

Abstract
The means used by enveloped viruses to bypass cellular membranes are well characterized; however, the mechanisms used by non‐enveloped viruses to deliver their genome inside the cell remain unresolved and poorly defined. The discovery of short, membrane interacting, amphipathic or hydrophobic sequences (known as membranotropic peptides) in both enveloped and non‐enveloped viruses suggests that these small peptides are strongly involved in breaching the host membrane and in the delivery of the viral genome into the host cell. Thus, in spite of noticeable differences in entry, this short stretches of membranotropic peptides are probably associated with similar entry‐related events. This review will uncover the intrinsic features of viral membranotropic peptides involved in viral entry of both naked viruses and the ones encircled with a biological membrane with the objective to better elucidate their different functional properties and possible applications in the biomedical field.
Keywords: enveloped viruses, fusion peptide, membranotropic peptides, non‐enveloped virus

1. INTRODUCTION
Significant improvements have been achieved in recent years in the understanding of the multiple alternative ways of virus entry into susceptible cells.1, 2 The strategies employed by viruses to enter cells are different according to the presence or absence of a lipid bilayer surrounding the virus. Enveloped viruses present a membrane bilayer while non‐enveloped viruses lack this membrane and present on their surface only capsid proteins. The mechanism of cell invasion by the two groups of viruses is rather diverse and the foremost difference is the direct consequence of their distinctive physicochemical state at the interface that occurs at the time of the encounter between the virus and the cell membrane; in particular, two lipid membranes confronting each other in the case of enveloped viruses as opposed to a layer consisting only of proteins that face a lipid layer in the entry of naked viruses. Enveloped viruses entry exploits direct fusion with a cellular membrane through the involvement of specialized viral fusion proteins, present on the viral membrane and the consequent transfer of the nucleocapsid into the cytoplasm.3 On the other hand, the entry of non‐enveloped viruses, which lacking the outer viral membrane are unable to take advantage of the cellular mechanism of membrane fusion, involves the activation of viral lytic factors that induce cell membrane rupture.4 The major features in membrane interaction by enveloped and non‐enveloped viruses are reported in Figure 1.
Figure 1.

Different mechanisms of membrane interaction exploited for viral entry by non‐enveloped and enveloped viruses. Non‐enveloped viruses are reported on the left; capsid proteins release lytic factors responsible of nucleic acid internalization. Envelope viruses exploit the mechanism of membrane fusion either on the plasma membrane or within an endocytic vesicle
Notwithstanding the different mechanisms of penetration, the key step is the entry process and the modification of the cell membrane which allows the viral genome to penetrate into the host cell and start the replication cycle in the appropriate cellular compartment.5, 6
The most critical barriers for viral penetration and replication are the plasma membrane, the cytoplasm, and any other membrane that needs to be crossed in order to have access to the sites where viral replication or assembly take place. The overall picture of the mechanism of viral entry is becoming increasingly complete; in fact, depending on their dimension and structure, they have acquired different strategies to penetrate and take control of cell functions.3, 4 The molecular details of the interactions at the interface of virus and cell surfaces are quite complex and highly variable, but there is a common idea that only a limited number of pathways allowing viruses to reach the sites of penetration exist, with enveloped and non‐enveloped viruses presenting different and unrelated processes, but with general principles driving all fusion events. The main difference between fusion peptides of enveloped viruses and lytic factors of non‐enveloped viruses is the fact that fusion peptides promote membrane fusion while lytic factors promote membrane disruption.
In this review, we summarize current knowledge on the mechanism of membrane fusion of both enveloped and non‐enveloped viruses with a focus on common principles. Fusion peptide derived from enveloped viruses and lytic peptides from non‐enveloped viruses are described with an effort to find undisclosed differences and similarities. We conclude reporting ground breaking applications of membranotropic peptides (both fusion and lytic peptides) which open a new exiting field of research.
2. VIRAL ENTRY
2.1. Enveloped viruses entry
Enveloped viruses depend on membrane fusion for cell penetration.7 The two main routes used by enveloped viruses to enter the cell are the endocytic and non‐endocytic pathways.8 Most enveloped viruses undergo endocytosis while only few are able to fuse directly to the plasma membrane.8 Viruses that use the endocytic route for cellular internalization are able to escape into the cytosol avoiding lysosomal degradation. Therefore, penetration invariably involves membrane fusion mediated by specific viral glycoproteins (catalysts) with the main difference being that viruses using endocytic pathways fuse their envelope with the endosomal membrane from the luminal side.8 Thus, membrane fusion constitutes the essential and ubiquitous mechanism of entry of enveloped viruses, irrespective of their route of entry.5, 9, 10
Viral fusion proceeds through a hemifusion stalk with merging of proximal leaflets, which culminates in the opening and expansion of a pore connecting the two sides of the membrane.7 The overall process is supported by a catalyst responsible of the lowering of the transition barriers and fusion proteins constitute the catalytic agents fulfilling this function.11 The entry involves several other critical steps which include cellular receptor or co‐receptor binding, internalization, uncoating, and release of viral nucleic acids at the proper site of replication.7, 12, 13 Various factors can mediate an efficient interaction between cells and viruses; for example, cholesterol rich domains are platforms for the entry of many enveloped viruses such as the influenza virus14, 15 and many viral membranes contain much more cholesterol than the mammalian plasma membranes from which they are derived.16, 17
Viral fusion proteins undergo significant rearrangements from the pre‐fusion to the post‐fusion conformations which are triggered by either receptor binding, proteolytic cleavage or low endosomal pH, and eventually determine the exposure of previously sequestered hydrophobic peptides, loops, or patches, able to interact with and destabilize one or both the opposing membranes.6, 7
Crystallographic data on pre‐ and post‐fusion structures of viral fusion proteins has resulted in the identification of at least three distinct classes based on their three‐dimensional organization and mechanism of fusion (Figure 2).11, 18
Figure 2.

Schematic representation of the fusion process promoted by the three different classes of fusion proteins of enveloped viruses: Class I (panel A), Class II (panel B), and Class III (Panel C). Pre‐ and post‐fusion representation of fusion glycoproteins is reported. Fusion peptides are shown in red
Class I fusion proteins includes many of the most studied human pathogens such as influenza virus, human immunodeficiency virus (HIV), and Ebola virus; the key features is the requirement of a proteolytic cleavage to initiate fusion,19 activating these proteins which form an extended intermediate on the surface of the virion with the fusion peptide at the N‐terminus of the protein engaged in the interaction with the target membrane and forming a link between the two fusing bilayers. The pre‐hairpin structure is a unique state in which the fusion protein is simultaneously connecting two distinct membranes, the target membrane through the fusion peptide or patches and its own viral membrane through its transmembrane domain (TM). Further conformational changes produce trimers of hairpins with a central α‐helical coiled‐coil structure.20, 21 The 6‐helix formation is related to the opening of the fusion pore and provides the major driving force for the process. Thus, peptides able to prevent 6‐helix formation, interfering with refolding of the fusion glycoproteins act as potent viral entry inhibitors.22, 23
Class II fusion proteins, which include flaviviruses, alphaviruses, and bunyaviruses, are consistently different from those of Class I and are mainly composed of β structures.24 These fusion proteins are often heterodimers or homodimers lying almost flat on the virion surface.24 They are organized in three globular domains: Domain I is characterized by a central β‐sandwich which supports the three domain fold; Domain II originates from Domain I and comprises the fusion loop at the tip of the molecule; and Domain III, lies at the opposite end of Domain I, has an Ig‐like structure and terminates in a stem which connects it with the transmembrane anchor. Cleavage of the accessory protein, leads to an irreversible rearrangement of the fusion protein into a homotrimer which protrudes from the viral envelope and allows the penetration of the internal fusion peptide into the cell membrane.
