

Abstract
There are at least eight categories of cilia in the human body and malfunctioning of any one or several of them will have different consequences for the patient. A genetic error of the respiratory cilia (9 + 2) is the cause of the airways disease named immotile‐cilia syndrome (or PCD), whereas defective ependymal cilia (9 + 2) carries an increased risk of hydrocephalus. When the so‐called nodal cilia (9 + 0) of the early embryo are malfunctioning, there is a random determination of asymmetry of the heart and visceral organs (‘a 50% risk of situs inversus’). Some genes are responsible for the synthesis, transport, and assembly of the cilia, and mutations in these genes may lead to progressive degeneration of ciliary structures, such as the connecting cilium (9 + 0) of the photoreceptor cells—this is the cause of retinitis pigmentosa. Ciliary malfunctions due to genetic errors tend to be systemic and life‐long, whereas acquired diseases are local and may be temporary only. Copyright © 2004 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Keywords: immotile‐cilia syndrome, primary ciliary dyskinesia, situs inversus, hydrocephalus, infertility, retinitis pigmentosa
Introduction
The well‐known ‘9 + 2’ structure of a cilium needs no detailed presentation. Briefly, it is an assembly of nine microtubular doublets surrounding two central microtubular singlets. Each doublet has two rows of ‘dynein arms’, which contain the motor protein ‘dynein’. The dynein arms are able to walk on the back of the neighbouring doublet, which results in bending of the cilium. There are also so‐called nexin links, which are assumed to restrict the extent of sliding and to keep the array of doublets intact, and there are spokes (also called radial links) between the nine doublets and a ‘central sheath’, both probably with skeletal functions (Figure 1). The cilium emerges as an outgrowth from a basal body, which it resembles. The basal body is sometimes also named the centriole. It consists of nine microtubular triplets but has no central singlets.
Figure 1.
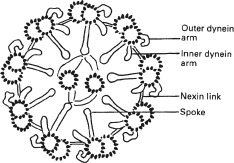
Diagram of a ciliary axoneme, ie the central structure of a cilium or of a sperm flagellum. The diameter of the axoneme is about 0.2 µm. Reprinted from Thorax, vol 53, BA Afzelius, Immotile cilia syndrome: past, present, and prospects for the future, pp 894–897, Copyright (1998), with permission from the British Medical Association
Diversity of cilia
There are at least eight categories of cilia or cilia‐derived organelles in the human body:
-
(1)
Mucus‐propelling cilia. These are motile, have a 9 + 2 structure with dynein arms, and a length of about 6 µm. Typically, a large number of cilia (200 or more) project from the upper surface of a ciliated cell. Their function is to propel a mucus blanket that rests on the ciliary tips. This is done with a to‐and‐fro movement in which cilia in their effective down‐stroke push the mucus blanket forward. The effected down‐stroke lasts a third to a fourth of the length of the beat cycle and moves at that stage with a speed of 1 mm/s. The so‐called respiratory cilia on the upper and lower airways are examples of mucus‐propelling cilia 1, 2.
-
(2)
Water‐propelling cilia. These have a 9 + 2 structure with dynein arms and are motile. Their length is about 10–15 µm. A ciliated cell has one, two, or many cilia. The cilia on the ependymal lining of the brain ventricles and those in the efferent ducts of the testis or in the oviduct are of this type.
-
(3)
Nodal cilia. These have a 9 + 0 structure with dynein arms and are motile. They are short, about 5 µm in length. There is only one cilium per cell and it moves with a vortical beating pattern, ‘like a propeller’ 3. They emerge and are active in the 7.5‐day post‐coitum embryo.
-
(4)
Monocilia. These have a 9 + 2 or a 9 + 0 structure, sometimes with dynein arms. They extend singly from the apex of a cell. In some cases, they appear on a cell that some days later will grow a number of mucus‐propelling cilia, as in the trachea 4, or grow water‐propelling cilia, as in the urethra 5; they are then called primary cilia. In other cases, they are constant features; the thyroid gland has cells with monocilia that extend into the follicles and that have been hypothesized to stir the colloid in the thyroid follicle 6. Another epithelium with monociliated cells is the corneal endothelium 7. Another name for them is ‘solitary cilia’.
-
(5)
Rudimentary cilia. These have a 9 + 0 structure without dynein arms and a length usually of about 2 or 3 µm. It is unlikely that they possess motility, also because they are often found cramped in solid tissues with no space for any movements. It is surprising to find single rudimentary cilia in the neurons and glia cells 8, chondrocytes 9, fibroblasts 10, etc. It has been suggested that their function is to move the centriole/basal body away from the central cytoplasm and thereby prevent the cell from dividing. Alternatively, they carry some receptor or they have no function.
-
(6)
Olfactory cilia. These have a short basal portion (about 1.5 µm) with a 9 + 2 structure, probably without dynein arms, and a long distal portion (maybe 50 µm) with only a few microtubular singlets 11. There are about 20 olfactory cilia on the apical portion of the olfactory cell. The olfactory cilia carry odour receptors in their cell membrane.
-
(7)
Photoreceptor cilia. These have a 9 + 0 structure without dynein arms and form a ‘connecting cilium’ between the inner and the outer rod segment (or between the inner and outer cone segments) and its continuation in the outer segment of the visual cell 12. The outer segment contains membrane disks with visual pigments.
-
(8)
Sperm flagellum. The sperm tail has a 45 µm long cilium with dynein arms. It moves by a meandering movement, a flagellar motility that differs from that of a cilium.
The terms ‘kinocilia’ and ‘stereocilia’ are used in the otological literature for some apical projections of the hair cells. The kinocilium is a regular monocilium, which in mammals is found only in the embryo and is lost in childhood. Whether it is motile at stage is not known with certainty. The term stereocilia is a misnomer, as they are long microvilli; the term stereovilli is recommended.
