

Abstract
Parasitic nematodes of livestock have major economic impact worldwide. Despite the diseases caused by these nematodes, some advances towards the development of new therapeutic agents and attempts to develop effective vaccines against some of them, there has been limited progress in the development of practical diagnostic methods. The specific and sensitive diagnosis of parasitic nematode infections of livestock underpins effective disease control, which is now particularly important given the problems associated with anthelmintic resistance in parasite populations. Traditional diagnostic methods have major limitations, in terms of sensitivity and specificity. This chapter provides an account of the significance of parasitic nematodes (order Strongylida), reviews conventional diagnostic techniques that are presently used routinely and describes advances in polymerase chain reaction (PCR)-based methods for the specific diagnosis of nematode infections. A particular emphasis is placed on the recent development of a robotic PCR-based platform for high-throughput diagnosis, and its significance and implications for epidemiological investigations and for use in control programmes.
Keywords: Parasitic nematodes, Livestock, Diagnosis, Molecular biology, Biotechnology
1. Introduction
The phylum Nematoda (roundworms) includes many parasites that are of major socio-economic importance. For example, grazing ruminants are frequently parasitized by multiple species of the order Strongylida, which can cause significant disease, known as parasitic gastroenteritis (PGE) (Kassai, 1999). Different species of strongylid nematodes can vary considerably in their pathogenicity, geographical distribution and susceptibility to anthelmintic drugs (Dobson et al., 1996). Mixed infections, involving multiple genera and species are common, and usually have a greater impact on the host than monospecific infections. Furthermore, the species composition of the parasites present in a host animal plays an important role in the severity of infection (Wimmer et al., 2004). Depending on the number, species and burden of parasitic nematodes, common symptoms of PGE include reduced weight gain or weight loss, anorexia, diarrhoea, reduced production and, in the case of blood-feeding species, anaemia and oedema, due to the loss of blood and/or plasma proteins (Kassai, 1999). Therefore, the knowledge of the nematode species present in a particular geographical area, their biology and epidemiology have important implications for the control of parasitism, particularly given the increasing problems of drug resistance in strongylid nematodes of livestock (Kaplan, 2004, Sangster, 1999, Wolstenholme et al., 2004).
Conventional coprological and immunological methods of diagnosis can be time consuming and have limitations, in terms of their sensitivity and specificity, and often do not achieve a specific diagnosis (Gasser, 2006). In particular, in the case of mixed infections, the specific detection and identification of parasites (and specific diagnosis of infection) can be laborious and time consuming using conventional methods, such as faecal egg counts (FECs) or larval culture (LC) and differentiation (Wimmer et al., 2004). Molecular techniques that rely on the amplification of nucleic acids, particularly those coupled to the polymerase chain reaction (PCR) (Saiki et al., 1988), are effective for the specific identification of parasites, and aid the diagnosis of infections from minute amounts of target template, if suitable markers are utilized. Such methods are likely to provide powerful alternative tools to traditional approaches, to underpin fundamental research into parasite epidemiology and to improve the control of parasitic disease (Gasser, 2006).
The knowledge of epidemiological factors, such as geographical and seasonal occurrence of different species of parasitic nematodes, has major implications for the control of parasitism and the development of better strategic, anthelmintic treatment regimens (Barger, 1999). For instance, in Australia, several studies have been carried out to investigate the epidemiology of livestock parasites, but there is a paucity of recent, published data for the south-eastern part of this continent; most studies were published in the 1970s (Anderson, 1972, Anderson, 1973). Furthermore, published data have been based largely on the use of traditional diagnostic approaches, and no detailed epidemiological investigations of gastrointestinal nematodes of sheep have been conducted using molecular tools.
The purpose of this chapter was to (i) concisely cover the economic impact of the diseases of livestock caused by gastrointestinal strongylid nematodes; (ii) provide a background on the biology and epidemiology of these important nematodes, and to review current information on anthelmintic resistance (AR); (iii) review conventional methods for the diagnosis of strongylid nematode infections and discuss their constraints; (iv) provide an account of key molecular-diagnostic methods, with an emphasis on recent advances through the development of robotic PCR-based technology and its significance and implications.
2. Economic Impact of Parasitic Diseases of Livestock
The livestock industry plays a major role in the economies of many developed and developing countries. The production of livestock animals provides food, animal products (e.g. leather, hides and wool), income, employment, a source of organic fertilizer and biogas as well as draught and work power (particularly important in developing countries in which modern machinery is not affordable or available). Globally, parasite infections are associated with significant economic losses to the livestock industry (Vercruysse and Claerebout, 2001). For example, nematode infections of sheep were estimated, in 2006, to cost the Australian grazing industry 369 million dollars per annum, making these parasites one of the greatest constraints on the Australian grazing industry (McLeod, 1995, Sackett and Holmes, 2006). The economic impact of parasitic diseases differs between the developed and developing worlds. In the developing world, the greatest impact of parasitic diseases relates to direct productivity losses and lost socio-economic potential. In the developed world, on the other hand, the main impact is in the costs of control (Hawkins, 1993, Perry and Randolph, 1999) associated with the use of anthelmintics, strategic anthelmintic programmes and improved pasture utilization, which all limit the impact of parasitism at the subclinical or economical levels (Corwin, 1997). How the economic impact of parasitism should be assessed, in order to aid decisions in disease control, is a topic of debate and has been reviewed in detail (Hawkins, 1993, Perry and Randolph, 1999).
In general, to measure the effects of subclinical parasitic infections, performance parameters, such as weight gain, feed conversion, forage utilization, conception rate, calving–breeding interval, milk production and disease resistance, are employed routinely (Corwin, 1997). Parasites can have various pathological effects on their host and may contribute to a decreased production performance (Hawkins, 1993).
Whilst lungworms, such as species of Dictyocaulus, Protostrongylus or Muellerius, are economically important, gastrointestinal nematodes that parasitize the abomasum, small and large intestines of ruminants are also responsible for major losses to the livestock industries (Hawkins, 1993, McLeod, 1995). Strongylid nematodes of the superfamily Trichostrongyloidea, including species of Haemonchus, Ostertagia, Teladorsagia, Trichostrongylus, and, to lesser extent, Nematodirus spp. are regarded to be of great importance, particularly in small ruminants (Berghen et al., 1993, Zajac, 2006). Other important gastrointestinal nematodes include species of Oesophagostomum, Chabertia and the bovine hookworm, Bunostomum, of which the latter two genera are regarded to be less common (Zajac, 2006). Usually, low intensities of infection do not cause a serious hazard to the health of livestock animals and may be tolerated (i.e. allowing the development of some immunity in the host), but as the numbers of worms increase, subclinical disease can manifest itself by signs of reduced appetite, weight gain, wool production and fertility, and is, therefore, of great economic importance (Fox, 1997, Zajac, 2006). These factors affect the herd productivity, the capacity to maintain and improve a herd (i.e. its health and genetic potential), human nutrition, community development and cultural issues relating to the use of livestock (Perry and Randolph, 1999). This is of particular relevance in regards to a rapidly growing human population and an associated increase in the global demand for products of animal origin, emphasizing the importance of maintaining and/or increasing the productivity of the livestock industry (Nonhebel and Kastner, 2011).
3. Strongylid Nematodes of Ruminant Livestock
The order Strongylida includes five superfamilies: the Diaphanocephaloidea, Ancylostomatoidea, Strongyloidea, Trichostrongyloidea and Metastrongyloidea. The Strongylida are characterized by the presence of a copulatory bursa and are, therefore, referred to as bursate nematodes (Anderson, 2000). The first four of these superfamilies are monoxenous (single host) and mainly inhabit the gastric and/or intestinal tracts of their vertebrate hosts. In contrast, members of the Metastrongyloidea are almost entirely heteroxenous (multiple hosts), relying in their transmission on intermediate hosts, such as earthworms and gastropods (Anderson, 2000). Adult metastrongyloids are usually found in the lungs of their hosts, but some species can be associated with the vascular system external to the lung (Anderson, 2000).
In general, with some exceptions (e.g. Nematodirus), the life cycle of the most important gastrointestinal strongylid nematodes follows a similar pattern (Fig. 4.1 ) (Levine, 1968). Sexually dimorphic adults are present in the digestive tract, where fertilized females produce large numbers of eggs which are passed in the faeces. Strongylid eggs (70–150 μm) usually hatch within 1–2 days. After hatching, larvae feed on bacteria and undergo two moults to then develop to ensheathed third-stage larvae (L3s) in the environment (i.e. faeces or soil). The sheath (which represents the cuticular layer shed in the transition from the L2 to L3 stage) protects the L3 stage from environmental conditions but prevents it from feeding. Infection of the host occurs by ingestion of L3s (with the exception of Nematodirus spp., for which the infective L3 develops within the egg). During its passage through the stomach, the L3 stage loses its protective sheath and has a histotrophic phase (tissue phase), depending on species, prior to its transition into the L4 and adult stages (Levine, 1968). Under unfavourable conditions, the larvae undergo a period of hypobiosis (arrested development; typical for species of Haemonchus and Teladorsagia). Hypobiotic larvae usually resume their activity and development in spring in the case of Haemonchus or autumn in the case of Teladorsagia. This may be synchronous with the start of the lambing season, manifesting itself in a peri-parturient increase in FECs in ewes (Salisbury and Arundel, 1970). The peri-parturient reduction of immunity increases the survival and egg production of existing parasites, increases susceptibility to further infections and contributes to the contamination of pasture with L3s when young, susceptible animals begin grazing (Hungerford, 1990).
Figure 4.1.
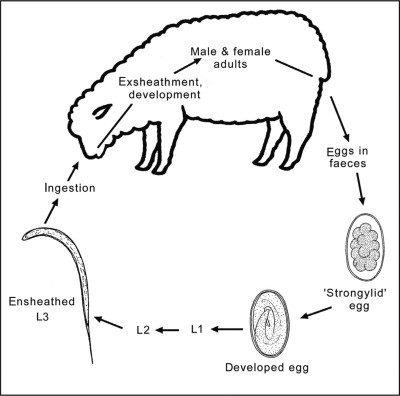
Life cycle representing gastrointestinal nematodes (order Strongylida) of small ruminants. First-, second- and third-stage larvae (L1, L2 and L3, respectively) are free-living in the environment. The fourth larval (L4) and adult stages (dioecious) are parasitic in the gastrointestinal tract of the ruminant host.
Adapted from Demeler (2005).
3.1. Key gastrointestinal nematode species of small ruminants
Of the numerous genera and species of gastrointestinal nematodes that infect small ruminants, the key representatives known to occur in Australasia are listed in Table 4.1 . The major species responsible for most disease in grazing sheep are Haemonchus contortus, Teladorsagia circumcincta and intestinal species of Trichostrongylus (Besier and Love, 2003).
Table 4.1.
The key morphological characteristics, pre-patent periods and locations in the host of the most important genera and species of gastrointestinal nematodes infecting sheep in Australasia (Anderson, 2000, Besier and Love, 2003, Gibbons, 2010, Levine, 1968, Taylor et al., 2007)
| Family | Species | Morphometrics/morphology |
Pre-patent period (days) | Location in the host | |
|---|---|---|---|---|---|
| Length (mm) | Features | ||||
| Trichostrongylidae | Haemonchus contortus | ♂ 10–20 | Red pseudocoelomic fluid and white, coiled uterus, giving a barber’s pole appearance. Presence of vulvar flap depends on strain | 18–21 | Abomasum |
| ♀ 18–30 | |||||
| Teladorsagia circumcincta | ♂ 7–8 | Small head and buccal cavity | 15–21 | Abomasum | |
| ♀ 10–12 | In females, a vulvar flap can be present | ||||
| Trichostrongylus axei | ♂ 2–6 | Dissimilar spicules of unequal length | 15–23 | Abomasum or stomach | |
| ♀ 3–8 | |||||
| T. colubriformis | ♂ 4–8 | Equal length spicules with triangular tip | 15–23 | Anterior small intestine | |
| ♀ 5–9 | |||||
| T. vitrinus | ♂ 4–7 | Equal length spicules with sharp tips | 15–23 | Anterior small intestine | |
| ♀ 5–8 | |||||
| T. rugatus | ♂ 4–7 | Dissimilar spicules with foot-like appearance | 15–23 | Small intestine | |
| ♀ 6–7 | |||||
| Cooperia curticei | ♂ 4–5 | Transverse striation of cuticle in all species | 14–15 | Small intestine | |
| ♀ 5–6 | Watch-spring-like body posture and the presence of a small cephalic vesicle are characteristic | ||||
| Molineidae | Nematodirus spathiger | ♂ 10–19 | Small but distinct cephalic vesicle | 18 | Small intestine |
| ♀ 15–29 | Very long spicules ending in a spoon-shaped terminal piece | ||||
| N. filicollis | ♂ 10–15 | Small but distinct cephalic vesicle | 18 | Small intestine | |
| ♀ 15–20 | Long and slender spicules with a narrow lanceolate membrane | ||||
| Ancylostomatidae | Bunostomum trigonocephalum | ♂ 12–17 | Anterior end is bend dorsally | 40–70 | Small intestine |
| ♀ 19–26 | Buccal capsule with is equipped with two cutting plates | ||||
| Chabertiidae | Oesophagostomum columbianum | ♂ 12–16 | Have two leaf crowns and a shallow buccal capsule. Position of cervical papillae used for species differentiation | 40–45 | Large intestine |
| ♀ 14–18 | |||||
| Oe. venulosum | ♂ 11–16 | Cervical papillae are situated posterior to the oesophagus | 40–45 | Large intestine | |
| ♀ 13–24 | |||||
| Chabertia ovina | ♂ 13–14 | Mouth is directed antero-ventrally | 42–50 | Large intestine | |
| ♀ 17–20 | Buccal capsule is subglobular without teeth | ||||
3.2. Pathogenesis and clinical signs of disease
Almost all sheep are infected with one or more gastrointestinal parasites, but the degree of severity of infection and the clinical signs associated with infections may vary considerably (e.g. Donald et al., 1978). The severity of disease is mainly influenced by factors such as the parasite species present, the number of worms present in the gastrointestinal tract, the general health and immunological state of the host, environmental factors, such as climate and pasture type, stress, stocking rate, management and/or diet (Kassai, 1999). Usually, three groups of animals are prone to heavy worm burdens: (i) young, non-immune animals; (ii) adult, immuno-compromised animals; and (iii) animals exposed to a high infection pressure from the environment (Zajac, 2006). Nematode populations in sheep are generally over-dispersed with only few sheep carrying heavy worm burdens, whilst the majority of sheep harbour low numbers of worms (Barger, 1985).
3.2.1. Haemonchus contortus
H. contortus is one of the most fecund strongyle nematodes; individual females are capable of producing thousands of eggs per day, which can lead to rapid larval pasture contamination and associated outbreaks of haemonchosis (Levine, 1968). In sheep, the pre-patent period of H. contortus is 18–21 days; adult worms are short-lived, surviving in their hosts for only a few months. The main pathogenic effects are caused by the L4s and adults, which both feed on blood, causing severe anaemia which usually becomes apparent after two weeks of infection (Baker et al., 1959). Acute disease is usually intensity-dependent and is associated with signs of haemorrhagic anaemia, dark-coloured faeces, oedema, weakness, reduced production of wool and muscle mass, or sometimes sudden death. In cases of chronic disease, decreased food intake, weight loss and anaemia are most commonly observed (Kassai, 1999). Unlike many other gastrointestinal parasites, H. contortus is not a primary cause of diarrhoea, and its effects on a flock are often not readily detected by routine observation (Zajac, 2006).
3.2.2. Teladorsagia circumcincta
Females of Te. circumcincta are less fecund than H. contortus, with an average egg production of 100–200 eggs per female per day (Cole, 1986). Teladorsagia does not feed on blood, and the main pathogenic effects are caused by its larval stages. Larval development takes place in the gastric glands, leading to nodule formation in abomasal mucosa and extensive damage to parietal cells, in turn causing a decrease in hypochloric acid production (Levine, 1968). Subsequently, the increase in abomasal pH causes a failure of pepsinogen to convert to the active form, pepsin, which results in elevated plasma pepsinogen levels and reduced protein digestion. The severity of the infection depends on concurrent infections, nutritional state of the host and also the ability to develop an immunogenic response (Stear et al., 2003). Commonly, moderate or subclinical infections occur, causing diarrhoea, poor weight gain, weight loss and reduced wool production (Zajac, 2006).
3.2.3. Trichostrongylus species
Species of Trichostrongylus represent an important group of parasites of grazing small ruminants. These parasites occur in the small intestine and mainly exert their pathogenic effects in lambs and weaners, but have also been reported to cause a significant depression of wool growth in old animals (Donald et al., 1978). In Australia, the three most common species are Trichostrongylus colubriformis, Trichostrongylus vitrinus and Trichostrongylus rugatus (see Beveridge et al., 1989a). The main pathogenic effects are caused by the exsheathed L3s of T. vitrinus, which burrow between the intestinal villi and lead to the formation of sub-epithelial tunnels (Beveridge et al., 1989b). Young nematodes developing in these tunnels are released 10–12 days following infection. The liberation of young adults is associated with extensive damage to the duodenal mucosa and with signs of generalized enteritis, including haemorrhage, oedema and plasma protein loss into the intestinal lumen, and subsequent hypoalbuminaemia and hypoproteinaemia (Taylor et al., 2007). Infections with Trichostrongylus are often difficult to distinguish from malnutrition in the case of low-intensity infections (Taylor et al., 2007) but, if present at high numbers, cause protracted watery diarrhoea which stains the fleece of the hindquarters “black scours” (Levine, 1968). Trichostrongylus axei, which inhabits the abomasum, is less common and occurs usually in smaller numbers (Donald et al., 1978).
3.2.4. Other species of veterinary significance
Cooperia curticei, Nematodirus spathiger, N. fillicollis, Oesophagostomum venulosum are common parasites of the small and/or large intestine, whilst Chabertia ovina and the hookworm, Bunostomum trigonocephalum, are less common (Zajac, 2006). Individually, these species have relatively low pathogenicity, but may contribute to PGE in grazing small ruminants. Nematodirus battus is of particular pathogenic significance in some temperate areas, such as the British Isles, where the mass-hatching of infective L3s occurs during spring, causing disease of young lambs (Taylor and Thomas, 1986); however, this species of Nematodirus has not been reported in Australasia.
4. Some General Aspects of the Epidemiology of Gastrointestinal Nematodes of Livestock
In general terms, infectious diseases can be transmitted horizontally (from one animal to another) or vertically (from one generation to the next). Horizontal transmission can occur either directly or indirectly (Thrusfield, 2005). In the case of gastrointestinal nematodes infecting sheep, transmission occurs exclusively horizontally and directly and can be captured as a relationship among the parasite, host and environment (Fig. 4.2 ) (Levine, 1968). Various factors linked to this relationship determine the type and severity of infection, and many interactions occur among these components. Host-related factors include species, genotype, age, sex and immunity; parasite-related factors include life history, duration of the histotrophic phase, survival of larvae in the environment and their location in the host; environmental factors include climate, weather, season, type of vegetation and microclimate. The interactions between host and parasite mainly determine the potential for disease to occur and the pattern/course of infection, whereas the interaction between host–environment and parasite–environment influence disease transmission (Levine, 1968). In addition, the relationship among host (e.g. genotype), parasites and the host microbiome (particularly at the infection site) could play a key role in governing parasite burdens, disease progression and also disease transmission, and warrants detailed investigation.
Figure 4.2.
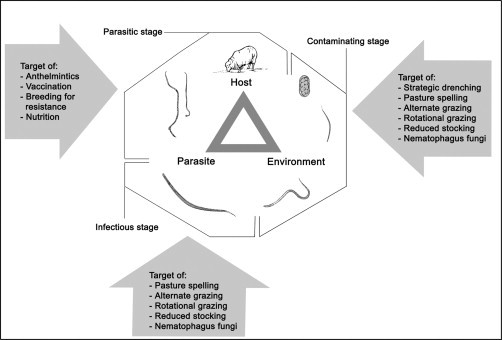
Relationship among host, parasites and environment, and factors that affect parasite control.
Adapted from Gordon (1948), Levine (1968) and Donald et al. (1978).
Regional differences in climate have major effects on the epidemiology of nematode infections and their geographical distribution (Beveridge et al., 1989a, Cole, 1986, De Chaneet and Dunsmore, 1988). Because species distributions and the seasonal availability of different parasites are largely determined by their ecological needs (e.g. for the successful development of their free-living stages), environmental conditions, in particular temperature and relative humidity, are of major importance (Beveridge et al., 1989a, O'Connor et al., 2006) (Table 4.2 ). Thus, climate impacts directly on the distribution of parasites. However, there are other exogenous factors, such as anthelmintic treatment regimens or host movement, which can influence the distribution and prevalence of these parasitic nematodes in a particular geographical environment (Blouin et al., 1995, Gordon, 1948).
Table 4.2.
Key features of major trichostrongylid nematodes of sheep and environmental influences on survival
| Nematode species | Life-cycle stage |
|||
|---|---|---|---|---|
| Unembryonated egg | Embryonated egg | Pre-infective larvae | Infective larvae | |
| H. contortus | High susceptibility to cold and desiccation. High mortality < 10 °C. |
Susceptible to cold and desiccation. Low hatching in the absence of moisture and/or < 10 °C. |
High susceptibility to cold and desiccation. | Optimum survival under warm and moist conditions. Poor survival in dry climates (warm or cool) and sub-freezing winter. |
| T. colubriformis | Intermediate susceptibility to cold and desiccation. High mortality < 5 °C. |
Intermediate susceptibility to cold. Low susceptibility to desiccation. | Susceptible to cold and desiccation. High mortality < 5 °C. |
Optimum survival under warm or cool moist conditions. Poor survival over sub-freezing winters. |
| Te. circumcincta | Low susceptibility to cold. Intermediate susceptibility to desiccation. High egg viability at 0–10 °C. |
Low susceptibility to cold and desiccation. Hatching < 5 °C. |
Intermediate susceptibility to cold. Susceptible to desiccation. |
Optimum survival under cool moist conditions and sub-freezing winters. Poor survival under warm, dry conditions. |
Adapted from O'Connor et al. (2006).
