

INTRODUCTION
Definition
Plague in humans is a severe febrile illness caused by the gram-negative bacillus, Yersinia pestis. Historically, it is synonymous with catastrophic epidemics. Y. pestis is maintained in nature as a zoonotic infection of rodents and their fleas in scattered foci in large areas of Asia, Africa, and the Americas. It is an incidental infection of humans and mammals other than rodents. Humans acquire infection most often by the bite of rodent fleas, occasionally by handling or ingesting infected animal tissues, or by inhaling contagious airborne particles. The vast majority of cases of human plague now occur in underserved Third World populations where sanitation is poor and persons live in close association with rodent reservoirs of infection. The principal clinical forms of plague are bubonic, septicemic, and pneumonic. Bubonic (lymphadenitic) plague accounts for 85% or more of primary plague cases. Septicemic and pneumonic plagues occur most often as secondary complications of bubonic plague, but may arise directly from primary infections. All forms of plague are curable if they are diagnosed and treated early. However, delays in treatment can rapidly lead to potentially fatal overwhelming infection and endotoxemia. Although plague has a high epidemic potential, preventive steps can be taken to reduce risks of exposure, and outbreaks can be readily halted using standard public health measures. Cases should be reported to health authorities immediately so that appropriate investigative and control measures can be implemented without delay. Reporting and control of the disease is mandated by national and international health regulations. Recently, concern has been raised over plague as a potential weapon of terrorism; Y. pestis has been classified as a Category A select agent subject to federal laws governing its management and transport. Medical and public health preparedness and response plans have been developed to counter the threat of its potential misuse.1, 2
History
Three pandemics of plague have been recorded. In the Justinian pandemic (circa 542–767 ad), it is thought that plague moved from upper Egypt to the Mediterranean and spread from there to Europe and Asia Minor, ultimately causing an estimated 40 million deaths. The second plague pandemic began in Central Asia early in the 14th century, caused epidemics in China and India, and moved along caravan routes to the Near and Middle East. Entering Messina by ship in 1347, plague swept swiftly through Europe and the British Isles, receiving the epithet “Black Death.” Medieval plague killed a quarter or more of the affected populations at its height and was followed by successive epidemics in the European region over the next several centuries. The last global surge of plague, the third (modern) pandemic, arose in the latter half of the 19th century in Yunnan Province, China, struck Hong Kong in 1894, and spread from there by rat-infested steamships to port cities throughout the world, including several in the United States.3, 4, 5 Within 30 years of its appearance in Hong Kong, the third plague pandemic had resulted in 26 million human cases and more than 12 million deaths, most of them in India.3
The plague bacillus was first identified in Hong Kong in 1894 by Alexandre Yersin, who isolated the organism from enlarged lymph nodes (buboes) of plague victims.6 In 1898, Paul-Louis Simond, a French scientist sent to investigate epidemic plague in Bombay, identified the plague bacillus in the tissues of dead rats and subsequently proposed that the organism was transmitted from rat to rat, and from rats to humans by rat fleas.
After 1900, the global spread of plague was limited by regulations that controlled rats in ports and imposed inspection and rat-proofing of ships. Y. pestis did, however, become newly established among urban and rural rodent populations in many previously unaffected areas of the Americas, Europe, Africa, and Asia, resulting in scattered zoonotic foci that still exist throughout the world today.7, 8 In San Francisco, between 1900 and 1908, major outbreaks of rat-borne plague killed more than 200 persons. By 1908, plague was epizootic in ground squirrels in counties surrounding the city,5 and in subsequent years spread to wild rodent populations throughout California and other states in the western third of the country. The last outbreak of urban plague and of person-to-person pneumonic transmission in the United States occurred in a slum area of Los Angeles in 1924–1925.
By the middle of the 20th century, cities in the United States and elsewhere had enforced higher sanitary standards and building codes, effective insecticides and rodenticides had become widely available, and several classes of antibiotics had been shown to be efficacious in treating plague. Most human plague since then has been sporadic and rural in distribution. Outbreaks have been relatively slow to develop and readily controlled by a combination of surveillance, early diagnosis and treatment, and flea and rat suppression.9 The major exceptions to this generalization were the large rat-borne plague epidemics that occurred from 1962 to 1975 in war-torn Vietnam and in the 1990s in Madagascar.6, 10
Plague is one of the three diseases (plague, cholera, yellow fever) subject to quarantine under World Health Organization (WHO) international health regulations.11 In 1994, articles of these regulations were used to prevent potential spread of plague from India after a reported outbreak of pneumonic plague there evoked local panic and an exaggerated international response to the perceived health emergency.12, 13
AGENT
General Microbiology
Y. pestis is a nonmotile, nonsporulating, gram-negative coccobacillus in the family Enterobacteriaceae.4, 14 It is microaerophilic, oxidase- and urease-negative, non–lactose-fermenting, and biochemically unreactive. The organism is nonfastidious and is highly infective to laboratory rodents. Y. pestis grows slowly but well on a wide variety of common media (e.g., brain-heart infusion broth, sheep blood agar, chocolate agar, and MacConkey agar). Routine specimens can be safely handled under BSL-2 procedures, but manipulation of isolates requires BSL-3 measures to adequately protect laboratory workers.14 Growth occurs across a wide range of temperature (4°C to 40°C) and pH (5.0 to 9.6), but is optimal at 28°C and pH 7.4. Incubation at 37°C for 24 hours on agar yields pinpoint, transparent colonies that are easily overlooked, especially if there is contamination of the culture by other bacteria. At 48 hours, colonies are typically gray, 1 mm to 2 mm in diameter, and have an irregular “hammered metal” appearance when viewed under magnification. In broth, Y. pestis grows in flocculent clumps, typically attached to the sides of the tube in downwardly projecting stalactite forms that leave a clear broth. With polychromatic stains such as Wayson, Wright, or Giemsa, plague bacilli from clinical specimens demonstrate a characteristic bipolar appearance resembling closed safety pins (Fig. 42-1 ). Although Y. pestis is not truly encapsulated, it produces an envelope that contains the unique fraction 1 (F1) glycoprotein surface antigen.
FIGURE 42-1.
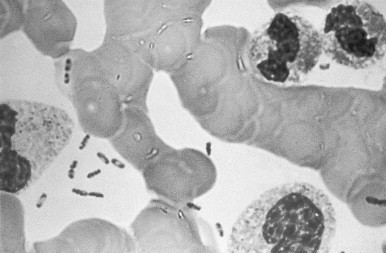
Peripheral blood smear from a patient with plague septicemia, showing characteristic bipolar-staining Yersinia pestis (Wright's stain, oil immersion).
Decoding of the entire genome of the plague bacillus shows that it only recently evolved from Y. pseudotuberculosis.15, 16 It is highly adapted to rodents and their fleas and does not long persist outside them. In the environment, it is rapidly killed by temperatures above 40°C and by desiccation.
Y. pestis Virulence Factors
Y. pestis expresses several virulence factors that are important for its survival in rodents, humans, and fleas.4, 17, 18, 19 Most genes encoding these virulence factors reside on three plasmids, Pst (9.5 kb), Lcr (approximately 70 kb), and pFra (approximately 110 kb). The Pst plasmid encodes a temperature-dependent plasminogen activator (Pla) and a bacteriocin (pesticin). The plasminogen activator, a bacterial surface protease, is important for survival of Y. pestis in fleas. This protease facilitates the formation of a bolus of blood and aggregated bacteria in the flea midgut, which blocks the flea's proventriculus and leads to regurgitation of infective material when the “blocked” flea attempts to feed (see Epizootic Plague). Pesticin promotes iron uptake by Y. pestis.
The low calcium response plasmid (Lcr) encodes gene products active in low calcium environments. The products include various outer surface proteins (Yops) and a soluble V antigen, thought to be essential to Y. pestis survival in macrophages. Factors associated with the approximately 70-kb plasmid inhibit the production by macrophages of interferon-gamma (IFN-γ) and tumor necrosis factor–alpha (TNF-α).
