

Summary
The entry of two dengue virus (DENV) serotypes into Vero cells was analysed using biochemical inhibitors, dominant negative mutants of cellular proteins involved in endocytic pathways, fluorescence microscopy and infectivity determinations. By treatment with dansylcadaverine and chlorpromazine and overexpression of a dominant negative form of the Eps15 protein, a clathrin‐mediated endocytosis for productive DENV‐1 internalization into Vero cells was demonstrated whereas the infectious entry of DENV‐2 in the same cell system was independent of clathrin. Treatment with the inhibitors nystatin and methyl‐β‐cyclodextrin, as well as transfection of Vero cells with dominant negative caveolin‐1, had no effect on DENV‐2 virus infection. It was also shown, by using the K44A mutant and the inhibitor dynasore, that dynamin was required for DENV‐2 entry. Consequently, the infectious entry of DENV‐2 into Vero cells occurs by a non‐classical endocytic pathway independent of clathrin, caveolae and lipid rafts, but dependent on dynamin. By contrast, DENV‐2 entry into A549 cells was clathrin‐dependent, as previously reported in HeLa, C6/36 and BS‐C‐1 cells. Our results conclusively show, for the first time, a differential mode of infective entry for DENV‐1 and DENV‐2 into a common host cell, Vero cells, as well as alternative entry pathways for a given serotype, DENV‐2, into different types of cells.
Introduction
Dengue virus (DENV) is an enveloped, positive single‐stranded RNA virus belonging to the genus Flavivirus, family Flaviviridae, responsible of the most prevalent arthropode‐borne viral infection in humans. There are four antigenically related serotypes (DENV‐1 to DENV‐4), which cocirculate in tropical and subtropical regions around the world between their vectors, the mosquitoes Aedes aegypti and A. albopictus, and the vertebrate hosts. DENV human infection can be manifested either by the benign self‐limited febrile illness called dengue fever (DF) or by the severe and potentially fatal disease named dengue haemorrhagic fever/dengue shock syndrome (DHF/DSS) (Gubler, 2002). Primary infection induces protective immunity only against the infecting serotype, and secondary infection with other serotype appears to be one of the risk factors to develop DHF/DSS through an immunopathological process of antibody‐dependent enhancement (ADE) (Kliks et al., 1989; Rothman, 2004). The failure to maintain programmes for controlling the mosquito vector has caused the global re‐emergence of this infection, turning DENV into a public health problem. Currently, it is estimated that the virus is endemic in more than 100 countries, producing about 50 million infections each year (Gubler, 2002).
Despite the increasing incidence of DENV as a human pathogen, there are no antiviral agents or vaccines for treatment or prevention, and little is known about the DENV multiplication cycle. In particular, the initial steps leading to DENV entry into the host cells for primary infection are presently very poorly understood. The events involved in virus entry comprise virion binding to the cell surface receptors followed by internalization into the cytoplasm. The envelope E glycoprotein has been identified as the DENV protein involved in both attachment and internalization, representing a key determinant for infection. By contrast, the identity of the cellular receptor interacting with E glycoprotein is, at present, controversial. Several recent reports have implicated a long list of receptor candidate molecules, including a wide diversity of proteins and heparan sulfate proteoglycans (reviewed in Acosta et al., 2008a). A probable reason for this complexity may be the ability of DENV to replicate in a wide spectrum of cells (Diamond et al., 2000), suggesting the possibility of divergent receptors according to the type of host cell. A plausible mechanism emerging from these conflicting data is a multistep process consisting of the sequential interaction of E with at least two target molecules: an abundant attachment receptor to concentrate virus particles on the cell surface and facilitate the subsequent interaction with a high affinity second receptor of protein nature which mediates virus internalization. According to the cell type, heparan sulfate and the lectin DC‐SIGN have been proposed as putative primary receptors (Martínez‐Barragán and del Angel, 2001; Lozach et al., 2005).
Upon receptor binding, enveloped viruses exploit two main pathways for internalization of the nucleocapsid into the cytoplasm: membrane fusion at the cell surface (pH‐independent) or within intracellular vesicles after endocytosis (pH‐dependent). Earlier studies showed by electron microscopy observation that DENV‐2 penetrated directly into the cytoplasm of human monocytes, mosquito and BHK cells by fusion of the virion envelope with the plasma membrane at physiological pH (Hase et al., 1989; Lim and Ng, 1999) whereas the formation of low pH‐induced syncytia in DENV‐2 infected cells was also published (Randolph and Stollar, 1990). More recent reports have provided evidence that uptake of DENV‐2 virions in HeLa, mosquito C6/36, BHK‐15, and BSC‐1 cells occurs through receptor‐mediated endocytosis, dependent on intravesicular low pH for membrane fusion activity (Krishnan et al., 2007; van der Schaar et al., 2007; Acosta et al., 2008b; Mosso et al., 2008), similarly as it was observed for other members of Flavivirus (Mizutani et al., 2003; Nawa et al., 2003; Chu and Ng, 2004; Chu et al., 2006).
Despite the general consensus about a pH requirement for viral fusion, the information about the precise intracellular pathway for DENV internalization is scanty and controversial. For productive infection of HeLa, mosquito and BS‐C‐1 cells, the functional entry of DENV‐2 is clathrin‐dependent (Krishnan et al., 2007; Acosta et al., 2008b; Mosso et al., 2008; van der Schaar et al., 2008), the most common and well‐known mode of endocytosis. But contrasting results have been reported about the role of membrane cholesterol for DENV‐2 entry in diverse cellular systems such as human monocytes (Reyes‐del Valle et al., 2005), mouse neuroblastoma cells (Lee et al., 2008) and mosquito cells (Acosta et al., 2008b; Mosso et al., 2008; Umashankar et al., 2008). Additionally, there are no reports addressing the functional entry mechanism utilized by serotypes other than DENV‐2. The clinical characteristics associated with the distinct serotypes cannot be precisely defined, but diverse epidemiological studies have reported that DENV‐2 caused more severe disease than DENV‐1 (Vaughn et al., 2000; Nisalak et al., 2003; Balmaseda et al., 2006). Thus, the knowledge of the virus–cell interactions leading to successful infection for different DENV serotypes may help to understand their varying in vivo pathogenic profile.
Here we have analysed comparatively the infectious entry of DENV‐1 and DENV‐2 serotypes in Vero cells by the systematic utilization of a variety of biochemical and molecular inhibitors. Our results conclusively demonstrate that alternative entry routes are taken by different DENV serotypes within a host cell and, also, by the same serotype in different cell systems.
Results
Kinetics and rate of viral internalization
To define accurately the conditions of the viral internalization assay required for monitoring DENV entry under different treatments, we determined first the kinetics and rate of virus penetration into Vero cells by measuring productive internalized virus particles. To this end, cells were infected and incubated at 4°C for 1 h to allow virus attachment. Then, cells were washed with PBS at 4°C and shifted at 37°C to permit internalization of the bound particles. At various times after the shift, a series of infected cultures were treated with proteinase K to remove adsorbed virus whereas internalized virus remained protected inside the cell; after inactivation of the enzyme, suspensions of infected cells were plated on Vero cell monolayers to quantify infectious internalized particles by plaque formation. Another series of cultures were treated with PBS after each time of temperature shift and a similar infectious centre assay was performed to quantify total cell‐associated virus (membrane‐bound and internalized virions). The percentage of PFU resistant to proteinase K treatment in comparison with control PBS treatment was calculated for each time point. As shown in Fig. 1A, at the beginning of the temperature shift, most DENV‐2 particles were detached from cells by proteinase K, but when the time of incubation at 37°C increased virus became rapidly resistant. After 60–90 min of incubation, 88–94% of virus was resistant to proteinase K treatment in comparison to control treatment, indicating a successful internalization of infectious virions. A similar profile in time and amount of internalized virus into Vero cells determined by the infectious centre assay was obtained for DENV‐1 (data not shown).
Figure 1.
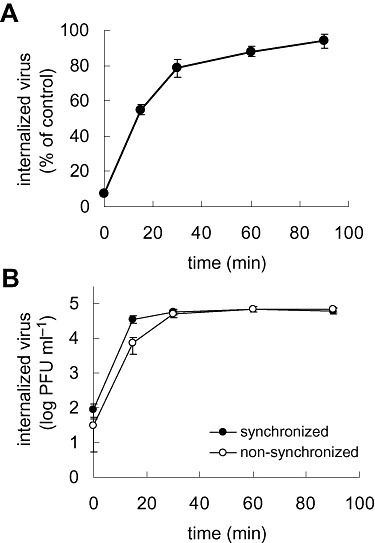
Kinetics and rate of DENV internalization into Vero cells. A. Vero cells were infected with DENV‐2 during 1 h at 4°C and then transferred to 37°C. At the indicated times after incubation at 37°C, extracellular virus was inactivated with proteinase K. Results are shown as a percentage of internalized virus when compared with controls in which PBS was substituted for proteinase K. B. Vero cells were infected with DENV‐2 at 37°C (non‐synchronized infection) or at 4°C and then transferred to 37°C (synchronized infection). At the indicated times extracellular virus was inactivated with proteinase K. Results are expressed as number of internalized PFU.
Most inhibitors of endocytic pathways to be tested lack cytotoxicity only when cells are treated for very short time periods in certain culture conditions (see Experimental procedures). In particular, cells treated with the inhibitors did not maintain fully viable when incubated at 4°C. Consequently, the kinetics and rate of virus internalization were tested in cells non‐pre‐incubated during 1 h at 4°C, a condition previously chosen to separate adsorption and internalization processes. Vero cells were infected with DENV, immediately incubated at 37°C, and at different times the amount of internalized virus particles was determined by proteinase K treatment and infectious centre assay as described above. The curve of virus internalization in these assay conditions was comparable to that obtained with 1 h of preincubation at 4°C (Fig. 1B). A slight difference in the number of infectious internalized virus was initially detected, probably due to the lack of synchronization for penetration of pre‐adsorbed virions. But, after 30 min at 37°C, the amount of proteinase K‐resistant virus penetrated into the cell was similar for both assays. Thus, the effect of inhibitors on virus internalization was measured by protease treatment and infectious centre assay after 1 h of DENV infection at 37°C.
DENV‐1 and DENV‐2 entry to Vero cells is low pH dependent
To test the role of low pH for DENV‐1 and DENV‐2 entry to Vero cells, we evaluated the effect of ammonium chloride, a lysosomotropic weak base that immediately raises the pH of intracellular acidic vesicles, and concanamycin A, a selective inhibitor of the vacuolar proton ATPase (Mizzen et al., 1985; Castilla et al., 1994; Guinea and Carrasco, 1994). Treatment of Vero cells with ammonium chloride and concanamycin A before infection and during the first hour of infection at 37°C significantly reduced the amount of internalized virus for both serotypes, with maximum effect at a concentration of 50 mM and 50 nM respectively (Fig. 2A). This low pH requirement was also assessed by inhibition of viral yield (Fig. S1A) and viral antigen expression detected by immunofluorescence assay (Fig. 2B). However, no inhibitory effect on virus multiplication was detected when ammonium chloride was added 1 h after infection, confirming that this compound effectively blocked an early event during the virus entry process (Fig. 2B). Furthermore, no virucidal effect of ammonium chloride was detected when the virus was pre‐incubated with the compound before infection (Fig. S1B) neither virus adsorption was affected (Fig. S1C). Similarly, lack of effect on virion inactivation and adsorption was detected with concanamycin A (data not shown). To ensure that the drug concentrations used effectively increased the pH of intracellular vesicles in Vero cells we performed acridine orange staining. Untreated cells showed the typical cytoplasmatic orange fluorescence of the acid compartments, whereas cells treated with ammonium chloride and concanamycin A did not exhibit this fluorescence pattern (Fig. S1D). Thus, it can be concluded that DENV‐1 and DENV‐2 require the acidic intracellular pH to penetrate into Vero cells indicating that virus entry occurs through a low pH‐dependent endocytic route.
