

Abstract
Human noroviruses inflict a significant health burden on society and are responsible for approximately 699 million infections and over 200 000 estimated deaths worldwide each year. Yet despite significant research efforts, approved vaccines or antivirals to combat this pathogen are still lacking. Safe and effective antivirals are not available, particularly for chronically infected immunocompromised individuals, and for prophylactic applications to protect high‐risk and vulnerable populations in outbreak settings. Since the discovery of human norovirus in 1972, the lack of a cell culture system has hindered biological research and antiviral studies for many years. Recent breakthroughs in culturing human norovirus have been encouraging, however, further development and optimization of these novel methodologies are required to facilitate more robust replication levels, that will enable reliable serological and replication studies, as well as advances in antiviral development. In the last few years, considerable progress has been made toward the development of norovirus antivirals, inviting an updated review. This review focuses on potential therapeutics that have been reported since 2010, which were examined across at least two model systems used for studying human norovirus or its enzymes. In addition, we have placed emphasis on antiviral compounds with a defined chemical structure. We include a comprehensive outline of direct‐acting antivirals and offer a discussion of host‐modulating compounds, a rapidly expanding and promising area of antiviral research.
Keywords: antivirals, direct‐acting antivirals, host‐targeting drugs, norovirus, therapy
1. INTRODUCTION
1.1. Norovirus and disease manifestation
Human noroviruses are a predominant etiological agent of acute gastroenteritis worldwide, estimated to cause over one‐fifth of all gastroenteritis cases globally 1 and approximately 699 million infections per annum.2 Norovirus is responsible for over 200 000 estimated deaths each year,3 with the largest proportion occurring in children from lower income nations.3
Norovirus‐associated disease is usually self‐limiting, and symptoms such as diarrhea, vomiting, nausea, low‐grade fever, and abdominal cramps, usually resolve within two to four days in healthy adults.4 Human norovirus affects people of all ages and demographics worldwide,5 however, increased infection rates occur within high‐risk groups such as neonates, the elderly and the immunocompromised. Immunocompromised and transplant recipients would benefit from antivirals, as norovirus infection can persist for years in some cases 6, 7 and norovirus infection the transplant setting is a major health concern.
Transmission of human norovirus primarily occurs from person‐to‐person and also through contaminated food and water.8 Because norovirus is highly contagious with a low infectious dose (estimated to range between 18 and 1000 viral particles),9 outbreaks are particularly common in semi‐enclosed and closed settings such as nursing homes, hospitals, restaurants, cruise ships, social catered events, child‐care centers, within the military,10 and navy.11, 12 Prolonged shedding of norovirus post infection could also contribute to norovirus spread, and has been reported as a mechanism for nosocomial transmission.13 To reduce norovirus transmission, prophylactic antivirals to prevent large‐scale outbreaks are essential to protect high‐risk groups.
1.2. Social, health, and economic burden
In 2010 the World Health Organization's “Global estimates of the burden of foodborne disease” found that norovirus was the most common cause of death from foodborne gastroenteritis14 and was also responsible for 70 000 deaths in children under five years of age.3 These statistics were associated with lower income nations, where basic sanitation and wastewater infrastructure is often lacking; contributing to high levels of viral transmission.
This social burden is reflected by a substantial economic impact, with norovirus infection estimated to incur over USD $4.2 billion in direct annual health costs and $60.3 billion in societal costs worldwide, from factors such as forced business closures, hospitalizations, and days off work.3 Additionally, norovirus infections have been recognized as the fourth greatest cause of disability‐adjusted life years across the globe.14 These social and economic impacts justify the intensive research undertaken to identify effective antiviral and vaccine solutions for human norovirus.
1.3. Virus genome and classification
Human norovirus is classified within the Caliciviridae, a diverse family of positive‐sense RNA viruses, currently divided into five accepted genera including; Norovirus, Sapovirus, Vesivirus, Lagovirus, and Nebovirus,15 with several additional genera proposed.16, 17, 18, 19 The Norovirus genus is further classified into seven genogroups (GI‐GVII), with each genogroup containing numerous genotypes, based on capsid and polymerase protein coding sequence diversity.20 Recombinant viruses with different polymerase and capsid genotypes are common21 and many viruses detected today are recombinant in nature.22
Genogroup I (GI), GII, and GIV noroviruses can infect humans, with GII noroviruses responsible for approximately 80% to 90% of norovirus infections worldwide.23 In particular, the genogroup II, genotype 4 (GII.4) strains are recognized as causing the majority (~70%) of GII norovirus global cases and outbreaks24, 25, 26 and have historically been responsible for six reported pandemics. In temporal order they included; 1995 (US 95‐96 variant), 2002 (Farmington Hills 2002 variant), 2004 (Hunter 2004), 2006 (Den Haag 2006b), 2009 (New Orleans, 2009), and 2012 (Sydney 2012), respectively.27 While GII.4 noroviruses persist as the dominant strain in circulation worldwide, a number of viruses from other genotypes have emerged in recent years. For example, a sudden increase and high prevalence of the GII.17 strain in Asian countries occurred between 2014 and 2015,28, 29, 30 although the same high prevalence of this strain was not reflected in other parts of the world, with lower levels detected in Australasia, Europe, and North America compared to the Asian outbreaks during that same period.31, 32, 33
The human norovirus positive‐sense, single‐stranded RNA genome is 7.5 to 7.7 kb (Figure 1) and encapsidated within a nonenveloped, icosahedral 27 to 35 nm virion. The genome has three open reading frames (ORFs). ORF1 encodes a polyprotein that is posttranslationally cleaved into seven nonstructural proteins (p48 [NS1/2], NTPase [NS3], p22 [NS4], VPg (NS5], a viral protease [Pro, 3C‐like, NS6], and a viral RNA‐dependent RNA polymerase [RdRp, NS7]), by the virus‐encoded 3C‐like cysteine protease (3CLpro) (Figure 1) (reviewed in Atmar34; Karst35; Karst et al36). ORF2 and ORF3 encode the proteins VP1 and VP2, respectively; VP1 is the major capsid protein and VP2 is the minor capsid protein, likely involved in capsid assembly and genome encapsidation.37 The VP1 protein structure comprises the shell (S) and protruding (P) domains; the S domain encloses the viral RNA, while the antigenically variable P domain forms the outer surface of VP1, and is also involved in cell attachment.38, 39 The VP1 protein can be expressed in baculovirus which then self‐assembles into virus‐like particles (VLPs). These VLPs are antigenically and structurally indistinguishable to virions produced by the complete virus.40
Figure 1.

Schematic of the human norovirus genome. The norovirus genome is a positive‐sense, single‐stranded RNA genome comprising three ORFs that encode the nonstructural proteins: p48/N‐terminal (NS1/2), NTPase (NS3), p22 (NS4), VPg (NS5), protease (NS6), and RNA polymerase (NS7); and the structural proteins: VP1 and VP2. The numbers at the edges of each domain indicate nucleotide positions. Genome illustration is based on the norovirus GII.4 Sydney 2012 sequence (GenBank accession number JX459908). ORF, open reading frames [Color figure can be viewed at wileyonlinelibrary.com]
1.4. Models for studying norovirus infection
Despite the clinical significance of norovirus infection, antiviral studies have been hindered, because until recently, human norovirus could not be successfully propagated in cell culture. Recent breakthroughs have enabled human norovirus to be cultured in B cells41 and intestinal enteroids,42 which represent milestones in the field of norovirus biology. However, the modest replication levels generated by these new systems (≤3.5 log increase in B cells41, 43 and ≤3.8 log increase in enteroids42) means that they require optimization before widespread use for antiviral screening and development.
The GI.1 (Norwalk virus) norovirus replicon system has been used to assess antiviral candidates against the human virus in lieu of a viral culture system (Figure 2). The Norwalk replicon consists of an intact ORF1, ORF3, and genomic 3′ end, however, ORF2 is disrupted by a neomycin gene, engineered into the VP1‐encoding region. As such, while the subgenomic promoter is preserved, the expression of an intact VP1 is disrupted. Self‐replicating and stably expressed in Huh‐7 cells or BHK‐21 cell lines,44 the Norwalk replicon has proven itself as a useful tool to screen potential antiviral compounds (Table 1). However, replication levels of the Norwalk replicon are relatively low (approximately 1 × 103 Norwalk RNA copies per cell44), when compared to other replicon systems such as those for hepatitis C virus (HCV). The HCV replicons typically yield more than 1 × 104 copies per cell (AA Eltahla, 2018 personal communication), and have been used to identify many successful antiviral candidates. Moreover, they are amenable to high‐throughput antiviral screening (reviewed in Horscroft et al45), which has directly led to many of the direct‐acting antivirals (DAAs) used to treat HCV infections today (reviewed in Asselah et al46).
Figure 2.
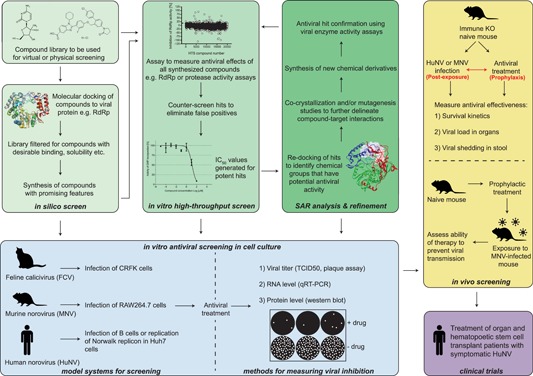
Current methods for the identification and characterization of norovirus antivirals. A flow chart depicting the methods and tools available for assessing the effectiveness of norovirus antivirals. Panels in green involve a combination of in silico and in vitro methods. Panels in blue and yellow represent in vitro and in vivo methods, respectively. The purple panel represents clinical testing in human patients. CRFK, Crandell Rees feline kidney; IC50, half maximal inhibitory concentration; qRT‐PCR, quantitative reverse‐transcription polymerase chain reaction; RdRp: RNA‐dependent RNA polymerase; SAR, structure‐activity relationship; TCID50, tissue culture infective dose [Color figure can be viewed at wileyonlinelibrary.com]
Table 1.
Antiviral compounds within this review with known structures that have been examined in more than one system

|

|

|

|

|
Abbreviation: FCV, feline calicivirus; HuNV, human norovirus; mAb, monoclonal antibody; MNV, murine norovirus; Mw, molecular weight (g/mol); Nb: nanobody; ^, denotes a representative from a series of compounds that displays the highest level of potency.
This article is being made freely available through PubMed Central as part of the COVID-19 public health emergency response. It can be used for unrestricted research re-use and analysis in any form or by any means with acknowledgement of the original source, for the duration of the public health emergency.
Other models have also been used to study antiviral efficacy against norovirus (Figure 2). For example, researchers have used in vitro enzyme activity assays, such as viral polymerase assays47 and protease assays48 (Figure 2) to screen compounds for antiviral activity in a high‐throughput format.49 In addition, X‐ray crystallography and in silico modeling can be used to examine ligand and viral protein interactions, to further elucidate antiviral mechanisms.50
Surrogate viruses from within the Caliciviridae family have also been exploited to screen for inhibitory activity of antiviral candidates across several calicivirus genera (Figure 2). These surrogates include murine norovirus (MNV; Norovirus), feline calicivirus (FCV; Vesivirus), porcine sapovirus (Sapovirus), rabbit hemorrhagic disease virus (RHDV; Lagovirus), and Tulane virus, from the proposed genus Recovirus.51 MNV, in particular, has been used as the predominant human norovirus surrogate as it is classified within the same genus and can be robustly propagated in cell culture,52, 53 making it amenable to viral disinfection and sterilization studies.54, 55, 56 Additional features that make MNV desirable for antiviral screening include its ability to be manipulated through reverse genetics, whilst in vivo studies in mice of many genetic backgrounds are straightforward.57, 58
Various animal models have also been used for human norovirus challenge studies and include: nonhuman primates such as chimpanzees, macaques, marmosets, and tamarins.59, 60, 61 Additionally, gnotobiotic or miniature pigs and gnotobiotic calves62, 63, 64, 65, 66 have also been used, as well as knockout and humanized mice for norovirus infection studies.67, 68, 69 However, no animal model has been deemed entirely suitable, due to obvious differences in clinical disease, gut physiology, microbiomes, naivety to norovirus infection compared to human populations, and the low genetic diversity of laboratory test animals.36
1.5. Viral replication cycle and antiviral targets
Every stage of the human norovirus replication cycle represents a unique target for antiviral development (Figure 3). However, the development of antiviral therapies that target viral replication requires a detailed understanding of norovirus biology and viral gene functionality, much of which is still to be elucidated. An overview of the human norovirus lifecycle and the specific antiviral targets are outlined in Figure 3. The stages of the replication cycle for antiviral targeting include: host cell attachment, internalization, genome release, viral genome replication mediated by the viral RdRp, translation of the genomic and subgenomic templates using the VPg and host cell machinery, viral protease cleavage of the viral polyprotein to yield mature viral proteins, followed by assembly, packaging, and cell egress (reviewed in Thorne and Goodfellow70).