The pFra plasmid contains genes for the fraction 1 (F1) envelope glycoprotein antigen and a murine exotoxin. F1 antigen is produced only by organisms growing at 30°C or greater. Y. pestis strains expressing F1 antigen are able to resist phagocytosis in the absence of opsonizing antibodies. The murine toxin (Ymt) is highly toxic to mice and rats, but is not known to be toxic in humans. In the flea midgut, Ymt (recently recognized to be phospholipidase D) plays a critical role in protecting Y. pestis from a cytotoxic digestion product of blood plasma. The emergence in Y. pestis of the single responsible gene (ymt) did, by adapting the bacillus to fleas, radically separate the natural history of the plague bacillus from that of the enteropathogenic Yersinia spp.20
Chromosomally encoded virulence factors include a lipopolysaccharide endotoxin and a factor controlling the absorption of exogenous iron. Endotoxin is responsible for complement activation and release of cytokines. When Y. pestis is grown on media with Congo red, a heme-analogue dye, iron absorption is indicated by pigment production. Strains that do not produce pigment have diminished virulence in mammals and are unable to induce blocking in the flea gut. Hemin storage locus (hms) products expressed in the low temperature (28°C or less) environment of the flea assist in forming blockages of the flea gut necessary for efficient transmission.18, 19, 20 A chromosomally encoded product, the pH 6 antigen (Psa), inhibits phagocytosis of Y. pestis in mammals.
Y. pestis Biotypes
Y. pestis isolates can be classified into biotypes based on their ability to ferment glycerol and reduce nitrate. Three biotypes have been associated with respective plague pandemics.21 Relict populations of the Antiqua biotype are found in Africa, southeastern Russia, and Central Asia; the Mediaevalis biotype is found around the Caspian Sea; and the Orientalis biotype in Asia and the Western Hemisphere. Typing based on ribosomal gene characteristics supports these distinctions. Considering their adaptation to diverse ecologies worldwide, wild Y. pestis strains are quite uniform, although finer genomic analyses have recently demonstrated subtle and evolving differences.22, 23
Y. pestis Life Cycle
Y. pestis is a zoonotic infection that principally involves wild and domestic rodents, and their various flea species (Fig. 42-2 ). Plague spreads from rodents to humans when humans intrude into the natural (wild) cycle, or when commensal rodents become infected and transmission is established in homes and other communal settings. Human-to-human transmission may result from close and unprotected exposure to persons with respiratory plague. Transmission from person to person by the human flea, Pulex irritans, is controversial. Studies of plague transmission in the 20th century suggest that spread by the human flea, if it occurs, is inefficient and uncommon; however, human fleas have been proposed as an important factor in the rapid spread of plague in Europe during the Black Death.
FIGURE 42-2.
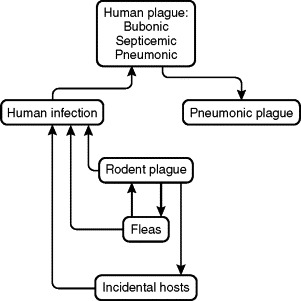
Yersinia pestis cycles, demonstrating the interrelationships of its rodent reservoir, flea vector, and incidental mammalian hosts, including humans. In the usual situation, Y. pestis is transmitted from rodent to rodent, and occasionally from rodent to human or other incidental mammalian host, by an infective rodent flea. Pet dogs and cats can bring infective rodent fleas into the home environment. Infection may be transmitted directly from rodent to rodent by cannibalism, and from infected animal to predator or to human by ingestion. Humans may also acquire infection by accidental percutaneous inoculation while handling contaminated tissues or fluids. Plague pneumonia can be transmitted from one person to another and, rarely, from cats to persons, by infective respiratory droplets.
EPIDEMIOLOGY
Ecology
More than 200 species of mammals and 150 species of fleas have been found to be naturally infected with Y. pestis. However, few of these hosts are important for maintaining enzootic or epizootic life cycles, and fewer still pose a significant risk to humans. The natural history of Y. pestis is complex, and an understanding of the organism's ecology and life cycle is helpful in defining its epidemiology and designing control and prevention strategies (Box 42-1 ).7, 8, 9 24, 25, 26
Box 42-1. Plague Foci.
North American Plague Foci
Wild rodent plague has been reported west of the 100th meridian in 17 contiguous western states and in some areas of adjacent Canada and Mexico.7, 8, 25 The major plague sites are the southwestern focus, comprising mainly semi-arid grassland plateaus, foothills, and forested uplands of northeastern Arizona, most of New Mexico, southern Utah, and southern Colorado; the Pacific Coast focus, comprising mostly valley grassland, foothills, and the montane habitat of California and southern Oregon; the Great Basin focus, encompassing parts of Utah, Nevada, and southern Idaho; and the Rocky Mountain and northern focus, comprising mostly northern Colorado, Wyoming, and Montana.
The principal rodent hosts in the southwestern focus are various burrowing ground squirrels (Spermophilus spp.), prairie dogs (primarily Cynomys gunnisoni), wood rats (Neotoma spp.), antelope ground squirrels (Ammospermophilus spp.), deer mice (Peromyscus maniculatis), and related species. The major rodent hosts in the various niches of the Pacific Coast focus include Spermophilus spp., especially the California ground squirrel (S. beecheyi), the golden-mantled ground squirrel (S. lateralis), chipmunks (various Tamias spp.), deer mice and other Peromyscus spp., and voles (Microtus spp.). Other important hosts in the United States include various small ground squirrels (S. elegans, S. beldingi, and S. townsendi) in the Rocky Mountain and Great Basin regions, and the black-tailed prairie dog (Cynomys ludovicianus) in the Great Plains region. Epizootics of plague have recently occurred in urban tree squirrel (Sciurus niger) populations in cities along the eastern foothills of the Colorado Rocky Mountains, but pose a small risk to humans because these squirrels are parasitized by nonanthropophagic fleas.
Since 1950, a few cities in the United States have rarely been found to have Y. pestis–infected rats (e.g., Tacoma, San Francisco, Los Angeles, Dallas). However, no widespread epizootics or human plague cases have resulted, possibly because the rats have been infested with only small numbers of fleas or with flea species that are inefficient vectors of Y. pestis.
The principal fleas transmitting plague among wild rodent epizootic hosts in the United States include various ground squirrel fleas (Oropsylla montana [Diamanus montanus], Hoplopsyllus anomalus, Thrassis spp., Opisocrostis spp., Oropsylla idahoensis), prairie dog fleas (Opisocrostis spp.), wood rat fleas (Orchopeas spp.), and chipmunk fleas (Eumolpianus eumolpi).7, 8, 25 Or. montana is the most important vector of Y. pestis to persons in the United States because it is a competent host that readily feeds on a wide range of rodents, and on other mammals, including humans.
South American Plague Foci
In South America, active enzootic plague foci exist in Brazil, Bolivia, Peru, and Ecuador, and have been described previously in Paraguay, Argentina, and Venezuela. Y. pestis infection in these foci has been variously found in commensal rats (Rattus spp.), cotton rats (Sigmodon spp.), rice rats (Oryzomys spp.), field mice (Akodon spp.), cane mice (Zygodontomys spp.), wild cavies, and domesticated guinea pigs (Cavia and Galea spp.).3, 26 Domestic guinea pigs are reared in homes for food in the Andean region and are considered a potential commensal risk of infection to humans; human plague outbreaks in the Andean region, including a recent extensive bubonic plague epidemic in northern Peru and a mixed bubonic/pneumonic plague outbreak in Ecuador, have been suspected to be associated in the domestic environment with infected guinea pigs as well as R. rattus. A previously active plague area in northeastern Brazil has been quiescent over the past 20 years. The various fleas that serve as principal vectors in South American foci are X. cheopis on Rattus spp. and Polygenis and Pleochaetis spp. on wild rodents. Pulex irritans, the human flea (which also parasitizes domesticated guinea pigs), has been implicated as a potential transmitter of plague to humans in some Andean outbreaks.
African Plague Foci
Widely scattered active plague foci exist in East and South Africa, including Democratic Republic of the Congo (previously Zaire), Uganda, Kenya, Tanzania, Zambia, Zimbabwe, Mozambique, Botswana, South Africa, Namibia, and Angola, and on the Indian Ocean island of Madagascar.3, 7, 26 Less active foci exist in some northern Africa states (e.g., Libya, Algiers). The principal wild rodent hosts in Africa include gerbils (Tatera and Desmodillus spp.), swamp rats (Otomys spp.), various grass mice (Arvicanthis spp.), multimammate mice (Mastomys spp.), and commensal rats (Rattus spp.). One scenario describes plague in grassland gerbil populations spreading to multimammate mice in agricultural fields, and then to commensal rats in villages, resulting in human plague outbreaks. The principal flea vectors of wild rodent hosts are Xenopsylla and Dinopsyllus spp., while X. cheopis and X. braziliensis are the principal flea species involved in transmission among commensal rats and to man.
