

Abstract
Multiple sclerosis (MS) is an immune‐mediated demyelinating disorder of unknown etiology. Both genetic‐susceptibility and environment exposures, including vitamin D deficiency, Epstein‐Barr viral and Herpesvirus (HHV‐6) infections are strongly implicated in the activation of T cells and MS‐pathogenesis. Despite precise knowledge of how these factors could be operating alone or in combination to facilitate and aggravate the disease progression, it is clear that prolonged induction of inflammatory molecules and recruitment of other immune cells by the activated T cells results in demyelination and axonal damage. It is imperative to understand the risk factors associated with MS progression and how these factors contribute to disease pathology. Understanding of the underlying mechanisms of what factors triggers activation of T cells to attack myelin antigen are important to strategize therapeutics and therapies against MS. Current review provides a detailed literature to understand the role of both pathogenic and non‐pathogenic factors on the impact of MS.
Keywords: autoimmune, demyelinated neurons, encephalitis, neurodegeneration
Significance
Despite major research efforts in the past few decades, the extent to which environmental factors (including external pathogens) and genetic susceptibility contribute to the pathogenesis of multiple sclerosis (MS) is not clearly identified. This review examine the viral and bacterial as well as non‐pathogentic environmental associations to MS progression and how these factors contribute to disease pathology.
1. INTRODUCTION
Multiple sclerosis (MS) is a multifaceted inflammatory disease of the central nervous system (CNS). It involves a gradual destruction of myelin due to an autoimmune response generated against self‐antigens in a genetically vulnerable patient (Donati & Jacobson, 2002). The attack against self‐antigen leads to a gradual demise of neurons, resulting in severe neurological deficits (Goldenberg, 2012). The myelin sheath is an insulating cover of axons maintained by oligodendrocytes (Compston & Coles, 2008), and facilitates electrical action potentials between neurons. Loss of oligodendrocytes and degeneration of the myelin sheath leads to the formation of plaques or lesions, commonly located in the optic nerve, brain stem, basal ganglia, and spinal cord white matter (Popescu & Lucchinetti, 2012). While the repair process of remyelination is possible early in disease, oligodendrocytes are incapable of complete repair due to subsequent attacks to the sheath, contributing to the plaque formation (Chari, 2007; Kuhlmann et al., 2008). Damage or death of neurons due to lesion accumulation causes astrogliosis adding to lesion creation (Compston & Coles, 2008) diminishing the communication between neurons and leading to a wide range of disorders, largely dependent on the location of the lesions, in the brain (Virtanen & Jacobson, 2012). Symptoms include loss of sensitivity, changes in sensation (tingling, or pins and needles), muscle weakness and spasms, blurred vision, ataxia, and speech disorders (Compston & Coles, 2008). MS affects people of almost all ages, although higher incidences are detected in younger adults as they are more susceptible to environmental factors. MS is more common in women, suggesting that hormones may also play a significant role in determining susceptibility (Milo & Kahana, 2010; Nylander & Hafler, 2012). Prevalence of MS is two to three times higher in women than in men (Figure 1) (Ahlgren, Oden, & Lycke, 2012; Compston & Coles, 2002; Orton et al., 2010; Wallin et al., 2012).
Figure 1.
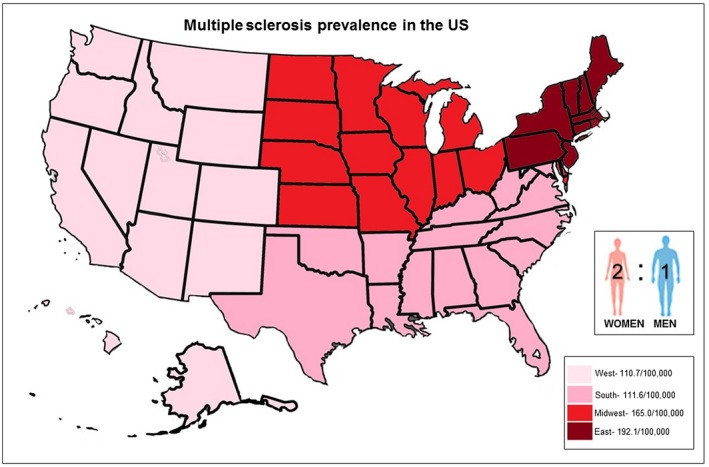
Multiple sclerosis (MS) epidemiology in the United States. Prevalence of MS on the map is illustrated by different color. Areas with low prevalence of MS are indicated in light pink, medium prevalence of MS (pink), moderate prevalence of MS (red), and high prevalence of MS (burgundy). Inset shows the ratio between genders and that women are twice as likely than men to develop MS
2. SUBTYPES OF MS
During the initial stages of the disease, patients primarily experience a neurological complication referred to as Clinically Isolated Syndrome (CIS). This is the first demyelinating event, lasting for approximately 24 hr and presenting as monofocal or multifocal lesions in the CNS (Marcus & Waubant, 2013). Possible symptoms of CIS include optic neuritis, a brain stem/cerebellar syndrome, a spinal cord syndrome, or hemispheric dysfunction (Miller, Barkhof, Montalban, Thompson, & Filippi, 2005). Later, the disease becomes more convoluted and characterized as progressing into four distinct variants. These subtypes are important for patient prognosis and treatment considerations. The most common type of MS is called Relapsing Remitting MS (RRMS) and occurs in approximately 87% of MS cases (Ransohoff, Hafler, & Lucchinetti, 2015). RRMS patients show neurological symptoms and have a short and temporary relapse phase where not only existing symptoms are aggravated, but new symptoms may also arise (Weiner, 2008). The inflammatory attacks during this period are unpredictable and are associated with visual and memory impairment, tingling and numbness and fatigue. This relapse stage is followed by a stage of remission (complete/incomplete) in which the patient may recover (fully or partially), resulting in asymptomatic disease for several months or years (Ghasemi, Razavi, & Nikzad, 2017). Another variant is Secondary Progressive MS (SPMS) occurring in ~65% of patients with RRMS, gradually transitioning into SPMS as neurological complications steadily advance over time. Patients may have intermittent relapses, but no remission is observed (Correale, Gaitan, Ysrraelit, & Fiol, 2017; Ghasemi et al., 2017). A more uncommon variant, Primary Progressive MS (PPMS), occurs in approximately 10%–15% of MS cases (Ransohoff et al., 2015). PPMS is distinguished by a gradual progression from onset with deteriorating symptoms and no occurring relapses or remissions. These patients are primarily affected at the nerves of the spinal cord and present with fewer brain lesions compared to other subtypes. Patients with PPMS show ataxia with problems in walking, weakness, stiffness, and trouble with balance (Ghasemi et al., 2017). Lastly, Progressive‐Relapsing MS, another rare subtype (occurring in ~5% of MS cases) is marked by complications starting from the initial stages of the disease, and progresses with critical relapsing cases without any remissions, mostly without recovery resulting in exacerbating neurological injuries. Symptoms in these patients include eye pain, double vision and depression, along with sexual, intestinal, and urinary system dysfunctions (Ghasemi et al., 2017).
Although MS is a disease common to adults, onset of the disease in children below 18 years of age is not uncommon, accounting for about 10% of total MS cases and is termed pediatric MS (PMS) (Ferreira, Machado, Dantas, Moreira, & Souza, 2008; Jancic et al., 2016; Schreiner, 2018). PMS is pathophysiologically distinct in terms of brain biopsy, cellular profiling in CSF, and the immune molecules present when compared with adult MS patients (Schreiner, 2018). The psychological consequences of childhood MS include depression, fatigue, anxiety, panic disorder, bipolar disorders, and cognitive impairment (Amato et al., 2010). Though the disability rate is lower than that of adult‐onset MS, children diagnosed with PMS pose a risk of developing neurological deficits during the active stage of life (Renoux et al., 2007).
3. IMMUNOPATHOLOGY
T‐cells mature may express, on their cell surface, either a CD4 glycoprotein, found in T‐helper (Th) cells, or they may express a CD8 glycoprotein, present in cytotoxic T‐cells (Golubovskaya & Wu, 2016). Exposure to specific cytokines, such as IL‐1, IL‐2, IL‐4, IL‐6, TNF, and INF‐γ result in the differentiation of naïve CD4+ cells into a variety of subsets: Th1, Th2, Th9, Th17, Th22, Tfh, and Treg. Each subset possesses a unique cytokine profile, releasing cytokines specific to that subset, which may provide protective or anti‐/pro‐inflammatory properties (Golubovskaya & Wu, 2016). Th1 provides protection against intracellular pathogens, Th2 defending against parasites, and Th17 focuses on defending the host against extracellular pathogens. For example: exposure to IL‐1 results in the differentiation of a naïve CD4+ cell to a Th17 subset which function as mediators of inflammation and produce IL‐17, protecting the host against fungi and bacteria (Golubovskaya & Wu, 2016; Raphael, Nalawade, Eagar, & Forsthuber, 2015). Th17 cells are known to mediate autoimmunity and play key roles in augmenting inflammation (Raphael et al., 2015).
Antigen‐presenting cell (APC) activated CD8+ T cells proliferate and produce effector cells which induce the apoptosis of target cells, holding a key role within the adaptive immune response. However, there are various subsets of these CD8+ effector cells: A naïve (CD45RA+/CCR7+) subset lacking effector functions, (a) an antigen‐experienced and differentiated (CD45RA+/CCR7−) subset providing effector functions (TEff), (b) an antigen‐experienced (CD45RA−/CCR7+) subset lacking effector functions, known as central memory cells (TCM), and (c) an effector function bearing (CD45RA−/CCR7−) memory cell subset, defined as effector memory cells (TEM) (Saxena, Martin‐Blondel, Mars, & Liblau, 2011). Proliferation and antigen‐driven activation of these effector C8+ cells is mediated by Th1 produced IL‐2, and may present as autoreactive T cells as a result of self‐antigen presentation by APCs, potentially resulting in T‐cell induced axonal damage (Hwang, Hong, & Glimcher, 2005; Rollings, Sinclair, Brady, Cantrell, & Ross, 2018; Saxena et al., 2011). Expansion of CD8+ cells, detection of identical antigen‐specificity in a population of T cell clones, and presence of predominantly IL‐17 producing CD8+ cells, all within active lesions in MS, support the concept of CD8+ cell mediated autoimmunity, in MS (Saxena et al., 2011).
An atypical response by immune cells against the myelin sheath is the common pathological factor between all MS subtypes. This response is facilitated by discriminatory activation of CD4+ helper and CD8+ killer T cells, accompanied by little to no activation of regulatory T cells (Gandhi, Laroni, & Weiner, 2010). As a result, inflammation is an alternate characteristic symptom of MS. T lymphocytes and activated macrophages infiltrate the brain and with the aid of activated microglia, induce immunological activation, leading to progressive myelin destruction (Kasper & Shoemaker, 2010). Disruption of the blood–brain barrier (BBB) also allows immune cells to enter the brain, where T lymphocytes attack the myelin sheath (Compston & Coles, 2002). This triggers a subsequent inflammatory cascade, leading to the secretion of cytokines, including interferon (IFN)‐γ, tumor necrosis factor (TNF)‐α, and interleukin (IL)‐17, and the production of antibodies by B lymphocytes which can secrete their own cytokines, TNFα and Transforming Growth Factor (TGF)‐β (Duddy et al., 2007). These soluble factors cause further recruitment of immune cells, perpetuating inflammation and damage (Nylander & Hafler, 2012). In addition, pro‐inflammatory Type 1 T helper (Th1) and Th17 cells display direct cytotoxic effects on oligodendrocyte precursor cells and obstruct remyelination (Baxi et al., 2015; Moore et al., 2015).
4. GENETIC FACTORS
Predisposition to genetic changes and its effect on MS susceptibility was derived from the characteristics of familial aggregation and the amount of genetic information they share (Gejman, Sanders, & Duan, 2010; Oksenberg, Baranzini, Sawcer, & Hauser, 2008). Although MS is not a hereditary disease, predisposition to genetic variations increases the risk. Studies have shown that the risk rate in monozygotic‐twins that have 100% genetic familiarity is approximately 25% (Ghasemi et al., 2017). These risks are reduced on decreased genetic familiarity, with 5% in siblings, 2% in parents, 2% in children, and 1% amongst distant relatives (Hassan‐Smith & Douglas, 2011).
