

Abstract
Introduction:
Coordination of multiple degrees of freedom in the performance of dynamic and complex motor tasks presents a challenging neuromuscular control problem. Experiments have inferred that humans exhibit self-organized, preferred coordination patterns, which emerge due to actor and task constraints on performance. The purpose of this study was to determine if the set of effective coordination strategies that exist for a task centers on a small number of robust, invariant patterns of behavior.
Methods:
Kinetic movement patterns computed from a cohort of 780 primarily female adolescent athletes performing a drop vertical jump task (DVJ) were analyzed in order to discover distinct groups into which individuals could be classified based on the similarity of movement coordination solutions.
Results:
Clustering of reduced-dimension joint moment of force time series revealed three very distinct, precisely delineated movement profiles that persisted across trials, and which exhibited different functional performance outcomes, despite no other apparent group differences. The same analysis was also performed on a different task—a single-leg drop landing (SLD)—which also produced distinct movement profiles; however, the three DVJ profiles did not translate to this task as group assignment was inconsistent between these two tasks.
Conclusion:
The task demands of the DVJ and SLD—successful landing, reversal of downward momentum, and, in the case of the DVJ, vertical propulsion toward a maximally positioned target—constrain movement performance such that only a few successful outcomes emerge. Discovery of the observed strategies in the context of associated task constraints may help our understanding of how injury risk movement patterns emerge during specific tasks, as well as how the natural dynamics of the system may be exploited to improve these patterns.
Keywords: injury risk, biomechanics, dynamical systems, self-organization
Introduction
How the central nervous system (CNS) controls and organizes the vast number of musculoskeletal degrees of freedom (DOF) presents a difficult biological control problem (1). Typical motor tasks, including behaviors like reaching for or grasping an object, involve the control of many DOF. Accordingly, there has been a focus on how the CNS “selects” an effective movement solution from the available DOF that will satisfy task demands (2–7). A related issue resulting from the redundancy among human motor system DOF is that this provides an abundance of potentially viable coordination strategies (8) and thus, there are many different dynamical configurations of motor DOF that will suffice to achieve a specified movement goal; however, while the set of potentially viable strategies is large, the set of feasible strategies that may emerge is likely to be far lower, constrained by the functional limitations relating to the individual performing the task, the environmental context in which the task is performed, and the task itself (9). Ultimately, the set of effective coordination strategies that exist for a given movement task may center around a relatively small number of robust, invariant patterns of behavior (4, 5, 10, 11), and closer examination of these structured motor patterns with respect to individual and task contexts may allow for more pointed insight into the process of “selecting” an effective coordination strategy for a specified movement task.
A fundamental question arising from this selection process is whether a small number of prototypical strategies for performing a specific task are widely adopted across individuals, or if different individuals instead settle upon distinct strategies. Identification of patterns that are consistent within individuals across multiple performances of the same task but vary across individuals suggest that, among the redundant strategies available, individuals may arrive at different strategies as a consequence of their own physical (e.g., neural, biomechanical, physiological) constraints (9) or their unique movement histories. A related question is whether the same individual consistently uses the same strategy for a given task or adopts a fresh strategy on each trial, as well as whether movement solution strategies that are stable for an individual in one movement task or context are transferred to other movement tasks or contexts. However, issues relating to the classification of such strategies affect the assessment and interpretation of the underlying, structured motor pattern(s) that manifest. A primary concern is the notion that “general” strategies exist for a specific task, as many motor DOF configurations can suffice to achieve a given task. Individual strategies tend to differ from average group responses to task demands (2), likely due to inherent variability in the neuromotor mechanisms underlying the execution and control of movement (12). Nevertheless, various approaches have been applied to classify individual performer strategies into general schemes. For example, performer strategies have been identified relative to load accommodation during walking (5) and drop landings (4, 6, 7). Other approaches have identified the underlying coordinative structures (i.e., synergies (13)) that characterize movement (14, 15). Thus, elucidating the process of identifying general performer strategies for a given task and their selection process by individuals can improve our understanding of how humans organize and execute effective, coordinated movement. Enhanced understanding of relationship of general performer strategies and movement coordination can improve both injury preventative and rehabilitative neuroscience and medicine.
In the present study, we attempted to identify coordination strategies, using analytic methods designed to extract invariant patterns from large and otherwise highly variable data sets (16, 17), by examining lower extremity joint moment profiles exhibited during performance of various landing tasks (7, 15). From the joint mechanical responses that characterize these profiles, inferences about their underlying neuromuscular control mechanisms (and collectively, the emergent coordination strategies that dictate them) can be made and interpreted in the context of performance and/or injury risk (18–21). Determining whether athletes select from a few distinct, general strategies for a given task, however, requires a method for detecting such strategies from movement time series data obtained from the athletes performing the task. In the present study, we employed a method to classify movement time series data into groups representing general coordination strategies (17). In principle, classification of movement time series data can be done by treating the entire time series as feature vector(s) for comparison; however, this feature space is too high-dimensional relative to the number of data points and also likely to contain a lot of redundancy (16). Therefore, approaches that emphasize relevant, higher-order features (e.g., ground reaction force profiles in load accommodation strategy identification (4–7)) or the use of dimensionality reduction techniques such as principal component analysis (PCA) (16) have been employed for classification of movement.
PCA, a multivariate statistical procedure that removes redundant information from high-dimensional data sets, has been applied to studies of motor variability as a means to distill invariant features of observed movement patterns via focus on the most relevant kinematic and kinetic aspects of these data (14, 17, 22–25). In particular, PCA has been used to classify performer strategies in healthy (17) and pathologic (22, 25) gait, vertical jumping (24), and weight lifting (14). In addition, these analytical approaches have been applied previously in studies of landing activities (6, 15). Thus, PCA-based analyses are a useful option to identify coordination strategies underlying performance of variety of movement tasks by discerning among the high-dimensional, highly variable configuration space(s) in which movement data resides.
There were two objectives of the present study. The first was to determine whether individuals use prototypical movement strategies for a particular task (a drop-vertical jump, DVJ) by assessing whether their motor patterns can be grouped into distinct and consistent profiles (i.e., representing the underlying dynamical signatures of task performance) by the application of a PCA-based clustering technique (17) to experimental data obtained from a large sample of participants (N = 780) performing the DVJ. Specifically, this approach aims to identify the resultant lower-dimensional representations of the DVJ task by attempting to cluster the principal components from relevant musculoskeletal degrees of freedom (DOF) extracted via PCA. Provided that distinct coordination strategies emerge from this analysis, this approach also aims to determine whether the set of observed strategies persists across trials and whether they could be associated with a different task (a single-leg drop landing, SLD). It was hypothesized that clustering of these components would result in a distinct clustering solution, and as such, would exhibit at least two clearly delineated groups and resultant coordination strategies. The second objective was to evaluate the significance of the observed set of strategies by identifying and comparing the functional consequences, in terms of task performance (specified by vertical mechanical power production) and other potential confounders (e.g., anthropometrics, strength, etc.), of being classified in each group. Using mechanical power as a proxy for task performance, it was hypothesized that each strategy would result in different performance outcomes; specifically, strategies would result in stratified (i.e., higher or lower) power production during performance of the DVJ.