Asian Plague Foci
The most important Eurasian plague hosts are gerbils (various Meriones spp.) in Iran, Kurdistan, Transcaucasia, other areas around the Caspian Sea, and the plains of southeastern Russia and Khazakstan; marmots (Marmota spp.) in Central Asia, including mountainous Khazakstan, northeastern China, Mongolia, Manchuria, and in Transbaikalia; and ground squirrels (Spermophilus spp.) in Mongolia, northern-central China, the central Asian plains and steppes, and some areas around the Caspian Sea.3, 7, 26 The primary flea vectors on gerbils are Xenopsylla and Nosopsyllus spp.; on marmots, various Oropsylla, Rhadinopsylla, and Citellophilus spp.; and on ground squirrels, Citellophilus and Neopsylla spp.
In India, the gerbil, Tatera indica, has been described as the principal wild rodent maintenance host.3, 7, 26 Although it lives principally in open grassland sites in its natural state, it does invade agricultural fields and village peripheries. Other maintenance hosts include Mellardia meltada, various field mice, and palm squirrels (Funambulus spp.). The important commensal rat species are Bandicota bengalensis, Bandicota indica, R. rattus, and R. norvegicus. The primary vectors of plague in India are X. cheopis and Xenopsylla astia.
The principal rodent hosts in Myanmar, Vietnam, and Indonesia are R. rattus subsp. diardii and the Polynesian rat (Rattus exulans).3, 7, 26 R. rattus subsp. flavipectus is an important host in southern China. R. norvegicus and the bandicoot, B. indicus, have been described as important hosts in Vietnam. The shrew, Suncus murinus, has been described as a possibly important host of plague in Vietnam and Indonesia. X. cheopis is the principal vector of plague in southern China, Myanmar, Vietnam, and Indonesia; X. astia (a less efficient vector) is also found on rats in Myanmar and Vietnam. Stivalius cognatus, a rat flea, is considered to be an important secondary vector of plague in Indonesia.
Enzootic Plague
In its natural reservoir hosts, infection with the plague bacillus is often unnoticed. This is characteristic of enzootic infections among rodents in remote, sparsely populated areas, such as the semiarid or arid grassland steppes of Central Asia and the savannahs of eastern Africa.7, 8, 9 25, 26 Fleas become infected by feeding on bacteremic rodents; although infection sickens or kills some enzootic hosts, most tolerate the infection and noticeable population die-offs do not usually occur. Although nonrodent mammals, such as predatory carnivores, do not serve as reservoirs of infection, they may contribute to the maintenance of zoonotic cycles by transporting infective fleas from one rodent population to another. Enzootic transmission generally places humans at a small risk of incidental infection, and associated cases are typically infrequent and sporadic.
Epizootic Plague
In epizootic plague, infection spreads rapidly among susceptible rodent populations transmitted by efficient flea vectors (both rodents and fleas serve as amplifying hosts of Y. pestis).7, 8, 24, 25, 26 Epizootics often result in die-offs of affected rodent populations and accelerated dispersal of their fleas. Spread of infective fleas to rodents living in close association with humans can pose a serious threat in residential areas. Prairie dogs are highly susceptible and sensitive epizootic hosts in the southwestern United States, and epizootics can quickly kill off colonies over wide areas, sometimes in close proximity to suburban developments.
Historically, urban epidemics and pandemic spread of plague have been linked to epizootics involving the commensal rodents Rattus rattus and R. norvegicus. The smaller, domestic, roof or black rat (R. rattus) is considered the most dangerous plague host. Since ancient times, this adaptable and clever rodent has lived in intimate domestic association with humans in Asia, the Middle East, and Africa. Its migration to Europe allowed the rapid spread of plague there during the medieval period. Black rats live in homes, adjacent grain fields, granaries, stables and barns, warehouses, and other places where human activity provides them with ready shelter and food supplies. They travel easily with human cargo and baggage and have (with their complement of fleas) been the principal agents of noncontiguous (per saltum) spread of plague. In contrast, the Norway rat (R. norvegicus) was probably not an important host in the first two great pandemics and is thought to have first migrated from Asia to the Middle East and Europe as late as the 18th century, when it displaced the roof rat as the principal plague host in urban areas of Europe and elsewhere. R. norvegicus is larger and more aggressive than R. rattus; it is a burrowing rodent that lives around the foundations of buildings and frequents cellars, drains, sewers, alleyways, brush heaps, and refuse dumps. The quartet of R. rattus, R. norvegicus, Y. pestis, and the oriental rat flea, Xenopsylla cheopis, was responsible for the global spread of plague during the third (modern) pandemic. Densities of rats (measured by trapping success rates) and of fleas on rats (rat flea index) and Y. pestis infection rates of both are important surveillance measurements.7, 9
Although domestic rats are the principal source of infected fleas in plague epidemics, they may not be able to sustain transmission of Y. pestis without reintroductions of the agent from the wild cycle. Recent investigations of bubonic plague in Maharashtra in western-central India suggested that local gerbils (Tatera indica) living in open, arid grasslands were the principal reservoirs of infection there, and that Y. pestis may have in part reached domestic R. rattus through intermediary populations of bandicoot rats (Bandicota bengalensis), which were numerous in adjacent croplands surrounding rural villages.27, 28
The widely distributed oriental rat flea, X. cheopis, and the related species, Xenopsylla braziliensis, are efficient vectors of the plague bacillus between rats and are the most dangerous vectors to humans.3, 7, 9 At ambient temperatures of 28°C and below, Y. pestis can multiply exponentially in the gut of these and a few other species of fleas and result in a clotted bolus of organisms that blocks the passage of further blood meals at the level of the foregut. The starving fleas avidly seek replenishing blood meals. Regurgitation by a “blocked” flea while it feeds enhances transmission of the plague bacillus to a new host. The ability of Y. pestis to cause blockage has recently been shown to be plasmid-mediated and dependent on a single gene change from Y. pseudotuberculosis (see Y. pestis Virulence Factors).19, 20
Incidental Animal Plague
A diverse range of wild and domestic mammals may become incidentally infected with Y. pestis, and humans can become infected when they have direct contact with their infected tissues or exudates, or when these incidental hosts transport rodent fleas into the domestic environment. Carnivores, especially felids, canids, and mustellids, are notable incidental hosts. Infected cats, in contrast to dogs, often become ill when infected with Y. pestis and have become an occasional source of human plague in the United States.29, 30 Lagomorphs (rabbits, hares) occasionally infect hunters and others who handle contaminated carcasses.31 Ungulates, including antelope, deer, camels, and goats, can sicken and die from Y. pestis infection. Handling of carcasses of infected goats, camels, and dromedaries, and ingestion of undercooked meat of these animals has been responsible for limited human plague outbreaks in northern Africa, the Middle East, and Central Asia.32
Distribution of Plague
Plague foci are distributed throughout the world. From 1989 to 2003, 38,359 human cases (approximately 2500 cases a year) and 2845 deaths (7%) were reported to the WHO by 25 countries, as required by the international health regulations.33 Nearly 80% of the cases were reported from the African region, 15% from Asia, and the remainder from the Americas. The countries that reported more than 1000 cases (from most to least) were Madagascar, Tanzania, Democratic Republic of Congo, Vietnam, Mozambique, Namibia, and Peru. The greatest recent emergence has occurred in eastern sub-Saharan Africa, especially in Tanzania and adjacent countries, and on the island of Madagascar. In Madagascar, recent outbreaks have occurred in coastal urban as well as rural highland areas.10, 34
In the United States, in the period 1989 to 2003, there were 101 reported cases of plague (∼7 cases per year) and 10 deaths (10%). In the United States, plague in animals occurs in 17 of the contiguous western states, in a region extending from the Pacific Coast to the Great Plains and central Texas. However, more than 80% of human cases occur in the southwestern states of New Mexico, Arizona, and Colorado, and approximately 10% in California.29, 35 In the United States, flea-borne plague is highly seasonal, occurring most often between May and October; winter plague cases are uncommon and most often arise from the handling of infected animals. In the tropics, plague incidence usually peaks in the cooler, moist months of the year.