4.1. Distribution of trichostrongylids of sheep according to climate zone
H. contortus is regarded as the most important nematode species in tropical (between 23.5N° and 23.5S°) and subtropical climates (north of 23.5N° and south of 23.5S°), where usually warm temperatures and summer-dominant or uniform rainfall favour the development of this species (O'Connor et al., 2006). In cool, temperate zones (between latitudes 45° and 65°), conditions throughout most of the year are limiting for the development of H. contortus, because its free-living stages are susceptible to low temperatures and desiccation (O'Connor et al., 2006). In the United States, for example, the climate is most suitable for H. contortus in the south-eastern to north-eastern and mid-western states, whilst in the western part of the country cold winters and dry summers reduce the success of this species (see Zajac, 2006). Trichostrongylus and Teladorsagia are more dominant in winter and uniform rainfall zones, because of their resistance to desiccation and their ability to develop at lower environmental temperatures (Beveridge et al., 1989a, Donald et al., 1978, Young and Anderson, 1981). In Mediterranean regions (between latitudes 30° and 40°), cool and wet winters favour the development of free-living stages of Teladorsagia and Trichostrongylus, whilst hot and dry summer conditions cause a decrease in larval pasture contamination (O'Connor et al., 2006). For example, in western regions of the United States, arid summers reduce the transmission rate of most gastrointestinal nematodes, and PGE is usually less important than in the eastern part of the United States (Zajac, 2006). In temperate zones (> 40°), the cooler environment creates a perfect climate for Teladorsagia and Trichostrongylus, and peak infections occur during summer and autumn (O'Connor et al., 2006). In warmer temperate zones, such as in south-eastern Australia, where milder winters prevail, peak infections occur in late winter to early spring (Brunsdon, 1980). In the tropics and subtropics, these two genera may be prevalent, but extend to regions with cooler, humid early spring or late autumn conditions (O'Connor et al., 2006). The three most common intestinal species of Trichostrongylus, namely T. vitrinus, T. colubriformis and T. rugatus, may occur sympatrically, but usually one species predominates in a particular geographical region and/or season (De Chaneet and Dunsmore, 1988). T. vitrinus predominates in lambs in Britain (Parnell et al., 1954), northern Europe (Eysker, 1978) or in the winter rainfall environments of south-eastern Australia (Anderson, 1972). T. colubriformis is most prevalent in North America (Levine, 1968), East and South Africa (Grant, 1981) and in the summer rainfall zones of eastern Australia (Southcott et al., 1976). In uniform rainfall zones of eastern Australia and in New Zealand, T. colubriformis and T. vitrinus occur together, but which species predominates depends on the season (Brunsdon, 1970, Waller et al., 1981). T. rugatus is most common in arid climates of South Africa or the inland of Australia (De Chaneet and Dunsmore, 1988).
5. The Control of Gastrointestinal Nematodes and Anthelmintic Resistance (AR)
Various approaches are used for the control of gastrointestinal helminths of livestock (Fig. 4.2). These are based on three basic principles (Hoste and Torres-Acosta, 2011):
-
i.
The first principle is to reduce the exposure of the host to the infective stages (L3s), mainly achieved by strategies of grazing management (e.g. rotational grazing, alternate grazing, pasture-spelling and/or reduced stocking rates).
-
ii.
The second principle targets the development of a more favourable response of the host to gastrointestinal parasite infection (e.g. achieved through vaccination, improved nutrition, genetic selection of hosts and breeding for resistance).
-
iii.
The third principle directly targets the elimination of worms from their hosts through the administration of conventional (synthetic formulations) or non-conventional (plant or mineral) anthelmintic compounds.
Following the introduction of phenothiazines in the 1950's, the control of gastrointestinal parasites has been achieved using chemical anthelmintics (Hoste and Torres-Acosta, 2011) and still predominantly relies on the treatment with broad-spectrum parasiticides, including the benzimidazoles (BZs), macrocyclic lactones (MLs) and imidazothiazoles/tetrahydropyrimidines (LVs) (Besier and Love, 2003) (Table 4.3 ). Although there has been a recent breakthrough with the development of a new drug, monepantel, from an alternative compound class (amino-acetonitrile derivatives) (Kaminsky et al., 2008), success in the discovery of new anthelmintics, more generally, has been extremely limited over the past two decades (Kaplan, 2004).
Table 4.3.
Main anthelmintics used for the treatment of nematode infections in livestock; their mode of action (if known) and proposed mechanisms of resistance
| Anthelmintics | Understood mode of action | Proposed mechanisms of resistance | References |
|---|---|---|---|
| Benzimidazoles | Bind to β-tubulin and prevent the formation of microtubules. Causes the inhibition of glucose uptake, protein secretion and microtubule production, leading to starvation of the parasite. | Mutations in the β-tubulin gene causing structural changes in β-tubulin. As a consequence the drug can no longer bind to its target site. |
Lacey (1988) Winterrowd et al. (2003) |
| Imidazothiazoles/tetrahydropyrimidines | Mimic the action of acetylcholine causing spastic paralysis of the worms. Paralysed worms are expelled by normal gut peristalsis leading to rapid removal of present worms. |
Poorly understood, possible involvement of structural changes in the nicotinic acetylcholine receptor preventing the binding of the drug. Also proposed have been changes in the sensitivity of the receptor towards acetylcholine which can lead to a cross-resistance with organophosphates. |
Richmond and Jorgensen (1999) Robertson et al. (1999) Martin et al. (2003) Martin and Robertson (2007) |
| Macrocyclic lactones (avermectins/milbemycins) | Causes an opening of glutamate-gated chloride channels (GluCl). This leads to an increased C1− ion influx into nerve cell causing flaccid paralysis of the worm. | Poorly understood, possible involvement of: Mutations in P-glycoprotein gene could cause a gain-of-function leading to a more rapid removal of the drug from the parasite. Selection at glutamate- and y-aminobutyric-acid-gated chloride channels. |
Blackhall et al. (1998) Blackhall et al. (2003) Kerboeuf et al. (2003) |
| Amino-acetonitrile derivatives | The hypothesized mode of action involves a nematode-specific clade of acetylcholine receptor subunits. | Full or partial loss of the gene which encodes the particular type of acetylcholine receptor. | Kaminsky et al. (2008) |
| Cycloocta-depsipeptides (emodepside/PFIO22A) | Binds to a presynaptic latrophilin receptor in nematodes. |
Harder et al. (2003) Harder et al. (2005) von Samson-Himmelstjerna et al. (2005) |
|
The frequent and often uncontrolled use of these drugs has led to widespread problems with AR in parasites of livestock (Taylor, M.A., Learmount, J., Lunn, E., Morgan, C., Craig, B.H., 2009). AR in parasites of veterinary importance has emerged as a major bionomic and economic problem worldwide, being currently most severe in parasitic nematodes of small ruminants (von Samson-Himmelstjerna, 2006, Waller, 1994, Waller, 1997). For instance, in Australia, it has been proposed that the prevalence and extent of resistance to all major classes of broad-spectrum anthelmintics is so widespread that it threatens the profitability of the entire sheep industry (Besier and Love, 2003). However, Larsen et al. (2006) showed that, despite widespread problems of drug resistance, farm productivity can be maintained or increased if sound farm management practices are put in place, emphasizing the need for integrated approaches of worm control.
Although there is hope for new, effective anthelmintics, there is also a major need to preserve those that we currently have at our disposal. Hence, monitoring the drug-susceptibility and -resistance status of strongylid nematode populations in livestock must be a high priority and should be an important component of integrated management strategies. Various methods, such as faecal egg count reduction test (FECRT) and egg hatch and larval development assays, have been used for estimating levels of drug-susceptibility/resistance in strongylid nematodes of small ruminants, cattle and horses (Coles et al., 1992) (Table 4.4 ).
Table 4.4.
Summary of in vivo and in vitro tests currently used for the diagnosis of anthelmintic resistance in gastrointestinal nematodes of livestock, and their principles and limitations
| Assay | Principle | Comments and existing limitations | References | |
|---|---|---|---|---|
| In vivo | Faecal egg count reduction test | Provides an estimate of anthelmintic efficacy by comparing faecal egg counts from sheep before and after treatment. Resistance is declared if reduction in the number of eggs counted is < 95% and the lower confidence interval for the percentage of reduction is below 90%. |
|
Martin et al. (1985) Presidente (1985) Coles et al. (1992) Jackson (1993) Grimshaw et al. (1996) McKenna, 1996, McKenna, 1997, McKenna, 2006 Taylor et al. (2002) Coles et al. (2006) Miller et al. (2006) Dobson et al. (2009) Levecke et al. (2011) |
| Controlled test | Involves the infection of worm free sheep. Infective larvae (of susceptible and resistant strains) are inoculated together with the tested anthelmintic at 0.5, 1 and 2 times the recommended dose. Resistance is declared if reduction in geometric mean worm counts is less than 90% or greater than 1000 worms surviving treatment. |
|
Presidente (1985) Taylor et al. (2002) |
|
| In vitro | Egg hatch test | Known numbers of undeveloped eggs are incubated in serial dilutions of the anthelmintic. The percentage of eggs that hatch at each concentration is then determined and the dose response calculated for the different concentrations of the drug tested (calculated as ED50 = the concentration of drug required to kill 50% of eggs). |
|
Le Jambre (1976) Boersema (1983) Weston et al. (1984) Borgsteede and Couwenberg (1987) Kerboeuf and Hubert (1987) Martin et al. (1989) Scott et al. (1989) von Samson-Himmelstjerna et al. (2009) |
| Larval paralysis test | Infective third-stage larvae are incubated for 24 h in serial dilutions of the anthelmintic. After 24 h the percentage of paralysed larvae is determined at each concentration of the drug and a dose response line is plotted and compared to known reference strains. |
|
Martin and Le Jambre (1979) Boersema (1983) Geerts et al. (1989) Sutherland and Lee (1990) |
|
| Larval motility test | Measures the motility of larval and adult nematodes after incubation together with dilutions of anthelmintics. A motility index is then calculated by a computer. |
|
Bennett and Pax, 1986, Bennett and Pax, 1987 Coles et al. (1989) Gill et al. (1991) |
|
| Migration test | Adult worms are used to differentiate between susceptible and resistant nematode strains. Worms are incubated for 30 min in anthelmintic dilutions and then transferred to migration chambers where their ability to migrate through a polyamide net is evaluated. A dose response is plotted based on the inhibited migratory capacity. |
|
Petersen et al., 1997, Petersen et al., 2000 | |
| Larval development test | Nematode eggs are cultured to the third-larval stage in the presence of a food source (E. coli or yeast extract and the anthelmintic to be tested. Larvae affected by the anthelmintic fail to develop to the third-stage larvae, expressed as LD50 (= the concentration of the drug required to kill 50% of larvae). |
|
Coles et al. (1988) Giordano et al. (1988) Taylor (1990) Gill et al. (1995) Gill and Lacey (1998) Demeler et al., 2010a, Demeler et al., 2010b |
|
Most recent advances in the diagnosis of AR have focused on the implementation of a standardized protocol for the egg hatch test (von Samson-Himmelstjerna et al., 2009) and the development and standardization of a larval migration inhibition test (Demeler et al., 2010a). However, many of these assays can be very time consuming to carry out, suffer from a lack of reliability and sensitivity as well as reproducibility of results (Taylor et al., 2002). Therefore, novel approaches of AR diagnosis are needed.
Molecular methods have been proposed to provide new alternatives to the most commonly applied in vivo and in vitro techniques for the diagnosis of AR and might be capable of overcoming some of their limitations (Beech et al., 2011, Demeler et al., 2010a, Demeler et al., 2010b, von Samson-Himmelstjerna, 2006). Critical to the development of molecular-diagnostic assays for AR is the in-depth knowledge of the mode of action of these chemicals, their target sites and mutations in the genome of parasitic helminths linked to a reduced susceptibility to these drugs (Beech et al., 2011, von Samson-Himmelstjerna, 2006, Wolstenholme et al., 2004). However, although the molecular basis of mechanisms of AR is currently best understood for BZ anthelmintics, concise information for other classes of broad-spectrum anthelmintics is far less elucidated (Taylor et al., 2002). In the case of BZ, a single-nucleotide polymorphism (SNP) at codon 200 of the β-tubulin isotype 1 is currently believed to be most closely linked to BZ resistance (Wolstenholme et al., 2004) and has been demonstrated in resistant strains of H. contortus (see Geary et al., 1992), T. colubriformis (see Silvestre and Humbert, 2002) and Te. circumcincta (see Elard and Humbert, 1999) in sheep. At least two more SNPs at position 167 and 198 have been identified, but appear to be less common amongst different species of trichostrongylid nematodes (Beech et al., 2011, Wolstenholme et al., 2004). Besides the sequence changes in β-tubulin, which are currently believed to be the major cause of BZ resistance, more recent investigations have also suggested a link to the drug transporter P-glycoprotein, which plays a role in the transport of the anthelmintic away from its site of action and may also select for resistance to MLs (Beech et al., 2011).
Based on the current knowledge of the mechanism of AR to BZ anthelmintics, conventional, allele-specific PCRs were developed to determine the genotype of adult worms of H. contortus (see Kwa et al., 1994) and Te. circumcincta (see Elard and Humbert, 1999). This work was further extended by Silvestre and Humbert (2000) by combining the previously described PCR assays with a RFLP procedure, which allowed the phenetic characterization and identification of L3s of H. contortus, T. colubriformis and Te. circumcincta. Alvarez-Sanchez et al. (2005) designed a real-time PCR (RT-PCR) method to determine the allele frequency of β-tubulin isotype 1 (codon 200) in nematode DNA samples. As stated by the authors, the diagnosis of BZ resistance using this assay showed a significant agreement with phenotypic tests, such as the egg hatch test and the faecal egg count reduction test, and allowed the analysis of allele frequencies in DNA samples from pooled larvae (von Samson-Himmelstjerna, 2006).
Despite these developments, there has been no assessment of the suitability of these assays for applications to field samples containing mixed species of gastrointestinal parasites, which limits their practical utility at this stage. Furthermore, all currently employed molecular assays involve adult nematodes (only available through necropsy of the host) or infective L3s (requires culturing of eggs for 7–10 days), but none of the existing assays has yet been assessed for the detection of AR directly from (mixed populations of) eggs, which would significantly reduce the time required for diagnosis. In contrast to the BZs, the molecular mechanisms associated with resistance to LEV and ML anthelmintics are not yet understood in detail, and recent research has suggested that in both cases multiple genes (Beech et al., 2011) are involved in resistance and that resistance is often the result of changes in the parasite other than the immediate drug target, including transporters and drug metabolism (Cvilink et al., 2009). Consequently, the polygenic nature of resistance to these anthelmintics and the lack of reliable or universal markers represent a major obstacle to the development of molecular-diagnostic tools for AR. No molecular test is yet available for these two groups of broad-spectrum anthelmintics.
6. Traditional Methods for the Diagnosis of Strongylid Nematode Infections and Their Limitations
Different species of nematodes can be variable in their pathogenicity, seasonal and geographical distribution and susceptibility towards different anthelmintics. Therefore, the accurate diagnosis of infections and the detailed knowledge of which parasites are present in a particular geographical environment are of major importance. Such information directly supports parasite control strategies and is of relevance for investigations into parasite biology, ecology and epidemiology.
6.1. Validation of diagnostic tests
The validation of a diagnostic test involves the determination of basic parameters (Table 4.5 ) and can be achieved in five steps (Table 4.6 ) (OIE, 2004). As a first step, a test suitable for a particular use has to be selected, developed and optimized. Subsequently, validation parameters have to be determined, such as analytical sensitivity and analytical specificity (Conraths and Schares, 2006). Following this initial assessment, the diagnostic sensitivity and specificity are determined by examining a larger number of samples for which the true disease or infection status of the animals being tested is known (determined by a ‘gold standard’). After a test has been evaluated, it may be considered validated (Conraths and Schares, 2006). However, a continuous monitoring of test performance during routine application is advisable.
Table 4.5.
Important validation parameters used in the assessment of diagnostic tests (Conraths and Schares, 2006, Pfeiffer, 2010, Thrusfield, 2005)
| Term | Definition | Method of assessment |
|---|---|---|
| Sensitivity | The proportion of animals with the disease and which test positive. | Assessment of these two parameters requires an independent, valid criterion termed a ‘gold standard’ used to define the true disease status of an animal. |
| Specificity | The proportion of animals without the disease and which test negative. | |
| Agreement | The agreement in results between two diagnostic tests, with one of the tests being a generally accepted diagnostic method. | Frequently assessed by the Kappa test, which measures the proportion of agreement beyond that to be expected by chance. |
| Accuracy | Refers to the concordance between test results and the ‘true’ clinical state. | Depends on the number of ‘false-positives’ and ‘false-negatives’, in comparison with the true infection state as determined by the ‘gold standard’. |
| Reliability | The extent to which test results are consistent in repeat experiments. | This includes the assessment of repeatability, reproducibility, inter- and intra-assay variability. Repeatability assessment can be done by running the test two or more times on the same samples in the same laboratory under the same conditions. Additionally, the intra-assay variability (between replicates within the same run) and inter-assay variability (replicates between different runs) can be assessed. |
| Reproducibility can be assessed in the same manner as described before, and assessed between different laboratories. | ||
Table 4.6.
Stages of test validation
| Stages of test validation |
|---|
|
Adapted from Conraths and Schares (2006).
6.2. Diagnosis in the live animal
Various methods are employed for the ante mortem diagnosis of gastrointestinal nematode infections. The most commonly employed methods are based on the observation of clinical signs indicative of disease, microscopic examination of faeces from infected hosts, biochemical and/or serological diagnostic approaches. However, widespread standardization of many laboratory techniques, including FECs or LCs, does not existent, and most diagnostic, research and teaching facilities apply their own modifications to published protocols (Kassai, 1999). Although these techniques are regarded to be standard diagnostic procedures, there is a lack of detailed studies of diagnostic performance, including the diagnostic sensitivity, specificity and/or repeatability.
6.2.1. Diagnosis based on clinical signs of disease
Disease caused by gastrointestinal nematodes manifests itself in a variety of clinical signs, including scouring, anaemia, loss of body condition and in severe cases death (Hungerford, 1990). The type and degree of clinical manifestation are also influenced by factors, such as the species of worms involved, the present worm burden, the plane of nutrition and reproductive/immunological status of the host animal (Hungerford, 1990, Levine, 1968). To aid the diagnosis of gastrointestinal nematode infections, a number of approaches have been developed for the interpretation of clinical signs linked to PGE. These approaches include, for example, body condition- (Russel et al., 1969), ‘dag’- (Larsen et al., 1994) or anaemia-scoring (van Wyk and Bath, 2002). Although useful as indicators for PGE, with applicability on a farm level, these approaches are subjective and lack specificity, as clinical signs can relate to an extremely wide range of diseases and problems affecting the host (van Wyk and Bath, 2002).
6.2.2. Faecal egg counts
The enumeration of eggs in faeces is the most common method for the diagnosis of gastrointestinal nematode infections and is routinely employed in parasitology. The technique is inexpensive, easy to perform and does not require specialized instrumentation, which makes it suitable for use in most diagnostic settings and countries. Important applications of this method include estimating infection intensity (McKenna and Simpson, 1987, McKenna, 1987), determining the degree of contamination with helminth eggs (Gordon, 1967), assessing the effectiveness of anthelmintics (Waller et al., 1989), determining the breeding value of an animal when selecting for worm resistance (Woolaston, 1992), and guiding control and treatment decisions (Brightling, 1988).
This method involves mixing faeces with a high-density solution (e.g. saturated sucrose, sodium chloride or sodium nitrate) to float parasite eggs (with the exception of trematode eggs) on the surface of the suspension. An aliquot of this suspension is withdrawn to count the eggs present and to then extrapolate the number to the volume of faeces used (in eggs per gram, EPG). Different methods have been developed for this purpose, such as the direct centrifugal flotation method (Lane, 1922), the Stoll dilution technique (Stoll, 1923), the McMaster method (Gordon and Whitlock, 1939) and the Wisconsin flotation method (Cox and Todd, 1962), of which the McMaster method is most widely used (Nicholls and Obendorf, 1994). For decades, numerous modifications of these methods have been described (Levine et al., 1960, Raynaud, 1970, Roberts and O'Sullivan, 1950, Whitlock, 1948), and most teaching and research institutions apply their own modifications to existing protocols (Kassai, 1999). Many of these modifications make use of different flotation solutions, sample dilutions and counting procedures, which achieve varying sensitivities and can complicate the comparison of FECs between laboratories. These inconsistencies make a pooled data analysis or retrospective studies of results between laboratories virtually impossible. The choice of flotation solution, sample dilution and volume examined can have a significant impact on the results obtained using this approach (Cringoli et al., 2004); similarly, biological factors can also limit the interpretation of these results, as described in the following.