Figure 2.
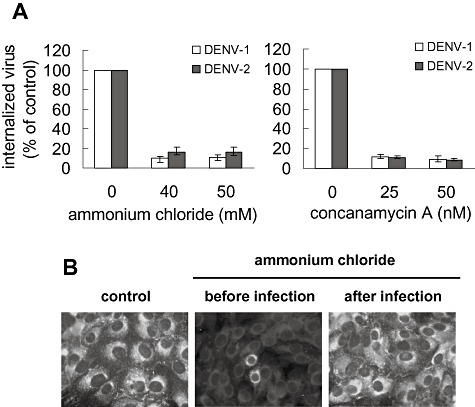
DENV‐1 and DENV‐2 entry into Vero cells is dependent of acidic pH. A. Cells were treated with ammonium chloride or concanamycin A and infected with DENV‐1 or DENV‐2. After 1 h internalization in the presence of the drugs, extracellular virus was inactivated with proteinase K and the cell pellets were plated onto Vero cells to determine internalized virus by an infectious centre assay. Results are expressed as percentage of internalized virus with respect to a control without drug treatment. Each point shows the mean ± SD of two independent experiments. B. Cells were treated with 50 mM ammonium chloride before or after infection with DENV‐2 or were left untreated (control). At 48 h p.i. immunostaining was carried out using mouse anti‐E glycoprotein antibody and FITC‐labelled anti‐mouse IgG.
Clathrin‐mediated endocytosis is required for DENV‐1 entry and infection of Vero cells, but not for DENV‐2
Clathrin‐mediated endocytosis is the most classical low pH‐dependent pathway used by different viruses to enter into the cells (Sieczkarski and Whittaker, 2002a; Marsh and Helenius, 2006). Additionally, the involvement of clathrin for DENV‐2 entry to mosquito C6/36 cells (Acosta et al., 2008b; Mosso et al., 2008), human HeLa cells (Krishnan et al., 2007) and BS‐C‐1 cells (van der Schaar et al., 2008) has been recently reported. To address the involvement of a clathrin‐mediated pathway in DENV‐1 and DENV‐2 entry to Vero cells, we analysed the effects on virus infection of chlorpromazine and dansylcadaverine, two pharmacological inhibitors of this pathway (Chu et al., 2006; Leppert et al., 2007; Zheng et al., 2008). A dose‐dependent inhibition of DENV‐1 internalization (Fig. 3A) and production at 48 h post‐infection (p.i.) (Fig. 3B) was obtained in Vero cells treated with both compounds during the first hour of infection, with a maximum reduction of virus internalization to 33% and 29% of control values at concentrations of 50 μM chlorpromazine and 500 μM dansylcadaverine respectively. Surprisingly, neither chlorpromazine nor dansylcadaverine treatment produced any inhibitory effect on DENV‐2 infection (Fig. 3A and B). By the contrary, a significant increase in the number of DENV‐2 internalized virions and PFU released from infected Vero cells was detected when clathrin‐dependent endocytosis was blocked (values higher than 100% of control in Fig. 3A and B). The internalization of TRITC‐labelled transferrin, a ligand known to enter into the cell by clathrin‐mediated endocytosis, was used as a functional control assay to assess that the action of the inhibitors was effectively exerted on receptor‐mediated endocytosis from clathrin‐coated pits. In control cells a dotted fluorescence pattern inside the cell cytoplasm was observed, while cultures treated with chlorpromazine exhibited a widespread fluorescence only at the cell surface, indicating that transferrin uptake was efficiently arrested (Fig. 3C). A similar staining pattern was observed after treatment with dansylcadaverine (data not shown). Consequently, results obtained with biochemical inhibitors suggest that DENV‐1 entry to Vero cells occurs through clathrin‐dependent endocytosis but DENV‐2 would use another endocytic route.
Figure 3.
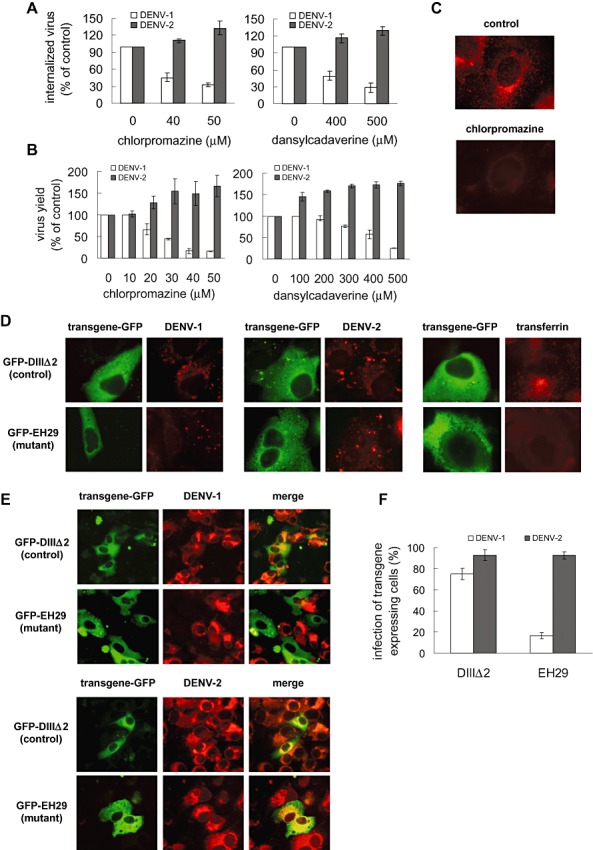
Entry of DENV‐1 into Vero cells requires clathrin‐mediated endocytosis while entry of DENV‐2 is clathrin‐independent. A. Cells were treated with chlorpromazine or dansylcadaverine and infected with DENV‐1 or DENV‐2. After 1 h internalization in the presence of the drugs, monolayers were treated with proteinase K and the cell pellets were plated onto Vero cells to determine internalized virus by an infectious centre assay. B. Cells were treated as in (A) and infected with DENV‐1 or DENV‐2. Virus yields were quantified at 48 h p.i. Results are expressed as percentage of virus multiplication with respect to a control without drug treatment. C. Cells were treated with 50 μM chlorpromazine or left untreated (control) and incubated with TRITC‐labelled transferrin. D. Cells transiently transfected with the constructs GFP‐DIIIΔ2 or GFP‐EH29 were infected with DENV‐1 or DENV‐2 or incubated with TRITC‐labelled transferrin. After 1 h cells were fixed and internalized viral particles were visualized by immunofluorescence staining using mouse anti‐E glycoprotein antibody and TRITC‐labelled anti‐mouse IgG. E. Cells transiently transfected with GFP‐DIIIΔ2 or GFP‐EH29 were infected with DENV‐1 or DENV‐2. After 24 h cells were fixed and viral antigen expression was visualized by immunofluorescence staining using mouse anti‐E glycoprotein antibody and TRITC‐labelled anti‐mouse IgG. F. For quantification of samples shown in (E), 250 transfected cells with similar levels of GFP expression were screened and cells positive for viral antigen were scored. In (A), (B) and (F) results are expressed as the mean ± SD of two independent experiments.
To assess the differential role of clathrin‐mediated endocytosis for the entry of both serotypes into Vero cells, a specific molecular inhibitor was next assayed. The dominant negative mutant of the clathrin coat‐associated cellular protein Eps15 named EH29 specifically interferes with clathrin‐coated pit assembly at the plasma membrane without affecting clathrin independent endocytic pathways (Benmerah et al., 1999). Vero cells were transfected with GFP‐tagged versions of the dominant negative mutant EH29 (GFP‐EH29) and the control construct that does not affect clathrin‐mediated uptake (GFP‐DIIIΔ2) (Benmerah et al., 1998). Twenty‐four hours post transfection, cells were infected with DENV‐1 or DENV‐2 for 1 h at 37°C and then processed to detect GFP expression and internalized viral antigen by indirect immunofluorescence staining. When Vero cells were transfected with the control construct GFP‐DIIIΔ2 both DENV‐1 and DENV‐2 virions were internalized in endocytic vesicles exhibiting a speckled virus antigen staining within the cytoplasm (Fig. 3D). A similar pattern was observed in mutant GFP‐EH29 transfected cells after DENV‐2 infection, indicating that DENV‐2 was capable of entering Vero cells when clathrin‐mediated uptake was blocked (Fig. 3D). In contrast, a slight and disperse red fluorescence at the cell surface, demonstrative of a blockade in virus entry, was detected in GFP‐EH29 transfected and DENV‐1‐infected cells, providing additional strong evidence that DENV‐1 internalization into Vero cells occurs via a clathrin‐dependent pathway. The functionality of the construct GFP‐EH29 was proved by an assay of transferrin incorporation in transfected cells. As expected, expression of the dominant negative protein, but not of the control, inhibited transferrin uptake (Fig. 3D).
To examine the effect of overexpression of Eps15 on productive infection, transfected cells were allowed to be infected during 24 h before proceeding to cell fixation and staining. Expression of the dominant negative mutant did not significantly affect infection with DENV‐2 as indicated by the similar signals for merge images in dominant and control transfected cells (Fig. 3E). The data for the 24 h infection were quantified by scoring cells positive for viral antigen in 250 cells with comparable levels of transgene expression. The DENV‐2‐positive cells represented 92.9% and 92.7% of transgene‐expressing cells in GFP‐EH29 and GFP‐DIIIΔ2 transfected cells respectively (Fig. 3F). When the same experimental procedure was performed after DENV‐1 infection, the ability of Vero cells to be productively infected with this serotype was blocked when clathrin dependent endocytosis was interrupted by the dominant negative mutant transfection, reducing the percentage of DENV‐1‐positive cells from 75.1% of GFP‐DIIIΔ2 transfected cells to 16.6% of GFP‐EH29 transfected cells (Fig. 3E and F). In conclusion, all these data confirm that DENV‐2 is capable of entering and causing a productive infection in Vero cells lacking a functional clathrin‐mediated endocytosis whereas the infection of Vero cells with DENV‐1 requires this endocytic pathway.
