

Abstract
The foundations of the science of medical virology are intertwined with the other life sciences, particularly microbiology and infectious diseases. Medical virology has a relatively brief history, spanning just over a century, but it is crowded with intriguing discoveries, stories of immense personal courage and numerous practical applications, many of which have had an overwhelmingly positive benefit on humankind. Its origins involved the replacement of centuries-old beliefs and theories with discoveries borne out of rigorous scientific investigation. Targeted prevention and control strategies could only be developed and implemented once the concept of the specificity of disease causation had been accepted, namely that infectious diseases are caused not by some common miasma (a mysteriously poisonous substance), but rather by specific agents. In a wider sense, the microbial sciences have played a pivotal role in the development of medical thought, particularly in applying scientific rigor in understanding pathological processes. Advances in understanding of infectious agents have led to improvements in human health and well-being that arguably have exceeded the contribution of any other branch of science. Indeed, many workers in this field have been awarded the Nobel Prize in Physiology or Medicine in recognition of their achievements.
Keywords: Virology, history, virosphere, nature of viruses, microorganisms
Infectious disease is one of the few genuine adventures left in the world. The dragons are all dead and the lance grows rusty in the chimney corner…. About the only sporting proposition that remains unimpaired by the relentless domestication of a once free-living human species is the war against those ferocious little fellow creatures, which lurk in the dark corners and stalk us in the bodies of rats, mice, and all kinds of domestic animals; which fly and crawl with the insects, and waylay us in our food and drink and even in our love.
So wrote the great microbiologist Hans Zinsser in his book Rats, Lice and History, written in 1935, as he reflected on his life in infectious disease research. Zinsser’s thoughts have stimulated generations of students and professionals ever since. Infectious diseases of today present challenges that are different but just as demanding as those facing Zinsser over 80 years ago.
This book presents the subject of medical virology from the perspective of its traditional base as a life science and its application to clinical practice and public health. It is the perspective established by Frank Fenner and David White, who in 1970 conceived the rationale for this book, and maintained it through the previous four editions. It is the perspective that many others have used to teach and learn medical virology.
The foundations of the science of medical virology are intertwined with the other life sciences, particularly microbiology and infectious diseases. Medical virology has a relatively brief history, spanning just over a century, but it is crowded with intriguing discoveries, stories of immense personal courage and numerous practical applications, many of which have had an overwhelmingly positive benefit on humankind. Its origins involved the replacement of centuries-old beliefs and theories with discoveries borne out of rigorous scientific investigation. Targeted prevention and control strategies could only be developed and implemented once the concept of the specificity of disease causation had been accepted, namely that infectious diseases are caused not by some common miasma (a mysteriously poisonous substance), but rather by specific agents. In a wider sense, the microbial sciences have played a pivotal role in the development of medical thought overall, particularly in applying scientific rigor in understanding pathological processes. Advances in understanding infectious agents have led to improvements in human health and well-being that arguably have exceeded the contribution of any other branch of science. Indeed, no less than 35 workers in this and closely related fields have been awarded the Nobel Prize in Physiology or Medicine in recognition of their achievements.
Infectious disease discoveries have had a profound effect on life expectancy and well-being across the world. For example, epidemics of smallpox, yellow fever, and poliomyelitis, commonplace until well into the 20th century, have been virtually eliminated by the application of various prevention and control strategies. However, hitherto unrecognized diseases have emerged over the past half-century at the rate of at least one per year. Many of the viruses dealt with in this edition were unknown when the first edition was published over 45 years ago. The epidemiology of other viruses has radically changed as humans continue to alter the environment in so many ways. Meeting the challenges posed by emerging diseases requires the medical virologist to acquire ever more increasing expertise and access to ever more complex technologies. Today diseases such as HIV/AIDS, hepatitis C, influenza, and diarrheal diseases represent significant threats to public health. Tomorrow it will be other diseases, the nature and means of control for which are largely unpredictable. One positive note is that all emerging viral diseases of recent years have been found to be caused by members of previously recognized families of viruses. Thus a thorough knowledge of representative members of each family is likely to facilitate and inform the rapid development of knowledge about any new pathogen.
Why Study Virology?
As many bacterial infections have succumbed to treatment with antibiotics, viral infections now pose proportionally a much greater threat to global public health than was the case, say, a half-century ago. Viral diseases exact a particularly heavy toll among young children and infants in the economically less developed nations where healthcare resources are limited. Ironically, there is a resurgence of interest in viruses that target bacteria (bacteriophages) as an alternative strategy for the control of some increasingly drug-resistant bacterial infections (e.g., cholera).
Although this book focuses on viral infections of medical significance, the reader needs to be aware that viruses are a major threat to livestock and plant species, and thereby of great importance in human nutrition and food supply. Human adaptation to diseases of livestock and crops has played a major role in the development of all civilizations.
