

Abstract
Nervous necrosis virus (NNV) is one of the fish pathogens that have caused mass mortalities of many marine and freshwater fishes in the world. To better comprehend the molecular immune mechanism of sea perch (Lateolabrax japonicus) against NNV infection, the comparative transcriptome analysis of red‐spotted grouper nervous necrosis virus (RGNNV)‐infected or mock‐infected L. japonicus brain (LJB) cells was performed via RNA sequencing technology. Here, 1,969 up‐regulated genes and 9,858 down‐regulated genes, which were widely implicated in immune response pathways, were identified. Furthermore, we confirmed that p53 signalling pathway was repressed at 48 hr post‐RGNNV infection, as indicated by up‐regulation of Mdm2 and down‐regulation of p53 and its downstream target genes, including Bax, Casp8 and CytC. Overexpression of L. japonicus p53 (Ljp53) significantly inhibited RGNNV replication and up‐regulated the expression of apoptosis‐related genes, whereas the down‐regulation caused by pifithrin‐α led to the opposite effect, suggesting Ljp53 might promote cell apoptosis to repress virus replication. Luciferase assay indicated that Ljp53 could enhance the promoter activities of zebrafish interferon (IFN)1, indicating that Ljp53 could exert its anti‐RGNNV activities by enforcing the type I IFN response. This study revealed the potential antiviral role of p53 during NNV infection.
Keywords: apoptosis, Lateolabrax japonicas, p53, red‐spotted grouper nervous necrosis virus, transcriptome
1. INTRODUCTION
Sea perch, Lateolabrax japonicus, an economically important species, is widely cultured in Asia, especially in China. It has been increasingly threatened by various diseases caused by bacteria and viruses due to its extensive farming (Jia, Jia, & Yi, 2015; Zhao, Fu, Wang, Jiao, & Qiu, 2016). Of which, viral nervous necrosis (VNN), caused by nervous necrosis virus (NNV), has been widely reported in more than 40 cultured fish, including sea perch and grouper, and resulted in significant economic losses (Nakai et al., 2009). However, there is still no effective method to prevent or control VNN. In teleost fish, the innate immune system plays a critical role in host against virus infection, but studies on the immune system of sea perch are still limited. In recent years, researches on immunity of sea perch against NNV infection were mainly focused on single genes, and only a few immune‐related genes, including LGP2 (Jia, Zhang et al., 2015), MDA5 (Jia, Jia et al., 2016), MAVS (Jia, Jin et al., 2016), TRAF3 (Zhang et al., 2018), HSP27 (Le, Jia et al., 2017) and RAVER1 (Jia et al., 2017), have been identified and characterized in sea perch. However, knowledge about the immunity system of sea perch, especially the immune response‐related signalling pathways implicated in NNV infection, still remains incomplete. Systematic study of sea perch immune‐related signalling pathway is necessary to provide the fundamental understanding of the interaction between sea perch and NNV.
Transcriptome analysis is a fast and cost‐effective way to understand the underlying pathways and mechanisms of host against pathogen infection by evaluating immune responses of gene expression (Liu, Wang, Kwang, Yue, & Wong, 2016; Zhong et al., 2017). Many aquatic species have been sequenced to study pathogenic processes during virus infection, including mandarin (Hu et al., 2015), orange‐spotted grouper (Huang et al., 2011), Pacific white shrimp (Zeng et al., 2013) and rainbow trout (Aquilino, Castro, Fischer, & Tafalla, 2014).
It was well known that the brain and retina were the main target organs of NNV (Poisa‐Beiro et al., 2008). Therefore, cells originated from sea perch brain should be a good material for studying sea perch–NNV interaction. In our previous study, a continuous cell line L. japonicus brain (LJB), derived from the brain of sea perch, was established and exhibited susceptibility to red‐spotted grouper nervous necrosis virus (RGNNV) (Le, Li et al., 2017). In this study, transcriptome sequencing libraries were constructed with RGNNV‐infected or mock‐infected LJB cells. Based on the analysis of differentially expressed genes (DEGs), we found that p53 signalling pathway might be involved in the immune response against RGNNV, and experimentally revealed the role of L. japonicus p53 (Ljp53) in the regulation of the type I IFN response and cellular apoptosis during RGNNV infection. This study provides insight into the immune response of sea perch against RGNNV infection and the important role of Ljp53 in inhibiting RGNNV infection.
2. MATERIALS AND METHODS
2.1. Cells, virus stock and plasmid
Lateolabrax japonicus brain cells were grown and maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 15% foetal bovine serum (FBS) (Gibco) at 28°C (Le, Li et al., 2017). HEK293T (human embryonic kidney 293) cells were maintained in DMEM supplemented with 10% FBS at 37°C in a 5% CO2 incubator (Russell, Graham, Smiley, & Nairn, 1977).
Red‐spotted grouper nervous necrosis virus was isolated from diseased sea perch in Guangdong Province of China and kept in our laboratory (Jia, Jia et al., 2015). RGNNV was propagated in LJB cells at 28°C, and virus titres were detected by 50% tissue culture infective dose (TCID50) (Jia, Zhang et al., 2015).
DrIFN1‐pro‐luc plasmid was kindly provided by Professor Yibing Zhang at the Institute of Hydrobiology, Chinese Academy of Sciences (Zhang & Gui, 2012).
2.2. RNA preparation, library construction and transcriptome sequencing
Lateolabrax japonicus brain cells were infected with RGNNV (multiplicity of infection [MOI] = 5) at 28°C for 4 hr. Then, the medium containing RGNNV was discarded and the same volume of growth medium with 15% FBS was added. The control (mock‐infected LJB cells) was treated with the same volume of medium. RGNNV‐infected or mock‐infected LJB cells were harvested for RNA isolation at 48 hr post‐infection (hpi), respectively. Total RNA was extracted using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. The concentration of total RNA was determined using NanoDrop 2000 UV‐Vis Spectrophotometer.
For cDNA library construction and sequencing, mRNA was first isolated from total RNA treated with DNase I using Magnetic Oligo (dT) Beads (Illumina) and was fragmented. Then, the double‐stranded cDNA was synthesized with random hexamer primers and was further subjected to end‐repair and adapter ligation using T4 DNA ligase. The products of ligation reaction were purified on 2% agarose gel, and cDNA fragments (about 200 bp) were recovered. PCR was carried out to enrich the purified cDNA template. Finally, the cDNA library was constructed. After validating on an Agilent Technologies 2100 Bioanalyzer, the library was sequenced using Illumina HiSeq 4000 according to the manufacturer's instruction. All data sets have been submitted to the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) database PRJNA497762.