DENV‐1 and DENV‐2 entry into Vero cells is independent of lipid rafts/caveolae but dependent on dynamin
To better characterize the clathrin‐independent endocytic route used by DENV‐2 to enter Vero cells, our next approach was to analyse if any step during the entry process required the integrity of lipid rafts and/or caveolae. Lipid rafts are cholesterol‐ and sphingolipid‐rich membrane microdomains used by certain viruses and toxins to enter into the cell (Nichols and Lippincott‐Schwartz, 2001; Kirkham and Parton, 2005). Caveolae are vesicles arising from specialized lipid rafts and associated to the protein caveolin (Pelkmans, 2005). The raft/caveolae‐dependent pathways are highly sensitive to cholesterol‐depleting agents that disrupt the lipid bilayer structure whereas clathrin‐dependent endocytosis is only affected at high doses (Anderson et al., 1996; Damm et al., 2005).
To determine if any step in DENV‐2 entry before trafficking to the acidic vesicles was sensitive to perturbation of lipid rafts, we evaluated the effects on virus infection of nystatin and methyl‐β‐cyclodextrin. These sterol‐binding drugs sequester cholesterol by extraction, in the case of methyl‐β‐cyclodextrin, or by complex formation at the membrane, in the case of nystatin, blocking the lipid raft‐dependent entry pathways (Ilangumaran and Hoessli, 1988; Anderson et al., 1996; Pelkmans et al., 2001). Since this class of compounds can also modify the lipid bilayer of the virus envelope leading to virus inactivation, we first analysed the virucidal effect of nystatin and methyl‐β‐cyclodextrin by incubation of DENV‐1 and DENV‐2 suspensions with different concentrations of the compounds during 1 h at 37°C. Then, samples were diluted and remaining infectivity was titrated by PFU in Vero cells. The sample dilution effectively reduced the drug concentration to be incubated with the cells at least 100‐fold to assess that any titre reduction was only due to cell‐free virion inactivation. As seen in Fig. 4A, methyl‐β‐cyclodextrin exerted a dose‐dependent inactivating activity against DENV‐1 and DENV‐2 virions as a probable consequence of envelope alteration by its interaction with cholesterol. Nystatin exerted an inactivating effect against DENV something lower than methyl‐β‐cyclodextrin (data not shown). Based on these results, the treatment conditions to test the effects of the drugs on DENV infection were chosen to affect lipid raft organization of cell membrane but without a direct contact between drug and virions. Thus, Vero cells were pre‐incubated with increasing concentrations of nystatin or methyl‐β‐cyclodextrin. Then, monolayers were extensively washed and infected with DENV‐2 or DENV‐1, probed as a negative control, in the absence of compound. The pretreatment with the drugs neither affected DENV‐2 internalization (Fig. 4B) nor DENV‐2 production in Vero cells at 48 h p.i. (data not shown). As expected for a virus using clathrin‐mediated endocytosis (Fig. 3), DENV‐1 internalization was not affected by both inhibitors at the tested concentrations (Fig. 4B). The positive control for the effectiveness of cell treatment with these drugs on lipid raft/caveolae pathways was performed assaying the entry of FITC‐labelled cholera toxin, a marker of internalization through this route, after a similar treatment to that used for virus entry assay. Untreated cells showed a clear dotted perinuclear cytoplasmic fluorescence due to the accumulation of the toxin in the endoplasmic reticulum and Golgi apparatus; whereas the treatment with nystatin or methyl‐β‐cyclodextrin resulted in a diffuse green fluorescence pattern indicative of the impairment of toxin incorporation (Fig. 4C). The blockade of cholera toxin entry showed that functional lipid rafts were destroyed by drug treatment, while the lack of inhibitory effect on DENV‐1 infection indicated that the drug concentrations were not as high to affect other endocytic pathways, like the clathrin‐mediated endocytosis. Consequently, it is valid to conclude from these experiments with pharmacological inhibitors that DENV‐2 entry is independent of lipid rafts/caveolae.
Figure 4.
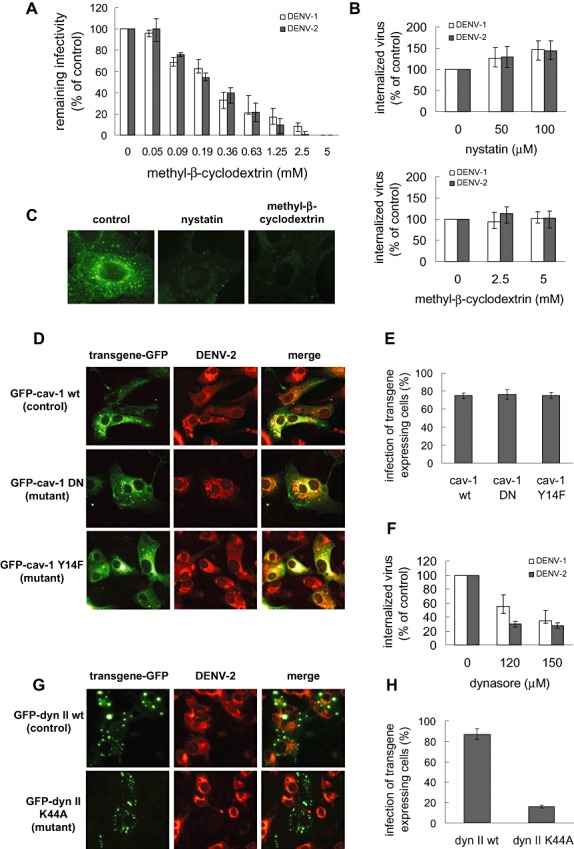
DENV‐2 entry into Vero cells is independent of caveolae/lipid‐rafts but is dependent on dynamin. A. DENV‐1 or DENV‐2 suspensions were incubated at 37°C with various concentrations of methyl‐β‐cyclodextrin. After 1 h remaining infectivity was determined. B. Vero cells were pretreated with nystatin or methyl‐β‐cyclodextrin. Then monolayers were washed with PBS and infected with DENV‐1 or DENV‐2 in culture medium without serum. After 1 h internalization, cultures were treated with proteinase K and the cell pellets were plated onto Vero cells to determine internalized virus by an infectious centre assay. Results are expressed as percentage of internalized virus with respect to a control without drug treatment. C. Cells were untreated (control) or treated with 100 μM nystatin or 5 mM methyl‐β‐cyclodextrin. Then cultures were incubated with FITC‐labelled cholera toxin. D. Cells transiently transfected with the constructs GFP‐cav‐1 wt, GFP cav‐1 DN or GFP‐cav‐1 Y14F were infected with DENV‐2. After 24 h infection cultures were fixed and immunofluorescence staining was performed. E. For quantification of samples shown in (D), 250 transfected cells with similar levels of GFP expression were screened and cells positive for viral antigen were scored. F. Vero cells were treated with dynasore, infected with DENV‐1 or DENV‐2 in the presence of the drug and then processed as in (B). G. Cells transiently transfected with the constructs GFP‐dyn II wt or GFP‐dyn II K44A were infected with DENV‐2. After 24 h infection cultures were fixed and immunofluorescence staining was performed. H. For quantification of samples shown in (G), 250 transfected cells with similar levels of GFP expression were screened and cells positive for viral antigen were scored. In (A), (B), (E), (F) and (H) values represent the mean ± SD of two independent experiments.
To address the lack of caveolae‐mediated endocytosis in DENV‐2 infection of Vero cells in a complementary manner, the impact of the expression of dominant negative mutants for caveolin‐1, the major structural protein of caveolae, was next studied. The fusion of GFP at the N‐terminal domain of caveolin‐1 sequence inhibits the functionality of this protein acting as a dominant negative mutant named GFP‐cav‐1 DN, while the GFP fusion at the C‐terminal end does not affect the protein (GFP‐cav‐1 wt) (Pelkmans et al., 2001). More recently, a new form of dominant negative mutant named GFP‐cav‐1 Y14F was generated by a point mutation that prevented caveolin‐1 phosphorylation (Coyne and Bergelson, 2006). Vero cells were transfected with the wild‐type and both mutant forms of caveolin‐1, and 24 h post‐transfection cells were infected with DENV‐2. After 24 h of infection, cells were fixed and analysed for GFP expression to quantify transfected cells and for immunofluorescence staining against DENV‐2 antigen to detect infected cells. As shown in Fig. 4D, the expression of the dominant negative mutants of caveolin GFP‐cav‐1 DN and GFP‐cav‐1 Y14F had no effect on the infection of cells with DENV‐2. The percentage of infection in transgene‐expressing cells, determined by scoring cells expressing viral antigens, was similar in cultures transfected with the three caveolin‐1 constructs (Fig. 4E), and thereby strengthened our conclusion that DENV‐2 entry was independent of caveolae‐mediated endocytosis. As expected, the dominant negative mutants of caveolin did not affect the entry of DENV‐1 into Vero cells (Fig. S2A and B).
Altogether, these results denote a route of infective entry for DENV‐2 into Vero cells through an endocytic pathway independent of clathrin and lipid rafts/caveolae. In recent years, certain viruses were reported to use similar routes not fully characterized (Sieczkarski and Whittaker, 2002b; Quirin et al., 2008; Rojek et al., 2008). In particular, the participation of dynamin is not clearly elucidated in non‐clathrin, non‐caveolar pathways (Conner and Schmid, 2003; Cao et al., 2007). Dynamin is a GTPase essential for pinching off vesicles in clathrin‐ and caveolae‐dependent endocytosis. To probe if dynamin is involved in DENV‐2 entry into Vero cells the effect of dynasore, a recently developed dynamin inhibitor (Macia et al., 2006), was evaluated. Although the infection of Vero cells with DENV‐2 was not clathrin‐ or caveolin‐dependent, treatment with dynasore before and during virus infection significantly reduced DENV‐2 internalization (Fig. 4F), as well as virus production at 48 h p.i. (data not shown). As a positive control, we determined that the internalization of DENV‐1 was inhibited by dynasore treatment (Fig. 4F). To confirm the participation of dynamin in DENV‐2 entry into Vero cells the GFP‐tagged versions of the wild‐type form of dynamin II GFP‐dyn II wt and the dominant negative mutant GFP‐dyn II K44A were used (Damke et al., 1994; Oh et al., 1998; Sauvonnet et al., 2005). Vero cells were transfected with both constructs and after 24 h of transfection cells were infected with DENV‐1 or DENV‐2. At 24 h p.i., cultures were processed for immunofluorescence. Cells expressing similar levels of GFP‐tagged wild‐type and mutant forms of dynamin were selected and those stained for viral antigen were scored. As seen in Fig. 4G, DENV‐2 multiplication was affected in cells expressing the dominant negative mutant, with percentages of infection in GFP‐dyn II wt and GFP‐dyn II K44A transfected cells of 87% and 16% respectively (Fig. 4H). The expression of mutant dynamin also significantly reduced the infection with DENV‐1 in the same cell line: in GFP‐dyn II wt transfected cells the percentage of infection was about 70% whereas in cells transfected with GFP‐dyn II K44A only 15% of cells were infected (Fig. S2C and D). Thus, we can conclude that the infective entry of both serotypes DENV‐1 and DENV‐2 into Vero cells is dependent on dynamin functionality.
