

Graphical abstract
Abbreviations: AIDS, acquired immune deficiency syndrome; Akt, protein kinase B (PKB); ALB, albumin; ALP, alkaline phosphatase; AmD, Assoc American Dietetic Association; Anti-B, anti-bacterial; Anti-C, anti-cancer; Anti-D, anti-diabetic; Anti−HC, anti-hypercholesterolemic; Anti-F, anti-fungal; Anti-P, anti-parasitic; Anti-O, anti-obesity; Anti-V, anti-viral; ApoA-I, apolipoprotein A; ApoB, apolipoprotein B; AS, antioxidant status; B, bone; BCOV, bis(curcumino)oxavanadyl; BEOV, bis(ethylmaltolato)oxovanadium; Bim, Blc-2 interacting mediator of cell death; BMOV, bis(maltolato)oxavanadium(IV); breakD, breakdown; BrOP, bromoperoxidase; C, cholesterol; Cardio-P, cardioprotective; CD4, CD4 receptor; C/EBPα, CCAAT-enhancer-binding protein α; CH, cerebral hemisphere; CHO-K1, Chinese hamster ovary cells; Citrate-T, citrate transporter; CoA, coenzyme A; CXCR-4, CXCR-4 chemokine co-receptor; Cyt c, cytochrome c; DM, diabetes mellitus; ELI, extra low interstitial; eNOS, endothelial nitric oxide synthase; ERK, extracellular regulated kinase; FasL, Fas ligand, FER: ferritin; FHR, fructose hypertensive rats; FKHR/FKHR1/AFX, class O members of the forkhead transcription factor family; FLIP, FLICE-inhibitory protein; FOXOs, forkhead box class O family member proteins; FPP, farnesyl-pyrophosphate; GI, gastrointestinal; GLU, glucose; GLUT-4, glucose transporter type 4; GPP, geranyl-pyrophosphate; GPT, glutamate-pyruvate transaminase; GR, glutathione reductase; GSH, reduced glutathione; GSSG, disulfide glutathione; Hb, hemoglobin; HbF, hemoglobin fraction; HDL, high-density lipoproteins; HDL-C, HDL cholesterol; HIV, human immunodeficiency virus; 3-HMG-CoA, 3-hydroxy-3-methyl-glutaryl-CoA; HMMF, high molecular mass fraction; HOMA-IR, insulin resistance index; Hyper-LEP, hyperleptynemia; IDDM, insulin-dependent diabetes mellitus; IGF-IR, insulin-like growth factor receptor; IgG, immunoglobulin G; INS, insulin; INS-R, insulin resistance; INS-S, insulin sensitivity; IL, interleukin; IPP, isopentenyl-5-pyrophosphate; IRS, insulin receptor tyrosine kinase substrate; JAK2, Janus kinase 2; K, kidney; L-AA, L-ascorbic acid; Lactate-T, lactate transporter; LDL, low-density lipoproteins; LDL-C, LDL cholesterol; LEP, leptin; LEP-R, leptin resistance; LEP-S, leptin sensitivity; LEPS, the concentration of leptin in the serum; L, liver; LMMF, low molecular mass fraction; LPL, lipoprotein lipase; LPO, lipid peroxidation; M, mitochondrion; MEK, ERK kinase activator; mo, months; MRC, mitochondrial respiratory chain; NAC, N-acetylcysteine; NaVO3, sodium metavanadate; NEP, neutral endopeptidase; Neuro-P, neuroprotective; n-HA, nano-hydroxyapatite; NIDDM, noninsulin-dependent diabetes mellitus; NO, nitric oxide; NPY, neuropeptide Y; Organic-AT, organic anion transporter; Over-W, over-weight; OXPHOS, oxidative phosphorylation; P, plasma; PANC-1, pancreatic ductal adenocarcinoma cells; PARP, poly (ADP-ribose) polymerase; Pi3K, phosphoinositide 3-kinase (phosphatidylinositol 3-kinase); PLGA, (Poly)Lactide-co-Glycolide copolymer; PO43−, phosphate ion; PPARγ, peroxisome-activated receptor γ; pRb, retinoblastoma protein; PTK, tyrosine protein kinase; PTP, protein tyrosine phosphatase; PTP-1B, protein tyrosine phosphatase 1B; RBC, erythrocytes; ROS, reactive oxygen species; RT, reverse transcriptase; Sa, mean roughness; SAcP, acid phosphatase secreted by Leshmania; SARS, severe acute respiratory syndrome; SC-Ti-6Al-4V, surface-coated Ti-6Al-4V; SHR, spontaneously hypertensive rats; SOD, superoxide dismutase; Sq, root mean square roughness; STAT3, signal transducer/activator of transcription 3; Sz, ten-point height; TC, total cholesterol; Tf, transferrin; TfF, transferrin fraction; TiO2, nHA:Ag-Ti-6Al-4V: titanium oxide-based coating containing hydroxyapatite nanoparticle and silver particles; TG, triglycerides; Top-IB, IB type topoisomerase; TS, transferrin saturation; V, vanadium; V5+/V4+, pentavalent/tetravalent vanadium; V-BrPO, vanadium bromoperoxidase; V-DLC, diamond-like layer with vanadium; VO4-/VO3-, vanadate anion; VO43-, vanadate ion; VO2+, vanadyl cation; VO2+-FER, vanadyl-ferritin complex; VS, vanadyl sulfate; WB, whole blood; wk, weeks; ZDF rats, Zucker diabetic fatty rats; ZF rats, Zucker fatty rats
Keywords: Vanadium, Biological role, Toxicological potential, Pharmacological activity, Industrial importance, Interactions
Abstract
Background
Vanadium (V) is an element with a wide range of effects on the mammalian organism. The ability of this metal to form organometallic compounds has contributed to the increase in the number of studies on the multidirectional biological activity of its various organic complexes in view of their application in medicine.
Objective
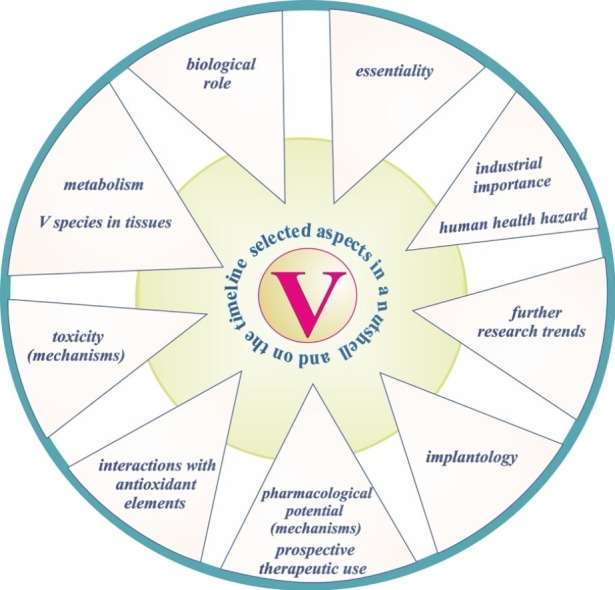
This review aims at summarizing the current state of knowledge of the pharmacological potential of V and the mechanisms underlying its anti-viral, anti-bacterial, anti-parasitic, anti-fungal, anti-cancer, anti-diabetic, anti-hypercholesterolemic, cardioprotective, and neuroprotective activity as well as the mechanisms of appetite regulation related to the possibility of using this element in the treatment of obesity. The toxicological potential of V and the mechanisms of its toxic action, which have not been sufficiently recognized yet, as well as key information about the essentiality of this metal, its physiological role, and metabolism with certain aspects on the timeline is collected as well. The report also aims to review the use of V in the implantology and industrial sectors emphasizing the human health hazard as well as collect data on the directions of further research on V and its interactions with Mg along with their character.
Results and Conclusions
Multidirectional studies on V have shown that further analyses are still required for this element to be used as a metallodrug in the fight against certain life-threatening diseases. Studies on interactions of V with Mg, which showed that both elements are able to modulate the response in an interactive manner are needed as well, as the results of such investigations may help not only in recognizing new markers of V toxicity and clarify the underlying interactive mechanism between them, thus improving the medical application of the metals against modern-age diseases, but also they may help in development of principles of effective protection of humans against environmental/occupational V exposure.
1. Introduction
1.1. A general overview
With its unique features, vanadium (V) receives a great deal of attention from chemists, biologists, biochemists, toxicologists, and pharmacologists. The biological activity of this element has prompted investigations of many organic V complexes and its inorganic compounds in terms of their potential use in the treatment of certain diseases in humans. Studies carried out so far on V have shown that the bioactive complexes/compounds of this metal can be therapeutically active at low concentrations [1,2].
Taking into account the clear interest in the anti-viral, anti-bacterial, anti-parasitic, anti-fungal, anti-cancer, anti-diabetic, anti-hypercholesterolemic, cardioprotective, and neuroprotective activity of V and in the possibility of using this element in the treatment of obesity, the present review focuses on mechanisms underlying the pharmacological potential of this metal and obstacles to use it as a metallopharmaceutical in the future. Key information about the mechanisms of the toxicity, essentiality, physiological role, and metabolism of V is collected as well along with selected aspects illustrated on the timeline. The review paper also summarizes the available literature data on the use of V in the tissue engineering and industrial sectors with a brief description of the risk for human health. In addition, our report draws attention to the directions of further research on V and to interactions of this metal with other elements, especially those with antioxidant potential. Comprehensive research on the interactions of V with antioxidant elements (summarized graphically in a separate chapter of the present report) is particularly important in view of minimization of the adverse effects of V resulting from its strong toxicological/pro-oxidative potential.
1.2. Vanadium – background
Vanadium (V) is a part of the 4th period, Group 5 (VB), of the periodic table of elements. It is a transition element (with atomic number 23 and relative atomic weight of 50.94) capable of forming various compounds and functioning as an anion or a cation [3]. It exists in +2, +3, +4, and +5 oxidation states, most commonly in the tetravalent and pentavalent form [2] (Fig. 1 ). In body fluids and extracellularly, V predominates in the pentavalent form, usually as a vanadate anion (VO3 −). Conversely, intracellularly, it exists mainly in the tetravalent form, as a vanadyl cation (VO2+) (Fig. 1) associated, inter alia, with proteins [4]. It is estimated that about 1% of intracellular VO2+ occurs in an unbound form [5]. It is known that the form in which V exists in the cell depends on the ratio of the total pool of reducers to cellular oxidizers [4].
Fig. 1.

Vanadium and its most common forms. Based on available literature data cited in section 1.1.
1.3. Vanadium – a brief historical outline with a Polish accent on the timeline
As presented in Fig. 2 , vanadium was first discovered at the beginning of the 19th century, in 1801, by Spanish mineralogist Andrés Manuel del Río, who named it “erythronium” from the red color of the compound. However, the discovery was not attributed to him. Due to a mistake made in 1805 by one of the members of the Commission of the French National Institute in Paris, i.e. chemist Collet-Descostills, the discovery was not accepted. This was followed by a rediscovery of the element in 1830 by Swedish physician and chemist Nils Gabriel Sefström, who named it „vanadium” [6]. In Polish chemical literature, Radwański described this element for the first time in 1839 giving it a Polish name "wanad", which is still used [7].
Fig. 2.

Historical outline of vanadium discovery. V: vanadium. Based on available literature data cited in section 1.2.
1.4. Vanadium: a summarizing note about biological/industrial importance and pharmacological applications
Vanadium is ubiquitously detected in the environment - in soil, water, and air [8]. It plays an important biological role [4] and has a very wide area of applications, which are graphically summarized in Fig. 3 and described in the further subsections of the present report.
Fig. 3.

Industrial, biological, and pharmacological importance of vanadium. V: vanadium, NI: normal intake.
2. Vanadium – industry
As illustrated in Fig. 3, vanadium (V) is commonly used in many industries, especially in glass, paint, ceramic, photographic, chemical, electrochemical, and refining industries. It is also used in metallurgical industry for production of steel and non-ferrous alloys [1,9,10]. Vanadium-containing steel (with high hardness, strength, elasticity, and abrasion resistance) has been widely used as spring steel, tool steel, and high-speed steel. Currently, steel with the addition of V and V-containing non-ferrous alloys are used to build jet engines, machine parts (including construction equipment), as well as aircraft and automobile components. Non-iron alloys containing V are also used in space technology and nuclear power industry. Moreover, steel with V was well suited for production of armored fighting vehicles used during the Second World War [[11], [12], [13], [14], [15], [16]]. More details about V-containing steel and alloys, which were and still are used to produce various structural components of aircraft, weapon, rocket engines, etc., are summarized in the following table (Table 1 ).
Table 1.
Use of vanadium in certain industrial sectors.
| Arm/military/other | Chemical composition of steel/alloy |
References | |||
|---|---|---|---|---|---|
| Elements | Content [%] | Vanadium | Content [%] |
||
| Steel with vanadium addition | |||||
| Armored fighting vehicles | |||||
| PzKw V (Panther) tank (Panzerkampfwagen V Panter) | C Cr |
0.50 2.13 |
V | 0.14 | [17] |
| PzKw V (Panther) tank (Panzerkampfwagen V Panter) | C Cr |
0.44 1.72 |
V | 0.10 | [18] |
| Panzerkampfwagen II (PzKpfw II) | C Cr Si Mo Cu Ni |
0.40 1.3 0.25 0.50 0.18 trace |
V | 0.17 | [19] |
| Combat aircraft/parts for aircraft and rockets/other | |||||
| Alloys with vanadium addition | |||||
| Ti-6Al-4V | |||||
| Components parts for weapons and aircraft (engine parts, fasteners) | Ti Al |
90 5.5−6.76 |
V | 3.5−4.5 | [12,13,14,15] |
| Ti-13V-11Cr-3Al | |||||
| SR-71 Blackbird Component parts aircraft, missile applications |
Ti Cr Al |
∼75 10-12 2.5−3.5 |
V | 12.5−14.5 | [13,14,20] |
| Ti-3Al-2.5 V | |||||
| Component parts for aircraft (pipes in hydraulic systems) | Ti Al |
95.755-95.5 2.5−3.5 |
V | 2−3 | [13,14] |
| Ti-8Al-1Mo-1V | |||||
| Component parts for aircraft (compressor blades) | Ti Al Mo |
90 7.35-8.35 0.75-1.25 |
V | 0.75−1.75 | [14] |
| Ti-6Al-6 V-2Sn | |||||
| Component parts for aircraft (jet engines), rocket engine housings, weapon components | Ti Al Sn |
82.89-87.8 5−6 1.5-2.5 |
V | 5−6 | [14,20] |
| Ti-10V-2Fe-3Al | |||||
| Component parts for aircraft (landing gear, parts responsible for take-off and landing) | Ti Al Fe |
82.8-86.8 2.6-3.4 1.6-2.2 |
V | 9.0−11.0 | [12,13,14,16] |
| Ti-15 V-3Cr-3Al-3Sn | |||||
| Component parts for aircraft (hulls, wires) | Ti Al Cr Sn |
76 2.5−3.5 2.5−3.5 2.5−3.5 |
V | 14−16 | [13,14,16] |
| Ti-3Al-8V-6Cr-4Mo-4Zr | |||||
| Component parts for aircraft (fasteners) | Ti Al Cr Mo Zr |
75 3-4 5.5−6.5 3.5−4.5 3.5−4.5 |
V | 7.5−8.5 | [14,16] |
| Ti-3.5Al-5Mo-6 V-3Cr-2Sn-0.5Fe | |||||
| Aerospace industry | Ti Al Mo Cr Sn Fe |
∼86 3.5 5 3 2 0.5 |
V | 6 | [16] |
| Ti-5Al-5Mo-5 V-1Cr-1Fe | |||||
| Aerospace industry | Ti Al Mo Cr Fe |
∼88 5 5 1 1 |
V | 5 | [16] |
| Ti-5Al-5 V-5Mo-3Cr-0.5Fe | |||||
| Aerospace industry | Ti Al Mo Cr Fe |
∼86 5 5 3 0,5 |
V | 5 | [16] |
| Ti-5Al-5Mo-5 V-3Cr-1Zr | |||||
| Aerospace industry | Ti Al Mo Cr Zr |
∼86 5 5 3 1 |
V | 5 | [16] |
| Ti-5 V-5Mo-5Al-3Cr | |||||
| Aerospace industry | Ti Mo Al Cr |
∼87 5 5 3 |
V | 5 | [16] |
| Ti-3Al-8V-6Cr-4Zr-4Mo | |||||
| Aerospace industry | Ti Al Cr Zr Mo |
∼83 3 6 4 4 |
V | 8 | [16,20] |
3. Vanadium – pharmacological aspect
As shown in Fig. 3, vanadium is also a potential candidate for therapeutic applications. Its anti-viral, anti-bacterial, anti-parasitic, anti-fungal, anti-cancer, anti-diabetic, and anti-hyper-cholesterolemic activity and cardioprotective, neuroprotective, and anti-obesity effects have been arousing interest of many research centers worldwide for many years. It has been reported that in human beings, pharmacologic amounts of V, i.e. 10 to 100 times higher than the normal intake, affect cholesterol and triglyceride metabolism, influence the erythrocyte shape, and stimulate hepatic glucose oxidation and glycogen synthesis [21]. In addition, V (in certain conditions) has been found to act as an antioxidant (Fig. 3), as evidenced in animal studies [[22], [23], [24], [25]]. More details about the mechanisms of the pharmacological activity of V are graphically illustrated and summarized in another part of the present report.
4. Vanadium – a physiological role: a summary of the most important issues
The research on certain aspects of the biological activity of V (presented in Fig. 4 ) has demonstrated an essential role of this element in the metabolism of carbohydrates (through the effects on the glycolysis, glycogenolysis, glycogenogenesis, and gluconeogenesis pathways), lipids (by stimulation of lipogenesis and inhibition of lipolysis), phospholipids, and cholesterol. The influence of V on bone mineralization, thyroid and erythrocyte metabolism, accumulation and transport of calcium in the cell, and the synthesis of secondary transmitters mediating in the transduction of intracellular signals is known as well. Furthermore, V also regulates the activity of key enzymes involved in the phosphorylation and dephosphorylation of proteins, kinases, and phosphatases, taking part not only in carbohydrate and lipid metabolism but also in cell proliferation and differentiation [1,2,4,26,27].
Fig. 4.

Role of vanadium in the mammalian organism. Based on available literature data cited in section 4.
5. Vanadium – a biphasic effect
5.1. Summarizing note with certain aspects of essentiality of V on the timeline
With its physiological duality, vanadium is essential in trace amounts (0.05 μM) and toxic in excess (>10 μM) [28,29] (Fig. 5 ). Its deficit and excessive concentrations can lead to a range of pathologies and cause irreversible damage to various tissues and organs.
Fig. 5.

Summary of positive and negative aspects of vanadium. AmDAssoc: American Dietetic Association. Based on available literature data cited in section 5.1 and other reports [[30], [31], [32], [33], [34]].
At low concentrations, V has been found to exert a beneficial effect on the growth and physiological functions of some microorganisms, plants, and fungi. The essentiality of this element has been demonstrated for bacteria (genus Azotobacter), cyanobacteria (genera Nostoc and Anabaena), certain algae (genera Scenedesmus, Chlorella, Fucus, and Bumilleriopsis), brown algae (Ascophyllum nodosum), fungi (genera Aspergillus, Culvularia, and Amanita), and lichens [[35], [36], [37]] (Fig. 5).
A positive effect of V on nitrogen (N) fixation (by Azotobacter) has been suggested by Bortels in 1933 (Fig. 6 ). In 1984 and in 1986, V turned out to be essential for the activity of certain enzymes [[42], [43], [44]] such as bromoperoxidase (BrOP), nitrogenases, and chloroperoxidase isolated from algae, nitrogen-fixing bacteria, and fungus Culvularia inaequalis, respectively [36,37,45] (Fig. 6).
Fig. 6.

Summary of selected aspects of vanadium toxicity/essentiality on the timeline. V: vanadium, N: nitrogen, V-BrPO: vanadium bromoperoxidase. Based on available literature data cited in section 5.1 and other reports; †after [33], **after [38], ††after [39], *after [40], ##after [41].
Furthermore, vanadium has also been recognized as essential for proper growth and development of certain animals [29,35]. The first suggestion about the essentiality of V for animals appeared in 1949, i.e. nearly 150 years after the first discovery of this metal [after 38] (Fig. 2). About 22 years later, in 1971, Schwarz and Milne, Hopkins and Mohr, and Strasia reported that V is necessary for rats and chicks (Fig. 6). In turn, in 1989, Anke and co-authors also reported that V is essential for goats (Fig. 6). In the meantime, in 1981, the first review about the physiological and biochemical effects of V was published (Fig. 6).
To date, symptoms of V deficiency have been described, inter alia, in birds, chickens, rats, guinea pigs, and goats [35,38,46]. The most substantive evidence for V essentiality was found in rats and goats [after 35]. On this basis, Nielsen and Uthus proposed considering vanadium as an essential trace element for proper growth and development of higher animals [38] In contrast, the essentiality of this metal for humans is unproven (Fig. 5), although some authors suggest that it is an essential trace element for man [38], mostly because vanadium has broad pharmacological activity, which points to the biological importance of this metal. The American Dietetic Association (AmDAssoc) considers V as essential in human nutrition [after 41] (Fig. 5). Nevertheless, no symptoms of V deficiency in humans have been described yet. The final proof for the essentiality of V for humans requires confirmation of the specific function of this element, as in the case of algae or fungi. However, it is definitely known that the high content of V in the environment and diet is harmful to animals and humans [1].
5.2. Summarizing note with certain aspects of toxicity of V on the timeline
As generally accepted, the toxicity of V is dependent on many different factors, including the composition of the diet, type of the V compound (inorganic/organic), nature of ligands attached to V complexes, valence, dose, route of entry of this metal into the organism, duration of action/exposure, and individual and species sensitivity (Fig. 7 ).
Fig. 7.