Fecundity. The biotic potential of different species of trichostrongylid nematodes varies (Gordon, 1981), and parasite density and immune-mediated ‘control’ by the host have been shown to influence the egg production of female worms in different species (Rowe et al., 2008, Stear and Bishop, 1999). The diagnostic value of FECs to estimate worm burdens for the highly fecund nematode species, H. contortus, has been shown (Le Jambre et al., 1971, Roberts and Swan, 1981). However, the correlation between FECs and worm burdens is much lower for genera with a low fecundity, such as species of Teladorsagia (Ostertagia) (Martin et al., 1985), Trichostrongylus (Sangster et al., 1979) and Nematodirus (Martin et al., 1985, McKenna, 1981).
Water content of faeces. Samples intended for faecal analysis can be of varying consistencies, being soft to watery (diarrhoeic) or hard and desiccated (mostly from animals following transport and without access to food or water) (Gordon, 1953, Gordon, 1981). These aspects are of importance, as the water content of the sample can either dilute or concentrate the numbers of eggs determined from 1 g of faeces, depending on the actual amount of dry matter (Le Jambre et al., 2007).
Storage of faeces. For practical reasons, faecal material requires proper storage prior to coprological examination. Storage conditions are of importance because they can cause a reduction in egg numbers. An artefactual reduction in FECs occurs primarily due to hatching of eggs or biological degradation (Nielsen et al., 2010). To circumvent this problem, different strategies, such as chemical preservation (Whitlock, 1943), airtight storage (Rinaldi et al., 2011) or refrigeration (Nielsen et al., 2010), have been recommended. Additional considerations are that FECs (i) only reflect patent but not pre-patent infections (Thienpont et al., 1986), (ii) do not provide any information regarding male or immature worms present (McKenna, 1981) and (iii) can be influenced by variation in times of egg excretion by adult worms (Villanua et al., 2006), age of the worm population and/or the immunity, age and sex of the host (Thienpont et al., 1986). Although there are morphological differences between the eggs of socio-economically important nematodes (Georgi and McCulloch, 1989), specific identification cannot be achieved by routine microscopy (with few exceptions, such as Nematodirus spp.) (Lichtenfels et al., 1997).
Therefore, FECs alone should not be used to make a diagnosis or guide treatment decisions, but should be interpreted in conjunction with information about the nutritional status, age and management of sheep in a flock (McKenna, 2002). However, according to common practice, an FEC of ≥ 200 EPG is regarded to indicate a significant worm burden and is used as basis for the decision for anthelmintic treatment (www.wormboss.com.au). The value of FEC results also depends on the hosts and/or parasite species involved. For example, FEC results for adult cattle are of limited diagnostic value, as they do not usually correlate with worm burden (McKenna, 1981); FECs in cattle are usually low and require more sensitive flotation techniques than for sheep (Mes et al., 2001); for species of Nematodirus, egg counts are also regarded to be of limited value, as most damage is caused by the immature stages before egg-laying commences (McKenna, 1981). In addition, the low sensitivity of FEC techniques, being in the range of 10–50 EPG for the McMaster technique, represents a limitation in relation to the diagnosis of AR by FECRT (Levecke et al., 2012).
Although it is unlikely that some of the current limitations of FEC will be resolved in the near future (i.e. those which relate to the fecundity of the nematodes), some recent developments have been made towards improving the procedure. Attempts have been made by the World Association for the Advancement of Veterinary Parasitology (WAAVP) to implement FEC protocols for the assessment of AR in different species of animals (Coles et al., 2006). Lectin staining for the identification of H. contortus eggs (Palmer and McCombe, 1996), computerized image recognition of strongylid eggs (Sommer, 1996) and automated egg counting (Mes et al., 2007) are interesting developments towards improved species identification and differentiation. However, the suitability of the latter two approaches needs rigorous assessment for routine applications because of their technical complexity. With the development of FECPAK, a diagnostic test-kit containing the necessary equipment for coproscopic examination (www.techiongroup.co.nz), efforts have been made to enable sheep farmers to carry out FECs themselves. However, the implementation of such a system requires a significant level of cooperation by farmers, adequate training and integrated quality assurance to ensure that correct diagnoses are made (McCoy et al. 2005). Despite the provision of training courses for farmers, many difficulties in the differentiation of structures observed during coproscopy were encountered for inadequately trained farm employees, limiting the value of the results obtained (McCoy et al., 2005). The development of the FLOTAC egg counting method (Cringoli et al., 2010), seems to be promising. Once validated for different host and parasite species, this method may deliver FECs with increased sensitivity (i.e. 1 EPG) and might represent an alternative to current flotation techniques.
6.2.3. Larval culture
To aid the identification of different nematode genera present in mixed infections, faecal culture methods have been developed to identify first-stage (L1) or infective L3s of strongylid nematodes. The most commonly used methods involve the incubation of faecal material, in order to produce infective L3s for subsequent morphological differentiation. Currently, a number of protocols have been published which differ in the temperatures, times and media used for culture and the approach of larval recovery (Dinaburg, 1942, Hubert and Kerboeuf, 1984, MAFF, 1986, Roberts and O'Sullivan, 1950, Whitlock, 1956). Probably, the most widely employed protocol suggests incubation at 27 °C for 7 days (MAFF, 1986).
However, studies investigating the ecology and developmental requirements of various species of gastrointestinal nematodes infecting livestock (Beveridge et al., 1989a, O'Connor et al., 2006) have shown that different species of strongylid nematodes require different conditions, such as environmental temperature and relative humidity, to enable adequate development. This is particularly important to consider when LC results are used to estimate the contribution of different species to mixed infections. One culture protocol is likely to favour the development of one species over others (Dobson et al., 1992). For instance, Whitlock (1956) observed that culture conditions (27 °C for 7 days) are suitable for most species, but that the free-living stages of Teladorsagia (Ostertagia) species develop better at somewhat lower temperatures. This statement was supported by the findings of Dobson et al. (1992) who demonstrated that the developmental success of the infective larvae in faecal cultures was lower for Te. circumcincta than for T. colubriformis when cultured alone or concurrently, thus indicating that LCs are unreliable for estimating the contribution of individual species in mixed infections. Similar observations were made by Berrie et al. (1988) for the bovine parasites Haemochus placei, Oesophagostomum radiatum and Cooperia pectinata. In this study, these authors observed that the recovery of larvae of H. placei was significantly lower compared with the other two species under the same culture conditions. Based on their findings, the authors stated that faecal culture and subsequent larval differentiation are unsuitable for an accurate estimation of the proportions of individual species in mixed infections and can only be used to provide an indication of the species present (Berrie et al., 1988).
Further variability in the results obtained from LCs have been attributed to differences in the composition of the culture medium used, which influences the moisture, oxygen availability and/or pH that larvae encounter during their development (Hubert and Kerboeuf, 1984, Roberts and O'Sullivan, 1950). Therefore, it had been suggested that providing a better-defined medium than faeces might help to obtain more consistent results (Hubert and Kerboeuf, 1984). To further examine this hypothesis, Hubert and Kerboeuf (1984) developed a modified method of LC using an ‘on-agar’ approach to provide well-defined and standardized conditions. Their results showed that the culture on agar medium led to higher recoveries of larvae compared with traditional faecal cultures. However, lengthy preparation times and increased laboratory requirements appear to limit the routine application of this method.
In addition to the variability of results related to the culture conditions employed, the specific identification of cultured larvae provides challenges (Fig. 4.3 ). For the identification of infective L3s to the species or genus level, a number of different approaches have been described. A commonly employed method for species differentiation involves the detection of particular morphological features of the larvae (e.g. the length of the tail sheath extension and total body length of L3s) and their comparison with published identification keys (Dikmans and Andrews, 1933, Gordon, 1933, MAFF, 1986, McMurtry et al., 2000, van Wyk et al., 2004). Various keys for the identification of L3s have been published (Dikmans and Andrews, 1933, Gordon, 1933, MAFF, 1986), and there is a substantial overlap between the body length measurements of different species, and a substantial variability in the length of L3s has been reported by different authors (McMurtry et al., 2000).
Figure 4.3.
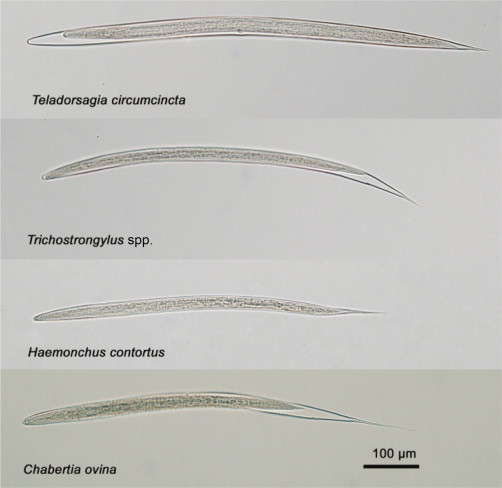
Third-stage larvae (L3s) of key species of gastrointestinal nematodes of sheep, encountered following larval culture (LC).
Van Wyk et al. (2004) developed a simplified approach which uses the mean length of the tail sheath extension to differentiate L3s of Teladorsagia and/or Trichostrongylus from the larvae of Haemonchus and Chabertia and/or Oesophagostomum. However, although useful for a differentiation of genera, without the requirement to measure every single larva (and thus being more time efficient), this approach has the disadvantage that it does not allow the unequivocal differentiation of all genera. For instance, Teladorsagia and Trichostrongylus (being the most common genera in winter rainfall areas) cannot be differentiated based on sheath extension length alone. To further refine their differentiation, additional morphological features are required. Lancaster and Hong (1987) proposed the presence of an inflexion (‘shoulder’) at the cranial extremity of Teladorsagia larvae as an informative morphological feature. However, this feature is very subtle and its detection is subjective.
Another approach to differentiate L3s of Teladorsagia from those of Trichostrongylus was proposed by Gordon (1933); it is based on the body length measurements of the larvae. According to this author, and based on the measurements of 1000 larvae of each genus, a body length ‘cut-off’ value of 720 μm allowed reliable differentiation of these genera, with Trichostrongylus larvae being ≤ 720 μm and those of Teladorsagia being > 720 μm. Although, practical, this technique requires the measurement of individual larvae and does not take into account variability in the length of developing larvae (e.g. influenced by culture conditions employed, climate/season in a particular environment, availability of appropriate food for developing L1s and L2s and/or immune status of the host) (McMurtry et al., 2000).
A third approach for the differentiation of Teladorsagia from Trichostrongylus L3s was described by McMurtry et al. (2000). The latter approach involves the treatment of cultured larvae with sodium hypochlorite to exsheath the larvae and count tubercles at the posterior end of the exsheathed L3. As claimed by the authors, this approach allows the differentiation among populations of T. axei, T. colubriformis, T. vitrinus and Te. circumcincta. However, the authors acknowledge that there is a degree of variability in the number of observed tubercles and that the tails of Te. circumcincta and T. axei lack these structures (McMurtry et al., 2000). Although being regarded as economically ‘less important’ parasites, the differentiation of L3s of Oesophagostomum and Chabertia, which infect the large intestine, is not considered possible using current techniques for larval differentiation, which makes this approach unsuitable if detailed epidemiological information on distribution and prevalence of genera and species is required. A less commonly used method for larval differentiation involves the culture and morphological identification of L1s (Whitlock, 1959). This technique has the advantage of being rapid, since the time required for the development of the L1 stage is shorter; however, the same limitations for the culture and identification of L3s exist for L1s and L2s (Lichtenfels et al., 1997).
In conclusion, although regarded as a standard diagnostic procedure, the technique of LC coupled to larval differentiation has the disadvantage of being time consuming and laborious, cannot readily be automated, and the morphological identification/differentiation, based on available publications, requires substantial expertise and training (Johnson et al., 1996, Lichtenfels et al., 1997).
6.2.4. Biochemical and immunological methods
In addition to commonly used coprodiagnostic methods, a number of biochemical and immunological approaches have been developed that aim at the specific diagnosis of infection. These methods are mainly based on the detection and measurement of parameters (e.g. detection of elevated serum pepsinogen and gastrin levels or circulating antibodies) that might be indicative of parasitic infections.
6.2.5. Pepsinogen and gastrin
Chief cells of the gastric fundus produce the pro-enzyme pepsinogen, which is converted to its active form by acid produced by parietal cells. When parasitized glands of the gastric mucosa are destroyed, the hydrochloric acid production of parietal cells decreases, causing a rise in abomasal pH and resulting in a failure to convert pepsinogen to active pepsin (Levine, 1968). Accumulating pepsinogen can escape between disrupted cell junctions into the blood. Therefore, an increase in serum pepsinogen concentration has been regarded to relate to mucosal damage by developing larval stages of Ostertagia (Levine, 1968). Berghen et al. (1993) reviewed the value and application of pepsinogen, gastrin and antibody responses as diagnostic indicators for ostertagiasis and identified a number of potentially limiting factors. The authors suggested that other parasitic or non-parasitic diseases can be responsible for a moderate rise in pepsinogen concentrations in blood, thus limiting the specificity of this approach.
Gastrin is a hormone produced by G-cells in the stomach. Gastrin stimulates parietal cells to secrete acid and also stimulates pepsinogen secretion, stomach motility and blood circulation in gastric vessels. It was suggested that strongylid nematodes can directly stimulate G-cells, causing an increased gastrin production (Berghen et al., 1993). However, as demonstrated for pepsinogen, the specificity of this approach was questioned (Berghen et al., 1993), because other parasites or factors, such as diet, lactation or abomasal lesions, can also affect gastrin levels. Furthermore, in an experimental context, it has been shown that high infective doses need to be administered to parasite-naïve calves to provoke a significant gastrin release (Berghen et al., 1993).
6.2.6. Immunological methods of diagnosis
A wide range of immunological methods, including those that are based on the detection of an immune response in an infected animals and those for the detection of parasite antigens, have been developed for the specific diagnosis of parasitic infections (e.g. Engvall and Ruitenberg, 1974, Fletcher, 1965, Ogunremi et al., 2008). Based on the target molecule (antigen or antibody), such immunological methods can be classified as being either direct or indirect.
Direct immunological methods are those that provide direct evidence of an infection and can be based on the detection of parasite antigens present in the circulation and/or excreta from infected hosts. Parasitic extracts have a complex composition and contain molecules which are sometimes shared by other parasites (Cohen and Sadun, 1976). Shared antigenic composition of closely related parasite species is a challenge, particularly for nematodes, and often leads to cross-reactivity in immunological tests (Eysker and Ploeger, 2000, Noordin et al., 2005). Also the presence of host materials associated with the parasite can complicate antigen purification and can interfere with the specificity of a diagnostic assay. Furthermore, the life-cycle stage of the parasite used as an antigen source can influence immuno-diagnostic results, as parasites undergo significant structural and biochemical changes during their development (Cohen and Sadun, 1976). As an example, the antigenic composition of larval stages differs from that of adults (Williams and Soulsby, 1970) and can give rise to variations in diagnostic sensitivity and specificity (McLaren et al., 1978).
Although limited research has been undertaken, to date, for livestock parasites, Johnson et al. (1996) described an immuno-diagnostic assay for the quantitative detection of excretory/secretory parasite antigens in host faeces (coproantigens). These authors evaluated the usefulness of this approach in a murine model system using Heligmosomoides polygyrus, a trichostrongyloid gastrointestinal nematode related to the common nematode species infecting ruminants. The authors also suggested that the enzyme-linked immunosorbent assay (ELISA) was useful to specifically detect parasite antigens in the host faeces and might have potential to detect pre-patent infections and make large scale, rapid screening possible. The diagnostic performance of the assay was promising under experimental conditions, but cross-reactivity, faecal components interfering with the reactivity and the loss of antigens in faeces have been reported (Johnson et al., 1996).
Indirect immunological methods are usually based on the detection of anti-parasite antibodies or cell-mediated immune responses in infected hosts. A variety of methods have been developed and applied to the diagnosis of nematode infections, such as the complement fixation test, indirect immunofluorescence, indirect haemagglutination and ELISA, of which the latter has been most commonly used (Doenhoff et al., 2004). However, parasitic helminths possess a huge variety of antigens, and there is limited information on which stages and antigens are actually responsible for eliciting immune responses (Berghen et al., 1993). Antibody detection from serum has several disadvantages, including that it cannot distinguish between a current and past infection, which is a major challenge when evaluating the effects of chemotherapy, does often not reflect infection intensity and sometimes achieves poor specificity, particularly in disease-endemic areas (Doenhoff et al., 2004).
The detection of anti-Ostertagia antibodies in the serum of cattle has been found to be useful for epidemiological and cross-sectional studies, but only of limited utility for diagnosis on an individual animal basis (Berghen et al., 1993). Although anti-Ostertagia antibodies are detectable in milk samples by ELISA, there are also some limitations to this approach (Charlier et al., 2010). The response to parasitic infections is variable among host individuals, and it has been shown that serum antibody levels can be influenced by factors, such as milk yield, season, mastitis, the number of pregnancies of a cow, stage of lactation and genetic constitution (Gasbarre et al., 1993, Kloosterman et al., 1993, Sanchez et al., 2004). Also the use of bulk milk samples has been investigated, which has the advantage of being an inexpensive and user-friendly approach (Charlier et al., 2010). However, bulk milk samples taken only a few weeks apart can show significant variation in test results, depending on calving patterns, number of cows contributing to the milk in a tank (i.e. dilution effect) and their relative milk yields (Pritchard, 2001).
6.3. Post-mortem diagnosis
The post-mortem diagnosis of infections is routinely employed in parasitology to determine the number of nematode parasites present in the gastrointestinal tract, for epidemiological studies or to assess anthelmintic efficacy. Generally, these techniques involve the opening and washing of the respective parts of the GI tract and the examination of subsamples of the washes to estimate the total numbers of nematodes present. A number of different techniques have been described (Eysker and Kooyman, 1993, MAFF, 1986, Robertson and Elliott, 1966). The differences among these techniques exist in the counting of nematode worms from either the chyme or digesta and/or the washes (separately or combined); the soaking or not of the organ in water or saline (mainly used to recover immature stages); and the proportion of the total volume and the number of aliquots examined (reviewed by Gaba et al., 2006). Other differences exist in the length of the intestinal section examined (proximal 10 m of small intestine vs. the entire length) and the mesh size of the sieve used to remove plant debris from the washes (McKenna, 2008). Most necropsy techniques for the estimation of the intensity of infection involve the counting of worms from pooled gut contents and washes.
Eysker and Kooyman (1993) described a method that involves three components (contents, immediate water wash of the organ and the saline wash after 5 h of soaking the organ). The disadvantage of this method is that it involves more labour at necropsy, but it has the distinct advantage that worms are separated from the bulk of the gut contents, allowing a more rapid worm count in the laboratory. Gaba et al. (2006) assessed their approach for H. contortus and Te. circumcincta and suggested that the estimation of infection intensity, based on gut washes alone, is reliable. However, a prerequisite is that abomasa are processed rapidly (within 15 min) after the death of the sheep, as worms progressively start migrating into the contents (Gaba et al., 2006). Gaba et al. (2006) also stated that immediate washing of the gut is insufficient for extracting T. axei or larvae from the mucosa and, therefore, may distort the results if necropsy is carried out for the purpose of assessing trichostrongylid diversity or to examine the gut for the presence of immature larval stages.
Similarly, the choice of mesh size of the sieves used is dictated by the purpose of the worm counting procedure, as the use of a smaller mesh size enables a higher recovery of early L4s, but has the disadvantage that more debris is held back in the subsample examined, resulting in a prolonged time for counting (McKenna, 2008). Therefore, a small sieve size (e.g. 38 μm aperture) is only necessary if information about early L4s is required, whereas, if studies are carried out to confirm AR (reflected by a reduced efficacy against adult worms), also larger mesh sizes (e.g. 250 μm aperture) can be used (McKenna, 2008).
The common practice of examining the proximal 10 m of the small intestine is based on the observation that most intestinal Trichostrongylus spp. occur within the first 6 m of the small intestine (Beveridge and Barker, 1983), whereas larger amounts of plant debris are present in the more distal part of the small intestine (McKenna, 2008). McKenna (2008) claimed that processing only the first 10 m of the small intestine led to a recovery of < 50% of the worms located in the entire length, resulting in serious underestimates of the total number of worms present. However, the data presented were based on the necropsy of only 15 sheep, and a recovery of less than 50% of the total number was observed only in a few individual sheep, whereas in most infected sheep trichostrongylid nematodes were located in the proximal 10 m of the small intestine (cf. McKenna, 2008). Therefore, it can be concluded that the improvement of accuracy achieved by processing the entire small intestine is marginal and involves a significant increase in the amount of labour and time required for processing.
7. DNA-Based Methods for Specific Diagnosis of Strongylid Nematodes Infections in Livestock: Sample Preparation, Markers and Developments
Clearly, traditional methods (reviewed in Section 6) have limitations, in terms of sensitivity and/or specificity. In addition, they can be time consuming and costly to carry out. Advances in molecular biological technologies have enabled the development of new, sensitive and specific diagnostic methods that have found applications in the field of veterinary parasitology. The ability to specifically identify and study parasites (irrespective of life-cycle stage) using such tools has provided new insights into parasite systematics, population genetics, ecology and epidemiology, and has important implications for the specific diagnosis, treatment and control of parasitic diseases (Gasser, 2006). In particular, methods that rely on the enzymatic amplification of nucleic acids can overcome some of the limitations of traditional approaches (Gasser, 2006). Techniques that employ the PCR (Mullis et al., 1986, Saiki et al., 1988) can selectively amplify in vitro target DNA sequences from complex genomes or matrices and have led to advances in many areas of the biological sciences. PCR involves the heat denaturation of double-stranded DNA, followed by a decrease in temperature to allow oligonucleotide primers to bind (= anneal) to their complementary sequence on sense and antisense strands of the target template. Then, the temperature is increased again to enhance the enzymatic activity of a thermostable DNA polymerase, which extends the complementary strands from the primer sites. These synthesis steps are usually repeated 20–40 times in an automated thermal cycler, resulting in an exponential increase in target DNA copies. The major advantage of this methodology is that it enables the study of parasite DNA from minute amounts of template, which would otherwise be insufficient for conventional analysis. The value of this technology in the field of diagnostic veterinary parasitology lies in its ability to specifically identify parasites, detect infection and analyse genetic variation, which are particularly important, given the increasing problems of AR in parasitic nematodes (Gasser, 2006, Gasser et al., 2008).