Virology is much broader than linking a particular disease to a specific pathogen: there are literally hundreds of new viruses being discovered that do not apparently relate to any known pathological condition of either animals or humans. Many of these may in the future be linked to human illnesses and thus the reader needs to be aware of the wider scope of the virological landscape, if not in detail at least to the point of “expecting the unexpected.” Conversely, the tantalizing goal remains to clarify what role, if any, viruses may play in well-known diseases of uncertain etiology, for example, multiple sclerosis.
The vast majority of new viral threats emerging annually either originate from an animal host (zoonosis) or are the result of host range extension (that is, “host species jumping”), or other changes in the epidemiology, ecology, and/or pathogenicity of the etiological agent. Since the last edition of this book, virus emergence has become a major focus of virological research.
The discovery of a new human pathogen often stimulates the discovery of related, but hitherto unidentified agents that may, or may not, present threats to human health at some point in the future. A prime example is the emergence of SARS virus, a coronavirus, and the subsequent explosion in our knowledge of coronaviruses of animals. This helped in the later rapid recognition of another human respiratory coronavirus—MERS coronavirus (Middle East Respiratory Syndrome coronavirus).
A Brief History of Virology
The history of virology can be divided into a number of eras: these span (1) the discovery of viruses as entities distinct from other disease-causing pathogens, (2) the association of many major human diseases with causative viruses, (3) the development of methods for virus isolation and characterization, (4) the defining of the chemical properties of viruses, and (5) the design and application of vaccines and therapeutics. A summary of the major milestones in the development of virology is given in Table 1.1 .
Table 1.1.
Some Milestones in the History of Virology
| Date | Discoverer(s) | Discovery(ies) |
|---|---|---|
| 1796 | E. Jenner | Application of cowpox virus for vaccination against smallpox |
| 1885 | L. Pasteur | Development of rabies vaccine |
| 1892 | D. Ivanovsky, M. Beijerinck | Ultrafiltration of tobacco mosaic virus |
| 1898 | F. Loeffler, P. Frosch | Ultrafiltration of foot-and-mouth disease virus—clear proof of virus etiology of disease—discovery of the first virus |
| 1898 | G. Sanarelli | Discovery of myxoma virus |
| 1900 | W. Reed, J. Carroll, A. Agramonte, J. Lazear, C. Finlay | Discovery of yellow fever virus and its transmission by mosquitoes |
| 1903 | M. Remlinger, Riffat-Bay, A. di Vestea | Discovery of rabies virus |
| 1907 | P. Ashburn, C. Craig | Discovery of dengue viruses |
| 1909 | K. Landsteiner, E. Popper | Discovery of polioviruses |
| 1911 | P. Rousa | Discovery of the first tumor virus: Rous sarcoma virus |
| 1911 | J. Goldberger, J. Anderson | Discovery of measles virus |
| 1915 | F. Twort, F. d’Herelle | Discovery of bacterial viruses (bacteriophages) |
| 1918 | Beginning of global pandemic of influenza | |
| 1919 | A. Löwenstein | Discovery of herpes simplex virus |
| 1930 | K. Meyer, C. Haring, B. Howitt | Discovery of Western equine encephalitis virus |
| 1931 | M. Theilera | Attenuation of yellow fever virus—vaccine development |
| 1933 | C. Andrews, P. Laidlaw, W. Smith | Isolation of human influenza viruses in ferrets |
| 1933 | R. Muckenfuss, C. Armstrong, H. McCordock, L. Webster, G. Fite | Discovery of St. Louis encephalitis virus |
| 1934 | C. Johnson, E. Goodpasture | Discovery of mumps virus |
| 1934 | M. Hayashi, S. Kasahara, R. Kawamura, T. Taniguchi | Discovery of Japanese encephalitis virus |
| 1935 | W. Stanleya | Purification/crystallization of tobacco mosaic virus |
| 1936 | C. Armstrong, T. Rivers, E. Traub | Discovery of lymphocytic choriomeningitis virus |
| 1937 | L. Zilber, M. Chumakov, N. Seitlenok, E. Levkovich | Discovery of tick-borne encephalitis virus (Russian spring summer encephalitis virus) |
| 1938 | B. von Borries, H. Ruska, E. Ruska | First electron micrograph of viruses (ectromelia, vaccinia viruses) |
| 1939 | E. Ellis, M. Delbrück | Development of one-step growth curve—bacteriophage |
| 1940 | K. Smithburn, T. Hughes, A. Burke, J. Paul | Discovery of West Nile virus |
| 1941 | G. Hirst | Discovery of agglutination of red blood cells by influenza virus |
| 1945 | M. Chumakov, G. Courtois, colleagues | Discovery of Crimean-Congo hemorrhagic fever virus |