Figure 3.
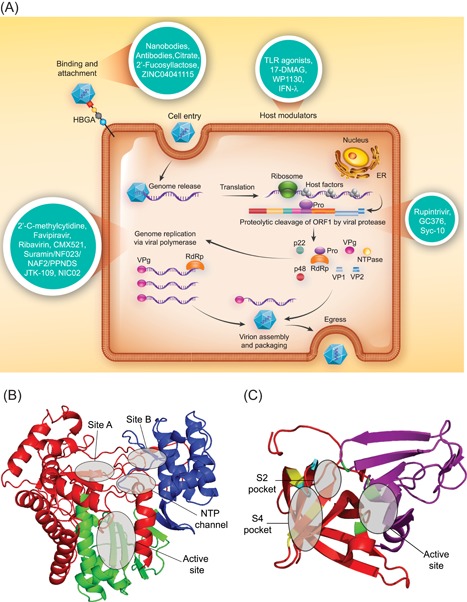
The replication cycle and antiviral targets for human norovirus. A, A schematic of the complete norovirus replication cycle is presented and antivirals that have been developed against norovirus are depicted in turquoise circles. The listed compounds represent only those antivirals which have a known target and a more extensive list of antivirals has been described in Table 1. B, A ribbon diagram of the human norovirus GII.4 Sydney 2012 RdRp with fingers (red), thumb (blue), and palm (green) domains color‐coded. The NNI binding sites (Site A, Site B, and the NTP channel) and the NA binding site (active site and Motif C) are labeled. C, A ribbon diagram of the GI.1 Norwalk virus protease (2FYQ) with N‐ and C‐terminal chains colored in magenta and red, respectively. Residues of the catalytic triad are colored in green. Residues for the S2 (cyan) and S4 (yellow) pockets 145 are shown as representative binding sites for protease inhibitors. NTP, nucleoside triphosphate [Color figure can be viewed at wileyonlinelibrary.com]
1.6. A review of norovirus antiviral therapies
While a number of other norovirus antiviral reports have been published in the last three years,71, 72, 73, 74, 75 there has been significant recent progress in the field, which now warrants an updated review. Herein we include a comprehensive overview of peer‐reviewed studies since 2010, including antiviral candidates examined in at least two different systems (e.g. viral enzyme inhibition assays and MNV cell culture). It should be noted that plant or food extracts that have antinorovirus effects have been omitted from this study when the active antiviral compound is unknown, with this review focusing on compounds that have a defined structure (as outlined in Table 1). We include studies using the recently developed human norovirus cell culture system in B cells.41 We also discuss recently identified broad‐spectrum antivirals with antinorovirus activity and provide a more comprehensive overview of host‐modulating compounds, which are a recent novel and exciting area within norovirus antiviral research (Table 1). Table 1 lists the individual antiviral compounds, or in the case where several very similar derivatives have been studied, it lists the most potent compound of the group as a representative derivative, with relevant citations. Each compound is indexed for clarity throughout the manuscript and in Table 1.
2. DRUGS WITH KNOWN TARGETS
2.1. Attachment and entry
Cell attachment and entry are features of the viral replication cycle that have been extensively investigated as antiviral targets for human immunodeficiency virus (HIV), dengue virus (DENV), and HCV,76 amongst others. To design compounds capable of targeting viral entry, knowledge of the viral entry receptor/s is usually required. Recently, CD300lf was identified as the entry receptor for MNV, however, the counterpart receptor for human norovirus has yet to be discovered.77, 78 As such, there is an absence of antivirals that target norovirus entry and a bias toward compounds that prevent norovirus cellular attachment.
The most widely studied attachment targets are the histo‐blood group antigens (HBGAs). HBGAs are complex carbohydrates that are presented abundantly on the surface of mucosal epithelia of the gastrointestinal tract. They interact with the surface P2 domain of the VP1 capsid protein79, 80 and are thought to aid viral attachment, and perhaps even entry.81, 82, 83 HBGAs are determined by host cell genetics (reviewed in Tan and Jiang82) and play an important role in terms of both virus and host interactions, and for noroviruses to initiate infection.81, 84
In one study, in silico screening was performed on a drug library (>2 million compounds) to identify molecules that interact strongly with HBGAs and prevent norovirus attachment.85 Potential hits (n = 160) were confirmed in vitro with enzyme‐linked immunosorbent assay (ELISA)‐based human norovirus VP1/HBGA blocking assays, which revealed that compounds with a cyclopenta‐a‐dimethyl phenanthrene base structure (n = 4) display potent inhibition (IC50 < 10 μM) of HBGA‐norovirus binding.85 The most potent compound in this series was ZINC04041115 [2.1.1] with an approximate EC50 of 2.5 μM.85 Studies by others using ELISA‐based VLP‐binding assays, have shown that the human milk oligosaccharides (HMOs) 2′‐fucosyllactose (2′FL) [2.1.2] and 3′‐fucosyllactose (3′ FL) inhibit the binding of GII.10 norovirus VLPs to HBGAs at low millimolar concentrations (5.5‐30.2 mM).86 2′FL was later shown to also inhibit binding of GII.17 and GI.1 VLPs to HBGAs with IC50 values ranging between 13 to 20 mM and 38 to 50 mM, respectively.87
In a similar fashion, citric acid [2.1.3] has been shown to bind norovirus GII.10 VLPs causing an altered morphology that prevents VLP‐HBGA binding.88 In addition, X‐ray crystallography has revealed that these HMOs interact directly with the P domain of the capsid, thus providing a clear mechanism for the binding inhibition observed.86, 87, 88 Despite these advances, attachment inhibitors have only been tested against VLPs from less common genotypes and thus their effectiveness against a broader spectrum of norovirus strains needs to be demonstrated.
Passive immunotherapy with monoclonal antibodies (mAbs) or nanobodies (Nbs) is an antiviral strategy to prevent norovirus attachment and entry, which would be particularly useful for immunocompromised patients. Therapeutic mAbs [2.1.4] generated from human and chimpanzee B cells have been described in several studies and were shown to block carbohydrate binding by norovirus VLPs.89, 90, 91 However, only mAb D8 was shown to provide protection against Norwalk virus infection in a chimpanzee model89 (Table 1). Despite this, mAbs generated thus far have limited cross‐genotypic activity, which makes them a less than optimal antiviral against the antigenically diverse norovirus.
More recently, Nbs have been shown to have broadly neutralizing activity against VLPs of important norovirus genotypes via interaction with the capsid P domain.92, 93 Nanobodies are recombinant, single‐domain, camelid heavy‐chain antibody fragments of ∼15 kDa which can bind with high affinity to a specific antigen (reviewed in Muyldermans94; Harmsen and De Haard95). The best‐described norovirus Nb is Nb‐85 [2.1.5] which has a broad binding capability against GII.1, GII.2, GII.4, GII.12, GII.17, and G1.11 VLPs, preventing attachment to HBGAs.92, 93 The cross‐genotypic activity displayed by Nbs illustrates that these molecules have the potential to overcome the narrow antigenic spectrum typically displayed by conventional mAbs. However, despite these findings, mAb and Nb studies have been based mostly on VLP‐binding and structural analysis of that binding (Table 1) and thus the effects of such compounds against norovirus in cell culture or in vivo need to be explored further before continued development toward clinical application.
2.2. Polymerase inhibitors
Of the human norovirus antiviral targets, the RdRp is one with numerous preclinical candidates identified that can inhibit its activity (Table 1). Critical for viral replication, the RdRp is a highly attractive antiviral target, as it largely lacks host homologs minimizing the chance of off‐target adverse effects.96 The human norovirus polymerase forms the canonical RdRp structure resembling a closed right hand, with fingers, palm, and thumb domains97 (Figure 3B), likely acting as a homodimer in it active state.97, 98, 99
The RdRp‐targeting antivirals are divided into two major classes; the nucleoside analogs (NAs) and the non‐nucleoside inhibitors (NNIs). NAs and/or NNIs have been successful in the treatment of HCV, HIV, herpesvirus and hepatitis B virus (HBV) infections (reviewed in Eltahla100; Zhang and Wang102; Fung et al103; Usachet al104), and candidates from both classes have been assessed for antiviral activity against norovirus within this review (outlined below).
2.2.1. Nucleoside analogs
NAs inhibit RNA synthesis through mimicry of incoming nucleoside triphosphates (NTPs), which upon incorporation subsequently cause chain termination,105 or less commonly, increase mutations during viral genome transcription that results in lethal mutagenesis (also known as error catastrophe).106 Since NAs bind in the highly conserved RdRp active site, they generally demonstrate broad‐spectrum antiviral activity compared to NNIs. Therefore, NAs developed against other RNA viruses have been examined for repurposing as human norovirus therapeutics. In terms of norovirus, 2′‐C‐methylcytidine (2CMC) [2.2.1] is the most intensely studied and was initially developed as an antiviral therapy against HCV. 2CMC is a chain terminating cytidine analog with broad‐spectrum in vitro activity against other flaviviruses including DENV, yellow fever virus, and West Nile virus.107 However, the development of the oral 2CMC prodrug, Valopicitabine, was halted for use against HCV following reports of undesirable gastrointestinal side effects.108
Despite 2CMC being discontinued for clinical development, it is still widely reported as a potential norovirus antiviral. In one study, MNV plaque formation and RNA synthesis were inhibited by 2CMC, with EC50 values of 2.0 µM and 1.6 µM, respectively.109 2CMC was also found to reduce RNA synthesis of the Norwalk replicon in a dose‐dependent fashion, with an EC50 of 18 µM.110 Another study reported a similar 2CMC potency for MNV RNA level reduction (6.9 µM), but potency against the Norwalk replicon was found to be 14‐fold higher (1.3 µM).111 This difference was proposed to be due to either varying methodology between studies, or differences in drug purity.110 In the human norovirus BJAB cell culture system, 2CMC inhibited human norovirus replication with an EC50 of 0.3 µM.67 Despite the variation in 2CMC potency observed between these in vitro systems, collectively these studies illustrate that the polymerase is an excellent antiviral target for cross‐genogroup inhibition of norovirus replication.
The NA 2CMC has also shown promise as a potential norovirus antiviral in mouse model studies. Knockout mice infected with MNV and treated with 2CMC were protected from mortality, diarrhea, and had reduced norovirus genome titers in tissues and stool (1.0‐1.5 log10 reduction), compared to mock‐treated animals.67, 110 Additionally, MNV‐infected mice treated with a high dose of 2CMC (100 mg·kg−1·day−1 for 5‐7 days) demonstrated reduced transmission to uninfected sentinel mice caged together, and offered prophylactic protection for up to 18 days.112
In an attempt to improve the safety and efficacy of 2CMC therapy, several derivatives have been examined for antinorovirus activity, for example, 2′‐F‐2′‐C‐methylcytidine (2FCMC). One study evaluated the inhibitory activity of 2CMC, 2FCMC, β‐d‐N(4)‐hydroxycytidine (NHC) and the HBV/HIV NA lamivudine against the replication of MNV and the Norwalk replicon.111 EC50 values for 2CMC and 2FCMC in MNV cell culture were 6.9 and 12.7 µM, respectively,111 while NHC and lamivudine had no inhibitory effect on replicon RNA synthesis. Indeed, both 2CMC and 2FCMC demonstrated dose‐ and time‐dependent inhibition of Norwalk replicon RNA levels, although 2CMC (EC50 1.3 µM) was found to be more than 2‐fold more potent than 2FCMC (EC50 3.2 µM).111 While the search continues for a safer derivative of 2CMC for the treatment of norovirus infection, no effective alternative has been identified thus far.