Requirement of the cytoskeleton network: DENV‐2 entry to Vero cells is independent of macropinocytosis and phagocytosis
The non‐clathrin non‐caveolar endocytic pathway for DENV‐2 internalization into Vero cells was further characterized by investigating the possible involvement of the cellular cytoskeleton in this process. To address the role of actin network in DENV‐2 entry, we analysed the effects of two drugs, cytochalasin D and latrunculin A, which disturb microfilament integrity by actin depolimerization (Rojek et al., 2008), and a third compound, jasplakinolide, which acts by inducing a rigid stabilization of the actin polymer and a blockade of the microfilament dynamics (Rojek et al., 2008). No inhibition was observed on DENV‐2 multiplication after treatment with these compounds (Fig. 5A). The lack of effect of actin‐disturbing agents on DENV‐2 entry was also assessed in an internalization assay, as shown in Fig. 5C for cytochalasin D, although the effectiveness of the compound to destroy the typical pattern of the actin microfilaments as bundles of thin fibers was observed by staining with FITC‐labelled phalloidin (Fig. 5D). Then, data with the different types of inhibitors allowed conclude that the structure and dynamics of the actin microfilaments are not involved in the endocytic DENV‐2 entry to Vero cells. Additionally, these results represent the first evidence towards the probable exclusion of macropinocytosis or phagocytosis as possible mechanisms of DENV‐2 internalization. Although neither macropinocytosis nor phagocytosis is frequently involved in virus entry, they have been reported as an alternative access for some viruses in certain cells (Maréchal et al., 2001; Clement et al., 2006; Amstutz et al., 2008). These two well‐known endocytic routes for cell entry independent of clathrin and caveolae are strictly dependent of the F‐actin network (Swanson and Watts, 1995). We intended to obtain further evidence about the role of macropinocytosis by using amiloride, an inhibitor of the Na+/H+ exchange at the plasma membrane (Swanson and Watts, 1995), but this compound exerted virucidal activity against DENV‐2 inactivating cell‐free virions (data not shown).
Figure 5.
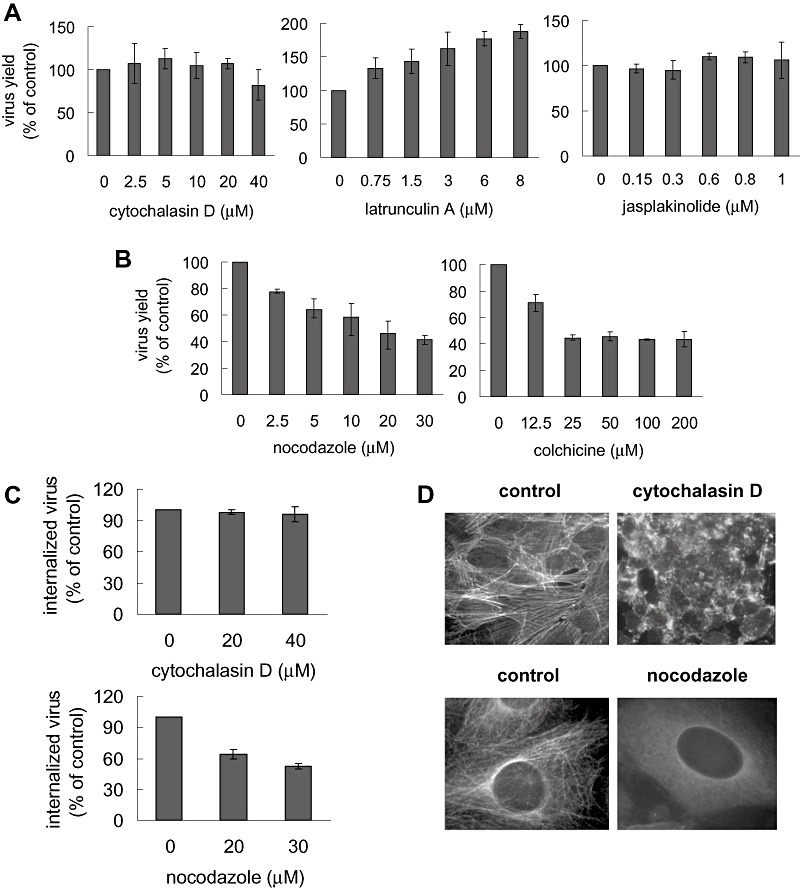
Effect of cytoskeleton‐disrupting agents on DENV‐2 entry into Vero cells. A and B. Cells were treated with increasing concentrations of cytochalasin D, latrunculin A, jasplakinolide, nocodazole or colchicine and infected with DENV‐2. Virus yields were determined at 48 h p.i. Results are expressed as percentage of virus multiplication with respect to a control without drug treatment. Each point shows the mean ± SD of two independent experiments. C. Cells were treated with cytochalasin D or nocodazole and infected with DENV‐2. After 1 h internalization in the presence of the drugs, monolayers were treated with proteinase K and the cell pellets were plated onto Vero cells to determine internalized virus by an infectious centre assay. D. Cells were untreated (control) or treated with 40 μM cytochalasin D or 30 μM nocodazole and then stained with FITC‐labelled phalloidin or mouse anti‐tubulin antibody and TRITC‐labelled anti‐mouse IgG.
The functionality of the microtubule cytoskeleton for DENV‐2 infection of Vero cells was studied by cell pretreatment with nocodazole and colchicine, both inhibitors of microtubule assembly through binding to tubulin (Hamel, 1996). The effect of these compounds on microtubule organization was verified using a monoclonal antibody to tubulin. The staining pattern observed as a network of tubules radiating from the perinuclear region to the cell periphery was transformed, after treatment with 30 μM nocodazole (Fig. 5D) or 200 μM colchicine (not shown), into an arrangement of condensed amorphous aggregates in the cytoplasm. The destabilization of the microtubule network by nocodazole or colchicine treatment during virus entry caused a significant reduction in virus production (Fig. 5B) as well as in virus internalization (Fig. 5C), suggesting the requirement of the structural integrity of the microtubule network for efficient DENV‐2 entry to Vero cells.
DENV‐2 entry to human A549 cells is mediated by clathrin‐dependent endocytosis
To compare the route of entry of DENV‐2 in other type of mammalian cells different from Vero, the infection of the human cell line A549 in the presence of pharmacological inhibitors of endocytosis was also assayed. The treatment of A549 cells with ammonium chloride, chlorpromazine, dansylcadaverine and nystatin was performed in the same conditions of incubation time and drug concentrations as previously described for Vero cells. The assayed range of concentrations was sufficient to induce essentially the same effects on the endocytic pathway in A549 cells in comparison to Vero cells, as determined by the corresponding staining markers (Fig. S3). After drug treatment, cells were infected with DENV‐2 and further incubated for 48 h, when supernatants were titrated by plaque formation on Vero cells. A dose‐dependent and significant reduction in DENV‐2 infection was detected in A549 cells when endosomal pH was increased by ammonium chloride pretreatment, indicating that a low pH‐dependent endocytic route of entry is also used by DENV‐2 in this mammalian cell line (Fig. 6A). A similar level of dose‐dependent inhibition of DENV‐2 infection was determined in A549 cells treated with both inhibitors of clathrin‐mediated endocytosis, chlorpromazine and dansylcadaverine (Fig. 6B and C). Accordingly, lipid rafts/caveolae were not required for the infection of A549 cells with DENV‐2 since no inhibitory effect was detected by nystatin treatment (Fig. 6D). These results indicate that the entry of DENV‐2 into A549 cells was dependent on clathrin‐mediated endocytosis and, consequently, differed from the pathway followed by the same viral serotype in Vero cells, as shown in Fig. 3.
Figure 6.
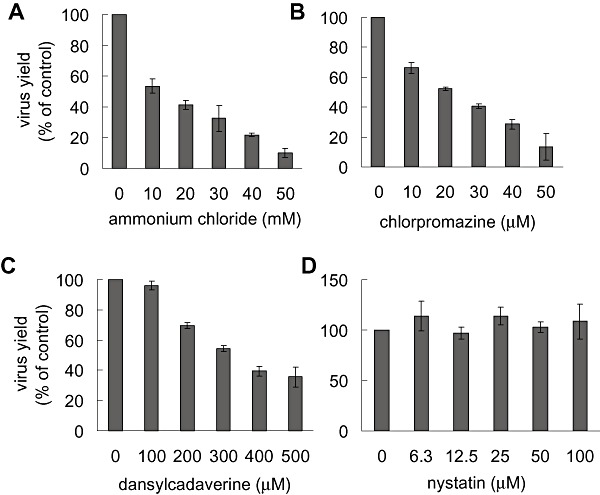
DENV‐2 infection of A549 cells is dependent on clathrin‐mediated endocytosis. A549 cells were treated with various concentrations of ammonium chloride (A), chlorpromazine (B), dansylcadaverine (C) or nystatin (D) and infected with DENV‐2. Virus yields were quantified at 48 h p.i. Results are expressed as percentage of virus multiplication with respect to a control without drug treatment. Each point shows the mean ± SD of two independent experiments.
Discussion
The study of the internalization pathway of two DENV serotypes reported here clearly shows the complexity of the mode of entry of this viral pathogen into vertebrate host cells. Our results conclusively show for the first time a differential mode of infective entry for the serotypes DENV‐1 and DENV‐2, into a common host cell, Vero cells, as well as alternative entry pathways for a given serotype, DENV‐2, into different types of cells.
The analysis of the effects of biochemical and molecular inhibitors demonstrated that DENV‐1 utilizes a clathrin‐mediated pathway of endocytosis whereas the clathrin‐coated vesicles were not required for the entry of DENV‐2 virions into Vero cells. In addition, infection assays in the presence of entry inhibitors quantitatively confirmed the results from the internalization assays, showing a concomitance in the levels of virus yields or viral antigen expressing cells for DENV‐1 and DENV‐2 with the proposed route of entry. Interestingly, a consistent and significative increase in DENV‐2 yield was observed when infection was performed in the presence of the two inhibitors of clathrin‐mediated endocytosis, chlorpromazine and dansylcadaverine. One possibility is that the clathrin pathway would be a non‐infective route of entry for DENV‐2 in Vero cells and, consequently, when it is blocked there is an enhancement in the utilization by DENV‐2 virions of the infective non‐clathrin pathway, leading to a higher production of infectious virus. Further research is required to ascertain this hypothesis or any other one to explain the results shown in Fig. 3A and B. In comparison, it has recently been shown that papillomaviruses could be internalized without inducing productive infection (Selinka et al., 2007) and, concomitantly, the inhibition of clathrin‐and caveolae‐mediated endocytosis led to an increase of human papillomavirus infectivity through a proposed alternative endocytic mechanism (Spoden et al., 2008). Therefore, the increased infectivity of DENV‐2 in cells with disturbed clathrin endocytosis may represent an additional support for usage of the non‐classical route.
The action of different agents perturbing the major clathrin‐independent endocytic processes on DENV‐2 infection of Vero cells suggested that neither caveolae nor lipid rafts appeared to be involved in virus entry. In apparent contradiction to our results, Reyes‐del Valle et al. (2005) and Lee et al. (2008) reported that disruption of lipid rafts by methyl‐β‐cyclodextrin depletion of cholesterol decreased DENV‐2 infection in human peripheral blood monocytes and mouse neuroblastoma N18 cells respectively. By contrast and in accordance with our results, Umashankar et al. (2008) demonstrated efficient membrane fusion by DENV‐2 in cholesterol‐depleted insect cells. These contrasting results could reflect variations in the properties of endocytosis or requirements for cholesterol during virus internalization among different cells as well as differences in treatment conditions to achieve cholesterol depletion and time of infection.