Risk Factors for Human Plague
The risk of plague varies with environmental circumstances and human behaviors, especially those that increase exposure to infective fleas. Low socioeconomic status is an important risk factor, because infestation by rats and their fleas is associated with poor housing and unsanitary conditions.
More than half of plague exposures in the United States are thought to occur in the general area of patients' homes.31 This is particularly true in the Southwest, where homes in rural and semi-rural areas are often situated near habitats of highly plague-susceptible animals, such as prairie dogs, rock squirrels, and woodrats.7, 8 In the Sierra Nevada mountains of California and Nevada, epizootic plague in chipmunks and ground squirrels is a risk to visitors to public parks. Hikers, campers, and hunters throughout the western United States have a small but definite risk of exposure, especially in the warmer months of the year.
Plague can also be transmitted to humans by direct inoculation while skinning or otherwise handling infected rodents, rabbits and hares, domestic and wild felids, and coyotes.7, 29, 31 Direct inoculation is associated with an increased risk of septicemia and a high fatality rate. In Mongolia and northeastern China, marmot hunting is the principal risk factor; further, when pneumonic cases arise, person-to-person transmission is facilitated in the cramped living quarters (yurts) of these nomadic hunters. Pharyngeal plague may result from the ingestion of undercooked contaminated meat32 and possibly by inhalation of infective respiratory droplets.
In the United States, relatively detailed information is available on human plague cases. Among 413 evaluable patients reported from 1960 to 2004, 333 (81%) presented with primary lymphadenitic (bubonic) plague, most thought to have arisen from infective flea bites; 64 (15%) of patients presented with primary septicemic plague, often after handling infected animal tissues; and 10 (2%) patients presented with primary pneumonic plague, 5 of whom had evidence of infective exposures from sick cats (CDC, unpublished data, 2005).
Following the extraordinary epidemics of pneumonic plague involving tens of thousands of cases in Manchuria in the period 1910–1926, pneumonic plague outbreaks around the world have occurred only sporadically, usually involving three or fewer generations of cases, and typically affecting family members, close friends, and care providers of pneumonic plague patients. The 1994 outbreak of reported pneumonic plague in Surat, India, probably involved fewer than 100 cases.13 Small clusters of cases of pneumonic plague have more recently been reported from Ecuador, Madagascar, and northern India. Because respiratory spread of Y. pestis occurs by infective droplets, only persons with close respiratory exposure have a high risk of infection. Primary plague pneumonia is thought to possibly be more contagious than secondary pneumonia because it is more likely to produce copious watery sputum, and the patient may be mobile and expose a wider circle of individuals in the early stage of contagiousness.
DISEASE
Bubonic Plague
Bubonic plague is the prototypic plague illness.3, 36, 37 The usual incubation period of bubonic plague is 2 to 6 days, occasionally longer. Illness typically begins with headache, fever of 38°C to 40°C, chills, myalgias, arthralgias, and a feeling of weakness. At the same time, or within 24 hours, the patient notices tenderness and pain in one or more regional lymph nodes proximal to the site of inoculation of the plague bacillus. The femoral and inguinal groups of nodes are most commonly involved, followed by the axillary and cervical nodes. Upper body sites may be relatively more involved in children than adults. The enlarging bubo(es) becomes progressively swollen, painful, and tender, sometimes exquisitely so. Typically, the patient guards against palpation and limits movement, pressure, and stretching around the bubo. The surrounding tissue often becomes edematous, sometimes markedly, and the overlying skin may be erythematous, warm, tense, and may desquamate (Fig. 42-3 ). Inspection of the skin surrounding or distal to the bubo sometimes reveals the site of a flea bite marked by a small papule, pustule, scab, or ulcer. Larger pustular lesions (furuncles or carbuncles), ulcers, and eschars may rarely occur and can be confused with those caused by tularemia or anthrax. The bubo of plague is distinguishable from lymphadenitis of most other causes by its rapid onset, extreme tenderness, accompanying signs of toxemia, and usual absence of cellulitis or obvious ascending lymphangitis. Treated in the uncomplicated state with an appropriate antibiotic, bubonic plague usually responds quickly, with disappearance of fever and resolution of other systemic manifestations over 2 to 5 days. Buboes often remain enlarged and tender for a week or more after treatment has begun and can occasionally become fluctuant. Without effective antimicrobial treatment, bubonic plague may progress to an increasingly toxic state of fever, tachycardia, lethargy leading to prostration, agitation and confusion, and, occasionally, convulsions and delirium. Mild forms of bubonic plague, called pestis minor, have been described in South America and elsewhere; in these cases, the patients are ambulatory, only mildly febrile, and have subacute buboes.
FIGURE 42-3.
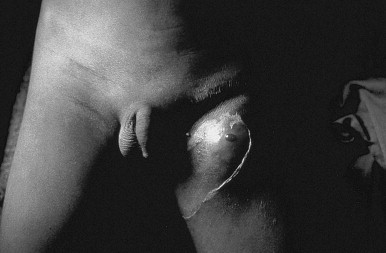
Patient with left inguinal and femoral buboes, demonstrating surrounding edema and overlying desquamation.
Plague patients typically have white blood cell counts of 12,000 to 25,000/μL with a predominance of immature polymorphonuclear leukocytes. Leukemoid reactions showing white blood cell counts as high as 50,000/μL3 or more can occur.36 Differential diagnostic possibilities for bubonic plague include streptococcal or staphylococcal adenitis, tularemia, cat scratch disease, mycobacterial infection, acute filarial lymphadenitis, chancroid and other sexually transmitted diseases that cause regional lymphadenitis, and strangulated inguinal hernia.
Septicemic Plague
Septicemic plague is characterized by a rapidly progressive, overwhelming endotoxemia and dissemination of infection.38, 39, 40 Primary septicemia occurs in the absence of an apparent regional lymphadenitis, and the diagnosis of plague is often not suspected until preliminary blood culture results are reported by the laboratory. Further, patients with sepsis often present with gastrointestinal symptoms such as nausea, vomiting, diarrhea, and abdominal pain, making a correct clinical diagnosis even more challenging.41 If not treated early with appropriate antibiotics and aggressive supportive care, septicemic plague is usually fulminating and fatal. In the United States from 1960 to 2004, 19 of 64 primary septicemic plague cases were fatal, yielding a case-fatality ratio of 30%. The systemic inflammatory response syndrome (SIRS) may progress rapidly. Petechiae and ecchymoses are manifestations of disseminated intravascular coagulation (DIC). In later stages, thrombi in the microvasculature of acral parts, such as tips of ears and digits, may result in gangrene of affected tissues. Refractory hypotension, acute renal failure, obtundation, and other signs of shock are preterminal events. Acute respiratory distress syndrome (ARDS) can occur in septicemic plague and be refractory to treatment.
Differential diagnostic possibilities include any other overwhelming systemic infection, including gram-negative sepsis with agents other than the plague bacterium, meningococcemia, and bacterial endocarditis. ARDS could be confused with other underlying conditions such as hantavirus pulmonary syndrome and severe acute respiratory syndrome (SARS).
Pneumonic Plague
Pneumonic plague is a highly virulent form of plague.3, 42 The incubation period for primary pneumonic plague is 1 to 7 days, but most cases arise 3 to 5 days after exposure. The onset is most often sudden, with chills, fever, headache, body pains, weakness, dizziness, and chest discomfort. Cough, sputum production, increasing chest pain, tachypnea, and dyspnea typically predominate on the second day of illness and may be accompanied by hemoptysis, increasing respiratory distress, cardiopulmonary insufficiency, cyanosis, and circulatory collapse. In primary plague pneumonia, principally an alveolar process, the sputum is most often watery or mucoid, frothy, and blood-tinged, but may become frankly bloody. Chest signs in primary plague pneumonia may indicate localized pulmonary involvement in the early stage, with rapidly developing segmental consolidation before bronchopneumonia spreads to other segments and lobes of the same and opposite lung (Fig. 42-4 ). Liquefaction necrosis and cavitation may develop at sites of consolidation and may leave significant residual scarring.