Class III fusion proteins have a mixed secondary structure with the central α‐helical trimeric core similar to Class I and two fusion loops located at the tip of an elongated β‐sheet similar to Class II fusion proteins; the main representatives of Class III fusion proteins are protein G of vesicular stomatitis virus (VSV),25, 26 gB protein of herpes simplex virus (HSV),27 and of Epstein‐Barr virus (EBV),28 and gp64 from insect‐cell baculovirus.29 The post fusion structure is characterized by the presence of an internal fusion peptide in Domain I organized in two hydrophobic fusion loops flanked by a β‐sheet domain with a pleckstrin‐like fold (Domain II). Domain II is nested with the largely α‐helical Domain III, which is composed of trimers that give rise to an elongated, rod‐like shape molecule. The α‐helical domain is inserted in Domain IV which is made of β‐sheets and a very long C‐terminal extension (Domain V).
Interestingly, the multicomponent herpesvirus fusion machinery requires the presence of gB, gH/gL, and gD and multiple cellular receptors are engaged in the entry pathway in a cascade of molecular interactions.30 gH/gL and gB are part of the core fusion machinery and need to cooperate in order to trigger the initial lipid destabilization which culminates in the fusion of the two bilayers.31, 32, 33 Despite the fact that it is still debated whether gH is merely a fusion regulator or it plays a more direct role in the fusion process, studies leave little doubt that also the gH/gL complex undergoes dynamic rearrangements during the fusion process.34 The crystallographic post‐fusion structure of gB shows that it is a canonical Class III fusion protein;27 and several synthetic peptides derived from gB induce the fusion of large unilamellar vesicles and inhibit herpes virus infection.33, 35, 36
Irrespective of their structural differences, the three classes of fusion proteins seem to induce membrane fusion by essentially the same generic mechanism and a common refolding pathway is highly suggestive for the conservation of several phases of the process. The key component of the fusion machinery is the fusion peptide, loop, or patches of sequences which are projected toward the foremost side of the fusion glycoproteins where they are able to insert into cell membranes and facilitate the fusion process.
2.2. Non‐enveloped virus entry
Non‐enveloped viruses exploit a different mechanism for entry because they lack a membrane which surrounds the protein capsid; as a consequence they require capsid‐dependent mechanisms for penetrating the cell membrane or for exiting from the endosome.37, 38 The entry process is much less known and involves a series of triggers producing conformational and structural rearrangements which end in the exposure and/or release of lytic factors. Low pH, receptor interactions, protease cleavage, chaperone‐assisted morphological changes, divalent cation chelation, or any combination of these factors which take place at the appropriate site of membrane penetration are essential triggers to produce the activated viral intermediate.39
The mechanism of membrane disruption involves proteolytic cleavage of the coat protein and programmed exposure/release of small membrane‐lytic peptides. In this scenario, viral penetration is mediated by short, membrane interacting, amphipathic and/or hydrophobic sequences present in proteins undergoing a conformational modification which allows the exposure of these domains and their interactions with membranes.40 Similarly to enveloped viruses, capsid proteins seem to be trapped in a metastable state waiting for a trigger to expose their membrane active peptides and the membrane penetrating ability of these sequences is fundamental for entry but at the same time their premature exposure has to be avoided before host cells provide the triggers.
Flock House virus (FHV) is one of the simplest and most studied non‐enveloped viruses. The infection starts with the binding to one or more host cell receptors leading to a receptor‐mediated endocytosis mechanism.41 The receptor binding induces a conformational change that initiates uncoating or viral disassembly once the particle is exposed to low pH within the endocytic pathway blocking this event often inhibits infection. The immature provirion FHV is initially assembled from 180 copies of the coat protein alpha and, subsequently, it undergoes autoproteolytic cleavage to generate the mature infectious virion which contains a large N‐terminal fragment, β (363 amino acids), and a small C‐terminal fragment, γ (44 amino acids).41 The capsid shell is constituted by the central region of β forms, while the γ peptides are the amphipathic helices non‐covalently associated with the capsid interior. The γ peptides are responsible of the interaction with the membrane and its local disruption which ends in viral entry. These peptides are composed of an N‐terminal amphipathic helix separated by a proline–glycine–proline turn from a hydrophobic C‐terminal region. The amphipathic γ helices are located in the interior of the capsid and when FHV enters cells through receptor mediated endocytosis, the γ peptides are exposed and determine the disruption of the membrane with consequent release of the viral genome in the cytosol.41 This is a dynamic process with γ peptides continuously, transiently, and reversibly exposed to the exterior of the capsid; γ peptides may not only be exposed but also released from the virus particle which may represent a common paradigm of non‐enveloped virus entry. In this dynamic process γ peptides may continuously “sample” the environment until they encounter the appropriate cellular trigger; at this point the virus undergoes an irreversible conformational change in which the γ amphipathic helices insert into the target membrane, allowing the viral RNA to enter the cytoplasm.40
3. MEMBRANOTROPIC PEPTIDES
3.1. Fusion peptides of enveloped viruses
Many studies are devoted to the comprehension of the mechanism of insertion of fusion peptides, loops, or patches into monolayers inducing nipple formation and curvature in the target cellular membrane through many synergic interactions. The insertion in one leaflet of a closed bilayer will cause the increase of the surface area of the leaflet and the formation of a spontaneous curvature, which is one of the driving forces to reduce the energetic barrier needed for the achievement of fusion.5 Fusion and/or membranotropic domains in viral fusion proteins contain aromatic residues which together with alanines and glycines6 contribute to the interaction with just one bilayer leaflet.
Many biophysical and structural techniques using synthetic analogues and model membranes have been used to determine physiologically relevant states during membrane partitioning.6, 42 Conformation, insertion depth and angle of insertion differ according to membrane lipid composition and peptide length and sequence. Insertion with an oblique orientation and the capability of changing conformation according to the environment is emerging as a common pattern of viral fusion peptides, although the most active membrane bound conformation for generating membrane destabilization may vary from virus to virus.