Discovered in mice in 1936, the major histocompatibility complex (MHC) and the human equivalent human leukocyte antigen (HLA) are extensively studied for both gene and allelic variation in MS (Matzaraki, Kumar, Wijmenga, & Zhernakova, 2017). HLA changes are responsible for approximately 20%–60% of genetic predisposition (International Multiple Sclerosis Genetics et al., 2011), and around 110 polymorphisms found in discrete loci outside the MHC gene are associated with the susceptibility to MS (Sawcer, Franklin, & Ban, 2014). According to a Genome‐wide association study, susceptibility to MS has been linked mainly to the HLA‐DRB1 locus class II region of the MHC, with the HLA‐DR15 (DRB1*1501‐DQA1*0102‐DQB1*0602‐DRB5*0101) haplotypes dominating MS risk in Caucasians and HLA‐DRB1*15:01 associated with MS risk in African Americans and northern Han Chinese populations (Compston & Coles, 2008). Additional alleles within the MHC locus have been associated to MS including the HLA‐DRB1*03:01, HLA‐DRB1*13:03, HLA‐DPB1*03:01 and HLA‐A*02:01, while HLA‐C554 and HLA‐DRB1*11 are believed to be protective against the disease (International Multiple Sclerosis Genetics et al., 2011). Polymorphisms in non‐HLA genes associated to MS include the interleukin 2 (IL2) receptor alpha (IL2RA) and interleukin 7 (IL7) receptor alpha (IL7RA) loci expressed on regulatory T cells, important for proliferation and survival of T‐ and B cells, and increase the vulnerability of developing MS (Giovannoni & Ebers, 2007). Single‐Nucleotide Polymorphisms (SNPs) in IL‐7R markers identified in MS patients in Sweden suggest that SNPs can lead to dysregulation in the activity of both T‐ and B‐cells (Lundmark et al., 2007). By analyzing MS patients possessing SNPs in the Serine‐Threonine Kinase 11 gene (encoding liver kinase B1—LKB1), Boullerne et. al. suggests it as a risk factor for MS (Boullerne et al., 2015). LKB1 has effects on T‐cell activation, Th1/Th17 cytokine production, and modulate adenosine monophosphate kinase (AMPK) activation. Dysregulation of LKB1 function is implicated toward MS pathogenesis (Germain et al., 2013). It has been demonstrated using GSAW that multiple non‐MHC loci—CLEC16A, IL2RA and IL7R are susceptibility loci for MS (De Jager et al., 2009). Other non‐MHC critical factors with links to MS include TNFRSF1A (Caminero, Comabella, & Montalban, 2011; Hoffjan et al., 2015), which contains two independent susceptible alleles responsible for dysregulated expression of TNF; IRF8 has a susceptible allele that encodes transcription factors, and CD6 is involved in T‐cell co‐stimulation and differentiation (De Jager et al., 2009).
5. ENVIRONMENTAL FACTORS
High rates of discordancy in monozygotic twin studies led to the conclusion that genetic and epigenetic factors are not the only factors (Giovannoni & Ebers, 2007), thus emphasizing the importance of environmental factors in developing MS. Environmental factors such as smoking, vitamin deficiency, diet, and UV radiation exposure act as modifiable risk factors (Sawcer et al., 2014). The expression of the Vitamin D receptor on most immune cells including APC is considered a driving force in MS progression (Disanto, Morahan, Barnett, Giovannoni, & Ramagopalan, 2012). Presence of 1,25‐dihydroxyvitamin D3 (1α,25‐(OH)2D3) modulates the expression of MHC II‐complexed antigens on the surface of immune cells, inhibits the production of Type 2 helper T cell cytokines, and affects T‐and B cell proliferation and antibody secretion (Aranow, 2011; van Etten & Mathieu, 2005). The International Multiple Sclerosis Genetics Consortium (IMSGC) and Welcome Trust Case Control Consortium 2 (WTCCC2) reported that the genes encoding the Cytochrome P450 Family 27 Subfamily B Member 1 (CYP27B1) enzyme for vitamin D production and Cytochrome P450 Family 24 Subfamily A Member 1 (CYP24A1) enzyme which degrades 1,25‐dihydroxyvitamin D contribute to MS progression by lowering the levels of active vitamin D. Moreover, RNA expression levels of the MS susceptibility gene, HLA‐DRB1*15:01 are controlled by vitamin D levels (International Multiple Sclerosis Genetics et al., 2011).
The risk of developing SPMS is 3‐fold higher in smokers than in non‐smokers who had relapsing–remitting clinical onset of the disease (Healy et al., 2009; Wingerchuk, 2012). Furthermore, tobacco affects the integrity of the BBB and triggers inflammatory responses that, lead to loss of membrane integrity (Hawkes, 2007; Hernan et al., 2005; Riise, Nortvedt, & Ascherio, 2003). Marabita et. al. showed that smoking affects DNA methylation in MS patients and gene expression of Aryl‐Hydrocarbon Receptor Repressor (AHRR) is higher in smokers and contained hypomethylation at several CpG sites (Marabita et al., 2017). Tobacco smoke induces the production of free radicals, such as nitric oxide (NO) which exacerbates the clinical progression of MS by causing inflammatory lesions and axonal degeneration (Hernan et al., 2005).
Risk for MS development intensifies due to complex interplay of different pathogenic environmental factors including viral and bacterial infections(Virtanen & Jacobson, 2012). Infection with measles, Herpesvirus (HHV‐6) and Epstein Barr virus (EBV) at a young age increase the chances of developing MS (Compston & Coles, 2008; Giovannoni et al., 2006). Virus‐derived peptide sequences such as the Epstein–Barr nuclear antigen 1 (EBNA1) (Owens, Gilden, Burgoon, Yu, & Bennett, 2011)and BRRF2 protein contribute to MS pathology partly by activation of memory B cells, increased frequency of oligoclonal IgG specificities, and by interacting with HLA through DR2b (DRB1*1501) and DR2a (DRB5*0101) (Abdelrahman, Selim, Hashish, & Sultan, 2014; Pender & Burrows, 2014). The production of EBV‐specific T cells or antibodies cross‐react with autoantigens in the CNS, and attack myelin sheath. Latent persistent EBV antigens bind to autoreactive B cells that facilitate EBV replication, and augment EBV‐specific T‐ and B‐cell responses (Ascherio et al., 2001; Lunemann, Kamradt, Martin, & Munz, 2007). Other viruses associated with MS include varicella zoster virus (VZV) and human endogenous retroviruses (HERVs).
A “multi‐hit” model study illustrated how the gut microbiota creates a suitable proinflammatory condition for multiple insults (genetic disposition and environmental factors) to act together reinforcing MS disease (Cekanaviciute et al., 2017). Akkermansia muciniphila and Acinetobacter calcoaceticus induce proinflammatory responses, and are found in higher numbers within the gut whereas Parabacteroides distasonis, which stimulates the anti‐inflammatory response, is found in lower numbers in MS patients (Mirza & Mao‐Draayer, 2017). It is also possible that individuals without early life exposure to infectious pathogens could mount abnormal responses during infectious exposure in their adult life (Baranzini et al., 2010). It is evident that multiple environmental factors play important roles toward increased risks of MS; however, the pathogenic contributions are still being unearthed and will be the focus of the remainder of this review.
6. PATHOGENS IN THE CNS
Most pathogens associated with MS are also considered causative agents in the development of other neurological disorders. By defining the mechanisms behind MS‐like symptoms linked to pathogenic infections, the similarities that may lead to onset of disease become more readily apparent. HHV‐6 infection is associated with neurological disorders such as febrile seizure, epilepsy, and encephalitis (Yao et al., 2009), while EBV infection has been associated with CNS vasculitis (Kano et al., 2017). These two pathogens share common mechanisms that can contribute to the onset of MS symptoms. Both EBV and HHV‐6 infection can trigger myelin‐reactive antibodies specifically targeting myelin basic protein (MBP), an ubiquitous myelin membrane protein (Deber & Reynolds, 1991; Tait & Straus, 2008). Low levels of MBP phosphorylation in both human MS patients as well as in mice models are detected (DeBruin et al., 2005; DeBruin, Haines, Bienzle, & Harauz, 2006); however, the direct effects of this have not yet been characterized. In contrast, VZV utilizes an endosomal/autophagic recycling pathway similar to that used by myelin proteins and the iron‐binding glycoprotein transferrin (Winterstein, Trotter, & Kramer‐Albers, 2008). It is through this pathway that VZV trafficks glycoprotein complexes from the endoplasmic reticulum (ER) to the outer cell membrane to release new virus (Buckingham, Jarosinski, Jackson, Carpenter, & Grose, 2016). Given that low serum transferrin levels have been linked to several chronic diseases, including MS (Zeman et al., 2000), trafficking of VZV alongside the transferrin receptor would aid in viral assembly, ultimately leading to increased immune activation.
Apart from demyelination, there is also evidence of axonal or neuronal injury in MS that have gained some importance in the MS field. The “inside‐out” concept of induced demyelination suggests that pathogenic damage occurs at the unmyelinated axon, often in combination with enhanced oligodendrocyte apoptosis (Tsunoda & Fujinami, 2002). Several viral infections are accompanied by axonal damage like HIV (An et al., 1997), HSV (Martin, 1984), and Theiler's murine encephalomyelitis virus (TMEV) (Tsunoda & Fujinami, 2002), which has also been associated with MS. The pathogenesis of this axonal injury still requires much study. One concept suggests direct immune attack on the axon based upon the presence of antibodies to neurofilament and tubulin in some MS patients (Silber & Sharief, 1999). Separately, some viruses can be transported through the axons leading to axonal degeneration and dissemination of virus throughout the CNS (Kristensson, 1996). In TMEV infection, for example, axonal injury precedes demyelination which indicates in some cases demyelination may come secondary to axonal injury (Das Sarma, Kenyon, Hingley, & Shindler, 2009). Additionally, as has been discussed, there is also evidence on axonal injury in silent inactive plaques where few axons are degraded at a given point in time but will instead persist for a longer time in the CNS (Lassmann, 2003).
Viral infections causing encephalitis (Morfopoulou et al., 2016), are also considered as a risk factor in developing MS (Weiss, 1983). Human beta‐coronaviruses, such as HCoV‐OC43 primarily linked with mid‐upper respiratory tract infections with neuro‐invasive properties leading to encephalitis, are associated with MS (Morfopoulou et al., 2016). Additionally, John Cunningham virus (JCV) induces progressive multifocal leukoencephalopathy (PML) in active RRMS patients undergoing immunosuppressant treatment with natalizumab (Schwab et al., 2016). The immunosuppression causes reactivation of JCV, which is normally kept dormant by the human immune system, contributing to the progression of PML (Schwab et al., 2016).
6.1. Viruses
Although no definitive association between viral infections and MS have been reported so far, compelling evidence for this hypothesis exists, such as the presence of viral DNA, RNA, or proteins, and immune response elements in the body fluids or tissue of patients (Donati & Jacobson, 2002). Attempts to establish a definitive association between MS and viral infections has been restricted by many factors, including asynchrony between viral infection and demyelination, genetic, and immunological susceptibility of the individuals, and the involvement of polymicrobials as the causative agent of MS. This is further complicated by differences in the pathogenic mechanisms of different individuals with varying genetic backgrounds, resulting in different forms of the disorder (Donati & Jacobson, 2002). Even though different animal models of MS, such as the experimental autoimmune encephalitis (EAE) model and TMEV model aided in the understanding of possible underlying mechanisms including molecular mimicry and bystander activation, animal models for the most commonly associated viruses HHV‐6 and EBV have yet to be developed (Virtanen & Jacobson, 2012).
Studies on viral induced demyelination show cellular mechanisms of myelin destruction through persistence, latency, reactivation, and tissue damage by viruses. Virus‐induced demyelination disorders have different underlying mechanisms related to direct toxic effects or immune activation by virus, molecular mimicry, bystander activation, and epitope spreading (Fujinami, von Herrath, Christen, & Whitton, 2006). Examples of direct toxic effects from an active infection is JCV which directly causes demyelination after infecting oligodendrocytes, and canine distemper virus which infects astrocytes in white matter leading to demyelination (Chalkias et al., 2014). Another mechanism underlying demyelination is molecular mimicry, where pathogenic proteins possess homologous amino acid sequences with self‐proteins subsequently causing viral induced autoimmunity (Fujinami & Oldstone, 1985). Below, various viruses and their associations to the etiology or pathogenesis of MS will be discussed.