Cilia‐related diseases of genetic causes
Immotile‐cilia syndrome
Eukaryotic cilia and flagella, contrary to their prokaryotic counterpart, are very complex organelles, possibly because their ancestors lived as free‐living prokaryotes before they entered a host cell to become first a symbiont and then an organelle 13. With two‐dimensional chromatography, the 9 + 2 axoneme can be seen to consist of hundreds of polypeptide species 14. The number of essential genes that specify a healthy cilium may hence also be in the hundreds. It is therefore futile to search for the one gene that will cause the ciliary apparatus to be malfunctioning—there are probably hundreds of such genes, some only rarely mutated, others more often so.
The first patients to be diagnosed with a ciliary disease were some men in Denmark and Sweden, who produced spermatozoa that lacked dynein arms and were immotile (Figure 2) 15, 16. When the lungs from one of the men were examined, they were found to be unable to transport inhaled marker particles 17. The bronchial cilia were later seen to lack dynein arms 18. The disease was named ‘immotile‐cilia syndrome’ 19. Not surprisingly, the patients had chronic or recurrent rhinitis, sinusitis, bronchitis, and in younger days also otitis—thus, illness in organs with mucus‐transporting, ciliated epithelia. More surprising is the fact that these men were not greatly handicapped by their disease; they came to the hospital because of fertility problems.
Figure 2.
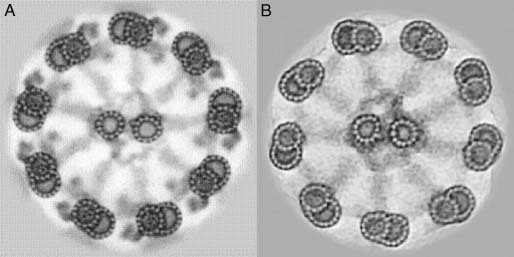
Computer averages of cross‐sectioned axonemes from a healthy man (A) and a man who suffers from the immotile‐cilia syndrome (B). The complete lack of dynein arms in axonemes from the man with the disease is striking. Reprinted from Tissue Cell, vol 27, BA Afzelius, R Dallai, S Lanzavecchia, PL Bellon, Flagellar structure in normal human spermatozoa and in spermatozoa that lack dynein arms, pp 241–247, Copyright (1995), with permission from Elsevier
Since that date, many more patients with the same clinical symptoms have been examined with respect to the motility or immotility of their nasal or bronchial cilia. Some persons were found to have immotile cilia; others had motile ones, although with a motility that was of insufficient strength or co‐ordination to be able to transport the mucus blanket in the normal fashion. The name ‘primary ciliary dyskinesia’, abbreviated PCD, was proposed for the syndrome. In a third category of people, no cilia were found at all and neither the term immotile‐cilia syndrome nor primary ciliary dyskinesia seemed suitable. For such cases, the term ‘acilia syndrome’ or ‘ciliary aplasia’ was suggested. It is, however, inconvenient to have separate names for each subgroup of the disease and the original term will be used. Thus, immotile‐cilia syndrome is considered here to be a group of diseases in which the respiratory cilia are immotile, dysmotile, or altogether missing.
Immotile‐cilia syndrome is clinically a uniform entity, but if examined by electron microscopy, it is seen to consist of a number of subgroups 20, 21, 22, 23. Further subgroups will undoubtedly be identified when suitable molecular genetic tools are applied in the search and identification of the genes responsible. The subgroups are listed in Table 1.
Table 1.
Overview of the molecular aberrations seen in different manifestations of the immotile‐cilia syndrome
| –No outer and inner dynein arms |
| –Few outer and inner dynein arms |
| –No outer dynein arms |
| –No inner dynein arms |
| –Short outer dynein arms |
| –No inner dynein arms and no spokes |
| –Short or no central microtubules |
| –No nexin links |
| –No or short spokes and eccentric central microtubules |
| –Basal bodies are present but have not grown any (or only a few) cilia |
| –No basal bodies and no cilia |
| –Cilia with a normal ultrastructure but unable to perform their work |
It is not always easy to see whether the electron micrographs show a normal amount of inner dynein arms and nexin links. Some measures can be taken to increase the reliability of the diagnosis. One is to add tannic acid to the glutaraldehyde solution when fixing the ciliated epithelia 24. This will enhance the contrast in the resulting electron micrographs and make diagnosis easier (Figure 2). Another method is to achieve an average imaging either of the circle of the nine microtubular doublets and their connecting links employing Markham's technique, or by performing a computer averaging in which not only the doublets of one cilium, but those of several cilia can be averaged 24.
It seems clear that electron microscopy—although ideal for the characterization of different subgroups of the syndrome—is not the perfect method to evaluate the functional capacity of ciliated epithelia. Several other diagnostic tools have been used, such as measuring the speed of mucociliary transport in situ by administrating an aerosol of small Teflon particles tagged with the short‐lived isotope 99Tc and measuring the location changes of the radioactivity in the lungs 17. A less time‐consuming (but also less accurate) method is to deposit small grains of saccharin or dye particles in the inferior concha and to measure the time required for the taste of the saccharin or for the dye particles to be visible in the throat 25.
These methods will not differentiate between genetically defective cilia and cilia with some acquired disorder—thus, between primary ciliary dyskinesia and secondary ciliary dyskinesia. The method by Jorissen et al is the only method that can make this very important distinction 26. In their method, they take a brush biopsy from the inferior concha of the nose and transfer it to a tissue culture vessel in which the medium is supplemented with pronase to make the individual cells separate from the epithelium and also supplemented with antibiotics to eliminate any bacterial growth. After about 1 day, the cilia are shed and after several more days, new cilia emerge. Healthy cilia are then recognized by their metachronal beats, which make the cell rotate in the culture medium, whereas cells that will not rotate are such that come from a person with the immotile‐cilia syndrome.