Summary of the mechanisms and factors of vanadium toxicity: vanadium; LPO: lipid peroxidation; ROS: reactive oxygen species; MRC: mitochondrial respiratory chain; OXPHOS: oxidative phosphorylation; M: mitochondrion; Cyt c: cytochrome c; PARP: poly (ADP-ribose) polymerase. Based on available literature data cited in section 5.2.
The history of research on the adverse effects of action of this metal dates back to 1876 when Priestly described toxicity of V in certain animals (Fig. 6). As regards animals, rabbits and guinea pigs were found to be more sensitive to this element than mice and rats [5,47,48] (Fig. 7). In turn, a classic paper about the toxic and pharmacologic action of vanadium was written in 1912 and analyses of the content of V in various organisms were initiated 18 years later in 1930 (Fig. 6).
It is known that, in some conditions, V can act as a strong pro-oxidant and interact synergistically with other oxidants enhancing oxidative stress [2,31,49], which in turn can result in many negative consequences including disintegration of cell membranes, denaturation of proteins, and degradation of DNA [50] (Fig. 7). It may also weaken the antioxidant barrier [49,[51], [52], [53]] and intensify lipid peroxidation (LPO) [49,[53], [54], [55], [56], [57], [58]] i.e. the free radical process underlying one of the mechanisms of cell damage [50] (Fig. 7). In addition, V can reduce the thiol status [49,59], release some transition metals [60], interact with other elements [49,53,61], and accumulate in certain internal organs including the liver and kidneys [46,56,[62], [63], [64]] evoking hepato- and nephrotoxic effects, respectively (Fig. 7). In higher concentrations, V can also act as a pro-apoptotic factor (by induction of oxidative stress) and lead to programmed cell death (apoptosis) with damage to mitochondrial membranes, cytochrome c (cyt c) outflow, and activation of caspases and poly (ADP-ribose) polymerase (PARP) [5] (Fig. 7), which plays a role of a cell death promoting factor [65]. Disorders of the electron transport chain and oxidative phosphorylation resulting in a decrease in the level of ATP [1] are the other adverse effects of V action (Fig. 7).
The mechanisms of toxicity of V require further experimental work, as they have not been fully elucidated yet. Some of them have been reported to be implicated in the adverse effects of V action. For example, the mechanism by which V produces a reprotoxic effect is believed to be linked to oxidative stress [66] and/or actin cytoskeleton damage [67] (Fig. 7). It has been suggested that the hepatotoxic, nephrotoxic [48,62,[68], [69], [70], [71]], cardiotoxic [72], genotoxic [73,74], neurotoxic [72,75,76], and carcinogenic [48,77] effects of this metal as well as inflammatory and fibrotic changes in the lungs [78] are associated with the pro-oxidative potential of V and its ability to intensify LPO (Fig. 7). It is known that LPO products can stimulate the expression of oncogenes [79], which in turn can lead to transformation of normal cells into cancerous cells (Fig. 7). Epidemiological studies suggest that exposure to elevated levels of V may be a risk factor for cancer development [80]. Studies on an animal model have revealed a positive correlation between the frequency of neoplasia and susceptibility to V-induced inflammation [80], which is involved in the pathogenesis of many diseases (not only malignant diseases) and results from V-induced oxidative stress. In turn, long-lasting oxidative stress with simultaneous weakening of the antioxidant system (present in every living organism) can lead to permanent cell/tissue damage and, consequently, to initiation of the disease process. Therefore, the generation of reactive oxygen species (ROS) under oxidative stress (one of the main factors affecting morbidity) can lead to damage to DNA and, consequently, to initiation of carcinogenesis.
6. Vanadium – health hazards
6.1. Environmental/occupational exposure to V: the most important aspects
The risk for animal and human health related to exposure to V is largely dependent on the degree of its oxidation state. It has been highlighted that V at the highest oxidation state (+5) is the most toxic vanadium form [9]. One of the compounds of this metal, vanadium pentoxide (V2O5), which contains V at the highest oxidation state +5, has been reported to easily reach the alveolar surface, subsequently enter circulation directly, and diffuse in the entire organism inducing diverse injuries [9]. The risk of poisoning with the pentavalent V form is constantly growing due to its extensive release into the environment by metallurgical industry, chemical plants, oil refineries, and coal and mazut-fired heating plants [8,9,45,81]. Moreover, relatively high pollution of the environment with V2O5 is also caused by dust derived from the combustion of liquid and solid fuels as well as municipal waste [10,81], resulting in an increased risk of exposure of anyone who lives near such areas. Consequently, the level of this metal in the soil increases. In turn, higher concentrations of V in the soil lead to more intensive uptake of this metal being by plants due to the lack of a selective mechanism protecting plants against excessive absorption of the element. Thus, V can enter the animal and human organism via the food chain. Additionally, the lack of proper industrial wastewater management containing this metal can be another risk factor for animal and human health [9,82].
In the case of occupational exposure, workers repairing thermal power boilers and high-pressure combustion engines as well as those responsible for production of V2O5 and replacement of V-based catalytic converters are the most vulnerable to the most toxic pentavalent form of this metal [9]. In the literature, occupational exposure to V2O5 is described as the etiological factor of the asthmatic syndrome known as "boilermaker disease" [83].
6.2. Basic symptoms of exposure to V via inhalation and ingestion: a brief note
The most important symptoms of exposure to V via the oral route (presented in Fig. 8 ) include gastrointestinal disorders, abdominal pain, nausea, vomiting, diarrhea, loss of appetite, weight reduction, and green-black tongue. In turn, the symptoms of exposure to this metal through inhalation (illustrated in the same Figure) include rhinitis, chest pain, pharyngitis, bronchitis, pneumonia, bradycardia, cough, dyspnoea, bronchial asthma, headache, dizziness, conjunctivitis, blurred vision, apathy, and depression [9,31,81,[84], [85], [86]].
Fig. 8.

Major symptoms of vanadium poisoning via ingestion and inhalation. Based on available literature data cited in section 6.2.
7. Vanadium - metabolism: a summary of key information
The most important issues of the metabolism of V and forms in which this metal is present in physiological fluids/tissues are collected in Fig. 9 . As illustrated, V consumption, which depends on the diet, is estimated to be around 15−20 μg/day (typical daily dose consumed by humans) or 10−60 μg/day (daily dose ingested by the U.S. population) [8,87]. After intake, vanadate (mainly as VO4 −) reaches the gastrointestinal (GI) tract and, in the acidic environment of the stomach, most of the anionic vanadate (absorbed more effectively than VO2+) is transformed into vanadyl (VO2+), precipitated to insoluble vanadyl hydroxides [VO(OH)2] in the slightly alkaline medium of the intestine, and excreted with feces (∼94−98 %) [33,39]. In turn, resorbed V is removed from the organism through kidneys [87,88] probably in the form of unidentified complexes with high-(protein-bound) and low-molecular-weight species (VO2+-complexes) [38,89]. The level of V in urine is estimated to be around 12 % of the intake amount [87].
Fig. 9.

Summary of metabolism of vanadium and species of this metal in physiological fluids and tissues. V: vanadium, V5+: pentavalent vanadium, V4+: tetravalent vanadium, VO4−/VO3−: vanadate anion, VO2+: vanadyl cation, V*: exchangeable vanadium (‘free’), VO(OH)2: insoluble vanadyl hydroxide, GSH: reduced glutathione, Hb: hemoglobin, RBC: erythrocytes, ALB: albumin, Tf: transferrin, IgG: immunoglobulin G, FER: ferritin, Citrate-T: citrate transporter, Lactate-T: lactate transporter, Organic-AT: organic anion transporter, LMMF: low molecular mass fraction, HMMF: high molecular mass fraction, TfF: transferrin fraction, HbF: hemoglobin fraction. Based on available literature data cited in section 7 and other reports [60,90,91].
As reported, the absorption of V from the GI tract is mainly estimated in the range of 0.2–2 %, although amounts greater than 10 % have been indicated to be absorbed as well (Fig. 9) [21,46,92,93]. Additionally, Sanchez et al. [94] found an absorption rate of V at the level of 52 % in rats treated with bis(maltolato)oxovanadium(IV) (BMOV, 6.22 mg/day) in drinking water for five weeks. It has been highlighted that such differences may be associated with several factors, inter alia, the diet composition and/or chemical form of ingested V [38]. After entering the bloodstream, 80–90 % of V is bound by albumin (ALB), which plays a pivotal role in the transport of various metals [95] or transferrin (Tf) (preferably), which is the main V transporter in the blood plasma [8,33]. Both Tf and ALB bind V in the 4+ and 5+ oxidation state [96].
It has been reported that V in the form of vanadyl (VO2+, tetravalent V4+ state) binds to Tf at the same binding site as the Fe3+ ion [97] and that V5+, for which the cationic form (dioxovanadium cation, VO2 +) has been indicated, also occupies the same pockets as Fe3+ [98]. Studies conducted by Azevedo et al. [99] have confirmed that both V4+ and V5+ can bind to apo-Tf and holo-Tf, thus they can be efficiently up-taken by cells through receptor-mediated endocytosis of Tf. In addition, Tf can bind VIII forming di-vanadium(III)-Tf [(VIII)2-Tf], which can be up-taken by cells via receptor-mediated endocytosis as well [100]. It has also been reported that V species can enter cells via membrane citrate transporters, the lactate transporter, and the organic anion transporter (Organic-AT) [101]. Additionally, VO2+ can be up-taken via passive diffusion [102,103] and bound by immunoglobulin G (IgG) or low-molecular components of plasma such as phosphate, citrate, lactate, and oxalate [33,39,91,104,105]. In turn, V5+ ions such as H2VO4 −/HVO4 2- can enter cells through anion channels, i.e. phosphate or sulfate channels [106]. In cells, V5+ is reduced to VO2+ by some reducing substances such as ascorbic acid and thiol-containing cysteine [107]. For instance, in erythrocytes (RBC), V5+ is principally reduced by glutathione (GSH) to VO2+, which binds to hemoglobin (Hb) [108,109] (Fig. 9).
The total content of V in the organism of adults is about 100−200 μg (equivalent to the mean tissue concentration of about 40 nM) [6]. Half of this amount is located in bones, which are the major storage pool for long-term V accumulation and where vanadate can substitute phosphate in the mineral hydroxyapatite [Ca10(PO4)6(OH)2] [33,37,106] (Fig. 9). The other amount is mainly deposited in the liver, kidney, and spleen [108] due to the importance of these organs in detoxification of the organism and excretion of harmful substances. Muscles, lungs, and brain are the other sites of V accumulation [108] (Fig. 9). As regards V speciation, most of this metal in the liver is present in the transferrin and low-molecular mass fraction (TfF and LMMF, respectively) [108] as well as a vanadyl-ferritin [(VO2+)-FER] complex [26]. The FER-V and LMMF-V fractions have also been identified in the spleen [108]. In kidneys, V was divided between a high molecular mass fraction (HMMF) and LMMF; additionally, an exchangeable fraction of this metal has been indicated [108] (Fig. 7). In turn, large amounts of V in the lungs have been identified in the hemoglobin and Tf fraction (Hb-F and TfF, respectively) and in the readily exchangeable fraction [108] (Fig. 9).
8. Vanadium – medical aspect in a nutshell
8.1. Biomedical materials with vanadium
While discussing V in terms of its medical application, it has to be mentioned that this metal is applied in tissue engineering to obtain biomaterials that allow regeneration of damaged tissues/organs and restoration of their lost functions [110]. As presented in another subsection of this paper, V is part of metallic biomaterials (mainly titanium alloys), bioactive coatings (including polymeric coatings), and diamond-like layers applied to the surface of some titanium alloys in order to increase the bioactivity of implants and improve osseointegration [11].
8.1.1. Metallic biomaterials and bioactive coatings with vanadium
Two titanium alloys with V such as Ti-6Al-4 V and Ti-6Al-4 V ELI are used as biomedical materials, mainly in bone surgery, due to their good mechanical properties, very high resistance to corrosion, and biocompatibility [11] (Table 2 ). In addition, 2V-49Co-49Fe (named Permendur) is another vanadium-containing alloy. 2V-49Co-49Fe with 2 % of V is mainly used for production of devices supporting the work of the heart (Table 2) [11].
Table 2.
Vanadium-containing biomaterials and their use in medical devices.
| Metallic biomaterials | |||||
|---|---|---|---|---|---|
| Type of alloy | Percentage composition [%] | Application | Reference | ||
| Ti-6Al-4V | Ti 90 | Al 6 | V 4 | hip joint replacement knee joint replacement dental implants (zygomatic implants), bridges/crowns, dental bridges orthodontic implants spine stabilizers trauma devices/bone fixation (intramedullary rods and nails, surgical screws/bone plates) encapsulation of cardiac pacemaker |
[11,13,111,112,113,114,115,116,117,118,119] |
| Ti-6Al-4V ELI | Ti 90 | Al 6 | V 4 | medical implants | [11,13] |
| Ti-3Al-2.5V | Ti 92.755−95.5 | Al 2.5−3.5 | V 2−3 | dental implants hip and knee joint replacement (orthopedic implants) trauma devices (intramedullary rods) |
[11] |
| Magnetic alloys | |||||
| 2V-49Co-49Fe (Permendur) | Co 49 | Fe 49 | V 2 | devices supporting the work of the heart | [11] |
| Biocompatible coatings applied on metallic alloys | |||||
| TiO2:nHA:Ag-Ti-6Al-4V SC-Ti-6Al-4V V-DLC |
Ti 90 Al 6 V 4 Ti 90 Al 6 V 4 – |
orthopedic/dental implants hip joint replacement trauma/orthopedic surgery |
[11,120,121] | ||
| Polymeric composite biomaterials | |||||
| Composition of polymeric coating | Properties | Application | Reference | ||
| V2O5/PLGA | antimicrobial activity | implant materials | [122] | ||
Ti: titanium; Al: aluminum; V: vanadium, ELI: extra low interstitial (max. Fe: 0.14 %, max. O: 0.13 %). TiO2:nHA:Ag-Ti-6Al-4V: titanium oxide-based coating containing hydroxyapatite nanoparticle and silver particles, SC-Ti-6Al-4V: surface-coated Ti-6Al-4 V, V-DLC: diamond-like layer with vanadium (DLC type carbon layer), V2O5: vanadium pentoxide; PLGA: (Poly)Lactide-co-Glycolide copolymer.
The presence of vanadium in biomedical materials requires our additional comment. Since implants made of titanium alloys (containing V) are exposed to body fluids, V may be released into surrounding tissues, exerting adverse effects. The fact that the implant surface has a significant impact on reactions that occur at the implant-tissue interface should also be taken into account. Therefore, the surface layer of the implant is often modified in order to induce a specific reaction of the tissue to the implant. In the case of some implants (e.g. orthopedic and dental implants), a surface with appropriate bioactivity and roughness is created to achieve rapid osseointegration and a durable connection between biomaterial and tissue [11].
As reported, layers with varied composition are often applied on the surface of metallic alloys in order to improve the biological activity of the implant and provide an additional barrier against the release of metals from metallic biomaterials [11]. For example, titanium oxide(TiO2)-based coating containing nano-hydroxyapatite (n-HA) and silver particles (Ag) is applied on the Ti-6Al-4 V alloy (Table 2) [120]. The TiO2: n-HA: Ag layers are used as bioactive coatings with antimicrobial properties to increase biological activity, osseointegration, and biochemical stability in implantable medical devices (Table 2).
Moreover, polymeric V-based coatings made of a (Poly)Lactide-co-Glycolide copolymer (PLGA) with antimicrobial activity (Table 2) [122] and a diamond-like layer including V (V-DLC) are also formed to be used as implant materials [11] (Table 2).
8.2. Vanadium as a potential drug in the treatment of certain modern-age diseases
Research on the pharmacological potential of V (in vitro and in vivo experimental models) has shown that some compounds/complexes of this element can be effective against: (a) viruses (including HIV-1 and HIV-2 immunodeficiency virus, dengue virus, SARS virus, and influenza virus) (Fig. 10 ) [33,123] responsible for acquired immune deficiency syndrome (AIDS), dengue fever, severe acute respiratory syndrome (SARS), and acute respiratory infection (influenza), respectively; (b) parasitic protozoa of the genus Trypanosoma responsible for American trypanosomiasis and African trypanosomiasis known as Chagas disease and sleeping sickness, respectively; protozoan parasites of the genera Leishmania and Entamoeba (Fig. 10) [39,45,[123], [124], [125], [126], [127], [128], [129]] responsible for the development of leishmaniasis and amoebiasis, respectively; (c) fungi from the genera Candida, Aspergillus, Trichophyton, and Microscopus (Fig. 10) [[130], [131], [132], [133], [134], [135]] responsible for the development of many fungal infections, and (d) gram-negative and gram-positive bacteria (Fig. 10) [26,123,124,[131], [132], [133],136,137] causing e.g. food poisoning, gastrointestinal and respiratory infections, typhoid fever, pneumonia, strep throat, tuberculosis, and skin diseases. It was also shown that certain V compounds/complexes may have anti-cancer, anti-diabetic, and anti-hypercholesterolemic activity and can act as cardioprotective and neuroprotective agents. More details about these issues along with relevant literature data are provided in sections 8.2.2.5 (anti-cancer), 8.2.2.6 (anti-diabetic), 8.2.2.7 (cardioprotective), 8.2.2.8 (neuroprotective), and 8.2.2.10 (anti-hypercholesterolemic activity). Moreover, the possibility of using this metal in prevention of obesity has been suggested as well [138] (Fig. 11 ).
Fig. 10.

Vanadium against certain viruses, bacteria, parasites, and fungi. Based on available literature data [26,33,39,45,[124], [125], [126], [127], [128], [129], [130], [131], [132], [133], [134], [135], [136], [137]].
Fig. 11.