Favipiravir (T705; 6‐fluoro‐3‐hydroxy‐2‐pyrazinecarboxamide) [2.2.2] is another broad‐spectrum NA that has been assessed as a potential antiviral against human norovirus. T705 is a purine analog,113 which was shown to induce lethal mutagenesis against MNV,114 consistent with results from other viruses.115 T705 is approved for treatment of influenza in Japan116 and also inhibits replication of several viruses using in vitro and mouse models, including; flaviviruses, arenaviruses, hantaviruses,113 and Ebola virus.117 T705 has poor antiviral activity against MNV replication in cell culture, inhibiting virus‐induced cytopathic effects with an EC50 of 250 µM, and RNA synthesis with an EC50 of 124 µM,118 with a therapeutic index of just 4.3.118 The human norovirus replicon has also been used to examine the antiviral efficacy of T705, which exhibited a modest EC50 of 21 µM and a CC50 of more than 100 µM.119 Despite clinical approval for influenza, T705 displays a level of potency that is likely too low to be pursued further as a norovirus therapeutic.
Another broad‐spectrum NA examined against norovirus is ribavirin (RBV; 1‐α‐d‐ribofuranosyl‐1,2,4‐triazole‐3‐carboxamide) [2.2.3]. RBV has broad‐spectrum antiviral activity against both DNA and RNA viruses and has been used clinically to treat HCV, hepatitis E virus, Lassa fever, and respiratory syncytial virus, amongst others (reviewed in Snell120). A guanoside analog, RBV was first shown to inhibit the human norovirus replicon and MNV replication in 2007, with an EC50 of around 40 µM for Norwalk replicon and an 82% reduction of replicon genome levels at 100 µM compared to mock‐treated controls.121 As expected for an NA, the binding site of RBV was shown to be the polymerase active site by cocrystallization studies in the MNV RdRp.122 However, RBV poorly inhibits MNV (EC50 of 63.5 μM) as well as the Norwalk replicon (EC50 of 40 μM).111
RBV exerts its inhibitory action early against MNV, with effects detectable at 8 hpi (MOI 0.01), suggesting chain termination as a possible mode of NA action.114 However, the sequencing of MNV populations after four serial passages within the RAW264.7 cell line and in the presence of 200 μM RBV showed a three‐fold increase in mutations per nucleotide compared to untreated populations.114 These observations are consistent with reports of RBV having multiple modes of action, including mutagenesis, direct inhibition of RNA polymerases, unbalancing intracellular NTP pools, and interference of 5′ cap structures for capped viruses.120, 123, 124, 125, 126, 127 Despite this, the modest inhibition of RBV against MNV and the Norwalk replicon, coupled with the numerous adverse effects reported with RBV treatment, suggests that this antiviral is not desirable as an antinorovirus agent.
CMX521 [2.2.4] is a novel NA discovered through an HTS, and according to the manufacturer's press releases, has potent and pan‐genotypic activity against norovirus.128 CMX521 is reportedly in the recruitment stage of phase I clinical trials to evaluate the safety, tolerability, and pharmacokinetics in less than or equal to 50 healthy adults. No peer‐reviewed publications were available when writing this review, with results projected to be released later in 2018.128
2.2.2. Non‐nucleoside inhibitors
NNIs generally exhibit narrow‐spectrum antiviral activity and bind allosterically to block conformational rearrangements of the viral polymerase required to form an active replication complex.129 Currently, there are three known NNI binding sites on the norovirus RdRp. One binding pocket is within the NTP access path located between the fingers and thumb domains,130 the second pocket is termed Site A, a positively charged NTP traversal channel with flexible amino acid side chains,131 and the third pocket is Site B, a highly conserved allosteric binding pocket present across the Caliciviridae and located within the thumb region131, 132 (Figure 3B).
Several therapeutics have been repurposed as NNIs against human norovirus. One example includes the sleeping sickness medication suramin [2.2.5], which is a large, symmetric polyanionic naphthylurea, with a molecular mass of 1297.29 g/mol. Suramin was found to potently inhibit in vitro activity of both the human and MNV polymerases130 with IC50 values of 24.6 and 70.0 nM, respectively. Similarly, the smaller suramin derivative NF023 [2.2.6] inhibited human and mouse norovirus RdRp activities with IC50 values of 71.5 and 200 nM, respectively.130 Cocrystallization studies of these compounds with the human norovirus polymerase identified the suramin and NF023 binding pocket within the NTP pathway, between the fingers and thumb domains.130
Other suramin derivatives that inhibit in vitro norovirus RdRp transcription are naphthalene disulfonate (NAF2; 286.27 g/mol) [2.2.7] and pyridoxal‐5′‐phosphate‐6‐(2′‐naphthylazo‐6′‐nitro‐4′,8′‐disulfonate) tetrasodium salt (PPNDS; 694.36 g/mol) [2.2.8].131, 132 NAF2 and PPNDS inhibited the human norovirus RdRp in an enzyme assay with IC50 values of 14 and 0.45 μM, respectively, and PPNDS also inhibited the MNV RdRp with an IC50 of 0.88 µM.131, 133 Cocrystallization studies revealed that NAF2 binds into both Site A and Site B, while PPNDS binds solely in Site B.131 Further X‐ray synchrotron crystallization studies by the same group confirmed that PPNDS also binds allosterically within Site B of the MNV RdRp by mimicking two stacking RNA bases.133
Despite the promising potency of suramin and its derivatives, including PPNDS, these compounds demonstrate limited bioavailability and exhibit poor cell permeability,134 greatly reducing their antiviral efficacy in viral culture.132, 135, 136 While suramin has been shown to inhibit MNV replication in cell culture (EC50 0.3 µM), the large structure and low cell permeability meant delivery required a liposome system to enter RAW264.7 cells.135 Similarly, PPNDS was found to be much less potent in MNV cell culture, demonstrating only 20.5% inhibition of MNV replication in a plaque assay at 10 µM, compared to 98.0% inhibition at the same concentration in the RdRp assays.132 Moreover, PPNDS has been eliminated from other enzyme inhibition studies due to nonspecific, off‐target effects,137 and is therefore likely to be an unsuitable drug candidate for further antinorovirus development.
Repurposing antivirals developed against other viruses can sometimes reveal drug candidates with broad‐spectrum activities for further development. A recent study evaluated the antiviral activity of six HCV NNIs against the human norovirus RdRp132 including; Filibuvir, JTK‐109, Lomibuvir, Nesbuvir, Setrobuvir, and Tegobuvir using a fluorescent polymerase assay. All HCV NNIs had an IC50 of more than 100 µM except JTK‐109. JTK‐109 [2.2.9], was shown to have broad‐spectrum activity across several genera of the Caliciviridae, inhibiting the human norovirus polymerase with an IC50 of 4.3 µM MNV replication in cell culture with an EC50 of 6.1 µM, and also132
Against calicivirus RdRps in an in vitro activity assay with EC50 values ranging from 0.1 to 2.3 µM.132 In the same study, PPNDS inhibited the human and mouse norovirus RdRps by 1.4 and 2.3 µM, respectively,132 which is less potent than previously reported above,133 with the variation likely due to different methodologies. Norovirus RdRp mutant studies, docking studies and combinational antagonism with PPNDS suggested the binding site of JTK‐109 is within the highly conserved Site B pocket.132
To further exploit the broad‐spectrum RdRp binding pocket Site B,132 a high‐throughput in silico screen of approximately 300 000 commercially available compounds was performed to identify ligands that bind to Site B of the human norovirus and MNV RdRps.50 Sixty‐two compounds were selected from the screen and assessed for inhibition of the human norovirus RdRp, revealing a hit (compound 11) with an IC50 of 5.0 µM. Synthesis of derivatives of this compound produced a novel candidate (compound 54 [2.2.10]) with IC50 values of 5.6 µM (human norovirus RdRp) and 12.1 µM (MNV RdRp). Further in silico molecular modeling and biological antagonism studies with PPNDS suggested the binding site of compound 54 was indeed within the targeted broad‐spectrum Site B pocket, however, it was relatively ineffective against MNV in cell culture, with an EC50 of more than 50 µM.50
These issues of cell permeability, specificity, compound toxicity, and potency could all be addressed by extensive medicinal chemistry to improve the “druggability” of these developmental NNIs.
Several potential norovirus NNIs have also been identified by HTS of compound libraries using viral enzyme activity assays. A case in point is the HTS of almost 20 000 compounds for RdRp inhibition using a fluorescent activity assay by Eltahla et al.47 Four lead compounds were identified (NIC02 [2.2.11], NIC04, NIC10, and NIC12) that had IC50 values against the human norovirus polymerase ranging from 5.0 to 9.8 µM, and inhibited both the Norwalk replicon and MNV replication in cell culture with EC50 values between 30.1 to >100 µM and 4.8 to 38.1 µM, respectively.47 These NNIs represent scaffolds with the potential to be structurally modified for increased potency.
While a growing body of research has been invested in identifying antivirals that target the norovirus polymerase, to date all candidates are still in early preclinical development and the search for a compound continues in earnest.
2.3. Protease inhibitors
Much like the RdRp, the norovirus protease (NS6; Figure 3C) represents a desirable antiviral target since it plays an essential role in viral replication, through cleavage of the NS polyprotein, which is necessary for the production of viral progeny. Norovirus protease inhibitors (PIs) have been tested over a wide range of model antiviral systems (Figure 2) and represent the class of calicivirus antivirals with the most number of compounds (Table 1). Rupintrivir [2.3.1] was one of the first PIs described for norovirus, and although this compound was originally developed for human rhinovirus, Kim et al48 demonstrated broad‐spectrum antiviral activity of Rupintrivir against other picornaviruses, coronaviruses, and caliciviruses, including; MNV, FCV, and the Norwalk replicon. Subsequent studies have supported these initial observations and showed that Rupintrivir inhibited Norwalk replicon with EC50 values of 0.3 and 1.3 μM.138, 139 However, in some cases PI potency does not translate in vivo, for example, Rupintrivir demonstrates poor pharmacokinetic properties and limited bioavailability,140 and is, therefore, not ideal as an antinorovirus therapy.
The remaining norovirus PIs described in the literature were mostly developed by one group of researchers and are broadly divided into transition state (TS) inhibitors and TS mimics (reviewed in Galasiti et al72). The TS mimics include α‐hydroxyphosphonates (e.g. compound 7d [2.3.2]141) which effectively inhibit the Norwalk replicon at low micromolar concentrations.141 Although potent, TS mimics are far less studied than TS inhibitors and represent a small proportion of the compounds described in Table 1.