7.1. Sample preparation prior to PCR
The main goals of sample preparation are to (i) concentrate the target organisms and, subsequently, the template for the PCR; (ii) eliminate possible PCR inhibitors; and (iii) produce a homogenous sample for specific and sensitive enzymatic amplification (Rådström et al., 2004). Complex biological (e.g. faecal) samples can contain a wide range of inhibitory substances (e.g. bile salts, collagen, haeme, humic acids and polysaccharides), which are capable of reducing or preventing PCR amplification (Rådström et al., 2004, Wilson, 1997). Different samples can have very different compositions, and the presence of substances potentially inhibitory to the PCR can vary depending on the sample type and composition (Hoorfar et al., 2004, Wilson, 1997). Therefore, the selection and evaluation of the sample preparation approach and a suitable reaction mixture, including polymerases and primers, are critical to obtain PCR-compatible samples of comparable composition, irrespective of the variation in the original matrix (e.g. batch-to-batch variation) (Hoorfar et al., 2004).
The selection of the most suitable method for sample preparation depends on the type of sample and the purpose of the PCR analysis, as there is no universal approach that would equally suit all sample matrices and/or applications (Hoorfar et al., 2004). Besides the choice of the adequate sample preparation approach, also the choice of the polymerase and the inclusion of ‘amplification enhancers’ (e.g. bovine serum albumin or dimethylsulphoxide) may improve enzymatic amplification and reduce the effects of inhibitory substances (Rådström et al., 2004). As an additional control for the presence of inhibitory substances in the amplification mixture and the efficiency of the DNA isolation and/or PCR reaction, the use of ‘spike-control DNA’ (natural or synthetic nucleotide sequences/fragments introduced into the sample) can be used (Ninove et al., 2011).
7.2. Specific genetic markers for strongylid nematodes
The key to the development of a reliable, PCR-based method for the specific diagnosis of infection is the definition of one or more suitable DNA target regions (genetic marker or locus) based on DNA sequencing. Since different genes evolve at different rates, the DNA region selected should be sufficiently variable in sequence to allow the identification of parasites to the taxonomic level required. For specific identification, the target DNA should display no or minor sequence variation within a species and differ sufficiently in sequence to consistently allow the delineation among species. In contrast, for the purpose of identifying population variants (‘strains’ or genotypes), a considerable degree of variation in the sequence should exist within a species. A range of target regions in the nuclear and mitochondrial genomes have been employed to achieve the identification of parasites to species or sub-specific genotypes (Anderson et al., 1998, Blouin, 2002, Chilton, 2004, Gasser, 2006). In nuclear ribosomal genes and spacers, there is often less sequence variation among individuals within a population and between populations, which makes them suitable as species-specific markers. Hence, in the case of genetic markers for the specific identification of strongylid nematodes of livestock, most of the focus has been on investigating nuclear ribosomal DNA (rDNA).
Although some success has been achieved with other DNA targets (e.g. Callaghan and Beh, 1994, Callaghan and Beh, 1996, Christensen et al., 1994, Roos and Grant, 1993, Zarlenga et al., 1994), most studies have consistently demonstrated that the first (ITS-1) and second (ITS-2) internal transcribed spacers (ITSs) of nuclear rDNA provide reliable genetic markers for the specific identification of a range of strongylid nematodes of livestock, including species of Haemonchus, Teladorsagia and Ostertagia (abomasum), Trichostrongylus (abomasum or small intestine), Cooperia, Nematodirus, Bunostomum (small intestine), Oesophagostomum and Chabertia (large intestine), Dictyocaulus, Protostrongylus and Metastrongylus (lung) (reviewed by Gasser, 2006, Gasser et al., 2008).
A comparison of the ITS sequences from a range of bursate nematodes has shown that the ITS-1 (364–522 bp) is usually larger in size than the ITS-2 (215–484 bp) (see Chilton, 2004). For instance, the ITS-1 region of the trichostrongylids Ostertagia ostertagi and Ostertagia lyrata (801 bp) (Zarlenga et al., 1998b) is longer than that of other trichostrongylids, including congeners, due to the presence of an internal 204 bp fragment, which is repeated twice (Zarlenga et al., 1998a, Zarlenga et al., 1998b, 2001). No major differences have been detected among species of Teladorsagia/Ostertagia in the lengths of their ITS-2 sequences (Chilton et al., 2001, Stevenson et al., 1996). The G + C content (39–50%) of the ITS-1 sequence of species studied is usually greater than of their ITS-2 (29–45%). The ITS-2 sequences of some species can be relatively A + T-rich (60–70%), which may relate to structural aspects of the precursor rRNA molecule. In addition, studies to date, show that the magnitude of sequence variation in both the ITS-1 and ITS-2 within a species is considerably less (usually < 1.5%) than the levels of sequence differences among species (Gasser, 2006), providing the basis for the specific identification of strongylids and diagnosis of infections of livestock.
8. Conventional PCR-Based Tools for the Identification of Species of Strongylid Nematodes and Diagnosis of Infections
The ITS-1 and/or ITS-2 rDNA regions provide useful genetic markers for the development of diagnostic PCR-based tools for strongylid nematodes (Gasser et al., 2006, Gasser et al., 2008); in addition to being species-specific in sequence, they are short (usually ≤ 800 bp), repetitive and undergo homogenization (Elder and Turner, 1995, Gasser, 2006), the latter factors ultimately determining the efficiency, ‘sensitivity’ and specificity of any PCR amplification procedure.
PCR-coupled SSCP analysis has provided a powerful approach for the specific identification of strongylid nematodes using markers ITS-1 and/or ITS-2 and for detecting cryptic (= morphologically similar but genetically distinct) species at any stage of development (Gasser et al., 2006, Gasser and Chilton, 2001). Although there has been a considerable focus on strongylid nematodes of humans, there have been some applications to those of livestock (reviewed by Gasser et al., 2006, Gasser et al., 2008).
Oligonucleotide primers have been designed to specific regions flanking and/or within the ITS-1 or ITS-2 for diagnostic applications (reviewed by Chilton, 2004, Gasser, 2006, Gasser et al., 2008). Using rDNA targets, this strategy has also been employed for the development of PCR assays for the genus- or species-specific identification of different developmental stages of strongylid nematodes. For instance, Zarlenga et al., 1998a, Zarlenga et al., 1998b described the development of a semi-quantitative PCR assay for the diagnosis of patent O. ostertagi infection in cattle. Conserved oligonucleotide primers were used in PCR to amplify a ~ 1 kb rDNA region, spanning the ITS-1 and part of the 5.8S rRNA gene, from O. ostertagi, whereas amplicons of ~ 600 bp were amplified from H. contortus, Cooperia oncophora and O. radiatum. When DNA samples derived from adult nematodes of the different genera were mixed and amplified simultaneously, there was no evidence of inhibition in the PCR, and O. ostertagi-specific amplicons were readily detected electrophoretically. There was a correlation between the intensity of the ~ 1-kb and 600-bp amplicons on gels and the percentage of O. ostertagi DNA within the mix of heterologous species. There was also a strong correlation between the percentage of O. ostertagi DNA and percentage of O. ostertagi eggs in the faeces. Effective amplification was achieved from one-twentieth of a single O. ostertagi egg. Hence, the establishment of this PCR assay had major implications for diagnosis of patent O. ostertagi infection in cattle as well as for studying the epidemiology of this parasite. Other studies (reviewed by Gasser, 2006, Gasser et al., 2008) demonstrated the diagnostic utility of PCR assays using species-specific ITS oligonucleotide primers or probes, even when the sequences of related species differ by a single nucleotide (Hung et al., 1999). For instance, Zarlenga et al. (2001) extended previous work to develop a multiplex PCR assay for the specific detection and differentiation of economically important gastrointestinal strongylid nematodes (including H. placei, O. ostertagi, Trichostrongylus spp., Cooperia oncophora/C. surnabada and O. radiatum) of cattle.
9. Real-Time PCR Coupled to High-Resolution Melting Analysis as a Diagnostic Tool
9.1. Principles of the technology
The method of RT-PCR, developed in the early 1990s (Higuchi et al., 1992), allows the enzymatic amplification to be monitored in real-time in vitro. All current RT-PCR systems detect the amplification using fluorescent dyes or probes. The predominant advantages of RT-PCR over-conventional PCR are that it allows high-throughput analysis in a ‘closed-tube’ format, not requiring handling or electrophoresis following amplification, that it can be employed for quantitation over a wide ‘dynamic range’ and that it can be used to differentiate amplicons of varying sequence(s) by melting-curve analysis.
The principle of the original method was to incorporate a specific, intercalating dye (e.g. ethidium bromide) into the PCR to measure the change in fluorescence after each cycle using a digital camera and a fluorometer coupled to the reaction tube (Higuchi et al., 1993). The technique has been modified to include other (non-carcinogenic) dyes, such as SYBR Green I (Becker et al., 1996), LCGreen (Wittwer et al., 2003), SYTO9 (Monis et al., 2005a) and EvaGreen (Wang et al., 2006). RT-PCR assays using such dyes enable the relative or absolute quantitation of amplicons by allowing the identification of the cycle (C t) at which the amplification commences. One or more DNA standards (of differing concentrations) and test samples are subjected to cycling at the same time and their C t values established and compared. Standard curves can be constructed based on the use of reference samples, and the relative amounts of template in test samples are calculated in relation to these curves.
Intercalating dyes, such as SYBR Green I, detect any double-stranded DNA, which is advantageous because they can be incorporated into any assay. However, a disadvantage is that the dye binds to all double-stranded DNA in a reaction, which includes primer dimers and non-specific products. This limitation can be overcome by acquiring fluorescence data at a temperature that denatures the non-specific products and leaves the specific products intact. The melting point of an amplicon is linked to the composition and length of the nucleotide sequence(s) within it, which means that a melting-curve analysis can be used to detect and/or characterize sequence variation within and among samples. Other recent advances include the complementary use of high resolution melting (HRM) analysis following RT-PCR (e.g. Jeffery et al., 2007). Melting analysis using the dye LCGreen or SYTO9 has been reported to achieve acceptable levels of reproducibility, attaining better mutation detection capacity than SYBR Green I (Monis et al., 2005a, Wittwer et al., 2003). Alternative, more expensive detection systems (other than intercalating dyes) include TaqMan probes (Heid et al., 1996), minor groove binder Eclipse probes (Afonina et al., 2002), molecular beacons (Piatek et al., 1998) and fluorescence resonance energy transfer (Chen and Kwok, 1999), ensuring specificity in the PCR through exclusive binding to the target sequence (Monis et al., 2005b).
9.2. RT-PCR assays for the diagnosis of infections with strongylid nematodes
Despite the promising results of RT-PCR for the diagnosis and quantification of selected protozoan and metazoan parasites (Bell and Ranford-Cartwright, 2002, Monis et al., 2005b, van Lieshout and Verweij, 2010, Zarlenga and Higgins, 2001), to date, relatively little research has focused on its use for the diagnosis of strongylid infections of livestock (cf. Gasser, 2006, Gasser et al., 2008). There have been attempts to use RT-PCR for the specific diagnosis and/or quantification of helminth eggs or larvae from the faeces from infected hosts. First efforts were made by von Samson-Himmelstjerna et al., 2002, von Samson-Himmelstjerna et al., 2003, who developed RT-PCR assays for the diagnosis and quantification of ovine gastrointestinal nematodes, including H. contortus, Ostertagia leptospicularis, T. colubriformis, C. curticei and for small strongyles (cyathostomins) of horses. However, these assays were used for the identification of larval or adult nematodes only, which limited their utility for routine diagnostic applications.
Harmon et al. (2007) evaluated the use of RT-PCR to quantify eggs of H. contortus from sheep faeces and examined various aspects, such as the influence of faecal inhibitors on PCR, the effects of competing and non-competing DNA in multiplex reactions and the impact of embryonic development within the egg on the PCR result. The assay developed showed linear quantifiable amplification of DNA obtained from egg quantities ranging from five to 75 eggs, whereas DNA from higher egg numbers of 75–1000 eggs did not show significant differences in C t, limiting the quantitative capacity of the assay to a narrow detection range (Harmon et al., 2007). During this study an impact of egg embryonic development on C t values has only been observed between 0 and 6 h of development at 21 °C, whilst later time periods at 6, 12 and 30 h did not show statistical differences in C t when compared to each other (Harmon et al., 2007). Non-competitive DNA, derived from environmental sources, did not appear to have a negative impact on amplification, but in multiplex reactions, the presence of high amounts of competing trichostrongyle DNA hindered the amplification of a different target species whose DNA is present at much smaller amounts (Harmon et al., 2007).
The storage of faecal samples is often necessary in a practical context, but the possible impact of egg embryonation during prolonged storage is known to be a critical factor relating to the accuracy of quantifying egg numbers by RT-PCR (Bott et al., 2009, Harmon et al., 2007). Therefore, strategies to circumvent this issue should be developed, which could possibly involve allowing maximum development to occur prior to DNA isolation (Harmon et al., 2007). It has been hypothesized that the technique used for DNA extraction and the presence of PCR inhibitors could be responsible also for discrepancies in the linear correlation between DNA amount and number of nematode eggs (Harmon et al., 2007). Harmon et al. (2007) suggested to account for the variability arising from DNA extraction and the presence of faecal inhibitors through the use of multiplex PCR systems that quantify, in relative terms, egg numbers using C t values, and include an exogenous DNA template to standardize C t values and assess every sample individually for faecal inhibitors (Harmon et al., 2007). Further work in this field had been done by two other research teams, who developed RT-PCR assays for the diagnosis of infections with the human hookworms Ancylostoma duodenale, Necator americanus and the nodule worm Oesophagostomum bifurcum (see Verweij et al., 2007) and the equine parasite Strongylus vulgaris (see Nielsen et al., 2008). These assays employed specific primers and TaqMan probes to target the ITS-2 region of nuclear rDNA. Verweij et al. (2007) suggested that false-negative RT-PCR results (n = 2) in relation to LC might be explained by the fact that the amount of faeces used for DNA isolation was 30 times less than that used to set up LC. However, both assays were reported by the authors to be of high analytical specificity and of a sensitivity superior to that of LC. Inhibition by faecal components was not evident. A potential limitation of these studies was that the specificity of these assays was based exclusively on the design and use of TaqMan probes. However, HRM or sequencing was not used to verify the identity or specificity of the amplicons produced.
Bott et al. (2009) established a combined microscopic-RT-PCR method that allows the semi-quantification of strongylid infections in sheep. During this study specific oligonucleotide primers were designed to ITS-2 and 28S rDNA regions of seven key genera or species of strongylids of sheep, including H. contortus, Te. circumcincta, Trichostrongylus spp., C. oncophora, C. ovina, Oesophagostomum columbianum and O. venulosum, and used in separate PCR reactions. To determine relative proportions of species/genera contributing to an FEC, standard curves were prepared for the RT-PCRs for individual species and demonstrated a log–linear relationship over four orders of magnitude. The C t values obtained from species-specific PCR reactions showed also a linear correlation to the numbers of eggs present per gram of faeces and demonstrated the applicability of this PCR approach for semi-quantification of target species. For some of the primer pairs used in this study, as little as 0.1–2 pg of DNA was sufficient to achieve specific amplification from the respective species, which equates to a proportion of genomic DNA which can be isolated from a single egg (Bott et al., 2009). During the evaluation of this PCR assay, all amplicons generated from specific primer pairs were examined by HRM and sequence analysis to confirm their identity. Designed primers were critically assessed for their analytical specificity (i.e. against a broad range of parasites that are known to be detectable from the faeces of infected sheep, including lungworms), and there was no evidence of non-specific amplification. However, Bott et al. (2009) stated that, due to possible sequence polymorphism or heterogeneity of ITS-2 among or within individuals from different geographical locations, the performance of the PCR platform might need additional assessment, if applied in different countries or regions.
Bott et al. (2009) also raised some points that needed consideration for future applications, such as the effect of faecal consistency on FEC and PCR results. The testing of loose/diarrhoeic and desiccated faecal samples can lead to an under- and over-estimation, respectively, of FECs (Le Jambre et al., 2007) and likely variability in semi-quantitative PCR results. Furthermore, Bott et al. (2009) discussed the need for rapid DNA isolation from faecal samples following their collection. The results of previous studies (Harmon et al., 2007) indicated that mitosis during the larval development leads to an increase of ITS-2 copy number and results in enhanced amplification during the RT-PCR. Because the storage and transport of samples at ambient temperatures is often necessary for practical reasons, approaches for the preservation of faecal material, for example, ethanol fixation might be applied. In addition, the direct extraction of DNA from faeces, using commercially available kits, has been proposed as an alternative to methods that involve the concentration of eggs by faecal flotation prior to DNA isolation (cf. Bott et al., 2009, Nielsen et al., 2008). However, such direct extraction methods need to be critically assessed for their ability to remove potential inhibitors (e.g. humic acids, phenolic compounds or blood) from faeces. A faecal flotation and egg isolation approach has been shown to remove such inhibitors (Bott et al., 2009) and has the advantage that it provides an FEC, which can be compared with a PCR result but which is not possible employing a direct DNA isolation-coupled PCR method. Furthermore, flotation allows for a concentration of eggs from multiple grams of faeces prior to DNA isolation and PCR, thus increasing the likelihood of amplifying DNA from very small numbers of nematode eggs (Bott et al., 2009, Nielsen et al., 2008). By contrast, only small amounts of faeces (e.g. ~ 0.25 g) are used in most commercially available, direct faecal DNA isolation methods, limiting the ‘sensitivity’ of subsequent PCR. Noting these limitations, the combined microscopic-molecular method (Bott et al., 2009) was established for the specific diagnosis of patent strongylid infections in sheep, and future work is required to evaluate the performance of this method for the specific diagnosis of infections with immature (pre-patent) or hypobiotic stages (e.g. Te. circumcincta or H. contortus) of nematodes and to compare it with direct amplification from DNA isolated directly from faecal samples.
9.3. Critical evaluation and application of a RT-PCR method to assess the composition of strongylid nematode populations in sheep with naturally acquired infections
Roeber et al. (2011) critically evaluated the performance of the RT-PCR method (Bott et al., 2009) for the diagnosis of naturally acquired strongylid nematode infections in sheep (n = 470; in a temperate climatic zone of south-eastern Australia), using a panel of 100 ‘negative control’ samples from sheep known not to harbour parasitic helminths. The authors compared the diagnostic sensitivity and specificity of this RT-PCR assay with a conventional faecal flotation method. They also established a system to rank the contribution of particular strongylid nematodes to the FECs from ‘mixed infections’ in individual sheep. The testing of faecal samples revealed that Te. circumcincta (80%) and Trichostrongylus spp. (66%) were most prevalent, followed by C. ovina (33%), O. venulosum (28%) and H. contortus (1%). For most sheep tested in this study, Te. circumcincta and Trichostrongylus spp. represented the largest proportion of strongylid eggs in faecal samples from individual sheep. This was the first large-scale prevalence survey of gastrointestinal nematodes in live sheep utilizing a molecular tool. The ability to rapidly rank strongylid nematodes according to their contribution to mixed infections represents a major advantage over routine flotation methods. The conclusion from this study was that this RT-PCR tool might be able to replace the conventional technique of LC.
This assessment showed that the RT-PCR assay achieved high diagnostic sensitivity (98%) and specificity (100%), and the test results were in ‘good’ agreement (i.e. Kappa: 0.95) with those achieved using conventional faecal flotation. In addition, of 53 field samples which were test-negative based on coproscopy, 23 tested positive by PCR for one or more target nematodes (a result which was confirmed by sequencing), demonstrating better sensitivity of the molecular approach compared with coproscopic examination, and reinforcing that microscopy is not an adequate reference technique (i.e. is an imperfect gold standard; cf. Conraths and Schares, 2006) for the detailed assessment of the sensitivity and specificity of a diagnostic assay. The results achieved demonstrated that the prevalences of key genera/species, such as Te. circumcincta and Trichostrongylus spp., known to be the dominant species infecting sheep in the winter rainfall environment of Victoria, Australia, were largely in accordance with information available in the published literature (e.g. Anderson, 1972, Anderson, 1973). The application of an ordinal ranking system to estimate the contribution of individual parasites to observed FEC results showed that these genera/species were also responsible for the largest proportion of strongylid eggs in the faecal samples tested. Although known to be abundant in winter rainfall environments, C. ovina and O. venulosum were found at high prevalence (33.6% and 28.7%, respectively). An unexpected finding was that O. venulosum was the main contributor to the observed FEC results for one of nine farms tested, which has important implications for the interpretation of FECs and anthelmintic control.
According to common practice (Brightling, 1988), FEC results of ≥ 200 EPG are considered to relate to a ‘significant’ worm burden, and without further considerations of the species present and their reproductive capacity, give an indication for anthelmintic treatment. This common practice, which involves frequent and, in many cases, unnecessary or excessive administration of anthelmintic drugs can promote AR development in gastrointestinal nematodes of sheep and other hosts, as recent evidence has shown (Kaplan, 2004, Wolstenholme et al., 2004). Given that O. venulosum is recognized to be less pathogenic than most strongylids of the upper alimentary tract (Donald et al., 1978) but has high fecundity (Gordon, 1981), FEC results (e.g. > 200 EPG) in which O. venulosum is the sole or main contributor would be misinterpreted, and sheep harbouring this relatively non-pathogenic would be treated unnecessarily. Therefore, the specific/generic identification of infecting nematodes assists the interpretation of FEC results and, subsequently, treatment decisions, thus, directly contributing to efforts of preserving the efficacy of currently available anthelmintics.