The two structural motif widely found in fusion proteins and able to produce membrane curvature are the amphipathic α‐helix and tilted peptides.6, 7 The segregation of hydrophobic and polar residues on the two opposite sides of the amphipathic α‐helix is responsible of the superficial insertion into the upper monolayer which modifies the packing of the lipid polar head groups and creates a positive curvature of the upper monolayer which eventually leads to a positive curvature of the whole membrane and triggers local fusion of the membrane leaflets, transient pore formation, cracks, and membrane fusion. The membrane curvatures necessary to achieve fusion is created by the insertion of a biologically relevant number of amphipathic helices. Similarly, also tilted peptides are characterized by an asymmetric distribution of hydrophobic residues, which again produces a modification of the organization of the membrane into which they insert.6
The membrane bound conformation of the influenza virus fusion peptide is characterized by the presence of a hairpin of two tightly packed, antiparallel α‐helices;43, 44 the asymmetric insertion is determined by the fact that hydrophobic residues insert into the proximal membrane leaflet and polar residues project outward. The precise angle of the kink between the two arms depends on the sequence, the pH, and the lipid environment and determines the functional features necessary for activity. As a matter of fact, the compact hairpin structure drives favorable insertion, while its expanded structures promote subsequent membrane destabilization.45
Canonical Class II and Class III fusion peptides correspond to loops which do not undergo conformational changes upon insertion into the target membrane; the interaction clearly involves few hydrophobic residues, and the insertion into the outer leaflet of the membrane is superficial and probably inadequate to destabilize membranes. Thus, a cooperative effect attained upon fusion activation is the only explanation for activity.6 The idea of a single fusion peptide being exclusively responsible for the membrane perturbing activity has been overwhelmed by evidences supporting the concerted action of different membranotropic peptides,46 which together with the canonical fusion peptide are involved in the modification of membrane curvature.46, 47, 48, 49, 50, 51
Many viral fusion proteins present an additional hydrophobic membrane proximal region at the intersection between the ectodomain and the transmembrane anchor (TM), the so‐called MPER (or pre‐TM);46 the unusual clustering of aromatic amino acid in these regions prompted the idea of their strong involvement in the fusion process. Indeed, peptides corresponding to the pre‐TM region partition into membrane interfaces and likely cooperate with fusion peptides and TM domains during apposition of membranes, enhancing the overall hydrophobicity of the environment and contributing to the distortion of the lipid membranes required for fusion.6 The pre‐TM of HSV‐1 gH interacts strongly with membranes;47 the pre‐TM of Gp47 of foamy virus induces fusion of model membranes;52 the aromatic domain of the glycoprotein S2 of severe acute respiratory syndrome virus (SARS) partitions into lipid membranes and perturbs their integrity;53 the pre‐TM region of the GP2 protein from Ebola promotes perturbations of membranes when in a helical structure.54
HSV fusion involves both gB and gH and several membranotropic sequences are present in both glycoproteins,48, 55 although the precise role played by each of these regions remains to be elucidated. The two fusion loops at the tip of the Domain II of gB constitute a structural subdomain with the hydrophobic amino acids forming a crest lined on both sides by charged residues;35 the concerted use of the two peptides produced a significant distortion of the target membrane bilayer, while when they were used separately they presented a lower membrane penetration.35 The two charged residues represent a novel feature of fusion peptides with the presence of hydrophilic residues on either side favoring insertion.35 Fusion of inner and outer monolayers is clearly involved but not formation of pores, indicating that bilayer perturbation not complemented by leakage is a typical feature of viral fusion peptides which violate the host membrane without compromising its integrity.35 Several gH peptides are able to interact with membranes and play a role in the process. Among these, gH625, represents a key achievement in grasping the role of hydrophobic viral peptides.55, 56, 57, 58 The peptide contains residues critical for interaction such as aromatic residues (tryptophan and tyrosines) which are known for their preferred location at the membrane interface and for their ability to facilitate oligomerization59 together with numerous hydrophobic residues (glycines, leucines, alanines) which are critical for membrane insertion;6 at the C‐terminus there is an arginine residue which is key for establishing peptide–lipid interactions. gH625 penetrates into membranes from its N‐terminal side, and assumes an amphipathic helical conformation.56 The N‐terminal histidine acts as a switch for triggering viral fusion and strongly enhances the fusion activity;56 the role of the histidine has been reported also for paramyxoviruses60 and togaviruses.61
Yao et al.62 in order to investigate how the fusion peptide (FP) and transmembrane domain (TMD) are positioned relative to each other in the post‐fusion state, expressed a chimeric peptide containing both the fusion peptide and the transmembrane domain connected by a Gly–Lys linker. They found that the two domains induce membrane curvature and transient dehydration and their assembly facilitates the transition of the membrane from hemifusion intermediates to the fusion pore.
Lipids also play a key role in this process, generating membrane curvature thanks to their physico‐chemical properties. Cholesterol is key as it selectively intercalates into the leaflet of the bilayer favoring its distortion without producing unfavorable hydrophobic/hydrophilic interactions.16 The secondary structure of the fusion peptide of HIV changes according to the cholesterol content in the membrane, being α‐helical in the absence of cholesterol, but shifting to a β conformation with the increase of cholesterol.63 Both the fully α‐helical and the fully β‐structured peptides are able to insert deeply inside the membrane, while the mixed secondary structures, present at intermediate cholesterol concentrations, are more superficially inserted into lipid bilayers and less effective in inducing membrane fusion.63 It is likely that different secondary structures and domains with different content of cholesterol might be involved in different stages of fusion.17 Probably, lipid phase discontinuities between liquid ordered and disordered domains containing cholesterol produce membrane defects with exposed hydrophobic surfaces favoring a deeper insertion of fusion peptides, and promoting membrane fusion.16
In conclusion, the clear view is that membrane fusion is a very complex process involving several domains of the fusion proteins which interact directly or indirectly with biological membranes, and contribute to the merging of the viral envelope and cell membrane.
3.2. Lytic peptides of non‐enveloped viruses
Lytic peptides have been identified in capsid proteins, as small, amphipathic and hydrophobic sequences responsible of the viral membrane breaching activity, similarly to fusion peptides present in enveloped viruses. The entry of non‐enveloped viruses is commonly started by a conformational change suggestive of the entry of enveloped viruses, which determines the release of viral components with membrane lytic activity. Lytic factors bind to the membrane, temporarily unsettle the integrity and deliver the viral particle across the membrane.4 Despite differences among non‐enveloped viruses, similar events characterize their entry: induced modifications of capsid proteins resulting in peptide exposure followed by outward projection of lytic peptides able to interact with host membranes and disrupt them, ensuing the delivery of the viral genome inside the host cell.
Although several lytic peptides have been identified and characterized, their mode of interaction with membranes remains mainly vague. Some peptides (as the lytic peptides of nodaviruses, picornaviruses, and reoviruses) can be generated by an autocatalytic cleavage step of a precursor, whereas others can be generated from the proteolytic activity of cellular enzymes.64, 65
Essentially we can classify lytic peptides in amphipathic α‐helices and myristoyl groups; or we can classify them according to their mechanism of membrane action in those causing transient modification of the cellular membrane, pore formation, and total disruption of the limiting membrane.4 Although being clearly different among themselves and with fusion peptides of enveloped viruses, they present notable similarities.
Incubation of HeLa cells with various peptides corresponding to the C terminus of the L2 protein of papillomavirus, determines the entrance of propidium iodide into cells.66 The incubation of the adenovirus internal protein VI with liposomes loaded with a fluorophore, caused the release of the entrapped fluorophore.67 Similarly, incubation of VP5* of rotavirus with liposomes entrapping a fluorophore caused its release, suggesting that VP5* is sufficient to perforate the lipid vesicles.68, 69 The membrane disrupting γ1 peptide of FHV also triggers the release of the fluorophore.70, 71 As for enveloped virus fusion peptides, the amphipathic α‐helix seems to be a key structural motif also for non‐enveloped viruses. The N‐terminal 21 amino acids of the FHV γ peptide form an amphipathic α‐helix which is commonly referred to as γ1,4 while the γ peptide comprises also a C‐terminal region which is commonly considered to play a supporting role for the correct positioning of γ1. The γ peptide assumes a random coil conformation in solution, while it adopts a kinked helical conformation in model membranes; similarly to other membranotropic peptides also viral lytic factors seem to be able to adopt a membrane‐active conformation when interacting with the lipid bilayer.38, 72 The γ1 peptide is able to spontaneously partition into lipid bilayers and increases the membrane permeability of liposomes;67, 69, 70, 71, 73 in particular, it mediates liposome lysis through the insertion only into the outer leaflet of the lipid bilayer, and locating parallel to the membrane surface, with the hydrophobic face of the helix packed against the membrane surface.70, 72 A concentration dependent membrane leakage process74 similar to other non‐enveloped viruses such as poliovirus VP475 and reovirus μ1N76 is observed.