6.1.1. Measles virus
Measles is caused by the highly contagious paramyxovirus and is transmitted via respiratory route (Laksono, de Vries, McQuaid, Duprex, & de Swart, 2016; Ludlow, McQuaid, Milner, de Swart, & Duprex, 2015). The clinical manifestation is relatively mild presenting as skin rashes, though, in rare cases, neurological complications can also occur. Acute disseminated encephalitis (Lundmark et al., 2007), measles inclusion body encephalitis, and subacute sclerosing panencephalitis are all severe cases of measles which have been associated with CNS disorders (Laksono et al., 2016). In MS patients, the most frequent anti‐viral antibodies produced intra‐thecally tend to be against measles, rubella and VZV, referred to as MRZ reaction phenomenon. Although this is not restricted to a direct association with these three viruses, it may be a response against different viruses such as HSV or EBV. Evidence supporting the possible direct interaction or causal association of measles with MS is not well documented; moreover, unchanged prevalence of MS after vaccination against measles, shed further doubt on the possible association between the virus and disease (Ahlgren, Odén, Torén, & Andersen, 2009; Donati & Jacobson, 2002; Jacobson, Flerlage, & McFarland, 1985; Norrby, Link, & Olsson, 1974).
6.1.2. Human Herpesvirus 6
Human Herpesvirus 6 (HHV‐6) is a DNA virus belonging to β‐Herpes virus family and is widely sero‐prevalent in adults. The two variants HHV‐6A and HHV‐6B have about 90% sequence homology, and are transmitted via saliva and close contact with infected parents during early years of life. HHV‐6B is the causative agent of exanthema subitum (or roseola), a benign febrile illness with skin rashes while HHV‐6A has not been definitively associated with any disease (Reynaud, Horvat, & Horvat, 2013). HHV‐6 is able to enter cells through the CD46 receptor, with viral antigen able to be detected in oligodendrocytes and rarely in astrocytes. However, HHV‐6 has been detected in the scar‐like plaques on the myelin sheath of MS patients (Reynaud et al., 2013). Accordingly, IgM antibody against the P41/38 antigen and DNA of HHV‐6A was reported in serum from RRMS patients. HHV‐6A persists and may reactivate CNS immune cells, leading to higher lymphoproliferative response against HHV6A which has been detected in MS patients. Mechanistically, HHV‐6 may be linked to pathogenesis of MS through a proline‐rich integral membrane protein known as U24 (Sullivan & Coscoy, 2010). The proline rich region of U24 possesses sequence identity with MBP at a key phosphorylation site which, if molecular mimicry were to occur, the mitogen‐associated kinase pathway and the glycogen synthase kinase may be recruited and activated, resulting in lower phosphorylation of MBP. The potential of the proline‐rich region of U24 acting as a molecular mimic may mediate cross‐reactivity of T‐cells and lead to targeting of MBP in MS patients.
6.1.3. Epstein barr virus
EBV causes infectious mononucleosis and remains latent in B cells (Thorley‐Lawson, 2001) for approximately 30 years after infection (Nielsen et al., 2007). Later onset of symptomatic EBV infection is extremely risky (Ascherio & Munger, 2007). CD4 and CD8 T cells are the primary line of defense against EBV‐infection. The association of EBV with MS is supported by numerous case reports where EBV infection was shown to cause axonal demyelination, and detectable EBV antibodies were reported in CSF and meningeal lymphoid follicles (Salvetti, Giovannoni, & Aloisi, 2009; Thacker, Mirzaei, & Ascherio, 2006; Tselis, 2012). The major EBV antigen in MS cases is EBNA1, with antibodies produced specifically in the CSF compartment forming oligoclonal bands, a characteristic of viral infection in MS. Epitope mapping techniques employed for testing the specificity of oligoclonal IgG showed that MS‐specific epitopes are derived from EBNA1 and BRRF2 (EBV lytic gene product) proteins, providing evidence of intrathecal production of EBNA1‐specific IgG in MS (Cepok et al., 2005). Viral capsid antigen (VCA) and the early antigens (EA) are also produced against EBV. Serum antibodies to VCA show more variation and higher seroprevalence than antibodies to EBNA‐1. EBV transforms B cells controlled by host immunological control leading to EBV‐associated malignancies. During the lytic stage, C‐terminus of EBNA1 binds to the episome (circular DNA of EBV) and transmits virus into progenies. Another link between EBV infection and MS corresponds to molecular mimicry for immunodominant myelin resultant motifs on T cells (Libbey & Fujinami, 2010). Peptides derived from EBV have sequences which are stimulatory for MBP derived CD4 T‐cell clones. A cross‐reactive T‐cell clone detects the MBP peptide similar to the HLA‐DR2b and EBV BSLF1 peptide similar to the HLA‐DR2a. T cells specific to EBNA1 will provide immunity against EBV, but in MS patients, an EBNA1‐specific response of T cells is distorted; yielding exacerbated T cell responses throughout infection. Molecular mimicry amongst viral antigens and autoantigens support autoreactive T cells, such as the accumulation of EBNA1‐targeting T cells leading to autoimmunity associated with infection. EBV genes in turn excite autoimmune B cells which are also cross‐reactive, serving as evidence that EBV has involvement in MS pathogenesis (Compston & Coles, 2008) (Figure 2).
Figure 2.
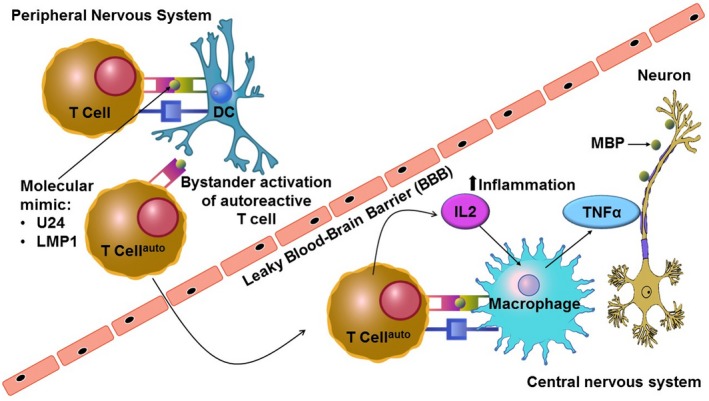
Activation of cross‐reactive T cells by pathogen‐derived peptides and self‐epitopes via molecular mimicry or by stander activation. Bystander activation involves a wide range of events that occur after viral infection such as production of cytokines and chemokines, expression of host otherwise inactive genes, and the unveiling of blocked self‐antigens (Donati & Jacobson, 2002). The process of epitope spreading implies the introduction of additional epitopes following an initial inflammatory response to one epitope, such as myelin basic protein (Lehmann, Forsthuber, Miller, & Sercarz, 1992). The mechanism can be explained by self‐sustained breakdown of myelin by continuous exposure of epitope during inflammatory reactions. The phenomenon of epitope spreading is well studied in the experimental autoimmune encephalitis and TME models for demyelination induced by viruses. B‐cells localized to the central nervous system present infectious agents on their surface and hence the infected epitope spreads to T cells and autoimmunity is promoted
6.1.4. Herpes zoster virus
VZV commonly causes encephalitis, meningitis, and myelitis. Immune responses against VZV, measles, and rubella have been found in patients with CNS vasculitis (Graef, Henze, & Reiber, 1994) and in the CSF of MS patients. The proposed pathway of VZV on CNS infection involve autophagy and retrograde trafficking of the virus primarily from vesicles on the trigeminal ganglion to cerebral arteries. Transport of the VZV within T‐cells and subsequent fusion with neurons has been postulated as a possible infectious mechanism (Horien & Grose, 2012). Despite this, the risk of developing MS upon VZV infection is less when compared to infection with EBV or HHV‐6 (Virtanen & Jacobson, 2012).
6.1.5. Retroviruses
There is growing evidence that retroviruses play a role in MS. It is suggested that two cofactors of the HERVs family, (a) MS‐associated retrovirus (MSRV) and (b) endogenous retroviral family W env(C7) member 1(ERVWE1), whose envelope proteins (Zappia E) show neuropathogenic features in vitro and in vivo, contribute to the inflammatory processes and MS pathogenesis (Sospedra & Martin, 2005). HERV‐W env gene encodes syncytin‐1 protein, which expresses in astrocytes, microglia and macrophages of MS patients. It promotes release of cytokines and reactive oxygen species in astrocytes leading to oligodendrocyte damage stimulating neuroinflammation.
6.2. Bacteria
6.2.1. Peptidoglycan
During the host response to infection, many bacterial components are released that can be recognized as targets for immune response. Heteropolymer peptidoglycan (PG) released by bacterial cells serves as a ligand to activate inflammatory immune response (Gupta, Wang, Vinson, & Dziarski, 1999; Weidemann et al., 1994). PG has been shown to be present in immunocompetent APC in the brain tissue of MS patients suggesting its role in MS pathogenesis (Schrijver et al., 2001). The presence of PG in the brain tissue of MS patients has been shown in the absence of active infection and thus indicates that peripheral PG may translocate with the APC to the CNS and serve to drive an inflammatory response in this location.
6.2.2. Chlamydia pneumoniae
Chlamydia pneumoniae causes common respiratory infections, with up to 50% of the population showing seropositivity in late adulthood (Grayston, Wang, Kuo, & Campbell, 1989). The first report of coincidental infection with C. pneumoniae in a patient with RPMS was reported in 1998 (Sriram, Mitchell, & Stratton, 1998). Upon broader examination it was found that over 97% of serum CSF in patients with MS tested positive for the C. pneumoniae momp gene. Additionally, C. pneumoniae was isolated from 64% of MS patients, showing coincidental infection (Sriram et al., 1999). C. pneumoniae has also been implicated in the pathogenesis of other neurodegenerative diseases such as Alzheimer's Disease and schizophrenia given that the pathogen can invade the blood stream, and when taken up by monocytes as carriers, can cross the BBB and access the CNS (Cahoon, 2009). It must be recognized that the direct relation of C. pneumoniae with MS is controversial. Likely, the bacteria acts as a co‐factor and contributes toward persistent and prolonged infection leading to MS disease progression(Fainardi, Castellazzi, Seraceni, Granieri, & Contini, 2008).
6.2.3. Mycobacterium avium subspecies paratuberculosis
Mycobacterium avium subspecies paratuberculosis (MAP), an infectious bacillus, has been previously associated with Crohn's disease (CD) in humans. To study possible associations of MAP infection with MS, the prevalence of MAP protein MAP2694 (a glycine and proline rich transmembrane protein conserved in the Mycobacteriaceae family), and the immune response to this protein in patients with MS compared to normal control subjects were studied. Patients with MS tested positive for MAP DNA (42%) when compared to healthy controls (12%). Additionally, MAP protein antibodies were detected more often in MS patients (32%) compared to healthy subjects (2%) (Cossu et al., 2011).
6.2.4. Helicobacter pylori
Helicobacter pylori has shown the histological isolation from 86.4% of MS patients versus 50% of healthy control subjects. (Gavalas et al., 2015). Gavalas et al. first reported a reverse correlation between H. pylori and MS with subsequent meta‐analysis, concluding that patients with H. pylori infection had lower rates of MS than those without infection. This suggests that H. pylori infection may provide some protection against MS development or progression (Jaruvongvanich, Sanguankeo, Jaruvongvanich, & Upala, 2016), mechanisms to which still remain unclear.
6.2.5. Staphylococcus aureus
Counter to observed associations between bacterial infection and MS, Staphylococcus aureus infection in a murine model of MS demonstrated a reduction in experimental autoimmune encephalomyelitis(Fainardi et al., 2008). This is counterintuitive to the model of bacterial inflammation driving MS development or progression as S. aureus produces systemic inflammation and stimulates the production of cytokines responsible for inflammatory response. This protective role was demonstrated by a reduction in demyelination and higher axon density in infected groups, as well as lower clinical scores, compared to uninfected groups. This protective role was mediated in part by an extracellular adhesion protein which was recovered in the blood of infected animals (Kumar et al., 2015).