In the present study, we were especially interested in dissecting whether cholesterol is important as a constituent of virus membrane, host cell membrane or both structures. To address this question, we used methyl‐β‐cyclodextrin to treat separately either DENV‐2 suspension or Vero cells prior to virus infection, and experimental conditions (concentration and time of exposure to the drug) were optimized to avoid a general perturbation of membrane properties. Clearly, our studies demonstrate that the drug‐induced release of cholesterol from virion membranes correlated with a dramatic loss of infectivity whereas cholesterol depletion of cell membranes did not affect subsequent virus infection. Thus, this virucidal activity of cholesterol depleting drugs, also reported for other flaviviruses (Aizaki et al., 2008), may induce a false inference about the role of cellular cholesterol‐rich microdomains for fusion and infection. In fact, the abovementioned reports of Reyes‐del Valle et al. (2005) and Lee et al. (2008) did not analyse the virucidal activity of methyl‐β‐cyclodextrin against DENV‐2. At present we do not know how the presence of cholesterol in the lipid viral envelope, acquired during the budding process, affects virion infectivity. It will be interesting in future studies to analyse at what stage the virus multiplication cycle is blocked after infection with methyl‐β‐cyclodextrin‐inactivated DENV virions.
We have also demonstrated, by using the K44A mutant and the inhibitor dynasore, that DENV‐2 entry is dependent on dynamin. Dynamin activity was initially thought to be specific to clathrin‐mediated endocytosis (Damke et al., 1994). But it is now evident that this GTPase is functional in a variety of endocytic vesicle scission events independent of clathrin (Henley et al., 1998; Gold et al., 1999; Pelkmans et al., 2002).
The sum of all our results demonstrate that the entry route of DENV‐2 to Vero cells is a non‐classical endocytic pathway independent of clathrin, caveolae and lipid rafts, but dependent on dynamin, differing from the clathrin‐mediated route used by DENV‐1 in the same cells and by DENV‐2 in other cell systems, as shown here for A549 cells (Fig. 6) and in recent reports for HeLa, C6/36 and BSC‐1 cells (Krishnan et al., 2007; Acosta et al., 2008b; Mosso et al., 2008; van der Schaar et al., 2008). Figure 7 shows the multiple portals of entry available in eukaryotic cells to be hijacked by viruses, indicating those used by DENV‐1 and DENV‐2, according to the present knowledge. There are four well‐known routes (phagocytosis, macropinocytosis, clathrin‐ and caveolae‐mediated endocytosis) whereas the other non‐clathrin‐ and non‐caveolar‐mediated pathways, with or without the participation of lipid rafts and dynamin, are less characterized. However, studies performed in recent years have increased the number of viruses involved in these non‐classical entry pathways, including influenza virus (Sieczkarski and Whittaker, 2002b), lymphocytic choriomeningitis virus (Quirin et al., 2008; Rojek et al., 2008), human immunodeficiency virus (Vidricaire and Tremblay, 2007), rotavirus (Sánchez‐San Martín et al., 2004), human papillomavirus (Spoden et al., 2008) and coronaviruses (Van Hamme et al., 2008; Wang et al., 2008). In all cases, the internalization pathways need further characterization to compare them and establish similarities and differences.
Figure 7.
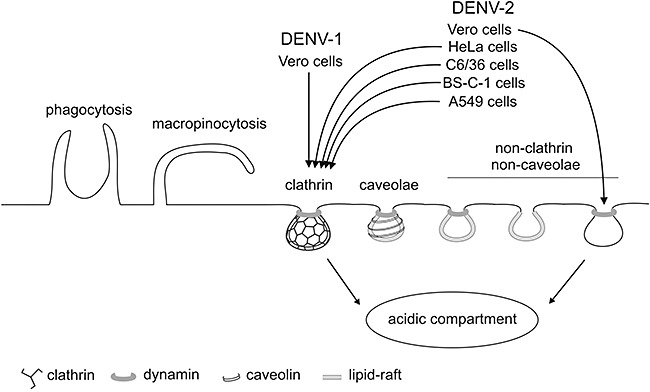
Endocytic pathways utilized by DENV‐1 and DENV‐2 to infect different cell lines. DENV‐1 infection of Vero cells occurs through a classical clathrin‐mediated dynamin‐dependent endocytosis, while infection of this same cell line by DENV‐2 takes place through a non‐classical endocytic pathway independent of clathrin, caveolin‐1 and lipid‐rafts, but dependent on dynamin. This entry route differs from the clathrin‐mediated pathway followed by DENV‐2 to infect A549 cells and, as previously shown, to infect C6/36 cells (Acosta et al., 2008b; Mosso et al., 2008), HeLa cells (Krishnan et al., 2007) and BS‐C‐1 cells (van der Schaar et al., 2008).
Furthermore, the use of alternative entry routes in different cells by a given virus, as appears to occur with DENV‐2, may provide an advantage by improving the chances of productive virus entry and expanding the host cell range of infection. There may be cell type differences in the relative action or abundance of the endocytic pathways discussed here, and such differences may have effects on the preferred route of entry for virus. Precisely, the wide spectrum of cell tropism is a well‐known property of DENV that may be related to the possibility here shown to exploit optional routes of intracellular transport.
At present, the mechanistic basis of the different clathrin‐dependence between DENV‐1 and DENV‐2 for entry into Vero cells cannot be fully understood. A different receptor usage may determine the consequential route of entry. A comparative study for DENV‐1 and DENV‐2 receptor in Vero cells has not been performed, but in human liver HepG‐2 cells two different proteins, the laminin receptor and GRP78, were identified as DENV‐1 and DENV‐2 specific receptors respectively (Jindadamrongwech et al., 2004; Thepparit and Smith, 2004), and also a higher saturation of infection was found for DENV‐1 respect of DENV‐2 (Suksanpaisan and Smith, 2003). Another factor that may affect DENV entry is the presence of sugar molecules on the viral surface. The utilization of the two potential N‐linked glycosylation sites of E (Asn‐67 and Asn‐153) has been determined for only a limited number of strains and the heterogeneity among circulating virus populations remains uncharacterized (Bryant et al., 2007). Diverse phenotypic changes, such as fusion activity, tropism, virulence and morphogenesis, have been associated with the lack of glycosylation at one or both sites, with a variable behavior in mosquito or mammalian cells (Guirakhoo et al., 1993; Johnson et al., 1994; Bryant et al., 2007; Mondotte et al., 2007). Further studies are needed to reveal if E glycosylation is related to the endocytic via utilized for DENV entry. Finally, after binding, both serotype virions need to be exposed at intravesicular low pH for functional entry (Fig. 2), in accordance with the conformational transition reported for DENV‐2, DENV‐3 and DENV‐1 E glycoprotein under acidic conditions that allow membrane fusion (Kuhn et al., 2002; Modis et al., 2004; Stiasny and Heinz, 2006; Nayak et al., 2009). However, differences between the DENV‐1 and DENV‐2 E post‐fusion conformations were reported (Nayak et al., 2009). Interestingly, van der Schaar et al. (2008) found distinct requirements in the endocytic trafficking for two strains of the serotype DENV‐2 in BSC‐1 cells: by real‐time imaging strain S1 particles fused from within Rab7‐positive late endosomes whereas virions of strain NGC fused from Rab5‐positive early endosomes. This may reflect a different pH threshold for fusion, a hypothesis that must be also evaluated among DENV serotypes. van der Schaar et al. (2008) also reported that the majority of DENV‐2 particles had fused from acidic vesicles within 17 min p.i., in accordance with our results showing inhibition of DENV‐1 and DENV‐2 entry after 1 h of infection by the pH‐rising effect of ammonium chloride and concanamycin A (Fig. 2A). Future experiments will allow to locate the precise vesicular pH for fusion of both DENV serotypes by monitoring the kinetics of low pH‐dependence for entry into Vero cells and the kind of acidic organelle where fusion takes place.
An understanding of DENV entry into cells will not only extend our understanding of entry mechanism but also help to exploit new targets for antiviral therapy. The blockade of DENV entry into the host cell is an interesting antiviral strategy because it represents a barrier to suppress the beginning of infection (Altmeyer, 2004; Perera et al., 2008). In particular, the studies about the anti‐DENV activity of sulfated polysaccharides indicated that virus binding and internalization were blocked by this class of compounds (Talarico and Damonte, 2007; Talarico et al., 2007). A differential susceptibility of DENV serotypes to polysulfates was observed: DENV‐2 was the most susceptible serotype whereas DENV‐1 showed very weak inhibition (Lin et al., 2002; Talarico et al., 2005; Talarico and Damonte, 2007). These variations may be ascribed to the differences in virus–cell interactions during entry of DENV‐1 and DENV‐2 into Vero cells here reported. The simultaneous cocirculation of the four DENV serotypes in diverse regions from Asia and America makes mandatory the requirement for a potential antiviral agent to be reactive againtst all serotypes. Therefore, the elucidation of the precise mechanism of internalization for DENV serotypes will provide the opportunity to identify the adequate target for development of effective therapeutic agents.
Experimental procedures
Cells and viruses
The cell lines Vero (African green monkey kidney) and A549 (lung carcinoma human cells) were grown in Eagle's minimum essential medium (MEM) (Gibco) supplemented with 5% fetal bovine serum. For maintenance medium (MM), the serum concentration was reduced to 1.5%. The C6/36 HT mosquito cell line from Aedes albopictus, adapted to grow at 33°C, was cultured in L‐15 Medium (Leibovitz) supplemented with 0.3% tryptose phosphate broth, 0.02% glutamine, 1% MEM non‐essential amino acids solution and 5% fetal bovine serum.
The stocks of DENV‐1, strain Hawaii, and DENV‐2, strain NGC, were prepared in C6/36 HT cells and titrated by plaque formation in Vero cells.
Antibodies and reagents
The anti‐DENV antibody was a mouse monoclonal antibody reactive against the E glycoprotein of the four dengue serotypes purchased from Abcam (Cambridge, UK). Monoclonal antibody to tubulin, goat anti‐mouse IgG conjugated to fluorescein isothiocyanate (FITC) or rhodamine (TRITC), FITC‐phalloidin, and FITC‐cholera toxin B subunit were purchased from Sigma‐Aldrich (USA). TRITC‐human transferrin was from Molecular Probes (USA).
Dansylcadaverine, chlorpromazine, nystatin, methyl‐β‐cyclodextrin, ammonium chloride, concanamycin A, dynasore, colchicine, nocodazole, cytochalasin D, latrunculin A, jasplakinolide, acridine orange and 3‐(4,5‐dimethylthiazol‐2‐yl)‐2,5‐diphenyl tetrazolium bromide (MTT) were purchased from Sigma‐Aldrich (USA).