Methods
Participants
Data were collected from a total of 780 basketball, soccer, and volleyball adolescent athletes (M = 93; F = 687; mean ± SD (range) age = 14.0 ± 1.8 (8.2 – 18.8) years; height = 1.61 ± 8.8 (1.32 – 1.92) m; weight = 54.8 ± 12.3 (28.6 – 96.6) kg) who performed a series of biomechanical screening tasks implemented as part of two large prospective, longitudinal studies conducted over the course of four years (18, 20). The sample included many more women than men because those studies focused on anterior cruciate ligament injury risk, which is substantially greater for women than men (26). This cohort was especially suitable for the present study as they provided a rich array of movement patterns to analyze, owing to the diversity of anthropometric factors that generally underlie the maturational process in these individuals. The recruitment process, target population, and assessment protocols were identical for both studies. Study participants were recruited prior to their respective competitive sports seasons, and each athlete’s testing session represented their first visit to the research laboratory (i.e., they had not been tested previously). Prior to data collection in both studies, the study protocol was approved by the Cincinnati Children’s Hospital Medical Center Institutional Review Board, and informed, written consent was obtained from each subject and their parent(s) or legal guardian(s), if under 18 years of age.
Three-Dimensional Motion Analysis
Three-dimensional motion capture data were collected on each subject using a 10-camera, high-speed, passive optical motion capture system (Motion Analysis Corp., Santa Rosa, California, USA) sampled at 240 Hz. Ground reaction forces (GRF) were collected using two embedded force platforms (AMTI, Watertown, Massachusetts, USA) that sampled at 1200 Hz and were synchronized with the motion capture data. All participants (N = 780) performed two trials of the DVJ task. The cohort assessed as part of the randomized clinical trial (N = 352) also performed the SLD task in addition to the DVJ task. The DVJ and SLD tasks were chosen because of their moderate difficulty and relative but not complete similarity to one another (i.e., both constitute a reversal of downward momentum of the center of mass but are executed differently primarily in that SLD involves just one leg while DVJ is bilateral). The same member of the research staff gave instructions and demonstrated to each subject on how to execute proper performance of both tasks. While it is unknown if participants had previously been trained in either the DVJ or SLD task, given that these tasks are highly specific and, along with the corresponding instructions, were developed specifically for these studies, it is very unlikely that participants were trained in either task. Prior to data collection, participants were not given any habituation trials, and all trials were performed one after another with little rest in between (i.e., < 10 s).
Each subject was instrumented with 37 retroreflective markers with a minimum of three tracking markers per segment (Figure 1A). Markers were placed on the lower back between the S5 and T1 vertebrae, and bilaterally on the acromio-clavicular joint, lateral epicondyle of the elbow, mid-wrist, anterior superior iliac spine, greater trochanter, mid-thigh, medial and lateral femoral condyles, tibial tubercle, lateral and distal aspects of the shank, medial and lateral malleoli, the heel, the dorsal surface of the midfoot, the lateral foot (fifth metatarsal) and central forefoot (between the second and third metatarsals). Before the dynamic trials, a static trial was conducted with the subject in anatomical pose and with foot direction and placement standardized to the laboratory’s coordinate system to define each subject’s neutral kinematic posture (Figure 1A).
Fig. 1.
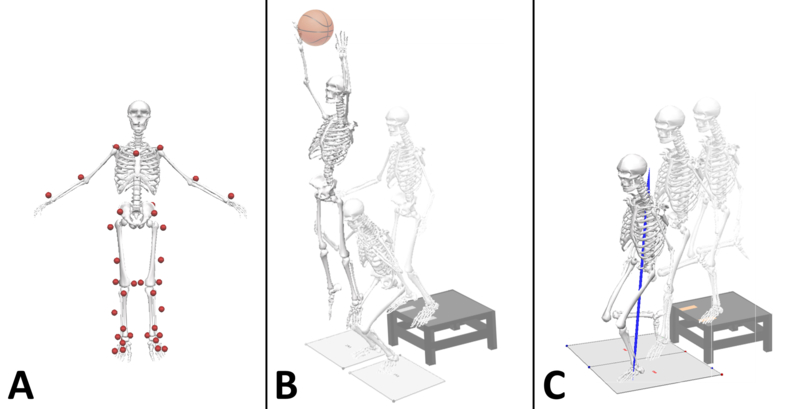
(A) Marker placement for each subject prior to data collection from the anterior view; (B) Illustration of a representative subject performing the drop vertical jump (DVJ) task; (C) Illustration of a representative subject performing the single-limb drop landing (SLD) task.
For the DVJ task, participants initially stood on top of a 31 cm tall box with their feet aligned with tape at the edge of the box (situated approximately shoulder-width apart). Participants were then instructed to drop off the box with both feet at the same time, land on the force platforms immediately in front of the box, and perform a maximum effort vertical leap to reach an overhead target (Figure 1B). The target height was set to their pre-recorded maximum vertical jump height. Trials were repeated if participants either led with one foot while dropping off the box, i.e., both feet did not leave the box at the same time, or did not immediately perform a maximum vertical leap upon landing, i.e., they paused upon landing before performing the jump. For the SLD task, participants initially aligned one foot with tape on the respective side on the edge of the 31 cm box and balanced on that foot. Participants were then instructed to drop off the box, land on the same leg, and hold the landing for a minimum of 2 s without shifting the body’s weight, hopping, or touching the ground with the opposite limb (Figure 1C). Participants performed one trial each on the left and right limbs separately in a randomized fashion. Trials were repeated if participants were unable to stabilize upon landing or if the opposite foot hit the box on the way down.
Marker trajectories and force data were match-filtered using a low-pass, fourth-order Butterworth filter with a cutoff frequency of 12 Hz. These data were match-filtered to reduce the potential for exceedingly large joint moments of force to be exhibited when marker and force data are filtered using unmatched frequencies. A 6-DOF skeletal model was applied to the marker trajectories to determine the position and orientation of each segment at each time sample, and the model was scaled to each subject’s height and weight. Cardan joint angles and joint moments of force were calculated during the stance phase—defined as the period of time from when participants made initial contact with the ground until toe-off occurred for the DVJ, and from when participants made initial contact with the ground until 500 ms after initial contact for the SLD (27)—via an inverse dynamics analysis using Visual3D software (C-Motion, Inc., Germantown, Maryland, USA). These data were then time-normalized to 101 data points (representing 0–100% of the participants’ stance phase).
Principal Component Analysis and Cluster Analysis
Clustering analysis was done separately for two trials of the DVJ task and one trial of the SLD task. For each task, the time-normalized joint moment of force data from the hip, knee, and ankle joints on both the left and right limbs were compiled to produce 18 time series in all (2 lower limbs × 3 joints × 3 axes of rotation). As the DVJ is a bilateral task, the left and right leg data were compiled from the same trial, whereas a single SLD trial was used for the SLD analysis. For the SLD analysis, the joint moment time series from the participants’ dominant limb was compiled. Each subject’s dominant limb was determined by asking the subject which limb they would use if they were to kick a ball as far as possible. Joint moment time series data were compiled for all participants, and PCA was performed separately for each DOF (18 in all for the DVJ; 9 for the SLD) to convert these time series to a feature vector of projection coefficients on the k leading principal components for that time series, with k < M (the number of data points in each time series). The resulting k-dimensional feature vectors were submitted to a hierarchical agglomerative clustering analysis.