The first TS inhibitors designed included a series of peptidyl aldehydes that incorporated a glutamine surrogate in their structure,142 and take advantage of the preference for the norovirus protease to cleave at glutamine‐glycine peptide sites.143 Within this original series of peptidyl aldehydes (n = 10), compounds 4 [2.3.3] and 5 displayed inhibition of MNV infection (EC50 of 5.5 and 20.3 μM, respectively) and the Norwalk replicon (EC50 of 2.1 and 7.8 μM, respectively).142 Several variants of the these primary hits were developed in later studies and included: dipeptidyl aldehydes (e.g. compound 16 [2.3.4]),48, 144 tripeptidyl aldehydes (e.g. syc10 [2.3.5]),145 peptidyl α‐ketoamides (e.g. compound 6d [2.3.6]),48, 141, 146 α‐ketoheterocycles (e.g. compound 8a [2.3.7],146 bisulfite adducts (e.g. GC376, [2.3.8]),48, 147, 148 and ester or carbamate prodrugs of bilsulfite adducts (e.g. compound 11 [2.3.9]).149 Each of these variants have displayed an equal or a greater level of antiviral activity when compared to the original peptidyl aldehyde inhibitors, with the most potent of these variants displaying low nanomolar potency against MNV (in vitro and in vivo) and the Norwalk replicon [2.3.4].144
Another subclass of TS inhibitors includes macrocyclic compounds150, 151, 152, 153 which are structurally modified versions of the aforementioned peptide‐based TS inhibitors. These macrocyclic compounds were originally designed to improve the membrane permeability and oral bioavailability of TS inhibitors to increase their suitability for the clinical application.153 Several studies have investigated the inhibitory effects of these macrocyclic TS inhibitors (EC50 range: 1.5‐>20 μM) such as compound 24 [2.3.10]),151, 154 as well as triazole‐based (EC50 range, 3.8‐88.3 μM) (e.g. compound 8 [2.3.11]),153 oxadiazole‐based (EC50 range, 2.5‐51.2 μM; e.g. compound 17 [2.3.12]),150 and oxazolidinone‐based (EC50 range, 6.7‐17.5 μM; e.g. compound 9 [2.3.13])152 variants against the Norwalk replicon. Although none of the macrocyclic TS inhibitors have reached the potency achieved by the peptide‐based TS inhibitors, several macrocyclic variants display broad‐spectrum antiviral activity against members of the picornavirus‐like supercluster similar to that of Rupintrivir.154 The effectiveness of PIs across multiple genera and norovirus genogroups illustrates that the protease is an excellent antiviral candidate for the treatment of norovirus infections.
2.4. Host‐factor drugs
A pitfall of using some DAAs is the emergence of resistance mutations which can undermine their effectiveness, although combinational therapy is a proven option in HIV and HCV therapy to circumvent this. Antivirals that target the host generally have a higher barrier to resistance than some classes of DAAs, particularly PIs and NNIs which are known to have a low barrier to resistance (reviewed in Kaufmann et al155). Thus host‐targeted therapies represent an important antiviral class to be considered for the treatment of norovirus infections. Host‐factor drugs can target individual cellular components that directly interact with the virus or aid viral replication. Alternatively, host‐factor drugs may influence multiple cellular components that culminate in antiviral defenses.
2.4.1. Protein targets
A recently explored class of host‐targeted norovirus antivirals include deubiquitinase (DUB) inhibitors.156, 157, 158 DUB inhibitors are a class of enzymes involved in regulation of the ubiquitin‐proteasome system which is commonly exploited by viruses for replication (reviewed in Luo159). WP1130 [2.4.1] is an example of a small synthetic DUB inhibitor (molecular weight of 384.2 g/mol) that was shown to effectively inhibit MNV and norovirus replication through induction of the unfolded protein response.158 However, this initial study revealed that inhibition of MNV in mice was limited to the small intestine due to poor bioavailability of WP1130.158 To address the poor bioavailability, libraries of WP1130 variants were developed and tested for improved antiviral efficacy.156, 157 Out of 59 derivatives, compound 9 [2.4.2] was found to be the most potent antiviral.156 Treatment at 2.5 µM with this derivative resulted in an 84.7% reduction in replication of the Norwalk replicon and a 2.5‐log reduction in MNV titer in infected cells; more than double the potency of the parental compound.156
Another host protein that has been identified as an antiviral target is heat‐shock protein 90 (Hsp90), a molecular chaperone involved in the maturation of proteins responsible for multiple biological processes (reviewed in Li et al160). Vashist et al161 initially showed using siRNA knockdown and overexpression studies that Hsp90 interacts with the 5′ and 3′ termini the MNV genome and plays a key role in MNV replication.161 The authors then showed that abolishment of Hsp90 activity using 17‐dimethylaminoethylamino‐17‐demethoxygeldanamycin (17‐DMAG [2.4.3]) potently inhibited MNV replication in vitro (IC50 of 60 nM and CC50 of 4.5 µM) and reduced MNV titer (~ 1‐log) in an in vivo mouse model.161
Despite potential cytotoxicity from some host‐targeted compounds (reviewed in Engel et al162), generally, therapeutics that target specific host proteins result in effective antiviral activity with minimal off‐target effects, which are desirable features for norovirus therapeutics.
2.4.2. Immunomodulators
Immunomodulators are an excellent therapeutic option for virus infections due to their ability to induce a powerful host response against intracellular parasites. The best example of immunomodulators are interferons (IFNs) and for over a decade, studies have shown that type I and II IFNs, as well as their receptors, provide protection against murine and human norovirus infections.44, 52, 53, 163, 164, 165, 166, 167, 168 However, the role of type III IFNs (IFN‐λ), in norovirus infection and their potential as norovirus antivirals has only recently been explored.169 IFN‐λ [2.4.4] binds to a heterodimeric receptor comprised of IFN‐λ receptor 1 (IFNLR1) and interleukin‐10R2 (IL‐10R2), which induces the expression of many of the same genes induced by type I and II IFNs.170, 171, 172, 173 However, unlike type I and II IFN, a single dose of IFN‐λ (1 μg) has been shown to both prevent and clear persistent MNV infection in mice.174 IFN‐λ was shown to target nonhematopoietic cells174 and intestinal epithelial cells which express high levels of IFNLR1.175 Using a MNV transmission model, endogenous IFN‐λ expression (induced using a transfected plasmid) was shown to block MNV transmission from mice infected with the acute CW3 MNV strain.176 IFN‐λ treatment also prevented intestinal CW3 replication, inflammation, and antibody responses in mice,176 which illustrates that it has an integral role in the prevention of norovirus transmission. Taken together, the antiviral activity displayed by IFN‐λ in these studies reveal that it has potential as a therapeutic to not only cure infections but also prevent norovirus spread if given prophylactically.
More recently the group of immunomodulators known as Toll‐like receptor (TLR) agonists have been explored as norovirus antivirals. These compounds have been used for many years as vaccine adjuvants, but have also been shown to inhibit the replication of RNA viruses HIV and HBV.177, 178 TLR agonists induce the innate immune response and stimulate IFN production, which is known to have antiviral activities against MNV and human norovirus in vitro.44, 52, 53, 163, 164, 165, 166, 167, 168 Several TLR7 agonists were shown to display potent inhibition of MNV infection, including R‐848 [2.4.5] (23.5 nM), GS‐9620 (0.59 μM), Gardiquimod (0.13 μM) and R‐837 (1.5 μM), with therapeutic indices more than 30 for each compound.179 Furthermore, conditioned media generated by stimulation of THP‐1 cells with 10 µM of R‐848 reduced replication of the Norwalk replicon by 50%.179 More recently the TLR4 agonist, poly‐γ‐glutamic acid (γ‐PGA) [2.4.6] was explored as an antiviral and when tested against MNV in vitro displayed an EC90 value < 100 nM.180 In addition, a 50 mg/kg dosage of γ‐PGA to mice before and following MNV infection resulted in ≥ 47% reduction in viral genomes.180 Given that the TLR4 agonist monophospholipid A is approved for use as an adjuvant for HPV and HBV vaccines (reviewed in Dowling and Mansell181) and the TLR7 agonist R‐837 is already approved for the treatment of HPV‐associated genital warts,182 repurposing TLR4 and TLR7 agonists could hugely expedite their use for norovirus clinical treatment.
2.5. Compounds with unknown targets
Several compounds have demonstrated antiviral activity where the mechanism of inhibition is unknown. These include flavonoid‐type molecules, such as theaflavin digallate [2.5.1], myricetin and epigallocatechin‐3‐gallate, among many others that display broad‐spectrum medicinal properties, although they display limited inhibition of norovirus infection in vitro.183, 184, 185, 186 The mechanism of action of flavonoids against norovirus is thought to be a result of anti‐inflammatory pathway activation. Similarly, phytochemicals such as curcumin [2.5.2] are a class of compounds that display inhibition of norovirus in vitro,187 however, the mechanism of inhibition is poorly understood. Vitamin A [2.5.3] has been shown to inhibit MNV in vitro, albeit poorly with less than 50% inhibition at 50 U/mL and ~50% inhibition of the Norwalk replicon at 100 U/mL. Vitamin A‐induced changes in the microflora are thought to be the mechanism responsible for these antiviral effects.188 Lastly, nitazoxanide (NTZ) [2.5.4] (covered in section 3) has shown promise as an antiviral therapy, but the defined mechanism of activity against norovirus has yet to be determined. Most recently NTZ was shown to potently inhibit FCV replication in cell culture with an EC50 of 0.6 µM,189 and the GI norovirus replicon at clinically relevant concentration (5 μg/mL),190 which was later shown to result in a broad antiviral response.191 Despite these observations, the latter study also showed that NTZ was ineffective against MNV suggesting that further antiviral investigations are warranted.189
3. DRUGS IN CLINICAL DEVELOPMENT
To date, no norovirus antiviral or vaccine is approved for medical use, and the only norovirus antiviral candidate to complete clinical trials is NTZ. This compound was originally developed in the 1970s, and is currently an FDA‐approved therapy for treating Giardia and Crytosporidium infections.192 NTZ has demonstrated broad‐spectrum antimicrobial activity against a range of bacterial, protozoan, and viral infections, including inhibition of the Norwalk replicon (EC50 1.6 µM) (reviewed in Rossignol193).
In phase II randomized double‐blind trial, NTZ therapy was administered to 25 of 50 patients (≥ 12 years) that tested positive for rotavirus or norovirus infection. Treatment resulted in a significant reduction in the duration of gastroenteritis symptoms (norovirus, P = 0.0295 and rotavirus, P = 0.0052) from 2.5 to 1.5 days when compared to the placebo.194
Additional anecdotal studies have supported the efficacy of NTZ treatment for norovirus. NTZ successfully treated one immunosuppressed transplant patient infected with norovirus that had experienced 10 consecutive days of gastroenteritis symptoms.195 Four days after commencing NTZ treatment, a complete resolution of symptoms was recorded without a reduction in immunosuppressive drugs.195 NTZ treatment was also shown to resolve diarrheal symptoms and clear norovirus in stool samples from a pediatric patient with chronic norovirus, following kidney transplantation.196
Although the above‐mentioned studies suggest that NTZ is a promising therapy for the treatment of norovirus infections, there is an equal amount of evidence revealing that NTZ is ineffective against norovirus infections.197, 198, 199, 200 Despite this contrary evidence, NTZ represents the only therapeutic option currently available apart from RBV, immunoglobulins and supportive care to patients with persistent infections.
4. CONCLUSIONS
Human norovirus is a pervasive pathogen that creates a significant social and economic impact and causes hundreds of thousands of deaths each year. Despite intensive research for safe and effective norovirus antivirals, none have yet been clinically approved, and the majority of candidates are still in the early stages of preclinical development. This review outlines recent therapeutic candidate studies to provide an updated overview of the current human norovirus antiviral development pipeline.
Optimization of human norovirus culture in enteroid and B cell systems offers the promise that in time, more robust and reliable culture methods will be developed allowing greater replication levels. These improved systems may enhance antiviral studies to provide a stronger platform for effective norovirus antiviral development.
The landscape of antiviral development for norovirus is changing. The rapidly developing field of host immunomodulatory therapies is opening the door to potential treatments for many viral infections, with promising results already published for norovirus. Moreover, recent discoveries have reported highly conserved binding pockets on critical viral enzymes, such as the norovirus polymerase, which could allow for the further development of broad‐spectrum antivirals.
CONFLICTS OF INTEREST
The authors declare that there are no conflicts of interest.
ACKNOWLEDGEMENTS
PAW, DET, and NEN acknowledge the support from National Health and Medical Research Council project grants (APP1083139 and APP1123135). NEN and DET acknowledge support through Research Training Program and Research Excellence Awards.