7. ANIMAL MODELS AND TREATMENT OPTIONS
Animal models are critical for diseases like MS, which have a complex etiology and lack a complete understanding of its mechanisms. Due to its complexity, there is currently no single animal model that can reflect the whole spectrum of the disease. Over the years, several animal models have been designed to study MS and reviewed effectively (Procaccini, De Rosa, Pucino, Formisano, & Matarese, 2015), however, the most frequently used model for etiological studies of MS is the EAE model. In EAE, an immune reaction to CNS components is induced in susceptible animals with self‐antigens that are derived from myelin protein. This was first described in monkeys stimulated with rabbit antigen (Rivers, Sprunt, & Berry, 1933), but later Freund's adjuvant (Freund & McDermott, 1942) and pertussis toxin (Munoz, Bernard, & Mackay, 1984) were added to induce a humoral immune reaction similar to as seen in RRMS (Kabat, Wolf, & Bezer, 1947), and analogous to what is found in MS patients. Although studies have been done on guinea pigs (Freund, Stern, & Pisani, 1947) and monkeys (Kabat et al., 1947), the best models resulted from mice (Olitsky & Yager, 1949) and rats (Lipton & Freund, 1952). The type of antigen introduced to the animal determined its particular presentation of EAE. Immunization of SJL/J mice with an epitope of proteolipid protein induces RRMS (Tuohy, Lu, Sobel, Laursen, & Lees, 1989), while MS induced by myelin oligodendrocyte glycoprotein in C57BL6/J mice is chronic MS (Tompkins et al., 2002). Apart from EAE models, viral infection models which most effectively aid in the study of EBV related MS are TMEV infection models (Richards et al., 2011; Tsunoda, Iwasaki, Terunuma, Sako, & Ohara, 1996). Utilizing TMEV models also assist in studying new therapeutic methodologies targeted toward adhesion molecules and axonal degeneration (Tsunoda & Fujinami, 2010). Independent of introducing immunogens and viruses, certain chemical compounds have been utilized to cause an immune reaction leading to MS‐like symptoms, like cuprizone and lysolecithin, which cause oligodendrocyte cell death and have been primarily used to study demyelination and remyelination in MS (Jeffery & Blakemore, 1995; Matsushima & Morell, 2001).
7.1. Epigenetic therapy
Epigenetics, encompasses DNA methylation, histone modifications, and small RNA based mechanisms, may be a potential therapeutic avenue in MS treatment (Huynh & Casaccia, 2013). Due to observed epigenetic changes in MS, this therapy has emerged as a novel treatment alternative to MS. Histone deacetylase inhibitors like Trichostatin A, Vorinostat, and Valproic acid have been used as therapeutic drugs, and have been shown to reduce cell infiltration in the CNS and reduce overall inflammation through the suppression of dendritic cells and dendritic cell costimulatory molecules (Camelo et al., 2005; Ge et al., 2013; Zhang, Zhang, Wu, & Schluesener, 2012). Further, histone acetyl transferase inhibitors like curcumin were found to repress infiltration of inflammatory cells in the spinal cord (Xie et al., 2009). More recently, 5‐azacytidine, a DNA methylation inhibitor, was shown to inhibit EAE by induction of regulatory T cells in a mouse EAE model (Chan et al., 2014).
7.2. Hormone therapy
Corticosteroids are commonly used drugs for treatment of acute MS relapses (Karussis, 2013). Glucocorticoids strongly downregulate inflammation by inhibiting genes that affect the production of cytokines including IL‐1, IL‐2, IL‐3, IL‐4, IL‐5, IL‐6, IL‐8, and TNF‐α. This in turn, reduces the proliferative ability of T cells (Abbruzzese et al., 1983; Barkhof, Hommes, Scheltens, & Valk, 1991; Beck et al., 1992; Miller et al., 1991; Rose et al., 1970), and induces the synthesis of lipocortin‐1 to minimize eicosanoid production and reduce the permeability of BBB with steroids (Barkhof et al., 1991; Kesselring et al., 1989; Miller et al., 1992). Studies have shown that both corticotrophin (ACTH) and intravenous methylprednisolone could be used in short‐term treatment of the acute exacerbations of MS (Berkovich et al., 2017). An alternative to corticosteroids are sex hormones, for example, testosterone supplements can be protective in males and combination of estradiol and progesterone is protective in case of females (Golden & Voskuhl, 2016).
7.3. Immunotherapy
7.3.1. T cell therapy
The first line of treatment targeting T cells in both the clinic and mouse EAE models was represented by type 1 interferons and glatiramer acetate (GA). IFN‐β reduces relapses and slows progression of the disease (Abreu, 1982). GA is a synthetic co‐polymer containing alanine, glutamine, lysine, and tyrosine (Norohna, Toscas, & Jensen, 1990; Rudick, Carpenter, Cookfair, Tuohy, & Ransohoff, 1993). This compound shares immunologic similarities with the MBP molecule and can reduce inflammation by preventing lymphocyte sensitization (Abreu, 1985). GA is capable of downregulating antigen presentation through competitive inhibition for binding to MHC/HLA, thereby inducing a Th2 shift, in regulatory and CD8 suppressor GA‐reactive cells (Comi, Filippi, Wolinsky, & Group, 2001).
7.3.2. B‐cell therapy
B cell immunity plays an integral part in the development of MS because of its role in antibody presentation, cytokine production, meningeal inflammation, axonal degeneration, and grey matter demyelination (McLauchlan & Robertson, 2017). Therefore, B cell‐depleting therapy as an alternative to T cell therapy is considered highly effective against relapsing MS, and disability worsening in PPMS. The B cell‐depleting therapy is divided into two categories; polyclonal and monoclonal antibody therapies. Polyclonal antibody therapies inhibit cell‐mediated immune reactions by causing general immunosuppression, albeit not commonly used in MS due to high toxicity (Walker, Hoehn, & Kashiwagi, 1976). On the other hand, numerous monoclonal antibodies including Natalizumab, Alemtuzumab and Daclizumab have proven effective in early RRMS(Nguyen, Gresle, Marshall, Butzkueven, & Field, 2017). In addition, three emerging monoclonal antibodies against CD20‐positive B cells (rituximab, ocrelizumab, and ofatumumab) have shown overall promising effects in clinical phase II and III trials (Abbruzzese et al., 1983; Reff et al., 1994; Stashenko, Nadler, Hardy, & Schlossman, 1980). Other developing therapies against relapsing forms of the disease targets B cell CD19 antigen and B cell cytokine signaling molecules (Schneider et al., 1999).
7.4. Stem cell therapy
Hematopoietic stem cell transplantation suppresses the immune system and mitigates the inflammatory responses generated by an insult (Burt et al., 2009). The entire immune system is reconstructed using the patient's own stem cells (Burt et al., 2009). In numerous clinical cases, the use of stem cell therapy was able to bring about an induction of remission and stabilization of MS (Burt et al., 2009). In addition, stem cell therapy using embryonic and other types of adult stem cells, particularly mesenchymal stem cells, are currently being studied using the EAE model (Kassis et al., 2008; Zappia et al., 2005).
7.5. Electrical stimulation
Electrical stimulation has emerged as a potential therapeutic technique for MS. A pilot study showed that intraluminal electrical pharyngeal stimulation can treat dysphagia caused by MS (Restivo et al., 2013). A randomized control trial showed that resistance training of light to moderate intensity over long periods of time increases muscle strength in MS patients, while concurrent use of electrical stimulation does not help to further improve the outcome (Broekmans et al., 2011). Additionally, functional electrical stimulation has tremendous potential as a therapeutic strategy to aid with gait in MS patients (Hausmann et al., 2015; Sampson et al., 2016; van der Linden, Scott, Hooper, Cowan, & Mercer, 2014).
8. CONCLUSIONS
Although substantial resources have been devoted on finding a cure for MS, based on current knowledge about the etiology of the disease, further studies are required to determine the association between virus and bacteria in the pathology of MS development (Figure 3). The most effective etiologic agent causing MS is still not known, though it is possible that multiple agents work in synchronization to hijack the immune system and aggravate the disease. As such, a deeper understanding of the underlying mechanisms is needed because a variety of factors associated with numerous diseases in completely different subsets of patients predisposed genetically for the disease have been shown. In addition, more studies on extent of interplay between MS and other non‐pathogenic environmental factors like vitamin D levels, exposure to UV‐light, consumption of tobacco, with respect to MS pathogenesis are required.
Figure 3.
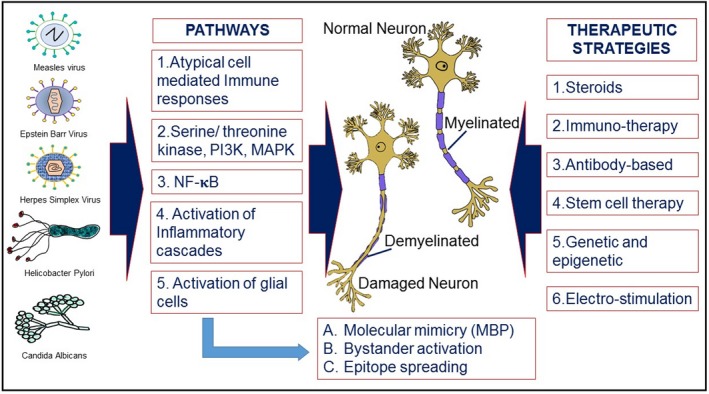
Schematic diagram of environmental factors and host derived pathways in causation of the disease and therapeutic strategies associated with multiple sclerosis (MS) progression. Although many correlations have been established between foreign pathogens and MS, there is no evidence for a direct association between any pathogen and the development or progression of MS. It is still unclear, whether bacterial or viral pathogens are capable of initiating MS in susceptible patients and exacerbate MS symptoms. As multiple species of bacteria and virus have been shown to associate with MS, it is possible that the mechanism of this association is a result of generalized inflammation caused by the immune response to these pathogens. The listed pathogens have been reported to cause one or more of the indicated pathways which culminate in the demyelination of neurons and progression of MS. The outlined therapeutic strategies include the existing treatment options as well as the focuses of current research
CONFLICT OF INTEREST
That authors declare no conflict of interest.
AUTHOR CONTRIBUTIONS
ST: Assisted in writing the manuscript and assistance with Figures 1 and 2. JL: Assisted in writing the manuscript and assistance with Figure 2. CRO: Assisted in writing the manuscript. KM: Assisted in writing the manuscript. TP: Assisted in writing the section related to treatment option and provided assistance with Figure 3. RKD: Assisted in writing the section related to viruses. AC: Assisted in writing the section related to bacteria. LG: Assisted in writing the section related to viruses, and provided assistance with Figure 2. YC: Assisted in writing the section related to animal model. ME‐L: Assisted in writing the section related to bacteria. NE‐H: Course director, assisted in writing the abstract and introduction, assisted with editing and provided assistance with Figures 1, 2 and 3.
ACKNOWLEDGMENTS
This review was written as part of the curriculum for the course GMS 6939, offered to the Ph.D. graduate students in the Biomedical Science Program at Florida International University, a minority serving institution. Authors contributed to the writing and editing equally.