The RT-PCR assay (Roeber et al., 2011), coupled to conventional coproscopy, and the microscopic detection of Nematodirus, Trichuris and Moniezia eggs in faecal samples revealed that the majority of sheep investigated were parasitized by two to four helminth taxa per animal. Data from this study provided new and important insights into the composition and distribution of nematodes, which would not have been achievable in such detail using any of the currently used coprological methods. The results indicated that this tool should be applicable in other climatic regions and/or major sheep producing countries in the world. Depending on the nematode species infecting sheep in a particular climatic zone, minor modifications could be made to the molecular assay to adapt it for the diagnosis of infections with other important parasites (e.g. hookworm or lungworm) and to provide opportunities to study their biology, prevalence and distributions. For instance, as the life cycles of some lungworms, such as Muellerius capillaris and Protostrongylus rufescens, involve invertebrate intermediate hosts, such as snails and slugs, an adapted RT-PCR assay could be used to examine the prevalence and relative intensity of these parasites in their intermediate hosts and to study their ecology.
The ability to identify helminth species and to rank them according to their contribution to FEC results (Roeber et al., 2011) represents a novel approach that is time- and cost-efficient compared with classical diagnostic techniques and enables a better interpretation of FEC results, particularly in relation to the anthelmintic treatment of infected sheep (cf. McKenna, 1996, McKenna, 1997). This advance in the diagnosis of gastrointestinal nematode infections could directly and significant contribute to enhanced parasite control.
9.4. Assessment of RT-PCR to replace larval culture (LC) and to support conventional faecal egg count reduction testing (FECRT) of sheep
Roeber et al. (2012a) assessed the RT-PCR assay to support the diagnosis of AR in nematodes, in conjunction with conventional FECRT; in addition, a direct comparison of PCR results with those of LC and selective total worm counts (TWCs; considered a ‘gold standard’) was undertaken. In this assessment, the molecular assay achieved a diagnostic sensitivity of 100% and specificity of 87.5% in relation to TWC. These percentages where similar to those achieved previously (Roeber et al., 2011) (diagnostic sensitivity 98% and specificity 100% in relation to FEC), demonstrating that the RT-PCR assay consistently achieved a high diagnostic performance. DNA sequencing results also demonstrated that this molecular assay had a better sensitivity than the routinely used TWC method and, together with FECRT, was of practical value for the detection of albendazole resistance in Te. circumcincta and T. colubriformis populations. However, although the PCR test results were in accordance with TWC, the direct comparison of molecular and LC results showed markedly different findings, depending on the recommended measurements used for larval differentiation (Dikmans and Andrews, 1933, Gordon, 1933, McMurtry et al., 2000). Using the morphometric criteria given by Dikmans and Andrews (1933), the majority of L3s from the LC of the albendazole treated group of sheep were identified as Trichostrongylus, whereas, using the measurements recommended by Gordon (1933) and McMurtry et al. (2000), the same larvae were identified as Teladorsagia. This discrepancy emphasizes the complications and errors associated with the use of LC, which can readily lead to misinterpretations as to which nematodes are resistant to a particular drug. Overall, this study (Roeber et al., 2012a) demonstrated clearly that the molecular assay coupled to FECRT provides a rapid, efficient and universally applicable tool for the diagnosis of AR and the early detection of residual populations of worms in sheep following treatment. Future studies should be conducted to test sheep on different farms and the response of gastrointestinal nematodes to the treatment with other main groups of anthelmintics, such as imidazothiazoles/tetrahydropyrimidines or MLs, and also the newly developed monepantel (Zolvix, Novartis) (Kaminsky et al., 2008).
The movement of sheep and their gastrointestinal parasites between or among farms favours the spread of drug resistance (Blouin et al., 1995). Therefore, the routine use of the RT-PCR assay, in conjunction with FECRT, could provide a universally applicable method to test sheep before transport to and/or introduction on to a new farm, in order to reduce the spread of drug-resistant populations of nematodes. In addition, the assay allows the identification of sheep shedding large numbers of parasite eggs in faeces and, in conjunction with information about the infecting helminth species, can be used to support ‘targeted, selective treatment’ approaches (Kenyon et al., 2009). Such a strategy focuses on treating only sheep that will benefit most from an anthelmintic treatment, whereas sheep with low-intensity infections remain untreated to provide refugia for the dilution of resistance genes within a parasite population. Furthermore, the present RT-PCR assay had a similar or improved sensitivity compared with post-mortem diagnosis, thus being able to replace the latter. In practice, this means that the presence of particular species/genera and their prevalence can be assessed reliably without the need to kill sheep.
10. A Robotic, Multiplexed-Tandem PCR Assay for the Diagnosis of Gastrointestinal Strongylid Nematode Infections of Sheep
10.1. Rationale, establishment and critical assessment of the multiplexed-tandem PCR assay
A limitation of RT-PCR developed (Roeber et al., 2011, Roeber et al., 2012a) was that it employs individual primer pairs for individual specific or generic amplifications and requires numerous, manual handling steps throughout the entire procedure. Therefore, the goal was to develop a user-friendly and practical platform that would allow the rapid testing of large numbers of samples at relatively low cost with limited manual intervention, and that could be introduced into a routine testing laboratory. Although conventional multiplex RT-PCR (e.g. Chamberlain et al., 1988) seemed to be a promising prospect at first, preliminary work conducted (Bott N.J. and Gasser R.B., unpublished) showed that primer sets used in individual PCRs (cf. Bott et al., 2009) could not be incorporated into a single reaction to achieve specific amplifications. Other restrictions of a conventional multiplex PCR approach are that each target sequence requires a probe for fluorescence-based detection at a particular wave length and that such probes are costly and require the use of multi-channel RT-PCR thermocyclers—which usually have only four to six distinct wave-length channels, thus limiting the number of species/genera that can be detected.
In order to circumvent these issues, Roeber et al. (2012b) explored the use of multiplexed-tandem PCR (MT-PCR) (Stanley and Szewczuk, 2005). MT-PCR consists of two amplification phases: (i) a primary ‘target enrichment’ phase (through a small number of PCR cycles) conducted using multiplexed primer sets and (ii) a subsequent analytical amplification phase (utilizing a diluted product from the primary amplification as a template), consisting of the targeted amplification, in tandem rather than by multiplex, of each genetic locus using specific, nested primers. Because the initial amplification phase is limited to 10–15 cycles, interactions between or among multiplexed primer sets is minimized, reducing competition or the generation of artefactual products and limiting amplification bias, which would otherwise prevent downstream quantification (Stanley and Szewczuk, 2005). Because the primary amplicons are diluted (e.g. 100-fold) prior to use as templates in the secondary phase, primer carry over and PCR inhibition are substantially reduced. By conducting the secondary (analytical) amplification phase in tandem, the method can be coupled to a single-channel, RT-PCR thermocycler, allowing rapid screening and quantification employing one fluorogenic dye (e.g. SYTO9), thus reducing the cost associated with detection.
Roeber et al. (2012b) established a high-throughput MT-PCR assay for the diagnosis of nematode infections in sheep, and critically assessed its diagnostic sensitivity and specificity relative to RT-PCR as well as conventional LC using faecal samples from different flocks of sheep from a broad geographical range in Australia. The MT-PCR achieved high diagnostic specificity (87.5%) and sensitivity (100%) based on the testing of a panel of 100 faecal DNA samples from helminth-free sheep and 30 samples from sheep with infections confirmed by necropsy. This MT-PCR assay was then used to test 219 faecal samples from sheep with naturally acquired infections from various geographical localities within Australia, and results were compared with those of LC, using 139 of the 219 samples. The MT-PCR and LC results correlated significantly for most nematodes examined, but parasites of the large intestine were significantly under-represented in the LC results. The findings showed that Trichostrongylus spp. (87%), Teladorsagia circumcincta (80%) and H. contortus (67%) had the highest prevalences, followed by O. venulosum (51%) and C. ovina (12%). Importantly, this MT-PCR allowed a species- or genus-specific diagnosis of patent nematode infections to be made within 24 h (compared with 7–10 days for LC).
The evaluation of two different pre-PCR genomic DNA isolation methods showed that a combined egg flotation and column-purification approach achieved better sensitivity, overall, compared with direct faecal DNA isolation (Roeber et al., 2012b). The testing of samples from sheep from different geographical locations in Australia showed that the prevalence of the key nematodes investigated were consistent with their presumed distribution, based on historical data and many years of studies conducted using routine diagnostic procedures (Donald et al., 1978). Epidemiological information had not been provided in such detail prior to the use of a molecular-diagnostic tool, and the results achieved by routine LC and the molecular-diagnostic platform showed significant correlations (r s = 0.69–0.83) for most nematodes. However, C. ovina and O. venulosum were both much less frequently detected in LC than by the molecular assays. Thus, the evidence showed that LC was unreliable for the diagnosis of infections with these two nematodes and did not allow their prevalence and distribution to be studied with any confidence. Importantly, the inability of LC to detect the latter two species subsequently led to an over-estimation of other nematodes, namely H. contortus, Trichostrongylus and Teladorsagia, present in the LCs examined, thus providing a biassed result. This finding also highlights the utility of the present molecular-diagnostic platform for epidemiological studies and suggests that results obtained previously using traditional diagnostic techniques might have led to inaccurate information on the prevalence and distribution of particular species of gastrointestinal nematodes. The findings of Roeber et al., 2012a, Roeber et al., 2012b showed that C. ovina and O. venulosum may contribute much more significantly to infections in sheep than currently acknowledged. Given that the pathogenicity of these parasites is considered low (Donald et al., 1978), it is likely that the inability of classical techniques to reliably identify and quantify these parasites has led to unnecessary anthelmintic treatments at increased cost to the farmer and an increased risk of AR in nematode populations.
10.2. Future applications of the assay and extensions
An important future application of the MT-PCR platform would be the testing of pasture samples for infective larvae of strongylid nematodes. This approach has significant implications for studying the epidemiology and ecology (i.e. the seasonal occurrence of species and length of larval survival) of gastrointestinal nematodes in livestock, and enables assessments of the degree of contamination and risk of infection. For this purpose, pasture foliage could be collected, washed and concentrated; DNA could then be isolated from the larval suspension and molecular testing conducted. However, as the number of cells increases during the mitotic division of developing eggs and larvae, the number of ITS-2 copies also increases, which means that the MT-PCR platform will need to be calibrated to accommodate this variation. This PCR platform permits different sensitivity settings (based on the number of amplification cycles in the primary PCR) and, whilst a medium sensitivity (15 cycles of primary amplification) was shown to produce the best results for the detection of eggs from faeces, this sensitivity setting could be adjusted for L3s from pasture samples.
There are numerous other possible applications of a high-throughput MT-PCR assay. For example, such a test could also be used to assist in evaluating the efficacy of vaccines by monitoring FEC, either at the stage of initial vaccine development or, later, in field trials, to predict levels of protection against particular parasite species in individual host animals of different genetic backgrounds (i.e. different breeds of sheep). Immunological methods could also be used in concert with FECRT to measure protective effects of anti-nematode vaccines for sheep following challenge infection/s (with reference to well-defined controls). Moreover, to further refine the diagnosis of AR, genomic DNA regions linked to resistance (e.g. β-tubulin gene) could be integrated as targets into the assay. However, as AR is frequently polygenic (Beech et al., 2011) and/or can also be associated with drug transport mechanisms (Cvilink et al., 2009), and the mechanisms of resistance are only partially understood for many drug classes (cf. Taylor et al., 2002), more research needs to focus on identifying reliable DNA markers for the specific detection of AR.
There is now major potential to extend the use of MT-PCR assay to other socio-economically important pathogens, including helminths (e.g. flukes and lungworms), protists (e.g. Cryptosporidium, Eimeria and Giardia), bacteria (e.g. Campylobacter, Escherichia, Salmonella and Yersinia) and viruses (e.g. Rota- and Corona-viruses). Furthermore, such a platform could also be adapted, with modifications being made to the DNA isolation procedure and primers used, for the diagnosis of infections with blood pathogens (e.g. Babesia and Theileria) or the detection of pathogens in environmental samples. In addition, although we have focused on the development of this platform for the detection and identification of pathogens of socio-economic importance in sheep (Roeber et al., 2012b), a similar approach would be of major benefit assessing such pathogens in other hosts, including other small ruminants, cattle, horses and even humans. For example, estimating the number of nematode eggs in the faeces from adult cattle is challenging, as eggs are generally present in low numbers and, thus, require more sensitive techniques of diagnosis (Agneessens et al., 2000, Mes et al., 2001). Therefore, the MT-PCR-coupled diagnostic platform could be further enhanced by automating the counting of eggs in faecal samples prior to DNA isolation. Approaches that involve the recovery of nematode eggs from faeces and the automated counting of all eggs present in a sample (as opposed to the counting of eggs from sub-aliquots of a sample and their extrapolation to the total sample volume) by image recognition have been developed (Mes et al., 2001, Mes et al., 2007). Consequently, with only minor modifications, the MT-PCR-based platform could provide a major advance in the diagnosis of nematode infections in cattle.
The MT-PCR platform (Roeber et al., 2012) could also be further enhanced through the use a robotic DNA isolation procedure. Automated DNA isolation platforms are readily available from different manufacturers and include easyMAG (BioMerieux, Mary l’Etoile, France), m2000sp (Abbott, Abbott Park, IL, USA), MagNA Pure LC 2.0 (Roche) and QiaSymphony (Qiagen, Hilden, Germany), but their suitability for the isolation of DNA from faecal samples would need to be critically assessed before they could be incorporated into the present MT-PCR system. Further advantages of this approach include the possibility for large-scale, targeted collections and long-term storage of DNA samples. Unlike traditional coprodiagnostic testing, in which samples are usually examined only once to enumerate parasite eggs or larvae, and are then discarded. The use of DNA samples has the advantage that the sample material collected can be stored (at − 80 °C) in a clean and space efficient manner (i.e. either in frozen or in dehydrated form), allowing the testing of the samples many years or even decades later. This provides the opportunity of carrying out (even retrospectively) studies of ‘new’ or ‘emerging’ pathogens. These are important applications in times of changing environmental conditions (e.g. climate change, urbanization and associated changes in pathogen transmission, distribution and epidemiology), increased problems with AR as well as the increased mobility of humans and/or animals between/among different regions or countries.
11. Concluding Remarks: Replacing Traditional with Molecular-Diagnostic Tools
The accurate diagnosis of gastrointestinal nematode infections of livestock underpins investigations of the biology, ecology and epidemiology of parasites and supports the monitoring of emerging problems with AR. Current, routinely used methods of diagnosis rely on the detection or morphological identification of the infective stages (eggs and/or larvae) of these nematodes from the faeces from their hosts. Until recently, these classical techniques, which are mostly time consuming and laborious, had not undergone any substantial technological advancement in the past decades. Because key stages (mainly eggs and larvae) of numerous genera and species of nematodes infecting livestock lack distinctive morphological features, traditional approaches are not able to achieve an accurate species- or even genus-specific diagnosis, making it difficult to conduct reliable studies of the biology, epidemiology and ecology of parasites, unless expensive and time-consuming post-mortem investigations are conducted. This situation has also hampered investigations of the occurrence and distribution of AR in strongylid nematodes of livestock, which represents a global problem.
Advances in PCR-based technologies and the availability of specific genetic markers in the ITSs of nuclear rDNA have provided the opportunity of developing enhanced PCR-based tools for diagnosis (reviewed by Gasser, 2006, Gasser et al., 2008). Current evidence (Roeber et al., 2011, 2012a,b) shows that RT-PCR and MT-PCR assays can replace the inaccurate and time-consuming method of LC in routine diagnostic laboratories. This high-throughput MT-PCR assay takes < 1 day to perform, compared with at least a week for LC, thus reducing the time that the sheep producer has to wait for a diagnosis. From a service provision perspective, this platform does not require detailed technical expertise of the operator, has high sensitivity and specificity and has broad applicability, in that it can be used to carry out large-scale epidemiological studies, to support the diagnosis of drug resistance and will be applicable or adaptable to other parasites and/or hosts. Furthermore, the MT-PCR platform delivers, rapidly, objective and detailed results (according to genus or species), which is of major value for the design of enhanced control strategies (Table 4.1). Overall, the evidence presented shows that the MT-PCR (Roeber et al., 2012b) essentially meets the international standards (Conraths and Schares, 2006, Office International des Epizooties, 2004) required for use in a laboratory setting for research or routine diagnostic purposes and has major advantages over-classical methods, particularly in relation to the interpretation of FEC results and recommendations about anthelmintic treatment. This assay enhances the diagnosis of infections with nematode species, which are problematic to detect or identify by traditional coprological techniques, either because of their unfavourable development under standard culture conditions or their high morphological/morphometric similarity with other species/genera (i.e. Teladorsagia and Trichostrongylus, C. ovina and O. venulosum). Consequently, this advance in diagnosis allows gaps in our current knowledge of the epidemiology and distribution of these parasites (being associated mainly with the inaccuracy of traditional diagnostic techniques) to be overcome.
If put into a commercial context and made available to diagnostic laboratories, the MT-PCR platform could also find applicability for the rapid screening for important diseases affecting livestock production systems, including those with zoonotic potential, and assist in the better surveillance and control of disease outbreaks related to such pathogens. DNA-based methods of diagnosis have significant advantages, in terms of sensitivity and specificity, time required to conduct the assay and can be standardized among and within laboratories. These features make them suitable to be incorporated into surveillance systems, which are based on a ‘from stable-to-table’ approach, such as the Hazard Analysis Critical Control Point (HACCP). The routine application of such high-throughput and automated diagnostic platforms provides major scope for better disease surveillance and for detailed epidemiological investigations, shorter response times to tackle and control disease outbreaks (as the diagnosis using molecular tools generally takes less time compared with culture- or microscopy-based approaches) and, ultimately, to provide greater protection for the consumer of animal products (Fig. 4.1). The data obtained using such methods would help government authorities, such as the Food and Agricultural Organization (FAO), Office International des Epizootis (OIE) and/or the World Health Organization (WHO) in the detailed tracking and mapping of disease outbreaks or spreads, to make forecasts about their occurrence and to implement appropriate contingency plans and guidelines for effective and sustainable pathogen control.
In conclusion, the high-throughput MT-PCR assay developed recently is considered to represent a major advance for the diagnosis of gastrointestinal nematode infections in small ruminants (sheep and goats). This assay has improved diagnostic performance over-traditional diagnostic techniques and is considered to provide a new benchmark. This robotic molecular assay delivers high performance for epidemiological surveys and can be used efficiently, in combination with FECRT, to support the detection and monitoring of AR. Due to its semi-automated, high-throughput nature, this method can now replace traditional diagnostic approaches and is suitable for routine application in service and research laboratories. The assay may need to be adapted to include the testing for additional parasites, depending on where the assay is to be deployed. Current evidence indicates that this robotic assay is highly adaptable, allowing the development of a wide range of next-generation diagnostic tools to underpin the control of socio-economically important infectious diseases of animals and the detection and monitoring of drug resistance. Could this be a turning point in the diagnosis of parasitic diseases of livestock?
Acknowledgements
This research has been supported through funds from the Australian Research Council (ARC). Other support from Melbourne Water Corporation is gratefully acknowledged. F. R. is the grateful recipient of scholarships from the University of Melbourne.