For a given task, the time-normalized vectors (M = 101 data points) were compiled from each subject (N = 780) to produce a 780 × 101 matrix for each of 18 kinetic variables considered. Each matrix was normalized to have a zero mean and unit variance for each of the 101 time points and was then submitted to a separate PCA to produce a N × M set of projection coefficients (i.e., principal component “scores”) for the given variable. These scores represent how strongly the data from a given subject correspond with each of the principal axes of the dataset (i.e., where most of the data variance lies). PCA uses an eigenvector decomposition method on the covariance matrix of an input N × M data matrix to produce an M-dimensional eigenvector matrix that represents the principal axes. These eigenvectors—which are sorted by decreasing magnitude of the associated eigenvalues—indicate the variance of the data along each axis: the first eigenvector represents the direction of highest variance, the second represents the next highest, and so forth. This eigenvector matrix is used to produce the N × M set of scores for a given variable by projecting the original N × M matrix back onto each principal axis.
By concentrating the most variance from the joint moment of force time series data into the fewest possible dimensions, PCA allows kinetic similarities across participants to be detected in far fewer dimensions than that of the time series themselves (i.e., 101 data points), making it possible to perform meaningful clustering even with datasets of moderate size. Thus, instead of comparing 101 points across all participants, only the first few principal components (which contain most of the information from the original time series) can be compared. For example, if the scores for hip joint moment of force in the sagittal plane were very similar between two participants, it can be inferred that they generated comparable hip extensor patterns of recruitment, as the pattern of variation between these two participants’ time series would be similar.
To perform the dimensionality reduction, only those eigenvectors that individually captured ≥ 1% of the variance of the input data matrix were retained. Each PCA produced between 8–13 eigenvectors that captured ≥ 1% of the variance, producing a total of 186 scores across the 18 variables for each subject (182 scores were retained for the second DVJ trial, while 87 scores were retained for the SLD trial). Together, the retained principal components accounted for about 97% of the variance for each variable. The pairwise Euclidean distance in this 186-dimensional feature space was calculated between each distinct pair of participants, who were then paired into binary clusters using hierarchical agglomerative clustering based on Ward’s linkage method (17, 28). Agglomerative clustering progressively groups the most similar participants together until a single group of all participants is formed, from which a clustering solution (i.e., determination of the most distinct groups) is determined quantitatively through the application of a stopping rule based on a plateauing of the agglomeration coefficient (17, 29); specifically, this coefficient—which indicates the overall dissimilarity of the clusters—begins to plateau when the groups exhibit maximal between-group dissimilarity. The original (full) time-series for all participants in a cluster were then grouped to visualize their similarity. This solution was qualitatively confirmed by visually inspecting the dendrogram and the average waveforms generated from each cluster. Each time-series cluster determined from this analysis represents a distinct mechanical response for that DOF in the specified task (DVJ or SLD, respectively), and from these responses, appropriate inferences about the varying neuromuscular control responses can be made. Therefore, identification of the corresponding clusters across all DOF captures the distinct neuromuscular coordination strategies being used by the participants.
Isokinetic Strength and Functional Performance Analysis
An important issue to consider is whether distinct strategies identified through the PCA correspond to distinct functional outcomes. This was done in the present study by computing mechanical power from the vertical ground reaction force (GRF) generated by participants in each trial and task to represent functional performance. This measure was used since the DVJ task relies on rapid production of vertical GRF via the hip, knee, and ankle joints (eccentric extension to effectively land and concentric extension to propel oneself upward (30)). Isokinetic knee flexor and extensor strength was also examined for its potential to cause a spurious association between group assignment and functional performance.
Mechanical power was determined for all of the participants in each group during landing and was calculated as
| [1] |
where was the average net vertical GRF produced by each subject, normalized to each subject’s weight and was the change in the vertical position of the body’s center of mass over the given sub-phase (i.e., “landing” or “propulsive”). The “landing” and “propulsive” sub-phases of the stance phase were differentiated by the point at which the subject’s center of mass was at a minimum. Knee flexor and extensor strength values were measured using an isokinetic dynamometer (Biodex Medical Systems Inc., Shirley, NY, USA). The collection procedures for these measures have been reported elsewhere (31). Peak knee flexor and extensor strength were normalized to each subject’s body weight.
Statistical Analysis of Performance for Clusters
To address the question of whether the clusters found correspond to different functional outcomes, pairwise two-sample Kolgomorov-Smirnov (K-S) tests were conducted to assess whether the mechanical power distributions between the detected groups were distinct. One-way analyses of variance (ANOVA) were used to assess cluster differences in anthropometry (i.e., height and weight), strength, and power during both the landing and propulsion sub-phases of the stance phase, and eta squared (η2) was computed to estimate effect sizes for each comparison. The thresholds for interpreting effect size as either small, medium, or large were indicated as 0.01, 0.06, and 0.14, respectively (32). Tukey’s Honestly Significant Difference criterion was used for post hoc comparisons between groups. The alpha level indicating significant differences between groups was selected a priori to be 0.05. Before conducting statistical analyses on strength and power measures, outliers were removed based on whether computed values fell outside of the range ± 2.5 SD from the median. Outliers were removed in order to limit bias by controlling for potentially unaccounted errors in the mechanical power values; for each measure, between 12–21 participants’ data were indicated as outliers and removed prior to analysis.
Results
Clustering Analysis and Movement Profiles
Clustering of kinetic data from the first DVJ trial revealed three groups with characteristically distinct patterns of movement coordination (Figure 2). No differences existed between the clusters in either subject height (F(2, 777) = 0.97, p = .381, η2 = .003) or weight (F(2, 777) = 0.20, p = .820, η2 = .001). All clusters also divided male and female participants in roughly equal proportions.
Fig. 2.
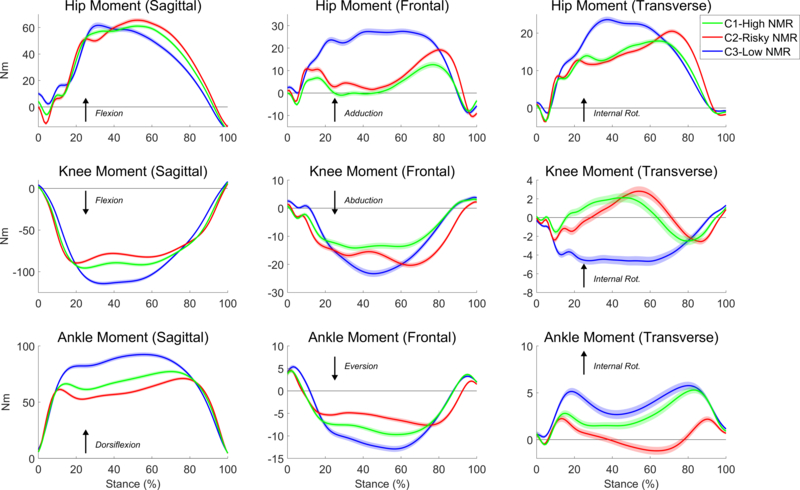
Mean hip, knee, and ankle joint moment of force time series across all participants in each of the three clusters detected from the clustering analysis of the first DVJ trial. Each waveform represents the mean external joint moment of force for the specified DOF, and the arrows indicate the interpretation convention for each moment. For DOF with an active joint torque mechanism (e.g., the hip in all three planes, the knee in the sagittal plane, and ankle in the sagittal plane), they are interpreted as internal moments of force, which correspond with the absolute magnitude of the moment. For example, external knee flexor moment is negative, so a higher internal knee extensor moment is interpreted as a more negative moment. Shaded regions indicate standard error regions between the groups.