Situs inversus totalis
It was clear from the beginning that the immotile‐cilia syndrome is a disease in which both the respiratory cilia and the spermatozoa were defective. More surprising was the finding that nearly half the number of patients have situs inversus totalis (or rarely have isolated dextrocardia or isolated reversed viscera). If they have developed bronchiectasis, as is often the case, as well as sinusitis, they are said to suffer from the Kartagener syndrome 27.
The hypothesis was put forward that some embryonal cilia are responsible for positioning the heart anlage to the left body side (in normal cases), but that without motile cilia (in embryos with the immotile‐cilia syndrome) there is a random determination of the heart to the left or right 18. In a paper from 1998, Nonaka et al 3 showed that the 7.5 post‐coitum mouse embryo indeed carries cilia with the function to determine the sidedness of the forming heart and that these cilia are located in the closing neural tube; they were named nodal cilia. The nodal cilia create a leftward stream over the node of a fluid that presumably contains a morphogenic factor, which specifies the location of the forming heart. In mutants of mice, which lack such nodal cilia, the heart will be formed at the left or right side in equal proportion. In a second, important paper, Nonaka et al showed that the positioning of the heart can be determined at will by applying a motor‐driven stream of the extra‐embryonic fluid over the node 28. An ‘inheritance of randomness’ is a rare but not unique phenomenon; it also acts in the left‐ or right‐handedness 29, 30.
It is of interest in this connection that 10–15% of all cases with diagnosed immotile‐cilia syndrome have respiratory cilia that lack the two central microtubules; thus, they are of the 9 + 0 type rather than of the common 9 + 2. They are of two kinds: cilia with one of the nine peripheral doublets transposed from the outer circle to the interior (so‐called 8 + 1 cilia) 31 and those with just 9 + 0 (and some 9 + 2 cilia) 32. All patients with these two types of respiratory cilia have the normal sidedness of heart and viscera (situs solitus), whereas in the many other subgroups a 50% incidence of situs inversus is found.
The mutant mice investigated by Nonaka et al 3 have, as mentioned, a 50% prevalence of situs inversus but have healthy airways, brain ventricles, oviducts, spermatozoa, etc. The mutation is likely to affect one of the proteins needed for normal development of the nodal cilia, but not one engaged in the formation of respiratory cilia, spermatozoa, etc. In this respect, the mutant mice resemble those children, women, and men who have situs inversus but not the other features of the Kartagener syndrome. Isolated situs inversus has a prevalence in Scandinavia of about 1 in 10 000. It is not known what proportion of these persons have situs inversus for genetic reasons and how many for accidental reasons, thus as an acquired trait. Isolated situs inversus has no health consequences.
Male infertility
Most men who suffer from the immotile‐cilia syndrome have immotile spermatozoa as well as immotile cilia. The ultrastructure of the central machinery of the flagellum and that of the cilium are very similar, although the axoneme of the sperm flagellum usually appears more distinct in electron micrographs than does that of the cilium—the nexin links and the inner dynein arms are normally well discernible in the sperm axoneme but not always so in cilia.
Many proteins in the sperm flagellum are identical to those of their ciliary counterpart and depend on the same genes. A patient with 100% immotile spermatozoa of the 9 + 0 type was thus reported to have cilia that likewise were immotile and of the 9 + 0 kind 33. The spermatozoa from another man were completely immotile in spite of the fact that they had a perfectly normal ultrastructure with well visible dynein arms, spokes, etc. His cilia had the same normal ultrastructure and were immotile 34. The sperm flagella from a third man had no inner dynein arms, nor had his cilia 35.
Some investigators have reported cases where the patient had all the typical clinical symptoms of Kartagener syndrome, yet had ultrastructurally normal and motile spermatozoa as well as normal, motile cilia; in one case, the man was unable to become a father 36; in another case, the man was the father of two children 37. These two cases raise the question of the factors responsible for the patients' organ asymmetry and respiratory tract disease.
There are also reported cases of discordance between cilia and flagella: either a total or nearly total lack of dynein arms in the immotile sperm axonemes and a normal structure and motility of the examined (nasal) cilia 38, or the reverse situation in which the patients are reported to have normal lung functions but to produce either motile or immotile spermatozoa that make them infertile 39, 40. Such cases show that pairs of genes exist that code for different proteins with analogous functions, one in cells carrying mucus‐propelling cilia and another in spermatozoa.
Because spermatozoa with poor or no motility are unable to penetrate the isthmus border of the oviduct and the zona pellucida of the egg, the spermatozoa will not reach the egg cytoplasm. However, with modern techniques of reproduction biology, men with sperm immotility are not necessarily doomed to be infertile. In particular, the subzonal insemination technique and the ICSI (intracellular sperm injection) technique have been successfully applied in a few cases 41, 42.
As fertilization success in coitus also depends on the number of spermatozoa in the ejaculate, another factor is of fundamental importance, namely the free passage of spermatozoa from the testes via the efferent ductuli to the two deferent ducts and the ureter. The epithelium of the ductuli efferentes is partly ciliated and its cilia beat in the direction of the epididymis; presumably they assist in the transportation out from the testes of spermatozoa, which at this stage are still immotile. There are only very few functional or electron microscopic studies of these cilia in health or disease. Phillips et al 43 have, however, described the cilia from the efferent ductuli, which lacked dynein arms, in a patient with Kartagener syndrome who had normal motile spermatozoa with dynein arms. There was thus concordance between the axonemes of the water‐propelling cilia, of the mucus‐propelling ones, and by inference the nodal cilia, but discordance between these three categories of cilia and the sperm axoneme.
In another case report, the man had situs inversus, immotile respiratory cilia, chronic bronchitis, and only very few spermatozoa 44. He was interpreted by the authors as a case of Young syndrome (=respiratory tract disease and obstructive azoospermia), but he is equally well interpreted as a case of immotile‐cilia syndrome.