FIGURE 42-4.
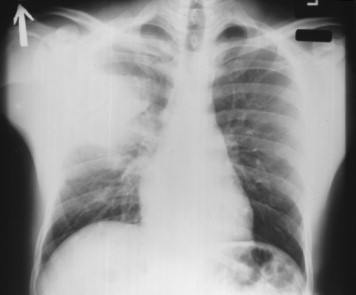
Chest radiograph of a patient with primary plague pneumonia, showing extensive infiltrates in the right upper and right middle lobes.
Secondary plague pneumonia manifests first as an interstitial pneumonitis in which sputum production is scant, and the sputum is more likely to be inspissated and tenacious in character than the sputum found in primary pneumonic plague. In the United States from 1960 to 2004, 48 cases of secondary plague pneumonia and 10 cases of primary plague pneumonia were reported to the Centers for Disease Control and Prevention (CDC), with no known secondary transmission to contacts and an overall case-fatality rate of 40% (CDC, unpublished data, 2005). Observers of pneumonic plague in the early 20th century remarked on minimal auscultatory findings, the appearance of toxemia, and the frequency of sudden death, as compared to patients with other bacterial pneumonias.42
Differential diagnostic possibilities include other bacterial pneumonias such as mycoplasma pneumonia, Legionnaire's disease, staphylococcal or streptococcal pneumonia, tularemia pneumonia, and Q fever. Severe viral pneumonia, including hantavirus pulmonary syndrome and SARS, could be confused with plague.
Meningeal Plague
Meningitis is an unusual manifestation of plague. In the United States, there were 18 (4%) meningitis cases among the total 413 evaluable plague cases reported to CDC from 1960 to 2004. Most cases were late complications of bubonic plague, and 14 patients (78%) survived. Although meningitis may be a part of the initial presentation of plague, its onset is often delayed and may be the result of insufficient antibiotic treatment of the primary illness.43 Chronic, relapsing meningeal plague over periods of weeks and even months was described in the preantibiotic era. Plague meningitis presents as a typical bacterial meningitis, with fever, headache, altered mental status, meningismus, and a polymorphonuclear leukocytic pleocytosis.
Pharyngeal Plague
Plague pharyngitis is an unusual condition and presents with fever, sore throat, and cervical lymphadenitis. In its early stages, it may be clinically indistinguishable from more common causes of pharyngitis.3 Cervical or submandibular buboes usually develop secondary to the pharyngeal involvement. Cases arise following respiratory droplet exposure, or from the ingestion of undercooked meat.3, 32 Pharyngeal plague may give rise to secondary plague pneumonia. Care providers working in plague-endemic areas should be alert to the possibility of plague in the differential diagnosis of acute bacterial pharyngitis. Pharyngeal colonization with Y. pestis sometimes occurs without symptoms among contacts of persons with pneumonic plague. Epidemiological observations do not suggest that persons with pharyngeal carriage present a contagious threat to others.
PATHOGENESIS AND IMMUNITY
Molecular Basis of Disease
Y. pestis is one of the most virulent bacteria known; both chromosomal- and plasmid-encoded gene products are involved (see Y. pestis Virulence). A lipopolysaccharide endotoxin is thought to be primarily responsible for the pathogenic effects of plague sepsis, including SIRS, ARDS, excessive cytokine activation, complement cascade with resultant DIC and bleeding, unresponsive shock, and organ failure.37, 38, 39, 40 44 Before antibiotics became available, nearly all persons with septicemic or pneumonic plague died rapidly from toxemia and organ failure.
Tissue Invasion and Pathology
Y. pestis organisms inoculated through the skin or mucous membranes are carried via lymphatics to regional lymph nodes, although it is possible that direct bloodstream dissemination may occur. In the early stages of infection, affected lymph nodes are edematous, congested, and have minimal inflammatory infiltrates and vascular injury. Fully developed buboes, however, contain large numbers of infectious plague organisms and show hemorrhagic necrosis and a mild neutrophilic infiltration.44 The affected nodes are usually surrounded by a serosanguineous effusion. When several adjacent lymph nodes are involved, a boggy, edematous mass can result. Spontaneous bubo rupture and suppurative drainage may occur.
Primary septicemia results when multiplication of Y. pestis occurs in the bloodstream in the absence of a bubo; secondary septicemic plague develops when primary lymphadenitic or pulmonary disease occurs, local defenses are breached, and infections spill over into the bloodstream. Newly inoculated plague bacilli are able to survive phagocytosis and be disseminated by mononuclear cells to distant sites. Y. pestis can invade and cause disease in almost any organ, and untreated infection usually results in widespread and massive tissue destruction. A diffuse interstitial myocarditis with cardiac dilation, multifocal necrosis of the liver, diffuse hemorrhagic splenic necrosis, and fibrin thrombi in renal glomeruli are common findings at autopsy.44, 45 If DIC occurs, the result is thrombosis and vascular necrosis with widespread cutaneous, mucosal, and serosal petechiae and ecchymoses.37, 38, 45
Pneumonic plague can result from inhalation of infective respiratory droplets from a person or animal with respiratory plague or secondary to hematogenous spread in a patient with bubonic or septicemic plague. It can result also from inhalation of Y. pestis in a laboratory accident.46 Primary plague pneumonia generally begins as a lobular process and then extends by confluence, becoming lobar and then multilobar. Typically, plague organisms are most numerous in the alveoli. Secondary plague pneumonia begins more diffusely as an interstitial process, with organisms initially most numerous in the interstitial spaces. In untreated cases of both primary and secondary plague pneumonia, the usual findings are diffuse pulmonary hemorrhage, necrosis, and scant neutrophilic leukocyte infiltration.42
Immunity
Yersinia pestis resists the innate host defense mechanisms such as phagocytosis and the induction of inflammatory responses by macrophages and neutrophils. On contact with eukaryotic cells, yersiniae release LcrV into the medium and deliver six effector Yops directly into the eukaryotic cell cytoplasm through a type III secretion system.47
Yersiniae have multiple mechanisms for suppression of the mammalian innate immune response. LcrV signals in a CD14- and toll-like receptor 2 (TLR2)–dependent fashion leading to immunosuppression by interleukin (IL)-10 induction.48, 49 LcrV together with YopB inhibit expression of phospho-p38, -p42/44, -JNK mitogen-activated protein kinases (MAPKs) and transcription factors nuclear factor (NF)-κB, c-fos, and c-jun in lipopolysaccharide (LPS)-treated macrophages.50 The inhibition in the expression of these signaling molecules correlates with the inhibition of TNF-α and nitric oxide production in macrophages.50 YopJ causes downregulation of NF-κB and mitogen-activated protein kinase pathways and also causes downregulated production of proinflammatory cytokines, including TNF-α. The plague virulence protein YopM targets the innate immune response by causing a global depletion of natural killer (NK) cells, possibly by blocking expression of functional IL-15R on NK cells and inhibiting IL-15 expression by macrophages. IL-15 is crucial for the maintenance and activation of circulating NK cells as well as for their development in the bone marrow.51 YopM, after delivery to host cells through a type III secretion mechanism, is transported to the nucleus.51 Production of IFN-γ by NK cells is necessary for early activation of macrophages to mediate robust bactericidal activity, production of pro-inflammatory cytokines, and strong expression of Th1 cytokines. The depletion of NK cells during Y. pestis infection results in an early decrease in production of Th1 cytokines such as IL-12 and IL-18, and TNF-α. The loss of IFN-γ expression in the Y. pestis–infected host may contribute to the absence of granulomas. In addition, decreased IFN-γ–mediated macrophage activation in Y. pestis infection favors greater bacterial growth.