On average, participants from the first cluster (blue waveforms in Figure 2; C1-High Neuromuscular Recruitment (NMR) cluster) exhibited increased and more sustained NMR of the lower extremities, as inferred by larger hip, knee, and ankle joint moments of force in the sagittal plane and larger hip moments in the frontal and transverse planes. In general, this profile is indicative of a more optimal coordination strategy, as these joint moments are critical to effective jumping performance. Hip, knee, and ankle joint extension are necessary to reverse the momentum of the center of mass, and hip abduction and external rotation maintain neutral proximal alignment of the lower limbs during landing, which result in improved control of the knee joint (18, 19, 30, 33). For these reasons, we adopted the label C1-High NMR for this group. Individuals from the second cluster (red waveforms in Figure 2; C2-Risky NMR) and third cluster (green waveforms in Figure 2; C3-Low NMR) exhibited decreased NMR compared to C1-High NMR, inferred by smaller hip joint moments in all three planes and the knee and ankle in the sagittal plane, which is associated with lower jumping performance. In addition to exhibiting the lowest NMR, the C2-Risky NMR group exhibited higher knee moments in the frontal and transverse planes. These biomechanical deficits are manifestations of poor knee joint control and have been associated with increased injury risk (18)—hence the use of the label “Risky” to describe this group. Qualitatively, C2-Risky NMR and C3-Low NMR were quite close, but as standard error-shaded regions around the curves in Figure 2 show, the differences between them are quantitatively significant.
Clustering of kinetic data from the second DVJ trial revealed clusters with movement profiles almost identical to those found in trial 1 (Figure 3). Notably, the resultant movement patterns exhibited by the groups, especially C1-High NMR and C2-Risky NMR, illustrated the same invariant features as revealed in the first DVJ trial. Thus, the three strategies represent a remarkable degree of stability across trials. Due to this, it was possible to analyze whether participants stayed with the same strategy in the two trials or utilized different strategies across the two DVJ trials, which were performed back-to-back in the same testing session. Table 1 shows some persistent associations among clusters. The majority of participants from C1-High NMR (55%) and C2-Risky NMR (87%) remained in their respective groups in the second DVJ trial. However, many participants from C3-Low NMR (48%) and many from C1-High NMR (34%) moved into C2-Risky NMR in the second trial, which contained an overwhelming majority of the participants as compared to the first DVJ trial.
Fig. 3.
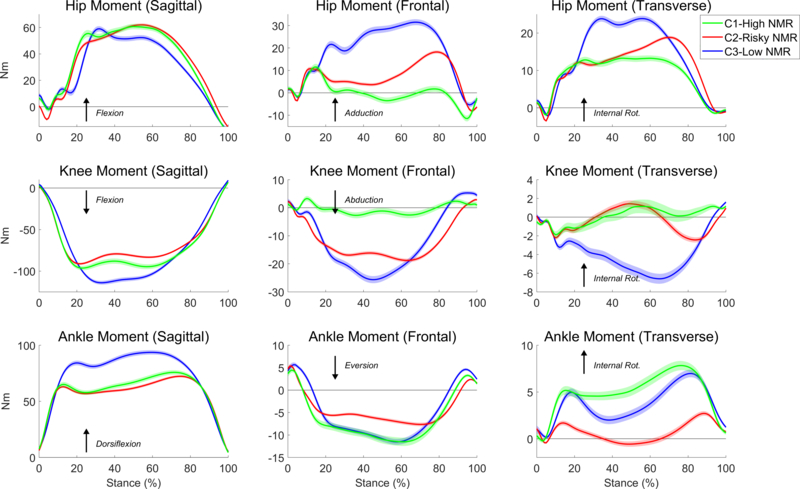
Average hip, knee, and ankle joint moment of force time series across all participants in each of the three clusters detected from the clustering analysis of the second DVJ trial. Each waveform represents the mean external joint moment of force for the specified DOF, and the arrows indicate the interpretation convention for each moment. Shaded regions indicate standard error regions between the groups.
Table 1.
Cluster association for each group detected in the clustering analyses of both DVJ trials and the SLD trial. Each column represents the number of subjects in the specified group from the first trial (left) that were associated with the specified group in the second trial (top) and their percentage of the total number of subjects. Majority associations are in bold. For example, 86.5% of subjects grouped in C2-Risky NMR on the first DVJ trial were grouped into C2-Risky NMR on the second DVJ trial (218/252).
| DVJ Trial 2 |
||||||
|---|---|---|---|---|---|---|
| N | C1-High NMR | C2-Risky NMR | C3-Low NMR | |||
| DVJ Trial 1 | C1-High NMR | 244 | 136 (55.7%) | 82 (33.6%) | 26 (10.7%) | |
| C2-Risky NMR | 252 | 19 ( 7.5%) | 218 (86.5%) | 15 ( 6.0%) | ||
| C3-Low NMR | 284 | 53 (18.7%) | 137 (48.2%) | 94 (36.4%) | ||
| Total | 780 | 208 (26.7%) | 437 (56.0%) | 135 (17.3%) | ||
| SLD Trial 1 |
||||||
| N | C1 | C2 | C3 | C4 | ||
| DVJ Trial 1 | C1-High NMR | 92 | 22 (23.9%) | 14 (15.2%) | 21 (22.8%) | 35 (38.0%) |
| C2-Risky NMR | 131 | 22 (16.8%) | 32 (24.4%) | 21 (16.0%) | 56 (42.7%) | |
| C3-Low NMR | 129 | 26 (20.2%) | 35 (27.1%) | 35 (27.1%) | 33 (25.6%) | |
| Total | 352 | 70 (19.9%) | 81 (23.0%) | 77 (21.9%) | 124 (35.2%) | |
| SLD Trial 1 |
||||||
| N | C1 | C2 | C3 | C4 | ||
| DVJ Trial 2 | C1-High NMR | 82 | 11 (13.4%) | 18 (22.0%) | 15 (18.3%) | 38 (46.3%) |
| C2-Risky NMR | 214 | 42 (19.6%) | 54 (25.2%) | 43 (20.1%) | 75 (35.0%) | |
| C3-Low NMR | 56 | 17 (30.4%) | 9 (16.1%) | 19 (33.9%) | 11 (19.6%) | |
| Total | 352 | 70 (19.9%) | 81 (23.0%) | 77 (21.9%) | 124 (35.2%) | |
Clustering of kinetic data from the SLD trial showed that four clusters of roughly equal size emerged from the data (Figure 4), and a post hoc comparison of the participant association in each of the detected groups from both tasks was made. However, in comparing the cluster associations between the DVJ and SLD tasks, no clear transfer was exhibited; specifically, no one group contained a majority of the participants as grouped from the results of either the first or second DVJ trial (Table 1). Another important observation from the SLD clusters is that the force patterns in all three cases are qualitatively quite similar. Thus, it may be appropriate to see the clusters as representing four subtle variants of the same overall strategy.