References
- Afonina I.A., Reed M.W., Lusby E., Shishkina I.G., Belousov Y.S. Minor groove binder-conjugated DNA probes for quantitative DNA detection by hybridization-triggered fluorescence. Biotechniques. 2002;32(940–944):946–949. doi: 10.2144/02324pf01. [DOI] [PubMed] [Google Scholar]
- Agneessens J., Claerebout E., Dorny P., Borgsteede F.H.M., Vercruysse J. Nematode parasitism in adult dairy cows in Belgium. Vet. Parasitol. 2000;90:83–92. doi: 10.1016/s0304-4017(00)00232-6. [DOI] [PubMed] [Google Scholar]
- Alvarez-Sanchez M.A., Perez-Garcia J., Cruz-Rojo M.A., Rojo-Vazquez F.A. Real time PCR for the diagnosis of benzimidazole resistance in trichostrongylids of sheep. Vet. Parasitol. 2005;129:291–298. doi: 10.1016/j.vetpar.2005.02.004. [DOI] [PubMed] [Google Scholar]
- Anderson N. Trichostrongylid infections of sheep in a winter rainfall region. I. Epizootiological studies in the Western District of Victoria, 1966–67. Aust. J. Agr. Res. 1972;23:1113–1129. [Google Scholar]
- Anderson N. Trichostrongylid infections of sheep in a winter rainfall region. II. Epizootiological studies in the western district of Victoria, 1967–68. Aust. J. Agr. Res. 1973;24:599–611. [Google Scholar]
- Anderson C.R. second ed. CAB international; Wallingford, UK: 2000. Nematode Parasites of Vertebrates. Their Development and Transmission. [Google Scholar]
- Anderson T.J.C., Blouin M.S., Beech R.N. Population biology of parasitic nematodes: applications of genetic markers. Adv. Parasitol. 1998;41:219–283. doi: 10.1016/s0065-308x(08)60425-x. [DOI] [PubMed] [Google Scholar]
- Baker N.F., Cook E.F., Douglas J.R., Cornelius C.E. The pathogenesis of trichostrongyloid parasites. III. Some physiological observations in lambs suffering from acute parasitic gastroenteritis. J. Parasitol. 1959;45:643–651. [PubMed] [Google Scholar]
- Barger I.A. The statistical distribution of trichostrongylid nematodes in grazing lambs. Int. J. Parasitol. 1985;15:645–649. doi: 10.1016/0020-7519(85)90010-4. [DOI] [PubMed] [Google Scholar]
- Barger I.A. The role of epidemiological knowledge and grazing management for helminth control in small ruminants. Int. J. Parasitol. 1999;29:41–47. doi: 10.1016/s0020-7519(98)00176-3. [DOI] [PubMed] [Google Scholar]
- Becker A., Reith A., Napiwotzki J., Kadenbach B. A quantitative method of determining initial amounts of DNA by polymerase chain reaction cycle titration using digital imaging and a novel DNA stain. Anal. Biochem. 1996;237:204–207. doi: 10.1006/abio.1996.0230. [DOI] [PubMed] [Google Scholar]
- Beech R.N., Skuce P., Bartley D.J., Martin R.J., Prichard R.K., Gilleard J.S. Anthelmintic resistance: markers for resistance, or susceptibility? Parasitology. 2011;138:160–174. doi: 10.1017/S0031182010001198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell A.S., Ranford-Cartwright L.C. Real-time quantitative PCR in parasitology. Trends Parasitol. 2002;18:338–342. [PubMed] [Google Scholar]
- Bennett J.L., Pax R.A. Micromotility meter: an instrument designed to evaluate the action of drugs on motility of larval and adult nematodes. Parasitology. 1986;93:341–346. doi: 10.1017/s0031182000051507. [DOI] [PubMed] [Google Scholar]
- Bennett J.L., Pax R.A. Micromotility meter: instrumentation to analyse helminth motility. Parasitol. Today. 1987;3:159–160. doi: 10.1016/0169-4758(87)90203-1. [DOI] [PubMed] [Google Scholar]
- Berghen P., Hilderson H., Vercruysse J., Dorny P. Evaluation of pepsinogen, gastrin and antibody response in diagnosing ostertagiasis. Vet. Parasitol. 1993;46:175–195. doi: 10.1016/0304-4017(93)90057-t. [DOI] [PubMed] [Google Scholar]
- Berrie D.A., East I.J., Bourne A.S., Bremner K.C. Differential recoveries from faecal cultures of larvae of some gastro-intestinal nematodes of cattle. J. Helminthol. 1988;62:110–114. doi: 10.1017/s0022149x00011330. [DOI] [PubMed] [Google Scholar]
- Besier R.B., Love S.C.J. Anthelmintic resistance in sheep nematodes in Australia the need for new approaches. Aust. J. Exp. Agric. 2003;43:1383–1391. [Google Scholar]
- Beveridge I., Barker I.K. Morphogenesis of Trichostrongylus rugatus and distribution during development in sheep. Vet. Parasitol. 1983;13:55–65. doi: 10.1016/0304-4017(83)90020-1. [DOI] [PubMed] [Google Scholar]
- Beveridge I., Pullman A.L., Martin R.R., Barelds A. Effects of temperature and relative humidity on development and survival of the free-living stages of Trichostrongylus colubriformis, T. rugatus and T. vitrinus. Vet. Parasitol. 1989;33:143–153. doi: 10.1016/0304-4017(89)90062-9. [DOI] [PubMed] [Google Scholar]
- Beveridge I., Pullman A.L., Phillips P.H., Martin R.R., Barelds A., Grimson R. Comparison of the effects of infection with Trichostrongylus colubriformis, T. vitrinus and T. rugatus in Merino lambs. Vet. Parasitol. 1989;32:229–245. doi: 10.1016/0304-4017(89)90123-4. [DOI] [PubMed] [Google Scholar]
- Blackhall W.J., Liu H.Y., Xu M., Prichard R.K., Beech R.N. Selection at a P-glycoprotein gene in ivermectin- and moxidectin-selected strains of Haemonchus contortus. Mol. Biochem. Parasitol. 1998;95:193–201. doi: 10.1016/s0166-6851(98)00087-5. [DOI] [PubMed] [Google Scholar]
- Blackhall W.J., Prichard R.K., Beech R.N. Selection at a γ-aminobutyric acid receptor gene in Haemonchus contortus resistant to avermectins/milbemycins. Mol. Biochem. Parasitol. 2003;131:137–145. doi: 10.1016/s0166-6851(03)00201-9. [DOI] [PubMed] [Google Scholar]
- Blouin M.S. Molecular prospecting for cryptic species of nematodes: mitochondrial DNA versus internal transcribed spacer. Int. J. Parasitol. 2002;32:527–531. doi: 10.1016/s0020-7519(01)00357-5. [DOI] [PubMed] [Google Scholar]
- Blouin M.S., Yowell C.A., Courtney C.H., Dame J.B. Host movement and the genetic structure of populations of parasitic nematodes. Genetics. 1995;141:1007–1014. doi: 10.1093/genetics/141.3.1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boersema J.H. Possibilities and limitations in the detection of anthelmintic resistance. Facts and reflections IV. Resistance of parasites to anthelmintics. A Workshop in the C.E.C. Animal Pathology Programme Held at the Central Veterinary Institute, Lelystad, The Netherlands, December 9–10, 1982; 1983. pp. 207–215. [Google Scholar]
- Borgsteede F.H.M., Couwenberg T. Changes in LC50 in an in vitro development assay during the patent period of Haemonchus contortus in sheep. Res. Vet. Sci. 1987;42:413–414. [PubMed] [Google Scholar]
- Bott N.J., Campbell B.E., Beveridge I., Chilton N.B., Rees D., Hunt P.W., Gasser R.B. A combined microscopic-molecular method for the diagnosis of strongylid infections in sheep. Int. J. Parasitol. 2009;39:1277–1287. doi: 10.1016/j.ijpara.2009.03.002. [DOI] [PubMed] [Google Scholar]
- Brightling A. Inkata Press; Melbourne, Australia: 1988. Sheep Diseases; pp. 3–10. [Google Scholar]
- Brunsdon R.V. Seasonal changes in the level and composition of nematode worm burdens in young sheep. N. Z. J. Agric. Res. 1970;13:126–148. [Google Scholar]
- Brunsdon R.V. Principles of helminth control. Vet. Parasitol. 1980;6:185–215. [Google Scholar]
- Callaghan M.J., Beh K.J. A middle-repetitive DNA sequence element in the sheep parasitic nematode, Trichostrongylus colubriformis. Parasitology. 1994;109:345–350. doi: 10.1017/s0031182000078379. [DOI] [PubMed] [Google Scholar]
- Callaghan M.J., Beh K.J. A tandemly repetitive DNA sequence is present at diverse locations in the genome of Ostertagia circumcincta. Gene. 1996;174:273–279. doi: 10.1016/0378-1119(96)00093-5. [DOI] [PubMed] [Google Scholar]
- Chamberlain J.S., Gibbs R.A., Ranier J.E., Nguyen P.N., Caskey C.T. Deletion screening of the Duchenne muscular dystrophy locus via multiplex DNA amplification. Nucleic Acids Res. 1988;16:11141–11156. doi: 10.1093/nar/16.23.11141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charlier J., Vercruysse J., Smith J., Vanderstichel R., Stryhn H., Claerebout E., Dohoo I. Evaluation of anti-Ostertagia ostertagi antibodies in individual milk samples as decision parameter for selective anthelmintic treatment in dairy cows. Prev. Vet. Med. 2010;93:147–152. doi: 10.1016/j.prevetmed.2009.10.002. [DOI] [PubMed] [Google Scholar]
- Chen X., Kwok P.Y. Homogeneous genotyping assays for single nucleotide polymorphisms with fluorescence resonance energy transfer detection. Genet. Anal. 1999;14:157–163. doi: 10.1016/s1050-3862(98)00016-3. [DOI] [PubMed] [Google Scholar]
- Chilton N.B. The use of nuclear ribosomal DNA markers for the identification of bursate nematodes (order Strongylida) and for the diagnosis of infections. Anim. Health Res. Rev. 2004;5:173–187. doi: 10.1079/ahr200497. [DOI] [PubMed] [Google Scholar]
- Chilton N.B., Newton L.A., Beveridge I., Gasser R.B. Evolutionary relationships of trichostrongyloid nematodes (Strongylida) inferred from ribosomal DNA sequence data. Mol. Phylogenet. Evol. 2001;19:367–386. doi: 10.1006/mpev.2001.0938. [DOI] [PubMed] [Google Scholar]
- Christensen C.M., Zarlenga D.S., Gasbarre L.C. Ostertagia, Haemonchus, Cooperia, and Oesophagostomum: construction and characterization of genus-specific DNA probes to differentiate important parasites of cattle. Exp. Parasitol. 1994;78:93–100. doi: 10.1006/expr.1994.1009. [DOI] [PubMed] [Google Scholar]
- Cohen S., Sadun E.H. Blackwell Scientific Publications; Oxford, UK: 1976. Immunology of Parasitic Infections. [Google Scholar]
- Cole V.G. Australian Government Publishing Service; Canberra, Australia: 1986. Helminth Parasites of Sheep and Cattle. [Google Scholar]
- Coles G.C., Bauer C., Borgsteede F.H.M., Geerts S., Klei T.R., Taylor M.A., Waller P.J. World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P.). Methods for the detection of anthelmintic resistance in nematodes of veterinary importance. Vet. Parasitol. 1992;44:35–44. doi: 10.1016/0304-4017(92)90141-u. [DOI] [PubMed] [Google Scholar]
- Coles G.C., Folz S.D., Tritschler J.P., II Motility response of levamisole benzimidazole-resistant Haemonchus contortus larvae. Vet. Parasitol. 1989;31:253–257. doi: 10.1016/0304-4017(89)90074-5. [DOI] [PubMed] [Google Scholar]
- Coles G.C., Jackson F., Pomroy W.E., Prichard R.K., von Samson-Himmelstjerna G., Silvestre A., Taylor M.A., Vercruysse J. The detection of anthelmintic resistance in nematodes of veterinary importance. Vet. Parasitol. 2006;136:167–185. doi: 10.1016/j.vetpar.2005.11.019. [DOI] [PubMed] [Google Scholar]
- Coles G.C., Tritschler J.P., II, Giordano D.J., Laste N.J., Schmidt A.L. Larval development test for detection of anthelmintic resistant nematodes. Res. Vet. Sci. 1988;45:50–53. [PubMed] [Google Scholar]
- Conraths F.J., Schares G. Validation of molecular-diagnostic techniques in the parasitological laboratory. Vet. Parasitol. 2006;136:91–98. doi: 10.1016/j.vetpar.2005.12.004. [DOI] [PubMed] [Google Scholar]
- Corwin R.M. Economics of gastrointestinal parasitism of cattle. Vet. Parasitol. 1997;72:451–460. doi: 10.1016/s0304-4017(97)00110-6. [DOI] [PubMed] [Google Scholar]
- Cox D.D., Todd A.C. Survey of gastrointestinal parasitism in Wisconsin dairy cattle. J. Am. Vet. Med. Assoc. 1962;141:706–709. [PubMed] [Google Scholar]
- Cringoli G., Rinaldi L., Veneziano V., Capelli G., Scala A. The influence of flotation solution, sample dilution and the choice of McMaster slide area (volume) on the reliability of the McMaster technique in estimating the faecal egg counts of gastrointestinal strongyles and Dicrocoelium dendriticum in sheep. Vet. Parasitol. 2004;123:121–131. doi: 10.1016/j.vetpar.2004.05.021. [DOI] [PubMed] [Google Scholar]
- Cringoli G., Rinaldi L., Maurelli M.P., Utzinger J. FLOTAC: new multivalent techniques for qualitative and quantitative copromicroscopic diagnosis of parasites in animals and humans. Nat. Protoc. 2010;5:503–515. doi: 10.1038/nprot.2009.235. [DOI] [PubMed] [Google Scholar]
- Cvilink V., Lamka J., Sklov L. Xenobiotic metabolizing enzymes and metabolism of anthelmintics in helminths. Drug Metab. Rev. 2009;41:8–26. doi: 10.1080/03602530802602880. [DOI] [PubMed] [Google Scholar]
- De Chaneet G.C., Dunsmore J.D. Climate and the distribution of intestinal Trichostrongylus spp. of sheep. Vet. Parasitol. 1988;26:273–283. doi: 10.1016/0304-4017(88)90096-9. [DOI] [PubMed] [Google Scholar]
- Demeler J. Institute für Parasitology, Tierärztliche Hochschule Hannover; Hannover, Germany: 2005. The Physiological Site of Action and the Site of Resistance to the Macrocyclic Lactone Anthelmintics in Sheep Parasitic Trichostrongyloid Nematodes. [Google Scholar]
- Demeler J., Küttler U., El-Abdellati A., Stafford K., Rydzik A., Varady M., Kenyon F., Coles G., Höglund J., Jackson F., Vercruysse J., von Samson-Himmelstjerna G. Standardization of the larval migration inhibition test for the detection of resistance to ivermectin in gastrointestinal nematodes of ruminants. Vet. Parasitol. 2010;174:58–64. doi: 10.1016/j.vetpar.2010.08.020. [DOI] [PubMed] [Google Scholar]
- Demeler J., Küttler U., von Samson-Himmelstjerna G. Adaptation and evaluation of three different in vitro tests for the detection of resistance to anthelmintics in gastrointestinal nematodes of cattle. Vet. Parasitol. 2010;170:61–70. doi: 10.1016/j.vetpar.2010.01.032. [DOI] [PubMed] [Google Scholar]
- Dikmans G., Andrews J.S. A comparative morphological study of the infective larvae of the common nematodes parasitic in the alimentary tract of sheep. Trans. Am. Microsc. Soc. 1933;52:1–25. [Google Scholar]
- Dinaburg A.G. The efficiency of the Baermann apparatus in the recovery of larvae of Haemonchus contortus. J. Parasitol. 1942;28:433–440. [Google Scholar]
- Dobson R.J., Barnes E.H., Birclijin S.D., Gill J.H. The survival of Ostertagia circumcincta and Trichostrongylus colubriformis in faecal culture as a source of bias in apportioning egg counts to worm species. Int. J. Parasitol. 1992;22:1005–1008. doi: 10.1016/0020-7519(92)90060-x. [DOI] [PubMed] [Google Scholar]
- Dobson R.J., LeJambre L., Gill J.H. Management of anthelmintic resistance: inheritance of resistance and selection with persistent drugs. Int. J. Parasitol. 1996;26:993–1000. doi: 10.1016/s0020-7519(96)80078-6. [DOI] [PubMed] [Google Scholar]
- Dobson R.J., Sangster N.C., Besier R.B., Woodgate R.G. Geometric means provide a biased efficacy result when conducting a faecal egg count reduction test (FECRT) Vet. Parasitol. 2009;161:162–167. doi: 10.1016/j.vetpar.2008.12.007. [DOI] [PubMed] [Google Scholar]
- Doenhoff M., Chiodini P., Hamilton J. Specific and sensitive diagnosis of schistosome infection: can it be done with antibodies? Trends Parasitol. 2004;20:35–39. doi: 10.1016/j.pt.2003.10.019. [DOI] [PubMed] [Google Scholar]
- Donald A.D., Southcott W.H., Dineen J.K. Commonwealth Scientific and Industrial Research Organisation; Melbourne, Australia: 1978. The Epidemiology and Control of Gastrointestinal Parasites of Sheep in Australia. p. xii + 153. [Google Scholar]
- Elard L., Humbert J.F. Importance of the mutation of amino acid 200 of the isotype 1 β-tubulin gene in the benzimidazole resistance of the small-ruminant parasite Teladorsagia circumcincta. Parasitol. Res. 1999;85:452–456. doi: 10.1007/s004360050577. [DOI] [PubMed] [Google Scholar]
- Elder J.F., Turner B.J. Concerted evolution of repetitive DNA sequences in eukaryotes. Q. Rev. Biol. 1995;70:297–320. doi: 10.1086/419073. [DOI] [PubMed] [Google Scholar]
- Engvall E., Ruitenberg E.J. ELISA, enzyme linked immunosorbent assay—a new technique for sero-diagnosis of trichinosis. Parasitology. 1974;69:xxiv. [PubMed] [Google Scholar]
- Eysker M. The population dynamics of Trichostrongylus vitrinus and T. colubriformis in sheep in the Netherlands. Parasitology. 1978;77:xvi. [Google Scholar]
- Eysker M., Kooyman F.N.J. Notes on necropsy and herbage processing techniques for gastrointestinal nematodes of ruminants. Vet. Parasitol. 1993;46:205–213. doi: 10.1016/0304-4017(93)90059-v. [DOI] [PubMed] [Google Scholar]
- Eysker M., Ploeger H.W. Value of present diagnostic methods for gastrointestinal nematode infections in ruminants. Parasitology. 2000;120:109–119. doi: 10.1017/s0031182099005752. [DOI] [PubMed] [Google Scholar]
- Fletcher S. Indirect fluorescent antibody technique in the serology of Toxoplasma gondii. J. Clin. Pathol. 1965;18:193–199. doi: 10.1136/jcp.18.2.193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox M.T. Pathophysiology of infection with gastrointestinal nematodes in domestic ruminants: recent developments. Vet. Parasitol. 1997;72:285–308. doi: 10.1016/s0304-4017(97)00102-7. [DOI] [PubMed] [Google Scholar]
- Gaba S., Chadoeuf J., Monestiez P., Sauve C., Cortet J., Cabaret J. Estimation of abomasum strongyle nematode infections in sheep at necropsy: tentative proposals for a simplified technique. Vet. Parasitol. 2006;140:105–113. doi: 10.1016/j.vetpar.2006.03.021. [DOI] [PubMed] [Google Scholar]
- Gasbarre L.C., Leighton E.A., Davies C.J. Influence of host genetics upon antibody responses against gastrointestinal nematode infections in cattle. Vet. Parasitol. 1993;46:81–91. doi: 10.1016/0304-4017(93)90049-s. [DOI] [PubMed] [Google Scholar]
- Gasser R.B. Molecular tools—advances, opportunities and prospects. Vet. Parasitol. 2006;136:69–89. doi: 10.1016/j.vetpar.2005.12.002. [DOI] [PubMed] [Google Scholar]
- Gasser R.B., Chilton N.B. Applications of single-strand conformation polymorphism (SSCP) to taxonomy, diagnosis, population genetics and molecular evolution of parasitic nematodes. Vet. Parasitol. 2001;101:201–213. doi: 10.1016/s0304-4017(01)00567-2. [DOI] [PubMed] [Google Scholar]
- Gasser R.B., Hu M., Chilton N., Campbell B., Jex A., Otranto D., Cafarchia C., Beveridge I., Zhu X. Single-strand conformation polymorphism (SSCP) for the analysis of genetic variation. Nat. Protoc. 2006;1:3121–3128. doi: 10.1038/nprot.2006.485. [DOI] [PubMed] [Google Scholar]
- Gasser R.B., Bott N.J., Chilton N.B., Hunt P., Beveridge I. Toward practical, DNA-based diagnostic methods for parasitic nematodes of livestock—bionomic and biotechnological implications. Biotechnol. Adv. 2008;26:325–334. doi: 10.1016/j.biotechadv.2008.03.003. [DOI] [PubMed] [Google Scholar]
- Geary T.G., Nulf S.C., Favreau M.A., Tang L., Prichard R.K., Hatzenbuhler N.T., Shea M.H., Alexander S.J., Klein R.D. Three [beta]-tubulin cDNAs from the parasitic nematode Haemonchus contortus. Mol. Biochem. Parasitol. 1992;50:295–306. doi: 10.1016/0166-6851(92)90227-b. [DOI] [PubMed] [Google Scholar]
- Geerts S., Brandt J., Borgsteede F.H.M., Van Loon H. Reliability and reproducibility of the larval paralysis test as an in vitro method for the detection of anthelmintic resistance of nematodes against levamisole and morantel tartrate. Vet. Parasitol. 1989;30:223–232. doi: 10.1016/0304-4017(89)90018-6. [DOI] [PubMed] [Google Scholar]
- Georgi J.R., McCulloch C.E. Diagnostic morphometry: identification of helminth eggs by discriminant analysis of morphometric data. Proc. Helminthol. Soc. Wash. 1989;56:44–57. [Google Scholar]
- Gibbons L.M. CAB International; Wallingford, UK: 2010. Keys to the Nematode Parasites of Vertebrates. Supplementary Volume; pp. 83–102. [Google Scholar]
- Gill J.H., Lacey E. Avermectin/milbemycin resistance in trichostrongyloid nematodes. Int. J. Parasitol. 1998;28:863–877. doi: 10.1016/s0020-7519(98)00068-x. [DOI] [PubMed] [Google Scholar]
- Gill J.H., Redwin J.M., van Wyk J.A., Lacey E. Avermectin inhibition of larval development in Haemonchus contortus—effects of ivermectin resistance. Int. J. Parasitol. 1995;25:463–470. doi: 10.1016/0020-7519(94)00087-5. [DOI] [PubMed] [Google Scholar]
- Gill J.H., Redwin J.M., Wyk J.A.v., Lacey E. Detection of resistance to ivermectin in Haemonchus contortus. Int. J. Parasitol. 1991;21:771–776. doi: 10.1016/0020-7519(91)90144-v. [DOI] [PubMed] [Google Scholar]
- Giordano D.J., Tritschler J.P., Coles G.C. Selection of ivermectin-resistant Trichostrongylus colubriformis in lambs. Vet. Parasitol. 1988;30:139–148. doi: 10.1016/0304-4017(88)90161-6. [DOI] [PubMed] [Google Scholar]
- Gordon H.M. Differential diagnosis of the larvae of Ostertagia spp. and Trichostrongylus spp. of sheep. Aust. Vet. J. 1933;9:223–227. [Google Scholar]
- Gordon H.M. The epidemiology of parasitic diseases, with special reference to studies with nematode parasites of sheep. Aust. Vet. J. 1948;24:17–45. [Google Scholar]
- Gordon H.M. The epidemiology of helminthosis in sheep in winter-rainfall regions of Australia. I. Preliminary observations. Aust. Vet. J. 1953;29:337–348. [Google Scholar]
- Gordon H.M. Some aspects of the control of helminthosis in sheep. Veterinary Inspector N.S.W. 1967;31:88–99. [Google Scholar]
- Gordon H.M. Epidemiology of helminthosis in sheep, refresher course for veterinarians. Proceedings No. 58: Refresher Course on Sheep, 10–14 August; University of Sydney, Post-Graduate Committee in Veterinary Science; 1981. pp. 551–566. [Google Scholar]
- Gordon H.M., Whitlock H.V. A new technique for counting nematode eggs in sheep faeces. J. Counc. Sci. Ind. Res. 1939;12:50–52. [Google Scholar]
- Grant J.L. The epizootiology of nematode parasites of sheep in a high-rainfall area of Zimbabwe. J. S. Afr. Vet. Assoc. 1981;52:33–37. [PubMed] [Google Scholar]
- Grimshaw W.T.R., Hong C., Hunt K.R. Potential for misinterpretation of the faecal egg count reduction test for levamisole resistance in gastrointestinal nematodes of sheep. Vet. Parasitol. 1996;62:267–273. doi: 10.1016/0304-4017(95)00874-8. [DOI] [PubMed] [Google Scholar]
- Harder A., Holden-Dye L., Walker R., Wunderlich F. Mechanisms of action of emodepside. Parasitol. Res. 2005;97(Suppl. 1):S1–S10. doi: 10.1007/s00436-005-1438-z. [DOI] [PubMed] [Google Scholar]