Summary of the mechanisms of the pharmacological activity of vanadium. Anti-V, Anti-B, Anti-P, Anti-F, Anti-C, Anti-D, Cardio-P, Neuro-P, Anti−HC, and Anti-O: anti-viral, anti-bacterial, anti-parasitic, anti-fungal, anti-cancer, anti-diabetic, cardioprotective, neuroprotective, anti-hypercholesterolemic, and anti-obesity, respectively, CD4: CD4 receptor, CXCR-4: CXCR-4 chemokine co-receptor, RT: reverse transcriptase, Na+/K+-ATPase: sodium-potassium pump, ROS: reactive oxygen species, ERK: extracellular regulated kinase, MEK: ERK kinase activator, SAcP: acid phosphatase secreted by Leshmania, pRb: retinoblastoma protein, AS: antioxidant status, GSH: reduced glutathione, GSSG: disulfide glutathione, Top-IB: IB type topoisomerase, NEP: neutral endopeptidase, VO43−: vanadate ion, PO43−: phosphate ion, PTP: protein tyrosine phosphatase, Pi3K: phosphoinositide 3-kinase (phosphatidylinositol 3-kinase), Akt: protein kinase B (PKB), GLUT-4: glucose transporter type 4, PTK: tyrosine protein kinase, eNOS: endothelial nitric oxide synthase, NO: nitric oxide, FLIP: FLICE-inhibitory protein; Bim: Blc-2 interacting mediator of cell death; FasL: Fas ligand; breakD: breakdown; Ca: calcium; K: potassium, FKHR/FKHR1/AFX: class O members of the forkhead transcription factor family, PTP-1B: protein tyrosine phosphatase 1B; JAK2: Janus kinase 2; STAT3: signal transducer/activator of transcription 3; NPY: neuropeptide Y; PPARγ: peroxisome-activated receptor γ; C/EBPα: CCAAT-enhancer-binding protein α; LPL: lipoprotein lipase. ↓: decrease, ↑: increase, ↓: inhibition/inactivation, ↑: activation, ↑↑: hyperactivation, ⊣ blockade of expression,→: modulation. Based on available literature data cited in section 8.2 and other reports [[139], [140], [141], [142]].
8.2.1. Mechanisms of the pharmacological potential of vanadium
8.2.1.1. Mechanisms of the anti-bacterial activity of vanadium: a summarizing note
The mechanisms of antibacterial activity of V, which have not been fully elucidated yet, include: (a) inhibition of the activity of Na+/K+-ATPase (Fig. 11) [26,123,137,143], (b) generation of ROS (Fig. 11), (c) impact on transport of substrates (e.g. thymidine, uridine, leucine, glucose) into the cell through the bacterial cell membrane (Fig. 11) along with induction of potassium (K+) outflow from the cell [137], (d) interaction with topoisomerase type II (gyrase) (Fig. 11), which is necessary for the proper functioning of the genome and bacterial growth, with the ATP-binding site used as a target for antibacterial drugs [144,145], (e) interaction with DNA in an intercalative manner [133] resulting in modification of the DNA structure (unwinding, stiffening, and elongation of the double helix) with formation of an intercalative complex [146], (f) interaction with components of the cytoskeleton, which results in morphological alterations of bacterial cells thereby preventing proper division [137], and (g) other nonspecific mechanisms of action (Fig. 11), with emphasis on the better bioavailability of V complexes due to the presence of a specific ligand allowing the complex to penetrate the hydrophobic lipid-rich bacteria wall [acc. to 123, 124].
8.2.1.2. Mechanisms of the anti-viral activity of vanadium: a summarizing note
Most data in this area were obtained in in vitro studies of the influence of V on human immunodeficiency HIV-1/HIV-2 virus [33]. Some complexes of this element have been shown to inhibit the activity of viral reverse transcriptase (RT) [after33,123], which is responsible for the synthesis of DNA on the viral RNA matrix in the reverse transcription process facilitating integration of the viral DNA into the genome of the host cell. Thus, by inhibiting RT activity, vanadium blocks viral replication. Furthermore, the binding of V complexes to the CD4 molecule (present on the surface of T-helper cells) and the chemokine CXCR-4 co-receptor has been suggested as well (Fig. 11). This leads to blockage of the passage of the virus to the host cell [after 123], thereby preventing its multiplication and protecting against the development of infection.
8.2.1.3. Mechanisms of the anti-fungal activity of vanadium: a summarizing note
Among the mechanisms of the anti-fungal activity of V complexes which, likewise the mechanisms of the anti-bacterial and anti-viral activity of this metal have not been fully recognized yet, interactions with DNA by intercalation have been proposed [133]. Additionally, inhibition of biosynthesis of an important component of the fungal cell membrane, i.e. ergosterol [134] (Fig. 11), by affecting the expression of genes involved in the synthesis of this compound [135] have been suggested.
8.2.1.4. Mechanisms of the anti-parasitic activity of vanadium: a summarizing note
The mechanisms of the anti-parasitic activity of vanadium (against trypanosomiasis, amoebiasis, and leishmaniasis) mainly suggested in in vitro studies include: (a) intercalation of V complexes into DNA (Fig. 11) [after 123] (anti-tryposomal and anti-amoebiasis activity), (b) inhibition of the activity of phosphatases [125] (Fig. 11) due to the structural similarity of vanadate to phosphate [39,45]; these enzymes are involved in gluconeogenesis - a metabolic pathway necessary for the growth of Leishmania spp. [126], (c) inhibition of the activity of acid phosphatase (SAcP) (Fig. 11) [127], which is a virulence factor most commonly secreted by Leishmania during growth [128], and (d) activation of macrophage/Th1-type response with the release of ROS and pro-inflammatory cytokines (including IL-1β and IL-6) [129] capable of destroying intracellular pathogens [147] (activity against Leishmania).
8.2.1.5. Mechanisms of the anti-neoplastic activity of vanadium: a summarizing note
Among the mechanisms of the antitumor activity of V compounds/complexes, which still require further studies, intensified ROS generation (including iron-induced ROS following ferritin disintegration due to V action) is suggested [33] (Fig. 11). ROS production is commonly known to lead to disturbance in cellular metabolism and damage to lysosomes and mitochondria (critical for maintaining cellular homeostasis) [148], resulting in activation of caspases and apoptosis [123,149,150]. Inhibition of the cell cycle by hyperactivation of the Ras-Raf-MEK-ERK pathway is also proposed [after 123] (Fig. 11). This pathway activates inhibitors of cyclin-dependent kinases and thus maintains the retinoblastoma protein (pRb) in the hypophosphorylated form capable of blocking the cell cycle [151]. The involvement of V in suppressing neoplastic transformation through apoptotic signaling and/or cell cycle arrest has also been suggested by Chakraborty et al. [152] as based on evidence from animal studies. Moreover, focus is placed on: (a) interactions of V with the components of the spindle [123], (b) intercalation with DNA [33] (Fig. 11), (c) potentiation of anti-proliferative activity [136], (d) inhibition of the activity of IB topoisomerase (Top-IB) [153] (Fig. 11) capable of relaxing highly twisted DNA molecules [154], (e) inhibition of the activity of neutral endopeptidase (NEP) [155], which inactivates anticancer enkephalins [156], (f) modulation of phase I and/or II xenobiotic metabolizing enzymes [[157], [158], [159], [160]], and (g) alteration of antioxidant status (AS) [158,160,161] (Fig. 11). A reduction in the GSH/GSSG ratio [153] (Fig. 11), which can lead to intensification of oxidative stress in the cell [49], is highlighted as well. Additionally, some V compounds have been reported to be able to counteract tumor metastasis [155].
Much attention is still focused on studies of the design of V compounds for cancer treatment, as evidenced by recently published articles. However, all V compounds/complexes (used in these studies) were mainly tested in different cell lines, not in vivo conditions (in an animal model). For example, the results from a study conducted by Ni et al. [162] showed that certain multidentate oxovanadium(IV) complexes have promising anti-cancer activity against human HepG2 and SMMC-7721 hepatocellular carcinoma cells. One of these compounds exhibited much more potent anti-tumor properties and turned out to be less toxic to normal human cells than the cisplatin complex. It suppressed tumor cell proliferation by causing cell cycle arrest and directly induced apoptosis in a dose-dependent manner. As emphasized by the authors, structural elements (i.e. metal components, variations of the coordination mode, labile water molecules, and chelated ligands) probably exert an essential cooperative effect on the anti-tumor activity. Another in vitro study demonstrated that V as oxido-vanadium(IV) complex [VOL(bipy)] was able to induce early apoptosis more efficiently in the HepG2 cell line than in normal L929 cells [163]. Moreover, the authors revealed that the rates of necrosis/late apoptosis were also induced in the HepG2 cells more potently than in the L929 cells. Based on these results, they suggested that the VOL(bipy) complex can be considered as a new strategy for treatment of hepatocellular carcinoma. In addition, they hypothesized (on the basis of studies conducted by other researchers) that the VOL(bipy) complex may be modified by co-application with an antioxidant to be safer than the administration thereof alone. In turn, a study reported by Kowalski et al. [164] revealed that oxidovanadium(IV) coordination complexes containing a 2-methylnitrilotriacetate ligand are good candidates for preclinical development of novel anticancer drugs targeting pancreatic cancer. As suggested, the molecular mechanisms of cytotoxicity of these complexes were dependent on generation of ROS and cell cycle arrest in the G2/M phase with simultaneous activation of the p53/p21 pathway. An earlier study conducted by the same author [165] showed a selective cytotoxic effect of V complexes containing phenanthroline and quinoline as organic ligands against a human pancreatic ductal adenocarcinoma cell line (PANC-1). The results from this work revealed that V complexes caused cell cycle arrest in the G2/M phase and induced a significant increase in ROS generation in a time- and concentration dependent manner. In addition, at a higher concentration, V complexes induced a mixed type of cell death in PANC-1 cells, including apoptotic and necroptotic processes [165]. The authors emphasize that the type of cell death induced by V complexes is mainly determined by ligands and recommend further studies supporting the therapeutic potential of V in pancreatic cancer treatment. In turn, data from another research group [166], who examined the effect of a substituent in the hydrazone ligand of a family of μ-oxidodivanadium(v) hydrazone complexes on the structure, DNA binding, and anticancer activity, have revealed that V complexes exhibited promising anti-cancer activity against SiHa cervical cancer cells. Their experiments demonstrated that these complexes acted in an apoptotic mode and were non-toxic to the normal T293 cell line [166]. More recently, the anticancer properties of a family of vanadium(V) complexes with a diaminotris(phenolato) chelating ligand have been explored by Reytman et al. [167], who demonstrated their promising in vitro efficacy against HT-29, OVCAR-3, A2780, A2780cis, and A2780adr cells. In addition, a new promising effect of V compounds involving oncolytic viruses and their potential to combat cancer has recently been suggested. The authors reported that the infection of oncolityc viruses in resistant tumor cell lines was enhanced by V compounds [168]. As emphasized, this is of particular interest because it offers new opportunities for the development of strategies for the treatment of cancer.
8.2.1.6. Mechanisms of the anti-diabetic activity of vanadium with a historical outline of issues related to its anti-diabetic effects: a summarizing note
One of the mechanisms suggested for the insulin-like effect of V is linked to the inhibitory action of this element on the activity of protein tyrosine phosphatases (PTP) [123] (Fig. 11). This effect results from the antagonism related to the structural similarity of vanadate anions (VO4 3−) to phosphate anions (PO4 3−) [39,45]. By inhibition of protein tyrosine phosphatase 1B (PTP-1B) responsible for inactivation of the insulin receptor (INS-IR), which plays a key role in glucose metabolism, vanadium contributes to activation of the receptor presumably by preventing dephosphorylation of tyrosine phosphorylated residues of the beta subunit of INS-IR [169]. This in turn activates the PI3K-Akt pathway (3-phosphatidylinositol kinase (PI3-K)/Akt protein kinase) responsible for the metabolism of carbohydrates and lipids [2] and thus leads to potentiation of insulin-induced signal transduction. In consequence, the transport of glucose into cells increases [39] (Fig. 11). The inhibitory effect of ROS (generated by V compounds) on the activity of PTP-1B is also proposed [170] (Fig. 11). In addition, vanadium (a) causes an increase in the number of glucose-type 4 (GLUT-4) transporters in the cell membrane [171] (Fig. 11), which allows glucose molecules to be transported to adipocytes and muscle cells [172], (b) affects the insulin-like growth factor receptor (IGF-IR) by inactivation of associated PTP, (c) activates receptor and non-receptor protein tyrosine kinases (PTK), (d) stimulates glycogenogenesis, and (e) inhibits glycogenolysis and gluconeogenesis [172,173] (Fig. 11). It has also been highlighted that the interaction with membranes may be important in the stabilization of V complexes, and structural changes in membrane proteins may contribute to their insulin-mimetic mechanisms and toxicities [102].
As illustrated in Fig. 12 , the first report describing certain effects of V action in patients with diabetes mellitus (DM) appeared in 1899. Some 80 years later, Tolman and co-workers [175] demonstrated that V can directly influence glucose (GLU) metabolism and suggested that it may play a role in the regulation of GLU in vitro. Over the same period, other authors reported that V in the form of vanadate and vanadyl can stimulate GLU oxidation in rat adipocytes [176,177]. In turn, in 1985, Heyliger et al. [178] described the antidiabetic properties of V in vivo for the first time. Since then, the number of experiments (in an animal model) in which some V compounds (i.e. vanadyl sulfate - VOSO4, VS; sodium metavanadate - NaVO3, SMV; sodium orthovanadate - Na3VO4) were used as potential antidiabetic drugs has been constantly growing. For example, in 1987, Meyerovitch and co-investigators [179] reported that V is able to normalize the blood GLU level (in vivo model) (Fig. 12). Besides, other authors described the use of V in diabetes mellitus (DM) (Fig. 12). In 2000, one of the organic V compounds, i.e. bis(ethylmaltolato)oxovanadium(IV) (BEOV) first synthesized in the late 1990s [180] was used in the first phase I clinical trial in non-diabetic volunteers (Fig. 12). A few years later, in 2007/08, the same V compound was used in a phase IIa trial in seven type 2 diabetic subjects [180,181] (Fig. 12). However, in 2009, the diabetes program was ceased (Fig. 12). Akesis Pharmaceuticals, Inc. (in January 21, 2009) announced it has discontinued its sole clinical development program for AKP-020, a Phase IIa drug candidate for the treatment of diabetes mellitus [182]. As reported, the renal changes resulting from the doses used in the preclinical safety program were the cause of this decision [174] (Fig. 12). Vanadium is well-known to be nephrotoxic in vivo. Data on the effects of this metal on the animal kidney are available in the literature. For example, a study conducted by De la Tore et al. [183] showed that certain markers of nephrotoxicity such as the serum creatinine and urea levels as well as the urinary creatinine concentration increased and decreased, respectively after V administration, compared to the V-untreated animals. The creatinine clearance was also found to be lowered in the V-supplied rats [183]. Moreover, V-induced morphologic changes in the kidney and a significant influence of age on the renal effects of this metal were observed as well [183]. All those findings gave rise to the following statement: “the obtained results can be of concern if in the future, vanadium compounds can be administered in the treatment of diabetic patients” [183]. In addition, another study in rats demonstrated V-induced kidney fibrosis [184] at glomerular tuft, preglomeruli, pretubules, and interstitium (cortex and medulla) [185]. In turn, an earlier study conducted by Boscolo et al. [186] also showed alterations in the urinary excretion of certain electrolytes in rats chronically treated with V.
Fig. 12.

Summary of the most important aspects related to vanadium anti-diabetic effects on the timeline. V: vanadium, DM: diabetes mellitus, GLU: glucose, INS: insulin, BEOV: bis(ethylmaltolato)oxovanadium(IV). ↓: decrease. Based on available literature data cited in section 8.2.2.6 and other reports; **after [34], #after [174].
Vanadium compounds/complexes are generally known to exhibit INS-mimetic properties through which they are involved in the regulation of GLU metabolism. As illustrated on the timeline (Fig. 13 ), vanadyl sulfate (VS) was one of the inorganic V forms most often used in studies with diabetic patients. Overall, the history of these studies dates back to 1995 when Cohen and co-workers [187] conducted the first trials on the effects of VS in patients with non-insulin-dependent diabetes mellitus (NIDDM). In 1996, Halberstam et al. [188] and Boden et al. [189] also used VS in NIDDM patients (Fig. 13). In turn, sodium metavanadate (SMV) was administered in patients with NIDDM and in those with insulin-dependent diabetes mellitus (IDDM) by Goldfine et al. [190,191]. The researchers used VS in studies with NIDDM patients in 2000 [192] (Fig. 13). In the subsequent years, i.e. in 2001 [193], 2008 [194], and 2013 [195], VS was applied in studies with NIDDM, obese, and IDDM patients, respectively (Fig. 13).
Fig. 13.

Summary of trials with inorganic vanadium compounds used in diabetic patients on the timeline. V: vanadium, VS: vanadyl sulfate, SMV: sodium metavanadate, NIDDM: noninsulin-dependent diabetes mellitus, IDDM: insulin-dependent diabetes mellitus, Over-W: over-weight, wk: weeks, mo: months. Based on available literature data cited in section 8.2.2.6.
It has been estimated that the dose of V capable of normalizing the GLU level in the blood of rodents is about 100 mg V/kg/day (at the level of V in the blood within the range of 10–20 μM). The same effect in humans was achieved at a dose of 1.5 mg V/kg/day (at the level of V in the blood within the range of 1–5 μM) [30] (Fig. 14 ).
Fig. 14.

Vanadium doses in normalization of glucose in rodents and humans. V: vanadium, GLU: glucose. Based on available literature data cited in section 8.2.2.6.
To sum up, the highest scientific interest was devoted to studies on the potential use of vanadium in the treatment of diabetes, which is related to its antidiabetic properties. Positive results of administration of V compounds were demonstrated both in diabetic animals and in human patients. With regard to various animal models of diabetes, many reports in this research area have been published and there are ongoing studies focused on this issue, as evidenced by the works by Adam et al. [196] and Krośniak et al. [197]. Briefly, Adam’s research group focused on the synthesis of a new anti-diabetic complex containing vanadium(IV) and vitamin A. The authors showed a reduced blood glucose level, a lowered creatinine concentration, and a decrease in the activity of glutamate-pyruvate transaminase (GPT) in the serum of diabetic mice treated with a V(IV)-vitamin A complex, compared to the untreated diabetic mice group. In turn, data from mouse studies conducted by Krośniak et al. [197], in which the influence of eight new V compounds on organs mass were tested, revealed that V complexes had a remarkably different effect on organ weight despite the similar composition and the same co-ligand. As stressed by the authors, this may point to separate metabolic pathways of these compounds in the body and a role of tridentate L Schiff base ligands. More recently, a report has been published by Szklarzewicz et al. [198] showing studies that compared V complexes with diverse structures and correlated them with their properties. The investigators synthesized three new complexes of this metal (at three different oxidation levels; III, IV, and V) with Schiff base ligands showing differences in the coordination of the metal with the ligand in order to compare their physicochemical and spectroscopic properties as potential antidiabetic drugs.
As far as application of V in diabetic patients is concerned, this may be a considerable problem and may pose a risk of toxicity, as this element is highly toxic and is able to accumulate and interact with other metals. A summary of its serious side effects, including tissue V accumulation in experimental animals (in rats with diabetes) treated with this metal orally, was reviewed in the work by Domingo et al. [62]. Other reports by Domingo [48,199] provided a summary of side effects derived from the oral administration of V compounds observed in diabetic patients. As known, such diseases as diabetes mellitus or cancer are chronic diseases requiring chronic treatment; therefore, as emphasized by the author, the long-term/chronic administration of V may be unavoidable and can lead to long-term side effects resulting from significant tissue V accumulation. Thus, the toxicity of V that could derive from the chronic treatment with this element is of particular concern. Hence, researchers [174,199] question the legitimacy of the possible application of V in oral diabetes therapy (Fig. 13). Moreover, as stressed by some investigators [168], it also has to be kept in mind that diabetes mellitus is sometimes characterized as a metabolic disease with which patients live for many years; therefore, low toxicity of potential drugs should be of particular concern.
8.2.1.7. Mechanisms of the cardio-protective action of vanadium: a summarizing note
The results of previous studies on both organic and inorganic V compounds have shown that this element can act as a cardioprotective agent. It may protect the heart against ischemia-reperfusion injury, prevent hypertension and hypertrophy of myocardium, and improve heart performance [200]. Inhibition of PTP activity and activation of the 3-phosphatidylinositol kinase (PI3-K)/Akt protein kinase signaling pathway (PI3K-Akt) have been proposed as mechanisms responsible for V-induced cardioprotection [200] (Fig. 11). Akt, known as protein kinase B (PKB), is responsible for the phosphorylation of many different proteins involved in basic cellular processes such as proliferation, growth, migration, and metabolism [201] and plays an important role in the regulation of myocardial hypertrophy and in angiogenesis [acc. to 33]. Activation of the PI3K-Akt pathway results in the phosphorylation of endothelial nitric oxide synthase (eNOS) responsible for catalyzing nitric oxide (NO) production (Fig. 11). In turn, as an initiator and mediator of cardioprotection, NO has anti-hypertrophic activity and plays an important role in the regulation of vasorelaxation [202,203]. Diffusion of NO from the endothelium to vascular smooth muscle activates cyclic guanosine monophosphate (cGMP), which inhibits the influx of Ca2+ ions into the cell and/or the activity of the calcium pump (Ca2+-ATPase) and activates potassium channels leading to vasodilation [202]. Furthermore, stimulation of glucose transport by the influence of V on the GLUT-4 glucose transporters (Fig. 11), resulting in normalization of their level in myocardial cells [204], is another mechanism of the V cardioprotective action [200] proposed in studies conducted on rats with pharmacologically induced diabetes. In addition, vanadium (as vanadate, which shows pro-hypertensive properties) [186] can induce vascular smooth muscle contraction (acting as a vasoconstrictive agent) by increasing the intracellular calcium (Ca2+) concentration [205] (Fig. 11). In vivo studies carried out on smooth muscle (isolated from guinea pig's airways) [206] have shown that the inhibition of the activity of the sodium-potassium pump (Na+/K+-ATPase) by this element [143] does not seem to be linked to the mechanism responsible for smooth muscle contractility [206]. A vasoconstrictive effect has also been demonstrated for vanadyl, for which a different mechanism of action was suggested, compared to that proposed for vanadate [207]. In addition, vanadium as vanadyl may also have an anti-hypertensive effect, as demonstrated by studies conducted on spontaneously hypertensive rats (SHR) and in those with hypertension induced by a fructose-rich diet (FHR rats) receiving vanadyl sulfate (VOSO4) or bis(maltolato)oxavanadium(IV) (BMOV) [[208], [209], [210]].
8.2.1.8. Mechanisms of the neuro-protective action of vanadium: a summarizing note
In terms of mechanisms underlying the neuroprotective effects of V, it has been speculated that this element activates the PI3K-Akt signaling pathway (survival signals) by inhibiting PTP, thereby inactivating forkhead box class O (FOXOs) family members (such as FKHR, FKHRL1, AFX) and finally resulting in suppression of expression of apoptosis-inducing factors such as Bim and Fas ligands (FasL). It has also been assumed to activate extracellular signal-regulated kinase (ERK) [[211], [212], [213], [214], [215]] (Fig. 11)
8.2.1.9. Involvement of vanadium in the mechanisms of appetite regulation in terms of anti-obesity effects: a summarizing note
In vivo studies in a rodent model (on rats) have shown that V administered in the form of BMOV leads to an increase in the intake of V and a decrease in the water and food intake, body weight, and serum leptin (LEP) concentration (LEPS) [216] (Fig. 15 ). The influence of this metal on the signaling pathway through LEP (JAK2/STAT3 pathway) [217,218] was suggested as a mechanism underlying these changes [216] (Fig. 15).
Fig. 15.