Female infertility or fertility
In 1983, the endometrial cilia from a woman with the classical Kartagener triad (situs inversus, bronchiectasis, and sinusitis) were described 45. An endometrial biopsy had been taken and revealed that its cilia lacked the inner and outer dynein arms. Nasal cilia were also defective; they were completely immotile. As the patient had not been able to become pregnant, it was suggested that whereas in normal cases the endometrial cilia are instrumental in transporting the egg along the oviduct, immotile endometrial cilia are incapable of such transport and the woman was infertile.
However, some years before that time, another case had been published: a woman also with Kartagener's triad and with endocervical cilia that lacked inner and outer dynein arms; however, she was the mother of two healthy children 46, 47. The authors concluded that ‘functional cilia in the endocervix are not essential for fertility’.
Also, in later studies, it is sometimes claimed that a woman with the complete Kartagener syndrome can be fertile 48 or sterile 49. Of the 17 women in Sweden with immotile‐cilia syndrome whom I have asked about their success or failure to become pregnant and who claim to have tried to become mothers for a minimum of 3 years, eight have children and nine have been unable to become pregnant. Hence female fertility is somewhat reduced when the cilia are immotile, dysmotile or missing, a fact that is not surprising considering the fact that the oviducts are ciliated and that egg transport is generally believed to be performed at least partially by these cilia.
Hydrocephalus
The ependyma of the brain ventricles is a ciliated epithelium and its cilia create a flow of cerebrospinal fluid in certain well‐defined directions. Those of the fourth ventricle, for instance, apparently propel cerebrospinal fluid from the aqueduct and from the plexus choroideus posteriorly towards the central canal and laterally towards the lateral apertures of the ventricle 50. Each ciliated cell carries an average of 16 cilia per cell and each cilium has a length of 13 µm 51. The cilia are described as having a uniform width rather than being tapering.
As these cilia a priori can be assumed to be of some importance, it was regarded of interest to find out whether patients with the immotile‐cilia syndrome have any abnormalities in their cerebral structure and function. In an early study, the brains of seven persons were examined, in this respect, by computer tomography, and in two or three cases, the ventricles were found to be slightly enlarged (with no consequences to their mental capacity) 52.
The team of Roth et al in Israel, who examined six persons with Kartagener syndrome, also by computer tomography, found no ventricular enlargement 53 and challenged this study. They concluded that the beat frequency of (nasal) cilia is not associated with brain abnormalities.
Yet, the great number of men, women, and, in particular, children who have been diagnosed with hydrocephalus (or with enlarged ventricles only) as well as with immotile, dysmotile, or lacking cilia make it reasonable to interpret these data as due to more than a coincidence 54, 55, 56, 57, 58, 59. In fact, slightly enlarged brain ventricles have been used as a prenatal sonographic marker of the immotile‐cilia syndrome, recommended when the fetus has either situs inversus or a sibling with the disease 59.
A mutation in the gene coding for the heavy chain of human axonemal dynein 5 (thus DNA5) has recently been identified in some patients with immotile‐cilia syndrome 58 and has been regarded as responsible for both immotile‐cilia syndrome and hydrocephalus. Whether this gene is the only culprit or the main one in these cases or just one of many genes that, when mutated, will have these consequences remains to be explored.
The water‐propelling cilia in man differ in some structural and functional respects from the mucus‐propelling ones. It is therefore conceivable that diseases exist which affect the water‐propelling cilia but not the mucus‐propelling ones. Evidently no such cases have yet been found. The water‐propelling cilia of the male efferent ducts, the female Fallopian tubes, and the brain ventricles all have water‐transporting ciliated epithelia but next to nothing is known of their ultrastructure in any cilia‐related genetic disease.
Anosmia
The olfactory receptor cells are ciliated, bipolar neurons. Each cell has a protruding olfactory knob carrying 8–30 olfactory cilia 60 that have a short basal segment and a long slender apical segment. The initial segment resembles an ordinary cilium, in that it has a 9 + 2 structure, although with no dynein arms and hence also with no capacity to beat. It is 1–2 µm long, whereas no data seem to be published for the length of the apical segment in man; maybe it is about 50 µm, as in several mammals 61, or as long as 200 µm, such as it is in the frog 62. The apical segment consists of a few microtubules surrounded by the cell membrane, in which the odour receptors are believed to be localized.
Patients with immotile‐cilia syndrome often complain of having a rather poor sense of smell, but that is explainable by their having a stuffed nose or a profuse nasal secretion. Usually they are not anosmic, however. It is possible that one subgroup or another of the immotile‐cilia syndrome contains or consists of anosmic persons—for instance, persons in the ciliary aplasia subgroup.
The olfactory epithelium and its disorders in one patient with congenital anosmia have been studied and published 63. Her receptor olfactory knobs were completely devoid of cilia, whereas they contained many centrioles and mitochondria; maybe this is a genetic disorder.
Retinitis pigmentosa
Once a sperm tail flagellum is synthesized and assembled, it remains permanent throughout its lifetime in epididymis and ejaculate; the spermatozoon has no repair mechanisms. The situation in the case of cilia is different. In particular, the components of the photoreceptor cilia of rods and cones have a turnover of the photopigment to an astonishing large extent: it has been estimated that 10% of the photoreceptor's outer segment (which is a modified cilium) is turned over every day 64. Large amounts of lipid and protein have to be synthesized in the inner segment of the rod and transported through the narrow connecting cilium to the outer segment, and equally large amounts will be transported from the outer segment to the inner one.
The protein complex responsible for the transport function has been identified and named the ‘intraflagellar transport protein’. There is evidence that this complex can move non‐membrane‐bound particles such assembled spokes from the cell body to the tip of the cilium, or transport the photoreceptor machinery in the case of rods and cones 65.
A defect in the intraflagellar transport protein seems to be the primary cause of some disease 66. If one of the polypeptides of the intraflagellar transport protein is mutated, the development of the outer segment of the rods will be abnormal and the retina will degenerate. It may lead to the particular form of gradual blindness that is named retinitis pigmentosa, which is one of the commonest causes of blindness. (Actually it is a group of diseases that are clinically identical but genetically different, just as is immotile‐cilia syndrome.) When combined with congenital deafness, it is called Usher syndrome.