Tiwari S, Lapierre J, Ojha CR, et al. Signaling pathways and therapeutic perspectives related to environmental factors associated with multiple sclerosis. J Neuro Res. 2018;96:1831–1846. 10.1002/jnr.24322
REFERENCES
- Abbruzzese, J. L. , Rocco, L. E. , Laskin, O. L. , Skubitz, K. M. , McGaughey, M. D. , & Lipsky, J. J. (1983). Prospective randomized double‐blind comparison of moxalactam and tobramycin in treatment of urinary tract infections. American Journal of Medicine, 74(4), 694–699. 10.1016/0002-9343(83)91029-X [DOI] [PubMed] [Google Scholar]
- Abdelrahman, H. S. , Selim, H. S. , Hashish, M. H. , & Sultan, L. I. (2014). Epstein‐Barr virus in multiple sclerosis. Journal of the Egyptian Public Health Association, 89(2), 90–95. 10.1097/01.EPX.0000452287.65133.fc [DOI] [PubMed] [Google Scholar]
- Abreu, S. L. (1982). Suppression of experimental allergic encephalomyelitis by interferon. Immunological Communications, 11(1), 1–7. 10.3109/08820138209050718 [DOI] [PubMed] [Google Scholar]
- Abreu, S. L. (1985). Interferon in experimental autoimmune encephalomyelitis (EAE): Effects of exogeneous interferon on the antigen‐enhanced adoptive transfer of EAE. International Archives of Allergy and Immunology, 76(4), 302–307. 10.1159/000233711 [DOI] [PubMed] [Google Scholar]
- Ahlgren, C. , Oden, A. , & Lycke, J. (2012). A nationwide survey of the prevalence of multiple sclerosis in immigrant populations of Sweden. Multiple Sclerosis Journal, 18(8), 1099–1107. 10.1177/1352458511433062 [DOI] [PubMed] [Google Scholar]
- Ahlgren, C. , Odén, A. , Torén, K. , & Andersen, O. (2009). Multiple sclerosis incidence in the era of measles‐mumps‐rubella mass vaccinations. Acta Neurologica Scandinavica, 119, 313–320. 10.1111/j.1600-0404.2008.01131.x [DOI] [PubMed] [Google Scholar]
- Amato, M. P. , Goretti, B. , Ghezzi, A. , Lori, S. , Zipoli, V. , Moiola, L. , … Trojano, M. (2010). Cognitive and psychosocial features in childhood and juvenile MS: Two‐year follow‐up. Neurology, 75(13), 1134–1140. 10.1212/WNL.0b013e3181f4d821 [DOI] [PubMed] [Google Scholar]
- An, S. F. , Giometto, B. , Groves, M. , Miller, R. F. , Beckett, A. A. , Gray, F. , … Scaravilli, F. (1997). Axonal damage revealed by accumulation of beta‐APP in HIV‐positive individuals without AIDS. Journal of Neuropathology and Experimental Neurology, 56(11), 1262–1268. [DOI] [PubMed] [Google Scholar]
- Aranow, C. (2011). Vitamin D and the immune system. Journal of Investigative Medicine, 59(6), 881–886. 10.2310/JIM.0b013e31821b8755 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ascherio, A. , & Munger, K. L. (2007). Environmental risk factors for multiple sclerosis. Part I: The role of infection. Annals of Neurology, 61, 288–299. 10.1002/ana.21117 [DOI] [PubMed] [Google Scholar]
- Ascherio, A. , Munger, K. L. , Lennette, E. T. , Spiegelman, D. , Hernan, M. A. , Olek, M. J. , … Hunter, D. J. (2001). Epstein‐Barr virus antibodies and risk of multiple sclerosis: A prospective study. JAMA, 286(24), 3083–3088. 10.1001/jama.286.24.3083 [DOI] [PubMed] [Google Scholar]
- Baranzini, S. E. , Mudge, J. , van Velkinburgh, J. C. , Khankhanian, P. , Khrebtukova, I. , Miller, N. A. , … Kingsmore, S. F. (2010). Genome, epigenome and RNA sequences of monozygotic twins discordant for multiple sclerosis. Nature, 464(7293), 1351–1356. 10.1038/nature08990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barkhof, F. , Hommes, O. R. , Scheltens, P. , & Valk, J. (1991). Quantitative MRI changes in gadolinium‐DTPA enhancement after high‐dose intravenous methylprednisolone in multiple sclerosis. Neurology, 41(8), 1219–1222. 10.1212/WNL.41.8.1219 [DOI] [PubMed] [Google Scholar]
- Baxi, E. G. , DeBruin, J. , Tosi, D. M. , Grishkan, I. V. , Smith, M. D. , Kirby, L. A. , … Gocke, A. R. (2015). Transfer of myelin‐reactive th17 cells impairs endogenous remyelination in the central nervous system of cuprizone‐fed mice. Journal of Neuroscience, 35(22), 8626–8639. 10.1523/JNEUROSCI.3817-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck, R. W. , Cleary, P. A. , Anderson, M. M. , Keltner, J. L. , Shults, W. T. , Kaufman, D. I. … Atwell, C. W. (1992). A randomized, controlled trial of cortico‐ steroids in the treatment of acute optic neuritis. New England Journal of Medicine, 326(9), 581–588. 10.1056/NEJM199202273260901 [DOI] [PubMed] [Google Scholar]
- Berkovich, R. , Bakshi, R. , Amezcua, L. , Axtell, R. C. , Cen, S. Y. , Tauhid, S. , … Steinman, L. (2017). Adrenocorticotropic hormone versus methylprednisolone added to interferon β in patients with multiple sclerosis experiencing breakthrough disease: A randomized, rater‐blinded trial. Therapeutic Advances in Neurological Disorders, 10(1), 3–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boullerne, A. I. , Skias, D. , Hartman, E. M. , Testai, F. D. , Kalinin, S. , Polak, P. E. , & Feinstein, D. L. (2015). A single‐nucleotide polymorphism in serine‐threonine kinase 11, the gene encoding liver kinase B1, is a risk factor for multiple sclerosis. ASN Neuro, 7(1). 10.1177/1759091415568914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broekmans, T. , Roelants, M. , Feys, P. , Alders, G. , Gijbels, D. , Hanssen, I. , … Eijnde, B. O. (2011). Effects of long‐term resistance training and simultaneous electro‐stimulation on muscle strength and functional mobility in multiple sclerosis. Multiple Sclerosis Journal, 17(4), 468–477. 10.1177/1352458510391339 [DOI] [PubMed] [Google Scholar]
- Buckingham, E. M. , Jarosinski, K. W. , Jackson, W. , Carpenter, J. E. , & Grose, C. (2016). Exocytosis of varicella‐zoster virus virions involves a convergence of endosomal and autophagy pathways. Journal of Virology, 90(19), 8673–8685. 10.1128/JVI.00915-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt, R. K. , Loh, Y. , Cohen, B. , Stefosky, D. , Balabanov, R. , Katsamakis, G. … Burns, W. H. (2009). Autologous non‐myeloablative haemopoi‐ etic stem cell transplantation in relapsing‐remitting multiple sclerosis: A phase I/II study. The Lancet Neurology, 8(3), 244–253. 10.1016/S1474-4422(09)70017-1 [DOI] [PubMed] [Google Scholar]
- Cahoon, L. (2009). The curious case of clioquinol. Nature Medicine, 15(4), 356–359. 10.1038/nm0409-356 [DOI] [PubMed] [Google Scholar]
- Camelo, S. , Iglesias, A. H. , Hwang, D. , Due, B. , Ryu, H. , Smith, K. , … Dangond, F. (2005). Transcriptional therapy with the histone deacetylase inhibitor trichostatin A ameliorates experimental autoimmune encephalomyelitis. Journal of Neuroimmunology, 164(1–2), 10–21. 10.1016/j.jneuroim.2005.02.022 [DOI] [PubMed] [Google Scholar]
- Caminero, A. , Comabella, M. , & Montalban, X. (2011). Role of tumour necrosis factor (TNF)‐alpha and TNFRSF1A R92Q mutation in the pathogenesis of TNF receptor‐associated periodic syndrome and multiple sclerosis. Clinical and Experimental Immunology, 166(3), 338–345. 10.1111/j.1365-2249.2011.04484.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cekanaviciute, E. , Yoo, B. B. , Runia, T. F. , Debelius, J. W. , Singh, S. , Nelson, C. A. , … Baranzini, S. E. (2017). Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Proceedings of the National Academy of Sciences USA, 114(40), 10713–10718. 10.1073/pnas.1711235114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cepok, S. , Zhou, D. , Srivastava, R. , Nessler, S. , Stei, S. , Büssow, K. , … Hemmer, B. (2005). Identification of Epstein‐Barr virus proteins as putative targets of the immune response in multiple sclerosis. The Journal of Clinical Investigation, 115, 1352–1360. 10.1172/JCI23661 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chalkias, S. , Dang, X. , Bord, E. , Stein, M. C. , Kinkel, R. P. , Sloane, J. A. , … Koralnik, I. J. (2014). JC virus reactivation during prolonged natalizumab monotherapy for multiple sclerosis. Annals of Neurology, 75, 925–934. 10.1002/ana.24148 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan, M. W. , Chang, C. B. , Tung, C. H. , Sun, J. , Suen, J. L. , & Wu, S. F. (2014). Low‐dose 5‐aza‐2'‐deoxycytidine pretreatment inhibits experimental autoimmune encephalomyelitis by induction of regulatory T cells. Molecular Medicine, 20, 248–256. 10.2119/molmed.2013.00159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chari, D. M. (2007). Remyelination in multiple sclerosis. International Review of Neurobiology, 79, 589–620. 10.1016/S0074-7742(07)79026-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Comi, G. , Filippi, M. , Wolinsky, J. S. , & European/Canadian Glatiramer Acetate Study Group . (2001). European/Canadian multicenter, double‐blind, randomized, placebo‐controlled study of the effects of glatiramer acetate on magnetic resonance imaging–measured disease activity and burden in patients with relapsing multiple sclerosis. Annals of Neurology, 49(3), 290–297. 10.1002/ana.64 [DOI] [PubMed] [Google Scholar]
- Compston, A. , & Coles, A. (2002). Multiple sclerosis. Lancet, 359(9313), 1221–1231. 10.1016/S0140-6736(02)08220-X [DOI] [PubMed] [Google Scholar]
- Compston, A. , & Coles, A. (2008). Multiple sclerosis. Lancet, 372(9648), 1502–1517. 10.1016/S0140-6736(08)61620-7 [DOI] [PubMed] [Google Scholar]
- Correale, J. , Gaitan, M. I. , Ysrraelit, M. C. , & Fiol, M. P. (2017). Progressive multiple sclerosis: From pathogenic mechanisms to treatment. Brain, 140(3), 527–546. 10.1093/brain/aww258 [DOI] [PubMed] [Google Scholar]
- Cossu, D. , Cocco, E. , Paccagnini, D. , Masala, S. , Ahmed, N. , Frau, J. , … Sechi, L. A. (2011). Association of Mycobacterium avium subsp. paratuberculosis with multiple sclerosis in Sardinian patients. PLoS One, 6(4), e18482 10.1371/journal.pone.0018482 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das Sarma, J. , Kenyon, L. C. , Hingley, S. T. , & Shindler, K. S. (2009). Mechanisms of primary axonal damage in a viral model of multiple sclerosis. Journal of Neuroscience, 29(33), 10272–10280. 10.1523/JNEUROSCI.1975-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Jager, P. L. , Jia, X. , Wang, J. , de Bakker, P. I. , Ottoboni, L. , Aggarwal, N. T. , … Oksenberg, J. R. (2009). Meta‐analysis of genome scans and replication identify CD6, IRF8 and TNFRSF1A as new multiple sclerosis susceptibility loci. Nature Genetics, 41(7), 776–782. 10.1038/ng.401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deber, C. M. , & Reynolds, S. J. (1991). Central nervous system myelin: Structure, function, and pathology. Clinical Biochemistry, 24(2), 113–134. 10.1016/0009-9120(91)90421-A [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeBruin, L. S. , Haines, J. D. , Bienzle, D. , & Harauz, G. (2006). Partitioning of myelin basic protein into membrane microdomains in a spontaneously demyelinating mouse model for multiple sclerosis. Biochemistry and Cell Biology, 84(6), 993–1005. 10.1139/o06-180 [DOI] [PubMed] [Google Scholar]
- DeBruin, L. S. , Haines, J. D. , Wellhauser, L. A. , Radeva, G. , Schonmann, V. , Bienzle, D. , & Harauz, G. (2005). Developmental partitioning of myelin basic protein into membrane microdomains. Journal of Neuroscience Research, 80(2), 211–225. 10.1002/jnr.20452 [DOI] [PubMed] [Google Scholar]