Similarly to the influenza77 and HIV gp4178 fusion peptides from enveloped viruses, the amphipathic region of γ peptide presents a kinked helical structure in solution. The rigid, boomerang like structure assumed by the influenza fusion peptide in lipid environment is required to promote membrane fusion;79 in fact, abolishing the kink in the structure or making it flexible eliminates membrane fusion79 probably for the failure of the fusion peptide to insert deep into the lipid bilayer and pack against the hydrocarbon moieties. At neutral pH the γ peptide is able to cause membrane disruption; while at low pH it is only able to alter its location relative to the capsid, but does not increase its membrane interacting ability.74 Surprisingly, thanks to a kinked structure and a tight alignment of the hydrophobic residues on one side of the peptide at low pH similar to influenza virus, FHV γ peptide shows localized perturbation of lipid arrangements with no proof of pore formation.38, 70 While membrane destabilization requires the simple insertion of the fusion peptide into the outer leaflet of the lipid bilayer, leakage needs both a deeper insertion and an interaction between peptides inside the membrane. The amphipathic region of γ peptide oligomerizes in bilayers71 with a low content of cholesterol, which being more fluid would promote association between peptides.74 The presence of the γ C‐terminus is absolutely necessary for virus entry;80 as a matter of fact, truncations, or point mutations in the C‐terminal region of γ determine a disordering of the pentameric bundle formed by the N‐terminal amphipathic helices of γ in FHV particles and hamper in vitro and in vivo membrane lysis. The pentameric bundles constitute the viral “membrane attack module,” and an essential function of the γ C‐terminal region is to maintain this module in its correct conformation4 (Figure 3). The γ and γ1 peptides cause a similar localized disorder of the target bilayer and the presence of the C‐terminal hydrophobic helical region in full‐length γ make the peptide effective at concentrations achievable in the context of viral infections.
Figure 3.
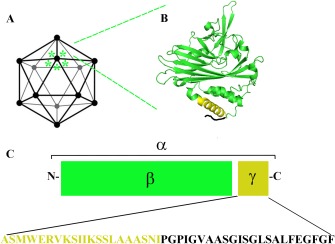
Schematic representation of FHV capsid (A). An expanded view of the crystallographic structure (pdb: 4FTB) of one subunit (α protein) showing the location of the amphipathic region of γ peptide in yellow (B). Schematic representation of α protein, which undergoes auto cleavage during maturation producing β and γ (C) with relative sequence of γ peptide
The kinked helical structure seems to be a common trait of enveloped and non‐enveloped viruses and differences in length and angularity of the helices as well as differences in the amino acid content may cause variations in the mechanism of interaction with the bilayer; the presence of this motif in viral proteins may signal a membrane associated role for this component during a certain step of the viral life cycle which adds to eventual other roles.
Adenovirus requires acidic pH for exposure of the amphipathic helix contained in its protein VI and disrupts endosomal membranes to release its nucleocapsid.67 Mutations in the amphipathic helix reduce infection and endosome escape, supporting the view that both the hydrophobic character and α‐helical structure are key to allow maximal membrane disruption. The N‐terminal amphipathic helix of protein VI, as FHV γ peptides, lies parallel to model membranes with the hydrophilic face interacting with the phospholipid head groups, and probably causes disruption of the bilayer by introducing positive curvature in membranes.73 Mutations in the amphipathic region of protein VI, which prevent insertion into the membranes, severely affects membrane penetration and cellular entry.81
Proteins VP1 and VP4 play a fundamental role in membrane penetration by poliovirus82 with the exposure of amphipathic α‐helical N‐terminal approximately 30 amino acid region of VP1 being necessary for liposome binding;83 while the N‐terminus of poliovirus VP4 contains a hydrophobic myristoyl (C14acyl) group for insertion into membranes.84 Following receptor binding, the amphipathic α‐helix within the N‐terminal portion of VP1, is exposed and probably forms pores to transfer the genome to the cytoplasm. Interestingly, VP4 also performs a role in membrane penetration and the hypothesis is that VP1 primarily function is to secure the particle to the limiting membrane, while VP4 participates directly in pore formation.
Mammalian orthoreovirus protein μ1 contains a small hydrophobic peptide (μ1N) with a myristoyl group at its N‐terminus.85 Disruption of cellular membranes, requires the complete dissociation of μ1N peptides from the particle; auto‐cleavage of μ1 during reovirus entry generates μ1N peptides that are linked to an N‐terminal myristoyl group. After lipid association, the μ1N peptide changes conformation from an extended to a β‐strand rich secondary structure.85 μ1N is able to generate size‐selective pores in erythrocyte or liposomal membranes. It is likely that the β‐hairpins in μ1N associate with the membranes forming a β‐barrel pore.85 Probably, membrane disruption caused by μ1N is similar to that of β‐barrel toxins.86
It is highly probable that a future more detailed knowledge of the mechanism used by lytic peptides of non‐enveloped viruses will lead to the discovery of more common features between enveloped and non‐enveloped membranotropic peptides.
4. APPLICATIONS OF MEMBRANOTROPIC PEPTIDES
Membranotropic peptides thanks to their adaptness to interact with the membrane, are opening the way for numerous applications.87, 88 Their ability to bind lipid membranes is correlated to their simultaneous hydrophobic and amphipathic nature, while their insertion into the bilayer is due to their capability to change conformation according to the environment; moreover, they are able to penetrate deep into the hydrophobic core but do not span the bilayer in a pore‐like manner; on the contrary, they tend to self‐associate at the interface between the membrane and the aqueous compartments.
Below are reported main applications.
4.1. Viral inhibitors
One of the main applications of fusion peptides of enveloped viruses is as inhibitors of viral penetration. Their ability to directly interact with the hydrophobic surfaces present on cell membranes and/or fusion proteins allows them to interfere with virus entry.89, 90 The inhibition mechanism is still unclear but it is likely that inhibition of infectivity is correlated to inactive aggregates formed between the fusogenic stretches present in the viral protein and in the peptides. In particular, the formation of aggregates is related to their ability to oligomerize or to mimic the mode of binding of their original domains in their partner protein; thus, stabilizing a pre‐fusion intermediate and preventing merging of the bilayers.55, 91, 92, 93
Self‐oligomerization of fusion peptides has been proposed to be responsible of inhibition by several groups.94, 95 HIV fusion peptides form structurally defined oligomeric complexes which have been considered responsible of inhibition;49, 96 moreover, mutants of the native sequence with a lower helical content and tendency to self‐associate into β‐sheets are able to inhibit membrane fusion with different magnitude and at various stages.97 VIRIP is a peptide designed to target gp41 fusion peptide and thus block HIV‐1 infection; it has undergone clinical studies and was demonstrated to be as active as peptides targeting the coiled‐coil.98, 99 Its clinical evaluation represents the proof of concept that membranotropic sequences could inhibit viral replication in infected individuals and may have potential clinical effectiveness. Moreover, the knowledge that several domains are implicated in the fusion mechanism and may interfere with the intramolecular interactions between the several domains, clearly demonstrates that they all represent potential targets for the design of entry inhibitors.100
4.2. Drug delivery vectors
Membranotropic peptides are emerging as delivery vectors.101, 102, 103 Until now, the most widely used delivery vectors are cationic cell penetrating peptides (CPPs), which enter essentially by endocytosis causing the entrapment of the cargo into endosomes with only minor quantities of the cargo able to reach the target where to exert the biological function. On the contrary, membranotropic peptides are internalized by direct penetration of the membrane and thus determine immediate bioavailability of the delivered molecule. Fusion membranotropic peptides are particularly noteworthy because they can physically interfere with the membrane hydrophobic interior forming bulges that protrude from the membrane and ease contacts between fusing bilayers; in particular, they are able to translocate molecules through the plasma membrane directly into the cell, promoting lipid–membrane reorganizing processes, and causing local and temporary membrane destabilization with subsequent reorganization, circumventing the endosomal entrapment by favoring the escape from the endosome.57, 104 The internalization mechanism is also related to the toxicity of the internalized drug and the development of resistance.