It has long been noted that fertility is decreased in men and women with this disease 67. In a search for the cause of the blindness, the axonemal ultrastructure from nasal cilia 68 and spermatozoa 69, as well as from the photoreceptor cilium 70, 71, have been examined. Statistically significant, yet only rather small deviations—or in one case, no deviation 71—from the normal structure have been recorded. The dysfunction evidently resides in the intraflagellar transport protein rather than in the tracts of transport.
Some cases of retinitis pigmentosa accompanied by immotile‐cilia syndrome have been published 72, 73, 74. Whether it is a mere coincidence or is due to a defect in the mechanism that is common for the functioning of the two (or more) kinds of cilia remains to be explored.
Acquired disorders of cilia
The ciliated cells of our airways form a first defence line against various agents and conditions such as viruses, bacteria, noxious or irritant gases, anaesthetics including halothane and dibucain, freezing temperature, or excessive heat. The cilia are wonderfully resistant to many of these agents but will undergo changes under other conditions. The cilia may thus become immotile, dysmotile, be shed from the cell, or they may become flaccid, matted, clumped together, or harmed in numerous other ways. It is difficult to decide, from an inspection of the cilia by light microscopy or electron microscopy, whether a certain biopsy of the respiratory epithelium shows cilia with primary or secondary immotility, dyskinesia, or loss. It seems, however, that certain defects, such as a complete lack of dynein arms, are diagnostic for the immotile‐cilia syndrome.
When infecting the respiratory epithelium, the bacterium Pseudomonas aeruginosa will secrete a toxic substance named pyocyanin that among others will immobilize the tracheal cilia 75. If infected with Mycoplasma pneumoniae, Bordetella pertussis or B. bronchiseptica bacteria, a derangement of the so‐called ‘ciliary necklace’ (four or five rows of intramembrane particles at the neck region of the cilia) will be seen 76, 77, 78. This disruption leads to ciliary dyskinesia or stasis, presumably due to an interference with the axonemal calcium ion control rather than due to a defect of the axoneme itself. A differential diagnosis of the ciliary immotility—inborn or acquired—can be performed with the technique by Jorissen et al 26, as explained above.
When infected with various common cold viruses, the respiratory epithelium will react by shedding individual cilia as well as entire ciliated cells 79. Cilia may also become withdrawn into the cell body, as was seen in the nasal epithelium during an episode of coronavirus infection 80. A substantial loss of cilia is likely to result in rhinorrhoea. A reduction to about half the normal number of ciliated cells has been recorded from the tracheae of hamsters that were deprived of vitamin A since birth 81.
The cilia in the ependyma of the brain are not exposed to the same harsh environment as the tracheal cilia. However, they can be affected by vitamin A deficiency 81 in the same way as the respiratory cilia and can also be infected by viruses such as the mumps virus or various types of influenza virus 82, 83. Evidently the first step of the infection is replication of the virus in close vicinity to the cilia. This leads to a loss of most cilia—to a considerable thinning of the ciliary forest. As a consequence hereof, there are no cilia to prevent particular matter from settling down on the ependymal surface 84. The epithelial lining of the aqueduct is therefore destroyed and in nature's attempt to heal the breach, glial cells will cover the aqueduct, which results in stenosis and, as a consequence hereof, in ventricular enlargements and in hydrocephalus. Possibly neuroamidase released from invading neutrophils and macrophages play a role here 85. Alternatively, the disturbances of the infected ependymal cells may cause early ventricular dilatations to appear before an aqueduct stenosis has occurred 83.
Conclusion
In conclusion, the eight or more categories of human cilia have an invariant basic pattern—nine microtubular doublets in a ring—but differ in several details, such as length, motility, the presence or absence of dynein arms or of central microtubules. It seems likely that some gene products are needed for building or maintaining all of the categories, others restricted to a single category, and/or to two, three or several ciliary categories. Examples hereof are given above.
References
- 1. Sanderson MJ, Sleigh MA. Ciliary activity of cultured rabbit tracheal epithelium. J Cell Sci 1981; 47: 331–347. [DOI] [PubMed] [Google Scholar]
- 2. Jeffery PK, Reid L. New observations on rat airway epithelium. A quantitative and electron microscopic study. J Anat 1975; 120: 295–320. [PMC free article] [PubMed] [Google Scholar]
- 3. Nonaka S, Tanaka Y, Okada Y, et al. Randomization of left–right asymmetry due to loss of nodal cilia generating a leftward flow of extraembryonic fluid in mice. Cell 1998; 95: 829–837. [DOI] [PubMed] [Google Scholar]