| 1948 | G. Dalldorf, G. Sickles | Discovery of Coxsackieviruses |
| 1949 | J. Endersa, T. Wellera, F. Robbinsa | Development of cell culture methodology for polio, measles, and other vaccines |
| 1950 | L. Florio, M. Miller, E. Mugrage | Discovery of Colorado tick fever virus |
| 1952 | R. Dulbecco, M. Vogt | Development of plaque assay for animal viruses—polioviruses, Western equine encephalitis virus |
| 1953 | W. Rowe | Discovery of human adenoviruses |
| 1954 | J. Salk, J. Youngner, T. Francis | Development of inactivated polio vaccine |
| 1958 | J. Lederberga | Discovery of genetic recombination and the organization of the genetic material of bacteria |
| 1959 | A. Sabin, H. Cox, H. Koprowski | Development of attenuated live-virus polio vaccine |
| 1962 | A. Lwoff, R. Horne, P. Tournier | Classification of the viruses based on virion characteristics |
| 1964 | M. Epstein, B. Achong, Y. Barr | Discovery of Epstein–Barr virus and its association with Burkitt’s lymphoma |
| 1965 | D. Tyrrell, M. Bynoe, J. Almeida | Discovery of human coronaviruses (B814 and 229E) |
| 1965 | F. Jacoba, A. Lwoffa, J. Monoda | Discoveries of genetic control of enzymes and virus synthesis: the operon |
| 1967 | B. Blumberga, H. Alter, A. Prince | Discovery of Australia antigen and its link to hepatitis B |
| 1969 | M. Delbrücka, A. Hersheya, S. Luriaa | Discoveries related to the replication mechanism and the genetic structure of viruses |
| 1970 | H. Temina, D. Baltimorea, R. Dulbeccoa | Discoveries related to the interaction between tumor viruses and the genetic material of the cell—reverse transcriptase |
| 1972 | A. Kapikian, colleagues | Discovery of Norwalk virus (norovirus) |
| 1973 | R. Bishop, G. Davidson, I. Holmes, T. Flewett, A. Kapikian | Discovery of human rotaviruses |
| 1973 | S. Feinstone, A. Kapikian, R. Purcell | Discovery of hepatitis A virus |
| 1975 | Y. Cossart, A. Field, A. Cant, D. Widdows | Discovery of parvovirus B-19 and its association with aplastic crisis in hemolytic anemia |
| 1975 | P. Sharpa, L. Chow, R. Robertsa, T. Broker | Discovery of RNA splicing and split genes (adenovirus) |
| 1976 | D. C. Gajduseka | Discovery of transmissible spongiform encephalopathies |
| 1976 | K. Johnson, P. Webb, J. Lange, F. Murphy, S. Pattyn, W. Jacob, G. Van der Groen, P. Piot, E. Bowen, G Platt, G. Lloyd, A. Baskerville, D. Simpson | Discovery of Ebola virus |
| 1976 | J. Bishopa, H. Varmusa | Discovery of the cellular origin of retroviral oncogenes |
| 1977 | D. Henderson, F. Fenner, I. Arita, many others | Global eradication of smallpox |
| 1978 | D. Nathansa, W. Arbera, H. Smitha | Discovery of restriction enzymes and their application to problems of molecular genetics |
| 1978 | S. Harrison, M. Rossman, N. Olson, R. Kuhn, T. Baker, J. Hogle, M. Chow, R. Rueckert, J. Johnson | Atomic structure of viruses (tomato bushy stunt virus, polioviruses, rhinoviruses) |
| 1980 | P. Berga | The development of recombinant-DNA technology |
| 1980 | R. Gallo, B. Poiesz, M. Yoshida, I. Miyoshi, Y. Hinuma | Discovery of human T lymphotropic viruses 1 and 2 |
| 1981 | V. Racaniello, D. Baltimore | Development of an infectious recombinant clone of a virus (poliovirus) |
| 1982 | S. Prusinera | Concept of the prion and their etiologic role in spongiform encephalopathies |
| 1982 | A. Kluga | Crystallographic electron microscopy and structural elucidation of biologically important nucleic acid–protein complexes |
| 1983 | F. Barré-Sinoussia, L. Montagniera, J. Chermann | Discovery of human immunodeficiency virus 1 (HIV1) |
| 1983 | M. Balayan | Discovery of hepatitis E virus and its transmission |
| 1985 | F. Barin, F. Clavel, M. Essex, P. Kanki, F. Brun-Vézinet | Discovery of human immunodeficiency virus 2 (HIV2) |
| 1988 | G. Hitchingsa, G. Eliona | Discoveries of important principles for drug treatment—acyclovir |
| 1989 | M. Houghton, Q.-L. Choo, G. Kuo, D. Bradley, H. Alter | Discovery of hepatitis C virus |
| 1993 | S. Nichol, C. Peters, P. Rollin, T. Ksiazek | Discovery of Sin Nombre virus and its association with hantavirus cardiopulmonary syndrome |
| 1994 | Y. Chang, P. Moore | Discovery of human herpesvirus 8—Kaposi sarcoma herpesvirus |
| 1995 | K. Murray, P. Hooper, A. Hyatt | Discovery of Hendra virus and its reservoir host fruit bats |
| 1996 | P. Dohertya, R. Zinkernagela | Discovery of the genetic specificity of the cell-mediated immune response |
| 1996 | R. Will, J. Ironside, J. Collinge, colleagues | Discovery that bovine spongiform encephalopathy prion is the cause of variant Creutzfeldt–Jakob disease in humans |
| 1999 | K. Chua, S. Lam, W. Bellini, T. Ksiazek, B. Eaton, colleagues | Discovery of Nipah virus |