The gH625 is able to directly translocate across the membrane bilayer and to transport several cargos such as quantum dots,105 liposomes,106 dendrimers,107, 108 nanoparticles;109 it is also able to cross the BBB in vivo.110, 111
MPG is an amphipathic peptide composed of the fusion peptide of HIV‐1 associated to an hydrophilic domain with positively charged residues derived from the Nuclear Localization Sequence (NLS) of Simian virus 40 (SV40) large T antigen (PKKKRKV), through a spacer (WSQ).112, 113 In vitro MPG is able to deliver both siRNA and DNA after just 1 h.114 The principal internalization mechanism was shown to be independent of the endosomal pathway and to involve membrane disorganization and folding into β‐structures within the membrane bilayer without any associated leakage or toxicity.
The study of the internalization of γ peptide derived from FHV40 revealed that this is mediated by relatively high cell surface adsorption leading to enhanced macropinocytic uptake and cytosolic distribution and also revealed a higher efficiency of internalization compared with Tat.115
Influenza virus fusion peptide has been used for increasing transfection efficiency, its pH‐dependent fusogenic and endosomolytic activities are able to enhance lysosomal degradation before the contents of the endosomes are delivered to lysosomes.116
The development of industrial applications in drug delivery is probably one of the most exciting and fastest growing fields, with the possibility of these peptides to pass through the BBB and become an important player in the fight against all pathologies correlated to neurosciences.
4.3. Antibacterial compounds
Scientists envisage also a possible use of viral membranotropic peptides as an alternative to classical antibiotics in order to combat the antibiotic resistance problem.117 Antimicrobial peptides (AMPs) are widely exploited and represent attractive candidates for the development of anti‐infective agents;117, 118, 119 recently, attention has been devoted also to the exploitation of membranotropic peptides derived from viral fusion proteins as antibacterial drugs. In fact, some AMPs with helical structure seem to share high sequence (preference for alanines and glycines) and structure (amphipathic α‐helix) similarity with fusion peptides and suggest a convergent evolution correlated to their ability to disturb lipid bilayers.120 The fusion domain of influenza virus was evaluated for its antibacterial activity; analysis showed that the amidation of the C‐terminus is a key factor to render the fusion peptide an antibacterial peptide and optimization of the amphiphilic balance can improve efficacy.120, 121 The antibacterial activity of viral membranotropic peptides is not yet widely evaluated and much work is still open in this field; in particular, their mechanism of perturbation of membrane bilayers may allow the design of novel sequences with the ability to denature the membrane bilayer of bacteria which will add to their many roles.
4.4. Vaccines
Development of effective vaccines against viruses is another worldwide concern. A potent vaccine needs to be able to induce both humoral and cellular immunity. Few literature data have been reported on this subject and the possible use of viral membranotropic peptides for this aim. Recently, the MPER domain of HIV gp41 was used for vaccine development against HIV;122 in vivo prime‐boost immunization enhanced humoral and cellular immune responses, suggesting the promising application of membranotropic peptides as vaccine candidates in future (Table 1).
Table 1.
Examples of fusion proteins and peptides
| Enveloped viruses | |||
|---|---|---|---|
| Class | Characteristic features | Virus | Fusion peptide sequence |
| Class I |
• Trimeric in pre‐and post‐fusion conformation; • synthesized as inactive precursor, proteolytic cleavage required for fusogenic activity; • N‐terminal fusion peptide; • formation/extension of an α‐helical coiled‐coil propels the fusion peptide toward the target membrane; • postfusion conformation contains a heptad repeat‐derived six helix bundle core structure. |
Influenza | GLFGAIAGFIENGWEGMIDGWYG |
| HIV | AVGIGALFLGFLGAAGSTMGARS | ||
| Ebola | GAAIGLAWIPYFGPAAE | ||
| Class II |
• Dimeric and parallel to the viral envelope in pre‐fusion conformation, and trimeric post‐fusion conformation; • fusion peptides are loops buried in the dimer interface; • post‐fusion trimeric complexes. |
Dengue | DRGWGNGCGLFGKGSL |
| Simian foamy | VYTGVYPFMWGGAYCFCDS | ||
| Class III |
• Trimeric in pre‐ and post‐fusion conformation; • fusion loops positioned toward the viral envelope; • postfusion structure resembles six‐helix bundle of class I proteins, without heptad repeat domains |
Rhabdovirus G | WY/YA |
| Herpes virus gB | VWFGHRY/RVEAFHRY | ||
| Baculovirus gp64 | YAYNGGSLDPNTRV/VKRQNNNHFAHHTCNK | ||
| Non‐enveloped viruses | |||
| Peptide name | Virus | Lytic peptide sequence | |
| γ peptide | Flock house virus | ASMWERVKSIIKSSLAAASNIPGPIGVAASGISGLSALFEGFGF | |
| γ1 peptide | Flock house virus | ASMWERVKSIIKSSLAAASNI | |
| VP4 peptide | Poliovirus | MNMSRQGIFQTVGSGLDHILSLA | |
| μ1N peptide | Reovirus | MGNASSIVQTINVTGDGNVFKPSAETSSTAVPSLSLSPGMLN | |
5. CONCLUSION
The membrane entry of enveloped and non‐enveloped viruses employs fundamentally different mechanisms, although common themes have emerged in the entry process. This similarity is essentially represented by the presence/exposure of small membranotropic peptides which cause membrane disruption and/or promote membrane fusion. Entry involves membrane fusion versus perforation, but cellular triggering factors and structural intermediates appear to share some similarities. Interestingly there is also some similarity with the mechanism used by bacterial toxins to cross biological membranes in order to reach the cytosol; in fact, many toxins, undergo conformational changes which allow them to initiate the translocation process.86, 123
How exactly both enveloped and non‐enveloped viruses overcome host cell membrane barriers to deliver their genomes remains an intriguing problem. Comprehensive structural and biochemical studies on enveloped viruses have brought to the conclusion that a unifying mechanism for host cell entry exists; where a membranotropic fusion loop, peptide, or patches catalyze fusion of the two membranes. In contrast, interaction of non‐enveloped viruses with host cells during entry is less defined; while membrane active peptides have been discovered as necessary elements for entry in several well‐studied non‐enveloped virus capsids.
In conclusion, it is now evident that the success of membranotropic peptides further stimulates challenging research on the unraveling of the many roles and applications that could be developed for both enveloped virus fusion peptides and small lytic peptides in non‐enveloped viruses; membranotropic peptides are attracting increasing attention from the scientific community and their future will be dictated by the progresses in their industrial applications.