- Harder A., Schmitt-Wrede H.P., Krücken J., Marinovski P., Wunderlich F., Willson J., Amliwala K., Holden-Dye L., Walker R. Cyclooctadepsipeptides—an anthelmintically active class of compounds exhibiting a novel mode of action. Int. J. Antimicrob. Agents. 2003;22(3):318–331. doi: 10.1016/s0924-8579(03)00219-x. (Review) [DOI] [PubMed] [Google Scholar]
- Harmon F.R., Williams Z.B., Zarlenga D.S., Hildreth M.B. Real-time PCR for quantifying Haemonchus contortus eggs and potential limiting factors. Parasitol. Res. 2007;101:71–76. doi: 10.1007/s00436-006-0428-0. [DOI] [PubMed] [Google Scholar]
- Hawkins J.A. Economic benefits of parasite control in cattle. Vet. Parasitol. 1993;46:159–173. doi: 10.1016/0304-4017(93)90056-s. [DOI] [PubMed] [Google Scholar]
- Heid C.A., Stevens J., Livak K.J., Williams P.M. Real time quantitative PCR. Genome Res. 1996;6:986–994. doi: 10.1101/gr.6.10.986. [DOI] [PubMed] [Google Scholar]
- Higuchi R., Dollinger G., Walsh P.S., Griffith R. Simultaneous amplification and detection of specific DNA sequences. Biotechnology. 1992;10:413–417. doi: 10.1038/nbt0492-413. [DOI] [PubMed] [Google Scholar]
- Higuchi R., Fockler C., Dollinger G., Watson R. Kinetic PCR analysis: real-time monitoring of DNA amplification reactions. Biotechnology. 1993;11:1026–1030. doi: 10.1038/nbt0993-1026. [DOI] [PubMed] [Google Scholar]
- Hoorfar J., Wolffs P., Rådström P. Diagnostic PCR: validation and sample preparation are two sides of the same coin. Acta Pathol. Microbiol. Immunol. Scand. 2004;112:808–814. doi: 10.1111/j.1600-0463.2004.apm11211-1207.x. [DOI] [PubMed] [Google Scholar]
- Hoste H., Torres-Acosta J.F. Non chemical control of helminths in ruminants: adapting solutions for changing worms in a changing world. Vet. Parasitol. 2011;180:144–154. doi: 10.1016/j.vetpar.2011.05.035. [DOI] [PubMed] [Google Scholar]
- Hubert J., Kerboeuf D. A new method for culture of larvae used in diagnosis of ruminant gastrointestinal strongylosis: comparison with fecal cultures. Can. J. Comp. Med. 1984;48:63–71. [PMC free article] [PubMed] [Google Scholar]
- Hung G.C., Gasser R.B., Beveridge I., Chilton N.B. Species-specific amplification by PCR of ribosomal DNA from some equine strongyles. Parasitology. 1999;119:69–80. doi: 10.1017/s0031182099004497. [DOI] [PubMed] [Google Scholar]
- Hungerford T.G. nineth ed. MacGraw-Hill Medical; Sydney, Australia: 1990. Diseases of Livestock. [Google Scholar]
- Jackson F. Anthelmintic resistance—the state of play. Br. Vet. J. 1993;149:123–138. doi: 10.1016/S0007-1935(05)80083-1. [DOI] [PubMed] [Google Scholar]
- Jeffery N., Gasser R.B., Steer P.A., Noormohammadi A.H. Classification of Mycoplasma synoviae strains using single-strand conformation polymorphism and high-resolution melting-curve analysis of the vlhA gene single-copy region. Microbiology. 2007;153:2679–2688. doi: 10.1099/mic.0.2006/005140-0. [DOI] [PubMed] [Google Scholar]
- Johnson M.J., Behnke J.M., Coles G.C. Detection of gastrointestinal nematodes by a coproantigen capture ELISA. Res. Vet. Sci. 1996;60:7–12. doi: 10.1016/s0034-5288(96)90122-8. [DOI] [PubMed] [Google Scholar]
- Kaminsky R., Ducray P., Jung M., Clover R., Rufener L., Bouvier J., Weber S.S., Wenger A., Wieland-Berghausen S., Goebel T., Gauvry N., Pautrat F., Skripsky T., Froelich O., Komoin-Oka C., Westlund B., Sluder A., Maser P. A new class of anthelmintics effective against drug-resistant nematodes. Nature. 2008;452:176–180. doi: 10.1038/nature06722. [DOI] [PubMed] [Google Scholar]
- Kaplan R.M. Drug resistance in nematodes of veterinary importance: a status report. Trends Parasitol. 2004;20:477–481. doi: 10.1016/j.pt.2004.08.001. [DOI] [PubMed] [Google Scholar]
- Kassai T. Butterworth Heinemann; Oxford, UK: 1999. Veterinary Helminthology. [Google Scholar]
- Kenyon F., Greer A.W., Coles G.C., Cringoli G., Papadopoulos E., Cabaret J., Berrag B., Varady M., Van Wyk J.A., Thomas E., Vercruysse J., Jackson F. The role of targeted selective treatments in the development of refugia-based approaches to the control of gastrointestinal nematodes of small ruminants. Vet. Parasitol. 2009;164:3–11. doi: 10.1016/j.vetpar.2009.04.015. [DOI] [PubMed] [Google Scholar]
- Kerboeuf D., Guégnard F., Vern Y.L. Detection of P-glycoprotein-mediated multidrug resistance against anthelmintics in Haemonchus contortus using anti-human mdr1 monoclonal antibodies. Parasitol. Res. 2003;91:79–85. doi: 10.1007/s00436-003-0942-2. [DOI] [PubMed] [Google Scholar]
- Kerboeuf D., Hubert J. Changes in the response of Haemonchus contortus eggs to the ovicidal activity of thiabendazole during the course of infection. Ann. Rech. Vet. 1987;18:365–370. [PubMed] [Google Scholar]
- Kloosterman A., Verhoeff J., Ploeger H.W., Lam T.J. Antibodies against nematodes in serum, milk and bulk milk samples as possible estimators of infection in dairy cows. Vet. Parasitol. 1993;47:267–278. doi: 10.1016/0304-4017(93)90028-l. [DOI] [PubMed] [Google Scholar]
- Kwa M.S.G., Veenstra J.G., Roos M.H. Benzimidazole resistance in Haemonchus contortus is correlated with a conserved mutation at amino acid 200 in beta-tubulin isotype 1. Mol. Biochem. Parasitol. 1994;63:299–303. doi: 10.1016/0166-6851(94)90066-3. [DOI] [PubMed] [Google Scholar]
- Lacey E. The role of the cytoskeletal protein, tubulin, in the mode of action and mechanism of drug resistance to benzimidazoles. Int. J. Parasitol. 1988;18:885–936. doi: 10.1016/0020-7519(88)90175-0. [DOI] [PubMed] [Google Scholar]
- Lancaster M.B., Hong C. Differentiation of third stage larvae of ‘ovine Ostertagia’ type and Trichostrongylus species. Vet. Rec. 1987;120:503. doi: 10.1136/vr.120.21.503. [DOI] [PubMed] [Google Scholar]
- Lane C. The mass diagnosis of ankylostome infestation (Part I) Trans. R. Soc. Trop. Med. Hyg. 1922;16:274–315. [Google Scholar]
- Larsen J.W.A., Anderson N., Vizard A.L., Anderson G.A., Hoste H. Diarrhoea in Merino ewes during winter: association with trichostrongylid larvae. Aust. Vet. J. 1994;71:365–372. doi: 10.1111/j.1751-0813.1994.tb00930.x. [DOI] [PubMed] [Google Scholar]
- Larsen J.W.A., Anderson N., Ware J.W., Fegely C.D. The productivity of Merino flocks in South-Eastern Australia in the presence of anthelmintic resistance. Small Rumin. Res. 2006;62:87–93. [Google Scholar]
- Le Jambre L.F. Egg hatch as an in vitro assay of thiabendazole resistance in nematodes. Vet. Parasitol. 1976;2:385–391. [Google Scholar]
- Le Jambre L.F., Dominik S., Eady S.J., Henshall J.M., Colditz I.G. Adjusting worm egg counts for faecal moisture in sheep. Vet. Parasitol. 2007;145:108–115. doi: 10.1016/j.vetpar.2006.11.017. [DOI] [PubMed] [Google Scholar]
- Le Jambre L.F., Ractliffe L.H., Uhazy L.S., Whitlock J.H. Fecal egg output of lambs in relationship to Haemonchus contortus burden. Int. J. Parasitol. 1971;1:157–160. doi: 10.1016/0020-7519(71)90010-5. [DOI] [PubMed] [Google Scholar]
- Levecke B., Rinaldi L., Charlier J., Maurelli M., Morgoglione M., Vercruysse J., Cringoli G. Monitoring drug efficacy against gastrointestinal nematodes when faecal egg counts are low: do the analytic sensitivity and the formula matter? Parasitol. Res. 2011;109:953–957. doi: 10.1007/s00436-011-2338-z. [DOI] [PubMed] [Google Scholar]
- Levecke B., Rinaldi L., Charlier J., Maurelli M.P., Bosco A., Vercruysse J., Cringoli G. The bias, accuracy and precision of faecal egg count reduction test results in cattle using McMaster, Cornell-Wisconsin and FLOTAC egg counting methods. Vet. Parasitol. 2012;188:194–199. doi: 10.1016/j.vetpar.2012.03.017. [DOI] [PubMed] [Google Scholar]
- Levine N.D. Burgess Publishing Company; Minneapolis, USA: 1968. Nematode Parasites of Domestic Animals and of Man. [Google Scholar]
- Levine N.D., Mehra K.N., Clark D.T., Aves I.J. A comparison of nematode egg counting techniques for cattle and sheep feces. Am. J. Vet. Res. 1960;21:511–515. [PubMed] [Google Scholar]
- Lichtenfels J.R., Hoberg E.P., Zarlenga D.S. Systematics of gastrointestinal nematodes of domestic ruminants: advances between 1992 and 1995 and proposals for future research. Vet. Parasitol. 1997;72:225–245. doi: 10.1016/s0304-4017(97)00099-x. [DOI] [PubMed] [Google Scholar]
- MAFF . Her Majesty’s Stationary Office; London, UK: 1986. Manual of Veterinary Parasitological Laboratory Techniques; pp. 20–27. [Google Scholar]
- Martin P.J., Anderson N., Jarrett R.G. Resistance to benzimidazole anthelmintics in field strains of Ostertagia and Nematodirus in sheep. Aust. Vet. J. 1985;62:38–43. doi: 10.1111/j.1751-0813.1985.tb14230.x. [DOI] [PubMed] [Google Scholar]
- Martin P.J., Anderson N., Jarrett R.G. Detecting benzimidazole resistance with faecal egg count reduction tests and in vitro assays. Aust. Vet. J. 1989;66:236–240. doi: 10.1111/j.1751-0813.1989.tb13578.x. [DOI] [PubMed] [Google Scholar]
- Martin P.J., Le Jambre L.F. Larval paralysis as an in vitro assay of levamisole and morantel tartrate resistance in Ostertagia. Vet. Sci. Commun. 1979;3:159–164. [Google Scholar]
- Martin R.J., Bai G.X., Clark C.L., Robertson A.P. Methyridine (2-[2-methoxyethyl]-pyridine) and levamisole activate different ACh receptor subtypes in nematode parasites: a new lead for levamisole-resistance. Br. J. Pharmacol. 2003;140:1068–1076. doi: 10.1038/sj.bjp.0705528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin R.J., Robertson A.P. Mode of action of levamisole and pyrantel, anthelmintic resistance, E153 and Q57. Parasitology. 2007;134:1093–1104. doi: 10.1017/S0031182007000029. [DOI] [PubMed] [Google Scholar]
- McCoy M.A., Edgar H.W., Kenny J., Gordon A.W., Dawson L.E., Carson A.F. Evaluation of on-farm faecal egg counting in sheep. Vet. Rec. 2005;156:21–23. doi: 10.1136/vr.156.1.21. [DOI] [PubMed] [Google Scholar]
- McKenna P.B. The diagnostic value and interpretation of faecal egg counts in sheep. N. Z. Vet. J. 1981;29:129–132. doi: 10.1080/00480169.1981.34821. [DOI] [PubMed] [Google Scholar]
- McKenna P.B. The estimation of gastrointestinal strongyle worm burdens in young sheep flocks: a new approach to the interpretation of faecal egg counts I. Development. NZ Vet. J. 1987;35:94–97. doi: 10.1080/00480169.1987.35396. [DOI] [PubMed] [Google Scholar]
- McKenna P.B. Potential limitations of the undifferentiated faecal egg count reduction test for the detection of anthelmintic resistance in sheep. N. Z. Vet. J. 1996;44:73–75. doi: 10.1080/00480169.1996.35938. [DOI] [PubMed] [Google Scholar]
- McKenna P.B. Further potential limitations of the undifferentiated faecal egg count reduction test for the detection of anthelmintic resistance in sheep. N. Z. Vet. J. 1997;45:244–246. doi: 10.1080/00480169.1997.36038. [DOI] [PubMed] [Google Scholar]
- McKenna P.B. Faecal egg counts as a guide for drench use. N. Z. Vet. J. 2002;50:123–124. doi: 10.1080/00480169.2002.36295. [DOI] [PubMed] [Google Scholar]
- McKenna P.B. A comparison of faecal egg count reduction test procedures. N. Z. Vet. J. 2006;54:202–203. doi: 10.1080/00480169.2006.36697. [DOI] [PubMed] [Google Scholar]
- McKenna P.B. Comparison of two worm counting procedures for the enumeration of abomasal and small intestinal nematode parasites of sheep. Vet. Parasitol. 2008;157:254–259. doi: 10.1016/j.vetpar.2008.08.003. [DOI] [PubMed] [Google Scholar]
- McKenna P.B., Simpson B.H. The estimation of gastrointestinal strongyle worm burdens in young sheep flocks: a new approach to the interpretation of faecal egg counts. II. Evaluation. N. Z. Vet. J. 1987;35:98–100. doi: 10.1080/00480169.1987.35397. [DOI] [PubMed] [Google Scholar]
- McLaren M.L., Draper C.C., Roberts E., Minter-Goedbloed E., Lighthart G.S., Teesdale C.H., Amin M.A., Omer A.H.S., Bartlett A., Voller A. Studies on the enzyme linked immunosorbent assay (ELISA) for Schistosoma mansoni infection. Ann. Trop. Med. Parasitol. 1978;72:243–253. doi: 10.1080/00034983.1978.11719312. [DOI] [PubMed] [Google Scholar]
- McLeod R.S. Costs of major parasites to the Australian livestock industries. Int. J. Parasitol. 1995;25:1363–1367. doi: 10.1016/0020-7519(95)00071-9. [DOI] [PubMed] [Google Scholar]
- McMurtry L.W., Donaghy M.J., Vlassoff A., Douch P.G.C. Distinguishing morphological features of the third larval stage of ovine Trichostrongylus spp. Vet. Parasitol. 2000;90:73–81. doi: 10.1016/s0304-4017(00)00230-2. [DOI] [PubMed] [Google Scholar]
- Mes T.H., Ploeger H.W., Terlou M., Kooyman F.N., Van der Ploeg M.P., Eysker M. A novel method for the isolation of gastro-intestinal nematode eggs that allows automated analysis of digital images of egg preparations and high throughput screening. Parasitology. 2001;123:309–314. doi: 10.1017/s0031182001008496. [DOI] [PubMed] [Google Scholar]
- Mes T.H.M., Eysker M., Ploeger H.W. A simple, robust and semi-automated parasite egg isolation protocol. Nat. Protoc. 2007;2:486–489. doi: 10.1038/nprot.2007.56. [DOI] [PubMed] [Google Scholar]
- Miller C.M., Waghorn T.S., Leathwick D.M., Gilmour M.L. How repeatable is a faecal egg count reduction test? NZ Vet J. 2006;54:323–328. doi: 10.1080/00480169.2006.36718. [DOI] [PubMed] [Google Scholar]
- Monis P.T., Giglio S., Saint C.P. Comparison of SYTO9 and SYBR Green I for real-time polymerase chain reaction and investigation of the effect of dye concentration on amplification and DNA melting curve analysis. Anal. Biochem. 2005;340:24–34. doi: 10.1016/j.ab.2005.01.046. [DOI] [PubMed] [Google Scholar]
- Monis P.T., Giglio S., Keegan A.R., Thompson R.C.A. Emerging technologies for the detection and genetic characterization of protozoan parasites. Trends Parasitol. 2005;21:340–346. doi: 10.1016/j.pt.2005.05.012. [DOI] [PubMed] [Google Scholar]
- Mullis K., Faloona F., Scharf S., Saiki R., Horn G., Erlich H. Specific enzymatic amplification of DNA in vitro: the polymerase chain reaction. Cold Spring Harb. Symp. Quant. Biol. 1986;51(1):263–273. doi: 10.1101/sqb.1986.051.01.032. [DOI] [PubMed] [Google Scholar]
- Nicholls J., Obendorf D.L. Application of a composite faecal egg count procedure in diagnostic parasitology. Vet. Parasitol. 1994;52:337–342. doi: 10.1016/0304-4017(94)90125-2. [DOI] [PubMed] [Google Scholar]
- Nielsen M.K., Peterson D.S., Monrad J., Thamsborg S.M., Olsen S.N., Kaplan R.M. Detection and semi-quantification of Strongylus vulgaris DNA in equine faeces by real-time quantitative PCR. Int. J. Parasitol. 2008;38:443–453. doi: 10.1016/j.ijpara.2007.07.014. [DOI] [PubMed] [Google Scholar]
- Nielsen M.K., Vidyashankar A.N., Andersen U.V., DeLisi K., Pilegaard K., Kaplan R.M. Effects of fecal collection and storage factors on strongylid egg counts in horses. Vet. Parasitol. 2010;167:55–61. doi: 10.1016/j.vetpar.2009.09.043. [DOI] [PubMed] [Google Scholar]
- Ninove L., Nougairede A., Gazin C., Thirion L., Delogu I., Zandotti C., Charrel R.N., De Lamballerie X. RNA and DNA bacteriophages as molecular diagnosis controls in clinical virology: a comprehensive study of more than 45,000 routine PCR tests. PLoS One. 2011;6:e16142. doi: 10.1371/journal.pone.0016142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nonhebel S., Kastner T. Changing demand for food, livestock feed and biofuels in the past and in the near future. Livest. Sci. 2011;139:3–10. [Google Scholar]
- Noordin R., Smith H.V., Mohamad S., Maizels R.M., Fong M.Y. Comparison of IgG-ELISA and IgG4-ELISA for Toxocara serodiagnosis. Acta Trop. 2005;93:57–62. doi: 10.1016/j.actatropica.2004.09.009. [DOI] [PubMed] [Google Scholar]
- O'Connor L.J., Walkden-Brown S.W., Kahn L.P. Ecology of the free-living stages of major trichostrongylid parasites of sheep. Vet. Parasitol. 2006;142:1–15. doi: 10.1016/j.vetpar.2006.08.035. [DOI] [PubMed] [Google Scholar]
- Office International des Epizooties . Quality management in veterinary testing laboratories. In: Office International des Epizooties (OIE), editor. OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Office International des Epizooties; Paris, France: 2004. pp. 1–31. [Google Scholar]
- Ogunremi O., Halbert G., Mainar Jaime R., Benjamin J., Pfister K. Accuracy of an indirect fluorescent-antibody test and of a complement-fixation test for the diagnosis of Babesia caballi in field samples from horses. Prev. Vet. Med. 2008;83:41–51. doi: 10.1016/j.prevetmed.2007.06.009. [DOI] [PubMed] [Google Scholar]
- Palmer D.G., McCombe I.L. Lectin staining of trichostrongylid nematode eggs of sheep: rapid identification of Haemonchus contortus eggs with peanut agglutinin. Int. J. Parasitol. 1996;26:447–450. doi: 10.1016/0020-7519(96)00009-4. [DOI] [PubMed] [Google Scholar]
- Parnell I.W., Rayski C., Dunn A.M., Mackintosh G.M. A survey of the helminths of Scottish hill sheep. J. Helminthol. 1954;28:53–110. doi: 10.1017/s0022149x00032715. [DOI] [PubMed] [Google Scholar]
- Perry B.D., Randolph T.F. Improving the assessment of the economic impact of parasitic diseases and of their control in production animals. Vet. Parasitol. 1999;84:145–168. doi: 10.1016/s0304-4017(99)00040-0. [DOI] [PubMed] [Google Scholar]
- Petersen M.B., Craven J., Bjoern H., Nansen P. Use of a migration assay for the separation of adult pyrantel-susceptible and -resistant Oesophagostomum dentatum. Vet. Parasitol. 2000;91:141–145. doi: 10.1016/s0304-4017(00)00264-8. [DOI] [PubMed] [Google Scholar]
- Petersen M.B., Friis C., Bjoern H. A new in vitro assay of benzimidazole activity against adult Oesophagostomum dentatum. Int. J. Parasitol. 1997;27:1333–1339. doi: 10.1016/s0020-7519(97)00104-5. [DOI] [PubMed] [Google Scholar]
- Presidente P.J.A. Methods for detection of resistance to anthelmintics. In: Anderson N., Waller P.J., editors. Resistance in nematodes to anthelmintic drugs. CSIRO and Australian Wool Corporation Technical Publication; 1985. pp. 13–27. [Google Scholar]
- Pfeiffer D.U. Wiley-Blackwell; Oxford, UK: 2010. Veterinary Epidemiology: An Introduction. [Google Scholar]
- Piatek A.S., Tyagi S., Pol A.C., Telenti A., Miller L.P., Kramer F.R., Alland D. Molecular beacon sequence analysis for detecting drug resistance in Mycobacterium tuberculosis. Nat. Biotechnol. 1998;16:359–363. doi: 10.1038/nbt0498-359. [DOI] [PubMed] [Google Scholar]
- Pritchard G. Milk antibody testing in cattle. In Pract. 2001;23:542–549. [Google Scholar]
- Rådström P., Knutsson R., Wolffs P., Lövenklev M., Löfström C. Pre-PCR processing: strategies to generate PCR-compatible samples. Mol. Biotechnol. 2004;26:133–146. doi: 10.1385/MB:26:2:133. [DOI] [PubMed] [Google Scholar]
- Raynaud J.P. Etude de l’efficacité d’une technique de coproscopie quantitative pour le diagnostic de routine et le contrôle des infestations parasitaires des bovins, ovins, équins et porcins. Ann. Parasitol. Hum. Comp. 1970;45:321–342. [PubMed] [Google Scholar]
- Richmond J.E., Jorgensen E.M. One GABA and two acetylcholine receptors function at the C. elegans neuromuscular junction. Nat. Neurosci. 1999;2:791. doi: 10.1038/12160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roeber F., Campbell A.J.D., Anderson N., Anderson G.A., Larsen J.W.A., Gasser R.B., Jex A.R. A molecular diagnostic tool to replace conventional faecal egg count reduction testing in sheep. PLoS One. 2012;7(5):e37327. doi: 10.1371/journal.pone.0037327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roeber F., Jex A.R., Campbell A.J.D., Nielsen R., Anderson G.A., Stanley K.K., Gasser R.B. Establishment of a robotic, high-throughput platform for the specific diagnosis of gastrointestinal nematode infections in sheep. Int. J. Parasitol. 2012;42(13–14):1151–1158. doi: 10.1016/j.ijpara.2012.10.005. [DOI] [PubMed] [Google Scholar]
- Roeber F., Jex A.R., Campbell A.J.D., Campbell B.E., Anderson G.A., Gasser R.B. Evaluation and application of a molecular method to assess the composition of strongylid nematode populations in sheep with naturally acquired infections. Infect. Genet. Evol. 2011;11:849–854. doi: 10.1016/j.meegid.2011.01.013. [DOI] [PubMed] [Google Scholar]
- Rinaldi L., Coles G.C., Maurelli M.P., Musella V., Cringoli G. Calibration and diagnostic accuracy of simple flotation, McMaster and FLOTAC for parasite egg counts in sheep. Vet. Parasitol. 2011;177:345–352. doi: 10.1016/j.vetpar.2010.12.010. [DOI] [PubMed] [Google Scholar]
- Roberts F.H.S., O'Sullivan P.J. Methods for egg counts and larval cultures for strongyles infesting the gastro-intestinal tract of cattle. Aust. J. Agr. Res. 1950;1:99–102. [Google Scholar]
- Roberts J.L., Swan R.A. Quantitative studies of ovine haemonchosis. I. Relationship between faecal egg counts and total worm counts. Vet. Parasitol. 1981;8:165–171. [PubMed] [Google Scholar]
- Robertson T.G., Elliott D.C. The laboratory assessment of worm parasite populations in sheep. N. Z. J. Agric. Res. 1966;9:350–358. [Google Scholar]
- Roos M.H., Grant W.N. Species-specific PCR for the parasitic nematodes Haemonchus contortus and Trichostrongylus colubriformis. Int. J. Parasitol. 1993;23:419–421. doi: 10.1016/0020-7519(93)90021-p. [DOI] [PubMed] [Google Scholar]
- Rowe A., McMaster K., Emery D., Sangster N. Haemonchus contortus infection in sheep: parasite fecundity correlates with worm size and host lymphocyte counts. Vet. Parasitol. 2008;153:285–293. doi: 10.1016/j.vetpar.2008.01.040. [DOI] [PubMed] [Google Scholar]
- Russel A.J.F., Doney J.M., Gunn R.G. Subjective assessment of body fat in live sheep. J. Agric. Sci. 1969;72:451–454. [Google Scholar]
- Sackett, D., Holmes, P., 2006. Assessing the economic cost of endemic disease on the profitability of Australian beef cattle and sheep producers. In: Meat and Livestock Australia Limited (Ed.), Sydney, Australia. ISBN 1741910021.