| 1999 | D. Asnis, M. Layton, W.I. Lipkin, R. Lanciotti | Extension of West Nile virus range to North America |
| 2001 | B. van den Hoogen, A. Osterhaus, colleagues | Discovery of human metapneumovirus |
| 2003 | C. Urbani, J. Peiris, S. Lai, L. Poon, G. Drosten, K. Stöhr, A. Osterhaus, T. Ksiazek, D. Erdman, C. Goldsmith, S. Zaki, J. DeRisi, others | Discovery of SARS coronavirus |
| 2003 | B. La Scola, D. Raoult, others | Discovery of mimivirus, the largest virus known at the time |
| 2005 | J. Taubenberger, P. Palese, T. Tumpey, A. Garcia-Sastre, others | 1918 influenza virus genome sequenced and the virus reconstructed |
| 2005 | Beginning of global pandemic of chikungunya | |
| 2005 | E. Leroy, J. Towner, R. Swanepoel, others | Discovery that the reservoir hosts of Ebola/Marburg viruses are bats |
| 2007 | T. Allander, D. Wang, Y. Chang, others | Discovery of human polyomaviruses KI, WU, MC |
| 2008 | H. zur Hausena | Discovery that human papilloma viruses cause cervical cancer |
| 2008 | B. La Scola, D. Raoult, others | Discovery of virophage, Sputnik |
| 2010 | W. Plowright and the FAO Global Rinderpest Eradication Programme | Global eradication of rinderpest |
| 2011 | B. Hoffmann, M. Beer, T. Mettenleiter, colleagues | Discovery of Schmallenberg virus |
| 2012 | A.M. Zaki, R. Fouchier, W.I. Lipkin | Discovery of MERS coronavirus |
| 2014 | Beginning of an Ebola hemorrhagic fever epidemic in West Africa, the largest ever | |
| 2015 | Beginning of a global epidemic of Zika virus disease—discovery of microcephaly as consequence of in utero infection |
Scientists who were awarded the Nobel Prize for their work—cited at date of the discovery rather than the date of award.
Virology has its foundations in the initial discoveries of bacteria and related diseases. Up to the 19th century the prevailing view was that diseases of humans and animals were the result of miasmas and other environmental influences. This was despite the thesis of Girolamo Fracastoro who suggested as early as 1546 that epidemic diseases were disseminated by minute particles carried over long distances. Anton van Leeuwenhoek first saw bacteria through his microscope in 1676 and Lazarro Spallanzani first grew bacteria in culture in 1775. Remarkably, Edward Jenner developed vaccination against smallpox in 1796 against a backdrop of prevailing opinion that such diseases were caused by environmental factors rather than specific microscopic agents.
The establishment of microbiology as a scientific discipline owes much to the work of Louis Pasteur, who in 1857 discovered the specificity of microbial fermentation, who then went on in 1865 to elaborate the nature of diseases of silkworms. But it was his work on rabies that signaled the start of the virus discovery era. In 1885, Pasteur looked on as his first rabies vaccine was given to a boy, Joseph Meister, bitten severely by a rabid dog, thus opening up the strategy of vaccine development through a process of virus attenuation (Fig. 1.1 ).
Figure 1.1.

In 1881 and 1882, Louis Pasteur, Charles Chamberland, Émile Roux, and Louis Thuillier began their research toward developing a rabies vaccine. They modified Pierre-Victor Galtier’s technique by inoculating nervous tissue from a rabid dog through a long series of dogs via subdural trephination. After many passages, they obtained a virus of maximum virulence and with a fixed incubation period of about 10 days. The degree of attenuation of virus recovered from each passage was measured and virus was then further attenuated in rabbits. This final attenuation procedure consisted of suspending the spinal cord of a rabid rabbit in a flask, in a warm dry atmosphere, to achieve slow desiccation. They succeeded in producing “attenuated viruses of different strengths,” the weakest of which could be used to prepare the first dose of a vaccine. Inoculating dogs with a sequence of spinal cords of increasing virulence rendered the recipients resistant to inoculation with fully virulent virus. Within a year, Pasteur and his colleagues reported the results of this treatment in 350 cases of rabies exposure—only one person developed rabies, and this a child who was treated 6 days after exposure. Over the next decades many thousands of people with potential rabies exposures were immunized with ever-improving animal nervous system (brain and spinal cord) vaccines, at the Institut Pasteur in Paris, which was founded in 1888, and in other locations throughout the world.
Louis Pasteur, 1822–95. Painting by Albert Edelfeldt, 1885. From Institut Pasteur, used with permission.