- 4. Kober HJ. Die lumenseitige Oberfläche der Rattentrachea während Ontogenese. Z Mikrosk‐Anat Forsch 1975; 3: 399–409. [PubMed] [Google Scholar]
- 5. Odor DL, Blandau RJ. Observations on the solitary cilium of rabbit oviductal epithelium. Its motility and ultrastructure. Am J Anat 1985; 174: 437–453. [DOI] [PubMed] [Google Scholar]
- 6. Martin A, Hedinger C, Häberlin‐Jakob M, Walt H. Structure and motility of primary cilia in the follicular epithelium of the human thyroid. Virchows Arch B Cell Pathol Mol Pathol 1988; 55: 159–166. [DOI] [PubMed] [Google Scholar]
- 7. Svedbergh B, Bill A. Scanning electron microscopic studies of the corneal endothelium in man and monkeys. Acta Ophthalmol 1972; 50: 321–336. [DOI] [PubMed] [Google Scholar]
- 8. Dahl H. Fine structure of cilia in rat cerebral cortex. Z Zellforsch 1963; 60: 369–386. [DOI] [PubMed] [Google Scholar]
- 9. Wilsman NJ, Farnum CE, Reed‐Aksamit DK. Incidence and morphology of equine and murine chondrocytic cilia. Anat Rec 1980; 197: 355–361. [DOI] [PubMed] [Google Scholar]
- 10. Poole CA, Flint MH, Beaumont BW. Analysis of the morphology and function of primary cilia in connective tissue: a cellular cybernetic probe? Cell Motil 1985; 5: 175–194. [DOI] [PubMed] [Google Scholar]
- 11. Moran DT, Rowley JC, Jafek BW, Lovell MA. The fine structure of the olfactory mucosa in man. J Neurocytol 1982; 11: 721–746. [DOI] [PubMed] [Google Scholar]
- 12. Röhlich P. The sensory cilium of retinal rods is analogous to the transition zone of motile cilia. Cell Tissue Res 1975; 161: 421–430. [DOI] [PubMed] [Google Scholar]
- 13. Margulis L. Symbiosis in Cell Evolution (2nd edn). Freeman: New York, 1993; 213–303. [Google Scholar]
- 14. Luck D. Genetic and biochemical dissection of the eucaryotic flagellum. J Cell Biol 1984; 98: 789–794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Pedersen H, Rebbe H. Absence of arms in the axoneme of immobile human spermatozoa. Biol Reprod 1975; 12: 541–544. [DOI] [PubMed] [Google Scholar]
- 16. Afzelius BA, Eliasson R, Johnsen O, Lindholmer C. Lack of dynein arms in immotile human spermatozoa. J Cell Biol 1975; 66: 225–232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Camner P, Mossberg B, Afzelius BA. Evidence for congenitally nonfunctioning cilia in the tracheobronchial tract of two subjects. Am Rev Respir Dis 1975; 112: 807–809. [DOI] [PubMed] [Google Scholar]
- 18. Afzelius BA. A human syndrome caused by immotile cilia. Science 1976; 193: 316–319. [DOI] [PubMed] [Google Scholar]
- 19. Eliasson R, Mossberg B, Camner P, Afzelius BA. The immotile‐cilia syndrome. A congenital abnormality as an etiological factor in chronic airway infection and male sterility. N Engl J Med 1977; 297: 1–6. [DOI] [PubMed] [Google Scholar]
- 20. Afzelius BA. The immotile‐cilia syndrome: a microtubule‐associated defect. CRC Crit Rev Biochem 1985; 19: 63–87. [DOI] [PubMed] [Google Scholar]
- 21. Sturgess JM, Chao J, Wong J, Aspin N, Turner JAP. Cilia with defective radial spokes. N Engl J Med 1979; 300: 53–56. [DOI] [PubMed] [Google Scholar]
- 22. Stannard W, Rutman A, Wallis C, O'Callaghan C. Central microtubular agenesis causing primary ciliary dyskinesia. Am J Respir Crit Care Med 2004; 164: 634–637. [DOI] [PubMed] [Google Scholar]
- 23. Carlén B, Lindberg S, Stenram U. Absence of nexin links as a possible cause of primary ciliary dyskinesia. Ultrastr Pathol 2003; 27: 123–126. [DOI] [PubMed] [Google Scholar]
- 24. Afzelius BA, Dallai R, Lanzavecchia S, Bellon PL. Flagellar structure in normal human spermatozoa and in spermatozoa that lack dynein arms. Tissue Cell 1995; 27: 241–247. [DOI] [PubMed] [Google Scholar]
- 25. Andersen I, Proctor DF. Measurement of nasal mucociliary clearance. Eur J Respir Dis 1983; 64(Suppl 127): 37–40. [PubMed] [Google Scholar]
- 26. Jorissen M, Willems T, Van der Schueren B, Verbeken E, De Boeck K. Ultrastructural expression of primary ciliary dyskinesia after ciliogenesis in culture. Acta Oto‐Rhino‐Laryngol Belg 2000; 54: 343–356. [PubMed] [Google Scholar]
- 27. Kartagener M. Zur Pathogenese der Bronchektasien bei Situs viscerum inversus. Beitr Klin Tuberk 1933; 83: 489–501. [Google Scholar]
- 28. Nonaka S, Shirotori H, Saijoh Y, Hamada H. Determination of left–right pattern of the mouse embryo by artificial nodal flow. Nature 2002; 419: 96–99. [DOI] [PubMed] [Google Scholar]
- 29. Afzelius BA. Inheritance of randomness. Med Hypoth 1996; 47: 23–26. [DOI] [PubMed] [Google Scholar]
- 30. McManus C. Right Hand, Left Hand. Orion Book: London, 2002. [Google Scholar]
- 31. Sturgess JM, Chao J, Turner JAP. Transposition of ciliary microtubules. N Engl J Med 1980; 303: 318–322. [DOI] [PubMed] [Google Scholar]
- 32. Stannard W, Rutman A, Wallis C, O'Callaghan C. Central microtubular agenesis causing primary ciliary dyskinesia. Am J Respir Crit Care Med 2004; 169: 634–637. [DOI] [PubMed] [Google Scholar]
