

Abstract
Cognitive control refers to the use of internal goals to guide how we process stimuli, and control can be applied proactively (in anticipation of a stimulus) or reactively (once that stimulus has been presented). The application of control can be guided by memory; for instance, people typically learn to adjust their level of attentional selectivity to changing task statistics, such as different frequencies of hard and easy trials in the Stroop task. This type of “control-learning” is highly adaptive, but its boundary conditions are currently not well understood. In the present study, we assessed how the presence of performance feedback shapes control-learning in the context of item-specific (reactive control, Experiments 1a and 1b) and list-wide (proactive control, Experiments 2a and 2b) proportion of congruency manipulations in a Stroop protocol. We found that performance feedback did not alter the modulation of the Stroop effect by item-specific cueing, but did enhance the modulation of the Stroop effect by a list-wide context. Performance feedback thus selectively promoted proactive, but not reactive, adaptation of cognitive control. These results have important implications for experimental designs, potential psychiatric treatment, and theoretical accounts of the mechanisms underlying control-learning.
Keywords: cognitive control, attention, memory, control-learning, performance feedback
Introduction
In many everyday scenarios (e.g., driving during heavy traffic), people need to engage cognitive control, using internal goals to guide how they process stimuli and select actions, rather than relying on routine behavior. Given repeated exposure to, or commonalities between, particular situations or contexts that require the recruitment of cognitive control, people can learn to link contextual cues (for instance, approaching a particular type of intersection) to the need for imposing a particular type of control “state” (e.g., a heightened focus of attention) (Abrahamse, Braem, Notebaert, & Verguts, 2016; Egner, 2014). This phenomenon, known as “control-learning,” occurs when previous experience (memory) guides control strategies and is typically assessed by analyzing how people learn to adjust their attentional priorities to context-dependent differences in frequencies of hard and easy trials on classic control tasks (for reviews, see Bugg, 2017; Bugg & Crump, 2012; Chiu & Egner, 2019), including conflict-control paradigms.
Conflict-control learning refers to how people adjust to interference (or conflict) in information processing when current goals conflict with overlearned, prepotent responses (Botvinick, Braver, Barch, Carter, & Cohen, 2001). For example, in the classic Stroop task (Stroop, 1935), participants must name the ink color of color-words and are typically slower and less accurate when the ink color is incongruent with the meaning of the color-word (e.g., the word BLUE printed in red) than when the ink color is congruent (e.g., the word BLUE printed in blue). This performance difference, referred to as the congruency effect, is thought to index the need for, and efficiency of, cognitive control. Per the influential “conflict-monitoring theory” (Botvinick et al., 2001), the cognitive system monitors the level of processing conflict to drive commensurate adjustments in the recruitment of control: more conflict (e.g., an incongruent trial) requires more control, and greater control results in more efficient attentional selection and thus a smaller congruency effect over time.
Within the conflict-control literature, research has focused on so-called proportion congruent (PC) effects as indices of learning-guided control. PC effects refer to the observation that congruency effects are reduced in contexts where conflict (incongruent trials) is more frequent or probable, i.e., in the presence of implicit cues, stimuli, or blocks that are more predictive of incongruent than congruent trials (e.g., Bugg & Crump, 2012; Logan & Zbrodoff, 1979). In an item-specific proportion congruent (ISPC) paradigm, for instance, the ratio of hard (incongruent) to easy (congruent) trials varies as a function of a particular stimulus (e.g., a specific animal category), which acts as the learning signal cueing the recruitment of specific control-states (Bugg, Jacoby, & Chanani, 2011). Thus, the ISPC is thought to reflect what is known as “reactive control,” or control that is recruited in the moment, in response to a particular stimulus-context configuration, because participants cannot anticipate which cue or stimulus will be shown from one trial to the next (Braver, 2012; Bugg, 2017). In contrast to the ISPC, the list-wide proportion congruent (LWPC) paradigm varies the ratio of hard to easy trials between blocks of trials and is thus thought to reflect “global” or “proactive control,” or control that is recruited in anticipation of upcoming demand, because here demand is temporally predictable.
Learning to apply control in a context-sensitive manner is highly useful. Accordingly, discovering ways to boost control-learning has important implications both for improving our understanding of high-level, adaptive cognition, as well as for potential interventions in populations for whom context-dependent control is known to be impaired (e.g., schizophrenia, Barch & Sheffield, 2017; aging, Braver et al., 2014).The present study therefore investigated a key factor that may promote the learning of reactive and/or proactive conflict-control strategies, namely, the provision of performance feedback, which can act as an intrinsic motivator of learning (Bejjani, DePasque, & Tricomi, 2019; Tricomi & DePasque, 2016).
Previous work has primarily investigated the effects of extrinsic reinforcers on the recruitment of proactive and reactive control, especially within the AX-Continuous Performance Task (AX-CPT) (Chiew & Braver, 2011, 2013; Fröber & Dreisbach, 2014, 2016; Qiao et al., 2018; Yee & Braver, 2018). Here, trials consist of contextual cues followed by response-probes, and particular responses are required for specific cue-probe combinations (e.g., A-X). Particular cue-target combinations occur with high frequency, thus biasing the responses associated with each probe. Proactive, preparatory control is indexed by the amount of interference experienced when the cue does not match its usual target on A-Y lure trials, while reactive control is indexed by the amount of interference experienced when inhibiting responses on B-X lure trials. Typical findings show that the recruitment of proactive control is enhanced with extrinsic reinforcers such as reward (e.g., see reviews of motivation-cognition interactions, Yee & Braver, 2018; Botvinick & Braver, 2015).
Thus, there is some evidence that performance feedback can selectively enhance proactive control. However, it is not clear whether these findings in the AX-CPT generalize to the type of experiential control-learning processes captured in PC paradigms. Specifically, cues in the AX-CPT are explicitly instructed, and the control requirements consist of overcoming pre-potent response priming. By contrast, PC paradigms tap into the implicit learning of varying cognitive control demands through interaction with the task. In the case of conflict-control tasks, this involves learned adjustments in attentional selection of target (over distracter) stimulus information, rather than response priming. Finally, most AX-CPT studies have focused exclusively on how extrinsic reward boosts control (Krug & Braver, 2014) rather than examining effects of intrinsic reinforcement triggered by performance feedback.
Another line of relevant research has indicated that incentivizing task performance can enhance the recruitment of cognitive control on conflict-control tasks (Bugg, Diede, Cohen-Shikora, & Selmeczy, 2015; Chiew & Braver, 2016; Kang, Wang, & Zhou, 2017; Kostandyan et al., 2019; Soutschek, Stelzel, Paschke, Walter, & Schubert, 2015; Soutschek, Strobach, & Schubert, 2014). For example, given sufficient preparation time, people show reduced flanker interference effects when cues are predictive of reward and informative of the upcoming trial congruency (Chiew & Braver, 2016). They also show reduced cross-modal conflict for target detection when predictive reward cues are predictable; if unpredictable, participants could only reduce cross-modal conflict given enough preparatory time and differentiation between the cue and target (Kang et al., 2017).
Because preparatory time is important for recruiting proactive control, researchers have investigated the temporal dynamics of when, and for how long, incentives can impact control recruitment. One study has shown that when participants are rewarded with cumulative point totals after twenty-trial reward blocks, they do not modulate their flanker interference effect, but do show increased pupil dilation, relative to no reward blocks; however, when pre-cues indicate the transient possibility of reward, on each trial, within those short blocks, participants show both increased pupil dilation and reduced flanker effects, suggesting that reward boosts sustained arousal and results in transient performance benefits (Kostandyan et al., 2019). Likewise, Bugg and colleagues (2015) reported transient performance changes whereby – irrespective of reward – participants used probabilistic cues that indicated whether twenty-trial blocks were mostly hard or easy to relax their control settings; reward mostly speeded responses and influenced the first trial within the block, indicating its primary effect on arousal and task expectations. Finally, Soutschek and colleagues (2014a, b) found independent effects of reward and conflict expectancy in boosting the recruitment of cognitive control. These studies collectively demonstrate that extrinsic reward can increase arousal and change task performance, contingent on whether the task timing and parameters allow for participants to utilize the cue information. Of note, however, all of these studies rewarded participants on a block-wise level, used extrinsic rewards with relatively transient effects, or investigated how reward cues impacted the recruitment of cognitive control rather than experiential learning of control.
Whether the type of learned reactive and proactive control-learning processes that are assessed in the ISPC and LWPC paradigms, respectively, are sensitive to motivational influences such as reward and performance feedback thus remains largely unknown. Current theories on control-learning suggest that individual perceptual and motor representations become bound in an associative network with goal representations that are co-activated during an event, allowing for contextually appropriate recruitment of control (e.g., Abrahamse et al., 2016; Egner, 2014). This associative binding then results in three primary predictions about control-learning: one, that control-learning is context-specific; two, that control-learning is primarily implicit; and three, that control-learning is sensitive to reward. The latter two, in particular, refer to the grounding of cognitive control in associative learning principles. When applying associative learning models to cognitive control data, researchers assume that instead of learning the likelihood of reward, participants implicitly learn the likelihood of upcoming control-demand and update their expectancies based on the control-demand they experience on each trial (Chiu, Jiang, & Egner, 2017; Jiang, Beck, Heller, & Egner, 2015; Muhle-Karbe, Jiang, & Egner, 2017). This process should be sensitive to implicit or explicit reward, because reward would be expected to reinforce these learned associations.
Here, we tested whether the mere addition of performance feedback (“correct”, “incorrect”) would modulate how participants learn to recruit reactive and proactive cognitive control. Performance feedback can motivate participants and provide valuable information, which, in itself, is also intrinsically rewarding (cf. Lempert & Tricomi, 2015). Although success on difficult control trials can act as its own reward (Schouppe et al., 2015), proactive control is thought to be costly to recruit (Braver, 2012). If intrinsic motivational factors can compensate for the costs of recruiting control, performance feedback should differentially impact control-learning, depending on the type of control-demand.
The overarching goal of this study was thus to test how performance feedback shapes control-learning. To this end, Experiments 1a and 1b employ a picture-word ISPC Stroop task as a measure of learned reactive, in-the-moment control, and Experiments 2a and 2b employ a picture-word LWPC Stroop task as a measure of learned proactive or global control. We manipulated the receipt of performance feedback as a between-subjects factor in both experiments, and tested for the interaction between trial type (difficult/easy), proportion congruent (i.e., the probability of high/low control-demand), and feedback group (no feedback/feedback) on reaction time and accuracy as evidence of learning.
Our results will shed light on motivation-cognition interactions as well as how learning modulates the application of cognitive control. Specifically, finding that performance feedback enhances the recruitment of proactive control in the LWPC but not reactive control in the ISPC would suggest that the dual mechanisms of control framework applies even when accounting for the role of learning in cognitive control (Chiu & Egner, 2019) and that control-learning theories need to account for the temporal demands of how control is recruited. If, on the other hand, we find that neither the LWPC nor the ISPC is sensitive to performance feedback, that would suggest that the temporal demands of proactive and reactive control matter little and that learned control recruitment is relatively automatic and unsusceptible to motivational influences.
Experiment 1a:
To assess how performance feedback affects the learning of associations between specific stimuli and control demands, we devised a picture-word item-specific proportion congruent (ISPC) Stroop task. In this task (Figure 1), image categories served as implicit probabilistic cues that predicted the likelihood of conflict-control demand: half of the images were more likely to appear with an incongruent overlaid distractor word (low proportion congruent; e.g., DEER on a picture of a bear), while the others were more likely to appear with a congruent overlaid distractor word (high proportion congruent; e.g., BEAR on a picture of a bear).
Figure 1. Summary of task procedure.
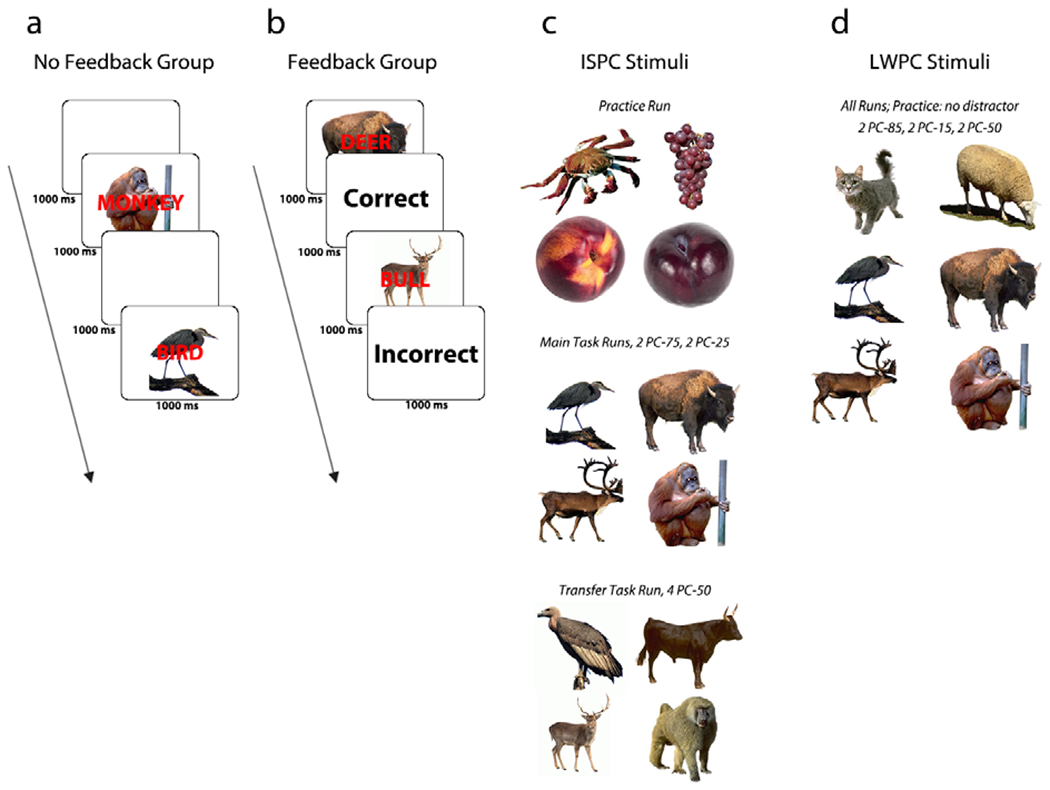
Across Experiments 1 and 2, participants categorized picture stimuli while ignoring overlaid distractor words. Some images were more often paired with incongruent distracters (low PC items: high control-demand), while others were more often paired with congruent distracters (high PC items: low control-demand). Some were also paired equally often with incongruent and congruent distracters (PC-50). The receipt of performance feedback was manipulated between groups (A, B). In Experiment 1, we used different stimuli for the practice run (which acclimated participants to the picture-word Stroop task demands), the main task runs, and the transfer run (C). In Experiment 2, four of the stimuli were biased at the item and list level, associated with either a lower or higher likelihood of control-demand, while two PC-50 stimuli were biased only at the list-level (PC-72/28). Participants were exposed to all these images during the practice run (D). Note that all images are from http://cabezalab.org/cabezalabobjects/, but do not match the stimuli used in the Experiments (1: bird, deer, bear, turtle; 2: bird, deer, bear, turtle, cat, and dog).