Kinetics of virus internalization
Vero cells were infected with DENV‐1 or DENV‐2 at a multiplicity of infection (moi) of 1 PFU cell−1 at 4°C (synchronized infection). After 1 h adsorption cells were washed with ice‐cold PBS, covered with MM and incubated at 37°C. At different times cells were washed with PBS and treated with proteinase K (1 mg ml−1, Invitrogen) for 45 min at 4°C to remove adsorbed but not internalized virus. Then, proteinase K was inactivated with 2 mM PMSF in PBS with 3% bovine seroalbumin (BSA), and cells were washed with PBS 0.2% BSA by low speed centrifugation. Finally, pellets were resuspended in MM and different serial dilutions of the cell suspensions were plated onto Vero cell monolayers and covered with MM containing 1% methylcellulose. Infectious centres were counted after 6–7 days according to virus serotype. To determine the rate of viral internalization a parallel set of cultures was processed in the same conditions except that proteinase K was replaced by PBS. For each time point the number of plaques formed in the PBS controls was considered 100%. To determine the kinetics of virus internalization in a non‐synchronized infection, the same experimental procedure was performed but excluding the 4°C incubation step.
Inhibition of DENV infection and internalization by pharmacological inhibitor treatment
The entry mechanism of DENV‐1 and DENV‐2 was studied evaluating the effect of inhibitors of different endocytic pathways. The time of treatment and range of concentrations adequate for each inhibitor were first determined by a cell viability assay in order to avoid any toxic effects. Monolayers of Vero and A549 cells grown in 24‐well microplates were incubated in MM with or without compound, and cell viability was measured after each treatment using the MTT assay, as described previously (Talarico et al., 2005). According to the viability data obtained (not shown), the treatment conditions for each inhibitor were chosen as follows: for chlorpromazine (10–50 μM), concanamycin A (3–50 nM), colchicine (12.5–200 μM), nocodazole (2.5–30 μM) and cytochalasin D (2.5–40 μM) cells were pretreated for 2 h at 37°C; for ammonium chloride (10–50 mM), methyl‐β‐cyclodextrin (0.62–5 mM in MM without serum) and dynasore (20–150 μM) cells were pretreated for 1 h at 37°C; for dansylcadaverine (100–500 μM), latrunculin A (0.62–8 μM) and jasplakinolide (0.15–1 μM), 30 min of pretreatment at 37°C; and for nystatin (6.3–100 μM in MM without serum) cells were pretreated for 5 h at 37°C. After pretreatment, cells were infected with DENV‐1 or DENV‐2 at an moi of 0.1 PFU cell−1 in the presence of the drug, except for nystatin and methyl‐β‐ciclodextrin where pretreated cultures were intensely washed with PBS before infection. In all cases, virus inocula were removed after 1 h of infection at 37°C, and then cultures were washed with PBS and further incubated at 37°C in MM without compound. At 48 h p.i., supernatants were harvested to determine extracellular virus yields by plaque assay. In order to quantify productive internalized virus particles Vero cells were treated with the drugs in the conditions mentioned above and then were infected with DENV‐1 or DENV‐2 at an moi of 1 PFU cell−1 in the presence or absence of the compound. After 1 h at 37°C cells were washed with PBS, treated with proteinase K and processed as abovementioned.
To assess the effect of ammonium chloride and concanamycin A on the pH of acid intracellular vesicles, Vero or A549 cells, treated or not with the compound during 1 h at 37°C, were stained with acridine orange (1 μg ml−1 in MM without serum) for 15 min at 37°C. Then, cells were washed twice with PBS, mounted on PBS and visualized on a fluorescence microscope Olympus BX51.
Transfections
The EGFP‐tagged constructs of Eps15 GFP‐EH29 (dominant negative mutant) and GFP‐DIIIΔ2 (control) were kindly provided by Dra. C. Shayo (IBYME, Argentina). The EGFP‐tagged constructs of wild‐type caveolin‐1 GFP‐cav‐1 wt and dominant negative mutants GFP‐cav‐1 DN and GFP‐cav‐1 Y14F were kindly provided by Dr J. M. Bergelson (University of Pennsylvania, USA). EGFP‐tagged wild‐type dynamin II GFP‐dyn II wt and dominant negative mutant GFP‐dynII‐K44A were kindly provided by Dr M. A. McNiven (Mayo Clinic, USA). Vero cells grown on cover slips until 70% confluency, were transfected with each construct using Lipofectamine 2000 reagent (Invitrogen, USA) as previously described (Acosta et al., 2008b). Briefly, 4 μg of each construct was diluted in 50 μl Opti‐MEM and combined with 50 μl Opti‐MEM containing 2.5 μl lipofectamine. After 40 min of incubation at room temperature, the DNA–liposome complexes were added to the cells and cultures were incubated for 6 h at 37°C. At this time, medium was replaced by MM and cells were incubated until 24 h p.i.
For visualization of internalized viral particles, 24 h‐transfected cells cultures were infected with DENV‐1 or DENV‐2 at an moi of 10 PFU cell−1 and after 1 h of infection cells were fixed with methanol for 10 min at −20°C. For visualization of viral antigen production 24 h‐transfected cell cultures were infected with DENV‐1 or DENV‐2 at an moi of 1 PFU cell−1 and after 24 h infection cultures were fixed as described above.
Immunofluorescence assays
After methanol fixation, cells were washed with PBS and stained for DENV internalization or multiplication with a monoclonal antibody against E glycoprotein followed by TRITC‐labelled goat anti‐mouse IgG. After a final washing with PBS, cells were mounted in a glycerol solution containing 1,4‐diazabicyclo[2,2,2]octane (DABCO). The percentage of infection of transgene‐expressing cells was calculated by scoring the number of cells positive for viral antigen from approximately 250 transfected cells with comparable levels of transgene expression.
To visualize the actin filaments, compound‐treated and untreated Vero cells were washed with PBS, fixed with 4% paraformaldehyde and permeabilized with 0.2% Triton X‐100. FITC‐conjugated phalloidin was used for 1 h at room temperature as a probe of actin filaments. Then, cells were washed and observed. To visualize the microtubule network, compound‐treated and untreated Vero cells were washed with PBS and fixed with methanol for 10 min at −20°C. Then, cells were incubated with mouse anti‐α‐tubulin for 1 h at 37°C. After washing with PBS, the primary antibody was recognized by anti‐mouse TRITC antibody for 1 h at 37°C. Then cells were washed, mounted and visualized.
Internalization assays with control ligands
To assess the effectiveness of drug treatment or dominant negative mutant expression transferrin or cholera toxin uptake studies were performed. Briefly, cells treated with clorpromazine or dansylcadaverine, or cultures transfected with Eps15 constructs were incubated with TRITC‐transferrin (15 μg ml−1) for 1 h at 37°C. Then cells were washed with PBS to remove any uninternalized ligand and fixed with methanol for 10 min at −20°C. For nystatin or methyl‐β‐cyclodextrin treatments, pretreated cell cultures were washed and incubated with FITC‐cholera toxin (0.3 μg ml−1) for 1 h at 37°C. Then cultures were washed with PBS and fixed with methanol as described above. Cells were visualized under a fluorescence microscope Olympus BX51.
Virucidal assay
A DENV‐2 or DENV‐1 suspension containing 1 × 106 PFU ml−1 was incubated with an equal volume of MM with o without different concentrations of compound for 1 h at 37°C. The samples were then chilled, diluted in MM and residual infectivity was determined by plaque formation.
Virus adsorption assay
To address any inhibitory effect of ammonium chloride or concanamycin A on virus adsorption, Vero cells were pretreated with the drugs and then infected with DENV‐2 at an moi of 1 PFU cell−1 in the presence or absence (control) of the drugs. After 1 h adsorption at 4°C, cells were washed with cold PBS to remove unadsorbed virus and cells were disrupted by two cycles of freezing and thawing. The amount of infectious bound virus was then measured by plaque formation.
Supporting information
Fig. S1. Effect of ammonium chloride on DENV multiplication, viral particles infectivity, virus adsorption and intracellular vesicle acidification. A. Cells were treated with increasing concentrations of ammonium chloride and infected with DENV‐1 orDENV‐2. Virus yields were quantified by plaque assay at 48 h p.i. B. Suspensions ofDENV‐2 virions were incubated at 37°C with various concentrations of ammonium chloride. After 1 h samples were diluted and remaining infectivity was determined by plaque formation assay. C. Vero cells were treated as in (A) and infected with DENV‐2 at 4°C. After 1 h adsorption in the presence or absence of the drug cells were disrupted and cell‐bound virus was determined by plaque formation assay. In (A), (B) and (C) results are expressed as percentage of virus multiplication with respect to a control without drug treatment. Each point shows the mean ± SD of two independent experiments. D. Cells were left untreated (control) or treated with 50 mM ammonium chloride or 50 nM concanamycin A. Then, samples were stained with acridine orange.
Fig. S2. DENV‐1 infection of Vero cells is independent of caveolae but dependent on dynamin. A. Cells transiently transfected with the constructs GFP‐cav‐1 wt, GFP cav‐1 DN or GFP‐cav‐1 Y14F were infected with DENV‐1. After 24 h infection cultures were fixed and immunofluorescence staining was performed. B. For quantification of samples shown in (A), 250 transfected cells with similar levels of GFP expression were screened and cells positive for viral antigen were scored. C. Cells transiently transfected with the constructs GFP‐dyn II wt or GFP‐dyn II K44A were infected withDENV‐1. After 24 h infection cultures were fixed and immunofluorescence staining was performed. D. For quantification of samples shown in (C), 250 transfected cells with similar levels of GFP expression were screened and cells positive for viral antigen were scored. In (B) and (D) results are expressed as the mean ± SD of two independent experiments.
Fig. S3. Effect of different inhibitors on A549 cells endocytic processes. A549 cells were left untreated (control) or treated with ammonium chloride 50 mM (A), chlorpromazine 50 μM (B) or nystatin 100 μM (C). Then cells were stained with acridine orange (A), or incubated with TRITC‐labelled transferrin (B) or FITC‐labelled cholera toxin (C).
Please note: Wiley‐Blackwell are not responsible for the content or functionality of any supporting materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
Supporting info item
Acknowledgements
We thank Dra. C. Shayo (IBYME, Argentina), Dr J.M. Bergelson (University of Pennsylvania, USA) and Dr M.A. McNiven (Mayo Clinic, USA) for their kind contributions of plasmid constructs. This work was funded by grants from Agencia Nacional para la Promoción Científica y Tecnológica (ANPCyT), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) and Universidad de Buenos Aires. E.B.D. is member of Research Career from CONICET and E.G.A. is a Fellow of the same institution.