The early pioneering work of the 19th century linking disease to specific bacteria was greatly assisted by the earlier development of the unglazed porcelain ultrafilter by Charles Chamberland who worked in Pasteur’s laboratory. These filters originally were used to sterilize water and other fluids by preventing the passage of bacteria. Dimitri Ivanovsky (1892) and Martinus Beijerinck (1898) showed that the agent causing mosaic disease in tobacco plants (now known to be tobacco mosaic virus [TMV]) passed through ultrafilters retaining bacteria. Beijerinck realized he was dealing with something other than a microbe but erroneously thought that the entity that passed through the ultrafilter was an infectious liquid and not a particle—he called it a “contagium vivum fluidum.” Friedrich Loeffler and Paul Frosch were the first to correctly conclude that an ultrafilterable infectious agent was indeed a submicroscopic particle. Studying the cause of foot-and-mouth disease of cattle, Loeffler and Frosch found that the causative agent passed through a Chamberland ultrafilter but not the finer Kitasato ultrafilter. Thus these first virologists saw ultrafiltration in a new way—they focused attention on what passed through the ultrafilter rather than what was retained, and thereby established an experimental methodology widely adopted in the early 20th century. In quick succession, further diseases were shown to be caused by ultrafilterable agents: in 1900 the first human virus, yellow fever virus, and its mosquito transmission cycle was discovered by Walter Reed, James Carroll and the US Army Yellow Fever Commission in Havana, Cuba, a discovery that was guided by the earlier work of the Cuban physician Carlos Findlay (Fig. 1.2 ).
Figure 1.2.

In 1900, Walter Reed and his colleagues discovered yellow fever virus, the first human virus, and its transmission cycle. This is a famous allegorical painting, entitled Conquerors of Yellow Fever by Dean Cornwell. It depicts Walter Reed (in white uniform) and Carlos Finlay (with white hair) looking on as Jesse Lazear, who died of yellow fever a month later, applies an infected mosquito to the arm of James Carroll. The painting includes Aristides Agramonte (behind Lazear), Leonard Wood (in brown helmet), Jefferson Kean (in white helmet), and several of the volunteers who subsequently were infected in the same way. Carroll became infected as a result of this experiment—he survived, and went on to have a distinguished career as a microbiologist, but suffered from chronic illness leading to an early death, said to be a consequence of his yellow fever infection.
Purchased copy, used with permission.
The concept of ultrafilterable infectious agents became more widely acceptable when Karl Landsteiner and Erwin Popper showed conclusively in 1909 that poliomyelitis was caused by an ultrafilterable agent. Importantly, as early as 1911 Peyton Rous also showed similar properties for the etiologic agent of a sarcoma of chickens: Rous sarcoma virus was to play an essential role in determining the basic mechanism by which viruses may trigger the onset of tumors.
The realization that oncogenesis and virus infection went hand in hand was an important milestone in the early days of virology, although it took many decades for its true significance to be appreciated. In 1970 Howard Temin and David Baltimore independently were able to show that oncogenic viruses contain a reverse transcriptase enzyme, thus explaining how an RNA virus could produce DNA copies of its genetic material.
Bacteriophages were independently discovered by Frederick Twort and Felix d’Herelle (1915) who investigated outbreaks of dysentery among troops of the First World War. Presciently, Twort foresaw that the clear plaques in plated Micrococcus cultures could be caused by “ultrafilterable viruses.”
During the following decades of the 20th century, it was thought by many that viruses represented infectious protein particles. This was a view reinforced by Wendell Stanley’s description in 1935 that crystals of pure TMV could be dissolved and transmit infection to healthy plants—he presumed that the crystals were pure protein. This was dispelled, however, when Frederick Bawden and Norman Pirie showed that TMV contained not only protein but also nucleic acid. The importance of this was shown by the classic studies of Oswald Avery, Colin MacLeod, and Maclyn McCarty (1944) and then Alfred Hershey and Martha Chase (1952), who proved DNA was linked to hereditary.
In 1933 the electron microscope was invented by Ernst Ruska and Max Knoll and in 1938 Bodo von Borries, Helmut Ruska, and Ernst Ruska published the first electron micrographs of ectromelia (mousepox) virus and vaccinia virus. It soon became clear that there was great diversity in the size and shape of the various viruses. A major advance was the development of negative-contrast electron microscopy in 1959 by Sydney Brenner and Robert Horne. Using this method, electron-dense stains surround virus particles to produce a negative image of the virus with remarkable resolution; importantly in those early days of medical virology, the method was simple to use. Figure 1.3, Figure 1.4 depict the diverse spectrum of morphological shapes represented by animal viruses. By the early 1960s, the fine structure of several viruses was unraveled by Aaron Klug, Donald Caspar, and others using X-ray crystallography—they showed that many viruses are constructed from uniform subunits, following the principles of icosahedral symmetry as first understood for the Platonic solids (regular polyhedra) by the ancient Greeks. Thus through the use of several different approaches the diversity of structural detail among various viruses began to emerge.
Figure 1.3.