This ISPC paradigm avoids stimulus-response learning confounds related to participants learning to use the distracters for predicting the response (e.g., Jacoby et al., 2003; Schmidt & Besner, 2008), because the task-relevant target image, rather than the distracter word, predicts congruency as well as responses in this design (cf. Bugg et al., 2011; Chiu et al., 2017). This is a commonly used design, and the current task thus allows us to determine to what degree the ISPC effect in a typical ISPC design may benefit from performance feedback. However, this design may be a suboptimal assessment of “pure” item-based control, since it involves different frequencies of specific stimulus-distracter compound stimuli (e.g., more frequent occurrences of the mostly-congruent and mostly-incongruent items) (Braem et al., 2019). To assess “pure” item-based control, we also included a transfer run without frequency confounds (Bugg et al., 2011; Bugg & Dey, 2018).
In the final run of the task, we tested whether these learned associations between image categories and control-demand also transferred to unbiased category exemplars, which were not predictive of proportion congruency. Transfer is not crucial for understanding the effect of performance feedback on the typical ISPC effect, but provides a cleaner measure of adaptive control, and is likely dependent on the strength of the ISPC effect due to its reliance on the category-level association (cf. Bugg & Dey, 2018). Crucially, in one group of participants, this task was administered without trial-by-trial feedback on response accuracy (Figure 1a), while for another group such feedback was supplied (Figure 1b).
Given that the ISPC effect is thought to reflect learned reactive control (e.g., Bugg, 2017), which in other contexts has been suggested to be insensitive to reward (Braver, 2012), we anticipated observing an ISPC effect of similar magnitude in both feedback groups. However, another plausible possibility is that, like most instances of associative learning, the acquisition of stimulus-control associations underlying the ISPC would benefit from performance feedback, which in turn would predict a larger ISPC effect for the feedback vs. no feedback group. Irrespective of these rival hypotheses, for the no feedback group, we expected to replicate previous ISPC studies, showing an interaction between proportion congruent (PC) and congruency whereby congruency effects are reduced for low PC relative to high PC images.
Method
Participants
The target sample size was determined based on previous ISPC studies (Bugg, Jacoby, and Chanani, 2011; Bugg, 2014; Bugg and Dey, 2018), which reported varying effect sizes (range ηp2 = [0.27, 0.50]). Given the uncertainty in the literature as to the true effect size, with a high power of 0.9, a Type I error of 0.05, and similar trial counts (e.g., 384 training trials for four images in Bugg et al., 2011 and Bugg, 2014), we required between eight (ηp2 = 0.50) and eighteen (ηp2 = 0.27) participants to observe the canonical PC by congruency interaction. To account for a new between-subjects factor, we aimed to run thirty participants per feedback group.
Sixty-seven Amazon Mechanical Turk (MTurk) workers consented to participate and were compensated $3.25 for spending approximately twenty-five minutes on this task. They were instructed to achieve above 70% accuracy. Seven workers were excluded because of poor accuracy (i.e., less than 70% accuracy, range [20.8%, 56.0%], M = 36.0% ± 15.5%), resulting in a final sample size of sixty (feedback (N = 30): meanage = 32.6 ± 8.8; 9F, 21M; no feedback (N = 30): meanage = 33.4 ± 11.3; 13F, 17M).
Stimuli
We obtained object images from an online database (http://cabezalab.org/cabezalabobjects/) and google searches of images with a license for noncommercial reuse with modification. These images were previously used in another study (Bejjani & Egner, 2018) and cropped to 500 x 300 pixels. Two image categories were used to create each item-specific proportion congruent context (e.g., PC-75 or PC-25; Figure 1c).
Experimental Procedure
The experimental procedure consisted of a practice run, three main task runs, and a transfer task run (Figure 1). First, in order to familiarize participants with the general idea of a picture-word Stroop task, they underwent a practice run with 24 trials consisting of images and stimulus-response mappings that were not included in the main task (Figure 1c). As in the main task, they classified picture stimuli while trying to ignore distractor category labels that were printed, in red, and overlaid on the pictures that they categorized. Participants categorized images by pressing category-specific keyboard response buttons (z, x, n, and m on a QWERTY keyboard, mapped onto left middle, left index, right index, and right middle finger, respectively). On each trial, an image stimulus was shown for 1000 ms, followed by performance feedback for 500 ms and then a blank intertrial interval screen for 500 ms. If participants did not achieve greater than 80% accuracy, they were required to redo the practice run. We ensured that participants understood the task instructions and demand without being overexposed to the stimulus-response mappings.
To establish an association between image categories and attentional control states, participants then performed three ISPC runs each consisting of 144 picture-word Stroop trials (Figure 1a and 1b). Unbeknownst to the participants, each image category (e.g., bear, deer, Figure 1c, Table 1) was associated with a 0.75 probability for either incongruent (high control-demand/low PC) or congruent (low control-demand/high PC) distracter words. These PC-75 and PC-25 items were presented with equal frequency in random order, producing an overall incidence of 50% congruent and incongruent trials. Our manipulation created two types of “control-demand cues”: one associated with a low state of attentional selectivity (mostly congruent trial cues) and another associated with a high state of attentional selectivity (mostly incongruent trial cues) (both at 0.75 probability). The trial timing followed that of the practice run, except that image stimuli were either followed by 1000 ms of a blank intertrial interval screen (no feedback group; Figure 1a) or 1000 ms of veridical performance feedback (feedback group; Figure 1b).
Table 1.
Item-specific Proportion Congruency Design Manipulation.
| Mostly Congruent (MC) | Mostly Incongruent (MIC) | |
|---|---|---|
| Main Task, PC-75/PC-25 Items (Runs 1-3) | ||
| Congruent | 75% (54 trials) | 25% (18 trials) |
| Incongruent | 25% (l8 trials) | 75% (54 trials) |
| Transfer, PC-50 Items (Run 4) | ||
| Congruent | 50% (36 trials) | 50% (36 trials) |
| Incongruent | 50% (36 trials) | 50% (36 trials) |
Finally, the transfer run followed the sequence and timing of the main task runs, but involved different category exemplar images that were non-predictive of trial congruency (i.e., PC-50 items; Figure 1c). This transfer tested whether participants generalized their learned control associations such that the “bear” and “deer” image categories, for example, cue higher control-demand, regardless of the specific bear and deer images that were shown. Specifically, this manipulation created separate, unbiased PC-50 items that could index learned stimulus-control at the category, rather than item-specific, level.
Across both main task and transfer runs, we minimized contingency learning and feature integration confounds by ensuring that each stimulus had only one congruent and incongruent pairing. In a typical four-choice Stroop task, participants could have twelve incongruent stimulus pairings and only four congruent stimulus pairings, but the current experiment matched the pairings so that there were only four incongruent stimuli and four congruent stimuli. For example, all incongruent trials for the bear picture, in one task version, involved the distractor word “deer” overlaid on the picture, and the deer picture was likewise only incongruent with the “bear” distractor word. For the transfer run, we used different image/word pairings for the incongruent trial than participants learned in the main task runs (cf. “far transfer”; Bugg et al., 2011). Unlike the main task runs, the transfer run also controls for stimulus frequency confounds, in which the frequently incongruent target-distractor combinations occur more often than the specific rarely incongruent target-distractor combinations, providing an index of pure control-learning without the possibility of lower-level stimulus-response learning contributions (see Braem et al., 2019).
Across feedback groups, we counterbalanced the assignment of image categories to proportion congruent such that each image pairing (e.g., bear + bird; bear + turtle; bear + deer) served as a mostly congruent and mostly incongruent context at least once. Note that although some of the image stimuli are oriented towards the left or right of the screen, counterbalancing image assignment to proportion congruent contexts controlled for any response incongruencies that stem from these spatial alignments.
If participants did not respond before the image disappeared from the screen, a feedback time-out was provided for 500 ms (no feedback) or 1000 ms (feedback) to encourage quicker responses.
All images were presented with equal frequency in the center of the screen, and all trial types were presented in random order.
Post-test Questionnaire
After the main experiment, participants filled out a surprise post-test questionnaire that assessed their explicit understanding of the ISPC manipulation. We first asked participants whether they noticed any systematic differences in how often an image was associated with an easy (congruent) trial or hard (incongruent) trial. We then assessed how well participants could cluster together images that were both predictive of hard or easy contexts (e.g., rating the bear and turtle together as being predictive). Finally, we asked the participants to match each picture to its associated trial type (hard/incongruent or easy/congruent). Fourteen participants did not answer these last two questions.
Data Analysis
We first analyzed reaction time (RT) and accuracy data for correct trials that were not a direct stimulus repetition from the previous trial (2.26% of trials) and that were not excessively fast (< 200 ms; 0.10% of trials) or slow (feedback time-out: > 1000 ms; 3.8% of trials). We then ran a 2 x 2 x 2 repeated-measures ANOVA with proportion congruent (mostly incongruent/mostly congruent) and current trial congruency (congruent/incongruent) as within-subjects factors and feedback group (feedback/no feedback) as a between-subjects factor. We ran separate ANOVAs for the PC-75/25 (main task runs) and PC-50 (transfer run) items. We include inclusion Bayes Factors for all data analyzed with repeated-measures ANOVAs, using the BFEffects R package (https://github.com/mattansb/BFEffects) via the Bayes Factor package (https://richarddmorey.github.io/BayesFactor/). Inclusion Bayes Factors, via this package, answer the question, “Are the observed data more probable under models with a particular predictor than they are under models without that particular predictor?” For example, an inclusion BF of 2 means that a model with that predictor is two times more likely than a model without that predictor.
All materials are available online at https://github.com/christinabeiiani/sclearn_fb.
Results and Discussion
Main Task, PC-75/25 Items
In Experiment 1, some image categories mostly predicted congruent trials (low control-demand) while others mostly predicted incongruent trials (high control-demand). The item-specific proportion congruent (ISPC) effect denotes the difference in Stroop congruency for low (PC-75) vs. high (PC-25) control-demand cues, i.e., the proportion congruent x congruency interaction. Tables S1–2 detail the RT and accuracy data. Because we detected strong speed/accuracy trade-offs within the data (see below), we focus our analyses on inverse efficiency scores and report analyses of RT and accuracy in the Supplementary Text for all experiments.
Inverse Efficiency
When examining the data, we detected strong trade-offs between reaction time and accuracy in both subject groups (overall: r(58) = −0.51, p < 0.001; no feedback group: r(28) = −0.60, p < 0.001; feedback group: r(28) = −0.52, p = 0.003). This was especially noticeable when examining mean RTs and accuracies within conditions (see Tables S1 and S2): sometimes participants were more accurate (but slower) on incongruent than congruent trials. Because of this RT/accuracy trade-off, we also computed (and analyzed) inverse efficiency scores (IES, Townsend & Ashby, 1983) by dividing RT within each condition by its respective accuracy (proportion correct) for each subject. This produces an RT score corrected for accuracy.
When taking both RT and accuracy into account in this manner, we observed an expected main effect of congruency (F(1,58) = 22.93, p < 0.001, ηp2 = 0.28, BF = 1870.64), as corrected RT was higher for incongruent (mean = 763 ms) than congruent trials (mean = 725 ms). This effect was qualified by an interaction with the PC factor (F(1,58) = 7.81, p = 0.007, ηp2 = 0.12, BF = 1.20) in the absence of a 3-way interaction involving the feedback group factor (F(1,58) = 0.08, p = 0.775, ηp2 = 0.00, BF = 0.02). Both the no feedback (mean: 35 ms) and feedback (mean: 28 ms) groups displayed significantly reduced congruency effects for low vs. high PC images (Figure 2a). No other effects reached significance (Fs < 0.33).
Figure 2. Experiment 1 control-learning results.