Fig. 4.
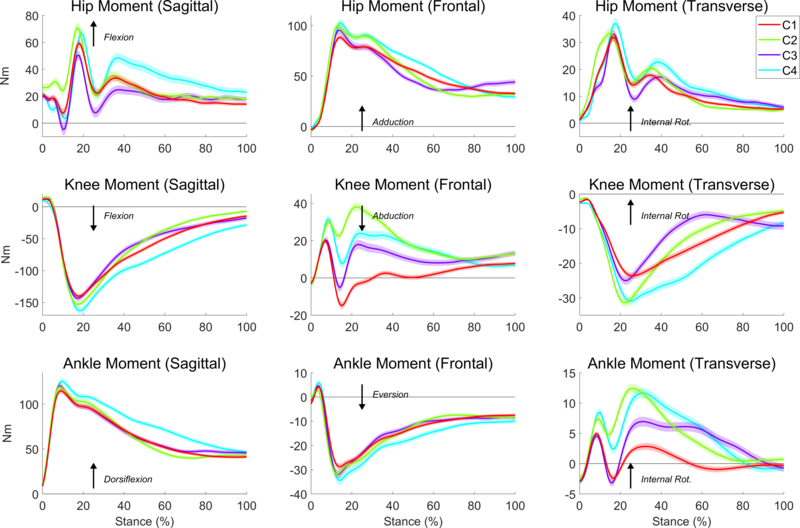
Average hip, knee, and ankle joint moment of force time series across all participants in each of the four clusters detected from the clustering analysis of the SLD trial. Each waveform represents the mean external joint moment of force for the specified DOF, and the arrows indicate the interpretation convention for each moment. Shaded regions indicate standard error regions between the groups.
Cluster Assignment, Isokinetic Strength, and Mechanical Power
During the landing phase for both DVJ trials, mechanical power measures from the participants in each cluster were determined to not have come from the same continuous distribution (pairwise K-S, all p < .05). However, during the propulsive phase of the second DVJ trial, mechanical power from clusters C2-Risky NMR and C3-Low NMR was found to have come from the same distribution (p = .520), but mechanical power from C1-High NMR did not come from the same distribution as C2-Risky NMR and C3-Low NMR (both p < .001).
Individuals among the three clusters exhibited differences in power during both of the landing and propulsion sub-phases during both DVJ trials (Table 2). Specifically, for the first DVJ trial, there was a significant main effect, with medium-to-large effect sizes, among the three groups during the landing (F(2, 756) = 78.56, p < .001, η2 = .172) and propulsion (F(2, 755) = 32.40, p < .001, η2 = .079) sub-phases. Post-hoc analyses revealed that C1-High NMR produced significantly higher power during landing than both C2-Risky NMR and C3-Low NMR (both p < .001); moreover, C2-Risky NMR produced significantly lower power than C3-Low NMR (p = .003). The same trends persisted during propulsion, with C1-High NMR producing higher power than the other two groups (both p < .001) and C2-Risky NMR producing lower power than C3-Low NMR (p < .001).
Table 2.
Mechanical power (Watts; mean ± SD) generated during the landing and propulsive phases of stance for both DVJ trials and isokinetic knee flexor and extensor strength (N × kg−1, mean ± SD) recorded for each of the three clusters detected in the clustering analysis.
| Task | Phase | C1-High NMR | C2-Risky NMR | C3-Low NMR |
|---|---|---|---|---|
| DVJ Trial 1 | Landing | −621.8 ± 227.0 | −378.1 ± 99.1 | −474.1 ± 118.4 |
| Propulsion | 482.9 ± 208.0 | 335.9 ± 176.4 | 402.8 ± 213.9 | |
| DVJ Trial 2 | Landing | −639.1 ± 223.5 | −405.1 ± 206.8 | 360.0 ± 187.0 |
| Propulsion | 497.2 ± 204.9 | −455.4 ± 234.4 | 390.0 ± 213.6 | |
| Left Knee Flexion | - | 0.62 ± 0.15 | 0.61 ± 0.15 | 0.59 ± 0.15 |
| Left Knee Extension | - | 0.91 ± 0.14 | 0.93 ± 0.14 | 0.89 ± 0.15 |
| Right Knee Flexion | - | 0.61 ± 0.15 | 0.59 ± 0.15 | 0.59 ± 0.15 |
| Right Knee Extension | - | 0.94 ± 0.14 | 0.95 ± 0.14 | 0.91 ± 0.16 |
Similarly, for the second DVJ trial, there were significant main effects, also with medium-to-large effect sizes, among the three groups during landing (F(2, 762) = 79.55, p < .001, η2 = .173) and propulsion (F(2, 756) = 33.20, p < .001, η2 = .081), with post-hoc analyses also revealing that C1-High NMR produced significantly higher power during both landing and propulsion than either C2-Risky NMR or C3-Low NMR (all p < .001). C2-Risky NMR and C3-Low NMR were not significantly different for either sub-phase (both p > .05).
There were no significant effects among the three groups in knee flexor strength for either the left (F(2, 763) = 1.71, p = .181, η2 = .004) or right (F(2, 768) = 1.56, p = .212, η2 = .004) limbs, respectively, as well as knee extensor strength for either the left limb (F(2, 757) = 0.83, p = .437, η2 = .002) or right limb (F(2, 761) = 0.93, p = .360, η2 = .002).
Discussion
The DVJ is one of the most commonly performed tasks in biomechanical assessments, having been used to identify deficits associated with acute, chronic, and secondary lower extremity injury (18, 19, 33). The most remarkable finding of the present study is that participants in the present cohort (i.e., primarily adolescent females and some adolescent male athletes) tended to select from one of only three very distinct and very precisely delineated general strategies for performing this task, rather than employing a large number of different strategies. Based on the large sample studied here, it appears that these three strategies represent the most common motor solutions that participants in this group used to perform this task, possibly reflecting strong influence of actor constraints on coordination and movement control (9). Analysis also showed that the strategies result in different task performance despite no other apparent anthropometric or strength differences among participants in the clusters.
