morgenstern B., Bauer G., Preiser W., Doerr H.W. Stability and inactivation of SARS coronavirus. Med. Microbiol. Immunol. 2005;194:1–6. doi: 10.1007/s00430-004-0219-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahman A., Bonny T.S., Stonesaovapak S., Ananchaipattans C. Yersinia enterocolitica studies and outbreaks. J. Pathogens. 2011 doi: 10.2061/2011/239391. Article ID 239391, 11 pages. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ranganathan S.C., Skoric B., Ramsay K.A., Carzino R., Gibson A.M., Hart E. Geographical differences in first acquisition of Pseudomonas aeruginosa in cystic fibrosis. Ann. Am. Thorac. Soc. 2013;10:108–114. doi: 10.1513/AnnalsATS.201209-077OC. [DOI] [PubMed] [Google Scholar]
- Reavis C. Rural health alert: Helicobacter pylori in well water. J. Am. Acad. Nurse Pract. 2005;17:283–289. doi: 10.1111/j.1745-7599.2005.0047.x. [DOI] [PubMed] [Google Scholar]
- Rebold J., Hoge C., Shlim D. Cyclospora outbreak associated with chlorinated drinking water. Lancet. 1994;344:1360–1361. doi: 10.1016/s0140-6736(94)90716-1. [DOI] [PubMed] [Google Scholar]
- Rolle-Kampczyk U.E., Fritz G.J., Diez U., Lehmann I., Richter M., Herbarth O. Well water—one source of Helicobacter pylori colonization. Int. J. Hyg. Environ. Health. 2004;207:363–368. doi: 10.1078/1438-4639-00301. [DOI] [PubMed] [Google Scholar]
- Rollins D.M., Colwell R.R. Viable but nonculturable stage of Campylobacter jejuni and its role in survival in the natural aquatic environment. Appl. Environ. Microbiol. 1986;52:531–538. doi: 10.1128/aem.52.3.531-538.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samuel M.C., Vugia D.J., Shallow S., Marcus R., Seglar S., McGivern T. Epidemiology of sporadic Campylobacter infection in the United States and declining trend in incidence. Clin. Infect. Dis. 2004;38:S165–S174. doi: 10.1086/381583. [DOI] [PubMed] [Google Scholar]
- Savioli L., Renganathan E., Montresor A., Davis A., Behbehani K. Control of schistosomiasis—a global picture. Parasitol. Today. 1997;13:444–448. doi: 10.1016/s0169-4758(97)01141-1. [DOI] [PubMed] [Google Scholar]
- Schaefer F.W. Waterborne Pathogens. 2nd ed. American Water Works Association; Denver, CO: 2006. Giardia lamblia. pp. 209–215. [Google Scholar]
- Schwartz D.R., Bryan R.T, Weber R., Visvesvara G. Microsporidiosis in HIV positive patients: current methods for diagnosis using biopsy, cytologic, ultrastructural, immunological, and tissue culture techniques. Folia Parasitol. 1994;41:101–109. [PubMed] [Google Scholar]
- Schmidt G., Roberts L., editors. Foundations of Parasitology. 4th ed. Times Mirror/Mosby; St. Louis: 1989. [Google Scholar]
- Soave R.N., Johnson W. Cyclospora: conquest of an emerging pathogen. Lancet. 1995;345:667–668. doi: 10.1016/s0140-6736(95)90863-3. [DOI] [PubMed] [Google Scholar]
- States S.J., Kuchta J.M., Conley L.F, Wolford R.S., Wadowsky R.M., Yee R.B. Factors affecting the occurrence of legionnaires’ disease bacterium in public water supplies. In: Larson R.A., editor. Biohazards of Drinking Water Treatment. Lewis; Chelsea, MI: 1989. pp. 67–83. [Google Scholar]
- Stephen C., Naines D., Bellinger T., Atkinson K., Schwantje H. Serological evidence of Toxoplasma infection in cougars on Vancouver Island, British Columbia. Can. Vet. J. 1996;37:241. [PMC free article] [PubMed] [Google Scholar]
- Sterling C.R., Marshall M.M. Waterborne Pathogens. 2nd ed. American Water Works Association; Denver, CO: 2006. Cryptosporidium parvum and Cryptosporidium hominis. pp. 193–197. [Google Scholar]
- Tacket C.O., Brenner F., Blake P.A. Clinical features and an epidemiological study of Vibrio vulnificus infections. J. Infect. Dis. 1984;149:558–561. doi: 10.1093/infdis/149.4.558. [DOI] [PubMed] [Google Scholar]