Biographies
Annarita Falanga received the Degree in Food Science and Technology at the University of Naples “Federico II”, in 2004. From 2004 to 2007 she attended the PhD course in Microbiological Science during which she was involved in the synthesis and characterization of fusogenic domains derived from membrane glycoproteins of Herpes Symplex Virus type I. From 2007 to 2012 she earned several Research Contracts at Consiglio Nazionale delle Ricerche (CNR) continuing her studies on peptides with antimicrobial activity. Her current research interests are in the development of stable formulation for radiopharmaceutical molecules.

Massimiliano Galdiero obtained his MD degree from the University of Naples ‘Federico II’ in 1992. He earned his PhD in Virology from the University of Cambridge (UK) in 1998. He was appointed lecturer in Microbiology in 1994 at the Department of Animal Health of the Faculty of Veterinary Medicine of the University of Naples ‘Federico II’. He moved to the Faculty of Medicine of the Second University of Naples where he was appointed associate professor and then full professor of Microbiology in 2004.

Giancarlo Morelli is a full professor of Chemistry at the University of Naples ‘Federico II’ (Napoli, Italy) and head of the Research Center on Bioactive Peptides. His research interest is devoted to the development of diagnostic and therapeutic agents based on bioactive peptides.

stefania galdiero received her PhD in Chemistry in the field of peptide chemistry at the University of Naples Federico II in 1998. She carried out research activities at the Columbia University of New York in 1996‐1997. In 1999 she earned the position of Assistant Professor of Inorganic Chemistry at the University of Naples and since 2015 she is Associate Professor. Since 2012, she is Adjunct Professor at Loyola University Chicago. Research activities focus on antimicrobial peptides and drug delivery mechanisms.

Falanga A, Galdiero M, Morelli G, Galdiero S. Membranotropic peptides mediating viral entry. Peptide Science. 2018;110:e24040. 10.1002/pep2.24040
REFERENCES
- 1. Yamauchi Y., Helenius A., J. Cell Sci. 2013, 126, 1289. [DOI] [PubMed] [Google Scholar]
- 2. White J. M., Whittaker G. R., Traffic 2016, 17, 593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Más V., Melero J. A., in Entry of Enveloped Viruses into Host Cells: Membrane Fusion (Ed: Mateu M. G.), Springer, Dordrecht, The Netherlands: 2013, 467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Moyer C. L., Nemerow G. R., Curr. Opin. Virol. 2011, 1, 44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Falanga A., Cantisani M., Pedone C., Galdiero S., Prot. Pept. Lett. 2009, 16, 751. [DOI] [PubMed] [Google Scholar]
- 6. Apellaniz B., Huarte N., Largo E., Nieva J. L., Chem. Phys. Lipids 2014, 181, 40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Harrison S. C., Virology 2015, 479–480, 498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Kalia M., Jameel S., Amino Acids 2011, 41, 1147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Kozlov M. M., Chernomordik L. V., Curr. Opin. Struct. Biol. 2015, 33, 61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Kozlov M. M., McMahon H. T., Chernomordik L. V., Trends Biochem. Sci. 2010, 35, 699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Podbilewicz B., Annu. Rev. Cell Dev. Biol. 2014, 30, 111. [DOI] [PubMed] [Google Scholar]
- 12. Lin L. T., Richardson C. D., Viruses 2016, 8, E250. 27657109 [Google Scholar]
- 13. Verrier E. R., Colpitts C. C., Sureau C., Baumert T. F., Hepatol. Int. 2016, 10, 567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Biswas S., Yin S.‐R., Blank P. S., Zimmerberg J., J. Gen. Physiol. 2008, 131, 503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Domanska M. K., Wrona D., Kasson P. M., Biophys. J. 2013, 105, 1383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Yang S.‐T., Kreutzberger A. J. B., Lee J., Kiessling V., Tamm L. K., Chem. Phys. Lipids 2016, 199, 136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Yang S. T., Kiessling V., Simmons J. A., White J. M., Tamm L. K., Nat. Chem. Biol. 2015, 11, 424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Plemper R. K., Curr. Opin. Virol. 2011, 1, 92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Munro J. B., Mothes W., J. Virol. 2015, 89, 5752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Chen L., Gorman J. J., McKimm‐Breschkin J., Lawrence L. J., Tulloch P. A., Smith B. J., Colman P. M., Lawrence M. C., Structure 2001, 9, 255. [DOI] [PubMed] [Google Scholar]
- 21. Yin H. S., Paterson R. G., Wen X., Lamb R. A., Jardetzky T. S., Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 9288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Lambert D. M., Barney S., Lambert A. L., Guthrie K., Medinas R., Davis D. E., Bucy T., Erickson J., Merutka G., S. R. Petteway, Jr. , Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 2186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Yi H. A., Fochtman B. C., Rizzo R. C., Jacobs A., Curr. HIV Res. 2016, 14, 283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Modis Y., Curr. Opin. Virol. 2014, 5, 34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Roche S., Bressanelli S., Rey F. A., Gaudin Y., Science 2006, 313, 187. [DOI] [PubMed] [Google Scholar]
- 26. Roche S., Rey F. A., Gaudin Y., Bressanelli S., Science 2007, 315, 843. [DOI] [PubMed] [Google Scholar]
- 27. Heldwein E. E., Lou H., Bender F. C., Cohen G. H., Eisenberg R. J., Harrison S. C., Science 2006, 313, 217. [DOI] [PubMed] [Google Scholar]
- 28. Backovic M., Longnecker R., Jardetzky T. S., Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 2880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Kadlec J., Loureiro S., Abrescia N. G., Stuart D. I., Jones I. M., Nat. Struct. Mol. Biol. 2008, 15, 1024. [DOI] [PubMed] [Google Scholar]
- 30. Cooper R., Heldwein E., Viruses 2015, 7, 6552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Matsuura H., Kirschner A. N., Longnecker R., Jardetzky T. S., Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 22641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Connolly S. A., Jackson J. O., Jardetzky T. S., Longnecker R., Nat. Rev. Microbiol. 2011, 9, 369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Galdiero S., Vitiello M., D'Isanto M., Falanga A., Collins C., Raieta K., Pedone C., Browne H., Galdiero M., J. Gen. Virol. 2006, 87, 1085. [DOI] [PubMed] [Google Scholar]