- Disanto, G. , Morahan, J. M. , Barnett, M. H. , Giovannoni, G. , & Ramagopalan, S. V. (2012). The evidence for a role of B cells in multiple sclerosis. Neurology, 78(11), 823–832. 10.1212/WNL.0b013e318249f6f0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donati, D. , & Jacobson, S. (2002). Viruses and multiple sclerosis In Brogden K. A., & Guthmiller J. M. (Eds.), Polymicrobial Diseases. Washington (DC): ASM Press; Available from: https://www.ncbi.nlm.nih.gov/books/NBK2494 [PubMed] [Google Scholar]
- Duddy, M. , Niino, M. , Adatia, F. , Hebert, S. , Freedman, M. , Atkins, H. , … Bar‐Or, A. (2007). Distinct effector cytokine profiles of memory and naive human B cell subsets and implication in multiple sclerosis. The Journal of Immunology, 178(10), 6092–6099. 10.4049/jimmunol.178.10.6092 [DOI] [PubMed] [Google Scholar]
- Freund, J. , Stern, E. R. , & Pisani, T. M. (1947). Isoallergic encephalomyelitis and radiculitis in guinea pigs after one injection of brain and Mycobacteria in water‐in‐oil emulsion. Journal of Immunology, 57(2), 179–194. [PubMed] [Google Scholar]
- Fainardi, E. , Castellazzi, M. , Seraceni, S. , Granieri, E. , & Contini, C. (2008). Under the microscope: Focus on Chlamydia pneumoniae infection and multiple sclerosis. Current Neurovascular Research, 5(1), 60–70. [DOI] [PubMed] [Google Scholar]
- Ferreira, M. L. , Machado, M. I. , Dantas, M. J. , Moreira, A. J. , & Souza, A. M. (2008). Pediatric multiple sclerosis: Analysis of clinical and epidemiological aspects according to National MS Society Consensus 2007. Arquivos De Neuro‐Psiquiatria, 66(3b), 665–670. 10.1590/S0004-282X2008000500011 [DOI] [PubMed] [Google Scholar]
- Freund, J. , & McDermott, K. (1942). Sensitization to horse serum by means of adjuvants. Proceedings of the Society for Experimental Biology and Medicine, 49(4), 548–553. 10.3181/00379727-49-13625 [DOI] [Google Scholar]
- Fujinami, R. S. , & Oldstone, M. B. (1985). Amino acid homology between the encephalitogenic site of myelin basic protein and virus: Mechanism for autoimmunity. Science (New York, N.Y.), 230, 1043–1045. [DOI] [PubMed] [Google Scholar]
- Fujinami, R. S. , von Herrath, M. G. , Christen, U. , & Whitton, J. L. (2006). Molecular mimicry, bystander activation, or viral persistence: Infections and autoimmune disease. Clinical Microbiology Reviews, 19(1), 80–94. 10.1128/CMR.19.1.80-94.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gandhi, R. , Laroni, A. , & Weiner, H. L. (2010). Role of the innate immune system in the pathogenesis of multiple sclerosis. Journal of Neuroimmunology, 221(1–2), 7–14. 10.1016/j.jneuroim.2009.10.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gavalas, E. , Kountouras, J. , Boziki, M. , Zavos, C. , Polyzos, S. A. , Vlachaki, E. , … Deretzi, G. (2015). Relationship between Helicobacter pylori infection and multiple sclerosis. Ann Gastroenterol, 28(3), 353–356. [PMC free article] [PubMed] [Google Scholar]
- Ge, Z. , Da, Y. , Xue, Z. , Zhang, K. , Zhuang, H. , Peng, M. , … Zhang, R. (2013). Vorinostat, a histone deacetylase inhibitor, suppresses dendritic cell function and ameliorates experimental autoimmune encephalomyelitis. Experimental Neurology, 241, 56–66. 10.1016/j.expneurol.2012.12.006 [DOI] [PubMed] [Google Scholar]
- Gejman, P. V. , Sanders, A. R. , & Duan, J. (2010). The role of genetics in the etiology of schizophrenia. Psychiatric Clinics of North America, 33(1), 35–66. 10.1016/j.psc.2009.12.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Germain, M. , Nguyen, A. P. , Khacho, M. , Patten, D. A. , Screaton, R. A. , Park, D. S. , & Slack, R. S. (2013). LKB1‐regulated adaptive mechanisms are essential for neuronal survival following mitochondrial dysfunction. Human Molecular Genetics, 22(5), 952–962. 10.1093/hmg/dds500 [DOI] [PubMed] [Google Scholar]
- Ghasemi, N. , Razavi, S. , & Nikzad, E. (2017). Multiple sclerosis: Pathogenesis, symptoms. Diagnoses and cell‐based therapy. Cell Journalournal, 19(1), 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giovannoni, G. , Cutter, G. R. , Lunemann, J. , Martin, R. , Munz, C. , Sriram, S. , … Gaydos, C. A. (2006). Infectious causes of multiple sclerosis. The Lancet Neurology, 5(10), 887–894. 10.1016/S1474-4422(06)70577-4 [DOI] [PubMed] [Google Scholar]
- Giovannoni, G. , & Ebers, G. (2007). Multiple sclerosis: The environment and causation. Current Opinion in Neurology, 20(3), 261–268. 10.1097/WCO.0b013e32815610c2 [DOI] [PubMed] [Google Scholar]
- Golden, L. C. , & Voskuhl, R. (2016). The importance of studying sex differences in disease: The example of multiple sclerosis. Journal of Neuroscience Research, 95(1–2), 633–643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldenberg, M. M. (2012). Multiple sclerosis review. Pharmacy and Therapeutics, 37(3), 175–184. [PMC free article] [PubMed] [Google Scholar]
- Golubovskaya, V. , & Wu, L. (2016). Different subsets of T cells, memory, effector functions, and CAR‐T immunotherapy. Cancers (Basel), 8(3), 36 10.3390/cancers8030036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graef, I. T. , Henze, T. , & Reiber, H. (1994). Polyspecific immune reaction in the central nervous system in autoimmune diseases with CNS involvement. Zeitschrift Fur Arztliche Fortbildung, 88(7–8), 587–591. [PubMed] [Google Scholar]
- Grayston, J. T. , Wang, S. P. , Kuo, C. C. , & Campbell, L. A. (1989). Current knowledge on Chlamydia pneumoniae, strain TWAR, an important cause of pneumonia and other acute respiratory diseases. European Journal of Clinical Microbiology and Infectious Diseases, 8(3), 191–202. 10.1007/BF01965260 [DOI] [PubMed] [Google Scholar]
- Gupta, D. , Wang, Q. , Vinson, C. , & Dziarski, R. (1999). Bacterial peptidoglycan induces CD14‐dependent activation of transcription factors CREB/ATF and AP‐1. Journal of Biological Chemistry, 274(20), 14012–14020. 10.1074/jbc.274.20.14012 [DOI] [PubMed] [Google Scholar]
- Hassan‐Smith, G. , & Douglas, M. R. (2011). Epidemiology and diagnosis of multiple sclerosis. British Journal of Hospital Medicine (Lond), 72(10), M146–M151. 10.12968/hmed.2011.72.Sup10.M146 [DOI] [PubMed] [Google Scholar]
- Hausmann, J. , Sweeney‐Reed, C. M. , Sobieray, U. , Matzke, M. , Heinze, H. J. , Voges, J. , & Buentjen, L. (2015). Functional electrical stimulation through direct 4‐channel nerve stimulation to improve gait in multiple sclerosis: A feasibility study. Journal of NeuroEngineering and Rehabilitation, 12, 100 10.1186/s12984-015-0096-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawkes, C. H. (2007). Smoking is a risk factor for multiple sclerosis: A metanalysis. Multiple Sclerosis Journal, 13(5), 610–615. 10.1177/1352458506073501 [DOI] [PubMed] [Google Scholar]
- Healy, B. C. , Ali, E. N. , Guttmann, C. R. , Chitnis, T. , Glanz, B. I. , Buckle, G. , … Ascherio, A. (2009). Smoking and disease progression in multiple sclerosis. Archives of Neurology, 66(7), 858–864. 10.1001/archneurol.2009.122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernan, M. A. , Jick, S. S. , Logroscino, G. , Olek, M. J. , Ascherio, A. , & Jick, H. (2005). Cigarette smoking and the progression of multiple sclerosis. Brain, 128(Pt 6), 1461–1465. 10.1093/brain/awh471 [DOI] [PubMed] [Google Scholar]
- Hoffjan, S. , Okur, A. , Epplen, J. T. , Wieczorek, S. , Chan, A. , & Akkad, D. A. (2015). Association of TNFAIP3 and TNFRSF1A variation with multiple sclerosis in a German case‐control cohort. International Journal of Immunogenetics, 42(2), 106–110. 10.1111/iji.12183 [DOI] [PubMed] [Google Scholar]
- Horien, C. , & Grose, C. (2012). Neurovirulence of varicella and the live attenuated varicella vaccine virus. Seminars in Pediatric Neurology, 19(3), 124–129. 10.1016/j.spen.2012.02.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huynh, J. L. , & Casaccia, P. (2013). Epigenetic mechanisms in multiple sclerosis: Implications for pathogenesis and treatment. The Lancet Neurology, 12(2), 195–206. 10.1016/s1474-4422(12)70309-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hwang, E. S. , Hong, J. H. , & Glimcher, L. H. (2005). IL‐2 production in developing Th1 cells is regulated by heterodimerization of RelA and T‐bet and requires T‐bet serine residue 508. Journal of Experimental Medicine, 202(9), 1289–1300. 10.1084/jem.20051044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- International Multiple Sclerosis Genetics, C., Wellcome Trust Case Control, C. , Sawcer, S. , Hellenthal, G. , Pirinen, M. , Spencer, C. C. A. , … Compston, A. (2011). Genetic risk and a primary role for cell‐mediated immune mechanisms in multiple sclerosis. Nature, 476(7359), 214–219. 10.1038/nature10251 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobson, S. , Flerlage, M. L. , & McFarland, H. F. (1985). Impaired measles virus‐specific cytotoxic T cell responses in multiple sclerosis. Journal of Experimental Medicine, 162(3), 839–850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jancic, J. , Nikolic, B. , Ivancevic, N. , Djuric, V. , Zaletel, I. , Stevanovic, D. , … Samardzic, J. (2016). Multiple sclerosis in pediatrics: Current concepts and treatment options. Neurology and Therapy, 5(2), 131–143. 10.1007/s40120-016-0052-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaruvongvanich, V. , Sanguankeo, A. , Jaruvongvanich, S. , & Upala, S. (2016). Association between Helicobacter pylori infection and multiple sclerosis: A systematic review and meta‐analysis. Multiple Sclerosis and Related Disorders, 7, 92–97. 10.1016/j.msard.2016.03.013 [DOI] [PubMed] [Google Scholar]
- Jeffery, N. D. , & Blakemore, W. F. (1995). Remyelination of mouse spinal cord axons demyelinated by local injection of lysolecithin. Journal of Neurocytology, 24(10), 775–781. 10.1007/BF01191213 [DOI] [PubMed] [Google Scholar]
- Kabat, E. A. , Wolf, A. , & Bezer, A. E. (1947). The rapid production of acute disseminated encephalomyelitis in rhesus monkeys by injection of heterologous and homologous brain tissue with adjuvants. Journal of Experimental Medicine, 85(1), 117–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kano, K. , Katayama, T. , Takeguchi, S. , Asanome, A. , Takahashi, K. , Saito, T. , … Hasebe, N. (2017). Biopsy‐proven case of Epstein‐Barr virus (EBV)‐associated vasculitis of the central nervous system. Neuropathology, 37(3), 259–264. 10.1111/neup.12356 [DOI] [PubMed] [Google Scholar]
- Karussis, D. (2013). Immunotherapy of multiple sclerosis: The state of the art. BioDrugs: Clinical Immunotherapeutics, Biopharmaceuticals and Gene Therapy, 27(2), 113–148. 10.1007/s40259-013-0011-z [DOI] [PubMed] [Google Scholar]
- Kasper, L. H. , & Shoemaker, J. (2010). Multiple sclerosis immunology: The healthy immune system vs the MS immune system. Neurology, 74(Suppl 1), S2–S8. 10.1212/WNL.0b013e3181c97c8f [DOI] [PubMed] [Google Scholar]
- Kassis, I. , Grigoriadis, N. , Gowda‐Kurkalli, B. , Mizrachi‐Kol, R. , Ben‐Hur, T. , Slavin, … D. (2008). Neuroprotection and immunomod‐ ulation with mesenchymal stem cells in chronic experimental autoimmune encephalomyelitis. Archives of Neurology, 65(5), 753–761. [DOI] [PubMed] [Google Scholar]
- Kesselring, J. , Miller, D. H. , MacManus, D. G. , Johnson, G. , Milligan, N. M. , Scolding, N. , … McDonald, W. I. (1989). Quantitative magnetic resonance imaging in multiple sclerosis: The effect of high dose intravenous methyl‐ prednisolone. Journal of Neurology, Neurosurgery and Psychiatry, 52(1), 14–17. 10.1136/jnnp.52.1.14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kristensson, K. (1996). Sorting signals and targeting of infectious agents through axons: An annotation to the 100 years' birth of the name "axon". Brain Research Bulletin, 41(6), 327–333. 10.1016/S0361-9230(96)00255-9 [DOI] [PubMed] [Google Scholar]