- 33. Neugebauer DC, Neuwinger Jockenhovel F, Nieschlag E. 9 + 0 axoneme in spermatozoa and some nasal cilia of a patient with totally immotile spermatozoa associated with thickend sheath and short midpiece. Hum Reprod 1990; 5: 981–986. [DOI] [PubMed] [Google Scholar]
- 34. Torikata C. The ciliary necklace—a transmission electron microscopic study using tannic acid‐containing fixation. J Ultrastr Mol Struct Res 1988; 101: 210–214. [DOI] [PubMed] [Google Scholar]
- 35. Toyama Y, Sumiya H, Fuse H, Shimazaki J. A case of an infertile man with short‐tailed spermatozoa. Andrologia 1996; 28: 81–87. [DOI] [PubMed] [Google Scholar]
- 36. Escudier E, Escalier D, Homasson J‐P, Pinchon MC, Bernaudin J‐F. Unexpectedly normal cilia and normal spermatozoa in an infertile man with Kartagener's syndrome. Eur J Respir Dis 1987; 70: 180–186. [PubMed] [Google Scholar]
- 37. Conraads VMA, Galdermans DJ, Kockx MM, Jacob WA, van Schaardenburg C, Coolen D. Ultrastructurally normal and motile spermatozoa in a fertile man with Kartagener's syndrome. Chest 1992; 102: 1616–1618. [DOI] [PubMed] [Google Scholar]
- 38. Walt H, Campana A, Balerna M, et al. Mosaicism of dynein in spermatozoa and cilia and fibrous sheath aberrations in an infertile man. Andrologia 1983; 14: 295–300. [DOI] [PubMed] [Google Scholar]
- 39. Jonsson MS, McCormick JR, Gillies CG, Gondos B. Kartagener's syndrome with mobile spermatozoa. N Engl J Med 1982; 307: 1131–1133. [DOI] [PubMed] [Google Scholar]
- 40. Wilton LJ, Teichtahl H, Temple‐Smith PD, DeKretser DM. Kartagener's syndrome with motile cilia and immotile spermatozoa: axonemal ultrastructure and function. Am Rev Respir Dis 1986; 134: 1233–1236. [DOI] [PubMed] [Google Scholar]
- 41. Wolf JP, Feneux D, Escalier D, Rodrigues D, Frydman R, Jouannet P. Pregnancy after subzonal insemination with spermatozoa lacking outer dynein arms. J Reprod Fertil 1993; 97: 487–492. [DOI] [PubMed] [Google Scholar]
- 42. Bongso TA, Sathananthan AH, Wong PC, et al. Human fertilization by micro‐injection of immotile spermatozoa. Hum Reprod 1989; 4: 175–179. [DOI] [PubMed] [Google Scholar]
- 43. Phillips DM, Jow WW, Goldstein M. Testis factors that may regulate gene expression: evidence from a patient with Kartagener's syndrome. J Androl 1995; 16: 158–162. [PubMed] [Google Scholar]
- 44. Smallman LA, Oates J, Proops DW. Young's syndrome (a case report). J Laryngol Otol 1988; 102: 460–463. [DOI] [PubMed] [Google Scholar]
- 45. Pedersen H. Absence of dynein arms in endometrial cilia: cause of infertility? Acta Obstet Gynecol Scand 1983; 62: 625–627. [DOI] [PubMed] [Google Scholar]
- 46. Bleau G, Richer C‐L, Bousquet D. Absence of dynein arms in cilia of endocervical cells in a fertile woman. Fertil Steril 1978; 30: 362–363. [DOI] [PubMed] [Google Scholar]
- 47. Jean Y, Langlais J, Roberts KD, Chapdelaine, Bleau G. Fertility of a woman with nonfunctional ciliated cells in the Fallopian tubes. Fertil Steril 1979; 31: 349–350. [DOI] [PubMed] [Google Scholar]
- 48. Eliyahu S, Shalev E. A fertile woman with Kartagener's syndrome and three consecutive pregnancies. Hum Reprod 1996; 11: 683–683. [DOI] [PubMed] [Google Scholar]
- 49. Lurie M, Tur‐Kaspa I, Weill S, Katz I, Rabinovici J, Goldenberg S. Ciliary ultrastructure of respiratory and Fallopian tube epithelium in a sterile women with Kartagener's syndrome. Chest 1989; 95: 578–581. [DOI] [PubMed] [Google Scholar]
- 50. Yamadori T. The directions of ciliary beat on the wall of the fourth ventricle in the mouse. Arch Histol Jpn 1977; 40: 283–296. [DOI] [PubMed] [Google Scholar]
- 51. Dempsey LC, Nielsen SL. Surface ultrastructure of human ependyma. J Neurosurg 1976; 45: 53–55. [DOI] [PubMed] [Google Scholar]
- 52. Afzelius BA. The immotile‐cilia syndrome and other ciliary diseases. Int Rev Exp Dis 1979; 19: 1–43. [PubMed] [Google Scholar]
- 53. Roth Y, Baum GL, Tadmor R. Brain dysfunction in primary ciliary dyskinesia. Acta Neurol Scand 1988; 78: 353–357. [DOI] [PubMed] [Google Scholar]
- 54. Jabourian Z, Zublin FD, Adler A, Gonzales C, Northrup B, Zwillingberg D. Hydrocephalus in Kartagener's syndrome. Ear Nose Throat J 1986; 65: 469–472. [PubMed] [Google Scholar]
- 55. De Santi MM, Magni A, Valletta EA, Gardi C, Lungarella G. Hydrocephalus, bronchiectasis and ciliary aplasia. Arch Dis Child 1990; 65: 543–544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Picco P, Leveratto L, Cama A, et al. Immotile cilia syndrome associated with hydrocephalus and precocious puberty. A case report. Eur J Pediatr Surg 1993; 3(Suppl 1): 20–21. [PubMed] [Google Scholar]
- 57. Al‐Shroef M, Karnik AM, Karnik AA, Longshore J. Ciliary dyskinesia associated with hydrocephalus and mental retardation in a Jordanian family. Mayo Clin Proc 2001; 76: 1219–1224. [DOI] [PubMed] [Google Scholar]
- 58. Ibanez‐Tallon I, Gorokhova S, Heintz N. Loss of function of axonemal dynein Mdnah5 causes primary ciliary dyskinesia and hydrocephalus. Hum Mol Genet 2002; 11: 715–721. [DOI] [PubMed] [Google Scholar]
- 59. Wessels MW, Den Hollander NS, Willems PJ. Mild fetal ventriculomegaly as a prenatal sonographic marker for Kartagener syndrome. Prenat Diagn 2003; 23: 239–242. [DOI] [PubMed] [Google Scholar]