While persistent strategies emerged in the DVJ trial, the general strategies underlying each group did not appear to cross over to a different task (i.e., the SLD) and even showed some variation over participants in the same task (DVJ Trials 1 and 2); however, a lack of data from multiple trials from each task limits this finding. Another key result was that, although the movement patterns of each of the three clusters for DVJ were relatively stable between the first and second DVJ trials, participants did not necessarily remain in their respective clusters between trials. Only participants from C2-Risky NMR consistently remained in this group from trial 1 to 2 (87%; Table 1). Moreover, in the first DVJ trial, the participants were split into each of the three clusters relatively evenly, whereas in the second DVJ trial, C2-Risky NMR contained a majority of the participants (56%; Table 1). These two observations may support the notion of the coordination strategy exhibited by C2-Risky NMR as the “default” strategy of the three observed strategies for effective DVJ performance. This implies that C2-Risky NMR is the strategy that all individuals who perform the DVJ may fall into at some point. This strategy could indicate some minimum in terms of neuromuscular performance or effort, or even more generally, it is the most likely of the three to occur given the constraints of the task. Because the trials were performed one after the other, it may be that some participants chose not to perform a maximal (to the best of their ability) vertical leap. Analyzing more than two trials of this task or computing jump height may have added support to one (or some combination) of these factors as contributing to the increased likelihood of this default strategy being exhibited; however, the nature of this particular dataset did not allow for the capture of peak jump height, as data collection was often stopped immediately following the end of the stance phase. Thus, these reasons remain merely speculative.
Interestingly, C3-Low NMR appears to blend elements of both C1-High NMR and C3-Risky NMR (Figures 2, 3). From this, it would appear that the clusters correspond simply to two “base” strategies (C1-High NMR and C2-Risky NMR) and a “mixed” strategy (C3-Low NMR). What is remarkable, however, is that the moment patterns of C3-Low NMR are extremely tightly clustered, suggesting that only a specific mixture of the two pure strategies is viable, and that combining the two in other proportions does not occur (or at least was not seen in this very large sample). This reinforces the notion that complex motor tasks are performed by selectively triggering one of a small set of discrete coordination modes rather than by a more diffuse repertoire of mixed strategies. Of course, the experiment tested this only in the context of a stereotypical task, and it is possible that a larger, more diffuse set of coordination modes would appear in more variable natural situations or emerge after several iterations of the task.
Clustering Assignment and Functional Performance
It is not unexpected that movement patterns employed during the DVJ task that exhibit higher joint moments of force are associated with objectively higher levels of performance (30), as evident in C1-High NMR who produced significantly higher power than those in C2-Risky NMR and C3-Low NMR. However, it is interesting that these patterns emerged between groups of participants for whom no apparent differences existed in anthropometric or muscular strength characteristics, where one might expect differences that could affect the internal joint moment generation, joint angular ranges of motion, and ultimately eccentric and concentric joint work that dictate landing and jumping success (30, 34). While there are other, more traditional biomarkers that may adequately explain the increased performance of this task, including training status (35), lower extremity joint coordination integrity (36), proprioceptive ability (37), and other physiological mechanisms like inhibition of the Golgi tendon reflex in agonist muscles, it may also be that improved performance was simply the result of more optimal, emergent coordination among the neuromuscular components responsible for executing the DVJ. While speculative, participants who land with a more neutral lower extremity alignment might facilitate vertical transfer of force and exhibit increased joint moments as a result. Regardless, it is clear that certain coordination strategies allow for increased performance over others.
Clustering Assignment and Injury Risk Assessment
High knee abduction moments in the frontal and transverse planes have previously been linked to increased dynamic knee valgus positioning and injury risk (18). Dynamic knee valgus, which is characterized by medial displacement and internal rotation of the knee (i.e., a knock-kneed, twisting position), is precipitated by the presence of knee frontal and transverse plane moments that drive the knee inward. Improved proximal control, particularly at the trunk and hip, have been shown to modulate this motor pattern by increasing lower extremity stiffness (38) and reducing the effect of the ground reaction forces that facilitate high knee moments. In other words, increased recruitment of the hip can lead to better absorption of these forces by the lower extremities which can negate the effects of landing that tend to push the knees inward (21). This appears to have been the case with the C1-High NMR group, who likely exhibited increased neuromuscular recruitment as suggested by higher lower extremity joint loadings in the sagittal plane.
Individuals in the C2-Risky NMR and C3-Low NMR groups, with their decreased sagittal plane joint moments and increased moments in the frontal and transverse planes, did not exhibit as much sagittal plane recruitment and, as a result, may have tended to passively rely on the elastic properties of the musculoskeletal system to absorb and return some of the mechanical energy upon landing (18). This is evident by the late recruitment of the hip musculature during the propulsive phase, as indicated by the upward trajectories of the frontal and transverse plane hip moments in the latter half of the stance phase (Figure 2, Figure 3). This may indicate that the lower extremities are compensating for the lack of a sufficient stretch-shortening mechanism during landing, which improves lower extremity stiffness (39, 40). Importantly, C2-Risky NMR appears to exhibit much higher frontal and transverse plane knee joint moments than C3-Low NMR, which is consistent with their characterization as having “Risky” NMR (18, 19, 21). While the C1-High NMR also has high knee moment in the frontal plane, this group’s transverse plane knee moment is strongly decreased relative to both C2-Risky NMR and C3-Low NMR, which is likely a manifestation of the protective mechanism of increased proximal control (21). It is important to note that the use of “Risky” to describe the second cluster’s NMR implies increased injury risk by those who exhibit this particular strategy. However, the present study did not prospectively examine the participants from any cluster and their risk for sustaining an injury and, thus, this interpretation is somewhat speculative. Future work should examine the association between the coordination strategies that emerge from these types of analyses and their propensity to increase or decrease injury risk.
Accordingly, the finding that different coordination strategies emerged during the DVJ despite no anthropometric differences implies that at least some non-modifiable factors, like height, weight, etc., may not preclude individuals from adopting coordination strategies that lead to more optimal functional outcomes, like higher power production or improved joint neuromuscular control. From the perspective of a practitioner, understanding body technical performance and coordination during task execution and the coordinative structures that govern them is especially salient given that certain tests (e.g., isokinetic lower extremity strength assessment, as observed in the present study) may not be effective to elicit differences relating to whole-body movement dynamics. Thus, it is essential to determine how and to what degree contextual constraints lead to the emergence of certain coordination strategies over others.
Crossover between the DVJ and SLD
It is not immediately clear why only three distinct coordination strategies reliably emerged for successful landing and jumping to reach an overhead target. One might assume that, given the morphological consistency of the human body across individuals, any strategies that emerge will be variations on a broad “base” strategy. This appears to be the case for the SLD task, where the four clusters produced from the analysis appear to exhibit temporally similar movement patterns (i.e., the timing and directionality of the joint loadings of each SLD strategy appear to be more similar than those of the DVJ) differentiated only by a weighted modulation of amplitude over time (Figure 4), although this is speculative. However, it is still surprising that four distinct variants should emerge rather than a diffuse set of variations. Also, this was not the case for the DVJ, where the strategies appear to be qualitatively different and each was very well-defined with extremely low intra-cluster variation. The demands of the DVJ—that participants were required to successfully land, reverse the momentum of the center of mass, and then propel themselves upward with enough power to reach a maximally positioned overhead target—may constrain motor solutions in such a manner that only a few possible outcomes would allow the individual to be successful (4, 7). Functionally speaking, successful DVJ strategies are ones that are ultimately able to produce a high vertical impulse that reverses the center of mass and allows the individual to leave the ground—a requirement not shared by the SLD. Specifically, higher NMR was likely achieved in the performance of the DVJ through the use of the stretch-shortening cycle to optimally restore elastic energy during landing and contribute to a higher vertical jump (36), whereas during performance of the SLD, the primary task constraint was not to jump, but to simply stabilize upon landing.