Biographies
AUTHOR’S BIOGRAPHIES
NE Netzler is a PhD candidate of molecular microbiology at the University of New South Wales (UNSW), Australia. Her project focuses on identifying antiviral compounds for norovirus and other caliciviruses. In particular, her interest lies in the discovery of broad‐spectrum antivirals with the potential to rapidly treat several clinically significant viruses for which we currently have no antivirals. During her PhD, Natalie was an author and editor for the book Foodborne Viral Pathogens published by Taylor and Francis (CRC Press). Natalie is also involved in teaching at UNSW and has been a tutor and demonstrator on the 2nd and 3rd year microbiology and virology courses. Following a BSc in biology (1998), Natalie completed an MSc in virology with first class Honors (2000), from the University of Waikato, New Zealand. Following graduation, Natalie worked as a research assistant at the biotechnology company, Genesis Research and Development, for several years, investigating ligands involved in plant flowering and siRNA inhibition targeting drug resistance.
D Enosi Tuipulotu is a PhD candidate of molecular virology at the University of New South Wales (UNSW). He completed a B.Med Sci with first class Honors majoring in Microbiology and Immunology from the University of New South Wales in 2014 and is currently a PhD candidate at UNSW focusing on the biology of norovirus infections. Specifically, he is interested in understanding the immune responses to norovirus infections to help identify host targets for antiviral therapy. Daniel has identified that agonists of Toll‐like receptors involved in antiviral defense are effective inhibitors or norovirus replication and represent a potential therapy for calicivirus infections. He is also involved in teaching at UNSW, including tutor and demonstration roles biology and microbiology.
PA White is a Professor of Microbiology and Molecular Biology at the University of New South Wales (UNSW), Australia. He is a microbiologist with interests in noroviruses, antiviral development, viral evolution, and antibiotic resistance. He has a breadth of experience in the development of novel molecular assay systems and next‐generation sequencing to investigate viral infections. Following a BSc (Hons) in biotechnology from King's College London (1992), Peter completed a PhD at the University College, London (1996) in molecular microbiology and biochemistry. Prof White commenced his postdoctoral research studies at the Macquarie University, Sydney, as a recipient of a Royal Society Fellowship and later worked as Hepatitis Group Leader at the Prince of Wales Hospital until joining the University of New South Wales in 2003. He is also an enthusiastic teacher, having convened the third‐year science course Viruses and Disease for 15 years at UNSW.
Netzler NE, Enosi Tuipulotu D, White PA. Norovirus antivirals: Where are we now? Med Res Rev. 2019;39:860–886. 10.1002/med.21545
References
REFERENCES
- 1. Lopman BA, Steele D, Kirkwood CD, Parashar UD. The vast and varied global burden of norovirus: Prospects for prevention and control. PLOS Med. 2016;13(4):e1001999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Pires SM, Fischer‐Walker CL, Lanata CF, et al. Aetiology‐specific estimates of the global and regional incidence and mortality of diarrhoeal diseases commonly transmitted through food. PLOS One. 2015;10(12):e0142927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Bartsch SM, Lopman BA, Ozawa S, Hall AJ, Lee BY. Global economic burden of norovirus gastroenteritis. PLOS One. 2016;11(4):e0151219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Atmar RL, Opekun AR, Gilger MA, et al. Norwalk virus shedding after experimental human infection. Emerg Infect Dis. 2008;14(10):1553‐1557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Ahmed SM, Hall AJ, Robinson AE, et al. Global prevalence of norovirus in cases of gastroenteritis: a systematic review and meta‐analysis. Lancet Infect Dis. 2014;14(8):725‐730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Ludwig A, Adams O, Laws HJ, Schroten H, Tenenbaum T. Quantitative detection of norovirus excretion in pediatric patients with cancer and prolonged gastroenteritis and shedding of norovirus. J Med Virol. 2008;80(8):1461‐1467. [DOI] [PubMed] [Google Scholar]
- 7. Bok K, Green KY. Norovirus gastroenteritis in immunocompromised patients. N Engl J Med. 2012;367(22):2126‐2132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Division of Viral Diseases, National Center for Immunization and Respiratory Diseases, Centers for Disease Control and Prevention . Updated norovirus outbreak management and disease prevention guidelines. MMWR Recomm Rep. 2011;60(RR‐3):1‐18. [PubMed] [Google Scholar]
- 9. Teunis PFM, Moe CL, Liu P, et al. Norwalk virus: how infectious is it? J Med Virol. 2008;80(8):1468‐1476. [DOI] [PubMed] [Google Scholar]
- 10. Harris JP, Lopman BA, O’Brien SJ. Infection control measures for norovirus: a systematic review of outbreaks in semi‐enclosed settings. J Hosp Infect. 2010;74(1):1‐9. [DOI] [PubMed] [Google Scholar]
- 11. Gonzaga VE, Ramos M, Maves RC, Freeman R, Montgomery JM. Concurrent outbreak of norovirus genotype I and enterotoxigenic Escherichia coli on a U.S. Navy ship following a visit to Lima, Peru. PLOS One. 2011;6(6):e20822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Farkas T, Thornton SA, Wilton N, Zhong W, Altaye M, Jiang X. Homologous versus heterologous immune responses to Norwalk‐like viruses among crew members after acute gastroenteritis outbreaks on 2 US Navy vessels. J Infect Dis. 2003;187(2):187‐193. [DOI] [PubMed] [Google Scholar]
- 13. Sukhrie FHA, Siebenga JJ, Beersma MFC, Koopmans M. Chronic shedders as reservoir for nosocomial transmission of norovirus. J Clin Microbiol. 2010;48(11):4303‐4305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Havelaar AH, Kirk MD, Torgerson PR, et al. World Health Organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLOS Med. 2015;12(12):e1001923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Clarke IN, Estes MK, Green KY, et al. Caliciviridae. 2012. International Committee on Taxonomy of Viruses. 9th Report, San Diego, CA.
- 16. Farkas T, Sestak K, Wei C, Jiang X. Characterization of a rhesus monkey calicivirus representing a new genus of Caliciviridae. J Virol. 2008;82(11):5408‐5416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Kemenesi G, Gellért Á, Dallos B, et al. Sequencing and molecular modeling identifies candidate members of Caliciviridae family in bats. Infect Genet Evol. 2016;41:227‐232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. L’homme Y, Sansregret R, Plante‐Fortier É, et al. Genomic characterization of swine caliciviruses representing a new genus of Caliciviridae. Virus Genes. 2009;39(1):66‐75. [DOI] [PubMed] [Google Scholar]
- 19. Mikalsen AB, Nilsen P, Frøystad‐Saugen M, et al. Characterization of a novel calicivirus causing systemic infection in Atlantic salmon (Salmo salar L.): proposal for a new genus of caliciviridae. PLOS One. 2014;9(9):e107132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Vinjé J. Advances in laboratory methods for detection and typing of norovirus. J Clin Microbiol. 2015;53(2):373‐381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Eden JS, Tanaka MM, Boni MF, Rawlinson WD, White PA. Recombination within the pandemic norovirus GII.4 lineage. J Virol. 2013;87(11):6270‐6282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ludwig‐Begall LF, Mauroy A, Thiry E. Norovirus recombinants: recurrent in the field, recalcitrant in the lab—a scoping review of recombination and recombinant types of noroviruses. J Gen Virol. 2018;99(8):970‐988. [DOI] [PubMed] [Google Scholar]
- 23. Atmar RL, Estes MK. The epidemiologic and clinical importance of norovirus infection. Gastroenterol Clin North Am. 2006;35(2):275‐290. [DOI] [PubMed] [Google Scholar]
- 24. Bull RA, Tu ETV, McIver CJ, Rawlinson WD, White PA. Emergence of a new norovirus genotype II.4 variant associated with global outbreaks of gastroenteritis. J Clin Microbiol. 2006;44(2):327‐333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Siebenga JJ, Vennema H, Zheng DP, et al. Norovirus illness is a global problem: emergence and spread of norovirus GII.4 variants, 2001‐2007. J Infect Dis. 2009;200(5):802‐812. [DOI] [PubMed] [Google Scholar]
- 26. Tu ETV, Bull RA, Greening GE, et al. Epidemics of gastroenteritis during 2006 were associated with the spread of norovirus GII.4 variants 2006a and 2006b. Clin Infect Dis. 2008;46(3):413‐420. [DOI] [PubMed] [Google Scholar]
- 27. White PA. Evolution of norovirus. Clin Microbiol Infect. 2014;20(8):741‐745. [DOI] [PubMed] [Google Scholar]
- 28. Koo HS, Lee MO, Ku PT, Hwang SJ, Park DJ, Baik HS. Molecular epidemiology of norovirus in asymptomatic food handlers in Busan, Korea, and emergence of genotype GII.17. J Microbiol. 2016;54(10):686‐694. [DOI] [PubMed] [Google Scholar]
- 29. Matsushima Y, Ishikawa M, Shimizu T, et al. Genetic analyses of GII.17 norovirus strains in diarrheal disease outbreaks from December 2014 to March 2015 in Japan reveal a novel polymerase sequence and amino acid substitutions in the capsid region. Euro Surveill. 2015;20(26):21173. [DOI] [PubMed] [Google Scholar]
- 30. Lu J, Sun L, Fang L, et al. Gastroenteritis outbreaks caused by norovirus GII.17, Guangdong Province, China, 2014‐2015. Emerg Infect Dis. 2015;21(7):1240‐1242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Medici MC, Tummolo F, Calderaro A, et al. Identification of the novel Kawasaki 2014 GII.17 human norovirus strain in Italy, 2015. Euro Surveill. 2015;20(35):30010. [DOI] [PubMed] [Google Scholar]
- 32. Parra GI, Green KY. Genome of emerging norovirus GII.17, United States, 2014. Emerg Infect Dis. 2015;21(8):1477‐1479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Lun JH, Hewitt J, Sitabkhan A, et al. Emerging recombinant noroviruses identified by clinical and waste water screening. Emerg Microbes Infect. 2018;7(1):50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Atmar RL. Noroviruses: state of the art. Food and environmental virology. 2010;2(3):117‐126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Karst SM. Pathogenesis of noroviruses, emerging RNA viruses. Viruses. 2010;2(3):748‐781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Karst SM, Wobus CE, Goodfellow IG, Green KY, Virgin HW. Advances in norovirus biology. Cell Host Microbe. 2014;15(6):668‐680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Vongpunsawad S, Venkataram prasad BV, Estes MK. Norwalk virus minor capsid protein VP2 associates within the VP1 shell domain. J Virol. 2013;87(9):4818‐4825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Prasad BV, Hardy ME, Dokland T, Bella J, Rossmann MG, Estes MK. X‐ray crystallographic structure of the Norwalk virus capsid. Science. 1999;286(5438):287‐290. [DOI] [PubMed] [Google Scholar]
- 39. Prasad BV, Rothnagel R, Jiang X, Estes MK. Three‐dimensional structure of baculovirus‐expressed Norwalk virus capsids. J Virol. 1994;68(8):5117‐5125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Jiang X, Wang M, Graham DY, Estes MK. Expression, self‐assembly, and antigenicity of the Norwalk virus capsid protein. J Virol. 1992;66(11):6527‐6532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Jones MK, Grau KR, Costantini V, et al. Human norovirus culture in B cells. Nat Protoc. 2015;10(12):1939‐1947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Ettayebi K, Crawford SE, Murakami K, et al. Replication of human noroviruses in stem cell‐derived human enteroids. Science. 2016;353(6306):1387‐1393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Jones MK, Watanabe M, Zhu S, et al. Enteric bacteria promote human and mouse norovirus infection of B cells. Science. 2014;346(6210):755‐759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Chang KO, Sosnovtsev SV, Belliot G, King AD, Green KY. Stable expression of a Norwalk virus RNA replicon in a human hepatoma cell line. Virology. 2006;353(2):463‐473. [DOI] [PubMed] [Google Scholar]