2.3. De novo assembly and functional annotation
After sequencing, we carried out a stringent filtering process of raw sequencing reads as described previously (Gui et al., 2013). The raw reads were cleaned by removing adapter sequences, non‐coding RNA, low‐quality sequences (reads with ambiguous bases “N” and the ratio of “N” > 10%) and reads with average length <20 bases. De novo transcriptome assembly was performed by Trinity program (v2.0.6) as described elsewhere (full‐length transcriptome assembly from RNA‐seq data without a reference genome). The function of unigenes was annotated depending upon following databases: NR (NCBI non‐redundant protein sequences), NT (NCBI nucleotide sequences), GO (Gene Ontology), COG (Clusters of Orthologous Groups of proteins), KEGG (Kyoto Encyclopedia of Genes and Genomes) and Swiss‐Prot (a manually annotated and reviewed protein sequence database).
2.4. Gene expression level analysis, DEG identification and classification
For differential gene expression analysis, reads per kilobase of exon model per million mapped reads value was used to normalize the gene expression levels (Poisa‐Beiro et al., 2008). Statistical comparison between two different groups was conducted using a web tool DESeq (http://www-huber.embl.de/users/anders/DESeq) (Lu et al., 2017). False discovery rate (FDR) <0.05 was used as the threshold of p‐value in multiple tests to judge the significance of gene expression difference. Genes were considered differentially expressed in a given library when the p‐value <0.05, and a greater than twofold change (absolute value of log2 ratio >1) in expression across libraries was observed. GO and KEGG pathway enrichment of differential unigenes were analysed. KO analysis of differential expression unigenes in metabolic pathways was conducted on website (http://www.genome.jp/kegg/tool/map_pathway2.html).
2.5. Expression analysis of DEGs involved in p53 signalling pathway post‐RGNNV challenge
Red‐spotted grouper nervous necrosis virus infection and sample collection were performed as described above. Total RNA was extracted using TRIzol reagent (Invitrogen) and reverse‐transcribed into cDNA by PrimeScript™ First Strand cDNA Synthesis Kit (Takara) according to the manufacturer's instructions. The relative expression levels of Mdm2, Mdm4, p53, Bax, Casp8 and Casp9 were analysed by quantitative real‐time PCR (qRT‐PCR). qRT‐PCR was performed as described previously (Le, Li et al., 2017). Primers for qRT‐PCR are listed in Supporting Information Table S1. Expression levels of target genes were normalized with β‐actin of sea perch by the 2−ΔΔCT methods. Data from each sample were shown as mean ± SD from three independent experiments in triplicates.
2.6. Effects of Ljp53 on RGNNV replication
To investigate the effects of Ljp53 overexpression on RGNNV replication, encoding region of Ljp53 was amplified by PCR using gene‐specific primers and sub‐cloned into pcDNA 3.1 (+) vectors (Invitrogen) to generate pcDNA‐Ljp53, which was confirmed through DNA sequencing analysis (Supporting Information Table S1). LJB cells in six‐well plates at 70%–80% confluence were transfected with pcDNA‐Ljp53 or pcDNA 3.1(+) using Lipofectamine 3000 (Invitrogen) according to the manufacturer's instruction. At 24 hr post‐transfection, cells were infected with RGNNV (MOI = 1) and harvested at 48 hpi for total RNA isolation. The expression of RNA‐dependent RNA polymerase (RDRP) was detected by qRT‐PCR as described above. Primers for qRT‐PCR are listed in Supporting Information Table S1.
To evaluate the effect of down‐regulation of Ljp53 on RGNNV replication, LJB cells in six‐well plates at 70%–80% confluence were treated with p53 inhibitor pifithrin‐α (MedChemExpress) at concentration of 20 μM or DMSO (0.1%) for 2 hr, and then, the treated cells were infected with RGNNV (MOI = 1) in the presence of pifithrin‐α or DMSO. Cells were harvested for qRT‐PCR at 48 hpi. qRT‐PCR was performed as described above. Primers for qRT‐PCR are listed in Supporting Information Table S1.
2.7. Luciferase activity assay
Luciferase activity assay was carried out as described previously (Jia et al., 2017). In brief, the plasmids pRL‐TK (Promega), pcDNA‐Ljp53 or pcDNA 3.1(+), and DrIFN1‐pro‐luc were co‐transfected into HEK293T cells using Lipofectamine 3000. At 48 hr post‐transfection, cells were harvested with passive lysis buffer and subjected to luciferase activity measurement using GloMax 20/20 Luminometer (Promega). Data were expressed as mean ± SD from three independent experiments performed in triplicates.
2.8. Expression analysis of apoptosis‐related genes
To evaluate the effect of Ljp53 on apoptosis‐related genes during RGNNV infection, the encoding region of Ljp53 was sub‐cloned into pEGFP‐N3 vectors (Invitrogen) to generate pEGFP‐Ljp53, which was confirmed through DNA sequencing analysis (Supporting Information Table S1). LJB cells were transfected with pEGFP‐Ljp53 or pEGFP‐N3 or treated with pifithrin‐α as above. Finally, cells were harvested for RNA isolation at 48 hpi. The relative expression levels of Casp3, Casp9 and CytC were analysed by qRT‐PCR. Primers for qRT‐PCR are listed in Supporting Information Table S1.
2.9. Statistics analyses
All statistics were calculated using SPSS version 19. Differences between control and treatment groups were assessed by one‐way ANOVA. p < 0.05 (*) and p < 0.01 (**) were, respectively, considered to be statistically significant and very significant.
3. RESULTS
3.1. De novo assembly and function annotation
To understand the gene expression patterns of LJB cells in response to RGNNV infection, RNA‐seq libraries were generated from RGNNV‐infected and mock‐infected LJB cells. A total of 4.751 × 107 and 4.525 × 107 raw reads were produced in mock‐infected and RGNNV‐infected samples, respectively. After filtration, 4.51 × 107 and 4.485 × 107 clean reads obtained from the mock‐infected sample (control group) and RGNNV‐infected group were used for de novo assembly (Table 1). In control group, transcriptome assembly yielded 38,354 unigenes with a mean length of 1,067 and N50 length of 1,979, and in RGNNV‐infected group, 30,209 unigenes with a mean length of 697 and N50 length of 1,110 were produced. Moreover, combined transcriptome assembly of the two groups generated 39,954 unigenes with a mean length of 1,038 and N50 length of 1,931 (Table 2). The length distribution of these unigenes is shown in Figure 1.