- Tamada Y., Yano K., Yatsuhashi H., Inoue O., Mawatari F., Ishibashi H. Consumption of wild boar linked to cases of hepatitis E. J. Hepatol. 2004;40:869–870. doi: 10.1016/j.jhep.2003.12.026. [DOI] [PubMed] [Google Scholar]
- Tate J.E., Burton A.H., Boschi-Pinto C., Steele A.D., Duque J., Parashar U.D. 2008 estimate of worldwide rotavirus-associated mortality in children younger than 5 years before the introduction of universal rotavirus vaccination programmes: a systematic review and meta-analysis. Lancet Infect. Dis. 2012;12:136–141. doi: 10.1016/S1473-3099(11)70253-5. [DOI] [PubMed] [Google Scholar]
- Thomas J.E., Gibson G.R., Darboe M.K., Dale A., Weaver L.T. Isolation of Helicobacter pylori from human feces. Lancet. 1992;340:1194–1195. doi: 10.1016/0140-6736(92)92894-l. [DOI] [PubMed] [Google Scholar]
- Tison D.L., Seidler J. Legionella incidence and density in potable drinking water supplies. Appl. Environ. Microbiol. 1983;45:337–339. doi: 10.1128/aem.45.1.337-339.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner P.C., Gammie A.J., Hollinrake K., Codd G.A. Pneumonia associated with contact with cyanobacteria. Br. Med. J. 1990;300:1440–1441. doi: 10.1136/bmj.300.6737.1440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Wielen P.W., van der Kooij D. Nontuberculous mycobacteria, fungi, and opportunistic pathogens in unchlorinated drinking water in the Netherlands. Appl. Environ. Microbiol. 2013;79:825–834. doi: 10.1128/AEM.02748-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wainwright K.E., Miller M.A., Barr B.C., Gardner I.A., Melli A.C., Essert T. Chemical inactivation of Toxoplasma gondii oocysts in water. J. Parasitol. 2007;93:925–931. doi: 10.1645/GE-1063R.1. [DOI] [PubMed] [Google Scholar]
- Waller T. Sensitivity of Encephalitozoon cuniculi to various temperatures, disinfectants and drugs. Lab. Anim. 1979;13:227–230. doi: 10.1258/002367779780937753. [DOI] [PubMed] [Google Scholar]
- Weber R., Bryan R. Microsporidial infections in immunodeficient and immunocompetent patients. Clin. Infect. Dis. 1994;19:517–521. doi: 10.1093/clinids/19.3.517. [DOI] [PubMed] [Google Scholar]
- Wenneras C., Erling V. Prevalence of enterotoxigenic Escherichia coli-associated diarrhea and carrier state in the developing world. J. Health Popul. Nutr. 2004;22:370–382. [PubMed] [Google Scholar]
- West P.M., Olds F.R. Clinical schistosomiasis. R. I. Med. J. 1992;75:179. [PubMed] [Google Scholar]
- Whiley H., Keegan A., Giglo S., Bentham R. Mycobacterium avium complex—the role of potable water in disease transmission. J. Appl. Microbiol. 2012;113:223–232. doi: 10.1111/j.1365-2672.2012.05298.x. [DOI] [PubMed] [Google Scholar]
- White D.O., Fenner F.J. Medical Virology. 4th ed. Academic Press; San Diego: 1994. [Google Scholar]
- Wu J.T, Juttner F. Effect of environmental factors on geosmin production by Fischerella muscicola. Wat. Sci. Technol. 1988;20:143–148. [Google Scholar]
- Yilmaz S.M., Hopkins S.H. Effects of different conditions on duration of infectivity of Toxoplasma gondii oocysts. J. Parasitol. 1972;58:938–939. [PubMed] [Google Scholar]
- Young W.E., Horth H., Crane R., Ogden T., Arnott M. Taste and odor threshold concentrations of potential potable water contaminants. Wat. Res. 1996;30:331–340. [Google Scholar]
- Zacheus O.M., Martikainen P.J. Occurrence of legionellae in hot water distribution systems of Finnish apartment buildings. Can. J. Microbiol. 1994;40:993–999. doi: 10.1139/m94-159. [DOI] [PubMed] [Google Scholar]
- Zeman D., Baskin G. Encephalitozoonosis in squirrel monkeys (Saimiri sciureus) Vet. Pathol. 1985;22:24–31. doi: 10.1177/030098588502200104. [DOI] [PubMed] [Google Scholar]