- 34. Chowdary T. K., Cairns T. M., Atanasiu D., Cohen G. H., Eisenberg R. J., Heldwein E. E., Nat. Struct. Mol. Biol. 2010, 17, 882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Falanga A., Tarallo R., Vitiello G., Vitiello M., Perillo E., Cantisani M., D'Errico G., Galdiero M., Galdiero S., PLoS One 2012, 7, e32186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Cantisani M., Falanga A., Incoronato N., Russo L., De Simone A., Morelli G., Berisio R., Galdiero M., Galdiero S., J. Med. Chem. 2013, 56, 8366. [DOI] [PubMed] [Google Scholar]
- 37. Bilkova E., Forstova J., Abrahamyan L., Viruses 2014, 6, 2899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Bajaj S., Dey D., Bhukar R., Kumar M., Banerjee M., J. Mol. Biol. 2016, 428, 3540. [DOI] [PubMed] [Google Scholar]
- 39. Suomalainen M., Greber U. F., Curr. Opin. Virol. 2013, 3, 27. [DOI] [PubMed] [Google Scholar]
- 40. Banerjee M., Johnson J. E., Curr. Prot. Pept. Sci. 2008, 9, 16. [DOI] [PubMed] [Google Scholar]
- 41. Odegard A., Banerjee M., Johnson J. E., Curr. Top. Microbiol. Immunol. 2010, 343, 1. [DOI] [PubMed] [Google Scholar]
- 42. Galdiero S., Falanga A., Cantisani M., Vitiello M., Morelli G., Galdiero M., Int. J. Mol. Sci. 2013, 14, 18758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Worch R., Acta Biochim. Pol. 2014, 61, 421. [PubMed] [Google Scholar]
- 44. Blijleven J. S., Boonstra S., Onck P. R., van der Giessen E., van Oijen A. M., Semin. Cell Dev. Biol. 2016, 60, 78. [DOI] [PubMed] [Google Scholar]
- 45. Larsson P., Kasson P. M., PLoS Comput. Biol. 2013, 9, e1002950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Peisajovich S. G., Shai Y., Biochim. Biophys. Acta 2003, 1614, 122. [DOI] [PubMed] [Google Scholar]
- 47. Galdiero S., Falanga A., Vitiello M., D'Isanto M., Collins C., Orrei V., Browne H., Pedone C., Galdiero M., Chembiochem 2007, 8, 885. [DOI] [PubMed] [Google Scholar]
- 48. Galdiero S., Vitiello M., D'Isanto M., Falanga A., Cantisani M., Browne H., Pedone C., Galdiero M., Chembiochem 2008, 9, 758. [DOI] [PubMed] [Google Scholar]
- 49. Kliger Y., Aharoni A., Rapaport D., Jones P., Blumenthal R., Shai Y., J. Biol. Chem. 1997, 272, 13496. [DOI] [PubMed] [Google Scholar]
- 50. Peisajovich S. G., Samuel O., Shai Y., J. Mol. Biol. 2000, 296, 1353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Samuel O., Shai Y., Biochemistry 2001, 40, 1340. [DOI] [PubMed] [Google Scholar]
- 52. Wang M., Zhang H., Liu Q. M., Sun Y., Li Z., Liu W. H., He X. H., Song J., Wang Y. X., Acta Virol. 2016, 60, 181. [DOI] [PubMed] [Google Scholar]
- 53. Mahajan M., Bhattacharjya S., Biochim. Biophys. Acta 2015, 1848, 721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Regula L. K., Harris R., Wang F., Higgins C. D., Koellhoffer J. F., Zhao Y., Chandran K., Gao J., Girvin M. E., Lai J. R., Biochemistry 2013, 52, 3393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Galdiero S., Falanga A., Vitiello M., D'Isanto M., Cantisani M., Kampanaraki A., Benedetti E., Browne H., Galdiero M., Peptides 2008, 29, 1461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Galdiero S., Falanga A., Morelli G., Galdiero M., Biochim. Biophys. Acta 2015, 1848, 16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Falanga A., Valiante S., Galdiero E., Franci G., Scudiero O., Morelli G., Galdiero S., Sci. Rep. 2017, 7, 9434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Galdiero S., Russo L., Falanga A., Cantisani M., Vitiello M., Fattorusso R., Malgieri G., Galdiero M., Isernia C., Biochemistry 2012, 51, 3121. [DOI] [PubMed] [Google Scholar]
- 59. Bakou M., Hille K., Kracklauer M., Spanopoulou A., Frost C. V., Malideli E., Yan L.‐M., Caporale A., Zacharias M., Kapurniotu A., J. Biol. Chem. 2017, 292, 14587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Krishnan A., Verma S. K., Mani P., Gupta R., Kundu S., Sarkar D. P., J. Virol. 2009, 83, 1727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Chanel‐Vos C., Kielian M., J. Virol. 2004, 78, 13543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Yao H., Lee M., Liao S. Y., Hong M., Biochemistry 2016, 55, 6787. [DOI] [PubMed] [Google Scholar]
- 63. Lai A. L., Moorthy A. E., Li Y., Tamm L. K., J. Mol. Biol. 2012, 418, 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Chevalier C., Galloux M., Pous J., Henry C., Denis J., Da Costa B., Navaza J., Lepault J., Delmas B., J. Virol. 2005, 79, 12253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Galloux M., Libersou S., Alves I. D., Marquant R., Salgado G. F., Rezaei H., Lepault J., Delmas B., Bouaziz S., Morellet N., J. Biol. Chem. 2010, 285, 19409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Kamper N., Day P. M., Nowak T., Selinka H. C., Florin L., Bolscher J., Hilbig L., Schiller J. T., Sapp M., J. Virol. 2006, 80, 759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Wiethoff C. M., Wodrich H., Gerace L., Nemerow G. R., J. Virol. 2005, 79, 1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Denisova E., Dowling W., LaMonica R., Shaw R., Scarlata S., Ruggeri F., Mackow E. R., J. Virol. 1999, 73, 3147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Dowling W., Denisova E., LaMonica R., Mackow E. R., J. Virol. 2000, 74, 6368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Bong D. T., Janshoff A., Steinem C., Ghadiri M. R., Biophys. J. 2000, 78, 839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Bong D. T., Steinem C., Janshoff A., Johnson J. E., Reza Ghadiri M., Chem. Biol. 1999, 6, 473. [DOI] [PubMed] [Google Scholar]
- 72. Janshoff A., Bong D. T., Steinem C., Johnson J. E., Ghadiri M. R., Biochemistry 1999, 38, 5328. [DOI] [PubMed] [Google Scholar]
- 73. Maier O., Galan D. L., Wodrich H., Wiethoff C. M., Virology 2010, 402, 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Maia L. F., Soares M. R., Valente A. P., Almeida F. C., Oliveira A. C., Gomes A. M., Freitas M. S., Schneemann A., Johnson J. E., Silva J. L., J. Biol. Chem. 2006, 281, 29278. [DOI] [PubMed] [Google Scholar]
- 75. Danthi P., Tosteson M., Li Q.‐H., Chow M., J. Virol. 2003, 77, 5266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Agosto M. A., Ivanovic T., Nibert M. L., Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 16496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Han X., Bushweller J. H., Cafiso D. S., Tamm L. K., Nat. Struct. Biol. 2001, 8, 715. [DOI] [PubMed] [Google Scholar]
- 78. Chang D. K., Cheng S. F., Chien W. J., J. Virol. 1997, 71, 6593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Lai A. L., Park H., White J. M., Tamm L. K., J. Biol. Chem. 2006, 281, 5760. [DOI] [PubMed] [Google Scholar]