Table 1.
Information and quality of transcriptome sequencing data
| Sample | Total raw reads (Mb) | Total clean reads (Mb) | Total clean bases (Gb) | Clean reads Q20 (%) | Clean reads Q30 (%) | Clean reads ratio (%) |
|---|---|---|---|---|---|---|
| Ljb_control | 47.51 | 45.1 | 4.51 | 94.25 | 87.07 | 94.93 |
| Ljb_nnv | 45.25 | 44.85 | 4.48 | 94.57 | 87.54 | 99.12 |
This article is being made freely available through PubMed Central as part of the COVID-19 public health emergency response. It can be used for unrestricted research re-use and analysis in any form or by any means with acknowledgement of the original source, for the duration of the public health emergency.
Table 2.
Summary of de novo assembly of transcriptomic profiles
| Sample | Total number | Total length | Mean length | N50 | N70 | N90 | GC (%) |
|---|---|---|---|---|---|---|---|
| Ljb_control | 38,354 | 40,935,337 | 1,067 | 1,979 | 1,119 | 404 | 48.37 |
| Ljb_nnv | 30,209 | 21,081,274 | 697 | 1,110 | 575 | 279 | 48.4 |
| All‐Unigenes | 39,954 | 41,498,627 | 1,038 | 1,931 | 1,085 | 391 | 48.18 |
This article is being made freely available through PubMed Central as part of the COVID-19 public health emergency response. It can be used for unrestricted research re-use and analysis in any form or by any means with acknowledgement of the original source, for the duration of the public health emergency.
Figure 1.

Length distribution of All‐unigenes
After assembly, 39,954 unigenes were annotated based on six databases including NR, NT, GO, COG, KEGG and Swiss‐Prot. A total of 32,685 unigenes were matched in at least one database. Of all the unigenes, 24,738 (61.92%), 21,854 (54.68%), 18,029 (45.12%) and 9,001 (22.53%) unigenes showed significant matches (E‐value ≤ 1E‐5) against the databases of NR, Swiss‐Prot, KEGG and COG, respectively (Table 3). GO analysis assigned 1,569 unigenes to biological process (BP), 1,078 to cellular component (CC) and 649 to molecular function (MF). For BP, the most assigned genes were cellular process with 294 unigenes. For CC, the most assigned genes were cell and cell part with 234 unigenes. For MF, binding with 292 unigenes was the most highly represented group (Figure 2). COG classification indicated that 9,001 unigenes were assigned to 25 different COG categories. The cluster for “General function prediction only” represented the largest group (3,237 unigenes), followed by “replication, recombination and repair” (1,426 unigenes) and “transcription” (1,375 unigenes) (Figure 3). A total of 18,029 unigenes were assigned to 301 KEGG pathways, covering six main categories, include cellular processes, environmental information processing, genetic information processing, human diseases, metabolism and organismal systems. Metabolic pathway (1,883 unigenes) was the most abundant KEGG pathway, followed by “pathway in cancer” (844 unigenes) and “focal adhesion” (678 unigenes) (Supporting Information Table S2).
Table 3.
Summary of annotation statistics of assembled transcripts
| Values | Total | Nr | Nt | Swiss‐Prot | KEGG | COG | GO | Overall |
|---|---|---|---|---|---|---|---|---|
| Number | 39,954 | 24,738 | 31,651 | 21,845 | 18,029 | 9,001 | 518 | 32,685 |
| Percentage | 100 | 61.92 | 79.22 | 54.68 | 45.12 | 22.53 | 1.30 | 81.81 |
COG: Clusters of Orthologous Groups; GO: Gene Ontology; KEGG: Kyoto Encyclopedia of Genes and Genomes.
This article is being made freely available through PubMed Central as part of the COVID-19 public health emergency response. It can be used for unrestricted research re-use and analysis in any form or by any means with acknowledgement of the original source, for the duration of the public health emergency.
Figure 2.

Gene Ontology function classification of All‐unigenes [Colour figure can be viewed at http://www.wileyonlinelibrary.com]
Figure 3.

Clusters of Orthologous Groups (COG) function classification of All‐unigenes [Colour figure can be viewed at http://www.wileyonlinelibrary.com]
3.2. Differentially expressed genes
To identify DEGs, the unigenes between LJB‐Control and LJB‐NNV groups were compared. A large number of genes were found to be differentially expressed in LJB cells post‐RGNNV infection, including 1,969 up‐regulated genes (DUGs) and 9,858 down‐regulated genes (DDGs) (FDR ≤ 0.001 and log2 ratio ≥ 1) (Supporting Information Table S3). Of these genes, many well‐known immune‐related genes were strongly inhibited after RGNNV infection, including NOD1, TLR2, IRF3, STAT1, IFN‐stimulated gene 15 (ISG15). Among all the DDGs, heat‐shock protein beta‐1 (HSPB1), also named HSP27, was mostly down‐regulated (decreased by 18.60 times).
Gene Ontology function and KEGG pathway enrichment analysis were performed to identify the biological function of DEGs. Based on GO classification, DEGs were mainly divided into three categories: BP (129 DEGs), CC (105 DEGs) and MF (158 DEGs). Furthermore, KEGG mapping annotated DEGs to 311 signalling pathways. Lots of DEGs were related to immune‐related signalling pathways, such as MAPK signalling pathway (228 DEGs), Epstein–Barr virus infection (199 DEGs), p53 signalling pathway (79 DEGs), NF‐κB signalling pathway (74 DEGs), NOD‐like receptor signalling pathway (59 DEGs) and RIG‐I‐like receptor signalling pathway (43 DEGs) (Supporting Information Table S4).