Mechanism of vanadium-induced weight loss – in vivo studies. V: vanadium, LEP: leptin, LEPS: the concentration of leptin in the serum, LEP-R: leptin resistance, Hyper-LEP: hyperleptynemia, LEP-S: leptin sensitivity, INS: insulin, INS-R: insulin resistance, INS-S: insulin sensitivity, NPY: neuropeptide Y, PTP-1B: protein tyrosine phosphatase 1B, JAK2: Janus kinase 2, STAT3: signal transducer/transcription factor, IRS: insulin receptor tyrosine kinase substrate ↓: decrease, ↑: increase, ↓: activity/pathway blockade, ↑: activation, ↓: signal blockade. Based on available literature data cited in section 8.2.2.8.
It is believed that the increase in the activity of PTP-1B blocks Janus tyrosine kinase 2 (JAK2), i.e. a substrate for PTP-1B [219,220], resulting in blockage of leptin signaling (JAK2/STAT3) and cascades of phosphatidylinositol 3-kinase (PI3-K) involved in the signaling pathway of both insulin (INS) and leptin (LEP) [220,221] (Fig. 15). Therefore, the signal from INS and LEP is blocked (Fig. 15). In turn, by inhibition of the activity of PTP-1B [222], vanadium leads to activation of the JAK2/STAT3 leptin signaling pathway [217,218] (Fig. 15) and thus inhibits the synthesis of neuropeptide Y (NPY) and its release in the hypothalamus. Consequently, appetite, body fat mass, and body weight are reduced [221,223] (Fig. 15). There is also an increase in leptin (LEP-S) and insulin (INS-S) sensitivity and a decrease in insulin (INS-R) and leptin (LEP-R) resistance [138,218,220,224] (Fig. 15). INS-R (which causes disturbances in the synthesis of lipoproteins and their changes in blood plasma) [225] along with LEP-R and LEP-R-related hyperleptinemia (Hyper-LEP) have been reported to be significant risk factors of hypertension, type II diabetes, dyslipidemia, atherosclerosis, and cardiovascular diseases [138,226,227] (Fig. 15). Thus, by affecting the activity of PTP-1B, V induces the JAK2/STAT3 signaling pathway, which improves sensitivity to LEP and INS and makes this element (as suggested) useful for treating obesity [138,217,218]. An increase in INS-S was found in diabetic rats receiving vanadyl sulfate (VOSO4) for 30 days [228] and in type I and II diabetes patients after administration of sodium metavanadate (NaVO3) for 2 weeks [190].
Noteworthy is also the fact that, through its direct influence on the organization of membrane lipids and cholesterol/sphingolipid-rich microdomains (where proteins involved in signal transduction are concentrated, e.g. the INS receptor), V enhances the effects caused by INS and, consequently, results in INS-R reduction [229]. A decrease in INS-R was demonstrated in obese rats (fa/fa Zucker rats) receiving V in the form of an organic derivative BMOV for 6 weeks [230] and in hypertensive rats chronically treated with VOSO4 [209]. Both V compounds were administered in drinking water.
8.2.1.10. Mechanisms of the anti-hypercholesterolemic activity of vanadium: a summarizing note
The mechanism of the anti-hypercholesterolemic action of V is related to the effect of this element on the cholesterol biosynthesis pathway shown in Fig. 16 . In vitro studies have demonstrated that V can inhibit the synthesis of this steroid [231] by blocking the utilization of mevalonate [232,233], which is formed from 3-hydroxy-3-methyl-glutaryl-coenzyme A (3-HMG-CoA) in a reaction catalyzed by HMG-CoA reductase and further transformed into isopentenyl-5-pyrophosphate (IPP) in phosphorylation/decarboxylation reactions [234] (Fig. 16I). An increase in the catabolism of cholesterol by V is another possible mechanism of action of this element [235] (Fig. 16I).
Fig. 16.

Cholesterol biosynthesis pathway with probable sites of vanadium action (I) along with changes in the lipid profile in humans occupationally exposed to vanadium (II) and in diabetic patients receiving inorganic vanadium salts (III). CoA: coenzyme A, 3-HMG-CoA: 3-hydroxy-3-methyl-glutaryl-CoA, IPP: isopentenyl-5-pyrophosphate, GPP: geranyl-pyrophosphate, FPP: farnesyl-pyrophosphate, C: cholesterol, HDL: high-density lipoproteins, LDL: low-density lipoproteins, HDL-C: HDL cholesterol, ApoA-I: apolipoprotein A, TC: total cholesterol, LDL-C: LDL cholesterol, ApoB: apolipoprotein B, VOSO4: vanadyl sulfate, NaVO3: sodium metavanadate. Based on available literature data cited in section 8.2.2.9.
The impact of V on stimulation of the lipoprotein lipase (LPL) activity has been highlighted as well [29,236]. The LPL enzyme controls many metabolic processes, e.g. it participates in the metabolism of lipids and lipoproteins [237]. Thus, any disorders in its proper functioning (during INS-R) [211] result in development of atherosclerosis, obesity, and type II diabetes [237].
Unfavorable changes have been found in the lipid profile of individuals that are occupationally exposed to V (working in production of V-containing steel) (Fig. 16II). There was an increase in the levels of high-density lipoprotein cholesterol (HDL-C) and apolipoprotein A (Apo-AI, main HDL apolipoprotein) in the serum and a decrease in such atherogenic indices as TC/HDL-C, LDL-C/HDL-C, and ApoB/ApoA-I [238] (Fig. 16II). A decrease in cholesterol and/or low-density lipoprotein cholesterol (LDL-C) have also been found in the serum of type II diabetes patients after 6 weeks of administration of VOSO4 [192,193] and in type I and II diabetes mellitus patients after 2-week treatment with sodium metavanadate (NaVO3) [191] (Fig. 16III).
A reduced cholesterol level has been demonstrated in the blood of healthy rats receiving V2O5 (0.56 mg V2O5/kg) once a month for 12 months [78], in obese rats (fa/fa Zucker rats) treated with sodium orthovanadate (Na3VO4, 0.8 mg/mL) in drinking water for 4 months [239], and in diabetic rats receiving VOSO4 (0.75 mg/mL, 1 mg/mL, 1.1 mg/mL or 0.5 mmol/kg/day) [[240], [241], [242], [243]], sodium metavanadate (NaVO3, 0.20 mg/mL), sodium orthovanadate (Na3VO4, 0.50 mg/mL) [241], or organic V complexes such as V5dipic-NH2 (0.1 mg/mL) [244] or bis(curcumino)oxavanadyl (BCOV, 0.05, 0.1 or 0.2 mmol/kg/day) [243] in drinking water for different periods (20 days, 2, 4, and 10 weeks, or 5 months). In turn, a decrease in the level of triglycerides (TG) in the blood was found: (a) in obese rats (fa/fa Zucker rats) receiving Na3VO4 (0.8 mg/mL) in drinking water for 4 months [239], (b) in obese non-diabetic Zucker fatty rats (ZF) and diabetic Zucker fatty rats (ZDF) after administration of organic (maltolic) vanadium derivatives, i.e. bis(maltolato)oxovanadium(IV) (BMOV, 0.19 mmol/kg/day, in drinking water) and bis(ethylmaltolato)oxovanadium(IV) (BEOV, 0.1 mmol/kg/day, with a probe), respectively, for 3 weeks [245], and (c) in diabetic rats receiving VOSO4 [240,[242], [243], [244]], V5dipic-NH2 [244], or BCOV [243]. BCOV has also been shown to reduce the serum LDL level significantly in these animals [243].
9. Vanadium - important research trends
9.1. Research on non-elucidated mechanisms
Given the increasing practical application of V in medicine and the fact that accumulation of this metal in certain internal organs/tissues during the treatment of chronic diseases may be a significant problem, it is still advisable to carry out studies on V toxicity and mechanisms of its toxic action (Fig. 17 ). It is also necessary to study the pharmacological activity of V as well as the mechanisms of absorption and excretion thereof. Moreover, the mechanisms of transport of V to target organs and its uptake by cells as well as the in vivo form of this metal and its influence on the immune system deserve further exploration (Fig. 17). The mechanisms of immune response upon V exposure should also be better clarified. Additionally, comprehensive research on the interactions of V with other elements is needed as well (Fig. 17).
Fig. 17.

Vanadium-associated issues – deserving more in-depth investigations.
9.2. Research on the interactions with elements having antioxidant potential: magnesium
Metal interactions affecting the cell metabolism, action of organs and, consequently, the function of the entire organism is still an important issue in toxicology, pharmacology, and medicine. Therefore, examination of the consequences, character, and mechanisms of interactions of V (which raises hopes for the use thereof in the treatment of certain diseases in humans) with elements, especially those with antioxidant potential, in an in vivo experimental model are still a current and vital issue. Moreover, considering the well-known pro-oxidant properties of V, the fact that the increased generation of ROS and oxidative stress play a crucial role in V-induced toxicity, and the suggestions of other authors that the hepatotoxic, nephrotoxic, cardiotoxic, genotoxic, neurotoxic, and carcinogenic effects of this metal as well as inflammatory and fibrotic changes in the lungs are associated with its redox-active nature and ability to intensify LPO (see Fig. 7), detailed investigations to evaluate the consequences of possible interactions of V with antioxidant elements during combined chronic treatment are even more desirable. This aspect is also important in the context of occupational exposure and growing environmental pollution with V.
The results of studies conducted in a rodent model by Ścibior et al. [57] showed protective action of Mg against the pro-oxidant activity of V. They also provided evidence that the beneficial Mg-induced limitation of the increase in the hepatic LPO during the administration of V may result from the independent action of Mg and from the antagonistic interaction with V. These effects were observed when healthy rats were exposed to V (as SMV) at the concentration of 0.125 mg V/L with supplementation with Mg (as MS at the concentration of 0.06 mg Mg/mL) for 18 weeks. However, further studies are needed to explain the exact mechanism(s) accounting for the protective effect of Mg (at the dose used) against the V-induced oxidative stress in the liver. The other findings reported by Ścibior et al. [52] demonstrated a markedly reduced level of LPO in the bone of healthy rats supplemented with Mg (as MS) during the 12-week SMV exposure (0.125 mg V/mL). These changes, as those mentioned previously for the liver, were also influenced by the independent action of Mg and by its antagonistic interaction with V. Moreover, the distinct trend toward the antagonistic interaction between V and Mg was revealed with regard to the level of LPO in the erythrocytes of healthy rats exposed to SMV (0.125 mg V/mL) during 12-week MS supplementation (0.06 mg Mg/mL) [53]. It should be added that the concentration of V mentioned above was selected based on earlier studies conducted by other researchers [[246], [247], [248], [249], [250]] who tested similar V concentrations in a rat model as well as reports of the levels of this metal in the blood and urine of occupationally exposed people [251,252], which were comparable with those noted in SMV-exposed rats. Based on the results obtained, it can be concluded that the issues related to the interactions of V with Mg (as one of the elements with antioxidant potential) are important not only for extending the knowledge of the mechanism of the effect of V on the organism. They are also essential to improve the understanding of the role of Mg in prevention of V toxicity and clarify the mechanisms underlying the potential protective action of this bioelement in V poisoning. This is even more relevant in view of the fact that long-lasting oxidative stress with simultaneous weakening of the antioxidant system can lead to permanent cell/tissue damage and, consequently, to initiation of the disease process. In addition, the examination of the combined V-Mg effects in mammalian organisms may also be helpful for the future safe use of V in medicinal applications. Undisputed is the fact that recognition of doses of elements showing synergistic/antagonistic action towards each other may play a role in both medicine and pharmacology. However, although some studies cited above reveal that supplementation with Mg during V exposure has beneficial effects reflected in reduction of the level of LPO in certain cells/tissues of healthy rats, further studies (in other animal models) have to be conducted to recognize whether the antagonistic effects of Mg on pharmacological amounts of V will allow V to exert beneficial effects, or whether their synergistic actions yield beneficial effects at lower amounts of V. Some time ago, Matsuda et al. [253] demonstrated for the first time that V and Mg administered in combination as SMV (0.3 mg/mL) and MS (0.3 mg/mL), respectively, to diabetic rats in drinking water for 3 weeks had a synergistic effect, as they augmented whole-body insulin sensitivity and glycogen synthesis. Based on these findings, the authors suggested a potential role for combination therapy with trace elements in type 2 diabetes mellitus and other insulin-resistant states. Diabetes is accompanied by changes in the level of Mg in tissues [254] and by weakening of the antioxidant barrier [255] and oxidative stress [256]. With regard to this issue, Sanchez et al. [94] showed that treatment with V of Mg-deficient rats corrected many alterations generated by Mg deficiency. Subsequently, Bermúdez-Peña et al. [257] evaluated whether V treatment might affect alterations in Mg metabolism associated with diabetes. The authors revealed the existence of interplay or other interactions between both elements in diabetic rats. As they stressed, these results might help to clarify the role of V as an anti-diabetic agent.
The existing literature comprises a few articles about the interactions between V and Mg [[51], [52], [53],57,63,94,253,[257], [258], [259], [260]], which has been reported to be able to protect against the harmful effects of ROS [[261], [262], [263], [264]]. A summary of data about the interactions between both metals (demonstrated to date in relation to parameters investigated in a rodent model) is presented in Fig. 18 .
Fig. 18.

Summary of vanadium-magnesium interactions – in vivo model: our studies (I) and studies of other authors (II). V: vanadium; Mg: magnesium, Fe: iron, Cu: copper: P: phosphorus, RBC: erythrocytes, GR: glutathione reductase, LPO: lipid peroxidation, CH: cerebral hemisphere, B: bone, WB: whole blood, L: liver, K: kidney, P: plasma, TS: transferrin saturation, L-AA: L-ascorbic acid, ALP: alkaline phosphatase, SOD: superoxide dismutase, Sa: mean roughness, Sq: root mean square roughness, Sz: ten-point height, Ins: insulin, HOMA-IR: insulin resistance index. Based on available literature cited in section 9.2.
Antagonistic interactions between V and Mg have been demonstrated for (a) the activity of glutathione reductase in erythrocytes (GRRBC) and the concentration of vanadium (VRBC), iron (FeRBC), and copper (CuRBC) in erythrocytes, (b) the level of lipid peroxidation in the liver (LPOL) and bone (LPOB), and (c) the concentration of vanadium in the kidney (VK) and cerebral hemisphere (VCH). In turn, a clear trend towards the V-Mg antagonistic interaction has been observed for (a) the concentration of magnesium in erythrocytes (MgRBC) and plasma (MgP), (b) the level of lipid peroxidation in erythrocytes (LPORBC), (c) the concentration of zinc (ZnB) and potassium (KB) in bone, (d) the level of iron in the kidney (FeK), (e) the concentration of V in whole blood (VWB), and (f) iron transferrin (Tf) saturation (TS) (Fig. 18I).
In contrast, synergistic interactions between V and Mg have been revealed for (a) the concentration of L-ascorbic acid in bone (L-AAB), (b) the activity of alkaline phosphatase (ALPB) and superoxide dismutase (SODB) in bone, (c) the concentration of calcium in erythrocytes (CaRBC) and in the liver (CaL). Synergy has also been determined for such roughness parameters as (d) mean roughness (Sa) and (e) root mean square roughness (Sq) (Fig. 18I), which were measured by optical profilometry in a three-dimensional (3D) scale in morphological studies of the femur of rats receiving V and Mg separately and in combination [260]. In turn, a trend towards synergistic interactions between both metals has been observed for (a) the level of sodium in erythrocytes (NaRBC) and bone (NaB), (b) the concentration of calcium (CaK) in the kidney, and (c) the ten-point height (Sz) defined as the arithmetic average height of the sum of five local maxima and five local minima (Fig. 18I).
The interactive effects between V and Mg (studied in an animal model) have also been revealed for such parameters as V intake, V absorption, urinary and fecal V excretion, serum Mg and insulin concentration, and insulin resistance index (HOMA-IR) (Fig. 18II).
As far as the historical outline of interactions is concerned, in 1970, Hill and Matrone proposed the theory of mineral interactions (Fig. 19 ) suggesting that metals with similar chemical and physical properties would interact with each other biologically [after 265]. In 1982, Georgievskii described synergistic and antagonistic interactions between certain elements, whereas O’ Del defined the term “interaction” in 1997 [after 265] (Fig. 19).
Fig. 19.

Historical outline of certain issues linked to mineral interactions and studies on the effects of combined administration of vanadium and elements with antioxidant properties to animals (mainly to rats). Mg: magnesium, Zn: zinc, Se: selenium, Cr: chromium. *after [265].
Studies on the influence of combined administration of V and elements with antioxidant potential date back to 1966 when Berg [258] described the effect of co-administration of V with Mg on the growth and mortality in chicks (Fig. 19). In 1989, Yamaguchi et al. [266] reported that Zn prevented the toxic effect of V (Fig. 19). In the late 1990s, two reports described the effect of Se on V toxicity in different regions of the rat brain [267] and a synergistic interaction between V and Mg on glucose metabolism in diabetic rats [253]. Additionally, the influence of combined administration of V with Zn or Se to rats [268] has been described (Fig. 19). The research on the interactions of V with antioxidant elements has been gaining popularity since 2005. Since that time, more reports have appeared about the effects of co-application of V with Cr(III) [59,[269], [270], [271], [272]] or Mg [[51], [52], [53], [54], [55], [56], [57], [58],63,259,260] to rats (Fig. 19). Moreover, there have also been reports on the effect of V administered as BMOV on bioavailability, biochemical parameters in serum, excretion and content of V and Mg in certain internal organs/tissues of healthy rats receiving V-containing feed and varying levels of Mg (magnesium oxide, MgO) [94], and the effect of V (BMOV) on the metabolism and distribution of Mg and some biochemical blood parameters in diabetic rats receiving feed containing V and Mg (MgO) [257].
It should also be added that more attention should be given to studies on treatment with V in association with compounds with antioxidant properties to optimize the therapeutic potential of V compounds and minimize their possible side effects. This is even more relevant in view of the work conducted by Wang et al. [273], who showed that the treatment with N-acetylcysteine (NAC), i.e. a thiol antioxidant reducing the accumulation of ROS and oxidative damage [274], inhibited V-induced ROS generation in human normal liver L02 cells but did not attenuate the antitumor activity of V in human hepatoma HepG2 cells. As emphasized by the authors, these results may be helpful in the therapeutic application of V compounds along with antioxidants as synergistic agents in order to reduce their potential toxicities in normal cells without affecting their antitumor activities in cancer cells. In addition, the same researchers suggest that combined administration of V with an antioxidant might be beneficial for diabetes therapy [273]. Recently, protective action of pyruvate against V-induced oxidative stress and cytotoxicity has been demonstrated in Chinese hamster ovary (CHO-K1) cells [275]. It has been suggested that the antioxidative effects of pyruvate, especially its ability to neutralize hydrogen peroxide, may be involved in the observed mechanism of protection.
To sum up, comprehensive investigations in an animal model on the interactions of V with antioxidant elements, such as Mg or certain compounds with antioxidant properties or dietary/plant-derived antioxidants, are particularly important. They would allow us not only to recognize a potential antidote minimizing the adverse effects of V resulting from its strong pro-oxidative activity but also to evaluate the potential efficacy of combined V-Mg treatment in certain modern-age diseases.
10. A brief summary and perspectives
Due to its multidirectional biological activity and a wide range of effects on the mammalian organism, vanadium has been arousing interest of many research centers worldwide for many years. The use of this metal in medicine is of particular concern. Some V compounds/complexes have been found to be promising for potential therapeutic use. Their beneficial therapeutic properties have been reported and diseases of interest against which they could be effective have been suggested. The number of studies on the use of V in medicine is constantly growing and its potential medical application remains an open question. Given the recent progress in the research on the pharmacological activity of different V compounds/complexes, it can be assumed that new V-based drugs will be available for therapeutic purposes in the future. Recently, much attention has been focused on the design of V compounds that can be useful for potential applications as anticancer agents. There are also ongoing studies on different anti-diabetic V complexes for their future safe use in the treatment of diabetes. On the other hand, however, many multidirectional studies on V (which has a narrow therapeutic index) have shown that further analyses are still required for this element to be used as a metallodrug in the fight against certain life-threatening modern-age diseases. It has been highlighted that the ability of V to accumulate and its strong toxicological potential are an important obstacle limiting the use of this metal in pharmacology. Therefore, it would be advisable to shift the balance towards the beneficial action of V, whose pharmacological potential has repeatedly been revealed, as evidenced by the results of many studies conducted in a variety of different cell lines and in animal models. However, this is a serious challenge and considerable research is still devoted to design V complexes with low toxicity and sufficiently high efficacy.
Besides the issues mentioned above and those presented in section 9 titled ‘Vanadium - important research trends’ and the need to identify the susceptibility of different cells/tissues to the effects of V, the following aspects draw researchers' attention: (a) identification of V species as an important factor in the assessment of toxicity of this element and its potential health risks to humans [276] (Fig. 20 ), (b) examination of the well-defined forms of this metal in terms of their availability, selectivity, and specificity [277] with recognition of key factors that may affect the mode of V action (Fig. 20), (c) biotransformation of V compounds in the organism and distribution between blood bioligands as a relevant issue of the drug metabolism and mechanism of action [103,109], (d) the role of the carrier ligand that may not be limited only to facilitating the transport/absorption of V compounds [278] with establishing the toxicity profiles of the ligand [167], (e) elucidation of the role of V species in interactions with immune system modulators and other transcription factors influencing immune signaling [277], and (f) recognition of therapeutic targets, pharmacokinetics, and pharmacodynamics in detail, which may help to design better and more effective V-based drugs [101].
Fig. 20.