- Saiki R.K., Gyllensten U.B., Erlich H.A. IRL Press; Oxford, UK: 1988. The Polymerase Chain Reaction. [Google Scholar]
- Salisbury J.R., Arundel J.H. Peri-parturient deposition of nematode eggs by ewes and residual pasture contamination as sources of infection for lambs. Aust. Vet. J. 1970;46:523–529. doi: 10.1111/j.1751-0813.1970.tb06637.x. [DOI] [PubMed] [Google Scholar]
- Sanchez J., Markham F., Dohoo I., Sheppard J., Keefe G., Leslie K. Milk antibodies against Ostertagia ostertagi: relationships with milk IgG and production parameters in lactating dairy cattle. Vet. Parasitol. 2004;120:319–330. doi: 10.1016/j.vetpar.2004.01.010. [DOI] [PubMed] [Google Scholar]
- Sangster N.C. Anthelmintic resistance: past, present and future. Second International Conference. Novel Approaches to the Control of the Helminth Parasites of Livestock. Baton Rouge, Louisiana, USA, 22–26 March; 1999. pp. 115–124. [Google Scholar]
- Sangster N.C., Whitlock H.V., Kelly J.D., Gunawan M., Hall C.A. The effect of single and divided dose administration on the efficacy of fenbendazole against adult stages of benzimidazole resistant sheep trichostrongylids. Res. Vet. Sci. 1979;26:85–89. [PubMed] [Google Scholar]
- Scott E.W., Mitchell E.S., Armour J., Bairden K., Soutar A., Bogan J.A. Level of benzimidazole resistance in a strain of Ostertagia circumcincta studied over several infections in lambs. Vet. Parasitol. 1989;30:305–314. doi: 10.1016/0304-4017(89)90100-3. [DOI] [PubMed] [Google Scholar]
- Silvestre A., Humbert J.F. Diversity of benzimidazole-resistance alleles in populations of small ruminant parasites. Int. J. Parasitol. 2002;32:921–928. doi: 10.1016/s0020-7519(02)00032-2. [DOI] [PubMed] [Google Scholar]
- Sommer C. Digital image analysis and identification of eggs from bovine parasitic nematodes. J. Helminthol. 1996;70:143–151. doi: 10.1017/s0022149x00015303. [DOI] [PubMed] [Google Scholar]
- Southcott W.H., Major G.W., Barger I.A. Seasonal pasture contamination and availability of nematodes for grazing sheep. Aust. J. Agr. Res. 1976;27:277–286. [Google Scholar]
- Stanley K., Szewczuk E. Multiplexed tandem PCR: gene profiling from small amounts of RNA using SYBR Green detection. Nucleic Acids Res. 2005;33:e180. doi: 10.1093/nar/gni182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stear M.J., Bishop S.C. The curvilinear relationship between worm length and fecundity of Teladorsagia circumcincta. Int. J. Parasitol. 1999;29:777–780. doi: 10.1016/s0020-7519(99)00019-3. [DOI] [PubMed] [Google Scholar]
- Stear M.J., Bishop S.C., Henderson N.G., Scott I. A key mechanism of pathogenesis in sheep infected with the nematode Teladorsagia circumcincta. Anim. Health Res. Rev. 2003;4:45–52. doi: 10.1079/ahrr200351. [DOI] [PubMed] [Google Scholar]
- Stevenson L.A., Gasser R.B., Chilton N.B. The ITS-2 rDNA of Teladorsagia circumcincta, T. trifurcata and T. davtiani (Nematoda: Trichostrongylidae) indicates that these taxa are one species. Int. J. Parasitol. 1996;26:1123–1126. [PubMed] [Google Scholar]
- Stoll N.R. Investigations on the control of hookworm disease. XV. An effective method of counting hookworm eggs in faeces. Am. J. Epidemiol. 1923;3:59–70. [Google Scholar]
- Sutherland I.A., Lee D.L. A larval paralysis assay for the detection of thiabendazole resistance in trichostrongyles. Parasitology. 1990;100:131–135. doi: 10.1017/s0031182000060200. [DOI] [PubMed] [Google Scholar]
- Taylor M.A. A larval development test for the detection of anthelmintic resistance in nematodes of sheep. Res. Vet. Sci. 1990;49:198–202. [PubMed] [Google Scholar]
- Taylor D.M., Thomas R.J. The development of immunity to Nematodirus battus in lambs. Int. J. Parasitol. 1986;16:43–46. doi: 10.1016/0020-7519(86)90063-9. [DOI] [PubMed] [Google Scholar]
- Taylor M.A., Hunt K.R., Goodyear K.L. Anthelmintic resistance detection methods. Vet. Parasitol. 2002;103:183–194. doi: 10.1016/s0304-4017(01)00604-5. [DOI] [PubMed] [Google Scholar]
- Taylor M.A., Coop R.L., Wall R.L. third ed. Blackwell Publishing; Oxford, UK: 2007. Veterinary Parasitology. [Google Scholar]
- Taylor M.A., Learmount J., Lunn E., Morgan C., Craig B.H. Multiple resistance to anthelmintics in sheep nematodes and comparison of methods used for their detection. Small Rumin. Res. 2009;86:67–70. [Google Scholar]
- Thienpont D., Rochette F., Vanparijs O.F.J. second ed. Janssen Research Foundation; Beerse: 1986. Diagnosing Helminthiasis by Coprological Examination. [Google Scholar]
- Thrusfield M.V. third ed. Blackwell Science Ltd.; Oxford, UK: 2005. Veterinary Epidemiology. [Google Scholar]
- van Lieshout L., Verweij J.J. Newer diagnostic approaches to intestinal protozoa. Curr. Opin. Infect. Dis. 2010;23:488–493. doi: 10.1097/QCO.0b013e32833de0eb. [DOI] [PubMed] [Google Scholar]
- van Wyk J.A., Bath G.F. The FAMACHA system for managing haemonchosis in sheep and goats by clinically identifying individual animals for treatment. Vet. Res. 2002;33:509–529. doi: 10.1051/vetres:2002036. [DOI] [PubMed] [Google Scholar]
- van Wyk J.A., Cabaret J., Michael L.M. Morphological identification of nematode larvae of small ruminants and cattle simplified. Vet. Parasitol. 2004;119:277–306. doi: 10.1016/j.vetpar.2003.11.012. [DOI] [PubMed] [Google Scholar]
- Vercruysse J., Claerebout E. Treatment vs. non-treatment of helminth infections in cattle: defining the threshold. Special Issue: Promoting Advancement, Preserving Tradition; Plenary Papers of the 18th International Conference of the World Association for the Advancement of Veterinary Parasitology, Stresa, Italy, 26–30 August; 2001. pp. 195–214. [DOI] [PubMed] [Google Scholar]
- Verweij J.J., Brienen E.A.T., Ziem J., Yelifari L., Polderman A.M., Van Lieshout L. Simultaneous detection and quantification of Ancylostoma duodenale, Necator americanus, and Oesophagostomum bifurcum in fecal samples using multiplex real-time PCR. Am. J. Trop. Med. Hyg. 2007;77:685–690. [PubMed] [Google Scholar]
- Villanua D., Perez-Rodriguez L., Gortazar C., Hofle U., Vinuela J. Avoiding bias in parasite excretion estimates: the effect of sampling time and type of faeces. Parasitology. 2006;133:251–259. doi: 10.1017/S003118200600031X. [DOI] [PubMed] [Google Scholar]
- von Samson-Himmelstjerna G. Molecular diagnosis of anthelmintic resistance. Vet. Parasitol. 2006;136:99–107. doi: 10.1016/j.vetpar.2005.12.005. [DOI] [PubMed] [Google Scholar]
- von Samson-Himmelstjerna G., Harder A., Schnieder T. Quantitative analysis of ITS2 sequences in trichostrongyle parasites. Int. J. Parasitol. 2002;32:1529–1535. doi: 10.1016/s0020-7519(02)00163-7. [DOI] [PubMed] [Google Scholar]
- von Samson-Himmelstjerna G., Buschbaum S., Wirtherle N., Pape M., Schnieder T. TaqMan minor groove binder real-time PCR analysis of beta-tubulin codon 200 polymorphism in small strongyles (Cyathostomin) indicates that the TAC allele is only moderately selected in benzimidazole-resistant populations. Parasitology. 2003;127:489–496. doi: 10.1017/s0031182003003974. [DOI] [PubMed] [Google Scholar]
- von Samson-Himmelstjerna G., Coles G., Jackson F., Bauer C., Borgsteede F., Cirak V., Demeler J., Donnan A., Dorny P., Epe C., Harder A., Höglund J., Kaminsky R., Kerboeuf D., Küttler U., Papadopoulos E., Posedi J., Small J., Várady M., Vercruysse J., Wirtherle N. Standardization of the egg hatch test for the detection of benzimidazole resistance in parasitic nematodes. Parasitol. Res. 2009;105:825–834. doi: 10.1007/s00436-009-1466-1. [DOI] [PubMed] [Google Scholar]
- Waller P.J. The development of anthelmintic resistance in ruminant livestock. Acta Trop. 1994;56:233–243. doi: 10.1016/0001-706x(94)90065-5. [DOI] [PubMed] [Google Scholar]
- Waller P.J. Anthelmintic resistance. Vet. Parasitol. 1997;72:391–412. doi: 10.1016/s0304-4017(97)00107-6. [DOI] [PubMed] [Google Scholar]
- Waller P.J., Donald A.D., Dobson R.J. Arrested development of intestinal Trichostrongylus spp. in grazing sheep and seasonal changes in the relative abundance of T. colubriformis and T. vitrinus. Res. Vet. Sci. 1981;30:213–216. [PubMed] [Google Scholar]
- Waller P.J., Donald A.D., Dobson R.J., Lacey E., Hennessy D.R., Allerton G.R., Prichard R.K. Changes in anthelmintic resistance status of Haemonchus contortus and Trichostrongylus colubriformis exposed to different anthelmintic selection pressures in grazing sheep. Int. J. Parasitol. 1989;19:99–110. doi: 10.1016/0020-7519(89)90027-1. [DOI] [PubMed] [Google Scholar]
- Wang W., Chen K., Xu C. DNA quantification using EvaGreen and a real-time PCR instrument. Anal. Biochem. 2006;356:303–305. doi: 10.1016/j.ab.2006.05.027. [DOI] [PubMed] [Google Scholar]
- Weston K.M., O'Brien R.W., Prichard R.K. Respiratory metabolism and thiabendazole susceptibility in developing eggs of Haemonchus contortus. Int. J. Parasitol. 1984;14:159–164. doi: 10.1016/0020-7519(84)90043-2. [DOI] [PubMed] [Google Scholar]
- Whitlock H.V. A method for preventing the development of strongylid eggs in sheep faeces during transport and storage. J. Counc. Sci. Ind. Res. 1943;16:215–216. [Google Scholar]
- Whitlock H.V. Some modifications of the McMaster helminth egg-counting technique and apparatus. J. Counc. Sci. Ind. Res. 1948;21:177–180. [Google Scholar]
- Whitlock H.V. An improved method for the culture of nematode larvae in sheep faeces. Aust. Vet. J. 1956;32:141–143. [Google Scholar]
- Whitlock H.V. The recovery and identification of the first stage larvae of sheep nematodes. Aust. Vet. J. 1959;35:310–316. [Google Scholar]
- Williams J.F., Soulsby E.J.L. Antigenic analysis of developmental stages of Ascaris suum. I. Comparison of eggs, larvae and adults. Exp. Parasitol. 1970;27:150–162. doi: 10.1016/s0014-4894(70)80019-4. [DOI] [PubMed] [Google Scholar]
- Wilson I.G. Inhibition and facilitation of nucleic acid amplification. Appl. Environ. Microbiol. 1997;63:3741–3751. doi: 10.1128/aem.63.10.3741-3751.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wimmer B., Craig B.H., Pilkington J.G., Pemberton J.M. Non-invasive assessment of parasitic nematode species diversity in wild Soay sheep using molecular markers. Int. J. Parasitol. 2004;34:625–631. doi: 10.1016/j.ijpara.2003.11.022. [DOI] [PubMed] [Google Scholar]
- Winterrowd C.A., Pomroy W.E., Sangster N.C., Johnson S.S., Geary T.G. Benzimidazole-resistant [beta]-tubulin alleles in a population of parasitic nematodes (Cooperia oncophora) of cattle. Vet. Parasitol. 2003;117:161–172. doi: 10.1016/j.vetpar.2003.09.001. [DOI] [PubMed] [Google Scholar]
- Wittwer C.T., Reed G.H., Gundry C.N., Vandersteen J.G., Pryor R.J. High-resolution genotyping by amplicon melting analysis using LCGreen. Clin. Chem. 2003;49:853–860. doi: 10.1373/49.6.853. [DOI] [PubMed] [Google Scholar]
- Wolstenholme A.J., Fairweather I., Prichard R., von Samson-Himmelstjerna G., Sangster N.C. Drug resistance in veterinary helminths. Trends Parasitol. 2004;20:469–476. doi: 10.1016/j.pt.2004.07.010. [DOI] [PubMed] [Google Scholar]
- Woolaston R.R. Selection of Merino sheep for increased and decreased resistance to Haemonchus contortus: peri-parturient effects on faecal egg counts. Int. J. Parasitol. 1992;22:947–953. doi: 10.1016/0020-7519(92)90052-m. [DOI] [PubMed] [Google Scholar]
- Young R.R., Anderson N. The ecology of the free-living stages of Ostertagia ostertagi in a winter rainfall region. Aust. J. Agr. Res. 1981;32:371–388. [Google Scholar]
- Zajac A.M. Gastrointestinal nematodes of small ruminants: life cycle, anthelmintics, and diagnosis. Vet. Clin. North Am. Food Anim. Pract. 2006;22:529–541. doi: 10.1016/j.cvfa.2006.07.006. [DOI] [PubMed] [Google Scholar]
- Zarlenga D.S., Barry Chute M., Gasbarre L.C., Boyd P.C. A multiplex PCR assay for differentiating economically important gastrointestinal nematodes of cattle. Vet. Parasitol. 2001;97:199–209. doi: 10.1016/s0304-4017(01)00410-1. [DOI] [PubMed] [Google Scholar]
- Zarlenga D.S., Gasbarre L.C., Boyd P., Leighton E., Lichtenfels J.R. Identification and semi-quantitation of Ostertagia ostertagi eggs by enzymatic amplification of ITS-1 sequences. Vet. Parasitol. 1998;77:245–257. doi: 10.1016/s0304-4017(98)00114-9. [DOI] [PubMed] [Google Scholar]
- Zarlenga D.S., Higgins J. PCR as a diagnostic and quantitative technique in veterinary parasitology. Vet. Parasitol. 2001;101:215–230. doi: 10.1016/s0304-4017(01)00568-4. [DOI] [PubMed] [Google Scholar]
- Zarlenga D.S., Hoberg E.P., Stringfellow F., Lichtenfels J.R. Comparisons of two polymorphic species of Ostertagia and phylogenetic relationships within the Ostertagiinae (Nematoda: Trichostrongyloidea) inferred from ribosomal DNA repeat and mitochondrial DNA sequences. J. Parasitol. 1998;84:806–812. [PubMed] [Google Scholar]
- Zarlenga D.S., Stringfellow F., Nobary M., Lichtenfels J.R. Cloning and characterization of ribosomal RNA genes from three species of Haemonchus (Nematoda: Trichostrongyloidea) and identification of PCR primers for rapid differentiation. Exp. Parasitol. 1994;78:28–36. doi: 10.1006/expr.1994.1003. [DOI] [PubMed] [Google Scholar]