3.3. qRT‐PCR verification for DEGs related to p53 signalling pathway
Comparative transcriptome analysis showed that 79 DEGs involved in p53 signalling pathway were regulated in RGNNV‐infected LJB cells compared to control cells (Supporting Information Table S5). A majority of these DEGs were down‐regulated, such as p53, Fas, CytC and Casp8, but only six DEGs, including Mdm2, Cdc2, Igf, P21, Gadd45 and Igf‐bp3, were up‐regulated. To verify the expression pattern of DEGs involved in p53 signalling pathway, the transcriptional levels of six randomly selected DEGs relating to p53 signalling pathway (Mdm2, Mdm4, p53, Bax, Casp8 and CytC) were detected by qRT‐PCR. As shown in Figure 4, the expression levels of these genes showed similar fold change to the transcriptome data. Meanwhile, in accordance with the high‐throughput sequencing data, the expressions of all these DEGs were significantly reduced post‐RGNNV infection, except for the expression of Mdm2 was up‐regulated by 5.2 times at 48 hpi (Supporting Information Figure S1). These results suggested that the expression profiling of DEGs determined by RNA‐seq was reliable. These results indicated that p53 signalling pathway was not only involved in RGNNV infection but also inhibited by RGNNV at 48 hpi.
Figure 4.
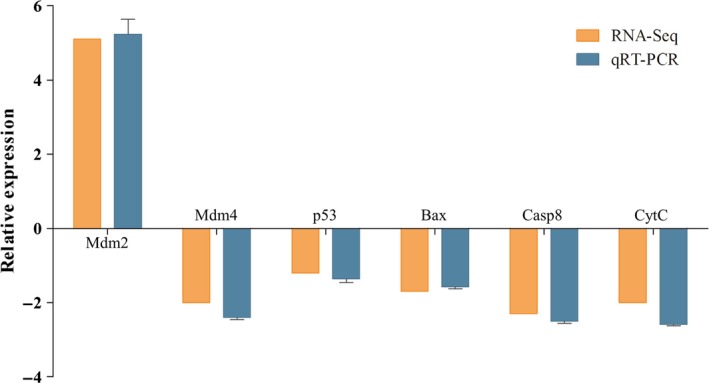
Comparison of the fold change expression of six differentially expressed genes involved in p53 signalling pathway as determined by RNA‐seq and qPCR. The results of qPCR were evaluated by normalizing to β‐actin gene [Colour figure can be viewed at http://www.wileyonlinelibrary.com]
3.4. Ljp53 functioned as an anti‐RGNNV factor
Given the important role of p53 in innate antiviral immunity, in the present work, Ljp53 was further cloned and analysed. Fortunately, the partial sequence of Ljp53 obtained from high‐throughput sequencing data contained the complete coding sequence of Ljp53 (GenBank accession number MH940221). To elucidate the role of Ljp53 during RGNNV infection in vitro, the transcription level of RDRP was detected in Ljp53 overexpressing LJB cells. As shown in Figure 5a,b, a significantly attenuated expression of RDRP was observed in Ljp53 expressing LJB cells, whereas pifithrin‐α caused down‐regulation of Ljp53 led to opposite results, suggesting the antiviral role of Ljp53 during RGNNV infection. Many works have demonstrated that p53 contributed to the innate antiviral response by enhancing type I IFN‐dependent antiviral activity (Ding et al., 2018; Muñoz‐Fontela et al., 2008). To further reveal the potential antiviral mechanism of Ljp53, the luciferase reporter assay was performed. Our results indicated that zebrafish IFN1 promoter was activated by Ljp53 overexpression (Figure 5c), indicating that Ljp53 might exert its anti‐RGNNV activity partially by activating the IFN signalling pathway.
Figure 5.

Effects of Ljp53 on RGNNV replication and zebrafish IFN1 promoter activity. (a) qRT‐PCR detection of RDRP in LJB cells transfected with pcDNA‐Ljp53 or control plasmid at 48 hr post‐RGNNV infection. (b) qRT‐PCR detection of RDRP in LJB cells treated with DMSO (control) or pifithrin‐α. (c) HEK293T cells were analysed at 48 hr after transfected with the pRL‐TK, pcDNA3.1(+) or pcDNA‐Ljp53 plasmids together with zebrafish IFN1 promoter‐driven reporter plasmid, and the luciferase activity was determined. Asterisks indicate significant differences between groups (**p < 0.01) [Colour figure can be viewed at http://www.wileyonlinelibrary.com].
3.5. Ljp53 promoted the expression of apoptotic genes
To validate the antiviral role of Ljp53 was associated with its pro‐apoptotic activity in RGNNV infection, the mRNA levels of Casp3, Casp9 and CytC were measured in Ljp53 overexpressing LJB cells. Our results showed that overexpression of Ljp53 up‐regulated the transcription of pro‐apoptotic Casp3, Casp9 and CytC at 48 hpi, whereas the suppression of Ljp53 caused by pifithrin‐α led to the opposite effect (Figure 6). Thus, our results suggested that the apoptosis induced by RGNNV infection was dependent to some extent on Ljp53, and Ljp53 might be involved in promoting apoptosis in LJB cells.
Figure 6.

Expression of apoptosis‐related genes. qRT‐PCR detection of Casp3 (a), Casp9 (b) and CytC (c) in LJB cells transfected with pEGFP‐N3 or pEGFP‐Ljp53 plasmid at 48 hr post‐RGNNV infection. qRT‐PCR detection of Casp3 (d), Casp9 (e) and CytC (f) in LJB cells treated with DMSO or pifithrin‐α. Asterisks indicate the significant differences between groups at 48 hpi (*p < 0.05; **p < 0.01) [Colour figure can be viewed at http://www.wileyonlinelibrary.com].
4. DISCUSSION
Sea perch is a commercially important marine fish widely cultured in Asia. However, the high mortality in a wide range of larvae and juveniles caused by NNV is an intensive threat. Nevertheless, there is limited transcriptomic information available in relation to the molecular immune mechanism of sea perch against NNV infection. In this study, transcriptome sequencing libraries were generated from RGNNV‐infected or mock‐infected LJB cells. In total, 4.51 × 107 and 4.485 × 107 clean reads were generated from the control group and RGNNV‐infected group, and 39,954 unigenes were annotated by six databases. And the number of unigenes was slightly different from previous reports in which the fish were infected with different pathogens (Lu et al., 2017; Zhao et al., 2016).