Key factors affecting the mode of V action.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
The indices presented in the current report in Fig. 18I were determined using some laboratory equipment included in the infrastructure of the Centre for Interdisciplinary Research of the John Paul II Catholic University of Lublin (Lublin, Poland). It was co-funded by the European Union from the European Regional Development Fund in the frame of the Operational Programme Development of Eastern Poland 2007-2013 (POPW.01.03.00-06-003/09-00).
Contributor Information
Agnieszka Ścibior, Email: cellbiol@kul.lublin.pl.
Łukasz Pietrzyk, Email: lukasz.pietrzyk@wp.pl.
Zbigniew Plewa, Email: plewa_z@o2.pl.
Andrzej Skiba, Email: a.ski@vp.pl.
References
- 1.Nriagu J.O. Part 2: Health Effects. John Wiley and Sons; New York: 1998. Vanadium in the environment. [Google Scholar]
- 2.Tracey A.S., Willsky G.R., Takeuchi E.S. CRC Press, Taylor and Francis Group; 2007. Vanadium: Chemistry, Biochemistry, Pharmacological, and Practical Applications. [Google Scholar]
- 3.Imtiaz M., Rizwan M.S., Xiong S., Li H., Ashraf M., Shahzad S.M., Shahzad M., Rizwan M., Tu S. Vanadium, recent advancements and research prospect: a review. Environ. Int. 2015;80:79–88. doi: 10.1016/j.envint.2015.03.018. [DOI] [PubMed] [Google Scholar]
- 4.Zaporowska H., Ścibior A. Hematological effects of vanadium on living organisms. In: Nriagu J.O., editor. Vol. 31. John Wiley and Sons; New York, Chichester, Weinheim, Brisbane, Singapore, Toronto: 1998. pp. 135–157. (Vanadium in the Environment. Part 2: Health Effects). [Google Scholar]
- 5.Evangelou A.M. Vanadium in cancer treatment. Cri. Rev. Oncol. Hematol. 2002;42:249–265. doi: 10.1016/S1040-8428(01)00221-9. [DOI] [PubMed] [Google Scholar]
- 6.Rehder D. Implications of vanadium in technical applications and pharmaceutical issues. Inorg. Chim. Acta Rev. 2017;455:378–389. doi: 10.1016/j.ica.2016.06.021. [DOI] [Google Scholar]
- 7.Eichstaedt I. Wiedza Powszechna; Warszawa: 1970. Księga Pierwiastków. [Google Scholar]
- 8.Barceloux D.G. Vanadium. J. Toxicol. Clin. Toxicol. 1999;37:265–278. doi: 10.1081/clt-100102425. [DOI] [PubMed] [Google Scholar]
- 9.2001. WHO: Vanadium Pentoxide and Other Inorganic Vanadium Compounds. Geneva. [Google Scholar]
- 10.Fortoul T.I., Rojas-Lemus M. Vanadium as an air pollutant. In: Fortoul T.I., Avila-Costa M.R., editors. Vanadium: Its Impact on Health. Nova Science Publishers; New York: 2007. pp. 1–6. Chapter 1. [Google Scholar]
- 11.Davis J.R. ASM International; 2003. Handbook of Materials for Medical Devices. [Google Scholar]
- 12.Melechow R., Tubielewicz K., Błaszczuk W. Wyd. Politechniki Częstochowskiej; Częstochowa: 2004. Tytan I Jego Stopy. [Google Scholar]
- 13.Veiga C., Davim J.P., Loureiro A.J.R. Properties and applications of titanium alloys: a brief review. Rev. Adv. Mater. Sci. 2012;32:133–148. [Google Scholar]
- 14.Dudek Ł., Hryniewicz T., Rokosz K. Zastosowanie tytanu i wybranych stopów tytanu w lotnictwie. Autobusy: technika, eksploatacja, systemy transportowe. 2016;17:62–66. [Google Scholar]
- 15.Vasudevan D., Balashanmugam P. Study of thermal behavior on titanium alloys (Ti-6Al-4V) J. Eng. Sci. Technol. 2017;12:2064–2077. [Google Scholar]
- 16.Antunes R.A., Salvador C.A.F., Lopes de Oliveira M.C. Materials selection of optimized titanium alloys for aircraft applications. Mat. Res. 2018;21 doi: 10.1590/1980-5373-mr-2017-0979. [DOI] [Google Scholar]
- 17.Riffin P.V. Watertown Arsenal Laboratory; 1945. Metallurgical Examination of a 3-1/4 Thick Armor Plate From a German PzKw V (Panther) Tank; pp. 1–4. Report No. WAL. 710/715, January. [Google Scholar]
- 18.Yoffa W., Hurlich A. Watertown Arsenal Laboratory; 1945. Metallurgical Examination of Armor and Welded Joints From the Side of a German PzKw V (Panther) Tank; pp. 1–9. Report No. WAL. 710/750, May. [Google Scholar]
- 19.Cady J.R., Gillett H.W., Grenell L.H. Vol. 47. Metal Progress; 1945. pp. 289–320. (Enemy Material from the Metallurgical Point of View (Report to War Metallurgy Committee)). No. 2, February. [Google Scholar]
- 20.Donachie M.J. second ed. ASM international; 2000. Titanium: A Technical Guide. [Google Scholar]
- 21.Harland B.F., Harden-Williams B.A. Is vanadium of human nutritional importance yet? J. Am. Diet. Assoc. 1994;94:891–894. doi: 10.1016/0002-8223(94)92371-X. [DOI] [PubMed] [Google Scholar]
- 22.Sekar N., Kanthasamy A., William S., Balasubramaniyan N., Govindasamy S. Antioxidant effect of vanadate on experimental diabetic rats. Acta Diabetol. Lat. 1990;27:285–293. doi: 10.1007/bf02580932. [DOI] [PubMed] [Google Scholar]
- 23.Yanardag R., Bolkent S., Karabulut-Bulan Ö, Tunali S. Effects of vanadyl sulfate on kidney in experimental diabetes. Biol. Trace Elem. 2003;95:73–85. doi: 10.1385/BTER:95:1:73. [DOI] [PubMed] [Google Scholar]
- 24.Tunali S., Yanardag R. Effect of vanadyl sulfate on the status of lipid parameters and on stomach and spleen tissues of streptozotocin-induced diabetic rats. Pharmacol. Res. 2006;53:271–277. doi: 10.1016/j.phrs.2005.12.004. [DOI] [PubMed] [Google Scholar]
- 25.Soares S.S., Gutiérrez-Merino C., Aureliano M. Mitochondria as a target for decavanadate toxicity in Sparus aurata heart. Aquat. Toxicol. 2007;83:1–9. doi: 10.1016/j.aquatox.2007.03.005. [DOI] [PubMed] [Google Scholar]
- 26.Pessoa J.C., Garribba E., Santos M.F.A., Santos-Silva T. Vanadium and proteins: uptake, transport, structure, activity and function. Coord. Chem. Rev. 2015;301-302:49–86. doi: 10.1016/j.ccr.2015.03.016. [DOI] [Google Scholar]
- 27.Zaporowska H., Ścibior A. Vanadium and its significance in animal cell metabolism. In: Nriagu J.O., editor. Vol. 31. John Wiley and Sons; New York, Chichester, Weinheim, Brisbane, Singapore, Toronto: 1998. pp. 121–133. (Vanadium in the Environment. Part 2: Health Effects). [Google Scholar]
- 28.Ramasarma T. A redox profile of vanadium. In: Aureliano M., editor. Vanadium Biochemistry, Research Signpost. 2007. pp. 45–74. [Google Scholar]
- 29.Das S., Chatterjee M., Janarthan M., Ramachandran H., Chatterjee M. Vanadium in cancer prevention. In: Michibata H., editor. Vanadium: Biochemical and Molecular Biological Approaches. Springer; 2012. pp. 163–185. Chapter 8. [Google Scholar]
- 30.Goc A. Biological activity of vanadium compounds. Cent. Eur. J. Biol. 2006;1:314–332. , doi: 0.2478/s11535-006-0029-z. [Google Scholar]
- 31.Mussali-Galante P., Rodriguez-Lara V., Avila-Costa M.R., Fortoul T.I. Vanadium as an air pollutant. In: Fortoul T.I., Avila-Costa M.R., editors. Vanadium: Its Impact on Health. Nova Science Publishers; New York: 2007. pp. 7–20. Chapter 2. [Google Scholar]
- 32.Etcheverry S.B., Di Virgilio A.L., Barrio D.A. Vanadium effects on bone metabolism. In: Michibata H., editor. Vanadium: Biochemical and Molecular Biological Approaches. Springer; 2012. pp. 145–162. Chapter 7. [Google Scholar]
- 33.Rehder D. Vanadium. Its role in humans. In: Sigel A., Sigel H., Sigel R.K., editors. Interrelations between Essential Metal Ions and Human Diseases. Springer; Dordrecht, Heidelberg, New York, London: 2013. pp. 139–167. [Google Scholar]
- 34.Gruzewska K., Michno A., Pawelczyk T., Bielarczyk H. Essentiality and toxicity of vanadium supplements in health and pathology. J. Physiol. Pharmacol. 2014;65:603–611. [PubMed] [Google Scholar]
- 35.Anke M., Illing-Günther H., Anke S., Müller R. Vanadium – an essential and toxic element? In: Ermidou- Pollet S., Pollet S., editors. Trace Element in Human: New Perspectives, Proceedings Book. 2013. pp. 1120–1153. Part II, Entypossis, Athens. [Google Scholar]
- 36.Rehder D. The potentiality of vanadium in medicinal applications. Future Med. Chem. 2012;4:1823–1837. doi: 10.4155/fmc.12.103. [DOI] [PubMed] [Google Scholar]
- 37.Rehder D. The role of vanadium in biology. Metallomics. 2015;7:730–742. doi: 10.1039/c4mt00304g. [DOI] [PubMed] [Google Scholar]
- 38.Nielsen F.H., Uthus E.O. The essentiality and metabolism of vanadium. In: Chasteen N., editor. Vanadium in Biological System. Kluwer Academic Press; Dordrecht: 1990. pp. 51–62. [Google Scholar]
- 39.Rehder D. John Wiley and Sons; Chichester, UK: 2008. Bioinorganic Vanadium Chemistry. [Google Scholar]
- 40.Michibata H. Springer; Dordrecht, Heidelberg, London, New York: 2012. Vanadium. Biochemical and Molecular Biological Approaches. [Google Scholar]
- 41.McDowell L.R. first ed. Design Publishing Inc.; 2017. Mineral History. The Early Years. [Google Scholar]
- 42.Vilter H. Peroxidases from phaeophyceae: a vanadium(V)-dependent peroxidase from Ascophyllum nodosum. Phytochemistry. 1984;23:1387–1390. doi: 10.1016/S0031-9422(00)80471-9. [DOI] [Google Scholar]
- 43.De Boer E., van Kooyk Y., Tromp M.G.M., Plat H., Wever R. Bromoperoxidase from Ascophyllum nodosum: a novel class of enzymes containing vanadium as a prosthetic group? Biochim. Biophys. Acta – Protein Structure Molecular Enzymology. 1986;869:48–53. doi: 10.1016/0167-4838(86)90308-0. [DOI] [Google Scholar]
- 44.Robson R.L., Eady R.R., Richardson T.H., Miller R.W., Jawkins M., Postgate J.R. The alternative nitrogenase of Azotobacter chroococcum is a vanadium enzyme. Nature. 1986;322:388–391. doi: 10.1038/322388a0. [DOI] [Google Scholar]
- 45.Rehder D. Vanadium in health issues. ChemTexts. 2018;4:20. doi: 10.1007/s40828-018-0074-z. [DOI] [Google Scholar]
- 46.French R.J., Jones P.J.H. Role of vanadium in nutrition: metabolism, essentiality and dietary consideration. Life Sci. 1992;52:339–346. doi: 10.1016/0024-3205(93)90146-t. [DOI] [PubMed] [Google Scholar]
- 47.Léonard A., Gerber G.B. Mutagenicity, carcinogenicity and teratogenicity of vanadium compounds. Mut. Res. 1994;317:81–88. doi: 10.1016/0165-1110(94)90013-2. [DOI] [PubMed] [Google Scholar]
- 48.Domingo J.L. Vanadium and tungsten derivatives as antidiabetic agents: a review of their toxic effects. Biol. Trace Elem. Res. 2002;88:97–112. doi: 10.1385/BTER:88:2:097. [DOI] [PubMed] [Google Scholar]
- 49.Ścibior A., Kurus J. Vanadium and oxidative stress markers – in vivo model: a review. Curr. Med. Chem. 2019;26:5456–5500. doi: 10.2174/0929867326666190108112255. [DOI] [PubMed] [Google Scholar]
- 50.Bartosz G. Wyd. Nauk. PWN; Warszawa: 2013. Druga Twarz Tlenu. Wolne Rodniki w Przyrodzie. [Google Scholar]
- 51.Ścibior A., Zaporowska H. Effects of combined vanadate and magnesium treatment on erythrocyte antioxidant defence system in rats. Environ. Toxicol. Pharmacol. 2010;30:153–161. doi: 10.1016/j.etap.2010.05.003. [DOI] [PubMed] [Google Scholar]
- 52.Ścibior A., Gołębiowska D., Adamczyk A., Kurus J., Staniszewska M., Sadok I. Evaluation of lipid peroxidation and antioxidant defense mechanisms in the bone of rats in conditions of separate and combined administration of vanadium (V) and magnesium (Mg) Chem. Biol. Interact. 2018;284:112–125. doi: 10.1016/j.cbi.2018.02.016. [DOI] [PubMed] [Google Scholar]
- 53.Ścibior A., Adamczyk A., Gołębiowska D., Kurus J. Evaluation of lipid peroxidation and the level of some elements in rat erythrocytes during separate and combined vanadium and magnesium administration. Chem. Biol. Interact. 2018;293:1–10. doi: 10.1016/j.cbi.2018.07.014. [DOI] [PubMed] [Google Scholar]
- 54.Ścibior A., Zaporowska H., Ostrowski J. Selected haematological and biochemical parameters of blood in rats after subchronic administration of vanadium and/or magnesium in drinking water. Arch. Environ. Contam. Toxicol. 2006;51:287–295. doi: 10.1007/s00244-005-0126-4. [DOI] [PubMed] [Google Scholar]
- 55.Ścibior A., Zaporowska H., Niedźwiecka I. Lipid peroxidation in the liver of rats treated with V and/or Mg in drinking water, J. Appl. Toxicol. 2009;29:619–628. doi: 10.1002/jat.1450. [DOI] [PubMed] [Google Scholar]
- 56.Ścibior A., Zaporowska H., Niedźwiecka I. Lipid peroxidation in the kidney of rats treated with V and/or Mg in drinking water. J. Appl. Toxicol. 2010;30:487–496. doi: 10.1002/jat.1520. [DOI] [PubMed] [Google Scholar]
- 57.Ścibior A., Gołębiowska D., Niedźwiecka I. Magnesium can protect against vanadium-induced lipid peroxidation in the hepatic tissue. Oxid. Med. Cell. Longev, special issue LPPHHD. 2013;2013:1–11. doi: 10.1155/2013/802734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ścibior A., Gołębiowska D., Niedźwiecka I., Adamczyk A. Inhibitory and stimulating effect of magnesium on vanadate-induced lipid peroxidation under in vitro conditions. Ind. J. Exp. Biol. 2013;51:721–731. [PubMed] [Google Scholar]
- 59.Ścibior A., Zaporowska H. Effects of vanadium(V) and/or chromium(III) on L-ascorbic acid and glutathione as well as iron, zinc, and copper levels in rat liver and kidney. J. Toxicol. Environ. Health Part A. 2007;70:696–704. doi: 10.1080/15287390601187906. [DOI] [PubMed] [Google Scholar]
- 60.Sabbioni E., Rade J., Bertolero F. Relationships between iron and vanadium metabolism: the exchange of vanadium between transferrin and ferritin. J. Inorg. Biochem. 1980;12:307–315. doi: 10.1016/S0162-0134(00)80271-8. [DOI] [PubMed] [Google Scholar]
- 61.Ścibior A. Vanadium (V) and magnesium (Mg) – in vivo interactions: a review. Chem. Biol. Interact. 2016;258:214–233. doi: 10.1016/j.cbi.2016.09.007. [DOI] [PubMed] [Google Scholar]
- 62.Domingo J.L., Gomez M., Sanchez D.J., Llobet J.M., Keen C.L. Toxicology of vanadium compounds in diabetic rats: the action of chelating agents on vanadium accumulation. Mol. Cell. Biochem. 1995;153:233–240. doi: 10.1007/bf01075942. [DOI] [PubMed] [Google Scholar]
- 63.Ścibior A., Adamczyk A., Gołębiowska D., Niedźwiecka I., Fornal E. The influence of combined magnesium and vanadate administration on the level of some elements in rat organs: V-Mg interactions and the role of iron essentials protein DMT-1 in the mechanism underlying altered tissues iron level. Metallomics. 2014;6:907–920. doi: 10.1039/c3mt00363a. [DOI] [PubMed] [Google Scholar]
- 64.Ścibior A., Gołębiowska D., Adamczyk A., Niedźwiecka I., Fornal E. The renal effects of vanadate exposure: potential biomarkers and oxidative stress as a mechanism of functional renal disorders – preliminary studies. Biomed Res. Int. 2014;2014:1–15. doi: 10.1155/2014/740105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kiliańska Z.M., Żołnierczyk J., Węsierska-Gądek J. Biologiczna aktywność polimerazy poli(ADP-rybozy)-1. Postępy Hig. Med. Dośw. 2010;64:344–363. [PubMed] [Google Scholar]
- 66.Altamirano-Lozano M.A., Álvarez-Barrera L., Mateos-Nava R.A., Fortoul T.I., Rodríguez-Mercado J.J. Potential for genotoxic and reprotoxic effects of vanadium compounds due to occupational and environmental exposures: an article based on a presentation at the 8th International Symposium on vanadium chemistry, biological chemistry, and toxicology, Washington DC, August 15-18, 2012. J. Immunotoxicol. 2014;11:19–27. doi: 10.3109/1547691X.2013.791734. [DOI] [PubMed] [Google Scholar]
- 67.Rodríguez-Lara V., Morales-Rivero A., Rivera-Cambas A.M., Fortoul T.I. Vanadium inhalation induces actin changes in mice testicular cells. Toxicol. Ind. Health. 2016;32:367–374. doi: 10.1177/0748233713501364. [DOI] [PubMed] [Google Scholar]
- 68.Donaldson J., Hemming R., LaBella F. Vanadium exposure enhances lipid peroxidation in the kidney of rats and mice. Can. J. Physiol. Pharmacol. 1985;63:196–199. doi: 10.1139/y85-037. [DOI] [PubMed] [Google Scholar]
- 69.Younes M., Strubelt O. Vanadate-induced toxicity towards isolated perfused rat livers: the role of lipid peroxidation. Toxicology. 1991;66:63–74. doi: 10.1016/0300-483X(91)90178-4. [DOI] [PubMed] [Google Scholar]
- 70.Younes M., Kayser E., Strubelt O. Effect of antioxidants on vanadate-induced toxicity towards isolated perfused rat livers. Toxicology. 1991;70:141–149. doi: 10.1016/0300-483X(91)90041-X. [DOI] [PubMed] [Google Scholar]
- 71.Hosseini M.J., Seyedrazi N., Shahraki J., Pourahmad J. Vanadium induces liver toxicity through reductive activation by glutathione and mitochondrial dysfunction. Adv. Biosci. Biotechnol. 2012;3:1096–1103. doi: 10.4236/abb.2012.38134. [DOI] [Google Scholar]
- 72.Donaldson J., LaBella F. Prooxidant properties of vanadate in vitro on catecholamines and on lipid peroxidation by mouse and rat tissues. J. Toxicol. Environ. Health. 1983;12:119–126. doi: 10.1080/15287398309530411. [DOI] [PubMed] [Google Scholar]
- 73.Beyersmann D., Hartwig A. Carcinogenic metal compounds: recent insight into molecular and cellular mechanisms. Arch. Toxicol. 2008;82:493–512. doi: 10.1007/s00204-008-0313-y. [DOI] [PubMed] [Google Scholar]
- 74.Fortoul T.I., Rodriguez-Lara V., González-Villalva A., Rojas-Lemus M., Cano-Gutiérrez G., Ustarroz-Cano M., Colín L., Bizarro-Nevares P., García-Pealez I., Montaño L.F., Jimenez-Martinez R.S., Lopez-Valdez N., Ruiz-Guerrero M.L., Meléndez-García N.A., García-Ibarra F.A., Martínez-Baez V., Zapata Alfaro D., Muñiz-Rivera-Cambas A., López-Zepeda L.S., Quezada E., Cervantes-Yépez S. Inhalation of vanadium pentoxide and its toxic effects in a mouse model. Inorg. Chim. Acta Rev. 2014;420:8–15. doi: 10.1016/j.ica.2014.03.027. [DOI] [Google Scholar]
- 75.Olopade J.O., Connor J.R. Vanadium and neurotoxicity: a review. Curr. Topics Toxicol. 2011;7:33–39. [Google Scholar]