- Kuhlmann, T. , Miron, V. , Cui, Q. , Wegner, C. , Antel, J. , & Bruck, W. (2008). Differentiation block of oligodendroglial progenitor cells as a cause for remyelination failure in chronic multiple sclerosis. Brain, 131(Pt 7), 1749–1758. 10.1093/brain/awn096 [DOI] [PubMed] [Google Scholar]
- Kumar, P. , Kretzschmar, B. , Herold, S. , Nau, R. , Kreutzfeldt, M. , Schutze, S. , … Hein, K. (2015). Beneficial effect of chronic Staphylococcus aureus infection in a model of multiple sclerosis is mediated through the secretion of extracellular adherence protein. Journal of Neuroinflammation, 12, 22 10.1186/s12974-015-0241-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laksono, B. M. , de Vries, R. D. , McQuaid, S. , Duprex, W. P. , & de Swart, R. L. (2016). Measles virus host invasion and pathogenesis. Viruses, 8, 210 10.3390/v8080210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lassmann, H. (2003). Axonal injury in multiple sclerosis. Journal of Neurology, Neurosurgery and Psychiatry, 74(6), 695–697. 10.1136/jnnp.74.6.695 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehmann, P. V. , Forsthuber, T. , Miller, A. , & Sercarz, E. E. (1992). Spreading of T‐cell autoimmunity to cryptic determinants of an autoantigen. Nature, 358, 155–157. 10.1038/358155a0 [DOI] [PubMed] [Google Scholar]
- Libbey, J. E. , & Fujinami, R. S. (2010). Potential triggers of MS. Results and Problems in Cell Differentiation, 51, 21–42. 10.1007/400_2008_12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipton, M. M. , & Freund, J. (1952). Encephalomyelitis in the rat following intracutaneous injection of central nervous system tissue with adjuvant. Proceedings of the Society for Experimental Biology and Medicine, 81(1), 260–261. 10.3181/00379727-81-19840 [DOI] [PubMed] [Google Scholar]
- Ludlow, M. , McQuaid, S. , Milner, D. , de Swart, R. L. , & Duprex, W. P. (2015). Pathological consequences of systemic measles virus infection. The Journal of Pathology, 235(2), 253–265. 10.1002/path.4457 [DOI] [PubMed] [Google Scholar]
- Lundmark, F. , Duvefelt, K. , Iacobaeus, E. , Kockum, I. , Wallstrom, E. , Khademi, M. , … Hillert, J. (2007). Variation in interleukin 7 receptor alpha chain (IL7R) influences risk of multiple sclerosis. Nature Genetics, 39(9), 1108–1113. 10.1038/ng2106 [DOI] [PubMed] [Google Scholar]
- Lunemann, J. D. , Kamradt, T. , Martin, R. , & Munz, C. (2007). Epstein‐barr virus: Environmental trigger of multiple sclerosis? Journal of Virology, 81(13), 6777–6784. 10.1128/JVI.00153-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marabita, F. , Almgren, M. , Sjoholm, L. K. , Kular, L. , Liu, Y. , James, T. , … Jagodic, M. (2017). Smoking induces DNA methylation changes in multiple sclerosis patients with exposure‐response relationship. Scientific Reports, 7(1), 14589 10.1038/s41598-017-14788-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcus, J. F. , & Waubant, E. L. (2013). Updates on clinically isolated syndrome and diagnostic criteria for multiple sclerosis. Neurohospitalist, 3(2), 65–80. 10.1177/1941874412457183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin, J. R. (1984). Intra‐axonal virus in demyelinative lesions of experimental herpes simplex type 2 infection. Journal of the Neurological Sciences, 63(1), 63–74. 10.1016/0022-510X(84)90109-6 [DOI] [PubMed] [Google Scholar]
- Matsushima, G. K. , & Morell, P. (2001). The neurotoxicant, cuprizone, as a model to study demyelination and remyelination in the central nervous system. Brain Pathology, 11(1), 107–116. 10.1111/j.1750-3639.2001.tb00385.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matzaraki, V. , Kumar, V. , Wijmenga, C. , & Zhernakova, A. (2017). The MHC locus and genetic susceptibility to autoimmune and infectious diseases. Genome Biology, 18(1), 76 10.1186/s13059-017-1207-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLauchlan, D. , & Robertson, N. P. (2017). B cell treatments for multiple sclerosis. Journal of Neurology, 264(4), 814–816. 10.1007/s00415-017-8442-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller, D. H. , Barkhof, F. , Berry, I. , Kappos, L. , Scotti, G. , & Thompson, A. J. (1991). Magnetic resonance imaging in monitoring the treatment of multiple sclerosis: Concerted action guidelines. Journal of Neurology, Neurosurgery and Psychiatry, 54(8), 683–688. 10.1136/jnnp.54.8.683 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller, D. , Barkhof, F. , Montalban, X. , Thompson, A. , & Filippi, M. (2005). Clinically isolated syndromes suggestive of multiple sclerosis, part 2: Non‐conventional MRI, recovery processes, and management. The Lancet Neurology, 4(6), 341–348. 10.1016/S1474-4422(05)70095-8 [DOI] [PubMed] [Google Scholar]
- Miller, D. H. , Thompson, A. J. , Morrissey, S. P. , MacManus, D. G. , Moore, S. G. , Kendall, B. E. , … McDonald, W. I. (1992). High dose steroids in acute relapses of multiple sclerosis: MRI evidence for a possible mechanism of therapeutic effect. Journal of Neurology, Neurosurgery and Psychiatry, 55(6), 450–453. 10.1136/jnnp.55.6.450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milo, R. , & Kahana, E. (2010). Multiple sclerosis: Geoepidemiology, genetics and the environment. Autoimmunity Reviews, 9(5), A387–A394. 10.1016/j.autrev.2009.11.010 [DOI] [PubMed] [Google Scholar]
- Mirza, A. , & Mao‐Draayer, Y. (2017). The gut microbiome and microbial translocation in multiple sclerosis. Clinical Immunology, 183, 213–224. 10.1016/j.clim.2017.03.001 [DOI] [PubMed] [Google Scholar]
- Moore, C. S. , Cui, Q. L. , Warsi, N. M. , Durafourt, B. A. , Zorko, N. , Owen, D. R. , … Bar‐Or, A. (2015). Direct and indirect effects of immune and central nervous system‐resident cells on human oligodendrocyte progenitor cell differentiation. The Journal of Immunology, 194(2), 761–772. 10.4049/jimmunol.1401156 [DOI] [PubMed] [Google Scholar]
- Morfopoulou, S. , Brown, J. R. , Davies, E. G. , Anderson, G. , Virasami, A. , Qasim, W. , … Breuer, J. (2016). Human coronavirus OC43 associated with fatal encephalitis. New England Journal of Medicine, 375(5), 497–498. 10.1056/NEJMc1509458 [DOI] [PubMed] [Google Scholar]
- Munoz, J. J. , Bernard, C. C. A. , & Mackay, I. R. (1984). Elicitation of experimental allergic encephalomyelitis (EAE) in mice with the aid of pertussigen. Cellular Immunology, 83(1), 92–100. 10.1016/0008-8749(84)90228-4 [DOI] [PubMed] [Google Scholar]
- Nguyen, A. L. , Gresle, M. , Marshall, T. , Butzkueven, H. , & Field, J. (2017). Monoclonal antibodies in the treatment of multiple sclerosis: Emergence of B‐cell‐targeted therapies. British Journal of Pharmacology, 174(13), 1895–1907. 10.1111/bph.13780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielsen, T. R. , Rostgaard, K. , Nielsen, N. M. , Koch‐Henriksen, N. , Haahr, S. , Sørensen, P. S. , & Hjalgrim, H. (2007). Multiple sclerosis after infectious mononucleosis. Archives of Neurology, 64, 72 10.1001/archneur.64.1.72 [DOI] [PubMed] [Google Scholar]
- Norohna, A. , Toscas, A. , & Jensen, M. (1990). Interferon beta augments suppressor cell function in multiple sclerosis. Annals of Neurology, 27, 207–210. [DOI] [PubMed] [Google Scholar]
- Norrby, E. , Link, H. , & Olsson, J. E. (1974). Measles virus antibodies in multiple sclerosis. Comparison of antibody titers in cerebrospinal fluid and serum. Archives of Neurology, 30, 285–292. 10.1001/archneur.1974.00490340013002 [DOI] [PubMed] [Google Scholar]
- Nylander, A. , & Hafler, D. A. (2012). Multiple sclerosis. Journal of Clinical Investigation, 122(4), 1180–1188. 10.1172/JCI58649 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oksenberg, J. R. , Baranzini, S. E. , Sawcer, S. , & Hauser, S. L. (2008). The genetics of multiple sclerosis: SNPs to pathways to pathogenesis. Nature Reviews Genetics, 9(7), 516–526. 10.1038/nrg2395 [DOI] [PubMed] [Google Scholar]
- Olitsky, P. K. , & Yager, R. H. (1949). Experimental disseminated encephalomyelitis in white mice. Journal of Experimental Medicine, 90(3), 213–224. 10.1084/jem.90.3.213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orton, S. M. , Ramagopalan, S. V. , Brocklebank, D. , Herrera, B. M. , Dyment, D. A. , Yee, I. M. , … Ebers, G. C. (2010). Effect of immigration on multiple sclerosis sex ratio in Canada: The canadian collaborative study. Journal of Neurology, Neurosurgery and Psychiatry, 81(1), 31–36. 10.1136/jnnp.2008.162784 [DOI] [PubMed] [Google Scholar]
- Owens, G. P. , Gilden, D. , Burgoon, M. P. , Yu, X. , & Bennett, J. L. (2011). Viruses and multiple sclerosis. Neuroscientist, 17(6), 659–676. 10.1177/1073858411386615 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pender, M. P. , & Burrows, S. R. (2014). Epstein‐Barr virus and multiple sclerosis: Potential opportunities for immunotherapy. Clinical & Translational Immunology, 3(10), e27 10.1038/cti.2014.25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popescu, B. F. , & Lucchinetti, C. F. (2012). Meningeal and cortical grey matter pathology in multiple sclerosis. BMC Neurology, 12, 11 10.1186/1471-2377-12-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Procaccini, C. , De Rosa, V. , Pucino, V. , Formisano, L. , & Matarese, G. (2015). Animal models of Multiple Sclerosis. European Journal of Pharmacology, 759, 182–191. 10.1016/j.ejphar.2015.03.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ransohoff, R. M. , Hafler, D. A. , & Lucchinetti, C. F. (2015). Multiple sclerosis‐a quiet revolution. Nature Reviews. Neurology, 11(3), 134–142. 10.1038/nrneurol.2015.14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raphael, I. , Nalawade, S. , Eagar, T. N. , & Forsthuber, T. G. (2015). T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine, 74(1), 5–17. 10.1016/j.cyto.2014.09.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reff, M. E. , Carner, K. , Chambers, K. S. , Chinn, P. C. , Leonard, J. E. , Raab, R. , … Anderson, D. R. (1994). Depletion of B cells in vivo by a chimeric mouse human monoclonal antibody to CD20. Blood, 83(2), 435–445. [PubMed] [Google Scholar]
- Renoux, C. , Vukusic, S. , Mikaeloff, Y. , Edan, G. , Clanet, M. , Dubois, B. , … Confavreux, C. (2007). Natural history of multiple sclerosis with childhood onset. New England Journal of Medicine, 356(25), 2603–2613. 10.1056/NEJMoa067597 [DOI] [PubMed] [Google Scholar]
- Restivo, D. A. , Casabona, A. , Centonze, D. , Marchese‐Ragona, R. , Maimone, D. , & Pavone, A. (2013). Pharyngeal electrical stimulation for dysphagia associated with multiple sclerosis: A pilot study. Brain Stimulation, 6(3), 418–423. 10.1016/j.brs.2012.09.001 [DOI] [PubMed] [Google Scholar]
- Reynaud, J. M. , Horvat, B. , & Horvat, B. (2013). Human herpesvirus 6 and neuroinflammation. ISRN Virology, 2013, 1–11. 10.5402/2013/834890 [DOI] [Google Scholar]
- Richards, M. H. , Getts, M. T. , Podojil, J. R. , Jin, Y. H. , Kim, B. S. , & Miller, S. D. (2011). Virus expanded regulatory T cells control disease severity in the Theiler's virus mouse model of MS. Journal of Autoimmunity, 36(2), 142–154. 10.1016/j.jaut.2010.12.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riise, T. , Nortvedt, M. W. , & Ascherio, A. (2003). Smoking is a risk factor for multiple sclerosis. Neurology, 61(8), 1122–1124. 10.1212/01.WNL.0000081305.66687.D2 [DOI] [PubMed] [Google Scholar]