References
- Acosta, E.G. , Talarico, L.B. , and Damonte, E.B. (2008a) Cell entry of dengue virus. Future Virol 3: 471–479. [Google Scholar]
- Acosta, E.G. , Castilla, V. , and Damonte, E.B. (2008b) Functional entry of dengue virus into Aedes albopictus mosquito cells is dependent on clathrin‐mediated endocytosis. J Gen Virol 89: 474–484. [DOI] [PubMed] [Google Scholar]
- Aizaki, H. , Morikawa, K. , Fukasawa, M. , Hara, H. , Inoue, Y. , Tani, H. , et al. (2008) Critical role of virion‐associated cholesterol and sphingolipid in hepatitis C virus infection. J Virol 82: 5715–5724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altmeyer, R. (2004) Virus attachment and entry offer numerous targets for antiviral therapy. Curr Pharm Des 10: 3701–3712. [DOI] [PubMed] [Google Scholar]
- Amstutz, B. , Gastaldelli, M. , Kälin, S. , Imelli, N. , Boucke, K. , Wandeler, E. , et al. (2008) Subversion of CtBP1‐controlled macropinocytosis by human adenovirus serotype 3. EMBO J 27: 956–969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson, H.A. , Chen, Y. , and Norkin, L.C. (1996) Bound simian virus 40 translocates to caveolin‐enriched membrane domains, and its entry is inhibited by drugs that selectively disrupt caveolae. Mol Biol Cell 7: 1825–1834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balmaseda, A. , Hammond, S.N. , Pérez, L. , Tellez, Y. , Saborío, S.I. , Mercado, J.C. , et al. (2006) Serotype‐specific differences in clinical manifestations of dengue. Am J Trop Med Hyg 74: 449–456. [PubMed] [Google Scholar]
- Benmerah, A. , Lamaze, C. , Bègue, B. , Schmid, S.L. , Dautry‐Varsat, A. , and Cerf‐Bensussan, N. (1998) AP‐2/Eps 15 interaction is required for receptor‐mediated endocytosis. J Cell Biol 140: 1055–1062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benmerah, A. , Bayrou, M. , Cerf‐Bensussan, N. , and Dautry‐Varsat, A. (1999) Inhibition of clathrin‐coated pit assembly by an Eps15 mutant. J Cell Sci 112: 1303–1311. [DOI] [PubMed] [Google Scholar]
- Bryant, J.E. , Calvert, A.E. , Mesesan, K. , Crabtree, M.B. , Volpe, K.E. , Silengo, S. , et al. (2007) Glycosylation of the dengue 2 virus E protein at N67 is critical for virus growth in vitro but not for growth in intrathoracically inoculated Aedes aegytpti mosquitoes. Virology 366: 415–423. [DOI] [PubMed] [Google Scholar]
- Cao, H. , Chen, J. , Awoniyi, M. , Henley, J.R. , and McNiven, M.A. (2007) Dynamin 2 mediates fluid‐phase micropinocytosis in epithelial cells. J Cell Sci 120: 4167–4177. [DOI] [PubMed] [Google Scholar]
- Castilla, V. , Mersich, S.E. , Candurra, N.A. , and Damonte, E.B. (1994) The entry of Junin virus into Vero cells. Arch Virol 136: 363–374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu, J.J.H. , and Ng, M.L. (2004) Infectious entry of West Nile virus occurs through a clathrin‐mediated endocytic pathway. J Virol 78: 10543–10555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu, J.J.H. , Leong, P.W.H. , and Ng, M.L. (2006) Analysis of the endocytic pathway mediating the infectious entry of mosquito‐borne flavivirus West Nile into Aedes albopictus mosquito (C6/36) cells. Virology 349: 463–475. [DOI] [PubMed] [Google Scholar]
- Clement, C. , Tiwari, V. , Scanlan, P.M. , Valyi‐Nagy, T. , Yue, B.Y.J.T. , and Shukla, D. (2006) A novel role for phagocytosis‐like uptake in herpes simplex virus entry. J Cell Biol 174: 1009–1021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conner, S.D. , and Schmid, S.L. (2003) Regulated portals of entry into the cell. Nature 422: 37–44. [DOI] [PubMed] [Google Scholar]
- Coyne, C.B. , and Bergelson, J.M. (2006) Virus‐induced Abl and Fyn kinase signals permit coxsackievirus entry through epithelial tight junctions. Cell 124: 119–131. [DOI] [PubMed] [Google Scholar]
- Damke, H. , Baba, T. , Warnock, D.E. , and Schmid, S.L. (1994) Induction of mutant dynamin specifically blocks endocytic coated vesicle formation. J Cell Biol 127: 915–934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Damm, E.‐M. , Pelkmans, L. , Kartenbeck, J. , Mezzacasa, A. , Kurzchalla, T. , and Helenius, A. (2005) Clathrin‐ and caveolin‐1‐independent endocytosis: entry of simian virus 40 into cells devoid of caveolae. J Cell Sci 168: 477–488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diamond, M.S. , Edgil, D. , Roberts, T.G. , Lu, B. , and Harris, E. (2000) Infection of human cells by dengue virus is modulated by different cell types and viral strains. J Virol 74: 7814–7823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gold, E.S. , Underhill, D.M. , Morrissette, N.S. , Guo, J. , McNiven, M.A. , and Aderem, A. (1999) Dynamin 2 is required for phagocytosis in macrophages. J Exp Med 190: 1849–1856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gubler, D.J. (2002) Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends Microbiol 10: 100–103. [DOI] [PubMed] [Google Scholar]
- Guinea, R. , and Carrasco, L. (1994) Concanamycin A: a powerful inhibitor of enveloped animal‐virus entry into cells. Biochem Biophys Res Commun 201: 1270–1278. [DOI] [PubMed] [Google Scholar]
- Guirakhoo, F. , Hubt, A.R. , Lewis, J.G. , and Roehrig, J.T. (1993) Selection and partial characterization of dengue 2 virus mutants that induce fusion at elevated pH. Virology 194: 219–223. [DOI] [PubMed] [Google Scholar]
- Hamel, E. (1996) Antimitotic natural products and their interaction with tubulin. Med Res Rev 16: 207–231. [DOI] [PubMed] [Google Scholar]
- Hase, T. , Summers, P.L. , and Eckels, K.H. (1989) Flavivirus entry into cultured mosquito cells and human peripheral blood monocytes. Arch Virol 104: 129–143. [DOI] [PubMed] [Google Scholar]
- Henley, J.R. , Krueger, E.W.A. , Oswald, B.J. , and McNiven, M.A. (1998) Dynamin‐mediated internalization of caveolae. J Cell Biol 141: 85–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ilangumaran, S. , and Hoessli, D.C. (1988) Effects of cholesterol depletion by cyclodextrin on the sphingolipid microdomains of the plasma membrane. Biochem J 335: 433–440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jindadamrongwech, S. , Thepparit, C. , and Smith, D.R. (2004) Identification of GRP 78 (BiP) as a liver cell expressed receptor element for dengue virus serotype 2. Arch Virol 149: 915–927. [DOI] [PubMed] [Google Scholar]
- Johnson, A.J. , Guirakhoo, F. , and Roehrig, J.T. (1994) The envelope glycoproteins of dengue 1 and dengue 2 viruses grown in mosquito cells differ in their utilization of potential glycosylation sites. Virology 203: 241–249. [DOI] [PubMed] [Google Scholar]
- Kirkham, M. , and Parton, R.G. (2005) Clathrin‐independent endocytosis: new insights into caveolae and non‐caveolar lipid raft carriers. Biochem Biophys Acta 1745: 273–286. [DOI] [PubMed] [Google Scholar]
- Kliks, S.C. , Nisalak, A. , Brandt, W.E. , Wahl, L. , and Burke, D.S. (1989) Antibody‐dependent enhancement of dengue virus growth in human monocytes as a risk factor for dengue hemorrhagic fever. Am J Trop Med Hyg 40: 444–451. [DOI] [PubMed] [Google Scholar]
- Krishnan, M.N. , Sukumaran, B. , Pal, U. , Agaise, H. , Murray, J.L. , Hodge, T.W. , and Fikrig, E. (2007) Rab 5 is required for the cellular entry of dengue and West Nile viruses. J Virol 81: 4881–4885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhn, R.J. , Zhang, W. , Rossmann, M.G. , Pletnev, S.V. , Corver, J. , Lenches, E. , et al. (2002) Structure of dengue virus: Implications for flavivirus organization, maturation, and fusion. Cell 108: 717–725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, C.J. , Lin, H.R. , Liao, C.L. , and Lin, Y.L. (2008) Cholesterol effectively blocks entry of flavivirus. J Virol 82: 6470–6480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leppert, B.J. , Mansfield, J.M. , and Paulnock, D.M. (2007) The soluble variant surface glycoprotein of African trypanosomes is recognized by a macrophage scavenger receptor and induces I kappa B alpha degradation independently of TRAF6‐mediated TLR signaling. J Immunol 179: 548–556. [DOI] [PubMed] [Google Scholar]
- Lim, H.Y. , and Ng, M.L. (1999) A different mode of entry by dengue‐2 neutralisation escape mutant virus. Arch Virol 144: 989–995. [DOI] [PubMed] [Google Scholar]
- Lin, Y.L. , Lei, H.Y. , Lin, Y.S. , Yeh, T.M. , Chen, S.H. , and Liu, H.S. (2002) Heparin inhibits dengue‐2 virus infection of five human liver cell lines. Antiviral Res 56: 93–96. [DOI] [PubMed] [Google Scholar]
- Lozach, P.Y. , Burleigh, L. , Staropoli, I. , Navarro‐Sanchez, E. , Harriague, J. , Virelizier, J. , et al. (2005) Dendritic cell‐specific intercellular adhesion molecule 3‐grabbing non‐integrin (DC‐SIGN) ‐mediated enhancement of dengue virus infection is independent of DC‐SIGN internalization signals. J Biol Chem 280: 23698–23708. [DOI] [PubMed] [Google Scholar]
- Macia, E. , Ehrlich, M. , Massol, R. , Boucrot, E. , Brunner, C. , and Kirchhausen, T. (2006) Dynasore: a cell‐permeable inhibitor of dynamin. Dev Cell 10: 839–850. [DOI] [PubMed] [Google Scholar]
- Maréchal, V. , Prevost, M.‐C. , Petit, C. , Perret, E. , Heard, J.‐M. , and Schwartz, O. (2001) Human immunodeficiency virus type 1 entry into macrophages mediated by macropinocytosis. J Virol 75: 11166–11177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsh, M. , and Helenius, A. (2006) Virus entry: open sesame. Cell 124: 729–740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martínez‐Barragán, J.J. , and Del Angel, R.M. (2001) Identification of a putative coreceptor on Vero cells that participates in dengue 4 virus infection. J Virol 75: 7818–7827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mizutani, T. , Kobayashi, M. , Eshita, Y. , Shirato, K. , Kimura, T. , Ako, Y. , et al. (2003) Involvement of the JNK‐like protein of the Aedes albopictus mosquito cell line, C6/36, in phagocytosis, endocytosis and infection with West Nile virus. Insect Mol Biol 12: 491–499. [DOI] [PubMed] [Google Scholar]
- Mizzen, L. , Hilton, A. , Cheley, S. , and Anderson, R. (1985) Attenuation of murine coronavirus infection by ammonium chloride. Virology 142: 378–388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Modis, Y. , Ogata, S. , Clements, D. , and Harrison, S.C. (2004) Structure of the dengue virus envelope protein after membrane fusion. Nature 427: 313–319. [DOI] [PubMed] [Google Scholar]
- Mondotte, J.A. , Lozach, P.Y. , Amara, A. , and Gamarnik, A.V. (2007) Essential role of dengue virus envelope protein N glycosylation at asparragine‐67 during viral propagation. J Virol 81: 7136–7148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mosso, C. , Galván‐Mendoza, I.J. , Ludert, J.E. , and Del Angel, R.M. (2008) Endocytic pathway followed by dengue virus to infect the mosquito cell line C6/36 HT. Virology 378: 193–199. [DOI] [PubMed] [Google Scholar]