- 45. Horscroft N, Lai VC, Cheney W, et al. Replicon cell culture system as a valuable tool in antiviral drug discovery against hepatitis C virus. Antivir Chem Chemother. 2005;16(1):1‐12. [DOI] [PubMed] [Google Scholar]
- 46. Asselah T, Boyer N, Saadoun D, Martinot‐Peignoux M, Marcellin P. Direct‐acting antivirals for the treatment of hepatitis C virus infection: optimizing current IFN‐free treatment and future perspectives. Liver Int. 2016;36(suppl 1):47‐57. [DOI] [PubMed] [Google Scholar]
- 47. Eltahla AA, Lim KL, Eden JS, Kelly AG, Mackenzie JM, White PA. Nonnucleoside inhibitors of norovirus RNA polymerase: scaffolds for rational drug design. Antimicrob Agents Chemother. 2014;58(6):3115‐3123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Kim Y, Lovell S, Tiew KC, et al. Broad‐spectrum antivirals against 3C or 3C‐like proteases of picornaviruses, noroviruses, and coronaviruses. J Virol. 2012;86(21):11754‐11762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Eltahla AA, Lackovic K, Marquis C, Eden JS, White PA. A fluorescence‐based high‐throughput screen to identify small compound inhibitors of the genotype 3a hepatitis C virus RNA polymerase. J Biomol Screen. 2013;18(9):1027‐1034. [DOI] [PubMed] [Google Scholar]
- 50. Ferla S, Netzler NE, Ferla S, et al. In silico screening for human norovirus antivirals reveals a novel non‐nucleoside inhibitor of the viral polymerase. Sci Rep. 2018;8(1):4129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Vashist S, Bailey D, Putics A, Goodfellow I. Model systems for the study of human norovirus Biology. Future Virol. 2009;4(4):353‐367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Karst SM, Wobus CE, Lay M, Davidson J, Virgin HWt. STAT1‐dependent innate immunity to a Norwalk‐like virus. Science. 2003;299(5612):1575‐1578. [DOI] [PubMed] [Google Scholar]
- 53. Wobus CE, Karst SM, Thackray LB, et al. Replication of norovirus in cell culture reveals a tropism for dendritic cells and macrophages. PLOS Biol. 2004;2(12):e432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Lim MY, Kim JM, Lee JE, Ko G. Characterization of ozone disinfection of murine norovirus. Appl Environ Microbiol. 2010;76(4):1120‐1124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Fraisse A, Temmam S, Deboosere N, et al. Comparison of chlorine and peroxyacetic‐based disinfectant to inactivate Feline calicivirus, Murine norovirus and Hepatitis A virus on lettuce. Int J Food Microbiol. 2011;151(1):98‐104. [DOI] [PubMed] [Google Scholar]
- 56. Lim MY, Kim JM, Ko G. Disinfection kinetics of murine norovirus using chlorine and chlorine dioxide. Water Res. 2010;44(10):3243‐3251. [DOI] [PubMed] [Google Scholar]
- 57. Chaudhry Y, Skinner MA, Goodfellow IG. Recovery of genetically defined murine norovirus in tissue culture by using a fowlpox virus expressing T7 RNA polymerase. J Gen Virol. 2007;88(Pt 8):2091‐2100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Yunus MA, Chung LMW, Chaudhry Y, Bailey D, Goodfellow I. Development of an optimized RNA‐based murine norovirus reverse genetics system. J Virol Methods. 2010;169(1):112‐118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Bok K, Parra GI, Mitra T, et al. Chimpanzees as an animal model for human norovirus infection and vaccine development. Proc Natl Acad Sci USA. 2011;108(1):325‐330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Subekti DS, Tjaniadi P, Lesmana M, et al. Experimental infection of Macaca nemestrina with a Toronto Norwalk‐like virus of epidemic viral gastroenteritis. J Med Virol. 2002;66(3):400‐406. [DOI] [PubMed] [Google Scholar]
- 61. Rockx BHG, Bogers WMJM, Heeney JL, Van Amerongen G, Koopmans MPG. Experimental norovirus infections in non‐human primates. J Med Virol. 2005;75(2):313‐320. [DOI] [PubMed] [Google Scholar]
- 62. Cheetham S, Souza M, Meulia T, Grimes S, Han MG, Saif LJ. Pathogenesis of a genogroup II human norovirus in gnotobiotic pigs. J Virol. 2006;80(21):10372‐10381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Cheetham S, Souza M, McGregor R, Meulia T, Wang Q, Saif LJ. Binding patterns of human norovirus‐like particles to buccal and intestinal tissues of gnotobiotic pigs in relation to A/H histo‐blood group antigen expression. J Virol. 2007;81(7):3535‐3544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Souza M, Azevedo MSP, Jung K, Cheetham S, Saif LJ. Pathogenesis and immune responses in gnotobiotic calves after infection with the genogroup II.4‐HS66 strain of human norovirus. J Virol. 2008;82(4):1777‐1786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Kocher J, Bui T, Giri‐Rachman E, et al. Intranasal P particle vaccine provided partial cross‐variant protection against human GII.4 norovirus diarrhea in gnotobiotic pigs. J Virol. 2014;88(17):9728‐9743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Seo DJ, Jung D, Jung S, et al. An experimental miniature piglet model for the infection of human norovirus GII. J Med Virol. 2017;90:655‐662. [DOI] [PubMed] [Google Scholar]
- 67. Kolawole AO, Rocha‐Pereira J, Elftman MD, Neyts J, Wobus CE. Inhibition of human norovirus by a viral polymerase inhibitor in the B cell culture system and in the mouse model. Antiviral Res. 2016;132:46‐49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Taube S, Kolawole AO, Höhne M, et al. A mouse model for human norovirus. MBio. 2013;4(4):e00450‐e00413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Guerrero RA, Ball JM, Krater SS, Pacheco SE, Clements JD, Estes MK. Recombinant Norwalk virus‐like particles administered intranasally to mice induce systemic and mucosal (fecal and vaginal) immune responses. J Virol. 2001;75(20):9713‐9722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Thorne LG, Goodfellow IG. Norovirus gene expression and replication. J Gen Virol. 2014;95(Pt 2):278‐291. [DOI] [PubMed] [Google Scholar]
- 71. Prasad BV, Shanker S, Muhaxhiri Z, et al. Antiviral targets of human noroviruses. Curr Opin Virol. 2016;18:117‐125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Galasiti Kankanamalage AC, Weerawarna PM, Kim Y, Chang KO, Groutas WC. Anti‐norovirus therapeutics: a patent review (2010‐2015). Expert Opin Ther Pat. 2016;26(3):297‐308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Thorne L, Arias A, Goodfellow I. Advances toward a norovirus antiviral: from classical inhibitors to lethal mutagenesis. J Infect Dis. 2016;213(suppl 1):S27‐S31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Kocher J, Yuan L. Norovirus vaccines and potential antinorovirus drugs: recent advances and future perspectives. Future Virol. 2015;10(7):899‐913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Kim Y, Galasiti Kankanamalage AC, Chang KO, Groutas WC. Recent advances in the discovery of norovirus therapeutics. J Med Chem. 2015;58(24):9438‐9450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Altmeyer R. Virus attachment and entry offer numerous targets for antiviral therapy. Curr Pharm Des. 2004;10(30):3701‐3712. [DOI] [PubMed] [Google Scholar]
- 77. Haga K, Fujimoto A, Takai‐Todaka R, et al. Functional receptor molecules CD300lf and CD300ld within the CD300 family enable murine noroviruses to infect cells. Proc Natl Acad Sci USA. 2016;113(41):E6248‐E6255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Orchard RC, Wilen CB, Doench JG, et al. Discovery of a proteinaceous cellular receptor for a norovirus. Science. 2016;353(6302):933‐936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Tan M, Hegde RS, Jiang X. The P domain of norovirus capsid protein forms dimer and binds to histo‐blood group antigen receptors. J Virol. 2004;78(12):6233‐6242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Choi JM, Hutson AM, Estes MK, Prasad BVV. Atomic resolution structural characterization of recognition of histo‐blood group antigens by Norwalk virus. Proc Natl Acad Sci USA. 2008;105(27):9175‐9180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Tan M, Jiang X. Norovirus gastroenteritis, carbohydrate receptors, and animal models. PLOS Pathog. 2010;6(8):e1000983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Tan M, Jiang X. Virus‐host interaction and cellular receptors of caliciviruses In Hansman GS, Jiang XJ and Green KY (Eds.), Caliciviruses molecular and cellular virology. 1st ed Norwich: Caister Academic Press; 2010:111‐129. [Google Scholar]
- 83. Hansman GS, Shahzad‐ul‐Hussan S, McLellan JS, et al. Structural basis for norovirus inhibition and fucose mimicry by citrate. J Virol. 2012;86(1):284‐292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Tan M, Jiang X. Norovirus and its histo‐blood group antigen receptors: an answer to a historical puzzle. Trends Microbiol. 2005;13(6):285‐293. [DOI] [PubMed] [Google Scholar]
- 85. Zhang X‐F, Tan M, Chhabra M, Dai Y‐C, Meller J, Jiang X. Inhibition of histo‐blood group antigen binding as a novel strategy to block norovirus infections. PLOS One. 2013;8(7):e69379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Weichert S, Koromyslova A, Singh BK, et al. Structural basis for norovirus inhibition by human milk oligosaccharides. J Virol. 2016;90(9):4843‐4848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Koromyslova A, Tripathi S, Morozov V, Schroten H, Hansman GS. Human norovirus inhibition by a human milk oligosaccharide. Virology. 2017;508:81‐89. [DOI] [PubMed] [Google Scholar]
- 88. Koromyslova AD, White PA, Hansman GS. Treatment of norovirus particles with citrate. Virology. 2015;485:199‐204. [DOI] [PubMed] [Google Scholar]
- 89. Chen Z, Sosnovtsev SV, Bok K, et al. Development of Norwalk virus‐specific monoclonal antibodies with therapeutic potential for the treatment of Norwalk virus gastroenteritis. J Virol. 2013;87(17):9547‐9557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Lindesmith LC, Beltramello M, Donaldson EF, et al. Immunogenetic mechanisms driving norovirus GII. 4 antigenic variation. PLOS Pathog. 2012;8(5):e1002705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Lindesmith LC, Beltramello M, Swanstrom J, Swanstrom J, Jones TA, Corti D, Lanzavecchia A, & Baric RS. Serum immunoglobulin A cross‐strain blockade of human noroviruses. In Open forum infectious diseases (Vol. 2, No. 3). Oxford University Press; 2015. 10.1093/ofid/ofv084 [DOI] [PMC free article] [PubMed]
- 92. Koromyslova AD, Hansman GS. Nanobodies targeting norovirus capsid reveal functional epitopes and potential mechanisms of neutralization. PLOS Pathog. 2017;13(11):e1006636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Koromyslova AD, Hansman GS. Nanobody binding to a conserved epitope promotes norovirus particle disassembly. J Virol. 2015;89(5):2718‐2730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Muyldermans S. Nanobodies: natural single‐domain antibodies. Annu Rev Biochem. 2013;82:775‐797. [DOI] [PubMed] [Google Scholar]
- 95. Harmsen MM, De Haard HJ. Properties, production, and applications of camelid single‐domain antibody fragments. Appl Microbiol Biotechnol. 2007;77(1):13‐22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Malet H, Massé N, Selisko B, et al. The flavivirus polymerase as a target for drug discovery. Antiviral Res. 2008;80(1):23‐35. [DOI] [PubMed] [Google Scholar]