- Rivers, T. M. , Sprunt, D. H. , & Berry, G. P. (1933). Observations on attempts to produce acute disseminated encephalomyelitis in monkeys. Journal of Experimental Medicine, 58(1), 39–53. 10.1084/jem.58.1.39 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rollings, C. M. , Sinclair, L. V. , Brady, H. J. M. , Cantrell, D. A. , & Ross, S. H. (2018). Interleukin‐2 shapes the cytotoxic T cell proteome and immune environment‐sensing programs. ScienceSignalling, 11(526). 10.1126/scisignal.aap8112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rose, A. S. , Kuzma, J. W. , Kurtzke, J. F. , Namerow, N. S. , Sibley, W. A. , & Tourtellotte, W. W. (1970). Cooperative study in the evaluation of therapy in multiple sclerosis. ACTH vs. placebo–final report. Neurology, 20(5), 1–59. [DOI] [PubMed] [Google Scholar]
- Rudick, R. , Carpenter, C. , Cookfair, D. , Tuohy, V. , & Ransohoff, R. (1993). In vitro and in vivo inhibition of mitogen‐driven T‐cell activation by recombinant interferon beta. Neurology, 43(10), 2080–2080. 10.1212/WNL.43.10.2080 [DOI] [PubMed] [Google Scholar]
- Salvetti, M. , Giovannoni, G. , & Aloisi, F. (2009). Epstein‐Barr virus and multiple sclerosis. Current Opinion in Neurology, 22, 201–206. 10.1097/WCO.0b013e32832b4c8d [DOI] [PubMed] [Google Scholar]
- Sampson, P. , Freeman, C. , Coote, S. , Demain, S. , Feys, P. , Meadmore, K. , & Hughes, A. M. (2016). Using functional electrical stimulation mediated by iterative learning control and robotics to improve arm movement for people with multiple sclerosis. IEEE Transactions on Neural Systems and Rehabilitation Engineering, 24(2), 235–248. 10.1109/tnsre.2015.2413906 [DOI] [PubMed] [Google Scholar]
- Sawcer, S. , Franklin, R. J. , & Ban, M. (2014). Multiple sclerosis genetics. The Lancet Neurology, 13(7), 700–709. 10.1016/S1474-4422(14)70041-9 [DOI] [PubMed] [Google Scholar]
- Saxena, A. , Martin‐Blondel, G. , Mars, L. T. , & Liblau, R. S. (2011). Role of CD8 T cell subsets in the pathogenesis of multiple sclerosis. FEBS Letters, 585(23), 3758–3763. 10.1016/j.febslet.2011.08.047 [DOI] [PubMed] [Google Scholar]
- Schneider, P. , MacKay, F. , Steiner, V. , Hofmann, K. , Bodmer, J. L. , Holler, N. , … Tschopp, J. (1999). BAFF, a novel ligand of the tumor necrosis factor family, stimulates B cell growth. Journal of Experimental Medicine, 189(11), 1747–1756. 10.1084/jem.189.11.1747 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schreiner, T. L. (2018). Pediatric multiple sclerosis: Background, pathophysiology, etiology. Retrieved from https://emedicine.medscape.com/article/2091406-overview [Google Scholar]
- Schrijver, I. A. , van Meurs, M. , Melief, M. J. , Wim Ang, C. , Buljevac, D. , Ravid, R. , … Laman, J. D. (2001). Bacterial peptidoglycan and immune reactivity in the central nervous system in multiple sclerosis. Brain, 124(Pt 8), 1544–1554. 10.1093/brain/124.8.1544 [DOI] [PubMed] [Google Scholar]
- Schwab, N. , Schneider‐Hohendorf, T. , Pignolet, B. , Breuer, J. , Gross, C. C. , Gobel, K. , … Wiendl, H. (2016). Therapy with natalizumab is associated with high JCV seroconversion and rising JCV index values. Neurol Neuroimmunol Neuroinflamm, 3(1), e195 10.1212/NXI.0000000000000195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silber, E. , & Sharief, M. K. (1999). Axonal degeneration in the pathogenesis of multiple sclerosis. Journal of the Neurological Sciences, 170(1), 11–18. 10.1016/S0022-510X(99)00178-1 [DOI] [PubMed] [Google Scholar]
- Sospedra, M. , & Martin, R. (2005). Immunology of multiple sclerosis. Annual Review of Immunology, 23, 683–747. 10.1146/annurev.immunol.23.021704.115707 [DOI] [PubMed] [Google Scholar]
- Sriram, S. , Mitchell, W. , & Stratton, C. (1998). Multiple sclerosis associated with chlamydia pneumoniae infection of the CNS. Neurology, 50(2), 571–572. 10.1212/WNL.50.2.571 [DOI] [PubMed] [Google Scholar]
- Sriram, S. , Stratton, C. W. , Yao, S. , Tharp, A. , Ding, L. , Bannan, J. D. , & Mitchell, W. M. (1999). Chlamydia pneumoniae infection of the central nervous system in multiple sclerosis. Annals of Neurology, 46(1), 6–14. 10.1002/1531-8249(199907)46:1aabbb6:AID-ANA4aaabb3.0.CO;2-M [DOI] [PubMed] [Google Scholar]
- Stashenko, P. , Nadler, L. M. , Hardy, R. , & Schlossman, S. F. (1980). Characterization of a human B lymphocyte‐specific antigen. The Journal of Immunology, 125(4), 1678–1685. [PubMed] [Google Scholar]
- Sullivan, B. M. , & Coscoy, L. (2010). The U24 protein from human herpesvirus 6 and 7 affects endocytic recycling. Journal of Virology, 84(3), 1265–1275. 10.1128/JVI.01775-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tait, A. R. , & Straus, S. K. (2008). Phosphorylation of U24 from human herpes virus type 6 (HHV‐6) and its potential role in mimicking myelin basic protein (MBP) in multiple sclerosis. FEBS Letters, 582(18), 2685–2688. 10.1016/j.febslet.2008.06.050 [DOI] [PubMed] [Google Scholar]
- Thacker, E. L. , Mirzaei, F. , & Ascherio, A. (2006). Infectious mononucleosis and risk for multiple sclerosis: A meta‐analysis. Annals of Neurology, 59, 499–503. 10.1002/ana.20820 [DOI] [PubMed] [Google Scholar]
- Thorley‐Lawson, D. A. (2001). Epstein‐Barr virus: Exploiting the immune system. Nature Reviews Immunology, 1, 75–82. 10.1038/35095584 [DOI] [PubMed] [Google Scholar]
- Tompkins, S. M. , Padilla, J. , Dal Canto, M. C. , Ting, J. P. , Van Kaer, L. , & Miller, S. D. (2002). De novo central nervous system processing of myelin antigen is required for the initiation of experimental autoimmune encephalomyelitis. The Journal of Immunology, 168(8), 4173–4183. 10.4049/jimmunol.168.8.4173 [DOI] [PubMed] [Google Scholar]
- Tselis, A. (2012). Epstein‐Barr virus cause of multiple sclerosis. Current Opinion in Rheumatology, 24, 424–428. 10.1097/BOR.0b013e3283542cf8 [DOI] [PubMed] [Google Scholar]
- Tsunoda, I. , & Fujinami, R. S. (2002). Inside‐out versus outside‐In models for virus induced demyelination: Axonal damage triggering demyelination. Springer Seminars in Immunopathology, 24(2), 105–125. 10.1007/s00281-002-0105-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsunoda, I. , & Fujinami, R. S. (2010). Neuropathogenesis of Theiler's murine encephalomyelitis virus infection, an animal model for multiple sclerosis. Journal of Neuroimmune Pharmacology, 5(3), 355–369. 10.1007/s11481-009-9179-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsunoda, I. , Iwasaki, Y. , Terunuma, H. , Sako, K. , & Ohara, Y. (1996). A comparative study of acute and chronic diseases induced by two subgroups of Theiler's murine encephalomyelitis virus. Acta Neuropathologica, 91(6), 595–602. 10.1007/s004010050472 [DOI] [PubMed] [Google Scholar]
- Tuohy, V. K. , Lu, Z. , Sobel, R. A. , Laursen, R. A. , & Lees, M. B. (1989). Identification of an encephalitogenic determinant of myelin proteolipid protein for SJL mice. The Journal of Immunology, 142(5), 1523–1527. [PubMed] [Google Scholar]
- van der Linden, M. L. , Scott, S. M. , Hooper, J. E. , Cowan, P. , & Mercer, T. H. (2014). Gait kinematics of people with multiple sclerosis and the acute application of functional electrical stimulation. Gait & Posture, 39(4), 1092–1096. 10.1016/j.gaitpost.2014.01.016 [DOI] [PubMed] [Google Scholar]
- van Etten, E. , & Mathieu, C. (2005). Immunoregulation by 1,25‐dihydroxyvitamin D3: Basic concepts. Journal of Steroid Biochemistry and Molecular Biology, 97(1–2), 93–101. 10.1016/j.jsbmb.2005.06.002 [DOI] [PubMed] [Google Scholar]
- Virtanen, J. O. , & Jacobson, S. (2012). Viruses and multiple sclerosis. CNS & Neurological Disorders: Drug Targets, 11(5), 528–544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker, J. E. , Hoehn, M. M. , & Kashiwagi, N. (1976). A trial of antilymphocyte globulin in the treatment of chronic progressive multiple sclerosis. Journal of the Neurological Sciences, 29(2–4), 303–309. 10.1016/0022-510X(76)90179-9 [DOI] [PubMed] [Google Scholar]
- Wallin, M. T. , Culpepper, W. J. , Coffman, P. , Pulaski, S. , Maloni, H. , Mahan, C. M. , … Veterans Affairs Multiple Sclerosis Centr es of Excellence Epidemiology, G. (2012). The gulf war era multiple sclerosis cohort: Age and incidence rates by race, sex and service. Brain, 135(Pt 6), 1778–1785. 10.1093/brain/aws099 [DOI] [PubMed] [Google Scholar]
- Weidemann, B. , Brade, H. , Rietschel, E. T. , Dziarski, R. , Bazil, V. , Kusumoto, S. , … Ulmer, A. J. (1994). Soluble peptidoglycan‐induced monokine production can be blocked by anti‐CD14 monoclonal antibodies and by lipid A partial structures. Infection and Immunity, 62(11), 4709–4715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiner, H. L. (2008). Oral laquinimod for treatment of relapsing‐remitting multiple sclerosis. The Lancet Neurology, 7(8), 672–673. 10.1016/S1474-4422(08)70148-0 [DOI] [PubMed] [Google Scholar]
- Weiss, S. R. (1983). Coronaviruses SD and SK share extensive nucleotide homology with murine coronavirus MHV‐A59, more than that shared between human and murine coronaviruses. Virology, 126(2), 669–677. 10.1016/S0042-6822(83)80022-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wingerchuk, D. M. (2012). Smoking: Effects on multiple sclerosis susceptibility and disease progression. Therapeutic Advances in Neurological Disorders, 5(1), 13–22. 10.1177/1756285611425694 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winterstein, C. , Trotter, J. , & Kramer‐Albers, E. M. (2008). Distinct endocytic recycling of myelin proteins promotes oligodendroglial membrane remodeling. Journal of Cell Science, 121(Pt 6), 834–842. 10.1242/jcs.022731 [DOI] [PubMed] [Google Scholar]
- Xie, L. , Li, X. K. , Funeshima‐Fuji, N. , Kimura, H. , Matsumoto, Y. , Isaka, Y. , & Takahara, S. (2009). Amelioration of experimental autoimmune encephalomyelitis by curcumin treatment through inhibition of IL‐17 production. International Immunopharmacology, 9(5), 575–581. 10.1016/j.intimp.2009.01.025 [DOI] [PubMed] [Google Scholar]
- Yao, K. , Honarmand, S. , Espinosa, A. , Akhyani, N. , Glaser, C. , & Jacobson, S. (2009). Detection of human herpesvirus‐6 in cerebrospinal fluid of patients with encephalitis. Annals of Neurology, 65(3), 257–267. 10.1002/ana.21611 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zappia, E. , Casazza, S. , Pedemonte, E. , Benvenuto, F. , Bonanni, I. , Gerdoni, E. … Mancardi, G. (2005). Mesenchymal stem cells ameliorate experi‐ mental autoimmune encephalomyelitis inducing T‐cell anergy. Blood, 106(5), 1755–1761. 10.1182/blood-2005-04-1496 [DOI] [PubMed] [Google Scholar]
- Zeman, D. , Adam, P. , Kalistova, H. , Sobek, O. , Kelbich, P. , Andel, J. , & Andel, M. (2000). Transferrin in patients with multiple sclerosis: A comparison among various subgroups of multiple sclerosis patients. Acta Neurologica Scandinavica, 101(2), 89–94. 10.1034/j.1600-0404.2000.101002089.x [DOI] [PubMed] [Google Scholar]
- Zhang, Z. , Zhang, Z. Y. , Wu, Y. , & Schluesener, H. J. (2012). Valproic acid ameliorates inflammation in experimental autoimmune encephalomyelitis rats. Neuroscience, 221, 140–150. 10.1016/j.neuroscience.2012.07.013 [DOI] [PubMed] [Google Scholar]