- 76.Fatola O.I., Olaolorun F.A., Olopade F.E., Olopade J.O. Trends in vanadium neurotoxicity. Brain Res. Bull. 2019;145:75–80. doi: 10.1016/j.brainresbull.2018.03.010. [DOI] [PubMed] [Google Scholar]
- 77.Stern A., Yin X., Tsang S.S., Davison A., Moon J. Vanadium as a modulator of cellular regulatory cascades and oncogene expression. Biochem. Cell Biol. 1993;71:103–112. doi: 10.1139/o93-018. [DOI] [PubMed] [Google Scholar]
- 78.Zychlinski L., Byczkowski J.Z., Kulkarni A.P. Toxic effects of long-term intratracheal administration of vanadium pentoxide in rats. Arch. Environ. Contam. Toxicol. 1991;20:295–298. doi: 10.1007/bf01064393. [DOI] [PubMed] [Google Scholar]
- 79.Catalá A. Lipid peroxidation of membrane phospholipids generates hydroxyl-alkenals and oxidized phospholipids active in physiological and/or pathological conditions. Chem. Phys. Lipids. 2009;157:1–11. doi: 10.1016/j.chemphyslip.2008.09.004. [DOI] [PubMed] [Google Scholar]
- 80.Rybakowska I., Stępińska M., Sein Anand J., Patock M. Korzystny i niekorzystny wpływ związków wanadu. Przegl. Lek. 2018;75:28–31. [Google Scholar]
- 81.Assem F.L., Levy L.S. Inhalation toxicity of vanadium. In: Michibata H., editor. Vanadium: Biochemical and Molecular Biological Approaches. Springer; 2012. pp. 209–224. Chapter 10. [Google Scholar]
- 82.Kabata-Pendias A., Pendias H. Wyd. Nauk. PWN; Warszawa: 1993. Biogeochemia Pierwiastków Śladowych. [Google Scholar]
- 83.Pałczyński C., Witczak T. Astma wywołana przez związki chemiczne o małej masie cząsteczkowej – część II. Alergia. 2010:12–17. [Google Scholar]
- 84.Musiałowicz E. Opis przypadku ostrego doustnego zatrucia pięciotlenkiem wanadu. Med. Pracy. 1976;27:395–397. [Google Scholar]
- 85.EPA United States Environmental Protection Agency . 2009. Provisional Peer-reviewed Toxicity Values for Vanadium and Its Soluble Inorganic Compounds Other Than Vanadium Pentoxide (CASRN 7440-62-2- and Others) EPA/690/R-09/070F. [Google Scholar]
- 86.Gawęda A. Pentatlenek wanadu w środowisku pracy – zagrożenia i oznaczanie. Bezpieczeństwo Pracy. 2001;7-8:18–21. [Google Scholar]
- 87.Sitprija V., Eiam-Ong S. Vanadium and metabolic problems. In: Nriagu J.O., editor. Vol. 31. John Wiley and Sons; New York, Chichester, Weinheim, Brisbane, Singapore, Toronto: 1998. pp. 91–120. (Vanadium in the Environment. Part 2: Health Effects). [Google Scholar]
- 88.Kučera J., Sabbioni E. Baseline vanadium levels in human blood, serum, and urine. In: Nriagu J.O., editor. Vol. 31. John Wiley and Sons; New York, Chichester, Weinheim, Brisbane, Singapore, Toronto: 1998. pp. 75–89. (Vanadium in the Environment. Part 2: Health Effects). [Google Scholar]
- 89.Baran E.J. Model studies related to vanadium biochemistry: recent advances and perspectives. J. Braz. Chem. Soc. 2003;14:878–888. doi: 10.1590/S0103-50532003000600004. [DOI] [Google Scholar]
- 90.Chasteen N.D., Lord E.M., Thompson H.J., Grady J.K. Vanadium complexes of transferrin and ferritin in the rat. Biochim. Biophys. Acta. 1986;884:84–92. doi: 10.1016/0304-4165(86)90230-8. [DOI] [PubMed] [Google Scholar]
- 91.Sanna D., Micera G., Garribba E. On the transport of vanadium in blood serum. Inorg. Chem. 2009;48:5747–5757. doi: 10.1021/ic802287s. [DOI] [PubMed] [Google Scholar]
- 92.Bogden J.D., Higashino H., Lavenhar M.A., Bauman J.W., Kemp F.W., Aviv A. Balance and tissue distribution of vanadium after short-term ingestion of vanadate. J. Nutr. 1982;112:2279–2285. doi: 10.1093/jn/112.12.2279. [DOI] [PubMed] [Google Scholar]
- 93.Azay J., Bres J., Krosniak M., Teissedre P.L., Cabanis J.C., Serrano J.J., Cros G. Vanadium pharmacokinetics and oral bioavailability upon single-dose administration of vanadyl sulfate to rats. Fundam. Clin. Pharmacol. 2001;15:313–324. doi: 10.1046/j.1472-8206.2001.00043.x. [DOI] [PubMed] [Google Scholar]
- 94.Sánchez C., Torres M., Bermúdez-Peña M.C., Aranda P., Montes-Bayón M., Sanz-Medel A., Llopis J. Bioavailability, tissue distribution and hypoglycaemic effect of vanadium in magnesium-deficient rats. Magnes. Res. 2011;24:196–208. doi: 10.1684/mrh.2011.0293. [DOI] [PubMed] [Google Scholar]
- 95.Roche M., Rondeau P., Singh N.R., Tarnus E., Bourdon E. The antioxidant properties of serum albumin. FEBS Lett. 2008;582:1783–1787. doi: 10.1016/j.febslet.2008.04.057. [DOI] [PubMed] [Google Scholar]
- 96.Chasteen N.D. Vanadium-protein interactions. In: Sigel H., Sigel A., editors. Vol. 31. Marcel Dekker, Inc.; New York: 1995. pp. 231–247. (Metal Ions in Biological Systems). [PubMed] [Google Scholar]
- 97.Correia I., Chorna I., Cavaco I., Roy S., Kuznetsov M.L., Ribeiro N., Justino G., Marques F., Santos-Silva T., Santos M.F.A., Santos H.M., Capelo J.L., Doutch J., Pessoa J.C. Interaction of [VIVO(acac)2] with human serum transferrin and albumin. Chem. Asian J. 2017;12:2062–2084. doi: 10.1002/asia.201700469. [DOI] [PubMed] [Google Scholar]
- 98.Harris W.R., Carrano C.J. Binding of vanadate to human serum transferrin. J. Inorg. Biochem. 1984;22:201–218. doi: 10.1016/0162-0134(84)80029-X. [DOI] [PubMed] [Google Scholar]
- 99.Azevedo C.G., Correia I., Dos Santos M.M.C., Santos M.F.A., Santos-Silva T., Doutch J., Fernandez L., Santos H.M., Capelo J.L., Pessoa J.C. Binding of vanadium to human serum transferrin - voltammetric and spectrometric studies. J. Inorg. Biochem. 2018;180:211–221. doi: 10.1016/j.jinorgbio.2017.12.012. [DOI] [PubMed] [Google Scholar]
- 100.Pessoa J.C., Goncalves G., Roy S., Correia I., Mehtab S., Santos M.F.A., Santos-Silva T. New insights on vanadium binding to human serum transferrin. Inorg. Chim. Acta Rev. 2014;420:60–68. doi: 10.1016/j.ica.2013.11.025. [DOI] [Google Scholar]
- 101.Treviño S., Díaz A., Sánchez-Lara E., Sanchez-Gaytan B.L., Perez-Aguilar J.M., González-Vergara E. Vanadium in biological action: chemical, pharmacological aspects, and metabolic implications in diabetic mellitus. Biol. Trace Elem. Res. 2019;188:68–98. doi: 10.1007/s12011-018-1540-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Yang X., Wang J., Lu J., Crans D.C. Membrane transport of vanadium compounds and the interaction with the erythrocyte membrane. Coord. Chem. Rev. 2003;237:103–111. doi: 10.1016/S0010-8545(02)00247-3. [DOI] [Google Scholar]
- 103.Sanna D., Serra M., Micera G., Garribba E. Uptake of potential anti-diabetic VIVO compounds of picolinate ligands by red blood cells. Inorg. Chim. Acta Rev. 2014;420:75–84. doi: 10.1016/j.ica.2013.12.038. [DOI] [Google Scholar]
- 104.Kiss T., Kiss E., Garribba E., Sakurai H. Speciation of insulin-mimetic VO (IV)-containing drugs in blood serum. J. Inorg. Biochem. 2000;80:65–73. doi: 10.1016/S0162-0134(00)00041-6. [DOI] [PubMed] [Google Scholar]
- 105.Sanna D., Micera G., Garribba E. Interaction of VO2+ ion and some insulin-enhancing compounds with immunoglobulin G. Inorg. Chem. 2011;50:3717–3728. doi: 10.1021/ic200087p. [DOI] [PubMed] [Google Scholar]
- 106.Rehder D. The future of/for vanadium. Dalton Trans. 2013;42:11749–11761. doi: 10.1039/c3dt50457c. [DOI] [PubMed] [Google Scholar]
- 107.Baran E.J. Vanadium detoxification. In: Nriagu J.O., editor. Vol. 31. John Wiley and Sons; New York, Chichester, Weinheim, Brisbane, Singapore, Toronto: 1998. pp. 317–345. (Vanadium in the Environment. Part 2: Health Effects). [Google Scholar]
- 108.De Cremer K., Van Hulle M., Chéry C., Cornelis R., Strijckmans K., Dams R., Lameire N., Vanholder R. Fractionation of vanadium complexes in serum, packed cells and tissues of Wistar rats by means of gel filtration and anion-exchange chromatography. J. Biol. Inorg. Chem. 2002;7:884–890. doi: 10.1007/s00775-002-0376-9. [DOI] [PubMed] [Google Scholar]
- 109.Sanna D., Serra M., Micera G., Garribba E. Interaction of antidiabetic vanadium compounds with hemoglobin and red blood cells and their distribution between plasma and erythrocytes. Inorg. Chem. 2014;53:1449–1464. doi: 10.1021/ic402366x. [DOI] [PubMed] [Google Scholar]
- 110.Laska A. Biomateriały stosowane w inżynierii tkankowej do regeneracji tkanek. Zesz. Nauk. Tow. Doktorantów UJ Nauki Ścisłe. 2017;14:187–196. [Google Scholar]
- 111.McCracken M. Dental implant materials: commercially pure titanium and titanium alloys. J. Prosthodont. 1999;8:40–43. doi: 10.1111/j.1532-849X.1999.tb00006.x. [DOI] [PubMed] [Google Scholar]
- 112.Gioka C., Bourauel C., Zinelis S., Eliades T., Silikas N., Eliades G. Titanium orthodontic brackets: structure, composition, hardness and ionic release. Dent. Mater. 2004;20:693–700. doi: 10.1016/j.dental.2004.02.008. [DOI] [PubMed] [Google Scholar]
- 113.Bombač D., Brojan M., Fajfar P., Kosel F., Turk R. Review of materials in medical applications. RMZ- Materials and Geoenvironment. 2007;54:471–499. [Google Scholar]
- 114.Elias C.N., Lima J.H.C., Valiev R., Meyers M.A. Biomedical applications of titanium and its alloys. JOM. 2008;60:46–49. [Google Scholar]
- 115.Geetha M., Singh A.K., Asokamani R., Gogia A.K. Ti based biomaterials, the ultimate choice for orthopaedic implants – a review. Prog. Mater. Sci. 2009;54:397–425. doi: 10.1016/j.pmatsci.2008.06.004. [DOI] [Google Scholar]
- 116.Palmquist A., Omar O.M., Esposito M., Lausmaa J., Thomsen P. Titanum oral implants: surface characteristics, interface biology and clinical outcome. J. R. Soc. Interface. 2010;7:515–527. doi: 10.1098/rsif.2010.0118.focus. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Oldani C., Dominguez A. Titanium as a biomaterial for implants. In: Fokter S., editor. Recent Advances in Arthroplasty. InTech; 2012. pp. 149–162. Chapter 9. [DOI] [Google Scholar]
- 118.Catalani S., Stea S.S., Beraudi A., Gilberti M.E., Bordini B., Toni A., Apostoli P. Vanadium release in whole blood, serum and urine of patients implanted with a titanium alloy hip prosthesis. Clin. Toxicol. 2013;51:550–556. doi: 10.3109/15563650.2013.818682. [DOI] [PubMed] [Google Scholar]
- 119.Chakravorty N., Jaiprakash A., Ivanovski S., Xiao Y. Implant surface modifications and osseointegration. In: Li Q., Mai Y.-W., editors. Biomaterials for Implants and Scaffolds. Springer Series in Biomaterials Science and Engineering 8; Germany: 2017. pp. 107–131. Chapter 4. [Google Scholar]
- 120.Dzhurinskiy D. Bioactive antimicrobial coatings for implantable medical devices formed by plasma electrolytic oxidation. Metal Forming. 2018;29:65–76. [Google Scholar]
- 121.Grill A. Diamond-like carbon coatings as biocompatible materials – an overview. Diam. Realt. Mater. 2003;12:166–170. doi: 10.1016/S0925-9635(03)00018-9. [DOI] [Google Scholar]
- 122.Anicic N., Vukomanovic M., Suvorov D. Design of a multifunctional vanadium pentoxide/polymer biocomposite for implant-coating applications. RSC Adv. 2017;7:38647–38658. doi: 10.1039/C7RA06471C. [DOI] [Google Scholar]
- 123.Pessoa J.C., Etcheverry S., Gambino D. Vanadium compounds in medicine. Coord. Chem. Rev. 2015;301-302:24–48. doi: 10.1016/j.ccr.2014.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Pessoa J.C. Thirty years through vanadium chemistry. J. Inorg. Biochem. 2015;147:4–24. doi: 10.1016/j.jinorgbio.2015.03.004. [DOI] [PubMed] [Google Scholar]
- 125.Turner T.L., Nguyen V.H., McLauchlan C.C., Dymon Z., Dorsey B.M., Hooker J.D., Jones M.A. Inhibitory effects of decavanadate on several enzymes and Leishmania tarentolae in vitro. J. Inorg. Biochem. 2012;108:96–104. doi: 10.1016/j.jinorgbio.2011.09.009. [DOI] [PubMed] [Google Scholar]
- 126.Naderer T., Ellis M.A., Sernee M.F., De Souza D.P., Curtis J., Handman E., McConville M.J. Virulence of Leishmania major in macrophages and mice requires the gluconeogenic enzyme fructose-1,6-bisphosphatase. Proc. Natl. Acad. Sci. 2006;103:5502–5507. doi: 10.1073/pnas.0509196103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Mendez R.S., Dorsey B.M., McLauchlan C.C., Beio M., Turner T.L., Nguyen V.H., Su A., Beynon W., Friesen J.A., Jones M.A. Vanadium complexes are in vitro inhibitors of Leishmania secreted acid phosphatases. Int. J. Chem. 2014;6:35–49. doi: 10.5539/ijc.v6n1p35. [DOI] [Google Scholar]
- 128.Fernandes A.C.S., Soares D.C., Saraiva E.M., Meyer-Fernandes J.R., Sauto-Padrón T. Different secreted phosphatase activities in Leishmania amazonensis. FEMS Microbiol. Lett. 2013;340:117–128. doi: 10.1111/1574-6968.12080. [DOI] [PubMed] [Google Scholar]
- 129.Adriazola I.O., Evangelista do Amaral A., Amorim J.C., Correia B.L., Petkowicz C.L., Mercê A.L., Noleto G.R. Macrophage activation and leishmanicidal activity by galactomannan and its oxovanadium (IV/V) complex in vitro. J. Inorg. Biochem. 2014;132:45–51. doi: 10.1016/j.jinorgbio.2013.09.017. [DOI] [PubMed] [Google Scholar]
- 130.Maqsood Z.T., Khan K.M., Ashiq U., Jamal R.A., Chohan Z.H., Mahroof-Tahir M., Supuran C.T. Oxovanadium(IV) complexes of hydrazides: potential antifungal agents. J. Enzyme Inhib. Med. Chem. 2006;21:37–42. doi: 10.1080/14756360500277459. [DOI] [PubMed] [Google Scholar]
- 131.Chohan Z.H., Sumrra S.H., Youssoufi M.H., Hadda T.B. Metal based biologically active compounds: design, synthesis, and antibacterial/antifungal/cytotoxic properties of triazole-derived Schiff bases and their oxovanadium(IV) complexes. Eur. J. Med. Chem. 2010;45:2739–2747. doi: 10.1016/j.ejmech.2010.02.053. [DOI] [PubMed] [Google Scholar]
- 132.Pahontu E., Julea F., Rosu T., Purcarea V., Chumakov Y., Petrenco P., Gulea A. Antibacterial, antifungal and in vitro antileukaemia activity of metal complexes with thiosemicarbazones. J. Cell. Mol. Med. 2015;19:865–878. doi: 10.1111/jcmm.12508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Jayaseelan P., Akila E., Rani M.U., Rajavel R. Synthesis, spectral characterization, electrochemical, anti-microbial, DNA binding and cleavage studies of new binuclear Schiff base metal(II) complexes derived from o-hydroxyacetophenone. J. Saudi Chem. Soc. 2016;20:625–634. doi: 10.1016/j.jscs.2013.07.001. [DOI] [Google Scholar]
- 134.Li H., Gong H., Qi Y., Li J., Ji X., Sun J., Tian R., Bao H., Song X., Chen Q., Liu G. In vitro and in vivo antifungal activities and mechanism of heteropolytungstates against Candida species. Sci. Rep. 2017;7:1–9. doi: 10.1038/s41598-017-17239-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Guo S., Yang W., Zjao M., Tian R., Zhang B., Qi Y. In vitro anticandidal activity and mechanism of polyoxovanadate functionalized by Zn-fluconazole complexes. Molecules. 2018;23:1–11. doi: 10.3390/molecules23051122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Correia I., Adão P., Roy S., Wahba M., Matos C., Maurya M.R., Marques F., Pavan F.R., Leite C.Q.F., Avecilla F., Pessoa J.C. Hydroxyquinoline derived vanadium(IV and V) and copper(II) complexes as potential anti-tuberculosis and anti-tumor agents. J. Inorg. Biochem. 2014;141:83–93. doi: 10.1016/j.jinorgbio.2014.07.019. [DOI] [PubMed] [Google Scholar]
- 137.Bijelic A., Aureliano M., Rompel A. The antibacterial activity of polyoxometalates: structures, antibiotic effects and future perspectives. Chem. Commun. 2018;54:1153–1169. doi: 10.1039/C7CC07549A. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Adachi Y., Yoshikawa Y., Yoshida J.Y., Kodera Y., Katoch A., Takada J., Sakurai H. Improvement of diabetes, obesity and hypertension in type 2 diabetic KKAy mice by bis(allixinato)oxovanadium(IV) complex. Biochem. Biophys. Res. Commun. 2006;345:945–950. doi: 10.1016/j.bbrc.2006.05.003. [DOI] [PubMed] [Google Scholar]
- 139.Takada Y., Hashimoto M., Kasahara J., Aihara K., Fukunaga K. Cytoprotective effect of sodium orthovanadate on ischemia/reperfusion-induced injury in the rat heart involves Akt activation and inhibition of fodrin breakdown and apoptosis. J. Pharmacol. Exp. Ther. 2004;311:1249–1255. doi: 10.1124/jpet.104.070839. [DOI] [PubMed] [Google Scholar]
- 140.Bhuiyan M.S., Shibuya M., Shioda N., Moriguchi S., Kasahara J., Iwabuchi Y., Fukunaga K. Cytoprotective effect of bis(1-oxy-2-pyridinethiolato)oxovanadium(IV) on myocardial ischemia/reperfusion injury elicits inhibition of Fas ligand and Bim expression and elevation of FLIP expression. Eur. J. Pharmacol. 2007;571:180–188. doi: 10.1016/j.ejphar.2007.05.046. [DOI] [PubMed] [Google Scholar]
- 141.Park S.J., Youn C.K., Hyun J.W., You H.J. The anti-obesity effect of natural vanadium-containing Jeju grand water. Biol. Trace Elem. Res. 2013;151:294–300. doi: 10.1007/s12011-012-9557-8. [DOI] [PubMed] [Google Scholar]
- 142.Liem D.A., Gho C.C., Gho B.C., Kazim S., Manintveld O.C., Verdouw P.D., Duncker D.J. The tyrosine phosphatase inhibitor bis(maltolato)oxovanadium attenuates myocardial reperfusion injury by opening ATP-sensitive potassium channels. J. Pharmacol. Exp. Ther. 2004;309:1256–1262. doi: 10.1124/jpet.103.062547. [DOI] [PubMed] [Google Scholar]
- 143.Aureliano M., Fraqueza G., Ohlin C.A. Ion pumps as biological targets for decavanadate. Dalton Trans. 2013;42:11770–11777. doi: 10.1039/c3dt50462j. [DOI] [PubMed] [Google Scholar]
- 144.Pietrusiński M., Stączek P. Badania struktury i funkcji prokariotycznych topoizomeraz klasy II szansą dla poszukiwania i syntezy nowych leków przeciwbakteryjnych. Postępy Biochem. 2006;52:271–282. [PubMed] [Google Scholar]