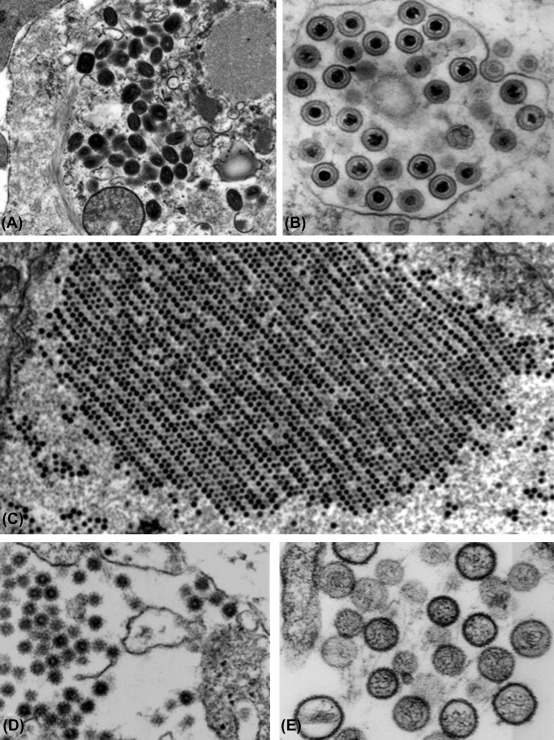
Thin-section electron microscopy of selected viruses. The remarkable diversity of the viruses is clearly revealed by thin-section electron microscopy of infected cells—and this technique provides important information about morphogenesis and cytopathology. (A) Family Poxviridae, genus Orthopoxvirus, variola virus. (B) Family Herpesviridae, genus Simplexvirus, human herpesvirus 1. (C) Family Adenoviridae, genus Mastadenovirus, human adenovirus 5. (D) Family Togaviridae, genus Alphavirus, Eastern equine encephalitis virus. (E) Family Bunyaviridae, genus Hantavirus, Sin Nombre virus. These images represent various magnifications; the details of the morphogenesis of the various viruses are given in the chapters of Part II of this book.
Figure 1.4.

Negative contrast electron microscopy of selected viruses. The remarkable diversity of the viruses is revealed by all kinds of electron microscopy methods, but none better than by negative staining. (A) Family Poxviridae, genus Orthopoxvirus, vaccinia virus. (B) Family Papovaviridae, genus Papillomavirus, human papillomavirus. (C) Family Filoviridae, Ebola virus. (D) Family Reoviridae, genus Rotavirus, human rotavirus. (E) Family Herpesviridae, genus Simplexvirus, human herpesvirus 1 (capsid only, envelope not shown). (F) Family Rhabdoviridae, genus Lyssavirus, rabies virus. (G) Family Caliciviridae, genus Norovirus, human norovirus. (H) Family Bunyaviridae, genus Phlebovirus, Rift Valley fever virus. (I) Family Orthomyxoviridae, genus Influenzavirus A, influenza virus A/Hong Kong/1/68 (H3N2). These images represent various magnifications; the size of the various viruses is given in Chapter 2: Classification of Viruses and Phylogenetic Relationships and in the chapters of Part II of this book.
Attempts to prevent virus disease using vaccines have paralleled the development of virology, beginning from the early pioneering days of Edward Jenner and Louis Pasteur. Notable developments included the attenuated yellow fever vaccine developed by Max Theiler in 1931, a vaccine that is still in widespread use today and has saved countless thousands of lives. Jonas Salk and Albert Sabin in 1954 and 1959 developed non-replicating (inactivated) and living attenuated virus vaccines against poliovirus, respectively, the use of which has been so extensive that poliovirus infection has all but been eradicated save for a few pockets of infection in remote parts of the world (Fig. 1.5 ). As is described in Chapter 11: Vaccines and Vaccination, vaccine research has often exploited novel concepts, for example, the use of plasma from chronically infected humans as a source of hepatitis B virus (HBV) envelope protein to stimulate immunity against hepatitis B virus (1976), and the use of genetically modified naked DNA preparations to induce the expression of antigens in the tissues of vaccine recipients.
Figure 1.5.

The World Health Organization Global Polio Eradication Initiative aims for global eradication of poliomyelitis by about 2018. The Initiative is led by the World Health Organization, UNICEF, and the Rotary and Gates Foundations; it has reduced the number of cases from the many thousands per year to less than 100 (359 cases in 2014; 74 cases in 2015). Polio will be the third disease globally eradicated, after smallpox and rinderpest. The most important step in polio eradication is interruption of endemic transmission by universal infant vaccination using oral vaccine (OPV; often by organizing “national immunization days”), supplementary IPV vaccination campaigns where needed, intensive surveillance of cases of flaccid paralysis, and in some places detection of virus in sewage. Figures (clockwise from top left). An Egyptian stele (slab) thought to depict a polio victim—18th Dynasty (1403–1365 BC); patients with permanent respiratory muscle paralysis after recovery from poliomyelitis would spend the rest of their lives requiring assisted respiration (immersed in an “iron lung”); patients with permanent lower limb weakness following poliomyelitis; those remaining countries reporting cases of poliomyelitis in 2014; oral administration of polio vaccine; in 1921, 39-year-old Franklin D. Roosevelt was diagnosed with poliomyelitis and was left with permanent paralysis from the waist down, but was rarely photographed in a wheelchair. He was elected US president in 1932.