Gene comparative screening identified a lot of DEGs, including several known immune‐related genes, such as IRF3, STAT1 and ISG15. Previous reports indicated that ISG15 could be induced by RGNNV infection in European sea bass and Senegalese sole (Álvarez‐Torres et al., 2017; Moreno, Garcia‐Rosado, Borrego, & Alonso, 2016). However, in this study, the expression of ISG15 was reduced at 48 hpi. ISG15, as an antiviral effector, was demonstrated to play important roles in defence against RGNNV infection in teleost (Liu et al., 2018; Moreno, Alvarez‐Torres, Garcia‐Rosado, Borrego, & Alonso, 2018). Many viruses have developed different mechanisms to escape the antiviral response, including counteracting the activity of ISG15 (Guerra, Cáceres, Knobeloch, Horak, & Esteban, 2008; Yuan & Krug, 2001). Several studies have addressed NNV persistent infections in many fish, indicating NNV has developed some mechanisms to antagonize host innate antiviral responses (Lu et al., 2016; Zhang et al., 2017). We speculated that RGNNV might counteract the antiviral activity of ISG15 to escape the host antiviral response in LJB cells. Zinc finger protein 395 (ZNF395) is one of the most down‐regulated genes (decreased by 18.11 times) in this study. ZNF395, as a novel hypoxia‐inducible transcription factor, contributes to the maximal stimulation of a subset of ISGs, such as ISG15, IFIT1/ISG56, IFI44, CXCL10 and CXCL11 (Schroeder, Herwartz, Jordanovski, & Steger, 2017). In this study, in line with the down‐regulation of ISG15, the expression of ZNF395 was also decreased. Given these results, it is possible that the declining level of ZNF395 may impair host antiviral responses by reducing the expression of ISGs involved in the innate immune response during RGNNV infection. It was known that ZNF395‐mediated activation of ISGs was depended on IKK signalling (Jordanovski et al., 2013). In present work, the expression levels of IKKα and IKKβ, the catalytic subunits of IKK, were also repressed post‐RGNNV infection. Further experiments would be necessary to determine the association of ISG15, ZNF395 and IKK signalling in sea perch. In addition, among these DEGs, HSPB1, also named HSP27, was down‐regulated, similar to our previous study (Le, Jia et al., 2017). HSPB1 has been reported to be widely involved in pathophysiology of oxidative stress and apoptosis. Overexpression of HSPB1 protected cells from oxidative damage and apoptosis triggered by Cd exposure (Mun̄oz‐Fontela, 2005). In our previous study, overexpression of LjHSP27 inhibited RGNNV‐induced apoptosis, supporting the vital role of HSP27 as an anti‐apoptosis protein (Le, Jia et al., 2017). It has been known that many DNA and RNA viruses can trigger oxidative stress and induce host cell death in infected cells (Casavant et al., 2006; Yuan et al., 2015). The role of LjHSP27 against oxidative stress still needs further research.
Published literature about the immune‐related signalling pathways involved in RGNNV infection was mainly focused either on the apoptosis pathway or on the innate immune pathways (Chen et al., 2017; Wang, Rajanbabu, & Chen, 2015). Recently, numbers of studies have shown that p53 signalling pathway is involved in the process of different virus infection and replication. For example, porcine circovirus type 2 infection induced S phase accumulation to promote viral replication in host cells via activation of the p53 signalling pathway (Xu et al., 2016). Classical swine fever virus Shimen regulated p53 signalling pathway to subvert host innate immune response (Ning et al., 2017). Given the important role of p53 signalling pathway during virus infection, in this study, we focused our analysis on the DEGs associated with p53 signalling pathway. qRT‐PCR results indicated that p53 signalling pathway was involved in RGNNV infection and inhibited by RGNNV. In p53 signalling pathway, Mdm2 and Mdm4 are major negative regulators of p53 (Chai et al., 2018; Raja, Ronsard, Lata, Trivedi, & Banerjea, 2017). A significant negative correlation was also found between Mdm2 and Ljp53 in this study. Thus, we speculated that RGNNV might repress the expression of Ljp53 by activating Mdm2 during RGNNV infection. As Mdm2 has been initially identified as a required host factor for viral replication (Raja et al., 2017), we cannot exclude an ambivalent role of Mdm2 during infection, as a result of its multiple functions and interactions with other host factors. In contrast, there was a positive correlation between Mdm4 and Ljp53. Previous study reported that Mdm2 targeted Mdm4 for proteasomal degradation in response to DNA damage (Kawai et al., 2003). Thus, we speculated that sea perch Mdm2 might also bind to Mdm4 for degradation during RGNNV infection. Nevertheless, to clarify the relationship of sea perch Mdm2 and Mdm4, more experiments will be required.
Tumour suppressor p53, primarily famous for its vital role in protecting against cancer development, has been proven to regulate various biological process, such as cell cycle arrest, cellular metabolism, cellular apoptosis and innate antiviral immunity (Liu, Zhang, Hu, & Feng, 2015). Several studies indicated that p53 functioned as a key player in innate antiviral immunity by both enforcing the type I IFN response and inducing apoptosis in virus‐infected cells (Rivas, Aaronson, & Munoz‐Fontela, 2010). Here, the expression of RDRP was significant reduced in Ljp53 overexpressing cells post‐RGNNV infection, whereas pifithrin‐α caused opposite results. Furthermore, the results of luciferase reporter assay showed that zebrafish IFN1 promoter was activated by Ljp53 overexpression. Thus, we speculated that Ljp53 inhibited RGNNV replication by activating the IFN signalling pathway.
It was known that the role of p53 in the control of virus infection was also associated with its ability to activate apoptosis during virus infection, which inhibited virus replication (Rivas et al., 2010). Several studies have reported that p53 can induce apoptosis through activating death receptor pathway or mitochondrial pathway (Benchimol, 2001; Moll & Zaika, 2001). Our previous study demonstrated that RGNNV infection caused apoptosis at 6 and 24 hpi in LJB cells (Le, Jia et al., 2017). Meanwhile, our results indicated that three apoptosis‐related genes, including Casp3, Casp9 and CytC, were notably enhanced as the ectopic expression of Ljp53 in vitro, whereas pifithrin‐α caused the opposite effect. In addition, significantly higher level of caspase 3 activities was observed in Ljp53 overexpressing cells post‐RGNNV infection. All these results indicated the pro‐apoptotic function of Ljp53 (Supporting Information Figure S2). It has been known that p53‐dependent apoptosis could be used by host as a useful mechanism to repress virus infection (Mun̄oz‐Fontela et al., 2005; Turpin et al., 2005). On the other hand, many viruses have also developed strategies to manipulate host p53 signalling pathways to increase virus replication (Casavant et al., 2006; Royds et al., 2006). For example, coronavirus inhibited p53‐mediated host apoptosis to ensure viral growth in infected cells (Yuan et al., 2015). In our study, RGNNV infection reduced the expression of Ljp53 at 48 hpi, which at least partially inhibited the host cell apoptosis. We speculated that RGNNV might repress the p53‐dependent apoptosis to promote virus replication in RGNNV‐infected LJB cells and facilitate transmission of newly formed viral particles to other cells.