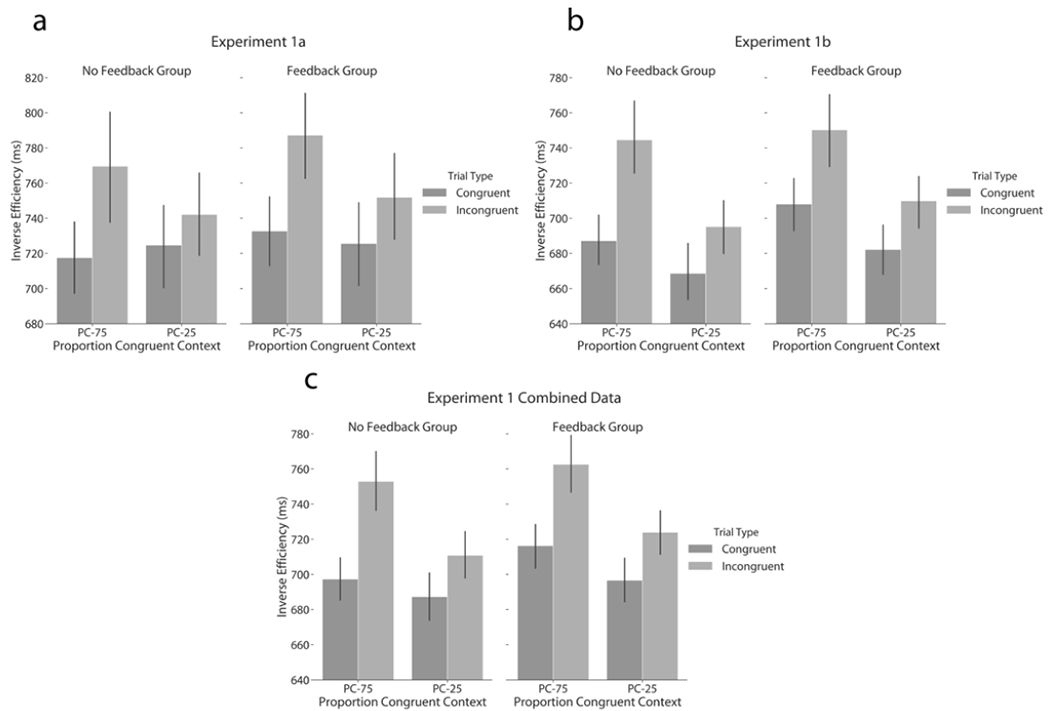
Mean Inverse Efficiency data (± SEM) are displayed as a function of proportion congruent context, current trial congruency, and feedback group for (a & b) the ISPC effect of Experiments 1a and b, which are averaged together in (c).
Overall, these analyses suggest that participants, irrespective of whether they received trial-by-trial performance feedback, learned the crucial item-specific associations between the image categories and specific control-states and adjusted their attentional priorities accordingly.
Transfer Run, PC-50/50 Items
Inverse Efficiency
Because there was also a speed-accuracy trade-off in the transfer run (overall: r(58) = −0.42, p < 0.001; no feedback group: r(28) = −0.48, p = 0.007; feedback group: r(28) = −0.47, p = 0.009), we computed inverse efficiency scores. We observed no significant effects, and thus little evidence for “far transfer” of the ISPC effect to new category members (congruency: F(1,58) = 2.69, p = 0.107, ηp2 = 0.04, BF = 0.32; congruency x feedback: F(1,58) = 2.05, p = 0.157, ηp2 = 0.03, BF = 0.23; all other effects, F < 1.52). Both the no feedback (mean: 19 ms) and feedback (mean: 4 ms) groups showed small, non-significant congruency differences between non-predictive category exemplars, suggesting that participants, as a whole, did not generalize previously learned item-specific control associations to unbiased category exemplars.
Explicit Understanding of Task Structure
We assessed whether participants had explicit knowledge of the task structure through a series of post-test questions. Twenty-five participants reported noticing that some images systematically varied in their proportion congruency. However, participants did not rate the possible image category pairings (e.g., bear + bird; bear + deer; bear + turtle) that could have been clustered together as a PC-75 or PC-25 “context” as being more predictive than chance (50%; t(45) = 1.71, p = 0.093, Cohen’s dz = 0.25; other ts < 0.67). Participants were also at chance (2/4) when matching the image categories to their respective congruencies (main task: t(46) = 0.84, p = 0.404, Cohen’s dz = 0.12; transfer run: t(46) = 0.45, p = 0.652, Cohen’s dz = 0.07). In sum, this suggests that most participants remained unaware of the task structure and learned the stimulus-control associations implicitly.
Experiment 1b:
Because Experiment 1a resulted in a speed-accuracy trade-off, which made estimation of the true effect size more difficult, and we sought to probe whether the findings of Experiment 1a were replicable, we ran a (larger) replication sample in Experiment 1b. To discourage a speed-accuracy trade-off, we added in an additional reminder in the task instructions about responding as quickly and accurately as possible, and restricted the participant sample to people who had a mailing address in the United States (Location: US qualification on MTurk), since participants with the largest overall inverse efficiency scores in Experiment 1a resided elsewhere.
Method
Participants
Determining the appropriate sample size for a replication of Experiment 1a is not straightforward. On the one hand, due to the strong speed-accuracy tradeoffs, the RT values (see Supplemental Text) alone likely do not represent an appropriate estimate of the true effect size. On the other hand, inverse efficiency scores are not commonly used in power analyses. We therefore eschewed formal effect size estimation and instead chose to double the sample size of Experiment 1a, aiming for a total of one hundred and twenty participants, which we hoped would provide reliable estimates and could be combined into a larger analysis.
One hundred and twenty-four MTurk workers consented to participate and were compensated $3.25 for spending approximately twenty-five minutes on this task. Four workers were excluded because of poor accuracy (i.e., less than 70% accuracy, range [9.0%, 38.0%], M = 24.1% ± 12.0%), resulting in a final sample size of one hundred and twenty (feedback (N = 60): meanage = 39.2 ± 12.4; 23F, 33M, 1 Genderqueer, 3 n/a; no feedback (N = 60): meanage = 34.6 ± 10.0; 28F, 31M, 1 n/a).
Experimental Procedure
The experimental procedure remained the same as in Experiment 1a, with one notable exception. Because we did not observe “far transfer” in Experiment 1a, we hypothesized that this was, in part, due to changing the word associated with each category-level exemplar from the main task runs to the transfer run. We therefore ensured that the same word associated with the category-level exemplars in the main task run was overlaid on the new, non-predictive category exemplar in the transfer run. Four participants did not answer the post-test / demographics questionnaire at the end of the learning task.
Results & Discussion
Main Task Runs, PC-75/25 Items
Inverse Efficiency
As with Experiment 1a, there were strong trade-offs between reaction time and accuracy data (overall: r(118) = −0.49, p < 0.001; no feedback group: r(58) = −0.48, p < 0.001; feedback group: r(58) = −0.53, p < 0.001). We therefore again computed inverse efficiency scores and reran the above analyses.
When combining RT and accuracy into a single metric, we observed significant main effects of congruency (F(1,118) = 74.21, p < 0.001, ηp2 = 0.38, BF = 3.58 x 1012) and PC (F(1,118) = 45.87, p < 0.001, ηp2 = 0.28, BF = 3.70 x 109). We found that participants were generally faster on congruent (mean = 686 ms) than incongruent (mean = 725 ms) trials, and in the low (mean = 689 ms) compared to high (mean = 722 ms) PC condition.
Importantly, we here also observed a significant interaction between congruency and proportion congruent context (Figure 2b; F(1,118) = 6.83, p = 0.010, ηp2 = 0.06, BF = 4.91), i.e., the typical ISPC effect, as congruency effects were smaller in the PC-25 (mean = 27 ms) than in the PC-75 (mean = 50 ms) condition. Combined with the absence of a significant 3-way interaction with feedback (F(1,118) = 0.89, p = 0.347, ηp2 = 0.01, BF = 0.03), this suggests that performance feedback had little effect on the adaptive modulation of the Stroop effect by proportion congruent context. The ISPC effect was 31 ms and 14 ms for the no feedback and feedback groups, respectively. No other effects were significant (F < 0.90).
Overall, for PC-75/25 items in Experiment 1, we observed only weak hints of a three-way interaction between congruency, context, and feedback for RT, but stronger evidence for a two-way interaction between congruency and context for both accuracy and IES. Together these results suggest that participants were able to modulate their congruency effects for low vs. high control-demand contexts irrespective of whether they received feedback.
Transfer Run, PC-50/50 Items
Inverse Efficiency
We observed significant main effects of congruency (F(1,118) = 20.14, p < 0.001, ηp2 = 0.15, BF = 4.74) and PC (F(1,118) = 8.29, p = 0.005, ηp2 = 0.07, BF = 13.55). Using this corrected RT measure, we found that participants responded faster on congruent (mean = 680 ms) than incongruent (mean = 695 ms) trials, and in the low (mean = 679 ms) compared to high (mean = 695 ms) PC condition. Congruency effects were also smaller when participants did not receive feedback (8 ms) than when they did (21 ms), congruency x feedback: F(1,118) = 4.27, p = 0.041, ηp2 = 0.04, BF = 0.30. No other effects were significant (feedback: F(1,118) = 1.47, p = 0.228, ηp2 = 0.01, BF = 0.38; all other effects, F < 0.26).
Overall, in Experiment 1, we found weak congruency effects in the transfer run and little evidence that participants modulated their congruency effects by proportion congruent context. This suggests that the transfer effect is weaker than the ISPC effect and that we may have been underpowered to detect any such small modulations.
Explicit Understanding of Task Structure
We assessed whether participants explicitly understood the task structure through a series of post-test questions. Thirty participants reported noticing that images systematically varied in their proportion congruency. However, participants did not rate the possible image category pairings (e.g., bear + bird; bear + deer; bear + turtle) that could have been clustered together as a PC-75 or PC-25 “context” as being more predictive than chance (50%; t(113) = 1.69, p = 0.093, Cohen’s dz = 0.16; other ts < 0.78). Participants were significantly worse than chance (2/4) at matching the image categories to their respective congruencies (main task: t(113) = 6.00, p < 0.001, Cohen’s dz = 0.56; transfer run: t(113) = 2.47, p = 0.015, Cohen’s dz = 0.23). As with Experiment 1a, most participants remained unaware of the task structure and learned the stimulus-control associations implicitly.
Across Experiment 1 Analysis
Because Experiments 1a and 1b shared the same task structure, with the exception of changing the word associations in the transfer task, we aggregated our data across experiments for exploratory analyses with enhanced power. We added experiment (1a/1b) as a between-subjects factor and then reran analyses across RT, accuracy, and inverse efficiency data to better evaluate the effect of performance feedback on the ISPC effect. Because we did not observe group-level transfer effects in either Experiment and because the transfer phase differed between the two cohorts, we did not include transfer run data here. Moreover, since apart from the transfer phase Experiment1a and 1b were identical in design and procedure, we treat Experiment as a factor of no interest in this analysis.
Main Task Runs, PC-75/25 Items
Inverse Efficiency
When combining RT and accuracy into a single metric to account for the speed-accuracy trade-off, we observe significant main effects of congruency (F(1,176) = 81.97, p < 0.001, ηp2 = 0.32, BF = 6.71 x 1016) and PC (F(1,176) = 25.24, p < 0.001, ηp2 = 0.13, BF = 8.22 x 108). Participants responded faster on congruent (mean = 699 ms) than incongruent (mean = 738 ms) trials, and in the low (mean = 705 ms) compared to high (mean = 732 ms) PC condition.
Finally, we again observed a significant ISPC effect irrespective of feedback group (Figure 2c; congruency x PC: F(1,176) = 13.70, p < 0.001, ηp2 = 0.07, BF = 14.89). The ISPC effect for inverse efficiency was 32 ms and 19 ms for the no feedback and feedback groups, respectively.
No other effects were significant (all other effects, F < 0.61).
Exploratory Correlations
Although we did not observe a significant transfer effect at the group-level, Bugg and Dey (2018) found evidence of a correlation between the ISPC and transfer effects, suggesting that individual differences in learning can play a role in item-based control and might prevent us from detecting a transfer ISPC at the group level in the current data set. Here, exploratory correlation analyses suggested that the ISPC RT effect in the main task was indeed predictive of an RT transfer effect (all participants: r(178) = 0.23, p = 0.002, corrected for overall RT: r(178) = 0.24, p < 0.001; no feedback group: r(88) = 0.19, p = 0.071, corrected: r(88) = 0.21, p = 0.051; feedback group: r(88) = 0.25, p = 0.018, corrected: r(88) = 0.27, p = 0.011).
Inverse efficiency scores were not predictive (all participants: r(178) = 0.00, p = 0.988, corrected for overall inverse efficiency: r(178) = 0.06, p = 0.444; no feedback group: r(88) = −0.03, p = 0.798, corrected: r(88) = 0.00, p = 0.969; feedback group: r(88) = 0.03, p = 0.776, corrected: r(88) = 0.13, p = 0.234). Notably, correlations tend to stabilize around 200 to 250 observations (Schönbrodt & Perugini, 2013), so any apparent differences between the feedback groups may not prove replicable.
Conclusion
In Experiments 1a and b, we conceptually replicated previous ISPC studies with a population of MTurk workers who had to respond manually rather than vocally (cf. Bugg, 2014; Bugg & Dey, 2018; Bugg et al., 2011). Specifically, we observed weak RT evidence (ηp2 = 0.03, BF = 0.00) suggesting that performance feedback reduces the ISPC effect and moderate Accuracy evidence (ηp2 = 0.05, BF = 4.10) suggesting that participants modulate Stroop congruency by proportion congruent (PC) context (see Supplementary Text), irrespective of performance feedback. However, when we combining RT and Accuracy into a single metric (inverse efficiency scores) to control for strong speed-accuracy tradeoffs in the data, we observed strong evidence (ηp2 = 0.07, BF = 14.89) for a reduced congruency effect for image categories that were predictive of higher compared to lower control-demand. Crucially, this ISPC effect was not modulated by feedback group (ηp2 = 0, BF = 0), indicating that the type of reactive control-learning captured by the ISPC effect is insensitive to performance feedback. Mechanistically, time demands involved in processing and incorporating performance feedback, along with the processing demands involved in responding and subsequently adjusting control settings on a moment-to-moment basis for reactive control, may not be compatible (cf. Chiew & Braver, 2013, 2016).