DIAGNOSIS
Except in epidemic situations, a high index of clinical suspicion and a careful clinical and epidemiologic history and physical examination are required to make a timely diagnosis of plague. A delayed or missed diagnosis of plague is associated with a high case-fatality rate, and infected travelers who seek medical care after they have left endemic areas (peripatetic plague cases) are especially at risk.52, 53 When plague is suspected, close communication between clinicians and the diagnostic laboratory, and between the diagnostic laboratory and a qualified reference laboratory is essential. Laboratory tests for plague are highly reliable when conducted by persons experienced in working with Y. pestis, but such expertise is usually limited to specialized reference laboratories. Because of recent concerns with possible bioterrorism, a network of participating laboratories across the United States has been developed with the ability to make rapid and confirmatory diagnoses. All state public health laboratories now have this capability and can, if necessary, forward materials to the CDC for rapid advanced procedures.54
Specimen Collection and Processing
When plague is suspected, specimens should be obtained promptly for microbiologic studies, chest radiographs taken, and specific antimicrobial therapy initiated pending confirmation. Appropriate diagnostic specimens for smears and culture include blood in all patients, lymph node aspirates in those with suspected buboes, sputum samples or tracheobronchial aspirates in those with suspected pneumonic plague, and cerebrospinal fluid in those with meningeal signs. A portion of each specimen should be inoculated onto suitable media (e.g., brain-heart infusion broth, sheep blood agar, chocolate agar, or MacConkey agar). Smears of each specimen should be stained with Wayson or Giemsa stain, and with Gram stain, and examined using light microscopy. If possible, the specimens should also be examined using direct fluorescent antibody (FA) testing.14 An acute phase serum specimen should be collected for Y. pestis antibody testing, followed by a convalescent phase specimen collected 3 to 4 weeks later. For diagnosis in fatal cases, autopsy tissues should be collected for culture, FA testing, and histological processing, including buboes, samples of solid organs (especially liver, spleen, and lung), and bone marrow. For culture, specimens should be sent to the laboratory either fresh or frozen on dry ice and not in preservatives or fixatives. Cary-Blair or a similar holding medium can be used to transport Y. pestis–infected tissues.
Laboratory Confirmation
Laboratory confirmation of plague depends on the isolation of Y. pestis from body fluids or tissues. When the patient's condition allows, several blood cultures taken over a 45-minute period prior to treatment will often result in successful isolation of the bacterium. Y. pestis strains are readily distinguished from other gram-negative bacteria by polychromatic and immunofluorescence staining properties, characteristics of growth on microbiologic media, biochemical profiles, and confirmatory lysis by the Y. pestis–specific bacteriophage.14 Laboratory mice and hamsters are susceptible to Y. pestis and are used in specialized laboratories to make isolations from contaminated materials and for virulence testing.
In the absence of cultural isolation of Y. pestis, plague cases can be confirmed by demonstrating a fourfold or greater change in serum antibodies to Y. pestis F1 antigen by passive hemagglutination (PHA) testing or by detecting a serum antibody titer of 128 or greater in a single serum sample from a patient with a compatible illness who has not received plague vaccine. The specificity of a positive PHA test is confirmed by F1 antigen hemagglutination inhibition (HI) testing. A small percentage of plague patients will develop diagnostic levels of antibodies within 5 days after the onset of illness, most seroconvert 1 to 2 weeks after onset, fewer seroconvert more than 3 weeks after onset, and less than 5% fail to seroconvert.55 Early specific antibiotic treatment may delay seroconversion by several weeks. Following conversion, serologic titers diminish gradually over months to years. Enzyme-linked immunosorbent assays (ELISAs) for detecting immunoglobulin M (IgM) and IgG antibodies to Y. pestis are useful in identifying antibodies in early infection and in differentiating them from antibodies developed in response to previous vaccination. Recently, antigen capture ELISA procedures, polymerase chain reaction assays, and handheld immunodiagnostic antigen detection tests for rapid, early diagnosis have been developed and are being evaluated. The handheld devices allow diagnostic testing of clinical materials at the bedside, even under primitive conditions.56
TREATMENT AND PROGNOSIS
Untreated, plague is fatal in over 50% of bubonic cases and in nearly all cases of septicemic or pneumonic plague. The overall plague case-fatality ratio in the United States in the past 50 years has been approximately 15%.57 Fatalities are almost always due to delays in seeking treatment, misdiagnosis, and delayed or incorrect treatment. Rapid diagnosis and appropriate antimicrobial therapy are essential. The case-fatality ratio is very high for pneumonic plague patients who begin treatment more than 18 to 24 hours after the onset of pulmonary symptoms.
Streptomycin has long been the treatment of choice for plague; this aminoglycoside is, however, no longer manufactured in the United States, is not widely available, and can cause irreversible ototoxicity in some patients. Because of its ready availability and ease of administration, gentamicin is replacing streptomycin as treatment for plague patients in the United States. Although comparative trials of safety and efficacy of streptomycin versus gentamicin in plague have not been conducted, case reports and a retrospective review of patients treated in New Mexico indicate that gentamicin is an acceptable substitute in the treatment of plague.58 Tetracycline and chloramphenicol are effective alternatives to the aminoglycosides. Doxycycline has become the tetracycline of choice because of its ease of administration, rapid and efficient absorption after ingestion, and its superior ability to achieve and maintain peak serum concentrations following oral administration. Chloramphenicol is indicated for conditions in which high tissue penetration is important, such as plague meningitis, pleuritis, endophthalmitis, and myocarditis. Chloramphenicol can be used separately or in combination with an aminoglycoside. Treatment with doxycycline, tetracycline, or chloramphenicol should begin with loading doses.
Ciprofloxacin, a fluoroquinolone, has shown promise in vitro and in laboratory animal studies,59, 60 but case series demonstrating its utility in human plague have not been reported. Penicillins, cephalosporins, and macrolides have poor efficacy and should not be used. Trimethoprim-sulfamethoxazole has been used successfully to treat bubonic plague, but is not considered a first-line choice and is not recommended for treating severe forms of the disease. In general, antimicrobial treatment should be continued for 7 to 10 days or for at least 3 days after the patient has become afebrile and has made a clinical recovery. Patients begun on intravenous antibiotics may be switched to oral regimens as indicated by clinical response. Improvement is usually evident 2 to 3 days from the start of treatment, even though lessening fever may continue for several more days.
Guidelines for medical and public health management of a potential Y. pestis terrorist attack have been developed that recommend streptomycin or gentamicin as the first-line treatment for pneumonic plague in a contained casualty situation, substituted if necessary by oral doxycycline or ciprofloxacin in the event of mass casualties.2
Consequences of delayed treatment of plague include DIC, ARDS, and other complications of bacterial sepsis and endotoxemia. Patients with these disorders require intensive monitoring and close physiologic support. Buboes may require surgical drainage if they threaten to rupture. Abscessed nodes rarely can be a cause of recurrent fever in patients who have otherwise had satisfactory recovery; the cause may be occult when intrathoracic or intra-abdominal nodes are involved. Viable Y. pestis organisms have occasionally been isolated from buboes 1 to 2 weeks into convalescence. Y. pestis strains resistant to one or more antibiotics used to treat plague have rarely been isolated from humans but have not been associated with treatment failures. Recently, however, an isolate of Y. pestis from a bubonic plague patient in Madagascar was found to have plasmid-mediated, transferable, high level in vitro antimicrobial resistance to all agents recommended as first-line agents in treating plague.61 The patient recovered after treatment with streptomycin and trimethoprimsulfamethoxazole, and no similar strains have been further identified from humans or from natural sources.
PREVENTION AND CONTROL
General Principles
Surveillance, environmental management, and personal protective measures are the cornerstones of prevention and control.9, 62, 63 Surveillance includes environmental monitoring to determine sites of plague activity, inspection of rodent habitat for signs of epizootics, collecting and testing of fleas from abandoned burrows, trapping and testing of live rodents and their fleas for Y. pestis infection, and testing of animals found sick or dead from suspected plague. In some circumstances, carnivore seropositivity is used as an indicator of rodent plague activity in an area. Dogs and wild canines readily seroconvert following exposure but retain elevated antibody levels for less than a year, making them useful sentinels of recent plague activity in an area.
Personal protective measures include avoidance of areas with known epizootic plague (these may be posted by government authorities); avoidance of sick or dead animals; use of repellents, insecticides, and protective clothing when there is a potential for exposure to rodent fleas; and use of gloves when handling animal carcasses.