- Nawa, M. , Takasaki, T. , Yamada, K.‐I. , Kurane, I. , and Akatsuka, T. (2003) Interference in Japanese encephalitis virus infection of Vero cells by a cationic amphiphilic drug, chlorpromazine. J Gen Virol 84: 1737–1741. [DOI] [PubMed] [Google Scholar]
- Nayak, V. , Dessau, M. , Kucera, K. , Anthony, K. , Ledizet, M. , and Modis, Y. (2009) Crystal structure of dengue virus type 1 envelope protein in the postfusion conformation and its implications for membrane fusion. J Virol 83: 4338–4344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nichols, B.J. , and Lippincott‐Schwartz, J. (2001) Endocytosis without clathrin coats. Trends Cell Biol 11: 406–412. [DOI] [PubMed] [Google Scholar]
- Nisalak, A. , Endy, T.P. , Nimmanniya, S. , Kalayanarooj, S. , Thisayakorn, U. , Scott, R.M. , et al. (2003) Serotype‐specific dengue virus circulation and dengue disease in Bangkok, Thailand from 1973 to 1979. Am J Trop Med Hyg 68: 191–202. [PubMed] [Google Scholar]
- Oh, P. , McIntosh, D.P. , and Schnitzer, J.E. (1998) Dynamin at the neck of caveolae mediates their budding to form transport vesicles by GTP‐driven fusion from the plasma membrane of endothelium. J Cell Biol 141: 101–114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelkmans, L. (2005) Secrets of caveolae‐ and lipid raft‐mediated endocytosis revealed by mammalian viruses. Biochem Biophys Acta 1746: 295–304. [DOI] [PubMed] [Google Scholar]
- Pelkmans, L. , Kartenbeck, J. , and Helenius, A. (2001) Caveolar endocytosis of simian virus 40 reveals a new two‐step vesicular‐transport pathway to the ER. Nat Cell Biol 3: 473–483. [DOI] [PubMed] [Google Scholar]
- Pelkmans, L. , Puntener, D. , and Helenius, A. (2002) Local actin polymerization and dynamin recruitment in SV40‐induced internalization of caveolae. Science 296: 535–539. [DOI] [PubMed] [Google Scholar]
- Perera, R. , Khalik, M. , and Kuhn, R.J. (2008) Closing the door on flaviviruses: Entry as a target for antiviral drug design. Antiviral Res 80: 11–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quirin, K. , Eschli, B. , Scheu, I. , Poort, L. , Kartenbeck, J. , and Helenius, A. (2008) Lymphocytic choriomeningitis virus uses a novel endocytic pathway for infectious entry via late endosomes. Virology 378: 21–33. [DOI] [PubMed] [Google Scholar]
- Randolph, V.B. , and Stollar, V. (1990) Low pH‐induced cell fusion in flavivirus‐infected Aedes albopictus cell cultures. J Gen Virol 71: 1845–1850. [DOI] [PubMed] [Google Scholar]
- Reyes‐del Valle, J. , Chávez‐Salinas, S. , Medina, F. , and Del Angel, R.M. (2005) Heat shock protein 90 and heat shock protein 70 are components of dengue virus receptor complex in human cells. J Virol 79: 4557–4567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rojek, J.M. , Perez, M. , and Kunz, S. (2008) Cellular entry of lymphocytic choriomeningitis virus. J Virol 82: 1505–1517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rothman, A.L. (2004) Dengue: defining protective versus pathologic immunity. J Clin Inv 113: 946–951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sánchez‐San Martín, C. , López, T. , Arias, C.F. , and López, S. (2004) Characterization of rotavirus cell entry. J Virol 78: 2310–2318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sauvonnet, N. , Dujeancourt, A. , and Dautry‐Varsat, A. (2005) Cortactin and dynamin are required for the clathrin‐independent endocytosis of (gamma) c cytokine receptor. J Cell Sci 168: 155–163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Der Schaar, H.M. , Rust, M.J. , Waarts, B.‐L. , Van Der Ende‐Metselaar, H. , Kuhn, R.J. , Wilschut, J. , et al. (2007) Characterization of the early events in dengue virus cell entry by biochemical assays and single‐virus tracking. J Virol 81: 12019–12028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Der Schaar, H.M. , Rust, M.J. , Chen, C. , Van Der Ende‐Metselaar, H. , Wilschut, J. , Zhuang, X. , et al. (2008) Dissecting the cell entry pathway of dengue virus by single‐particle tracking in living cells. PloS Pathog 4: 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selinka, H.C. , Florin, L. , Patel, H.D. , Freitag, K. , Schmidtke, M. , Makarov, V.A. , and Sapp, M. (2007) Inhibition of transfer to secondary receptors by heparan sulfate‐binding drug or antibody induces noninfectious uptake of human papillomaviruses. J Virol 81: 10970–10980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sieczkarski, S.B. , and Whittaker, G.R. (2002a) Dissecting virus entry via endocytosis. J Gen Virol 83: 1535–1545. [DOI] [PubMed] [Google Scholar]
- Sieczkarski, S.B. , and Whittaker, G.R. (2002b) Influenza virus can enter and infect cells in the absence of clathrin‐mediated endocytosis. J Virol 76: 10455–10464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spoden, G. , Freitag, K. , Hausmann, M. , Boller, K. , Sapp, M. , Lambert, C. , and Florin, L. (2008) Clathrin‐ and caveolin‐independent entry of human papillomavirus type 16‐Involvement of tetraspanin‐enriched microdomains (TEMs). PloS ONE 3: 1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiasny, K. , and Heinz, F.X. (2006) Flavivirus membrane fusion. J Gen Virol 87: 2755–2766. [DOI] [PubMed] [Google Scholar]
- Suksanpaisan, L. , and Smith, D.R. (2003) Analysis of saturation binding and saturation infection for dengue serotypes 1 and 2 in liver cells. Intervirology 46: 50–55. [DOI] [PubMed] [Google Scholar]
- Swanson, J.S. , and Watts, C. (1995) Macropinocytosis. Trends Cell Biol 5: 424–428. [DOI] [PubMed] [Google Scholar]
- Talarico, L.B. , and Damonte, E.B. (2007) Interference in dengue virus adsorption and uncoating by carrageenans. Virology 363: 473–485. [DOI] [PubMed] [Google Scholar]
- Talarico, L.B. , Pujol, C.A. , Zibetti, R.G.M. , Faría, P.C.S. , Noseda, M.D. , Duarte, M.E.R. , and Damonte, E.B. (2005) The antiviral activity of sulfated polysaccharides against dengue virus is dependent on virus serotype and host cell. Antiviral Res 66: 103–110. [DOI] [PubMed] [Google Scholar]
- Talarico, L.B. , Duarte, M.E.R. , Zibetti, R.G.M. , Noseda. M.D., and Damonte, E.B. (2007) An algal‐derived DL‐galactan hybrid is an efficient preventing agent for in vitro dengue virus infection. Planta Med 73: 1464–1468. [DOI] [PubMed] [Google Scholar]
- Thepparit, C. , and Smith, D.R. (2004) Serotype‐specific entry of dengue virus into liver cells: identification of the 37‐kilodalton/67‐kilodalton high‐affinity laminin receptor as a dengue virus serotype 1 receptor. J Virol 78: 12647–12656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umashankar, M. , Sánchez San Martín, C. , and Liao, M. (2008) Differential colesterol binding by class II fusion proteins determines membrane fusion properties. J Virol 82: 9245–9253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Hamme, E. , Dewerchin, H.L. , Cornelissen, E. , Verhasselt, B. , and Nauwynck, H.J. (2008) Clathrin‐ and caveolae‐independent entry of feline infectious peritonitis virus in monocytes depends on dynamin. J Gen Virol 89: 2147–2156. [DOI] [PubMed] [Google Scholar]
- Vaughn, D.W. , Green, S. , Kalayanarooj, S. , Innis, B.L. , Nimmannitya, S. , and Suntayakorn, S. (2000) Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J Infect Dis 181: 2–9. [DOI] [PubMed] [Google Scholar]
- Vidricaire, G. , and Tremblay, M.J. (2007) A clathrin, caveolae, and dynamin‐independent endocytic pathway requiring free membrane cholesterol drives HIV‐1 internalization and infection in polarized trophoblastic cells. J Mol Biol 368: 1267–1286. [DOI] [PubMed] [Google Scholar]
- Wang, H. , Yang, P. , Liu, K. , Guo, F. , Zhang, Y. , Zhang, G. , and Jiang, C. (2008) SARS coronavirus entry into host cells through a novel clathrin‐ and caveolae‐independent endocytic parhway. Cell Res 18: 290–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng, J. , Shen, W.H. , Lu, T.J. , Zhou, Y. , Chen, Q. , Wang, Z. , et al. (2008) Clathrin‐dependent endocytosis is required for TrkB‐dependent Akt‐mediated neuronal protection and dendritic growth. J Biol Chem 9: 13280–13288. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1. Effect of ammonium chloride on DENV multiplication, viral particles infectivity, virus adsorption and intracellular vesicle acidification. A. Cells were treated with increasing concentrations of ammonium chloride and infected with DENV‐1 orDENV‐2. Virus yields were quantified by plaque assay at 48 h p.i. B. Suspensions ofDENV‐2 virions were incubated at 37°C with various concentrations of ammonium chloride. After 1 h samples were diluted and remaining infectivity was determined by plaque formation assay. C. Vero cells were treated as in (A) and infected with DENV‐2 at 4°C. After 1 h adsorption in the presence or absence of the drug cells were disrupted and cell‐bound virus was determined by plaque formation assay. In (A), (B) and (C) results are expressed as percentage of virus multiplication with respect to a control without drug treatment. Each point shows the mean ± SD of two independent experiments. D. Cells were left untreated (control) or treated with 50 mM ammonium chloride or 50 nM concanamycin A. Then, samples were stained with acridine orange.
Fig. S2. DENV‐1 infection of Vero cells is independent of caveolae but dependent on dynamin. A. Cells transiently transfected with the constructs GFP‐cav‐1 wt, GFP cav‐1 DN or GFP‐cav‐1 Y14F were infected with DENV‐1. After 24 h infection cultures were fixed and immunofluorescence staining was performed. B. For quantification of samples shown in (A), 250 transfected cells with similar levels of GFP expression were screened and cells positive for viral antigen were scored. C. Cells transiently transfected with the constructs GFP‐dyn II wt or GFP‐dyn II K44A were infected withDENV‐1. After 24 h infection cultures were fixed and immunofluorescence staining was performed. D. For quantification of samples shown in (C), 250 transfected cells with similar levels of GFP expression were screened and cells positive for viral antigen were scored. In (B) and (D) results are expressed as the mean ± SD of two independent experiments.
Fig. S3. Effect of different inhibitors on A549 cells endocytic processes. A549 cells were left untreated (control) or treated with ammonium chloride 50 mM (A), chlorpromazine 50 μM (B) or nystatin 100 μM (C). Then cells were stained with acridine orange (A), or incubated with TRITC‐labelled transferrin (B) or FITC‐labelled cholera toxin (C).
Please note: Wiley‐Blackwell are not responsible for the content or functionality of any supporting materials supplied by the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
Supporting info item