One limitation in this study is the speed-accuracy tradeoff, which likely stems from the particular task demands (cf. Ruitenberg, Braem, Du Cheyne, & Notebaert, 2019). Previous ISPC studies using vocal responses (cf. Bugg, 2014; Bugg & Dey, 2018; Bugg et al., 2011) have mostly detected very small error rates (e.g., <1% for congruent trials across contexts). A recent mouse-tracking version of the ISPC paradigm (Bundt, Ruitenberg, Abrahamse, & Notebaert, 2018) also had overall error rates below 1% (Personal Communication, January 24th). It is thus possible that previous studies have not detected speed-accuracy trade-offs due to ceiling level accuracy. Moreover, participants in the current study clearly followed instructions, as they passed a strict accuracy threshold for inclusion in the analyses. This suggests that the speed-accuracy trade-off is not likely result of low motivation/poor performance.
A second limitation in this study was the consistent but comparably small congruency effects. Having participants categorize the pictures using an arbitrary manual response mapping may reduce the level of processing conflict experienced relative to using vocal responses. Indeed, vocal keys and typewritten responses have been shown to produce larger congruency effects (Logan & Zbrodoff, 1998) than manual key presses. Although this study produced highly significant and consistent congruency effects, having smaller overall congruency effects may make observing modulation of these effects by proportion congruent manipulations more difficult, which may have contributed to the unreliable ISPC effects in RT data. However, ISPC effects were robust in both the accuracy (see Supplementary Text) and inverse efficiency data.
Finally, almost all other ISPC studies involved undergraduate students who likely have less familiarity with psychological tasks than experienced MTurk workers (Bugg et al., 2011; Bugg & Dey, 2018; Bundt et al., 2018; see, though, older participants in Bugg, 2014 and for review of MTurk and nonnaivete, Chandler, Mueller, & Paolacci, 2014; Stewart, Chandler, & Paolacci, 2017). Notably, we did not ask participants how much experience they have with cognitive tasks, so we are unable to determine to what extent population differences or response modality each contributed to the results we’ve observed. However, we primarily view the difference in populations as a strength, since we cannot make claims about generalizability of control-learning findings if most ISPC studies that control for associative confounds are performed on WEIRD (Western, Educated, and from Industrialized, Rich, and Democratic countries) undergraduate students.
Unlike previous research (Bugg & Dey, 2018; Bugg et al., 2011), learned control-demand associations did not transfer to non-predictive category exemplars, perhaps due to particular task demands as noted earlier. We hypothesized in Experiment 1b that changing the word associated with each category-level exemplar from the main task runs to the transfer run (i.e., far transfer) may have disrupted the memory mechanisms (cf. Egner, 2014; Hommel, 2004) needed to retrieve the appropriate control settings, resulting in a lack of transfer across the category-level association. However, even in Experiment 1b, we did not find evidence of transfer at the group-level across all our performance metrics. Notably, because we did not find group-wide evidence of the transfer of learned control-demand associations, we also cannot claim that our ISPC results are unbiased by stimulus-specific associative learning confounds (cf. Braem et al., 2019). Indeed, although the proportion congruency manipulation was intended for image categories, because participants do not show transfer, we assume that they formed stimulus-specific rather than categorical associations with control-demand.
Finally, there was a significant, positive correlation between the RT ISPC effect (PC-75/25 items) and an RT transfer effect (PC-50 items) (cf. Bugg & Dey, 2018). Even when the ISPC effect is robust, the relationship between pure control-learning (transfer), with no associative learning confounds, and learned item-specific associations is weak (here: r(178) = 0.23; Bugg & Dey (2018): r(214) = 0.18; 3-5% of the variance explained). Future research should investigate under which conditions transfer is most optimal and be designed more explicitly to assess how performance feedback impacts pure control-learning.
In sum, Experiment 1 provided strong evidence that when correcting RT for accuracy via inverse efficiency scores, an intrinsic motivator such as performance feedback does not noticeably impact the ISPC effect.
Experiment 2a:
The aim of Experiment 2 was to ask the same question in the context of the list-wide proportion congruent (LWPC) paradigm, examining the effect of performance feedback on global or proactive, anticipatory control. In LWPC paradigms, control-demand is probabilistically manipulated across blocks of trials, which may facilitate control-learning through list-wide, global control; stimulus-driven, item-specific control; or a combination of both, as well as allowing for possible contingency-learning effects. For instance, individual image categories can predict stimulus congruency and be influenced by an overall list-wide, mostly congruent or incongruent context (both stimulus-driven and list-wide control). To dissociate between the contribution of the list-wide and item-specific influences on modulating Stroop congruency, while also controlling for contingency learning confounds, we here included both items that were non-predictive (PC-50) and strongly predictive (PC-85/15) (cf. Bugg & Chanani, 2011). The predictive items created an overall PC-75/25 list context, while the non-predictive items indexed control-learning biased at only the list-level. We subsequently refer to any differences in Stroop congruency for PC-50 items in low vs. high control-demand lists as the unbiased list-wide proportion congruent (LWPC) effect and any differences in Stroop congruency for PC-85/15 items as the biased ISPC/LWPC effect.
If the LWPC paradigm reflected learned proactive control (Bugg, 2017) and proactive control were costly yet sensitive to reward (Braver, 2012), we should observe a stronger LWPC effect for the feedback group. However, if the primary mechanism for control-learning is associative learning, sensitive to any instance of reinforcement, we should observe a comparable LWPC effect across feedback groups. Irrespective of these rival hypotheses, we expected to replicate the basic LWPC effect in the no-feedback group, which corresponds to the design of previous work (Bugg & Chanani, 2011), and we also expected to replicate the proportion congruent and congruency interaction that we observed in Experiment 1, specifically for the images that were biased at both the item and list level (the ISPC/LWPC effect).
Method
Participants
Seventy-two MTurk workers consented to participate and were compensated $3.64 for approximately twenty-eight minutes. Twelve workers were excluded because of poor accuracy (i.e., less than 70% accuracy, range [17.0%, 65.1%], M = 45.6% ± 14.7%), resulting in a final sample size of sixty (feedback (N = 30): meanage = 30.6 ± 8.4; 19F, 11M; no feedback (N = 30): meanage = 34.6 ± 10.5; 10F, 20M).
Stimuli
Stimuli consisted of six separate category exemplars rather than the four categories of Experiment 1, as shown in Figure 1d and Table 2.
Table 2.
List-wide Proportion Congruency Design Manipulation.
| Mostly Congruent (MC) | Mostly Incongruent (MIC) | |
|---|---|---|
| ISPC/LWPC, PC-85/PC-15 Items | ||
| Congruent | 85% (44 trials) | 15% (8 trials) |
| Incongruent | 15% (8 trials) | 85% (44 trials) |
| LWPC, PC-50 Items | ||
| Congruent | 50% (10 trials) | 50% (10 trials) |
| Incongruent | 50% (10 trials) | 50% (10 trials) |
Experimental Procedure
The experimental procedure largely followed that of Experiment 1, except with regards to the practice run, button mappings, probability manipulation, and counterbalancing. The trial timing was the same, with four main task blocks/runs of 144 trials.
The practice run in Experiment 2 consisted of 120 trials with no distractor words overlaid on the pictures. Given the inclusion of 6 stimulus types, we sought to primarily ensure that participants learned the stimulus-response mappings during the practice run (z, x, c and b, n, m respectively matched to left ring, middle, and index and right index, middle, and ring fingers). Participants responded to each mapping twenty times (randomized in order), but because this practice run was longer in duration than that of Experiment 1, we did not impose an accuracy requirement in order to continue onto the main task. Nonetheless, after the practice and each subsequent task run, participants were reminded of the six button mappings.
In the main task, PC-50 items were intermixed with PC-85 or PC-15 items to create an overall list-wide bias of PC-75/25. Specifically, each PC-85/15 item was presented forty-four and eight times in its frequent and rare trial types and each PC-50 item was presented ten times per trial type within a block/run. With two image items per probability, four of the six mappings were thus relevant per task block context, and we ensured that each probability (i.e., PC 85, 50, 15) was mapped to each hand (e.g., z, x, and c represented either PC 85, 50, or 15).
Unlike Experiment 1, we randomized image assignment to proportion congruent context and counterbalanced for block order. Block order is particularly important in list-wide manipulations of congruency, because having the same block type repeated between runs allows for longer blocks requiring a similar amount of control-demand, which are more effective at producing canonical control adaptation effects than shorter blocks requiring constant monitoring of the current attentional demand. Pilot data suggested that block order confounded the results (cf. Abrahamse, Duthoo, Notebaert, & Risko, 2013) when we used all six possible orders, so we instead only ran the block orders that extended the influence of the list-wide manipulation, i.e., two blocks of mostly congruent (MC) trials, followed by two blocks of mostly incongruent (MIC) trials (MC-MC-MIC-MIC) and two blocks of mostly incongruent trials, followed by two blocks of mostly congruent trials (MIC-MIC-MC-MC). We added block order as a between-subjects factor to the ANOVA analyses. Finally, we also controlled for contingency learning: each PC-85 was incongruent with the other PC-85 item only; the same was the case for PC-50 and PC-15 items. Because the PC-50 items occurred with the same frequency in the two PC contexts, they are therefore unconfounded with event frequency and other associative learning effects, and may index pure control-learning (cf. Braem et al., 2019).
Results and Discussion
ISPC/LWPC influenced, PC-85/15 Items
In Experiment 2a, PC-85/15 images were strongly predictive at the item-specific level of either congruent trials (low control-demand) or incongruent trials (high control-demand) and primarily served to create the overall list-wide manipulation (PC 75/25) while serving as a replication test of the Experiment 1 results. We here refer to the interaction between proportion congruent and congruency as an index of combined item-specific and list-wide control for these images (while acknowledging that this interaction may also reflect contributions from frequency-based stimulus-response learning effects). Tables S5–8 detail the RT and accuracy data.
Inverse Efficiency
When examining the overall means for each individual, we again detected strong trade-offs between reaction time and accuracy data (overall: r(58) = −0.60, p < 0.001; no feedback group: r(28) = −0.65, p < 0.001; feedback group: r(28) = −0.63, p < 0.001). Examining overall mean RTs and accuracies within conditions (see Tables S5 and S6), we observed that participants were sometimes more accurate but slower on incongruent trials. Because of this RT/accuracy trade-off, we also computed and analyzed inverse efficiency scores.
When correcting for the speed-accuracy trade-off, we observed an expected main effect of congruency (F(1,56) = 32.41, p < 0.001, ηp2 = 0.38, BF = 2438.91). Participants responded faster on congruent (mean = 688 ms) than incongruent (mean = 755 ms) trials. We found the expected interaction between PC and block order, F(1,56) = 8.25, p = 0.006, ηp2 = 0.13, BF = 14.94. Participants who experienced the mostly congruent context first had larger differences in corrected RT between the contexts (771 (PC-85) vs. 695 (PC-15) ms) than participants who experienced the mostly incongruent context first (698 vs. 722 ms). Participants also showed a trend towards responding more slowly for the high vs. low PC context when they had received feedback and experienced the mostly congruent context first (MC first: 48 (feedback) vs. 104 ms (no feedback); MIC first: 9 vs. −56 ms) (PC x feedback x block order: F(1,56) = 3.04, p = 0.087, ηp2 = 0.05, BF = 0.30). Participants further showed a trend towards smaller congruency differences when they received feedback (47 ms) than when they did not (87 ms) (congruency x feedback: F(1,56) = 2.84, p = 0.098, ηp2 = 0.05, BF = 0.19).
Participants across feedback groups also displayed significantly reduced congruency effects for high vs. low control-demand cues (PC x congruency: F(1,56) = 9.56, p = 0.003, ηp2 = 0.15, BF = 8.26). This ISPC/LWPC effect interacted with block order; specifically, participants who experienced mostly incongruent blocks first (mean: 14 ms) had a smaller congruency difference between low vs. high control-demand contexts than participants who experienced mostly congruent blocks first (mean: 148 ms; PC x congruency x block order: F(1,56) = 6.62, p = 0.013, ηp2 = 0.11, BF = 2.73), irrespective of their feedback group (PC x congruency x feedback: F(1,56) = 0.00, p = 0.967, ηp2 = 0.00, BF = 0.07). This is consistent with previous literature suggesting that there are asymmetrical costs when participants switch from mostly incongruent to congruent lists and mostly congruent to incongruent lists and that these costs result from different attentional recruitment (Abrahamse et al., 2013).
No other effects were significant (PC: F(1,56) = 2.31, p = 0.134, ηp2 = 0.04, BF = 10.21; all other effects, F < 0.76).
Overall, the two-way interaction between PC and congruency for RT, accuracy, and inverse efficiency, in the absence of the three-way interaction including feedback, suggests that we here replicate Experiment 1 and find that the ISPC is invariant to performance feedback.
LWPC, PC-50/50 Items
In Experiment 2, PC-50 images were non-predictive at the item-specific level and could therefore be influenced only by the overall list-wide manipulation (PC 75/25). The list-wide proportion congruent effect (LWPC) is denoted by the interaction between PC and congruency for these non-predictive PC-50 items, which were only biased on a global level due to the presence of strongly predictive PC-85/15 items within the list.