- 60. Jafek BW. Ultrastructure of human nasal mucosa. Laryngoscope 1983; 93: 1576–1599. [DOI] [PubMed] [Google Scholar]
- 61. Seifert K. The ultrastructure of the olfactory epithelium in macrosmatics. An electron microscopic investigation. In Normale und Pathologische Anatomie, Bargmann W, Doers W (eds), Heft 21. Georg Thieme Verlag: Stuttgart, 1970; pp 1–10. [Google Scholar]
- 62. Reese TS. Olfactory cilia in the frog. J Cell Biol 1965; 25: 209–230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Douek E, Bannister LH, Dodson HC. Olfaction and its disorders. Proc R Soc Med 1975; 69: 467–470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Pazour GJ, Baker SA, Deane JA, et al. The intraflagellar transport protein, IFT88, is essential for vertebrate photoreceptor assembly and maintenance. J Cell Biol 2002; 157: 103–113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Quin H, Diener DR, Geimer S, Cole DG, Rosenbaum JL. Intraflagellar transport (IFT) cargo: IFT transports flagellar precursors to the tip and turnover products to the cell body. J Cell Biol 2004; 164: 255–266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Rosenbaum JL, Witman GB. Intraflagellar transport. Nature Rev Mol Cell Biol 2002; 3: 813–825. [DOI] [PubMed] [Google Scholar]
- 67. Hallgren B. Retinitis pigmentosa combined with congenital deafness. Acta Psychiatr Neurol Scand 1959; 34(Suppl 138): 1–101. [PubMed] [Google Scholar]
- 68. Arden GB, Fox B. Increased incidence of abnormal nasal cilia in patients with retinitis pigmentosa. Nature 1979; 279: 534–536. [DOI] [PubMed] [Google Scholar]
- 69. Finkelstein D, Reissig M, Kashima H, Massof R, Hillis A, Proctor D. Nasal cilia in retinitis pigmentosa. Birth Defects 1982; 18: 197–208. [PubMed] [Google Scholar]
- 70. Hunter DG, Fishman GA, Mehta RS, Kretzer FL. Abnormal sperm and photoreceptor axonemes in Usher's syndrome. Arch Ophthalmol 1986; 104: 385–389. [DOI] [PubMed] [Google Scholar]
- 71. Szczesny PJ. Retinitis pigmentosa and the question of photoreceptor connecting cilium defects. Graefes Arch Clin Exp Ophthalmol 1995; 233: 275–283. [DOI] [PubMed] [Google Scholar]
- 72. Ohga H, Suzuki T, Fujiwara H, Furutani A, Koga H. A case of immotile cilia syndrome accompanied by retinitis pigmentosa. Acta Soc Ophthalmol Jpn 1991; 95: 795–801. [PubMed] [Google Scholar]
- 73. Van Dorp DB, Wright AF, Carothers AD, Bleeker‐Wagemakers EM. A family with RP3 type of X‐linked retinitis pigmentosa: an association with ciliary abnormalities. Hum Genet 1992; 88: 331–334. [DOI] [PubMed] [Google Scholar]
- 74. Bonneau D, Raymond F, Kremer C, Klossek JM, Kaplan J, Patte F. Usher syndrome type‐1 associated with bronchiectasis and immotile nasal cilia in two brothers. J Med Genet 1993; 30: 253–255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Reimer A, Klementsson K, Ursing J, Wretlind B. The mucociliary activity of the respiratory tract. Acta Otolaryngol 1980; 90: 462–469. [DOI] [PubMed] [Google Scholar]
- 76. Carson JL, Collier AM, Hu DC. Ultrastructural observations on cellular and subcellular aspects of experimental Mycoplasma pneumoniae disease. Infect Immun 1980; 29: 11 117–11 124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Muse K. Host cell membrane perturbations during experimental Bordetella pertussis infection. Proc 7th Eur Conf El Micr Haag 1980; 2: 432–433. [Google Scholar]
- 78. Bemis DA, Wilson SA. Influence of potential virulence determinants on Bordetella bronchiseptica induces ciliostasis. Infect Immun 1985; 50: 3–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Rautiainen M, Nuutinen J, Kiukaanniemi H, Collan Y. Ultrastructural changes in human nasal cilia caused by the common cold and recovery of ciliated epithelium. Ann Otol Rhinol Laryngol 1992; 101: 982–987. [DOI] [PubMed] [Google Scholar]
- 80. Afzelius BA. Ultrastructure of human nasal epithelium during an episode of coronavirus infection. Virchows Arch 1994; 424: 295–300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. McDowell EM, Keenan KP, Huang M. Effects of vitamin A‐deprivation on hamster tracheal epithelium. Virchows Arch [Cell Pathol] 1984; 45: 197–219. [DOI] [PubMed] [Google Scholar]
- 82. Johnson RT. Hydrocephalus and viral infection. Dev Med Child Neurol 1975; 17: 807–816. [DOI] [PubMed] [Google Scholar]
- 83. Tanaka T, Mekata Y, Yamano T, Shimada M. Early ependymal changes in experimental hydrocephalus after mumps virus inoculation in hamsters. Acta Neuropathol (Berlin) 1993; 85: 521–525. [DOI] [PubMed] [Google Scholar]
- 84. Go KG, Stokroos I, Blaauw EH, Molenaar I. Changes in the ventricular ependyma and choroid plexus in experimental hydrocephalus as observed by scanning electron microscopy. Acta Neuropathol (Berlin) 1976; 34: 55–64. [DOI] [PubMed] [Google Scholar]
- 85. Grondona JM, Pérez‐Martin M, Cifuentes M, et al. Ependymal denudation, aqueductal obliteration and hydrocephalus after a single injection of neuraminidase into the lateral ventricle of adult rats. J Neuropathol Exp Neurol 1996; 55: 999–1008. [DOI] [PubMed] [Google Scholar]