Moreover, it is possible that the limited number of trials analyzed in the present study (i.e., 2 for the DVJ, 1 for the SLD) did not allow for a comprehensive assessment of all the strategies that may be employed for either task. Consequently, this analysis would likely have benefitted from having analyzed several trials to discover consistent strategies, and thus, the insight into between-trial variability is limited. Another possibility is that additional coordination strategies were not exhibited because the present study examined only successful trials; examination of unsuccessful trials may provide critically valuable insight into why relatively few strategies were observed by highlighting other motor strategies that may be employed by individuals performing the task but lead to failed action. Still another possibility is the fact that the cohort in the present study was limited in both age and sex (i.e., the group was primarily adolescent female athletes), and thus other strategies might exist for primarily-male cohorts and/or adult individuals. Thus, future work should examine strategies underlying both successful and unsuccessful task performance, as well as expand these analyses to include more males or older individuals, to increase the generalizability of these results.
That virtually no crossover existed between groupings of the DVJ and SLD tasks indicated that task constraints are likely highly influential on each task’s performance (6, 9) and appear to limit individuals to produce a single, base coordination strategy with variation, as opposed to the three distinct strategies seen in the DVJ. It is possible, given that successful unilateral landing effectively doubles the influence of body weight as a constraint on successful task performance (as opposed to a bilateral task such as the DVJ), that the space of coordination strategies which are ultimately successful in this task is more narrowly defined, which is consistent with the findings of Nordin et al. (6, 7) and James et al. (4) in their examination of the Load Accommodation Strategies Model (3, 5). Thus, the postulate that only one base strategy with variation exists for the SLD aligns with our observation that coordination in the DVJ is not strongly predictive of coordination in the SLD. Another potential explanation is that this is not a maximal effort task, but a relatively passive one. It is worth noting that the comparison between a bilateral and unilateral movement itself may contribute to this discrepancy, and as such, constitutes a limitation to the present study. A different but related bilateral task (e.g., a standing vertical jump) or a variation of the same task (e.g., performing the DVJ from a higher box) may have made for a more meaningful comparison, and future work should explore this. Nevertheless, it is also worth noting that, of all possible variations of the general SLD strategy, only four clearly defined variants were actually observed.
Conclusions and Future Work
Modern, large-scale data mining and analysis techniques may prove to be crucial to the discovery of how human movement is organized and coordinated, with implications for understanding human motor control and informing research efforts within rehabilitation and neuroscience. Large samples of participants can provide a rich data set of potentially variable movement patterns, and techniques such as PCA (16) may be able to overcome the inherent variation in this data and uncover the invariant features of various tasks (14, 15, 17, 22–25), which can allow researchers and practitioners to build robust profiles of injury risk and performance of individuals who perform those tasks (4–7, 17, 25). In the present study, a small set of robust coordination strategies was detected in a large cohort of primarily female adolescent athletes during execution of a dynamic landing and jumping task and those strategies were associated with different functional outcomes, despite no apparent anthropometric or other functional differences among the individuals exhibiting them.
Future work should examine other athletic tasks, such as hopping or cutting, and perhaps extend the observations from the present study to uncover exactly how and under what circumstances certain coordination patterns emerge and evolve. This research strategy could verify whether distilled patterns of movement, such as those observed in the present study, can emerge as a function of various actor, task, and environmental constraints. In addition, examining the observed strategies in the context of associated control variables (i.e., motor primitives) may help to uncover the processes by which the CNS may exploit the natural dynamics of the system that leads to the emergence of robust movement patterns during this task.
Acknowledgments
The authors would like to acknowledge funding support from the National Institutes of Health/NIAMS Grants R21AR065068-01A1, U01AR067997, R01-AR056259, R01-AR049735, R01-AR055563 and R03AR057551.
The authors would like to acknowledge funding support from the National Institutes of Health/NIAMS Grants R21AR065068-01A1, U01AR067997, R01-AR056259, R01-AR049735, R01-AR055563 and R03AR057551. The study sponsors held no role in the design, data collection, analysis or interpretation in the present study. The authors have no conflicts of interest to report. The data presented in this manuscript have not been falsified or manipulated in any way, and the authors have agreed on its presentation. The results of the presented study do not constitute endorsement by the American College of Sports Medicine.
Footnotes
Conflict of Interest Statement
The study sponsors held no role in the design, data collection, analysis or interpretation in the present study. The authors have no conflicts of interest to report. The data presented in this manuscript have not been falsified or manipulated in any way, and the authors have agreed on its presentation. The results of the presented study do not constitute endorsement by the American College of Sports Medicine.
References
- 1.Bernstein NA. The Co-ordination and Regulation of Movements. Oxford, United Kingdom: Pergamon Press; 1967. [Google Scholar]
- 2.Dufek JS, Bates BT, Stergiou N, James CR. Interactive effects between group and single-subject response patterns. Hum Movement Sci. 1995;14(3):301–23. [Google Scholar]
- 3.Bates BT. Single-subject methodology: An alternative approach. Medicine & Science in Sports & Exercise. 1996;28(5):631–8. [DOI] [PubMed] [Google Scholar]
- 4.James CR, Bates BT, Dufek JS. Classification and comparison of biomechanical response strategies for accommodating landing impact. Journal of Applied Biomechanics. 2003;19(2):106–18. [DOI] [PubMed] [Google Scholar]
- 5.James CR, Atkins LT, Dufek JS, Bates BT. An exploration of load accommodation strategies during walking with extremity-carried weights. Hum Movement Sci. 2014;35:17–29. [DOI] [PubMed] [Google Scholar]
- 6.Nordin AD, Dufek JS. Load accommodation strategies and movement variability in single-leg landing. J Appl Biomech. 2017;33(4):241–7. [DOI] [PubMed] [Google Scholar]
- 7.Nordin AD, Dufek JS, James CR, Bates BT. Classifying performer strategies in drop landing activities. J Sports Sci. 2017;35(18):1–6. [DOI] [PubMed] [Google Scholar]