Postexposure treatment for 7 days with doxycycline or other tetracycline, chloramphenicol, or ciprofloxacin is recommended for persons who have had a known close exposure to a pneumonic plague patient in the prior 7 days. Oral doxycycline or ciprofloxacin has been recommended for postexposure prophylaxis in the event of a terrorist attack with Y. pestis.2 Pre-exposure prophylaxis may occasionally be recommended for persons who are unable to avoid visiting or residing in an area where a plague outbreak is in progress or who are screening or caring for plague patients in unusual circumstances, such as an outbreak. To reduce the risk of airborne droplet spread, plague patients should have a chest radiograph to rule out pulmonary involvement. Patients with suspected pneumonic plague should be managed in isolation under respiratory droplet precautions until the patient has responded clinically and sputum cultures are negative (sputum typically is sterile within 24 to 48 hours of beginning treatment). Persons caring for sick animals (especially cats) in plague-endemic areas should take precautions to avoid contamination with infectious exudates or expelled respiratory secretions.30
Sources of rodent food (garbage, feed for livestock and pets) and shelter (brushpiles, junk heaps, woodpiles) should be eliminated in domestic, peridomestic, and working environments, and buildings and food stores should be rodent-proofed. Controlling fleas with insecticides is a principal public health measure in situations where epizootic plague activity places humans at high risk. This includes insecticidal dusting and spraying of rodent burrows, rodent runs, and other sites where rodents and their fleas are found. In known plague foci, pet owners should keep their dogs and cats free of fleas and restrained. The decision to control plague by killing rodents should be left to public health authorities and should only be carried out in conjunction with effective flea control. Killing rodents has no lasting benefit without environmental sanitation.
In the event of a plague epidemic, measures should rapidly be taken to control spread, as described in international regulations and guidelines for plague control.9, 11, 62, 63 These measures include delineation of infected areas, rapid detection and treatment of cases and exposed contacts, isolation and monitoring of suspected human plague cases and case contacts, and control of fleas and rodents in plague-infected areas, in port facilities, and on ships and other conveyances as indicated. A rapid surveillance system to identify and contain the possible introduction of pneumonic plague into the United States was established at the time of the reported outbreaks of plague in India in 1994.64
Plague Vaccine
No plague vaccine is currently available in the United States. Much research is under way to develop safe, rapidly acting, and efficacious vaccines using advanced molecular approaches.65 The most promising of these is a vaccine that combines recombinant F1 and V antigens of Y. pestis, and early testing in humans is under way. Another approach in early stages of research is passive immunization with monoclonal antibodies against targeted Y. pestis antigens.
REFERENCES
- 1.Centers for Disease Control and Prevention Biological and chemical terrorism: Strategic plan for preparedness and response. Recommendations of the CDC Strategic Planning Workgroup. MMWR. 2000;49:1. [PubMed] [Google Scholar]
- 2.Inglesby TV, Dennis DT, Henderson DA. Plague as a biological weapon: Medical and public health management. JAMA. 2000;283:2281. doi: 10.1001/jama.283.17.2281. [DOI] [PubMed] [Google Scholar]
- 3.Pollitzer R. WHO Monograph Series 22. World Health Organization; Geneva: 1954. Plague; p. 1. [PubMed] [Google Scholar]
- 4.Perry RD, Fetherston JD. Yersinia pestis—Etiologic agent of plague. Clin Microbiol Rev. 1997;10:35. doi: 10.1128/cmr.10.1.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Link VB. A History of Plague in the United States of America. Public Health Monograph No. 26. Government Printing Office; Washington, DC: 1955. [PubMed] [Google Scholar]
- 6.Butler T. Plague and Other Yersinia Infections. Plenum Press; New York: 1983. [Google Scholar]
- 7.Gage KL. Plague. In: Collier L, Balows A, Sussman M, editors. 9th ed. Vol. 3. Arnold Publications; London: 1998. p. 885. (Topley and Wilson's Microbiology and Microbial Infections). [Google Scholar]
- 8.Poland JD, Barnes AM. Plague. In: Steele JH, editor. Vol. 2. CRC Press; Boca Raton, FL: 1979. p. 515. (CRC Handbook Series in Zoonoses, Section A: Bacterial, Rickettsial, and Mycotic Diseases). [Google Scholar]
- 9.Gratz NG. Plague Manual: Epidemiology, Distribution, Surveillance and Control. World Health Organization; Geneva: 1999. Control of plague transmission; p. 97. [Google Scholar]
- 10.Boisier P, Rahalison L, Rasolomaharo M. Epidemiological features of four successive annual outbreaks of bubonic plague in Mahajanga, Madagascar. Emerg Infect Dis. 2002;8:311. doi: 10.3201/eid0803.010250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.International Health Regulations (1969) World Health Organization; Geneva: 1983. [Google Scholar]
- 12.Dennis DT. Plague in India. BMJ. 1994;309:893. doi: 10.1136/bmj.309.6959.893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Campbell GL, Hughes JM. Plague in India: A new warning from an old nemesis. Ann Intern Med. 1995;122:151. doi: 10.7326/0003-4819-122-2-199501150-00014. [DOI] [PubMed] [Google Scholar]
- 14.Chu MC: Laboratory Manual of Plague Diagnostic Tests. Atlanta, GA, USPHS Centers for Disease Control and Prevention, 2000. Centers for Disease Control and Prevention: Bioweapons Laboratory Issues. 2001. Full text available on the Internet: www.bt.cdc.gov/Agent/Plague/ype_1a_cp/123010.pdf.
- 15.Achtman M, Zurth K, Morelli G. Yersinia pestis, the cause of plague, is a recently emerged clone of Yersinia pseudotuberculosis. Proc Natl Acad Sci USA. 1999;24:14043. doi: 10.1073/pnas.96.24.14043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Parkhill J, Wren BW, Thompson NR. Genome sequence of Yersinia pestis, the causative agent of plague. Nature. 2001;413:523. doi: 10.1038/35097083. [DOI] [PubMed] [Google Scholar]
- 17.Brubaker RR. Factors promoting acute and chronic diseases caused by yersiniae. Clin Microbiol Rev. 1991;4:309. doi: 10.1128/cmr.4.3.309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Carniel E. Evolution of pathogenic Yersinia, some lights in the dark. Adv Exp Med Biol. 2003;529:3. doi: 10.1007/0-306-48416-1_1. [DOI] [PubMed] [Google Scholar]
- 19.Hinnebusch BJ. Bubonic plague: A molecular genetic case history of the emergence of an infectious disease. J Mol Med. 1997;75:645. doi: 10.1007/s001090050148. [DOI] [PubMed] [Google Scholar]
- 20.Hinnebusch BJ, Rudolph AE, Cherepenov P. Role of murine toxin in survival of Yersinia pestis in the midgut of the vector flea. Science. 2002;296:733. doi: 10.1126/science.1069972. [DOI] [PubMed] [Google Scholar]
- 21.Guiyoule A, Grimont F, Iteman I. Plague pandemics investigated by ribotyping of Yersinia pestis strains. J Clin Microbiol. 1994;32:634. doi: 10.1128/jcm.32.3.634-641.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Guiyoule A, Rasoamanana B, Buchrieser C. Recent emergence of new variants of Yersinia pestis is Madagascar. J Clin Microbiol. 1997;35:2826. doi: 10.1128/jcm.35.11.2826-2833.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Radnedge L, Agron PG, Worsham PL. Genome plasticity in Yersinia pestis. Microbiology. 2002;148:1687. doi: 10.1099/00221287-148-6-1687. [DOI] [PubMed] [Google Scholar]
- 24.Pollitzer R, Meyer KF. The ecology of plague. In: May JH, editor. Studies in Disease Ecology. Hefner; New York: 1961. p. 433. [Google Scholar]