- 97. Hogbom M, Jager K, Robel I, Unge T, Rohayem J. The active form of the norovirus RNA‐dependent RNA polymerase is a homodimer with cooperative activity. J Gen Virol. 2009;90(Pt 2):281‐291. [DOI] [PubMed] [Google Scholar]
- 98. Bull RA, Hyde J, Mackenzie JM, et al. Comparison of the replication properties of murine and human calicivirus RNA‐dependent RNA polymerases. Virus Genes. 2011;42(1):16‐27. [DOI] [PubMed] [Google Scholar]
- 99. Ng KKS, Cherney MM, Vázquez AL, et al. Crystal structures of active and inactive conformations of a caliciviral RNA‐dependent RNA polymerase. J Biol Chem. 2002;277(2):1381‐1387. [DOI] [PubMed] [Google Scholar]
- 100. Eltahla A, Luciani F, White P, Lloyd A, Bull R. Inhibitors of the hepatitis C virus polymerase; mode of action and resistance. Viruses. 2015;7(10):5206‐5224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Zhang F, Wang G. A review of non‐nucleoside anti‐hepatitis B virus agents. Eur J Med Chem. 2014;75:267‐281. [DOI] [PubMed] [Google Scholar]
- 102. Fung J, Lai C‐L, Seto W‐K, Yuen M‐F. Nucleoside/nucleotide analogues in the treatment of chronic hepatitis B. J Antimicrob Chemother. 2011;66(12):2715‐2725. [DOI] [PubMed] [Google Scholar]
- 103. De Clercq E. The nucleoside reverse transcriptase inhibitors, nonnucleoside reverse transcriptase inhibitors, and protease inhibitors in the treatment of HIV infections (AIDS) In De Clercq, E. (ed), Advances in pharmacology. Vol 67 Amsterdam, Netherlands: Elsevier; 2013:317‐358. [DOI] [PubMed] [Google Scholar]
- 104. Usach I, Melis V, Peris JE. Non‐nucleoside reverse transcriptase inhibitors: a review on pharmacokinetics, pharmacodynamics, safety and tolerability. J Int AIDS Soc. 2013;16(1):18567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Galmarini C, Mackey J, Dumontet C. Nucleoside analogues: mechanisms of drug resistance and reversal strategies. Leukemia. 2001;15(6):875‐890. [DOI] [PubMed] [Google Scholar]
- 106. Loeb LA, Essigmann JM, Kazazi F, Zhang J, Rose KD, Mullins JI. Lethal mutagenesis of HIV with mutagenic nucleoside analogs. Proc Natl Acad Sci USA. 1999;96(4):1492‐1497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Pierra C, Amador A, Benzaria S, et al. Synthesis and pharmacokinetics of valopicitabine (NM283), an efficient prodrug of the potent anti‐HCV agent 2′‐C‐methylcytidine. J Med Chem. 2006;49(22):6614‐6620. [DOI] [PubMed] [Google Scholar]
- 108. Gardelli C, Attenni B, Donghi M, et al. Phosphoramidate prodrugs of 2′‐C‐methylcytidine for therapy of hepatitis C virus infection. J Med Chem. 2009;52(17):5394‐5407. [DOI] [PubMed] [Google Scholar]
- 109. Rocha‐Pereira J, Jochmans D, Dallmeier K, et al. Inhibition of norovirus replication by the nucleoside analogue 2′‐C‐methylcytidine. Biochem Biophys Res Commun. 2012;427(4):796‐800. [DOI] [PubMed] [Google Scholar]
- 110. Rocha‐Pereira J, Jochmans D, Debing Y, Verbeken E, Nascimento MSJ, Neyts J. The viral polymerase inhibitor 2′‐C‐methylcytidine inhibits Norwalk virus replication and protects against norovirus‐induced diarrhea and mortality in a mouse model. J Virol. 2013;87(21):11798‐11805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Costantini VP, Whitaker T, Barclay L, et al. Antiviral activity of nucleoside analogues against norovirus. Antivir Ther. 2012;17(6):981‐991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Rocha‐Pereira J, Jochmans D, Neyts J. Prophylactic treatment with the nucleoside analogue 2′‐C‐methylcytidine completely prevents transmission of norovirus. J Antimicrob Chemother. 2015;70(1):190‐197. [DOI] [PubMed] [Google Scholar]
- 113. Furuta Y, Gowen BB, Takahashi K, Shiraki K, Smee DF, Barnard DL. Favipiravir (T‐705), a novel viral RNA polymerase inhibitor. Antiviral Res. 2013;100(2):446‐454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. Arias A, Thorne L, Goodfellow I. Favipiravir elicits antiviral mutagenesis during virus replication in vivo. ELife. 2014;3:e03679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Baranovich T, Wong SS, Armstrong J, et al. T‐705 (favipiravir) induces lethal mutagenesis in influenza A H1N1 viruses in vitro. J Virol. 2013;87(7):3741‐3751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Kiso M, Takahashi K, Sakai‐Tagawa Y, et al. T‐705 (favipiravir) activity against lethal H5N1 influenza A viruses. Proc Natl Acad Sci USA. 2010;107(2):882‐887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Oestereich L, Lüdtke A, Wurr S, Rieger T, Muñoz‐Fontela C, Günther S. Successful treatment of advanced Ebola virus infection with T‐705 (favipiravir) in a small animal model. Antiviral Res. 2014;105:17‐21. [DOI] [PubMed] [Google Scholar]
- 118. Rocha‐Pereira J, Jochmans D, Dallmeier K, Leyssen P, Nascimento MSJ, Neyts J. Favipiravir (T‐705) inhibits in vitro norovirus replication. Biochem Biophys Res Commun. 2012;424(4):777‐780. [DOI] [PubMed] [Google Scholar]
- 119. Jin Z, Tucker K, Lin X, et al. Biochemical evaluation of the inhibition properties of favipiravir and 2′‐C‐methyl‐cytidine triphosphates against human and mouse norovirus RNA polymerases. Antimicrob Agents Chemother. 2015;59(12):7504‐7516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Snell NJC. Ribavirin—current status of a broad spectrum antiviral agent. Expert Opin Pharmacother. 2001;2(8):1317‐1324. [DOI] [PubMed] [Google Scholar]
- 121. Chang KO, George DW. Interferons and ribavirin effectively inhibit Norwalk virus replication in replicon‐bearing cells. J Virol. 2007;81(22):12111‐12118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Alam I, Lee JH, Cho KJ, et al. Crystal structures of murine norovirus‐1 RNA‐dependent RNA polymerase in complex with 2‐thiouridine or ribavirin. Virology. 2012;426(2):143‐151. [DOI] [PubMed] [Google Scholar]
- 123. Feld JJ, Hoofnagle JH. Mechanism of action of interferon and ribavirin in treatment of hepatitis C. Nature. 2005;436(7053):967‐972. [DOI] [PubMed] [Google Scholar]
- 124. Crotty S, Cameron C, Andino R. Ribavirin's antiviral mechanism of action: lethal mutagenesis? J Mol Med. 2002;80(2):86‐95. [DOI] [PubMed] [Google Scholar]
- 125. Crotty S, Cameron CE, Andino R. RNA virus error catastrophe: direct molecular test by using ribavirin. Proc Natl Acad Sci USA. 2001;98(12):6895‐6900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Crotty S, Andino R. Implications of high RNA virus mutation rates: lethal mutagenesis and the antiviral drug ribavirin. Microb Infect. 2002;4(13):1301‐1307. [DOI] [PubMed] [Google Scholar]
- 127. Crotty S, Maag D, Arnold JJ, et al. The broad‐spectrum antiviral ribonucleoside ribavirin is an RNA virus mutagen. Nature Med. 2000;6(12):1375‐1379. [DOI] [PubMed] [Google Scholar]
- 128.Chimerix. Chimerix announces discovery and demonstrated preclinical activity supporting ongoing phase 1 study of new antiviral for treatment and prevention of norovirus. 2018. http://ir.chimerix.com/news-releases/news-release-details/chimerix-announces-discovery-and-demonstrated-preclinical. Accessed June 30, 2018.
- 129. Caillet‐Saguy C, Simister PC, Bressanelli S. An objective assessment of conformational variability in complexes of hepatitis C virus polymerase with non‐nucleoside inhibitors. J Mol Biol. 2011;414(3):370‐384. [DOI] [PubMed] [Google Scholar]
- 130. Mastrangelo E, Pezzullo M, Tarantino D, et al. Structure‐based inhibition of norovirus RNA‐dependent RNA polymerases. J Mol Biol. 2012;419(3‐4):198‐210. [DOI] [PubMed] [Google Scholar]
- 131. Tarantino D, Pezzullo M, Mastrangelo E, et al. Naphthalene‐sulfonate inhibitors of human norovirus RNA‐dependent RNA‐polymerase. Antiviral Res. 2014;102:23‐28. [DOI] [PubMed] [Google Scholar]
- 132. Netzler NE, Enosi Tuipulotu D, Eltahla AA, et al. Broad‐spectrum non‐nucleoside inhibitors for caliciviruses. Antiviral Res. 2017;146:65‐75. [DOI] [PubMed] [Google Scholar]
- 133. Croci R, Tarantino D, Milani M, et al. PPNDS inhibits murine Norovirus RNA‐dependent RNA‐polymerase mimicking two RNA stacking bases. FEBS Lett. 2014;588(9):1720‐1725. [DOI] [PubMed] [Google Scholar]
- 134. Beindl W, Mitterauer T, Hohenegger M, Ijzerman AP, Nanoff C, Freissmuth M. Inhibition of receptor/G protein coupling by suramin analogues. Mol Pharmacol. 1996;50(2):415‐423. [PubMed] [Google Scholar]
- 135. Mastrangelo E, Mazzitelli S, Fabbri J, et al. Delivery of suramin as an antiviral agent through liposomal systems. ChemMedChem. 2014;9(5):933‐939. [DOI] [PubMed] [Google Scholar]
- 136. Croci R, Pezzullo M, Tarantino D, et al. Structural bases of norovirus RNA dependent RNA polymerase inhibition by novel suramin‐related compounds. PLOS One. 2014;9(3):e91765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137. Simeonov A, Kulkarni A, Dorjsuren D, et al. Identification and characterization of inhibitors of human apurinic/apyrimidinic endonuclease APE1. PLOS One. 2009;4(6):e5740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Rocha‐Pereira J, Nascimento MSJ, Ma Q, Hilgenfeld R, Neyts J, Jochmans D. The enterovirus protease inhibitor rupintrivir exerts cross‐genotypic anti‐norovirus activity and clears cells from the norovirus replicon. Antimicrob Agents Chemother. 2014;58(8):4675‐4681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Kitano M, Hosmillo M, Emmott E, Lu J, Goodfellow I. Selection and characterization of rupintrivir‐resistant Norwalk virus replicon cells in vitro. Antimicrob Agents Chemother. 2018;62:pii: e00201‐18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Hayden FG, Turner RB, Gwaltney JM, et al. Phase II, randomized, double‐blind, placebo‐controlled studies of ruprintrivir nasal spray 2‐percent suspension for prevention and treatment of experimentally induced rhinovirus colds in healthy volunteers. Antimicrob Agents Chemother. 2003;47(12):3907‐3916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Mandadapu SR, Gunnam MR, Galasiti Kankanamalage AC, et al. Potent inhibition of norovirus by dipeptidyl α‐hydroxyphosphonate transition state mimics. Bioorg Med Chem Lett. 2013;23(21):5941‐5944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142. Tiew K‐C, He G, Aravapalli S, et al. Design, synthesis, and evaluation of inhibitors of Norwalk virus 3C protease. Bioorg Med Chem Lett. 2011;21(18):5315‐5319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Hardy ME, Crone TJ, Brower JE, Ettayebi K. Substrate specificity of the Norwalk virus 3C‐like proteinase. Virus Res. 2002;89(1):29‐39. [DOI] [PubMed] [Google Scholar]
- 144. Galasiti Kankanamalage AC, Kim Y, Weerawarna PM, et al. Structure‐guided design and optimization of dipeptidyl inhibitors of norovirus 3CL protease. Structure–activity relationships and biochemical, X‐ray crystallographic, cell‐based, and in vivo studies. J Med Chem. 2015;58(7):3144‐3155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145. Muhaxhiri Z, Deng L, Shanker S, et al. Structural basis of substrate specificity and protease inhibition in Norwalk virus. J Virol. 2013;87(8):4281‐4292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Mandadapu SR, Weerawarna PM, Gunnam MR, et al. Potent inhibition of norovirus 3CL protease by peptidyl α‐ketoamides and α‐ketoheterocycles. Bioorg Med Chem Lett. 2012;22(14):4820‐4826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Mandadapu SR, Gunnam MR, Tiew K‐C, et al. Inhibition of norovirus 3CL protease by bisulfite adducts of transition state inhibitors. Bioorg Med Chem Lett. 2013;23(1):62‐65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Takahashi D, Kim Y, Lovell S, Prakash O, Groutas WC, Chang K‐O. Structural and inhibitor studies of norovirus 3C‐like proteases. Virus Res. 2013;178(2):437‐444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Galasiti Kankanamalage AC, Kim Y, Rathnayake AD, et al. Design, synthesis, and evaluation of novel prodrugs of transition state inhibitors of norovirus 3CL protease. J Med Chem. 2017;60(14):6239‐6248. [DOI] [PubMed] [Google Scholar]