In 1957, Alick Isaacs and Jean Lindemann discovered interferons, molecules that represent the initial mammalian response to infection. Great hope was placed on the use of interferons in the treatment of a wide spectrum of human virus infections: although of proven use in certain conditions, however, the use of interferons has not lived up to the earlier wide promise suggested by laboratory studies.
The sciences of immunology and cell and molecular biology have been intertwined with that of virology: landmark discoveries were made by Peter Doherty and Rolf Zinkernagel, who in 1974 discovered how the cellular immune system recognizes virus-infected cells; and Georges Kohler and Cesar Milstein, who in 1975 developed the first monoclonal antibodies.
The Virosphere
We live in what many now describe as the virosphere, since almost all living multicellular and unicellular organisms are susceptible to virus infection. Take as an example the oceans: every liter of seawater is populated with up to 10 billion viruses. It is estimated that there are around 5×1030 bacteria on planet Earth, and that viruses are numerically at least more common; this means there are more viruses in the world than all life forms. The vast majority are most likely viruses of bacteria (bacteriophages) serving to aid the recycling of organic matter, but some have a more sophisticated role in the environment, for example, determining insect behavior as an essential part of an arthropod life cycle. This staggeringly large repertoire of the virosphere is not restricted to inhabiting non-human life forms: we are only recently beginning to study the range of different viruses that humans appear to carry permanently (the human “virome,” see Chapter 39: Viral Syndromes), yet appear to cause no harmful effects. One example is the Torque teno (TT) virus, discovered by chance in 1997 during studies of “transfusion-transmitted” infection.
The Nature of Viruses
The unicellular microorganisms can be arranged in the order of decreasing size and complexity: protozoa, fungi, and bacteria (the latter including mycoplasmas, rickettsiae, and chlamydiae). These microorganisms, however small and simple, are cells. Such microorganisms contain DNA as the repository of genetic information, and also contain various species of RNA and most, if not all, of the machinery for producing energy and macromolecules. These microorganisms grow by synthesizing macromolecular constituents (nucleic acids, proteins, carbohydrates, and lipids), and most multiply by binary fission.
Viruses, on the other hand, are neither cellular nor microorganisms. The key differences between viruses and microorganisms are listed in Table 1.2 . Viruses do not possess functional organelles (e.g., mitochondria, Golgi, chloroplasts, and endoplasmic reticulum), and thus are totally dependent on the host for the machinery of energy production and synthesis of macromolecules. Viruses contain only one type of functional nucleic acid, either DNA or RNA, never both, and differ from microorganisms in having a life cycle divisible into two clearly defined phases. Outside of the host cell, the viruses are metabolically inert and can be considered as complexes of large macromolecules; during this extracellular phase of the viral life cycle, virus transmission is dependent upon movements of air and fluid, and in some cases the life cycle of insect vectors. Once inside the host cell, however, viruses behave with many of the properties of living organisms; viruses are metabolically active in that the viral genome exploits the machinery of the host to produce progeny genome copies, viral messenger RNA, and viral proteins (often along with carbohydrates and lipids), all of which are then assembled to form new virions (virion, the complete virus particle). This assembly from pools of precursor molecules is in contrast to the multiplication of cellular organisms by binary fission. In contrast to any microorganism, many viruses can reproduce even if only the viral DNA or RNA genome is introduced into the host cell. These qualities have been used to argue the question, “Are viruses alive?” One answer is to envision viruses “at the edge of life,” in some ways fulfilling the criteria we use to define life, but mostly not.
Table 1.2.
Contrasting Properties of Unicellular Microorganisms and Viruses
| Property | Bacteria | Rickettsiae | Mycoplasma | Chlamydiae | Viruses |
|---|---|---|---|---|---|
| >300 nm diametera | Yes | Yes | Yes | Yes | No |
| Growth on non-living mediab | Yes | No | Yes | No | No |
| Binary fission | Yes | Yes | Yes | Yes | No |
| Contain both DNA and RNA | Yes | Yes | Yes | Yes | Noc |
| Infectious nucleic acid | No | No | No | No | Many |
| Functional ribosomes | Yes | Yes | Yes | Yes | No |
| Sensitivity to antibiotics | Yes | Yes | Yes | Yes | Nod |
Some mycoplasmas and chlamydiae are less than 300 nm in diameter, and mimiviruses and the other new “giant DNA viruses” are greater than 300 nm in diameter.
Chlamydiae and most rickettsiae are obligate intracellular parasites.
A few viruses contain both types of nucleic acid, but one of these types acts as the main functional molecule and the other plays a minor role.
With very few exceptions.