- 25.Barnes AM. Surveillance and control of bubonic plague in the United States. Symp Zool Soc London. 1982;50:237–270. [Google Scholar]
- 26.Gratz NG. Plague Manual: Epidemiology, Distribution, Surveillance and Control. World Health Organization; Geneva: 1999. Rodent reservoirs and flea vectors of natural foci of plague; p. 63. [Google Scholar]
- 27.Dennis DT, Orloski K. Plague. In: Bernstein E, Tomchuck L, Brant J, editors. 1996 Medical and Health Annual. Encyclopaedia Brittanica; Chicago: 1996. p. 160. [Google Scholar]
- 28.Saxena VK, Vergese T. Ecology of flea-transmitted zoonotic infection in village Mamla, District Beed. Curr Sci. 1996;71:800. [Google Scholar]
- 29.Craven RB, Maupin GO, Beard ML. Reported cases of human plague infections in the United States, 1970–1991. J Med Entomol. 1993;30:758. doi: 10.1093/jmedent/30.4.758. [DOI] [PubMed] [Google Scholar]
- 30.Gage KL, Dennis DT, Orloski KA. Cases of cat-associated plague in the western U.S., 1977–1998. Clin Infect Dis. 2000;30:893. doi: 10.1086/313804. [DOI] [PubMed] [Google Scholar]
- 31.Mann JM, Martone WJ, Boyce JM. Endemic human plague in New Mexico: Risk factors associated with infection. J Infect Dis. 1979;140:397. doi: 10.1093/infdis/140.3.397. [DOI] [PubMed] [Google Scholar]
- 32.Christie AB, Chen TH, Elberg SS. Plague in camels and goats: Their role in human epidemics. J Infect Dis. 1980;141:724. doi: 10.1093/infdis/141.6.724. [DOI] [PubMed] [Google Scholar]
- 33.World Health Organization Human plague in 2000 and 2001. Wkly Epidemiol Rec. 2003;78:130. [PubMed] [Google Scholar]
- 34.Chanteau S, Ratsifasoamanana L, Rasoamanana B. Plague, a reemerging disease in Madagascar. Emerg Infect Dis. 1998;4:101. doi: 10.3201/eid0401.980114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Dennis DT. Plague as an emerging disease. In: Scheld WM, Craig WA, Hughes JM, editors. Emerging Infections 2. ASM Press; Washington, DC: 1998. p. 169. [Google Scholar]
- 36.Butler T, Bell WR, Nguyen NL. Yersinia pestis infection in Vietnam. I. Clinical and hematological aspects. J Infect Dis. 1974;129(Suppl):S78. doi: 10.1093/infdis/129.supplement_1.s78. [DOI] [PubMed] [Google Scholar]
- 37.Butler T. A clinical study of bubonic plague: Observations on the 1970 Vietnam epidemic with emphasis on coagulation studies, skin histology and electrocardiograms. Am J Med. 1972;53:268. doi: 10.1016/0002-9343(72)90168-4. [DOI] [PubMed] [Google Scholar]
- 38.Hull HF, Montes JM, Mann JM. Septicemic plague in New Mexico. J Infect Dis. 1987;155:113. doi: 10.1093/infdis/155.1.113. [DOI] [PubMed] [Google Scholar]
- 39.Butler T, Levin J, Nguyen NL. Yersinia pestis infection in Vietnam. II. Quantitative blood cultures and detection of endotoxin in the cerebrospinal fluid of patients with meningitis. J Infect Dis. 1976;133:493. doi: 10.1093/infdis/133.5.493. [DOI] [PubMed] [Google Scholar]
- 40.Crook LD, Tempest B. Plague—A clinical review of 27 cases. Arch Intern Med. 1992;152:1253. doi: 10.1001/archinte.152.6.1253. [DOI] [PubMed] [Google Scholar]
- 41.Hull HF, Montes JM, Mann JM. Plague masquerading as gastrointestinal illness. West J Med. 1986;145:485. [PMC free article] [PubMed] [Google Scholar]
- 42.Wu Lien-Teh: A Treatise on Pneumonic Plague. Geneva, League of Nations Health Organization, 1926.
- 43.Becker TM, Poland JD, Quan TJ. Plague meningitis—A retrospective analysis of reported cases in the United States, 1970–1979. West Med J. 1987;147:554. [PMC free article] [PubMed] [Google Scholar]
- 44.Dennis D, Meier F. Plague. In: Horsburgh CR, Nelson AM, editors. Pathology of Emerging Infections. ASM Press; Washington, DC: 1997. p. 21. [Google Scholar]
- 45.Finegold MJ. Pathogenesis of plague: A review of plague deaths in the United States during the last decade. Am J Med. 1968;45:549. doi: 10.1016/0002-9343(68)90171-x. [DOI] [PubMed] [Google Scholar]
- 46.Burmeister RW, Tigertt WD, Overholt EL. Laboratory-acquired pneumonic plague: Report of a case and review of previous cases. Ann Intern Med. 1962;56:789. doi: 10.7326/0003-4819-56-5-789. [DOI] [PubMed] [Google Scholar]
- 47.Cornelis GR. Yersinia type III secretion: Send in the effectors. J Cell Biol. 2002;158:401. doi: 10.1083/jcb.200205077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sing A, Rost D, Tvardovskaia N. Yersinia V-antigen exploits toll-like receptor 2 and CD14 for interleukin 10-mediated immunosuppression. J Exp Med. 2002;196:1017. doi: 10.1084/jem.20020908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kopp E, Medzhitov R. A plague on host defense. J Exp Med. 2002;196:1009. doi: 10.1084/jem.20021311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sodhi A, Sharma RK, Batra HV. Mechanism of rLcrV and rYopB mediated immunosuppression in murine peritoneal macrophages. Mol Immunol. 2004;41:767. doi: 10.1016/j.molimm.2004.04.029. [DOI] [PubMed] [Google Scholar]
- 51.Kerschen EJ, Cohen DA, Kaplan AM. The plague virulence protein YopM targets the innate immune response by causing a global depletion of NK cells. Infect Immun. 2004;72:4589. doi: 10.1128/IAI.72.8.4589-4602.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mann JM, Schmid GP, Stoetz PA. Peripatetic plague. JAMA. 1982;247:46. [PubMed] [Google Scholar]
- 53.Centers for Disease Control and Prevention Imported plague– New York City, 2002. MMWR. 2003;52:725. [Google Scholar]
- 54.Morse SA, Kellogg RB, Perry S. Detecting biothreat agents: The laboratory response network. ASM News. 2003;69:433. [Google Scholar]
- 55.Butler T, Hudson BW. The serological response to Yersinia pestis infection. Bull WHO. 1977;55:39. [PMC free article] [PubMed] [Google Scholar]
- 56.Chanteau S, Rahalison L, Ralafiarisoa L. Development and testing of a rapid diagnostic test for bubonic and pneumonic plague. Lancet. 2003;361:211. doi: 10.1016/S0140-6736(03)12270-2. [DOI] [PubMed] [Google Scholar]
- 57.Centers for Disease Control and Prevention. Fatal human plague— Arizona and Colorado, 1996. MMWR. 1997;46:617. [PubMed] [Google Scholar]
- 58.Boulanger L, Ettestad P, Fogarty J. Gentamicin and tetracyclines for the treatment of human plague: Review of 75 cases in New Mexico, 1985–1999. Clin Infect Dis. 2004;38:663. doi: 10.1086/381545. [DOI] [PubMed] [Google Scholar]
- 59.Frean JA, Arntzen L, Capper T. In vitro activities of 14 antibiotics against 100 human isolates of Yersinia pestis from a southern African plague focus. Antimicrob Agents Chemother. 1996;40:2646. doi: 10.1128/aac.40.11.2646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Russell P, Eley SM, Bell DL. Doxycycline or ciprofloxacin prophylaxis and therapy against experimental Yersinia pestis infection in mice. J Antimicrob Chemother. 1996;37:769. doi: 10.1093/jac/37.4.769. [DOI] [PubMed] [Google Scholar]
- 61.Galimand M, Guiyoule A, Gerbaud G. Multidrug resistance in Yersinia pestis mediated by a transferable plasmid. N Engl J Med. 1997;337:677. doi: 10.1056/NEJM199709043371004. [DOI] [PubMed] [Google Scholar]
- 62.Centers for Disease Control and Prevention. Prevention of plague. Recommendations of the Advisory Committee on Immunization Practices (ACIP) MMWR. 1996;45:1. [PubMed] [Google Scholar]
- 63.World Health Organization Technical guide for a system of plague surveillance. Wkly Epidemiol Rec. 1973;14:149. [Google Scholar]
- 64.Fritz CL, Dennis DT, Tipple MA. Surveillance for pneumonic plague in the United States during an international emergency: A model for control of imported emerging diseases. Emerg Infect Dis. 1996;2:30. doi: 10.3201/eid0201.960103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Titball RW, Williamson ED, Dennis DT. Plague. In: Plotkin SA, Orenstein WA, editors. Vaccines. 4th ed. WB Saunders; Philadelphia: 2004. p. 999. [Google Scholar]