- 150. Damalanka VC, Kim Y, Alliston KR, et al. Oxadiazole‐based cell permeable macrocyclic transition state inhibitors of norovirus 3CL protease. J Med Chem. 2016;59(5):1899‐1913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Damalanka VC, Kim Y, Galasiti kankanamalage AC, et al. Design, synthesis, and evaluation of a novel series of macrocyclic inhibitors of norovirus 3CL protease. Eur J Med Chem. 2017;127:41‐61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Damalanka VC, Kim Y, Galasiti kankanamalage AC, et al. Structure‐guided design, synthesis and evaluation of oxazolidinone‐based inhibitors of norovirus 3CL protease. Eur J Med Chem. 2017;127:41‐61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Weerawarna PM, Kim Y, Galasiti kankanamalage AC, et al. Structure‐based design and synthesis of triazole‐based macrocyclic inhibitors of norovirus protease: Structural, biochemical, spectroscopic, and antiviral studies. Eur J Med Chem. 2016;119:300‐318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Mandadapu SR, Weerawarna PM, Prior AM, et al. Macrocyclic inhibitors of 3C and 3C‐like proteases of picornavirus, norovirus, and coronavirus. Bioorg Med Chem Lett. 2013;23(13):3709‐3712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155. Kaufmann SHE, Dorhoi A, Hotchkiss RS, Bartenschlager R. Host‐directed therapies for bacterial and viral infections. Nat Rev Drug Discov. 2018;17(1):35‐56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156. Charbonneau M‐E, Gonzalez‐Hernandez MJ, Showalter HD, Donato NJ, Wobus CE, O’Riordan MXD. Small molecule deubiquitinase inhibitors promote macrophage anti‐infective capacity. PLOS One. 2014;9(8):e104096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157. Gonzalez‐Hernandez MJ, Pal A, Gyan KE, et al. Chemical derivatives of a small molecule deubiquitinase inhibitor have antiviral activity against several RNA viruses. PLOS One. 2014;9(4):e94491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Perry JW, Ahmed M, Chang K‐O, Donato NJ, Showalter HD, Wobus CE. Antiviral activity of a small molecule deubiquitinase inhibitor occurs via induction of the unfolded protein response. PLOS Pathog. 2012;8(7):e1002783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159. Luo H. Interplay between the virus and the ubiquitin–proteasome system: molecular mechanism of viral pathogenesis. Curr Opin Virol. 2016;17:1‐10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160. Li J, Soroka J, Buchner J. The Hsp90 chaperone machinery: conformational dynamics and regulation by co‐chaperones. Biochim Biophys Acta. 2012;1823(3):624‐635. [DOI] [PubMed] [Google Scholar]
- 161. Vashist S, Urena L, Gonzalez‐Hernandez MB, et al. Molecular chaperone Hsp90 is a therapeutic target for noroviruses. J Virol. 2015;89(12):6352‐6363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162. Engel AL, Holt GE, Lu H. The pharmacokinetics of Toll‐like receptor agonists and the impact on the immune system. Expert Rev Clin Pharmacol. 2011;4(2):275‐289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163. Hwang S, Maloney NS, Bruinsma MW, et al. Nondegradative role of Atg5‐Atg12/Atg16L1 autophagy protein complex in antiviral activity of interferon gamma. Cell Host Microbe. 2012;11(4):397‐409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164. Maloney NS, Thackray LB, Goel G, et al. Essential cell‐autonomous role for interferon (IFN) regulatory factor 1 in IFN‐γ‐mediated inhibition of norovirus replication in macrophages. J Virol. 2012;86(23):12655‐12664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165. Thackray LB, Duan E, Lazear HM, et al. Critical role for interferon regulatory factor 3 (IRF‐3) and IRF‐7 in type I interferon‐mediated control of murine norovirus replication. J Virol. 2012;86(24):13515‐13523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166. Changotra H, Jia Y, Moore TN, et al. Type I and type II interferons inhibit the translation of murine norovirus proteins. J Virol. 2009;83(11):5683‐5692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167. Chang K‐O, George DW. Interferons and ribavirin effectively inhibit Norwalk virus replication in replicon‐bearing cells. J Virol. 2007;81(22):12111‐12118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168. McCartney SA, Thackray LB, Gitlin L, Gilfillan S, Virgin iv HW, Colonna M. MDA‐5 recognition of a murine norovirus. PLOS Pathog. 2008;4(7):e1000108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169. Nice TJ, Robinson BA, Van Winkle JA. The role of interferon in persistent viral infection: insights from murine norovirus. Trends Microbiol. 2017;26:510‐524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170. Kohli A, Zhang X, Yang J, et al. Distinct and overlapping genomic profiles and antiviral effects of Interferon‐λ and‐α on HCV‐infected and noninfected hepatoma cells. J Viral Hepat. 2012;19(12):843‐853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171. Bolen CR, Ding S, Robek MD, Kleinstein SH. Dynamic expression profiling of type I and type III interferon‐stimulated hepatocytes reveals a stable hierarchy of gene expression. Hepatology. 2014;59(4):1262‐1272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172. Sheppard P, Kindsvogel W, Xu W, et al. IL‐28, IL‐29 and their class II cytokine receptor IL‐28R. Nat Immunol. 2003;4(1):63‐68. [DOI] [PubMed] [Google Scholar]
- 173. Kotenko SV, Gallagher G, Baurin VV, et al. IFN‐λs mediate antiviral protection through a distinct class II cytokine receptor complex. Nat Immunol. 2003;4(1):69‐77. [DOI] [PubMed] [Google Scholar]
- 174. Nice TJ, Baldridge MT, McCune BT, et al. Interferon‐λ cures persistent murine norovirus infection in the absence of adaptive immunity. Science. 2015;347(6219):269‐273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175. Baldridge MT, Lee S, Brown JJ, et al. Expression of Ifnlr1 on intestinal epithelial cells is critical to the antiviral effects of interferon lambda against norovirus and reovirus. J Virol. 2017;91(7):pii: e02079‐16‐. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176. Rocha‐Pereira J, Jacobs S, Noppen S, Verbeken E, Michiels T, Neyts J. Interferon lambda (IFN‐λ) efficiently blocks norovirus transmission in a mouse model. Antiviral Res. 2018;149:7‐15. [DOI] [PubMed] [Google Scholar]
- 177. Lanford RE, Guerra B, Chavez D, et al. GS‐9620, an oral agonist of Toll‐like receptor‐7, induces prolonged suppression of hepatitis B virus in chronically infected chimpanzees. Gastroenterology. 2013;144(7):1508‐1517. e1510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178. Bam RA, Hansen D, Irrinki A, et al. TLR7 agonist GS‐9620 is a potent inhibitor of acute HIV‐1 infection in human peripheral blood mononuclear cells. Antimicrob Agents Chemother. 2017;61(1):pii: e01369‐16‐. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179. Tuipulotu DE, Netzler NE, Lun JH, Mackenzie JM, White PA. TLR7 agonists display potent antiviral effects against norovirus infection via innate stimulation. Antimicrob Agents Chemother. 2018;62: pii: e02417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180. Lee W, Kim M, Lee S‐H, Jung H‐G, Oh J‐W. Prophylactic efficacy of orally administered Bacillus poly‐γ‐glutamic acid, a non‐LPS TLR4 ligand, against norovirus infection in mice. Sci Rep. 2018;8(1):8667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181. Dowling JK, Mansell A. Toll‐like receptors: the swiss army knife of immunity and vaccine development. Clin Transl Immunol. 2016;5(5). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182. Moore RA, Edwards JE, Hopwood J, Hicks D. Imiquimod for the treatment of genital warts: a quantitative systematic review. BMC Infect Dis. 2001;1(1):3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183. Ohba M, Oka T, Ando T, et al. Antiviral effect of theaflavins against caliciviruses. J Antibiot (Tokyo). 2017;70(4):443‐447. [DOI] [PubMed] [Google Scholar]
- 184. Seo DJ, Choi C. Inhibitory mechanism of five natural flavonoids against murine norovirus. Phytomedicine. 2017;30:59‐66. [DOI] [PubMed] [Google Scholar]
- 185. Seo DJ, Jeon SB, Oh H, et al. Comparison of the antiviral activity of flavonoids against murine norovirus and feline calicivirus. Food Control. 2016;60:25‐30. [Google Scholar]
- 186. Su X, D’Souza DH. Naturally occurring flavonoids against human norovirus surrogates. Food Environ Virol. 2013;5(2):97‐102. [DOI] [PubMed] [Google Scholar]
- 187. Yang M, Lee G, Si J, Lee S‐J, You H, Ko G. Curcumin shows antiviral properties against norovirus. Molecules. 2016;21(10):1401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188. Lee H, Ko G. Antiviral effect of vitamin A on norovirus infection via modulation of the gut microbiome. Sci Rep. 2016;6:25835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189. Fumian TM, Tuipulotu DE, Netzler NE, et al. Potential therapeutic agents for feline calicivirus infection. Viruses. 2018;10(8). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190. Dang W, Yin Y, Peppelenbosch MP, Pan Q. Opposing effects of nitazoxanide on murine and human norovirus. J Infect Dis. 2017;216(6):780‐782. [DOI] [PubMed] [Google Scholar]
- 191. Dang W, Xu L, Ma B, et al. Nitazoxanide inhibits human norovirus replication and synergizes with ribavirin by activation of cellular antiviral response. Antimicrob Agents Chemother. 2018:AAC.00707‐18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192. Fox LM, Saravolatz LD. Nitazoxanide: a new thiazolide antiparasitic agent. Clin Infect Dis. 2005;40(8):1173‐1180. [DOI] [PubMed] [Google Scholar]
- 193. Rossignol JF. Nitazoxanide: a first‐in‐class broad‐spectrum antiviral agent. Antiviral Res. 2014;110:94‐103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194. Rossignol JF, El‐Gohary YM. Nitazoxanide in the treatment of viral gastroenteritis: a randomized double‐blind placebo‐controlled clinical trial. Aliment Pharmacol Ther. 2006;24(10):1423‐1430. [DOI] [PubMed] [Google Scholar]
- 195. Siddiq DM, Koo HL, Adachi JA, Viola GM. Norovirus gastroenteritis successfully treated with nitazoxanide. J Infect. 2011;63(5):394‐397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196. Haubrich K, Gantt S, Blydt‐Hansen T. Successful treatment of chronic norovirus gastroenteritis with nitazoxanide in a pediatric kidney transplant recipient. Pediatr Tranplant. 2018;22:e13186. [DOI] [PubMed] [Google Scholar]
- 197. Capizzi T, Makari‐Judson G, Steingart R, Mertens WC. Chronic diarrhea associated with persistent norovirus excretion in patients with chronic lymphocytic leukemia: report of two cases. BMC Infect Dis. 2011;11(1):131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198. Jurgens PT, Allen LA, Ambardekar AV, McIlvennan CK. Chronic norovirus infections in cardiac transplant patients: considerations for evaluation and management. Prog Transplant. 2017;27(1):69‐72. [DOI] [PubMed] [Google Scholar]
- 199. Kempf B, Edgar JD, Mc Caughey C, Devlin LA. Nitazoxanide is an ineffective treatment of chronic norovirus in patients with x‐linked agammaglobulinemia and may yield false‐negative polymerase chain reaction findings in stool specimens. J Infect Dis. 2017;215(3):486‐487. [DOI] [PubMed] [Google Scholar]
- 200. Echenique IA, Stosor V, Gallon L, Kaufman D, Qi C, Zembower TR. Prolonged norovirus infection after pancreas transplantation: a case report and review of chronic norovirus. Tranpl Infect Dis. 2016;18(1):98‐104. [DOI] [PubMed] [Google Scholar]