Given the unique characteristics of viruses, where might viruses have originated? There are three principal theories that have been argued for many years. First, viruses may have originated as escaped eukaryotic genes, that is nucleic acid sequences, that evolved to encode protective protein coats to allow survival outside of the environment of the cell (transposons and retrotransposons have been suggested as the progenitors of retroviruses). Second, viruses may be degenerate forms of intracellular parasites, having lost most cellular functions (bacteria have been suggested as the progenitors of mitochondria, chloroplasts, and poxviruses); and third, viruses may have originated independently along with other primitive molecules and developed with self-replicating capabilities.
In the absence of fossil remains, insight as to virus evolution relies almost entirely on sequence analyses of virus genomes. For example, the genome of a plant viroid (a subviral agent comprised of infectious naked RNA), potato spindle tuber viroid, seems to be a self-replicating RNA copy of a part of the host potato DNA. Many of the genes of poxviruses are similar to those of eukaryote hosts. In any case, it seems certain from sequence analyses of viral genomes that all presently recognized viruses did not evolve from a single progenitor; rather, different kinds of viruses likely arose independently from different origins, and then continued to diversify and adapt survival and transmission qualities to better fit particular niches by the usual Darwinian process of mutation and natural selection.
It should be stressed that the genetic blueprint of all viruses is under continuing evolutionary pressure, sometimes showing dramatic examples of genetic change and natural selection of those variants that survive the best. Some viruses have continued to evolve in long association with each associated hosts (e.g., herpesviruses, some retroviruses); others have evolved by “host species jumping” (e.g., influenza viruses), and yet others by developing zoonotic transmission schemes (e.g., rabies virus).
Several important practical consequences follow from understanding that viruses are different from microorganisms and all life forms: for example, some viruses can persist for the lifetime of the host cell by the integration of the DNA genome (or a DNA copy of the RNA genome) into the genome of the host cell, or by the carriage of viral DNA genomes by the host cell in episomal form. Since viruses use the replicative machinery of the host, virus infections present major challenges to antiviral drug development. Drugs that interfere with viral replication nearly always interfere with essential host cell functions. This is in contrast to bacteria, which have unique metabolic pathways different from those of the host, enabling these to be exploited as targets for antibiotics.
The simplest viruses consist of a DNA or RNA genome contained within a protein coat, but there are classes of even simpler infectious agents: (1) satellites, which are defective viruses, dependent upon a helper virus to supply essential functions such as nucleic acid replication functions or structural elements such as capsid proteins; (2) viroids, which as noted above consist of a naked RNA molecule that is infectious; and (3) prions, the agents of the spongiform encephalopathies, consisting of an infectious protein without any associated nucleic acid.
Scope of This Book
From its beginning medical virology has been intertwined with many related sciences. Even though this book deals with medical virology per se—the viruses infecting humans and the diseases so caused—understanding the full scope of the subject requires a continuing appreciation and integration of related sciences, from cell biology to medical epidemiology and extending to human social behavior. The perspective represented by this book, of medical virology as an infectious disease science, is meant to provide a starting point, an anchor, for those who must relate the subject to clinical practice, public health practice, scholarly research, and other endeavors.
Part I of this book thus deals with the properties of viruses, how viruses replicate, and how viruses cause disease. These chapters are then followed by an overview of the principles of diagnosis, epidemiology, and how virus infections can be controlled. This first section is concluded by a discussion on emergence and attempts to predict the next major public health challenges. These form a guide for delving into the specific diseases of interest to the reader as described in Part II.
Further Reading
- Crawford D.H. A Very Short Introduction to Viruses. Oxford University Press; Oxford: 2011. [Google Scholar]
- Flint S.J., Racaniello V.R., Rall G.F., Skalka A.M., Enquist L. Principles of Virology, two volumes. fourth ed. ASM Press; Washington, DC: 2015. ISBN-10: 1555819516. [Google Scholar]
- Kaslow R.A., Stanberry L.R., LeDuc J.W. Viral Infections of Humans, Epidemiology and Control. fifth ed. Springer; New York: 2014. ISBN 978-1-4899-7447-1. [Google Scholar]
- Knipe D.M., Howley P.M. Field’s Virology. sixth ed. Lippincott Williams and Wilkins; Philadelphia: 2013. ISBN 978-145-110563-6. [Google Scholar]
- MacLachlan N.J., Dubovi E.J. Fenner’s Veterinary Virology. fourth ed. Academic Press; London: 2011. ISBN 978-0-12-375158-4. [Google Scholar]
- Waterson A.P., Wilkinson L. An Introduction to the History of Virology. Cambridge University Press; Cambridge: 1978. ISBN 978-0-521-21917-5. [Google Scholar]
- Zimmer C. A Planet of Viruses. University of Chicago Press; Chicago: 2011. ISBN 978-0-226-98335-6. [Google Scholar]
- Zinsser H. Rats, Lice and History. Transaction Publishers; New Brunswick, USA: 1935, reprinted 2007. ISBN 978-1-4128-0672-5. [Google Scholar]