- 80. Banerjee M., Khayat R., Walukiewicz H. E., Odegard A. L., Schneemann A., Johnson J. E., J. Virol. 2009, 83, 6929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Moyer C. L., Wiethoff C. M., Maier O., Smith J. G., Nemerow G. R., J. Virol. 2011, 85, 2631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Tosteson M. T., Wang H., Naumov A., Chow M., J. Gen. Virol. 2004, 85, 1581. [DOI] [PubMed] [Google Scholar]
- 83. Fricks C. E., Hogle J. M., J. Virol. 1990, 64, 1934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Chow M., Newman J. F., Filman D., Hogle J. M., Rowlands D. J., Brown F., Nature 1987, 327, 482. [DOI] [PubMed] [Google Scholar]
- 85. Zhang L., Agosto M. A., Ivanovic T., King D. S., Nibert M. L., Harrison S. C., J. Virol. 2009, 83, 7004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Galdiero S., Galdiero M., Pedone C., Curr. Prot. Pept. Sci. 2007, 8, 63. [DOI] [PubMed] [Google Scholar]
- 87. Ravi J., Ryadnov M. G., Biophysical Modulation of Peptide‐Membrane Interactions, The Royal Society of Chemistry, London: 2017, 188. [Google Scholar]
- 88. Falanga A., Galdiero M., Galdiero S., Int. J. Mol. Sci. 2015, 16, 25323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Falanga A., Tarallo R., Carberry T., Galdiero M., Weck M., Galdiero S., PLoS One 2014, 9, e112128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Badani H., Garry R. F., Wimley W. C., Biochim. Biophys. Acta (BBA) ‐ Biomembr. 2014, 1838, 2180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Rapaport D., Ovadia M., Shai Y., Embo J. 1995, 14, 5524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Franci G., Falanga A., Zannella C., Folliero V., Martora F., Galdiero M., Galdiero S., Galdiero M., J. Pept. Sci. 2017, 23, 311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Xu Y., Rahman N. A. B. D., Othman R., Hu P., Huang M., Prot.: Struct. Funct. Bioinform. 2012, 80, 2154. [DOI] [PubMed] [Google Scholar]
- 94. Chang D. K., Cheng S. F., Lin C. H., Kantchev E. B., Wu C. W., Biochim. Biophys. Acta 2005, 1712, 37. [DOI] [PubMed] [Google Scholar]
- 95. Lau W. L., Ege D. S., Lear J. D., Hammer D. A., DeGrado W. F., Biophys. J. 2004, 86, 272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Pritsker M., Jones P., Blumenthal R., Shai Y., Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 7287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Li Y., Han X., Tamm L. K., Biochemistry 2003, 42, 7245. [DOI] [PubMed] [Google Scholar]
- 98. Forssmann W. G., The Y. H., Stoll M., Adermann K., Albrecht U., Tillmann H. C., Barlos K., Busmann A., Canales‐Mayordomo A., Gimenez‐Gallego G., Hirsch J., Jimenez‐Barbero J., Meyer‐Olson D., Munch J., Perez‐Castells J., Standker L., Kirchhoff F., Schmidt R. E., Sci. Transl. Med. 2010, 2, 63re63. [DOI] [PubMed] [Google Scholar]
- 99. Münch J., Ständker L., Adermann K., Schulz A., Schindler M., Chinnadurai R., Pöhlmann S., Chaipan C., Biet T., Peters T., Meyer B., Wilhelm D., Lu H., Jing W., Jiang S., Forssmann W. G., Kirchhoff F., Cell 2007, 129, 263. [DOI] [PubMed] [Google Scholar]
- 100. Tarallo R., Carberry T. P., Falanga A., Vitiello M., Galdiero S., Galdiero M., Weck M., Int. J. Nanomed. 2013, 8, 521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Galdiero S., Falanga A., Vitiello M., Grieco P., Caraglia M., Morelli G., Galdiero M., J. Pept. Sci. 2014, 20, 468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Galdiero S., Vitiello M., Falanga A., Cantisani M., Incoronato N., Galdiero M., Curr. Drug Metab. 2012, 13, 93. [DOI] [PubMed] [Google Scholar]
- 103. Kalafatovic D., Giralt E., Molecules 2017, 22, 1929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Falanga A., Lombardi L., Tarallo R., Franci G., Perillo E., Palomba L., Galdiero M., Pontoni D., Fragneto G., Weck M., Galdiero S., RSC Adv. 2017, 7, 9106. [Google Scholar]
- 105. Falanga A., Vitiello M. T., Cantisani M., Tarallo R., Guarnieri D., Mignogna E., Netti P., Pedone C., Galdiero M., Galdiero S., Nanomedicine 2011, 7, 925. [DOI] [PubMed] [Google Scholar]
- 106. Perillo E., Allard‐Vannier E., Falanga A., Stiuso P., Vitiello M. T., Galdiero M., Galdiero S., Chourpa I., Int. J. Pharm. 2015, 488, 59. [DOI] [PubMed] [Google Scholar]
- 107. Borchmann D. E., Tarallo R., Avendano S., Falanga A., Carberry T. P., Galdiero S., Weck M., Macromolecules 2015, 48, 942. [Google Scholar]
- 108. Carberry T. P., Tarallo R., Falanga A., Finamore E., Galdiero M., Weck M., Galdiero S., Chemistry 2012, 18, 13678. [DOI] [PubMed] [Google Scholar]
- 109. Guarnieri D., Falanga A., Muscetti O., Tarallo R., Fusco S., Galdiero M., Galdiero S., Netti P. A., Small 2013, 9, 853. [DOI] [PubMed] [Google Scholar]
- 110. Valiante S., Falanga A., Cigliano L., Iachetta G., Busiello R. A., La Marca V., Galdiero M., Lombardi A., Galdiero S., Int. J. Nanomed. 2015, 10, 1885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Galdiero E., Falanga A., Siciliano A., Maselli V., Guida M., Carotenuto R., Tussellino M., Lombardi L., Benvenuto G., Galdiero S., Int. J. Nanomed. 2017, 12, 2717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Morris M. C., Chaloin L., Mery J., Heitz F., Divita G., Nucleic Acids Res. 1999, 27, 3510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Morris M. C., Vidal P., Chaloin L., Heitz F., Divita G., Nucleic Acids Res. 1997, 25, 2730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. Gallaher W. R., Cell 1987, 50, 327. [DOI] [PubMed] [Google Scholar]
- 115. Nakase I., Hirose H., Tanaka G., Tadokoro A., Kobayashi S., Takeuchi T., Futaki S., Mol. Ther. 2009, 17, 1868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Pichon C., Freulon I., Midoux P., Mayer R., Monsigny M., Roche A. C., Antisense Nucleic Acid Drug Dev. 1997, 7, 335. [DOI] [PubMed] [Google Scholar]
- 117. Galdiero S., Falanga A., Berisio R., Grieco P., Morelli G., Galdiero M., Curr. Med. Chem. 2015, 22, 1665. [DOI] [PubMed] [Google Scholar]
- 118. Falanga A., Lombardi L., Franci G., Vitiello M., Iovene M. R., Morelli G., Galdiero M., Galdiero S., Int. J. Mol. Sci. 2016, 17, 785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Maselli V., Siciliano A., Giorgio A., Falanga A., Galdiero S., Guida M., Fulgione D., Galdiero E., Environ. Pollut. 2017, 224, 597. [DOI] [PubMed] [Google Scholar]
- 120. Zhu S., Aumelas A., Gao B., J. Med. Chem. 2011, 54, 1091. [DOI] [PubMed] [Google Scholar]
- 121. Wang J., Zhong W., Lin D., Xia F., Wu W., Zhang H., Lv L., Liu S., He J., Chem. Biol. Drug Design 2015, 86, 487. [DOI] [PubMed] [Google Scholar]
- 122. Bolhassani A., Kardani K., Vahabpour R., Habibzadeh N., Aghasadeghi M. R., Sadat S. M., Agi E., Immunol. Lett. 2015, 168, 366. [DOI] [PubMed] [Google Scholar]
- 123. Galdiero S., Gouaux E., Prot. Sci. 2004, 13, 1503. [DOI] [PMC free article] [PubMed] [Google Scholar]