Inverse Efficiency
When examining the overall means for each individual, we again detected strong trade-offs between reaction time and accuracy data (overall: r(58) = −0.66, p < 0.001; no feedback group: r(28) = −0.68, p < 0.001; feedback group: r(28) = −0.65, p < 0.001). Examining overall mean RTs and accuracies within conditions (see Tables S7 and S8), we observed that participants were sometimes more accurate but slower on incongruent trials.
When correcting for the speed-accuracy trade-off with inverse efficiency scores, we observed an expected main effect of congruency (F(1,56) = 13.10, p < 0.001, ηp2 = 0.19, BF = 0.51) and a trending main effect of context (PC: F(1,56) = 2.88, p = 0.095, ηp2 = 0.05, BF = 7.18). Participants responded more quickly on congruent (mean = 797 ms) than incongruent (mean = 840 ms) trials, and in the low (mean = 797 ms) compared to high (mean = 840 ms) PC condition.
We replicate Abrahamse et al., (2013), observing a marginal modulation of the LWPC effect by block order (PC x congruency x block order: F(1,56) = 3.13, p = 0.082, ηp2 = 0.05, BF = 0.08). Participants who experienced mostly congruent blocks first tended to display larger LWPC effects (means: 63 ms vs. −15 ms). We also found the expected effect of block order (PC x block order: F(1,56) = 5.87, p = 0.019, ηp2 = 0.10, BF = 14.47), whereby participants had smaller differences in inverse efficiency across contexts when they first experienced the mostly incongruent context (809 (PC-25) vs. 790 ms (PC-75)) rather than mostly congruent context (785 vs. 889 ms). Participants also tended to have larger congruency differences when they did not receive feedback and experienced the mostly incongruent context first (MC first: 40 (feedback) vs. 35 (no feedback); MIC first: 9 vs. 86 ms) (congruency x feedback x block order: F(1,56) = 3.19, p = 0.080, ηp2 = 0.05, ηp2 = 0.05, BF = 0.02).
As shown in Figure 3a, we also observed significant modulation of the LWPC effect by feedback group (PC x congruency x feedback: F(1,56) = 5.20, p = 0.026, ηp2 = 0.09, BF = 0.02). Specifically, the feedback group showed strong modulation of congruency effects by PC context (mean: 74 ms, PC x congruency: F(1,28) = 8.40, p = 0.007, ηp2 = 0.23, BF = 0.28), while the no feedback group showed little evidence of the critical contextual interaction (mean: −26 ms, PC x congruency: F(1,28) = 0.54, p = 0.469, ηp2 = 0.02, BF = 0.27). Because we observed a greater modulation of Stroop congruency across low vs. high control-demand contexts for the feedback group than for the no-feedback group for non-predictive PC-50 items, we conclude that the LWPC, and thus proactive control, is sensitive to performance feedback.
Figure 3. Experiment 2 control-learning results.
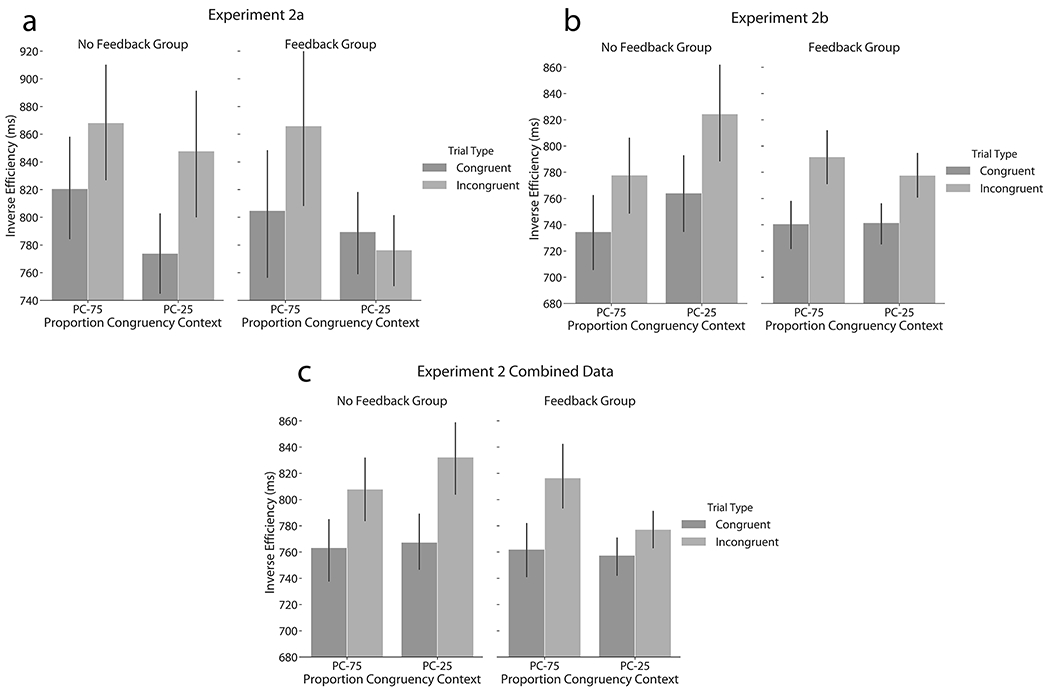
Mean Inverse Efficiency data (± SEM) are displayed as a function of proportion congruent context, current trial congruency, and feedback group for (a & b) the LWPC effect of Experiments 2a and b, which are averaged together in (c).
No other effects were significant (congruency x feedback: F(1,56) = 2.46, p = 0.122, ηp2 = 0.04, BF = 0.10; all other effects, F < 1.18).
Explicit Understanding of Task Structure
Twenty-five participants reported noticing that some images systematically varied in their proportion congruency, while twenty-eight reported noticing differences in difficulty across task blocks. Twenty-three participants reported noticing differences for both questions. Although participants did not rate the predictive image category pairings as being more predictive than chance (50%; ts < −0.26), they were able to match the image categories to their respective congruencies (easy/neutral/hard) above chance (i.e., test against 2 for response option chance level given 6 images; t(59) = 2.32, p = 0.024, Cohen’s dz = 0.30, M = 2.42). However, this was primarily driven by better matching of the biased ISPC/LWPC images to their trial type (i.e., test against 4/3 for response option chance level given 4 images; t(59) = 2.71, p = 0.009, Cohen’s dz = 0.35, M = 1.75) relative to the unbiased LWPC images (i.e., test against 2/3 for response option chance level given 2 images; t(59) = 0.07, p = 0.948, Cohen’s dz = 0.01, M = 0.67), which were below chance recognition. In short, although there was an increased level of awareness surrounding the PC manipulation relative to Experiment 1 (cf. Blais, Harris, Guerrero, & Bunge, 2012), perhaps due to the increased PC frequencies (e.g., 85 vs. 75), the unbiased LWPC effect likely remained implicit for participants.
Experiment 2b
Because we expected to replicate standard LWPC effects for the no feedback group in Experiment 2a and because we observed a speed-accuracy trade-off, we opted to run a larger replication sample. Like Experiment 1b, to discourage the speed-accuracy trade-off, we added in an additional line about responding as quickly and accurately and restricted the participant sample to people who had a mailing address in the United States.
Method
Participants
We followed the approach of Experiment 1b, opting to double the sample size in Experiment 2a for this replication sample.
One hundred and forty-two MTurk workers consented to participate and were compensated $3.64 for approximately twenty-eight minutes. Twenty-one workers were excluded because of poor accuracy (i.e., less than 70% accuracy, range [1.4%, 65.2%], M = 37.3% ± 20.1%). Another worker was excluded for having the same IP address as another participant; we could not unambiguously determine that these were different individuals. The final sample size was one hundred and twenty (feedback (N = 60): meanage = 36.2 ± 10.2; 25F, 35M; no feedback (N = 60): meanage = 34.7 ± 9.2; 30F, 29M, 1 n/a).
Results and Discussion
Tables S9–12 detail the RT and accuracy data.
ISPC/LWPC influenced, PC-85/15 Items
Inverse Efficiency
However, when examining the overall means for each individual, we detected strong trade-offs between reaction time and accuracy data (overall: r(118) = −0.56, p < 0.001; no feedback group: r(58) = −0.54, p < 0.001; feedback group: r(58) = −0.52, p < 0.001). Examining overall mean RTs and accuracies within conditions (see Tables S9 and S10), we observed that participants were sometimes more accurate but slower on incongruent trials. Because of a strong RT/accuracy trade-off, we computed and analyzed inverse efficiency scores.
When correcting for the speed-accuracy trade-off, we observed an expected main effect of congruency (F(1,116) = 41.58, p < 0.001, ηp2 = 0.26, BF = 1.02 x 108). Participants responded more quickly on congruent (mean = 668 ms) than incongruent (mean = 720 ms) trials. We found expected effects of block order and context (PC x block order: F(1,116) = 5.07, p = 0.026, ηp2 = 0.04, BF = 1.97), whereby participants had smaller differences in inverse efficiency across contexts when they first experienced the mostly incongruent context (680 (PC-15) vs. 672 (PC-85)ms) rather than mostly congruent context (692 vs. 732 ms).
Participants across feedback groups also displayed a significant ISPC/LWPC effect, showing reduced congruency effects for high vs. low control-demand contexts (PC x congruency: F(1,116) = 29.04, p < 0.001, ηp2 = 0.20, BF = 4091.42). Unlike Experiment 2a, this ISPC/LWPC effect did not interact with block order (PC x congruency x block order: F(1,116) = 1.51, p = 0.222, ηp2 = 0.01, BF = 0.25) or feedback group (PC x congruency x feedback: F(1,116) = 0.97, p = 0.328, ηp2 = 0.01, BF = 0.02). The ISPC/LWPC effect for inverse efficiency was 72 and 104 ms for the no feedback and feedback groups, respectively.
No other effects were significant (PC: F(1,116) = 2.14, p = 0.146, ηp2 = 0.02, BF = 1060.78; congruency x feedback: F(1,116) = 1.93, p = 0.168, ηp2 = 0.02, BF = 0.09; block order: F(1,116) = 2.23, p = 0.138, ηp2 = 0.02, BF = 0.58; all other effects, F < 0.53).
Overall, in Experiment 2b, we replicate the congruency by proportion congruent context interaction we observed with RT, accuracy, and IES in Experiment 2a, providing further evidence that the ISPC effect is insensitive to performance feedback.
LWPC, PC-50/50 Items
Inverse Efficiency
When examining the overall means for each individual, we again detected strong trade-offs between reaction time and accuracy data (overall: r(118) = −0.61, p < 0.001; no feedback group: r(58) = −0.71, p < 0.001; feedback group: r(58) = −0.43, p < 0.001).
When correcting for the speed-accuracy trade-off with inverse efficiency scores, we observed an expected main effect of congruency (F(1,116) = 31.83, p < 0.001, ηp2 = 0.22, BF = 1292.18). Participants responded more quickly on congruent (mean = 745 ms) than incongruent (mean = 793 ms) trials. We found expected effects of block order (PC x block order: F(1,116) = 5.65, p = 0.019, ηp2 = 0.05, BF = 5.18; block order: F(1,116) = 4.01, p = 0.047, ηp2 = 0.03, BF = 2.28), as participants responded more quickly when they first experienced the mostly incongruent context (738 ms) than when they experienced the mostly congruent context (800 ms), and had smaller differences in inverse efficiency across contexts when they first experienced the mostly congruent context (792 (PC-25) vs. 808 (PC-75) ms) rather than mostly incongruent context (761 vs. 714 ms). Participants also showed a slight tendency to respond slower to the low vs. high PC context when they did not receive feedback (−38 ms) than when they did (7 ms) (PC x feedback: F(1,116) = 2.82, p = 0.096, ηp2 = 0.02, BF = 0.61).
As shown in Figure 3b, unlike Experiment 2a, we did not observe significant modulation of the LWPC effect by feedback group (PC x congruency x feedback: F(1,116) = 1.79, p = 0.183, ηp2 = 0.02, BF = 0.01), but did by block order (PC x congruency x block order: F(1,116) = 5.27, p = 0.024, ηp2 = 0.04, BF = 0.05). The LWPC effect for the respective feedback groups was −17.2 and 15.0 ms. Participants who experienced mostly congruent blocks first tended to display larger LWPC effects (means: 27 ms (18 for no feedback, 35 for feedback group) vs. −29 ms (−53 ms for no feedback, −5 ms for feedback group)).
No other effects were significant (all other effects, F < 1.40).
Overall, we have found some evidence that the LWPC effect is sensitive to performance feedback, detecting the three-way congruency x context feedback interaction with accuracy and IES in Experiment 2a and here again with RT in Experiment 2b. Both groups also showed expected effects of block order, with larger LWPC effects, if they experienced the mostly congruent context first.
Explicit Understanding of Task Structure
Fifty-one participants reported noticing that some images systematically varied in their proportion congruency, while forty-eight reported noticing differences in difficulty across task blocks. Forty participants reported noticing differences for both questions. Participants rated the predictive image category pairings as being less predictive than chance (50%; all ts > 2.49). They were also able to match the image categories to their respective congruencies (easy/neutral/hard) above chance (i.e., test against 2 for response option chance level given 6 images; t(116) = 3.33, p = 0.001, Cohen’s dz = 0.31, M = 2.40). As in Experiment 2a, this was primarily driven by better matching of the biased ISPC/LWPC images to their trial type (i.e., test against 4/3 for response option chance level given 4 images; t(116) = 4.31, p < 0.001, Cohen’s dz = 0.40, M = 1.81) relative to the unbiased LWPC images (i.e., test against 2/3 for response option chance level given 2 images; t(116) = 1.07, p = 0.287, Cohen’s dz = 0.10, M = 0.59), which were below chance recognition. Replicating Experiment 2a, the unbiased LWPC effect likely remained implicit for participants.