In this study, transcriptome sequencing libraries were constructed with LJB cells mock‐infected or infected with RGNNV at 48 hpi. A large number of DEGs in response to RGNNV infection were identified. The DEGs involved in p53 signalling pathway were further investigated. Our results demonstrated that Ljp53 played an essential role in inhibiting RGNNV replication. Mechanistically, we confirmed that the antiviral role of Ljp53 was involved in its activity in mediating the IFN response and its pro‐apoptotic activity.
CONFLICT OF INTEREST
The authors declare no competing financial interests.
Supporting information
ACKNOWLEDGEMENTS
This work was supported by the National Natural Science Foundation of China (31502195, 31602191), the Pearl River S & T Nova Program of Guangzhou (201806010047), the Science and Technology Planning Project of Guangdong Province (2017A030303010), the Natural Science Foundation of Guangdong Province (2015A030308012), the Zhuhai Scholar Professor Program (2015) and the Fundamental Research Funds for the Central Universities (17lgpy61).
Xiang Y, Jia P, Liu W, Yi M, Jia K. Comparative transcriptome analysis reveals the role of p53 signalling pathway during red‐spotted grouper nervous necrosis virus infection in Lateolabrax japonicus brain cells. J Fish Dis. 2019;42:585–595. 10.1111/jfd.12960
Contributor Information
Meisheng Yi, Email: yimsh@mail.sysu.edu.cn.
Kuntong Jia, Email: jiakt3@mail.sysu.edu.cn.
REFERENCES
- Álvarez‐Torres, D. , Podadera, A. M. , Alonso, M. C. , Bandín, I. , Béjar, J. , & García‐Rosado, E. (2017). Molecular characterization and expression analyses of the Solea senegalensis interferon‐stimulated gene 15 (isg15) following NNV infections. Fish & Shellfish Immunology, 66, 423–432. 10.1016/j.fsi.2017.05.040 [DOI] [PubMed] [Google Scholar]
- Aquilino, C. , Castro, R. , Fischer, U. , & Tafalla, C. (2014). Transcriptomic responses in rainbow trout gills upon infection with viral hemorrhagic septicemia virus (VHSV). Developmental & Comparative Immunology, 44, 12–20. 10.1016/j.dci.2013.11.006 [DOI] [PubMed] [Google Scholar]
- Benchimol, S. (2001). p53‐dependent pathways of apoptosis. Cell Death & Differentiation, 8, 1049–1051. 10.1038/sj.cdd.4400918 [DOI] [PubMed] [Google Scholar]
- Casavant, N. C. , Luo, M. H. , Rosenke, K. , Winegardner, T. , Zurawska, A. , & Fortunato, E. A. (2006). Potential role for p53 in the permissive life cycle of human cytomegalovirus. Journal of Virology, 80, 8390–8401. 10.1128/JVI.00505-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chai, K. , Ning, X. , Nguyễn, T. , Zhong, B. , Morinaga, T. , Li, Z. , … Tagawa, M. (2018). Heat shock protein 90 inhibitors augment endogenous wild‐type p53 expression but down‐regulate the adenovirally‐induced expression by inhibiting a proteasome activity. Oncotarget, 9, 26130–26143. 10.18632/oncotarget.25452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, W. , Yi, L. , Feng, S. , Liu, X. , Asim, M. , Zhou, Y. , … Lin, L. (2017). Transcriptomic profiles of striped snakehead fish cells (SSN‐1) infected with red‐spotted grouper nervous necrosis virus (RGNNV) with an emphasis on apoptosis pathway. Fish & Shellfish Immunology, 60, 346–354. 10.1016/j.fsi.2016.11.059 [DOI] [PubMed] [Google Scholar]
- Ding, L. , Li, J. , Li, W. , Fang, Z. , Li, N. , Guo, Q. , … Hong, M. (2018). p53 mediated IFN‐β signaling to affect viral replication upon TGEV infection. Veterinary Microbiology, 227, 61–68. 10.1016/j.vetmic.2018.10.025 [DOI] [PubMed] [Google Scholar]
- Guerra, S. , Cáceres, A. , Knobeloch, K. , Horak, I. , & Esteban, M. (2008). Vaccinia virus E3 protein prevents the antiviral action of ISG15. PLoS Pathogens, 4, e1000096 10.1371/journal.ppat.1000096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gui, D. , Jia, K. , Xia, J. , Yang, L. , Chen, J. , Wu, Y. , … Yi, M. (2013). De novo assembly of the indo‐pacific humpback dolphin leucocyte transcriptome to identify putative genes involved in the aquatic adaptation and immune response. PLoS ONE, 8, e72417 10.1371/journal.pone.0072417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu, X. , Fu, X. , Li, N. , Dong, X. , Zhao, L. , Lan, J. , … Lin, L. (2015). Transcriptomic analysis of mandarin fish brain cells infected with infectious spleen and kidney necrosis virus with an emphasis on retinoic acid‐inducible gene 1‐like receptors and apoptosis pathways. Fish & Shellfish Immunology, 45, 619–629. 10.1016/j.fsi.2015.05.007 [DOI] [PubMed] [Google Scholar]
- Huang, Y. , Huang, X. , Yan, Y. , Cai, J. , Ouyang, Z. , Cui, H. , … Qin, Q. (2011). Transcriptome analysis of orange‐spotted grouper (Epinephelus coioides) spleen in response to Singapore grouper iridovirus. BMC Genomics, 12, 556 10.1186/1471-2164-12-556 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jia, P. , Jia, K. , Chen, L. , Le, Y. , Jin, Y. , Zhang, J. , … Yi, M. (2016). Identification and characterization of the melanoma differentiation – Associated gene 5 in sea perch, Lateolabrax japonicus . Developmental & Comparative Immunology, 61, 161–168. 10.1016/j.dci.2016.03.029 [DOI] [PubMed] [Google Scholar]