- 145.Szafran M., Zakrzewska-Czerwińska J., Jakimowicz D. Bakteryjne topoizomerazy typu I – rola biologiczna i zastosowanie jako potencjalnych celów dla antybiotyków. Postępy Hig. Med. Dośw. 2013;67:130–142. doi: 10.5604/17322693.1038352. [DOI] [PubMed] [Google Scholar]
- 146.Suh D., Chaires J.B. Criteria for the mode of binding of DNA binding agents. Bioorg. Med. Chem. 1995;3:723–728. doi: 10.1016/0968-0896(95)00053-J. [DOI] [PubMed] [Google Scholar]
- 147.Halliwell B. Phagocyte-derived reactive species: salvation or suicide? Trends Biochem. Sci. 2006;31:509–515. doi: 10.1016/j.tibs.2006.07.005. [DOI] [PubMed] [Google Scholar]
- 148.Wong Y.C., Kim S., Peng W., Krainc D. Regulation and function of mitochondria-lysosome membrane contact sites in cellular homeostasis. Trends Cell Biol. 2019;29:500–513. doi: 10.1016/j.tcb.2019.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Korbecki J., Baranowska-Bosiacka I., Gutowska I., Chlubek D. Biochemical and medical importance of vanadium compounds. Acta Biochim. Pol. 2012;59:195–200. [PubMed] [Google Scholar]
- 150.Cunha-de Padua M.M., Suter Correia Cadena S.M., de Oliveira Petkowicz C.L., Martinez G.R., Merlin Rocha M.E., Mercê A.L.R., Noleto G.R. Toxicity of native and oxovanadium (IV/V) galactomannan complexes on HepG2 cells is related to impairment of mitochondrial functions. Carbohydr. Polym. 2017;173:665–675. doi: 10.1016/j.carbpol.2017.06.027. [DOI] [PubMed] [Google Scholar]
- 151.Szemraj J., Rozpończyk E., Bartkowiak J., Greger J., Oszajca K. Znaczenie białka MDM2 w cyklu komórkowym. Postępy Biochem. 2005;51:44–51. [PubMed] [Google Scholar]
- 152.Chakraborty T., Ghosh S., Datta S., Chakraborty P., Chatterjee M. Vanadium suppresses sister-chromatid exchange and DNA-protein crosslink formation and restores antioxidant status and hepatocellular architecture during 2-acetylaminofluorene-induced experimental rat hepatocarcinogenesis. J. Exp. Ther. Oncol. 2003;3:346–362. doi: 10.1111/j.1533-869X.2003.01107.x. [DOI] [PubMed] [Google Scholar]
- 153.León I.E., Porro V., Di Virgilio A.L., Bollati-Fogolin M., Castelli S., Desideri A., Etcheverry S.B. Book of Abstract. 9th International Vanadium Symposium; Padova, Italy: 2014. Anticancer drug development. Effects of a complex of oxidovanadium(IV) with silibinin in osteoblast cell lines. Relationship with the Inhibition of Topoisomerase IB; p. 72. [Google Scholar]
- 154.Mańkowska D. Ludzkie topoizomerazy jako cel komórkowy współczesnej chemioterapii. Farm. Przegl. Nauk. 2009;9:30–33. [Google Scholar]
- 155.Papaioannou A., Manos M., Karkabounas S., Liasko R., Evangelou A.M., Correia I., Kalfakakou V., Pessoa J.C., Kabanos T. Solid state and solution studies of a vanadium(III)-L-cysteine compound and demonstration of its antimetastatis, antioxidant and inhibition of neutral endopeptidase activities. J. Inorg. Biochem. 2004;98:959–968. doi: 10.1016/j.jinorgbio.2004.02.011. [DOI] [PubMed] [Google Scholar]
- 156.Sobocińska M., Kamysz E. Potencjalne zastosowanie enkefalin w diagnostyce i terapii różnych chorób. Wiad. Chem. 2017;71:33–44. [Google Scholar]
- 157.Bishayee A., Chatterjee M. Selective enhancement of glutathione S-transferase activity in liver and extrahepatic tissues of rat following oral administration of vanadate. Acta Physiol. Pharmacol. Bulg. 1993;19:83–89. [PubMed] [Google Scholar]
- 158.Bishayee A., Chatterjee M. Time course effects of vanadium supplement on cytosolic reduced glutathione level and glutathione S-transferase activity. Biol. Trace Elem. Res. 1995;48:275–285. doi: 10.1007/bf02789409. [DOI] [PubMed] [Google Scholar]
- 159.Bishayee A., Roy S., Chatterjee M. Characterization of selective induction and alteration of xenobiotic biotransforming enzymes by vanadium during diethylnitrosamine-induced chemical rat liver carcinogenesis. Oncol. Res. 1999;11:41–53. [PubMed] [Google Scholar]
- 160.Bishayee A., Oinam S., Basu M., Chatterjee M. Vanadium chemoprevention of 7,12-dimethylbenz(a)anthracene-induced rat mammary carcinogenesis: probable involvement of representative hepatic phase I and II xenobiotic metabolizing enzymes. Breast Cancer Res. Treat. 2000;63:133–145. doi: 10.1023/a:1006476003685. [DOI] [PubMed] [Google Scholar]
- 161.Kanna P.S., Mahendrakumar C.B., Chatterjee M., Hemalatha P., Datta S., Chakraborty P. Vanadium inhibits placental glutathione S-transferase (GST-P) positive foci in 1,2-dimethyl hydrazine induced rat colon carcinogenesis. J. Biochem. Mol. Toxicol. 2003;17:357–365. doi: 10.1002/jbt.10099. [DOI] [PubMed] [Google Scholar]
- 162.Ni L., Zhao H., Tao L., Li X., Zhou Z., Sun Y., Chen C., Wei D., Liu Y., Diao G. Synthesis, in vitro cytotoxicity, and structure-activity relationships (SAR) of multidenate oxidovanadium(IV) complexes as anticancer agents. Dalton Trans. 2018;47:10035–10045. doi: 10.1039/C8DT01778F. [DOI] [PubMed] [Google Scholar]
- 163.Aliabad H.B., Falahati-Pour S.K., Ahmadirad H., Mohamadi M., Hajizadeh M.R., Mahmoodi M. Vanadium complex: an appropriate candidate for killing hepatocellular carcinoma cancerous cells. Biometals. 2018;31:981–990. doi: 10.1007/s10534-018-0139-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Kowalski Sz., Wyrzykowski D., Hac S., Rychlowski M., Radomski M.W., Inkielewicz-Stępniak I. New oxidovanadium(IV) coordination complex containing 2-methylnitrilotriacetate ligands induces cell cycle arrest and autophagy in human pancreatic ductal adenocarcinoma cell lines. Int. J. Mol. Sci. 2019;20:261. doi: 10.3390/ijms20020261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Kowalski Sz., Hać S., Wyrzykowski D., Zauszkiewicz-Pawlak A., Inkielewicz-Stępniak I. Selective cytotoxicity of vanadium complexes on human pancreatic ductal adenocarcinoma cell line by inducing necroptosis, apoptosis and mitotic catastrophe process. Oncotarget. 2017;8:60324–60341. doi: 10.18632/oncotarget.19454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Patra D., Paul S., Majumder I., Sepay N., Bera S., Kundu R., Drew M.G.B., Gosh T. Exploring the effect of substituent in the hydrazone ligand of a family of u-oxidodivanadium(V) hydrazone complexes on structure, DNA binding and anti-cancer activity. Dalton Trans. 2017;46:16276–16293. doi: 10.1039/c7dt03585c. [DOI] [PubMed] [Google Scholar]
- 167.Reytman L., Hochman J., Tshuva E.Y. Anticancer diaminotris(phenolato) vanadium(V) complexes: ligand-metal interplay. J. Coord. Chem. 2018;71:2003–2011. doi: 10.1080/00958972.2018.1461848. [DOI] [Google Scholar]
- 168.Crans D.C., Yang L., Haase A., Yang X. Health benefits of vanadium and its potential as an anticancer agent. In: Sigel A., Sigel H., Freisinger E., Sigel R.K.O., editors. Vol. 18. Walter de Gruyter GmbH; Berlin/Boston: 2018. pp. 251–279. (Metal Ions in Life Sciences). [DOI] [Google Scholar]
- 169.Boduła A., Wdowczyk M., Adamiec R. Rola białkowej fosfatazy tyrozynowej 1B (PTP-1B) w rozwoju insulinooporności. Postępy Hig. Med. Dośw. 2005;59:203–207. [PubMed] [Google Scholar]
- 170.Buglyo P., Crans D.C., Nagy E.M., Lindo R.L., Yang L., Smee J.J., Jin W., Chi L.H., Godzala M.E., Willsky G.R. Aqueous Chemistry of the VanadiumIII (VIII) and the VIII−Dipicolinate systems and a comparison of the effect of three oxidation states of vanadium compounds on diabetic hyperglycemia in rats. Inorg. Chem. 2005;44:5416–5427. doi: 10.1021/ic048331q. [DOI] [PubMed] [Google Scholar]
- 171.Mohammad A., Sharma V., McNeil J.H. Vanadium increases GLUT4 in diabetic rat skeletal muscle. Mol. Cell. Biochem. 2002;233:139–143. doi: 10.1023/a:1015558328757. [DOI] [PubMed] [Google Scholar]
- 172.Korbecki J., Baranowska-Bosiacka I., Gutowska I., Chlubek D. Insulino-mimetyczne właściwości związków wanadu. Postępy Biochem. 2016;62:60–65. [PubMed] [Google Scholar]
- 173.Kordowiak A.M., Holko P. Pochodne wanadu jako związki o istotnym znaczeniu biologicznym. Część I. Działanie przeciwcukrzycowe, Postępy Biol. Kom. 2009;36:361–376. [Google Scholar]
- 174.Domingo J.L., Gómez M. Vanadium compounds for the treatment of human diabetes mellitus: a scientific curiosity? A review of thirty years of research. Food Chem. Toxicol. 2016;95:137–141. doi: 10.1016/j.fct.2016.07.005. [DOI] [PubMed] [Google Scholar]
- 175.Tolman E.L., Barris E., Burns M., Pansini A., Partridge R. Effects of vanadium on glucose metabolism in vitro. Life Sci. 1979;25:1159–1164. doi: 10.1016/0024-3205(79)90138-3. [DOI] [PubMed] [Google Scholar]
- 176.Dubyak G.R., Kleinzeller A. The insulin-mimetic effects of vanadate in isolated rat adipocytes. Dissociation from effects of vanadate as a (Na+-K+)ATPase inhibitor. J. Biol. Chem. 1980;255:5306–5312. [PubMed] [Google Scholar]
- 177.Shechter Y., Karlish S.J. Insulin-like stimulation of glucose oxidation in rat adipocytes by vanadyl (IV) ions. Nature. 1980;284:556–558. doi: 10.1038/284556a0. [DOI] [PubMed] [Google Scholar]
- 178.Heyliger C.E., Tahiliani A.G., McNeill J.H. Effect of vanadate on elevate blood glucose and decreased cardiac performance of diabetic rats. Science. 1985;227:1474–1477. doi: 10.1126/science.3156405. [DOI] [PubMed] [Google Scholar]
- 179.Meyerovitch J., Farfel Z., Sack J., Shechter Y. Oral administration of vanadate normalizes blood glucose levels in streptozotocin-treated rats. Characterization and mode of action. J. Biol. Chem. 1987;262:6658–6662. [PubMed] [Google Scholar]
- 180.Thompson K.H., Lichter J., LeBel C., Scaife M.C., McNeill J.H., Orvig C. Vanadium treatment of type 2 diabetes: a view to the future. J. Inorg. Biochem. 2009;103:554–558. doi: 10.1016/j.jinorgbio.2008.12.003. [DOI] [PubMed] [Google Scholar]
- 181.Thompson K.H., Orvig C. Vanadium in diabetes: 100 years from phase 0 to phase I. J. Inorg. Biochem. 2006;100:1925–1935. doi: 10.1016/j.jinorgbio.2006.08.016. [DOI] [PubMed] [Google Scholar]
- 182.2009. Akesis Pharmaceuticals Discontinues Sole Clinical Development Program and Announces Intent to File for. Chapter 7 Bankruptcy, January 21. [Google Scholar]
- 183.De la Tore A., Granero S., Mayayo E., Corbella J., Domingo J.L. Effect of age on vanadium nephrotoxicity in rats. Toxicol. Lett. 1999;105:75–82. doi: 10.1016/s0378-4274(98)00385-3. [DOI] [PubMed] [Google Scholar]
- 184.Al-Bayati M.A., Xie Y., Mohr F.C., Margolin S.B., Giri S.N. Effect of pirfenidone against vanadate-induced kidney fibrosis in rats. Biochem. Pharmacol. 2002;64:517–525. doi: 10.1016/s0006-2952(02)01213-3. [DOI] [PubMed] [Google Scholar]
- 185.Al-Bayati M.A., Giri S.N., Raabe O.G., Rosenblatt L.S., Shifrine M. Time and dose-response study of the effects of vanadate on rats: morphological and biochemical changes in organs. J. Environ. Pathol. Toxicol. Oncol. 1989;9(5–6):435–455. [PubMed] [Google Scholar]
- 186.Boscolo P., Carmignani M., Volpe A.R., Felaco M., Del Rosso G., Porcelli G., Giuliano G. Renal toxicity and arterial hypertension in rats chronically exposed to vanadate. Occup. Environ. Med. 1994;51:500–503. doi: 10.1136/oem.51.7.500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Cohen N., Halberstam M., Shilimovich P., Chang C.J., Shamoon H., Rossetti L. Oral vanadyl sulfate improves hepatic and peripheral insulin sensitivity in patients with non-insulin-dependent diabetes mellitus. J. Clin. Invest. 1995;95:2501–2509. doi: 10.1172/JCI117951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Halberstam M., Cohen N., Shlimovich P., Rossetti L., Shamoon H. Oral vanadyl sulfate improves insulin sensitivity in NIDDM but not in obese nondiabetic subjects. Diabetes. 1996;45:659–666. doi: 10.2337/diab.45.5.659. [DOI] [PubMed] [Google Scholar]
- 189.Boden G., Chen X., Ruiz J., van Rossum G.D., Turco S. Effects of vanadyl sulfate on carbohydrate and lipid metabolism in patients with non-insulin-dependent diabetes mellitus. Metabolism. 1996;45:1130–1135. doi: 10.1016/s0026-0495(96)90013-x. [DOI] [PubMed] [Google Scholar]
- 190.Goldfine A.B., Simonson D.C., Folli F., Patti M.E., Kahn C.R. In vivo and in vitro studies of vanadate in human and rodent diabetes mellitus. Mol. Cell. Biochem. 1995;153:217–231. doi: 10.1007/bf01075941. [DOI] [PubMed] [Google Scholar]
- 191.Goldfine A.B., Simonson D.C., Folli F., Patti M.E., Kahn C.R. Metabolic effects of sodium metavanadate in humans with insulin-dependent and noninsulin-dependent diabetes mellitus in vivo and in vitro studies. J. Clin. Endocrinol. Metab. 1995;80:3311–3320. doi: 10.1210/jcem.80.11.7593444. [DOI] [PubMed] [Google Scholar]
- 192.Goldfine A.B., Patti M.E., Zuberi L., Goldstein B.J., LeBlanc R., Landaker E.J., Jiang Z.Y., Willsky G.R., Kahn C.R. Metabolic effects of vanadyl sulfate in humans with non-insulin-dependent diabetes: in vivo and in vitro studies. Metabolism. 2000;49:400–410. doi: 10.1016/S0026-0495(00)90418-9. [DOI] [PubMed] [Google Scholar]
- 193.Cusi K., Cukier S., DeFronzo R.A., Torres M., Puchulu F.M., Redondo J.C.P. Vanadyl sulfate improves hepatic and muscle insulin sensitivity in type 2 diabetes. J. Clin. Endocrinol. Metab. 2001;86:1410–1417. doi: 10.1210/jcem.86.3.7337. [DOI] [PubMed] [Google Scholar]
- 194.Jacques-Camarena O., González-Ortiz M., Martínez-Abundis E., López-Madrueno J.F., Medina-Santillán R. Effect of vanadium on insulin sensitivity in patients with impaired glucose tolerance. Ann. Nutr. Metab. 2008;53:195–198. doi: 10.1159/000175844. [DOI] [PubMed] [Google Scholar]
- 195.Soveid M., Dehghani G.A., Omrani G.R. Long-term efficacy and safety of vanadium in the treatment of type 1 diabetes. Arch. Iran. Med. 2013;16:408–411. doi:013167/AIM.009. [PubMed] [Google Scholar]
- 196.Adam A.M.A., Naglah A.M., Al-Omar M.A., Refat M.S. Synthesis of a new insulin-mimetic antidiabetic drug containing vitamin A and vanadium(IV) salt: chemico-biological characterizations. Int. J. Immunopathol. Pharmacol. 2017;30:272–281. doi: 10.1177/0394632017719601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Krośniak M., Szklarzewicz J., Gryboś R., Tatar B., Yildirim M., Sahin B., Yuksek N.D., Ustundag M. The influence of chronic supply of vanadium compounds on organ weights and body mass in animals diabetes model (NZO) Sci. Tech. Innov. 2019;4:63–73. doi: 10.5604/01.3001.0013.1563. [DOI] [Google Scholar]
- 198.Szklarzewicz J., Jurowska A., Olszewska A., Hodorowicz M., Gryboś R., Kruczała K. Synthesis, structure and properties of V(IIIIV and V) complexes with ONO Schiff bases. Sci. Tech. Innov. 2019;4:37–46. doi: 10.5604/01.3001.0013.1505. [DOI] [Google Scholar]
- 199.Domingo J.L. Vanadium and diabetes. What about vanadium toxicity? Mol. Cell. Biochem. 2000;203:185–187. doi: 10.1023/a:1007067011338. [DOI] [PubMed] [Google Scholar]
- 200.Bhuiyan M.S., Fukunaga K. Cardioprotection by vanadium compounds targeting Akt-mediated signaling. J. Pharmacol. Sci. 2009;110:1–13. doi: 10.1254/jphs.09r01cr. [DOI] [PubMed] [Google Scholar]
- 201.Szydłowski M., Jabłońska E., Juszczyński P. Rola ścieżki sygnałowej PI3K-AKT w ontogenezie limfocytów B i patogenezie nowotworów B-komórkowych. Cześć I. Hematologia. 2013;4:103–113. [Google Scholar]
- 202.Strijdom H., Chamane N., Lochner A. Nitric oxide in the cardiovascular system: a simple molecule with complex actions. Cardiovasc. J. Afr. 2009;20:303–310. [PMC free article] [PubMed] [Google Scholar]
- 203.Pasławska U., Kurzok H., Kiczak L., Bania J. Rola tlenku azotu w regulacji czynności układu krążenia. Med. Wet. 2012;68:349–352. [Google Scholar]
- 204.Kopp S.J., Daar J., Paulson D.J., Romano F.D., Laddaga R. Effects of oral vanadyl treatment on diabetes-induced alterations in the heart GLUT-4 transporter. J. Mol. Cell. Cardiol. 1997;29:2355–2362. doi: 10.1006/jmcc.1997.0469. [DOI] [PubMed] [Google Scholar]
- 205.Sandirasegarane L., Gopalakrishnan V. Vanadate increases cytosolic free calcium in rat aortic smooth muscle cells. Life Sci. 1995;56:PL169–PL174. doi: 10.1016/0024-3205(94)00487-D. [DOI] [PubMed] [Google Scholar]
- 206.Nayler R.A., Sparrow M.P. Mechanism of vanadate-induced contraction of airways smooth muscle of the guinea-pig. Br. J. Pharmacol. 1983;80:163–172. doi: 10.1111/j.1476-5381.1983.tb11062.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Cadène A., Grigorescu F., Serrano J.J., Cros G. Characterization of vanadyl sulfate on vascular contraction: roles of calcium and tyrosine phosphorylation. J. Pharmacol. Exp. Ther. 1997;281:491–498. [PubMed] [Google Scholar]
- 208.Bhanot S., McNeill J.H. Vanadyl sulfate lowers plasma insulin and blood pressure in spontaneously hypertensive rats. Hypertension. 1994;24:625–632. doi: 10.1161/01.hyp.24.5.625. [DOI] [PubMed] [Google Scholar]
- 209.Bhanot S., McNeill J.H., Bryer-Ash M. Vanadyl sulfate prevents fructose-induced hyperinsulinemia and hypertension in rats. Hypertension. 1994;23:308–312. doi: 10.1161/01.hyp.23.3.308. [DOI] [PubMed] [Google Scholar]
- 210.Bhanot S., Michoulas A., McNeill J.H. Antihypertensive effects of vanadium compounds in hyperinsulinemic, hypertensive rats. Mol. Cell. Biochem. 1995;153:205–209. doi: 10.1007/bf01075939. [DOI] [PubMed] [Google Scholar]
- 211.Kawano T., Fukunaga K., Takeuchi Y., Morioka M., Yano S., Hamada J., Ushio Y., Miyamoto E. Neuroprotective effect of sodium orthovanadate on delayed neuronal death after transient forebrain ischemia in gerbil hippocampus. J. Cereb. Blood Flow Metab. 2001;21:1268–1280. doi: 10.1097/00004647-200111000-00003. [DOI] [PubMed] [Google Scholar]
- 212.Hasegawa Y., Hamada J., Morioka M., Yano S., Kawano T., Kai Y., Fukunaga K., Ushio Y. Neuroprotective effect of postischemic administration of sodium orthovanadate in rats with transient middle cerebral artery occlusion. J. Cereb. Blood Flow Metab. 2003;23:1040–1051. doi: 10.1097/01.WCB.0000085160.71791.3F. [DOI] [PubMed] [Google Scholar]
- 213.Hasegawa Y., Morioka M., Hasegawa S., Matsumoto J., Kawano T., Kai Y., Tano S., Fukunaga K., Kuratsu J. Therapeutic time window and dose dependence of neuroprotective effects of sodium orthovanadate following transient middle cerebral artery occlusion in rats. J. Pharmacol. Exp. Ther. 2006;317:875–881. doi: 10.1124/jpet.105.096677. [DOI] [PubMed] [Google Scholar]
- 214.Shioda N., Ishigami T., Han F., Moriguchi S., Shibuya M., Iwabuchi Y., Fukunaga K. Activation of phosphatidylinositol 3-kinase/protein kinase B pathway by a vanadyl compound mediates its neuroprotective effect in mouse brain ischemia. Neuroscience. 2007;148:221–229. doi: 10.1016/j.neuroscience.2007.05.040. [DOI] [PubMed] [Google Scholar]
- 215.Han F., Shioda N., Moriguchi S., Qin Z.H., Fukunaga K. The vanadium (IV) compound rescues septo-hippocampal cholinergic neurons from neurodegeneration in olfactory bulbectomized mice. Neuroscience. 2008;15:671–679. doi: 10.1016/j.neuroscience.2007.11.011. [DOI] [PubMed] [Google Scholar]
- 216.Sánchez-González C., Rivas-García L., López-Chaves C., Rodríguez-Nogales A., Algieri F., Gálvez J., Gómez-Aracena J., Vera-Ramírez L., Montes-Bayon M., Sanz-Medel A., Llopis J. Exposure to bis(maltolato)oxovanadium(IV) increases levels of hepcidin mRNA and impairs the homeostasis of iron but not that of manganese. Food Chem. Toxicol. 2014;73:113–118. doi: 10.1016/j.fct.2014.08.011. [DOI] [PubMed] [Google Scholar]
- 217.Kita A., Uotani S., Kuwahara H., Takahashi R., Oshima K., Yamasaki H., Mizuguchi H., Hayakawa T., Nagayama Y., Yamaguchi Y., Eguchi K. Vanadate enhances leptin-induced activation of JAK/STAT pathway in CHO cells. Biochem. Biophys. Res. Commun. 2003;302:805–809. doi: 10.1016/S0006-291X(03)00264-X. [DOI] [PubMed] [Google Scholar]
- 218.Wilsey J., Matheny M.K., Scarpace P.J. Oral vanadium enhances the catabolic effects of central leptin in young adult rats. Endocrinology. 2006;147:493–501. doi: 10.1210/en.2004-1358. [DOI] [PubMed] [Google Scholar]
- 219.Myers M.P., Andersen J.N., Cheng A., Tremblay M.L., Horvath C.M., Parisien J.P., Salmeen A., Barford D., Tonks N.K. TYK2 and JAK2 are substrates of protein-tyrosine phosphatase 1B. J. Biol. Chem. 2001;276:47771–47774. doi: 10.1074/jbc.C100583200. [DOI] [PubMed] [Google Scholar]
- 220.Zabolotny J.M., Bence-Hanulec K.K., Stricker-Krongrad A., Haj F., Wang Y., Minokoshi Y., Kim Y.B., Elmquist J.K., Tartaglia L.A., Kahn B.B., Neel B.G. PTP1B regulates signal transduction in vivo. Dev. Cell. 2002;2:489–495. doi: 10.1016/S1534-5807(02)00148-X. [DOI] [PubMed] [Google Scholar]
- 221.Gogga P., Karbowska J., Meissner W., Kochan Z. Rola leptyny w regulacji metabolizmu lipidów i węglowodanów. Postępy Hig. Med. Dośw. 2011;65:255–262. doi: 10.5604/17322693.940259. [DOI] [PubMed] [Google Scholar]
- 222.Mohammad A., Wang J., McNeil J.H. Bis(maltolato)oxovanadium(IV) inhibits the activity of PTP-1B in Zucker rat skeletal muscle in vivo. Mol. Cell. Biochem. 2002;229:125–128. doi: 10.1023/a:1017984930836. [DOI] [PubMed] [Google Scholar]
- 223.Nogalska A., Świerczyński J. Leptyna – hormon o wielu funkcjach. Postępy Biochem. 2001;47:200–211. [PubMed] [Google Scholar]
- 224.Zabolotny J.M., Haj F.G., Kim Y.B., Kim H.J., Shulman G.I., Kim J.K., Neel B.G., Kahn B.B. Transgenic overexpression of protein-tyrosine phosphatase 1B in muscle causes insulin resistance but overexpression with leukocyte antygen-related phosphatase doses not additively impair insulin action. J. Biol. Chem. 2004;279:24844–24851. doi: 10.1074/jbc.M310688200. [DOI] [PubMed] [Google Scholar]
- 225.Czyżewska M., Wolska A., Ćwiklińska A., Kortas-Stempak B., Wróblewska M. Zaburzenia metabolizmu lipoprotein w zespole metabolicznym. Postępy Hig. Med. Dośw. 2010;64:1–10. [PubMed] [Google Scholar]
- 226.Dubiński A., Zdrojewicz Z. Rola leptyny w rozwoju nadciśnienia tętniczego. Postępy Hig. Med. Dośw. 2006;60:447–452. [PubMed] [Google Scholar]
- 227.Karbowska J., Kochan Z. Leptyna jako hormon łączący otyłość z dysfunkcją mięśnia sercowego. Postępy Hig. Med. Dośw. 2012;66:267–274. doi: 10.5604/17322693.997817. [DOI] [PubMed] [Google Scholar]
- 228.Missaoui S., Rhouma K.B., Yacoubi M.T., Sakly M., Tebourbi O. Vanadyl sulfate treatment stimulates proliferation and regeneration of beta cells in pancreatic islets. J. Diabetes Res. 2014;2014:1–7. doi: 10.1155/2014/540242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Roess D.A., Smith S.M.L., Winter P., Zhou J., Dou P., Baruah B., Trujillo A.M., Levinger N.E., Yang X., Barisas B.G., Crans D.C. Effects of vanadium-containing compounds on membrane lipids and on microdomains used in receptor-mediated signaling. Chem. Biodiver. 2008;5:1558–1570. doi: 10.1002/cbdv.200890144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Yuen V.G., Pederson R.A., Dai S., Orvig C., McNeill J.H. Effects of low and high dose administration of bis(maltolato)oxovanadium(IV) on fa/fa Zucker rats. Can. J. Physiol. Pharmacol. 1996;74:1001–1009. doi: 10.1139/y96-102. [DOI] [PubMed] [Google Scholar]
- 231.Curran G.L. Effect of certain transition group elements on hepatic synthesis of cholesterol in the rat. J. Biol. Chem. 1954;210:765–770. [PubMed] [Google Scholar]
- 232.Azarnoff D.L., Curran G.L. Site of vanadium inhibition of cholesterol biosynthesis. J. Am. Chem. Soc. 1957;79:2968–2969. doi: 10.1021/ja01568a074. [DOI] [Google Scholar]
- 233.Curran G.L., Azarnoff D.L., Bolinger R.E. Effect of cholesterol synthesis inhibition in normocholesteremic young men. J. Clin. Invest. 1959;38:1251–1261. doi: 10.1172/JCI103900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Wieczorek B., Gniot-Szulżycka J. HMG-CoA reduktaza – kluczowy enzym w biosyntezie izoprenoidów i cholesterolu. Kosmos. 2000;49:191–206. [Google Scholar]
- 235.Rezk Naser L., Rezk Salma A. Vanadium in therapy and toxicity; mini–review and recommendation of further study, MOJ Bioequiv. Availab. 2017;3:105–106. doi: 10.15406/mojbb.2017.03.00042. [DOI] [Google Scholar]
- 236.Braun J.E., Severson D.L. Lipoprotein lipase release from cardiac myocytes is increased by decavanadate but not insulin. Am. J. Physiol. 1992;262:E663–E670. doi: 10.1152/ajpendo.1992.262.5.E663. [DOI] [PubMed] [Google Scholar]
- 237.Mead J.R., Irvine S.A., Ramji D.P. Lipoprotein lipase: structure, function, regulation, and role in disease. J. Mol. Med. 2002;80:753–769. doi: 10.1007/s00109-002-0384-9. [DOI] [PubMed] [Google Scholar]
- 238.Zhang Y., Zhang Q., Feng C., Ren X., Li H., He K., Wang F., Zhou D., Lan Y. Influence of vanadium on serum lipid and lipoprotein profiles: a population-based study among vanadium exposed workers. Lipids Health Dis. 2014;13:1–6. doi: 10.1186/1476-511X-13-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Pugazhenthi S., Hussain A., Yu B., Brownsey R.W., Angel J.F., Khandelwal R.L. Vanadate induces normolipidemia and a reduction in the levels of hepatic lipogenic enzymes in obese Zucker rat. Mol. Cell. Biochem. 1995;153:211–215. doi: 10.1007/bf01075940. [DOI] [PubMed] [Google Scholar]
- 240.Ramanadham S., Mongold J.J., Brownsey R.W., Cros G.H., McNeill J.H. Oral vanadyl sulfate in treatment of diabetic mellitus in rats. Am. J. Physiol. 1989;257:H904–H911. doi: 10.1152/ajpheart.1989.257.3.H904. [DOI] [PubMed] [Google Scholar]
- 241.Domingo J.L., Gómez M., Llobet J.M., Corbella J., Keen C.L. Improvement of glucose homeostasis by oral vanadyl or vanadate treatment in diabetic rats is accompanied by negative side effects. Pharmacol. Toxicol. 1991;68:249–253. doi: 10.1111/j.1600-0773.1991.tb01233.x. [DOI] [PubMed] [Google Scholar]
- 242.Cam M.C., Pederson R.A., Brownsey R.W., McNeill J.H. Long-term effectiveness of oral vanadyl sulphate in streptozotocin-diabetic rats. Diabetologia. 1993;36:218–224. doi: 10.1007/bf00399953. [DOI] [PubMed] [Google Scholar]
- 243.Majithiya J.B., Balaraman R., Giridhar R., Yadav M.R. Effect of bis(curcumino)oxovanadium complex on non-diabetic and streptozotocin-induced diabetic rats. J. Trace Elem. Med. Biol. 2005;18:211–217. doi: 10.1016/j.jtemb.2004.12.001. [DOI] [PubMed] [Google Scholar]
- 244.Li M., Smee J.J., Ding W., Crans D.C. Anti-diabetic effects of sodium 4-amino-2,6-dipicolinatodioxovanadium(V) dihydrate in streptozotocin-induced diabetic rats. J. Inorg. Biochem. 2009;103:585–589. doi: 10.1016/j.jinorgbio.2008.11.011. [DOI] [PubMed] [Google Scholar]
- 245.Wasan K.M., Risovic V., Yuen V.G., McNeill J.H. Differences in plasma homocysteine levels between Zucker fatty and Zucker diabetic fatty rats following 3 weeks oral administration of organic vanadium compounds. J. Trace Elem. Med. Biol. 2006;19:251–258. doi: 10.1016/j.jtemb.2005.10.001. [DOI] [PubMed] [Google Scholar]
- 246.Zaporowska H., Wasilewski W. Wpływ wanadu na układ krwiotwórczy i wybrane wskaźniki krwi obwodowej szczurów szczepu Wistar. Bromat. Chem. Toksykol. 1989;22:121–125. [Google Scholar]
- 247.Zaporowska H., Wasilewski W. Some selected peripheral blood and haemopoietic system indices in Wistar rats with chronic vanadium intoxication. Comp. Biochem. Physiol. C. 1989;93:175–180. doi: 10.1016/0742-8413(89)90030-3. [DOI] [PubMed] [Google Scholar]
- 248.Zaporowska H., Wasilewski W. Haematological results of vanadium intoxication in Wistar rats. Comp. Biochem. Physiol. C. 1992;101:57–61. doi: 10.1016/0742-8413(92)90199-H. [DOI] [PubMed] [Google Scholar]
- 249.Zaporowska H. Effect of vanadium on L-ascorbic acid concentration in rat tissues. Gen. Pharmacol. 1994;25:467–470. doi: 10.1016/0306-3623(94)90199-6. [DOI] [PubMed] [Google Scholar]
- 250.Russanov E., Zaporowska H., Ivancheva E., Kirkova M., Konstantinova S. Lipid peroxidation and antioxidant enzymes in vanadate-treated rats. Comp. Biochem. Physiol. Pharmacol. Toxicol. Endocrinol. 1994;107:415–421. doi: 10.1016/1367-8280(94)90070-1. [DOI] [PubMed] [Google Scholar]
- 251.Kučera J., Lener J., Mñuková J., Bayerová E. Vanadium exposure tests in humans: hair, nails, blood, and urine. In: Nriagu J.O., editor. Vol. 31. John Wiley and Sons; New York, Chichester, Weinheim, Brisbane, Singapore, Toronto: 1998. pp. 55–73. (Vanadium in the Environment. Part 2: Health Effects). [Google Scholar]
- 252.Lees R.E.M. Changes in lung function after exposure to vanadium compounds in fuel oil ash. Br. J. Ind. Med. 1980;37:253–256. doi: 10.1136/oem.37.3.253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Matsuda M., Mandarino L., DeFronzo R.A. Synergistic interaction of magnesium and vanadate on glucose metabolism in diabetic rats. Metabolism. 1999;48:725–731. doi: 10.1016/S0026-0495(99)90171-3. [DOI] [PubMed] [Google Scholar]
- 254.Barbagallo M., Dominguez L.J. Magnesium and type 2 diabetes. World J. Diabetes. 2015;6:1152–1157. doi: 10.4239/wjd.v6.i10.1152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Ayepola O.R., Brooks N.L., Oguntibeju O.O. Oxidative stress and diabetic complications: the role of antioxidant vitamins and flavonoids. In: Oguntibeju O., editor. Antioxidant-Antidiabetic Agents and Human Health. Intech; 2014. pp. 25–58. Chapter 2. [DOI] [Google Scholar]
- 256.Dos Santos J.M., Tewari S., Mendes R.H. The role of oxidative stress in the development of diabetes mellitus and its complications. J. Diab. Res. 2019;4189813:3. doi: 10.1155/2019/4189813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Bermúdez-Peña M.C., López-Chaves C., Llopis J., Guerrero-Romero F., Montes-Bayón M., Sanz-Medel A., Sánchez-González C. Aggravation by vanadium of magnesium deficiency in STZ-induced diabetic rats. Magnes. Res. 2013;26:74–82. doi: 10.1684/mrh.2013.0342. [DOI] [PubMed] [Google Scholar]
- 258.Berg L.R. Effect of diet composition on vanadium toxicity for the chick. Poult. Sci. 1966;45:1346–1352. doi: 10.3382/ps.0451346. [DOI] [PubMed] [Google Scholar]
- 259.Ścibior A., Adamczyk A., Gołębiowska D., Niedźwiecka I. Effect of 12-week vanadate and magnesium co-administration on chosen haematological parameters as well as on some indices of iron and copper metabolism and biomarkers of oxidative stress in rats. Environ. Toxicol. Pharmacol. 2012;34:235–252. doi: 10.1016/j.etap.2012.04.006. [DOI] [PubMed] [Google Scholar]
- 260.Ścibior A., Adamczyk A., Mroczka R., Niedźwiecka I., Gołębiowska D., Fornal E. Effects of vanadium (V) and magnesium (Mg) on rat bone tissue: mineral status and micromorphology. Consequences of V-Mg interactions, Metallomics. 2014;6:2260–2278. doi: 10.1039/c4mt00234b. [DOI] [PubMed] [Google Scholar]
- 261.Garcia L.A., Dejong S.C., Martin S.M., Smith R.S., Buettner G.R., Kerber R.E. Magnesium reduces free radicals in an in vivo coronary occlusion-reperfusion model. J. Am. Coll. Cardiol. 1998;32:536–539. doi: 10.1016/s0735-1097(98)00231-9. [DOI] [PubMed] [Google Scholar]
- 262.Mak I.T., Komarov A.M., Kramer J.H., Weglicki W.B. Protective mechanisms of MG-gluconate against oxidative endothelial cytotoxicity. Cell. Mol. Biol. 2000;46:1337–1344. [PubMed] [Google Scholar]
- 263.Hans C.P., Chaudhary D.P., Bansal D.D. Effect of magnesium supplementation on oxidative stress in alloxanic diabetic rats. Magnes. Res. 2003;16:13–19. [PubMed] [Google Scholar]
- 264.Zhang Y., Davies L.R., Martin S.M., Bawaney I.M., Buettner G.R., Kerber R.E. Magnesium reduces free radical concentration and preserves left ventricular function after direct current shocks. Resuscitation. 2003;56:199–206. doi: 10.1016/S0300-9572(02)00353-2. [DOI] [PubMed] [Google Scholar]
- 265.Henry P.R., Miles R.D. Interactions among the trace minerals. Ciencia Anim. Bras. 2000;1:95–106. [Google Scholar]
- 266.Yamaguchi M., Sishi H., Suketa Y. Effect of vanadium on bone metabolism in weanling rats: zinc prevents the toxic effect of vanadium. Res. Exp. Med. 1989;189:47–53. doi: 10.1007/bf01856029. [DOI] [PubMed] [Google Scholar]
- 267.Haider S.S., Abdel-Gayoum A.A., El-Fakhri M., Ghwarsha K.M. Effect of selenium on vanadium toxicity in different regions of rat brain. Hum. Exp. Toxicol. 1998;17:23–28. doi: 10.1177/096032719801700104. [DOI] [PubMed] [Google Scholar]
- 268.Ścibior A. Wybrane parametry układu antyoksydacyjnego szczurów w interakcji wanad-selen oraz wanad-cynk. Dr. dissertation. 1999:1–141. [Google Scholar]
- 269.Ścibior A. Some selected blood parameters in rats exposed to vanadium and chromium via drinking water. Trace Elem. Electroly. 2005;22:40–46. doi:10.5414/TEP22040. [Google Scholar]
- 270.Ścibior A., Zaporowska H., Ostrowski J., Banach A. Combined effect of vanadium(V) and chromium(III) on lipid peroxidation in liver and kidney of rats. Chem-Biol. Interact. 2006;159:213–222. doi: 10.1016/j.cbi.2005.11.008. [DOI] [PubMed] [Google Scholar]
- 271.Ścibior A., Zaporowska H. Effects of combined treatment with vanadium(V) and chromium(III) on hematological parameters and plasma mineral status in rats. Trace Elem. Electroly. 2009;26:104–113. , doi:10.5414/TEP26104. [Google Scholar]
- 272.Ścibior A., Zaporowsk H., Wolińska A., Ostrowski J. Antioxidant enzyme activity and lipid peroxidation in the blood of rats co-treated with vanadium(V5+) and chromium (Cr3+) Cell Biol. Toxicol. 2010;26:509–526. doi: 10.1007/s10565-010-9160-8. [DOI] [PubMed] [Google Scholar]
- 273.Wang Q., Liu T.T., Fu Y., Wang K., Yang X.G. Vanadium compounds discriminate hepatoma and normal hepatic cells by differential regulation of reactive oxygen species. J. Biol. Inorg. Chem. 2010;15:1087–1096. doi: 10.1007/s00775-010-0668-4. [DOI] [PubMed] [Google Scholar]
- 274.Aldini G., Altomare A., Baron G., Vistoli G., Carini M., Borsani L., Sergio F. N-acetylcysteine as an antioxidant and disulphide breaking agent: the reasons why. Free Rad. Res. 2018;52:751–762. doi: 10.1080/10715762.2018.1468564. [DOI] [PubMed] [Google Scholar]
- 275.Zwolak I., Gołębiowska D. Protective activity of pyruvate against vanadium-dependent cytotoxicity in Chinese hamster ovary (CHO-K1) cells. Toxicol. Ind. Health. 2018;34:283–292. doi: 10.1177/0748233718754979. [DOI] [PubMed] [Google Scholar]
- 276.Hu M., Coetzee P.P. Vanadium speciation by chromatographic separation of V(IV) and V(V) in acidic solution followed by ICP-OES determination. Water SA. 2007;33:291–296. doi: 10.4314/wsa.v33i2.49097. [DOI] [Google Scholar]
- 277.Tsave O., Petanidis S., Kioseoglou E., Yavropoulou M.P., Yovos J.G., Anestakis D., Tsepa A., Salifoglou A. Role of vanadium in cellular and molecular immunology: association with immune-related inflammation and pharmacotoxicology mechanisms. Oxid. Med. Cell. Longev. 2016;2016:1–10. doi: 10.1155/2016/4013639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278.Rozzo C., Sanna D., Garribba E., Serra M., Cantara A., Palmieri G., Pisano M. Antitumoral effect of vanadium compounds in malignant melanoma cell lines. J. Inorg. Biochem. 2017;174:14–24. doi: 10.1016/j.jinorgbio.2017.05.010. [DOI] [PubMed] [Google Scholar]