- 8.Latash ML. The bliss (not the problem) of motor abundance (not redundancy). Experimental Brain Research. 2012;217(1):1–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Newell KM. Constraints on the development of coordination In: Wade MG, Whiting HTA, editors. Motor Development in Children: Aspects of Coordination and Control. Dordrecht, Netherlands: Martinus Nijhoff Publishers; 1986. p. 341–60. [Google Scholar]
- 10.Dingwell JB, Cusumano JP. Nonlinear time series analysis of normal and pathological human walking. Chaos. 2000;10(4):848–63. [DOI] [PubMed] [Google Scholar]
- 11.Hollman JH, Watkins MK, Imhoff AC, Braun CE, Akervik KA, Ness DK. Complexity, fractal dynamics and determinism in treadmill ambulation: Implications for clinical biomechanists. Clinical biomechanics. 2016;37:91–7. [DOI] [PubMed] [Google Scholar]
- 12.Newell KP, Slifkin AB. The nature of movement variability In: Piek JP, editor. Motor Behavior and Human Skill: A Multidisciplinary Approach. Champaign, IL: Human Kinetics Publishers, Inc.; 1998. p. 143–60. [Google Scholar]
- 13.Kelso JAS. Synergies: Atoms of brain and behavior In: Sternad D, editor. Progress in Motor Control. 629 New York, NY: Springer Science+Business Media; 2009. p. 83–91. [DOI] [PubMed] [Google Scholar]
- 14.Kipp K, Redden J, Sabick M, Harris C. Kinematic and kinetic synergies of the lower extremities during the pull in Olympic weightlifting. Journal of Applied Biomechanics. 2012;28(3):271–8. [DOI] [PubMed] [Google Scholar]
- 15.Nordin AD, Dufek JS. Neuromechanical synergies in single-leg landing reveal changes in movement control. Hum Mov Sci. 2016;49:66–78. [DOI] [PubMed] [Google Scholar]
- 16.Daffertshofer A, Lamoth CJ, Meijer OG, Beek PJ. PCA in studying coordination and variability: A tutorial. Clinical biomechanics. 2004;19(4):415–28. [DOI] [PubMed] [Google Scholar]
- 17.Phinyomark A, Osis S, Hettinga BA, Ferber R. Kinematic gait patterns in healthy runners: A hierarchical cluster analysis. Journal of Biomechanics. 2015;48(14):3897–904. [DOI] [PubMed] [Google Scholar]
- 18.Hewett TE, Myer GD, Ford KR, et al. Biomechanical measures of neuromuscular control and valgus loading of the knee predict anterior cruciate ligament injury risk in female athletes: A prospective study. Am J Sport Med. 2005;33(4):492–501. [DOI] [PubMed] [Google Scholar]
- 19.Paterno MV, Schmitt LC, Ford KR, et al. Biomechanical measures during landing and postural stability predict second anterior cruciate ligament injury after anterior cruciate ligament reconstruction and return to sport. Am J Sport Med. 2010;38(10):1968–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hewett TE, Ford KR, Xu Y, Khoury J, Myer GD. Utilization of ACL injury biomechanical and neuromuscular risk profile analysis to determine the effectiveness of neuromuscular training. Am J Sport Med. 2016;44(12):3146–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Galloway RT, Xu Y, Hewett TE, et al. Age-dependent patellofemoral pain: Hip and knee risk landing profiles in prepubescent and postpubescent female athletes. Am J Sport Med. 2018;46(11):2761–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Deluzio KJ, Wyss UP, Zee B, Costigan PA, Sorbie C. Principal component models of knee kinematics and kinetics: Normal vs. pathological gait patterns. Hum Movement Sci. 1997;16:201–17. [Google Scholar]
- 23.Federolf PA, Boyer KA, Andriacchi TP. Application of principal component analysis in clinical gait research: Identification of systematic differences between healthy and medial knee-osteoarthritic gait. Journal of Biomechanics. 2013;46(13):2173–8. [DOI] [PubMed] [Google Scholar]
- 24.Cushion EJ, Warmenhoven J, North JS, Cleather DJ. Principal component analysis reveals the proximal to distal pattern in vertical jumping Is governed by two functional degrees of freedom. Frontiers in Bioengineering and Biotechnology. 2019;7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Watari R, Kobsar D, Phinyomark A, Osis S, Ferber R. Determination of patellofemoral pain sub-groups and development of a method for predicting treatment outcome using running gait kinematics. Clinical biomechanics. 2016;38:13–21. [DOI] [PubMed] [Google Scholar]
- 26.Alentorn-Geli E, Myer GD, Silvers HJ, et al. Prevention of non-contact anterior cruciate ligament injuries in soccer players. Part 1: Mechanisms of injury and underlying risk factors. Knee Surgery Sports Traumatology Arthroscopy. 2009;17(7):705–29. [DOI] [PubMed] [Google Scholar]
- 27.Ford KR, Myer GD, Smith RL, Vianello RM, Seiwert SL, Hewett TE. A comparison of dynamic coronal plane excursion between matched male and female athletes when performing single leg landings. Clinical biomechanics. 2006;21(1):33–40. [DOI] [PubMed] [Google Scholar]
- 28.Jr Ward JH. Hierarchical grouping to optimize an objective function. Journal of the American Statistical Association. 1963;58(301):236–44. [Google Scholar]
- 29.Hair JF, Black WC, Babin BJ, Anderson RE, Tatham RL. Multivariate Data Analysis. Upper Saddle River, NJ: Prentice Hall; 1998. [Google Scholar]
- 30.Ford KR, Myer GD, Brent JL, Hewett TE. Hip and knee extensor moments predict vertical jump height in adolescent girls. Journal of Strength and Conditioning Research. 2009;23(4):1327–231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Myer GD, Ford KR, Barber Foss KD, Liu C, Nick TG, Hewett TE. The relationship of hamstrings and quadriceps strength to anterior cruciate ligament injury in female athletes. Clinical Journal of Sports Medicine. 2009;19(1):3–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cohen J. Statistical power analysis for the behavioral sciences. Hillsdale, NJ: Larence Earlbaum Associates; 1988. [Google Scholar]
- 33.Myer GD, Ford KR, Barber Foss KD, et al. The incidence and potential pathomechanics of patellofemoral pain in female athletes. Clinical biomechanics. 2010;25(7):700–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Moran KA, Wallace ES. Eccentric loading and range of knee joint motion effects on performance enhancement in vertical jumping. Hum Movement Sci. 2007;26(6):824–40. [DOI] [PubMed] [Google Scholar]
- 35.Krol H, Mynarski W. A comparison of mechanical parameters between the counter movement jump and drop jump in biathletes. J Hum Kinet. 2012;34:59–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Taube W, Leukel C, Gollhofer A. How neurons make us jump: The neural control of stretch-shortening cycle movements. Exerc Sport Sci Rev. 2012;40(2):106–15. [DOI] [PubMed] [Google Scholar]
- 37.Struzik A, Pietraszewski B, Kawczynski A, Winiarski S, Juras G, Rokita A. Manifestations of proprioception during vertical jumps to specific heights. Journal of Strength and Conditioning Research. 2017;31(6):1694–701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cannon J, Cambridge EDJ, McGill SM. Anterior cruciate ligament injury mechanisms and the kinetic chain linkage: The effect of proximal joint stiffness on distal knee control during bilateral landings. Journal of Orthopaedic & Sports Physical Therapy. 2019;59(8):601–10. [DOI] [PubMed] [Google Scholar]
- 39.Butler RJ, Crowell HP 3rd, Davis IM Lower extremity stiffness: Implications for performance and injury. Clinical biomechanics. 2003;18(6):511–7. [DOI] [PubMed] [Google Scholar]
- 40.Ford KR, Myer GD, Hewett TE. Longitudinal effects of maturation on lower extremity joint stiffness in adolescent athletes. Am J Sport Med. 2010;38(9):1829–237. [DOI] [PMC free article] [PubMed] [Google Scholar]