- Jia, P. , Jia, K. T. , & Yi, M. (2015). Complete genome sequence of a fish nervous necrosis virus isolated from sea perch (Lateolabrax japonicus) in China. Genome Announcements, 3, e00048–15. 10.1128/genomeA.00048-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jia, P. , Jin, Y. , Chen, L. , Zhang, J. , Jia, K. , & Yi, M. (2016). Molecular characterization and expression analysis of mitochondrial antiviral signaling protein gene in sea perch, Lateolabrax japonicus . Developmental & Comparative Immunology, 55, 188–193. 10.1016/j.dci.2015.10.014 [DOI] [PubMed] [Google Scholar]
- Jia, P. , Liu, W. , Chen, L. , Jin, Y. , Zhang, J. , Jia, K. , … Yi, M. (2017). Identification of sea perch (Lateolabrax japonicus) ribonucleoprotein PTB‐Binding 1 involved in antiviral immune response against RGNNV. Fish & Shellfish Immunology, 60, 119–128. 10.1016/j.fsi.2016.11.047 [DOI] [PubMed] [Google Scholar]
- Jia, P. , Zhang, J. , Jin, Y. , Zeng, L. , Jia, K. , & Yi, M. (2015). Characterization and expression analysis of laboratory of genetics and physiology 2 gene in sea perch, Lateolabrax japonicus . Fish & Shellfish Immunology, 47, 214–220. 10.1016/j.fsi.2015.09.004 [DOI] [PubMed] [Google Scholar]
- Jordanovski, D. , Herwartz, C. , Pawlowski, A. , Taute, S. , Frommolt, P. , & Steger, G. (2013). The hypoxia‐inducible transcription factor ZNF395 is controlled by IĸB kinase‐signaling and activates genes involved in the innate immune response and cancer. PLoS ONE, 8, e74911 10.1371/journal.pone.0074911 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawai, H. , Wiederschain, D. , Kitao, H. , Stuart, J. , Tsai, K. K. C. , & Yuan, Z. M. (2003). DNA damage‐induced MDMX degradation is mediated by MDM2. Journal of Biological Chemistry, 278, 45946–45953. 10.1074/jbc.M308295200 [DOI] [PubMed] [Google Scholar]
- Le, Y. , Jia, P. , Jin, Y. , Liu, W. , Jia, K. , & Yi, M. (2017). The antiviral role of heat shock protein 27 against red spotted grouper nervous necrosis virus infection in sea perch. Fish & Shellfish Immunology, 70, 185–194. 10.1016/j.fsi.2017.08.032 [DOI] [PubMed] [Google Scholar]
- Le, Y. , Li, Y. , Jin, Y. , Jia, P. , Jia, K. , & Yi, M. (2017). Establishment and characterization of a brain cell line from sea perch, Lateolabrax japonicus . Vitro Cellular & Developmental Biology – Animal, 53, 834–840. 10.1007/s11626-017-0185-7 [DOI] [PubMed] [Google Scholar]
- Liu, J. , Zhang, C. , Hu, W. , & Feng, Z. (2015). Tumor suppressor p53 and its mutants in cancer metabolism. Cancer Letters, 356, 197–203. 10.1016/j.canlet.2013.12.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, P. , Wang, L. , Kwang, J. , Yue, G. H. , & Wong, S.‐M. (2016). Transcriptome analysis of genes responding to NNV infection in Asian seabass epithelial cells. Fish & Shellfish Immunology, 54, 342–352. 10.1016/j.fsi.2016.04.029 [DOI] [PubMed] [Google Scholar]
- Liu, W. , Xiang, Y. , Zhang, W. , Jia, P. , Yi, M. , & Jia, K. (2018). Expression pattern, antiviral role and regulation analysis of interferon‐stimulated gene 15 in black seabream, Acanthopagrus schlegelii . Fish & Shellfish Immunology, 82, 60–67. 10.1016/j.fsi.2018.07.041 [DOI] [PubMed] [Google Scholar]
- Lu, K. , Ji, Z. , Rahimnejad, S. , Zhang, C. , Wang, L. , & Song, K. (2017). De novo assembly and characterization of seabass Lateolabrax japonicus transcriptome and expression of hepatic genes following different dietary phosphorus/calcium levels. Comparative Biochemistry and Physiology Part D: Genomics and Proteomics, 24, 51–59. 10.1016/j.cbd.2017.08.001 [DOI] [PubMed] [Google Scholar]
- Lu, L. , Li, S. , Lu, X. , LaPatra, S. E. , Zhang, N. , Zhang, X. , … Zhang, Y. (2016). Spring viremia of carp virus N protein suppresses fish IFNφ1 production by targeting the mitochondrial antiviral signaling protein. Journal of Immunology, 196, 3744–3753. 10.4049/jimmunol.1502038 [DOI] [PubMed] [Google Scholar]
- Moll, U. M. , & Zaika, A. (2001). Nuclear and mitochondrial apoptotic pathways of p53. FEBS Letters, 493, 65–69. 10.1016/S0014-5793(01)02284-0 [DOI] [PubMed] [Google Scholar]
- Moreno, P. , Alvarez‐Torres, D. , Garcia‐Rosado, E. , Borrego, J. J. , & Alonso, M. C. (2018). Differential antiviral activity of European sea bass interferon‐stimulated 15 protein (ISG15) against RGNNV and SJNNV betanodaviruses. Fish & Shellfish Immunology, 83, 148–157. 10.1016/j.fsi.2018.09.022 [DOI] [PubMed] [Google Scholar]
- Moreno, P. , Garcia‐Rosado, E. , Borrego, J. J. , & Alonso, M. C. (2016). Genetic characterization and transcription analyses of the European sea bass (Dicentrarchus labrax) isg15 gene. Fish & Shellfish Immunology, 55, 642–646. 10.1016/j.fsi.2016.06.043 [DOI] [PubMed] [Google Scholar]
- Muñoz‐Fontela, C. , Macip, S. , Martínez‐Sobrido, L. , Brown, L. , Ashour, J. , García‐Sastre, A. , … Aaronson, S. (2008). Transcriptional role of p53 in interferon‐mediated antiviral immunity. Journal of Experimental Medicine, 205, 1929–1938. 10.1084/jem.20080383 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mun̄oz‐Fontela, C. , Angel Garcia, M. , Garcia‐Cao, I. , Collado, M. , Arroyo, J. , Esteban, M. , … Serrano, M. , (2005). Resistance to viral infection of super p53 mice. Oncogene, 24, 3059–3062. 10.1038/sj.onc.1208477 [DOI] [PubMed] [Google Scholar]