Across Experiment 2 Analysis
Because Experiments 2a and 2b shared similar task structures, we aggregated our data across experiments for exploratory analyses with enhanced power. We added experiment (2a/2b) as a between-subjects factor and reran analyses across RT, accuracy, and inverse efficiency data to better evaluate the effect of performance feedback on the unbiased LWPC effects. We do not analyze the biased ISPC/LWPC effect, since it already conceptually replicated the Experiment 1 results. As with the combined analysis of Experiment 1, since the design and procedure were identical across Experiments 2a and b, we treat Experiment as a factor of no interest.
Inverse Efficiency
Finally, when correcting for the speed-accuracy trade-offs by computing inverse efficiency scores for PC-50 items, we found a standard congruency effect (F(1,172) = 38.35, p < 0.001, ηp2 = 0.18, BF = 2054.76). Participants responded more quickly on congruent (mean = 762 ms) than incongruent (mean = 808 ms) trials. We also found expected effects of block order (PC x block order: F(1,172) = 12.87, p < 0.001, ηp2 = 0.07, BF = 1024.64; block order: F(1,172) = 3.19, p = 0.076, ηp2 = 0.02, BF = 217.41). Participants responded slightly more quickly when they first experienced the mostly incongruent context (758 ms) than mostly congruent context (813 ms), and had smaller differences in inverse efficiency across contexts when they first experienced the mostly incongruent context (777 (PC-25) vs. 739 (PC-75) ms) rather than mostly congruent context (790 vs. 835 ms). Participants also had slightly larger congruency differences across feedback groups when participants experienced the mostly incongruent context first (27 (feedback) vs. 65 ms (no feedback)) than when they experienced the mostly congruent context first (47 vs. 44 ms) (congruency x feedback x block order: F(1,172) = 3.16, p = 0.077, ηp2 = 0.02, BF = 0).
Crucially, we also detected a significant unbiased LWPC effect that separately depended on feedback group and block order (Figure 3c; PC x congruency x feedback: F(1,172) = 8.31, p = 0.004, ηp2 = 0.05, BF = 0.01; PC x congruency x block order: F(1,172) = 8.38, p = 0.004, ηp2 = 0.05, BF = 0.02). The LWPC effect was −20 ms and 35 ms for the no feedback and feedback groups. When the participants experienced the mostly congruent context, the LWPC effect was, respectively, 16 and 61 ms for the no feedback group and feedback groups and decreased to −57 and 8 ms when the participants experienced the mostly incongruent context first.
No other effects were significant (congruency x feedback: F(1,172) = 2.38, p = 0.125, ηp2 = 0.01, BF = 0.04; all other effects of interest, F < 1.21).
Overall, when we combined the data from Experiments 2a and 2b, we found weak RT evidence that the LWPC was insensitive to feedback, and accuracy and IES evidence that the LWPC was sensitive to feedback and block order, with both groups showing larger congruency effects if they experienced the mostly congruent context first and the feedback group showing a larger LWPC effect than the no feedback group.
Conclusion
In Experiments 2a and 2b, we replicated the item-specific proportion congruent (ISPC) effects we observed in Experiments 1a and 1b, finding that Stroop congruency for the PC-85 and PC-15 items was reduced for high compared to low control-demand, irrespective of the feedback group. However, performance feedback significantly impacted list-wide proportion congruent (LWPC) effects (cf. Bugg & Chanani, 2011). After correcting for speed-accuracy trade-offs, participants within the feedback group showed reduced congruency differences for the PC-50 items within blocks predictive of higher compared to lower control-demand. This is consistent with previous research suggesting that proactive control, here represented by the LWPC, is costly (Braver, 2012; Bugg, 2017) and that intrinsic motivation manipulations can boost proactive, but not reactive, adaptation of cognitive control. Finally, we also replicated previous research suggesting that there are asymmetrical congruency effects when participants switch from a high attentional state (mostly incongruent context) to a low attentional state (mostly congruent context), and vice versa (cf. Abrahamse et al., 2013).
Unlike previous research (e.g., Bugg & Chanani, 2011), however, participants within the no feedback group showed little modulation of Stroop congruency. We do not view this as inconclusive evidence for the LWPC effect, because as shown in our compiled analysis, we found a significant modulatory effect of block order on the LWPC. Specifically, participants who received no feedback and experienced the mostly congruent context first had a positive LWPC effect. Moreover, as we speculated in Experiment 1, the speed-accuracy trade-off, as well as reduced congruency effects relative to previous LWPC studies that used vocal responses and tested undergraduate students, may impact the LWPC we observed here in unknown ways. Although participants were reminded of the picture-response mappings between each block and four mappings were only used during each block, having participants keep in mind six picture-mappings could have created additional working memory demand that distracted from the learned recruitment of costly control.
One notable limitation in Experiment 2 is the complexity of the design, with four to five factors, half of which were between-subjects (block order, feedback, Experiment). This is especially evident in the Bayes Factor analysis, which assigns greater prior weight when there are more supporting data (e.g., 0.978 for main effect of block type, but 0.265 for a three-way interaction in the combined Experiment analysis). Although we replicated the congruency x feedback x context interaction with a large sample size (N=180) and the asymmetrical congruency effects observed in another study (Abrahamse et al., 2013), the Bayes Factors for both of these interactions effects are relatively small compared to those observed in Experiment 1. Design-wise, between-subjects effects increase variability within the analysis cells, leading to more variable estimates as to the strength of evidence within these Experiments. Moreover, to deconfound the unbiased LWPC effect from the biased ISPC/LWPC effect means that there are fewer trials for the PC-50 items relative to the PC-85/15 items, which causes additional variance within each participant and the analysis. Together these pieces add additional nuance to the story.
Finally, one crucial factor unexamined within this study is the presence of individual differences. Previous research has found that proactive control is subject to inter-individual differences such as working memory capacity and reward responsiveness (Braver, 2012). The analysis of inverse efficiency scores across Experiments 2a and 2b suggested that only 48% of the variance in behavior was explained by the ANOVA model, highlighting how individual differences could also play a key role in how performance feedback impacts the learned recruitment of reactive and proactive control.
In sum, when correcting RT for accuracy via inverse efficiency scores, we observed a significant unbiased LWPC effect that was modulated by performance feedback. Taking Experiment 1 into account, these Experiment 2 results additionally suggest that intrinsic motivators like performance feedback can promote the recruitment of proactive control and have little impact on reactive control.
Exploratory across Experiments 1 and 2 Analysis
In Experiment 1, we observed no modulation of the ISPC effect by feedback, whereas in Experiment 2, we found that the LWPC effect was significantly larger when participants received feedback. The results in Experiments 1 and 2 thus suggest that performance feedback promotes proactive, but not reactive, adaptation of cognitive control. Because the paradigms both involve similar manipulations of proportion congruency with comparable stimuli, we were able to formally test whether feedback does in fact differentially affect proactive and reactive control by pooling the data and treating experiment as a between-group factor.
To test this dissociation explicitly, we combined the data across Experiments and ran a 2 x 2 x 2 x 2 repeated-measures ANOVA on inverse efficiency scores, with proportion congruent (mostly incongruent/mostly congruent) and congruency (congruent/incongruent) as within-subjects factors and experiment (1/2) and feedback (feedback/no feedback) as between subjects factors. For Experiment 1, we used the data from the PC-75/25 items, which reflect the ISPC, and for Experiment 2, we used the data from the PC-50 items, which reflect the LWPC. Notably, we could not include the block order factor from Experiment 2 and account for the additional response demands in the LWPC (i.e., 6 response buttons), which may result in some unexplained variance in this analysis.
We observed a significant 4-way interaction, indicative of a dissociation between the effect of feedback on reactive vs. proactive control learning on inverse efficiency scores (PC x congruency x feedback x Experiment: F(1,356) = 6.85, p = 0.009, ηp2 = 0.02, BF = 0; PC x congruency x feedback: F(1,356) = 2.60, p = 0.108, ηp2 = 0.01, BF = 0; PC x congruency x Experiment: F(1,356) = 1.98, p = 0.160, ηp2 = 0.01, BF = 0.02; PC x congruency: F(1,356) = 6.36, p = 0.012, ηp2 = 0.02, BF = 0). Performance feedback selectively promoted the recruitment of proactive vs. reactive control in the list-wide relative to item-specific paradigm.
General Discussion
This study examined whether an intrinsic reinforcer (performance feedback) has differential effects on the learned recruitment of reactive and proactive control within a variant of the picture-word Stroop task. In Experiment 1, we deployed an item-specific proportion congruent (ISPC) paradigm, manipulating in-the-moment, reactive control by ensuring that some picture images were more often overlaid with incongruent distractor words (low proportion congruent, high control-demand) while others were more often overlaid with congruent distractor words (high proportion congruent, low control-demand). In Experiment 2, we deployed a list-wide proportion congruent (LWPC) paradigm, manipulating anticipatory, proactive control by ensuring that some images were equally congruent and incongruent at the item-level but biased at the global list-level within the context of low or high control-demand blocks. For both experiments, the receipt of performance feedback was manipulated between groups.
In the context of explicitly instructed cues in the AX-CPT task, it has previously been shown that proactive control is costly to recruit, but sensitive to reward, while reactive control is insensitive to reward (Braver, 2012). We here tested rival hypotheses about how performance feedback would impact control-learning in response to implicit manipulations of task statistics. If control-learning is grounded in reinforcement learning, sensitive to reinforcement but not to the type of control-demand, we would observe enhanced recruitment of control in both Experiments. Specifically, for both feedback groups, we would observe a standard proportion congruent (PC) effect, whereby the congruency effect is reduced for high vs. low control-demand contexts. If, however, the type of control-demand learned is important, with proactive control sensitive to reinforcement and reactive control insensitive to reinforcement, we would observe a PC effect for both feedback groups in Experiment 1 and a PC effect only for the feedback group in Experiment 2.
We found that performance feedback promoted the learned recruitment of proactive but not reactive conflict-control when we corrected for substantial speed-accuracy trade-offs detected in the data. Analyzing inverse efficiency scores, we observed a PC effect that was invariant to feedback group in the ISPC for Experiment 1 and a PC effect only for the feedback group in the LWPC for Experiment 2. Our combined analyses of the experiments indeed suggested a significant dissociation between how performance feedback shapes control-learning.
We now discuss the theoretical importance of these results in the following sections on associative learning of cognitive control, interactions between motivation and cognitive control, and brief speculation on the neural systems involved in control-learning.
Associative Learning of Cognitive Control
Some research has suggested that PC effects are not control-based and that associative learning of stimulus-responses associations may instead account for the observed data patterns (i.e., contingency-learning, Schmidt & Besner, 2008; Schmidt, 2013). For example, in a Stroop task with four color-word stimuli, there are more possible unique incongruent than congruent stimuli (e.g., for the color-word BLUE, blue in RED, PURPLE, and BROWN vs. blue in BLUE). When researchers ensure that the overall congruency is equal (50/50) while manipulating proportion congruency, each congruent stimulus (total: 4) appears more often than each incongruent stimulus (total: 12). This then means that participants can learn stimulus-response associations, because the irrelevant dimension (e.g., the meaning of the color-word) more often predicts congruent stimuli. Participants could simply learn to read the color-words instead of learning to adjust their attentional selectivity, because the color-word RED is more often paired with the color red. The contingency learning account thus predicts that participants are slower to respond on low contingency (incongruent) trials vs. high contingency (congruent) trials, resulting in smaller congruency differences in a low-PC context that stems from pure S-R learning instead of control adjustments.
However, recent accounts have highlighted how, when the target-relevant dimension is predictive of proportion congruency, the ISPC avoids such contingency learning confounds (Bugg et al., 2011). Using the target (here, a picture), instead of the task-irrelevant word, as the ISPC signal for proportion congruency results in equivalent contingency across all possible combinations of the congruency and proportion congruency conditions. Moreover, we controlled for the formation of stimulus-response associations by ensuring that there was only one incongruent and congruent stimulus pair so that participants could not associate one picture more often with a particular response. While this particular design does still allow for participants to modulate word-reading rather than the attentional state recruited, the pattern of results are more consistent with a control-based account (Experiment 1 vs. Experiment 3 within Bugg et al., 2011). Specifically, if participants were modulating word-reading, the results would show modulation of Stroop congruency primarily with regard to congruent trials, where this strategy would be most efficient. The modulation of Stroop congruency across both Experiments, however, seems to have resulted from more efficient responding to incongruent trials than to congruent trials, suggesting a control-based account prevailed.
Other control-based accounts have also emphasized that a pure associative S-R learning explanation cannot account for transfer items and asymmetrical congruency effects in list-wide paradigms. If participants were merely modulating word-reading or using stimulus-response associations, we should not observe carry-over effects when changing between contexts for a list-wide proportion congruency task because the associations and strategy have remained the same (Abrahamse et al., 2013). However, we do, in fact, observe modulation by block order for both LWPC and ISPC effects, suggesting that participants recruit different control states across the high and low control-demand blocks. Finally, we also find list-wide effects even when controlling for item-specific associations (Bugg & Chanani, 2011). Altogether these results suggest that particular contexts (items/pictures or blocks) facilitated the retrieval of context-appropriate attentional control-states (e.g., high or low attentional focus).
Of note, we also found greater explicit awareness of the item-level manipulation when the proportion congruency increased (cf. Blais et al., 2012). The cognitive mechanisms and strategies at play between individuals who do and do not notice the underlying task structure may differ substantially. Indeed, in a recent preregistered direct replication study (Bejjani, Dolgin, Zhang, and Egner, 2019, https://osf.io/7jfbp/), we observed that participants were most effective at adjusting their cognitive control when they could consciously perceive predictive cues and had explicit knowledge of what the cues predicted. However, while most control-based accounts of item- or list-level imply that control adaptation occurs implicitly, these frameworks still allow for explicit control adaptation.
Indeed researchers have more recently posited that associative learning and cognitive control are inextricably linked (Abrahamse et al., 2016; Botvinick et al., 2001; Egner, 2014). Participants can learn to recruit more attentional focus for stimuli or contexts that are more often associated with incongruent/hard trials (i.e., higher control-demand), which facilitates greater processing efficiency and results in smaller congruency effects relative to stimuli or contexts that are more often associated with congruent trials (i.e., lower control-demand). The nature of the learning process is thought to be domain-specific (task-switching, conflict-control, attentional control, etc.), while incorporating a domain-general associative learning mechanism.
How exactly participants learn to generalize specific stimulus features and responses into abstract, context-appropriate attentional control-states and bring about strategic processing adjustments nonetheless remains poorly understood. In the expected value of control account (Shenhav, Botvinick, & Cohen, 2013), which incorporates a domain-general and domain-specific perspective, participants could learn to recruit proactive control based on the context and cost-benefit analysis, but how does the cognitive system determine the costs and benefits? If reactive control, represented by the ISPC, is recruited in the moment, do people still calculate the costs and benefits, or does the temporal constraint cause the process to become more automatic? Here, the receipt of performance feedback does not result in additional information for the participant, because participants already know the correct buttons to press, yet performance feedback profoundly changes the learned recruitment of proactive vs. reactive control. These results highlight that even intrinsic motivation states have little effect on the learned recruitment of reactive control, which suggests that different cognitive mechanisms, beyond expected value calculations, are at work between learning adjustments of control for the ISPC and LWPC.
Interactions between Motivation and Cognition
While a wealth of research has suggested that incentivizing task performance can enhance the recruitment of cognitive control (Botvinick & Braver, 2015; Chiew & Braver, 2016; Fröber & Dreisbach, 2014; Kang et al., 2017; Kostandyan et al., 2019; Soutschek, Stelzel, et al., 2014; Soutschek, Strobach, et al., 2014; Yee & Braver, 2018), most work has focused on incentivizing task performance with external reinforcers like monetary reward (Krug & Braver, 2014) or involved reward cues to detect whether participants use additional information in adjusting the recruitment of cognitive control. One critical difference between the motivational factors involved in previous studies and the current study is that we not only used a deterministic, intrinsic reinforcer (performance feedback) but also that the intrinsic reinforcer held no inherent value; specifically, performance feedback usually provides participants with additional valuable information, but in this study, participants already knew the correct responses for each particular stimulus. This is also evident in the fact that performance feedback is not necessary to observe control-learning effects (for reviews, see Abrahamse et al., 2016; Bugg, 2017; Bugg & Crump, 2012; Egner, 2014), but would otherwise help participants encode information in other cognitive tasks (e.g., learning paired-associates). Thus, unlike extrinsic probabilistic reward cues, performance feedback either enhanced learning by strengthening control-demand associations or boosted intrinsic motivation (see below).
We can note how the differences in study design impacted the results of the few proportion congruent studies that investigated the interaction between motivation and control-learning (Bugg et al., 2015; Soutschek et al., 2015, 2014). Experiment 4 within Bugg et al.(2015) tested whether high or low point value incentives could motivate participants to use probabilistic cues that indicated whether short, twenty-trial lists were mostly congruent or mostly incongruent blocks. The researchers found similar results to their previous experiments, whereby participants used the probabilistic cues to relax their control settings on mostly congruent lists relative to mostly incongruent lists. However, performance-contingent reward impacted the recruitment of cognitive control for the first trial on mostly incongruent blocks, i.e., when participants had no learned expectations for the upcoming control-demand. This is mostly in accord with Chiew and Braver (2016), who found that participants reduced flanker interference effects the most following informative, incentivized trials, although there are potential differences in sustained vs. transient effects. Nonetheless, rather than examine how reinforcers impact how participants learn to adjust the recruitment of cognitive control, the focus of these studies was to investigate how reward impacted the use of informative cues. With additional cues that inform participants of what to likely expect, participants may feel less inclined to modulate their attentional settings for high control-demand blocks.
Soutschek and colleagues (2014a, b) investigated how reward cues informing participants whether they could earn or lose money impacted the effect of conflict expectancy in a list-wide proportion congruent paradigm with blocks consisting of ten or forty trials. They found that participants had smaller congruency effects for high vs. low conflict expectancy for blocks where they could neither earn nor lose money and that participants had smaller congruency effects for high stakes blocks across conflict expectancy. The researchers interpret these results to mean that motivational factors like reward and conflict expectancy could both reduce congruency effects independently, but not so much as to eliminate congruency effects altogether. However, like Bugg et al. (2015), Soutschek and colleagues (2014a, b) also used a reward/punishment framework with high and low motivation cues on a block-wise level, within-participants, primarily investigating how motivation changes expectancy or expected value judgements for recruiting cognitive control, subject to framing effects. The current study, by contrast, focused on how the experience of success or failure itself, on a trial-by-trial basis, could motivate participants to modulate learned adjustments of control across contextual blocks longer than one hundred trials. Thus, rather than finding independent effects of conflict expectancy (proportion congruent) and motivation, we found that the two interacted to determine sustained, learned adjustments to cognitive control.
All these studies together, however, suggest that control-learning is subject to framing effects at large. The present study cannot adjudicate between two rival hypotheses: performance feedback may have either enhanced the learning of control-demand associations or boosted intrinsic motivation for applying learned control. For instance, boosting motivation could have occurred by increasing the intrinsic sense of achievement upon performing well (cf. Schouppe et al., 2015) or lessening the subjective aversion that participants feel towards conflict (cf. Dreisbach & Fischer, 2012), ultimately causing participants to recruit proactive control in Experiment 2. Future studies will need to adjudicate between these possibilities or the degree to which feedback promotes both of these processes (which are not mutually exclusive). Moreover, while researchers have trained reinforcement learning models to predict control-demand rather than reward, future research could also test whether models trained to predict probabilistic feedback on control-learning tasks result in similar learned adjustment of control or whether the additional uncertainty dampens the effect of control-learning altogether.
Of note, although we refer to the LWPC effect as reflecting proactive control, we cannot be certain that participants actually recruit control in an anticipatory fashion prior to each stimulus onset. Most studies of proactive control using the AX-CPT or other congruency tasks (e.g., flanker task) manipulate whether trial-by-trial cues are informative or non-informative of upcoming control-demand, thus investigating whether participants will recruit control proactively during the cue-to-target interval. Even though it is clear that the LWPC effect is “global” in the sense of generalizing across stimuli/trials within a block, it is not currently clear whether it is driven by the same kind of anticipatory control engagement probed by trial-by-trial curing paradigms. Answering this question represents an interesting direction for future research. A couple of studies have suggested that explicit precues can either enhance or diminish control-learning effects (Bugg & Diede, 2018; Hutchison, Bugg, Lim, & Olsen, 2016), especially in contexts of lower control demand (e.g., mostly congruent blocks).
How do these processes interact in the brain?
Recent studies have implicated the caudate nucleus, a region typically associated either with a domain-general valuation system that learns stimulus-reward outcome or stimulus-action contingencies (Balleine, Delgado, & Hikosaka, 2007; Delgado, 2007) or with a working memory gating system (Frank, Loughry, & O’Reilly, 2001), in control-learning (Chiu et al., 2017; Jiang, Beck, et al., 2015; Jiang, Brashier, & Egner, 2015; for review, see Chiu & Egner, 2019). While the exact cognitive mechanism performed by the caudate in control-learning is not known, the caudate is nonetheless known to be sensitive to performance feedback (Tricomi, Delgado, McCandliss, McClelland, & Fiez, 2006; Tricomi & Fiez, 2008; Bejjani, DePasque, & Tricomi, 2019; for review, see Tricomi & DePasque, 2016).
Of note, in this study, we found that the ISPC was invariant to performance feedback. However, Chiu et al., (2017) trained a reinforcement learning model on data from a similar, no feedback item-specific proportion congruent paradigm and found that prediction error estimates obtained from the model correlated with activation in the caudate nucleus. If the caudate is indeed sensitive to performance feedback yet also tracks the kind of item-specific control-learning reported here, which was invariant to performance feedback, future research will need to determine the exact role of the caudate in control-learning. Grandjean et al. (2013) and Blais & Bunge (2009) also investigated the neural correlates of the ISPC and found mostly prefrontal regions’ activation track the interaction of context and congruency, suggesting that perhaps coordination between cortical and subcortical regions determines the learned control adjustments across control-demand contexts. Future work will need to disentangle cortical and subcortical contributions to control-learning across domains (cf. Chiu & Egner, 2019).
Understanding the exact contributions of the reward and executive function systems will be important to determining how control-learning is impaired in psychiatric populations. Many clinical conditions, such as schizophrenia (Barch & Sheffield, 2017), are characterized by context-inappropriate behavior. These are typically attributed to a lack of control, but might instead arise from a deficit in control-learning, whereby afflicted individuals may not learn to associate certain cues or contexts with the need for control, resulting in inflexible behavior. A better understanding of the interaction between control, reward, and memory mechanisms in the healthy brain will thus lay the foundations for understanding potential control-learning failure models – and possible therapeutic approaches – in psychiatric and neurodegenerative disorders.
Generalization of Control-Learning
Notably, the results reported in this study were observed using an online population of Amazon Mechanical Turk (MTurk) workers. Studies that deployed variants of the ISPC and LWPC (Bugg, 2014; Bugg & Chanani, 2011; Bugg & Dey, 2018; Bugg et al., 2011) have typically used vocal responses within an in-person, undergraduate participant population (see Bundt et al., 2018; Ruitenberg et al., 2019 for mouse-tracking ISPC). Having developed an ISPC and LWPC for a crowd-sourced population, we propose that using performance feedback is generally beneficial for all control-learning studies: performance feedback does not affect the recruitment of reactive control (ISPC effect), lowers the attrition rate based on accuracy demands (i.e., fewer excluded participants), and boosts the recruitment of proactive control (LWPC effect) so that researchers can investigate the boundary conditions and individual differences that might affect control-learning. One notable limitation in adapting the item-specific and list-wide proportion congruency paradigms for online populations, however, was the speed-accuracy trade-offs we observed in both Experiments. Future work could examine the potential causes of the speed-accuracy trade-off (e.g., not having ceiling accuracy, participant population differences, response demand characteristics). However, despite the speed-accuracy trade-offs, we believe that these data are as representative of the ISPC and LWPC effects, since participants clearly followed instructions and performed above strict accuracy thresholds.
Conclusion
This study examined how performance feedback shapes control-learning across the item-specific and list-wide proportion congruent paradigms. We found that performance feedback promoted learned proactive adjustment to control-demand across blocks with a greater proportion of hard vs. easy trials, but had little effect on learned reactive adjustment for particular items associated with a higher likelihood of high vs. low control-demand. These results have important implications for our basic understanding of adaptive control, and potentially for treating clinical populations in which proactive control is impaired, but can be boosted with intrinsic reinforcers like performance feedback.
Supplementary Material
Public Significance Statement.
Control-learning describes the process through which memory guides how humans adjust their attentional priorities across various contexts, and it is typically assessed by analyzing how people learn to adjust their attentional priorities to different frequencies of hard and easy trials on tasks that challenge attention. Here, we manipulated how often particular images (item-specific) or blocks of trials (list-wide) were predictive of difficult trials and found that performance feedback selectively promoted the recruitment of anticipatory, proactive control, represented by the list-wide proportion congruent paradigm, and had little impact on the recruitment of in-the-moment, reactive control, represented by the item-specific proportion congruent paradigm. These results suggest that intrinsic motivation manipulations can enhance the recruitment of costly proactive control, which may prove especially useful for psychiatric disorders like schizophrenia in which proactive control is impaired.
Acknowledgments
This research was supported in part by NIMH R01 MH 087610. All authors report no conflicts of interest.
References
- Abrahamse E, Braem S, Notebaert W, & Verguts T (2016). Grounding cognitive control in associative learning. Psychological Bulletin, 142(7), 693. [DOI] [PubMed] [Google Scholar]
- Abrahamse EL, Duthoo W, Notebaert W, & Risko EF (2013). Attention modulation by proportion congruency: The asymmetrical list shifting effect. Journal of Experimental Psychology: Learning, Memory, and Cognition, 39(5), 1552–1562. 10.1037/a0032426 [DOI] [PubMed] [Google Scholar]
- Balleine BW, Delgado MR, & Hikosaka O (2007). The Role of the Dorsal Striatum in Reward and Decision-Making. Journal of Neuroscience, 27(31), 8161–8165. 10.1523/JNEUROSCI.1554-07.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barch DM, & Sheffield JM (2017). Cognitive Control in Schizophrenia: Psychological and Neural Mechanisms In Egner T (Ed.), The Wiley Handbook of Cognitive Control (pp. 556–580). Oxford: Wiley-Blackwell. [Google Scholar]
- Bejjani C, DePasque S, & Tricomi E (2019). Intelligence mindset shapes neural learning signals and memory. Biological Psychology, 146, 107715 10.1016/j.biopsycho.2019.06.003 [DOI] [PubMed] [Google Scholar]
- Bejjani C, & Egner T (2018). Spontaneous task structure formation results in a cost to incidental memory of task stimuli. Front Psychol. 2019 December 17;10:2833. doi: 10.3389/fpsyg.2019.02833. eCollection 2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bejjani C, Dolgin J, Zhang Z, Egner T. (2020) Disentangling the Roles of Cue Visibility and Knowledge in Adjusting Cognitive Control: A Preregistered Direct Replication of the Farooqui and Manly (2015) Study. Psychol Sci. 2020 March 30:956797620904045. doi: 10.1177/0956797620904045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blais C, & Bunge S (2009). Behavioral and Neural Evidence for Item-specific Performance Monitoring. Journal of Cognitive Neuroscience, 22(12), 2758–2767. 10.1162/jocn.2009.21365 [DOI] [PubMed] [Google Scholar]
- Blais C, Harris MB, Guerrero JV, & Bunge SA (2012). Rethinking the role of automaticity in cognitive control. The Quarterly Journal of Experimental Psychology, 65(2), 268–276. 10.1080/17470211003775234 [DOI] [PubMed] [Google Scholar]
- Botvinick M, & Braver T (2015). Motivation and Cognitive Control: From Behavior to Neural Mechanism. Annual Review of Psychology, 66(1), 83–113. 10.1146/annurev-psych-010814-015044 [DOI] [PubMed] [Google Scholar]
- Botvinick MM, Braver TS, Barch DM, Carter CS, & Cohen JD (2001). Conflict monitoring and cognitive control. Psychological Review, 108(3), 624–652. 10.1037/0033-295X.1083.624 [DOI] [PubMed] [Google Scholar]
- Braem S, Bugg JM, Schmidt JR, Crump MJC, Weissman DH, Notebaert W, & Egner T (2019). Measuring Adaptive Control in Conflict Tasks. Trends in Cognitive Sciences, 23(9), 769–783. 10.1016/j.tics.2019.07.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braver TS (2012). The variable nature of cognitive control: A dual-mechanisms framework. Trends in Cognitive Sciences, 16(2), 106–113. 10.1016/j.tics.2011.12.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braver TS, Krug MK, Chiew KS, Kool W, Westbrook JA, Clement NJ, … Carver CS. (2014). Mechanisms of motivation-cognition interaction: challenges and opportunities. Cognitive, Affective, & Behavioral Neuroscience, 14(2), 443–472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bugg JM (2014). Evidence for the sparing of reactive cognitive control with age. Psychology and Aging, 29(1), 115–127. 10.1037/a0035270 [DOI] [PubMed] [Google Scholar]
- Bugg JM (2017). Context, Conflict, and Control In Egner T (Ed.), The Wiley Handbook of Cognitive Control (pp. 79–96). Oxford: Wiley-Blackwell. [Google Scholar]
- Bugg JM, & Chanani S (2011). List-wide control is not entirely elusive: Evidence from picture-word Stroop. Psychonomic Bulletin & Review, 18(5), 930–936. 10.3758/s13423-011-0112-y [DOI] [PubMed] [Google Scholar]
- Bugg JM, & Crump MJ (2012). In support of a distinction between voluntary and stimulus-driven control: a review of the literature on proportion congruent effects. Frontiers in Psychology, 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bugg JM, & Dey A (2018). When stimulus-driven control settings compete: On the dominance of categories as cues for control. Journal of Experimental Psychology: Human Perception and Performance. 10.1037/xhp0000580 [DOI] [PubMed] [Google Scholar]
- Bugg JM, & Diede NT (2018). The effects of awareness and secondary task demands on Stroop performance in the pre-cued lists paradigm. Acta Psychologica, 189, 26–35. 10.1016/j.actpsy.2016.12.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bugg JM, Diede NT, Cohen-Shikora ER, & Selmeczy D (2015). Expectations and experience: Dissociable bases for cognitive control? Journal of Experimental Psychology: Learning, Memory, and Cognition, 41(5), 1349–1373. 10.1037/xlm0000106 [DOI] [PubMed] [Google Scholar]
- Bugg JM, Jacoby LL, & Chanani S (2011). Why it is too early to lose control in accounts of item-specific proportion congruency effects. Journal of Experimental Psychology: Human Perception and Performance, 37(3), 844. [DOI] [PubMed] [Google Scholar]
- Bundt C, Ruitenberg MFL, Abrahamse EL, & Notebaert W (2018). Early and late indications of item-specific control in a Stroop mouse tracking study. PLOS ONE, 13(5), e0197278 10.1371/journal.pone.0197278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chandler J, Mueller P, & Paolacci G (2014). Nonnaivete among Amazon Mechanical Turk workers: Consequences and solutions for behavioral researchers | SpringerLink. Behavioral Research Methods, 46(1), 112–130. [DOI] [PubMed] [Google Scholar]
- Chiew KS, & Braver TS (2011). Positive affect versus reward: emotional and motivational influences on cognitive control. Frontiers in Psychology, 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiew KS, & Braver TS (2013). Temporal dynamics of motivation-cognitive control interactions revealed by high-resolution pupillometry. Frontiers in Psychology, 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiew KS, & Braver TS (2016). Reward favours the prepared: incentive and task-informative cues interact to enhance attentional control. Journal of Experimental Psychology. Human Perception and Performance, 42(1), 52–66. 10.1037/xhp0000129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu Y-C, & Egner T (2019). Cortical and subcortical contributions to context-control learning. Neuroscience & Biobehavioral Reviews, 99, 33–41. 10.1016/j.neubiorev.2019.01.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu Y-C, Jiang J, & Egner T (2017). The Caudate Nucleus Mediates Learning of Stimulus-Control State Associations. The Journal of Neuroscience, 37(4), 1028–1038. 10.1523/JNEUROSCI.0778-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delgado MR (2007). Reward-Related Responses in the Human Striatum. Annals of the New York Academy of Sciences, 1104(1), 70–88. 10.1196/annals.1390.002 [DOI] [PubMed] [Google Scholar]
- Dreisbach G, & Fischer R (2012). Conflicts as aversive signals. Brain and Cognition, 78(2), 94–98. 10.1016/j.bandc.2011.12.003 [DOI] [PubMed] [Google Scholar]
- Egner T (2014). Creatures of habit (and control): a multi-level learning perspective on the modulation of congruency effects. Frontiers in Psychology, 5 10.3389/fpsyg.2014.01247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank MJ, Loughry B, & O’Reilly RC (2001). Interactions between frontal cortex and basal ganglia in working memory: A computational model. Cognitive, Affective, & Behavioral Neuroscience, 1(2), 137–160. 10.3758/CABN.1.2.137 [DOI] [PubMed] [Google Scholar]
- Fröber K, & Dreisbach G (2014). The differential influences of positive affect, random reward, and performance-contingent reward on cognitive control. Cognitive, Affective, & Behavioral Neuroscience, 14(2), 530–547. 10.3758/s13415-014-0259-x [DOI] [PubMed] [Google Scholar]
- Fröber K, & Dreisbach G (2016). How performance (non-)contingent reward modulates cognitive control. Acta Psychologica, 168, 65–77. 10.1016/j.actpsy.2016.04.008 [DOI] [PubMed] [Google Scholar]
- Grandjean J, D’Ostilio K, Fias W, Phillips C, Balteau E, Degueldre C, … Collette F.(2013). Exploration of the mechanisms underlying the ISPC effect: Evidence from behavioral and neuroimaging data. Neuropsychologia, 51(6), 1040–1049. 10.1016/j.neuropsychologia.2013.02.015 [DOI] [PubMed] [Google Scholar]
- Hommel B (2004). Event files: feature binding in and across perception and action. Trends in Cognitive Sciences, 8(11), 494–500. 10.1016/j.tics.2004.08.007 [DOI] [PubMed] [Google Scholar]
- Hutchison KA, Bugg JM, Lim YB, & Olsen MR (2016). Congruency precues moderate item-specific proportion congruency effects. Attention, Perception, & Psychophysics, 78(4), 1087–1103. 10.3758/s13414-016-1066-y [DOI] [PubMed] [Google Scholar]
- Jiang J, Beck J, Heller K, & Egner T (2015). An insula-frontostriatal network mediates flexible cognitive control by adaptively predicting changing control demands. Nature Communications, 6, 8165 10.1038/ncomms9165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang J, Brashier NM, & Egner T (2015). Memory Meets Control in Hippocampal and Striatal Binding of Stimuli, Responses, and Attentional Control States. Journal of Neuroscience, 35(44), 14885–14895. 10.1523/JNEUROSCI.2957-15.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang G, Wang L, & Zhou X (2017). Reward interacts with modality shift to reduce cross-modal conflict. Journal of Vision, 17(1), 19–19. 10.1167/17.1.19 [DOI] [PubMed] [Google Scholar]
- Kostandyan M, Bombeke K, Carsten T, Krebs RM, Notebaert W, & Boehler CN(2019). Differential effects of sustained and transient effort triggered by reward – A combined EEG and pupillometry study. Neuropsychologia, 123, 116–130. 10.1016/j.neuropsychologia.2018.04.032 [DOI] [PubMed] [Google Scholar]
- Krug MK, & Braver TS (2014). Motivation and Cognitive Control: Going Beyond Monetary Incentives In Bijleveld E & Aarts H (Eds.), The Psychological Science of Money (pp. 137–162). 10.1007/978-1-4939-0959-9_7 [DOI] [Google Scholar]
- Lempert KM, & Tricomi E (2015). The Value of Being Wrong: Intermittent Feedback Delivery Alters the Striatal Response to Negative Feedback. Journal of Cognitive Neuroscience, 28(2), 261–274. 10.1162/jocn_a_00892 [DOI] [PubMed] [Google Scholar]
- Logan GD, & Zbrodoff NJ (1979). When it helps to be misled: Facilitative effects of increasing the frequency of conflicting stimuli in a Stroop-like task. Memory & Cognition, 7(3), 166–174. 10.3758/BF03197535 [DOI] [Google Scholar]
- Logan GD, & Zbrodoff NJ (1998). Stroop-type interference: Congruity effects in color naming with typewritten responses. Journal of Experimental Psychology: Human Perception and Performance, 24(3), 978–992. 10.1037/0096-1523.24.3.978 [DOI] [Google Scholar]
- Muhle-Karbe PS, Jiang J, & Egner T (2017). Causal evidence for learning-dependent frontal-lobe contributions to cognitive control. Journal of Neuroscience, 1467–17. 10.1523/JNEUROSCI.1467-17.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qiao L, Xu L, Che X, Zhang L, Li Y, Xue G, … Chen A (2018). The Motivation-Based Promotion of Proactive Control: The Role of Salience Network. Frontiers in Human Neuroscience, 12 10.3389/fnhum.2018.00328 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruitenberg MFL, Braem S, Du Cheyne H, & Notebaert W (2019). Learning to be in control involves response-specific mechanisms. Attention, Perception, & Psychophysics. 10.3758/s13414-019-01753-0 [DOI] [PubMed] [Google Scholar]
- Schmidt JR, & Besner D (2008). The Stroop effect: Why proportion congruent has nothing to do with congruency and everything to do with contingency. Journal of Experimental Psychology: Learning, Memory, and Cognition, 34(3), 514–523. 10.1037/0278-7393.34.3.514 [DOI] [PubMed] [Google Scholar]
- Schönbrodt FD, & Perugini M (2013). At what sample size do correlations stabilize? Journal of Research in Personality, 47(5), 609–612. 10.1016/j.jrp.2013.05.009 [DOI] [Google Scholar]
- Schouppe N, Braem S, Houwer JD, Silvetti M, Verguts T, Ridderinkhof KR, & Notebaert W (2015). No pain, no gain: the affective valence of congruency conditions changes following a successful response. Cognitive, Affective, & Behavioral Neuroscience, 15(1), 251–261. 10.3758/s13415-014-0318-3 [DOI] [PubMed] [Google Scholar]
- Shenhav A, Botvinick MM, & Cohen JD (2013). The Expected Value of Control: An Integrative Theory of Anterior Cingulate Cortex Function. Neuron, 79(2), 217–240. 10.1016/j.neuron.2013.07.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soutschek A, Stelzel C, Paschke L, Walter H, & Schubert T (2015). Dissociable Effects of Motivation and Expectancy on Conflict Processing: An fMRI Study. Journal of Cognitive Neuroscience, 27(2), 409–423. 10.1162/jocn_a_00712 [DOI] [PubMed] [Google Scholar]
- Soutschek A, Strobach T, & Schubert T (2014). Motivational and cognitive determinants of control during conflict processing. Cognition and Emotion, 28(6), 1076–1089. 10.1080/02699931.2013.870134 [DOI] [PubMed] [Google Scholar]
- Stewart N, Chandler J, & Paolacci G (2017). Crowdsourcing Samples in Cognitive Science. Trends in Cognitive Sciences, 21(10), 736–748. 10.1016/j.tics.2017.06.007 [DOI] [PubMed] [Google Scholar]
- Stroop JR (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18(6), 643–662. 10.1037/h0054651 [DOI] [Google Scholar]
- Townsend JT, & Ashby FG (1983). Stochastic modeling of elementary psychological processes. Cambridge: Cambridge University. [Google Scholar]
- Tricomi E, Delgado MR, McCandliss BD, McClelland JL, & Fiez JA (2006).Performance Feedback Drives Caudate Activation in a Phonological Learning Task. Journal of Cognitive Neuroscience, 18(6), 1029–1043. 10.1162/jocn.2006.18.6.1029 [DOI] [PubMed] [Google Scholar]
- Tricomi E, & DePasque S (2016). The Role of Feedback in Learning and Motivation. In Advances in Motivation and Achievement: Vol. 19. Recent Developments in Neuroscience Research on Human Motivation (Vol. 19, pp. 175–202). 10.1108/S0749-742320160000019015 [DOI] [Google Scholar]
- Tricomi E, & Fiez JA (2008). Feedback signals in the caudate reflect goal achievement on a declarative memory task. NeuroImage, 41(3), 1154–1167. 10.1016/j.neuroimage.2008.02.066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yee DM, & Braver TS (2018). Interactions of motivation and cognitive control. Current Opinion in Behavioral Sciences, 19, 83–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.