- Nakai, T. , Mori, K. , Sugaya, T. , Nishioka, T. , Mushiake, K. , & Yamashita, H. (2009). Current knowledge on viral nervous necrosis (VNN) and its causative betanodaviruses. Israeli journal of Aquaculture – Bamidgeh, 61(3), 198–207. [Google Scholar]
- Ning, P. , Hu, C. , Li, X. , Zhou, Y. , Hu, A. , Zhang, Y. , … Zhang, Y. (2017). Classical swine fever virus shimen infection increases p53 signaling to promote cell cycle arrest in porcine alveolar macrophages. Oncotarget, 8, 55938–55949. 10.18632/oncotarget.18997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poisa‐Beiro, L. , Dios, S. , Montes, A. , Aranguren, R. , Figueras, A. , & Novoa, B. (2008). Nodavirus increases the expression of Mx and inflammatory cytokines in fish brain. Molecular Immunology, 45, 218–225. 10.1016/j.molimm.2007.04.016 [DOI] [PubMed] [Google Scholar]
- Raja, R. , Ronsard, L. , Lata, S. , Trivedi, S. , & Banerjea, A. C. (2017). HIV‐1 Tat potently stabilises Mdm2 and enhances viral replication. Biochemical Journal, 474, 2449–2464. 10.1042/BCJ20160825 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivas, C. , Aaronson, S. A. , & Munoz‐Fontela, C. (2010). Dual role of p53 in innate antiviral immunity. Viruses, 2, 298–313. 10.3390/v2010298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Royds, J. A. , Hibma, M. , Dix, B. R. , Hananeia, L. , Russell, I. A. , Wiles, A. , … Braothwaite, A. W. (2006). p53 promotes adenoviral replication and increases late viral gene expression. Oncogene, 25, 1509–1520. 10.1038/sj.onc.1209185 [DOI] [PubMed] [Google Scholar]
- Russell, W. C. , Graham, F. L. , Smiley, J. , & Nairn, R. (1977). Characteristics of a human cell line transformed by DNA from human adenovirus type 5. Journal of General Virology, 36, 59–72. 10.1099/0022-1317-36-1-59 [DOI] [PubMed] [Google Scholar]
- Schroeder, L. , Herwartz, C. , Jordanovski, D. , & Steger, G. (2017). ZNF395 is an activator of a subset of IFN‐stimulated genes. Mediators of Inflammation, 2017, 1–15. 10.1155/2017/1248201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turpin, E. , Luke, K. , Jones, J. , Tumpey, T. , Konan, K. , & Schultz‐Cherry, S. (2005). Influenza virus infection increases p53 activity: Role of p53 in cell death and viral replication. Journal of Virology, 79, 8802–8811. 10.1128/JVI.79.14.8802-8811.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, Y. D. , Rajanbabu, V. , & Chen, J. Y. (2015). Transcriptome analysis of medaka following epinecidin‐1 and TH1‐5 treatment of NNV infection. Fish & Shellfish Immunology, 42, 121–131. 10.1016/j.fsi.2014.10.040 [DOI] [PubMed] [Google Scholar]
- Xu, D. , Du, Q. , Han, C. , Wang, Z. , Zhang, X. , Wang, T. , … Tong, D. (2016). p53 signaling modulation of cell cycle arrest and viral replication in porcine circovirus type 2 infection cells. Veterinary Research, 47, 120 10.1186/s13567-016-0403-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan, L. , Chen, Z. , Song, S. , Wang, S. , Tian, C. , Xing, G. , … Zhang, L. (2015). p53 degradation by a coronavirus papain‐like protease suppresses type I interferon signaling. Journal of Biological Chemistry, 290, 3172–3182. 10.1074/jbc.M114.619890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan, W. , & Krug, R. M. (2001). Influenza B virus NS1 protein inhibits conjugation of the interferon (IFN)‐induced ubiquitin‐like ISG15 protein. EMBO Journal, 20, 362–371. 10.1093/emboj/20.3.362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng, D. , Chen, X. , Xie, D. , Zhao, Y. , Yang, C. , Li, Y. , … Chen, X. (2013). Transcriptome analysis of pacific white shrimp (Litopenaeus vannamei) hepatopancreas in response to taura syndrome virus (TSV) experimental infection. PLoS ONE, 8, e57515 10.1371/journal.pone.0057515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, H. , Ye, H. , Liu, S. , Deng, C. , Li, X. , Shi, P. , … Zhang, B. (2017). West nile virus NS1 antagonizes interferon beta production by targeting RIG‐I and MDA5. Journal of Virology, 91, e02396–16. 10.1128/JVI.02396-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, W. , Jia, P. , Liu, W. , Li, Y. , Yi, M. , & Jia, K. (2018). Functional characterization of tumor necrosis factor receptor‐associated factor 3 of sea perch (Lateolabrax japonicas) in innate immune. Fish & Shellfish Immunology, 75, 1–7. 10.1016/j.fsi.2018.01.039 [DOI] [PubMed] [Google Scholar]
- Zhang, Y. , & Gui, J. (2012). Molecular regulation of interferon antiviral response in fish. Developmental & Comparative Immunology, 38, 193–202. 10.1016/j.dci.2012.06.003 [DOI] [PubMed] [Google Scholar]
- Zhao, C. , Fu, M. , Wang, C. , Jiao, Z. , & Qiu, L. (2016). RNA‐Seq analysis of immune‐relevant genes in Lateolabrax japonicus during Vibrio anguillarum infection. Fish & Shellfish Immunology, 52, 57–64. 10.1016/j.fsi.2016.02.032 [DOI] [PubMed] [Google Scholar]
- Zhong, S. , Mao, Y. , Wang, J. , Liu, M. , Zhang, M. , & Su, Y. (2017). Transcriptome analysis of Kuruma shrimp (Marsupenaeus japonicus) hepatopancreas in response to white spot syndrome virus (WSSV) under experimental infection. Fish & Shellfish Immunology, 70, 710–719. 10.1016/j.fsi.2017.09.054 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials