

Abstract
Background
R2R3-MYB transcription factors (TFs) play important roles in plant growth and development, and response to biotic and abiotic stresses. However, their regulatory mechanisms in wound-induced anthocyanin biosynthesis in woody plants are largely unknown.
Results
In this work, we report that expression of anthocyanin biosynthesis genes (ABGs) were activated by PdMYB118, a MYB TF encoding gene from Populus deltoids, and the activation of PdMYB118 was significantly enhanced by PdTT8, a bHLH protein, through its direct interaction with PdMYB118. PdMYB118 and some ABGs were evidently induced by wound induction and methyl jasmonate (MeJA) treatment. Overexpression of PdMYB118 promoted anthocyanin accumulation in transgenic poplar upon wound induction. Furthermore, a poplar JASMONATE ZIM-domain (JAZ) protein, PtrJAZ1, repressed the transcriptional function of PdMYB118/PdTT8 complex by binding to PdTT8, and wound stimulated the biosynthesis of jasmonic acid (JA) and the degradation of PtrJAZ1.
Conclusions
Based on these observations, we proposed that PtrJAZ1 degradation triggered the expression of ABGs, leading to increased biosynthesis of anthocyanins in the wounded leaves of transgenic poplar. Therefore, our findings not only illustrate the crucial role of PdMYB118 in wound-induced anthocyanin biosynthesis in poplar, but also provide a molecular basis for the genetic engineering of colorful tree species.
Keywords: Anthocyanin, JAZ1, PdMYB118, PdTT8, Poplar
Background
As the major pigments, anthocyanins not only provide colors to plant flowers and fruits [1], but also function in plant resistance to different biotic and abiotic stresses [2–12]. In plants, anthocyanins are biosynthesized via the specific branch of flavonoid pathway [1, 13, 14]. In poplar, the biosynthesis of anthocyanins is catalized by a series of enzymes, including chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), flavanone 3′-hydroxylase (F3’H), flavonoid 3′5’-hydroxylase (F3’5’H), dihydroflavonol 4-reductase (DFR) and anthocyanidin synthase (ANS) [7, 8]. The expression of anthocyanin biosynthetic genes (ABGs) are regulated by the MBW complexes, which is composed of two kinds of transcriptional factors (TFs), R2R3-MYB and basic helix-loop-helix (bHLH), and WD40-repeat proteins [15–17]. In Arabidopsis, the MBW complexes are composed of the R2R3-MYB factors PAP1, PAP2, MYB113 and MYB114, the bHLH factors TT8, GL3 and EGL3, and a WD40 protein TTG1 [16]. So far, similar MBW complexes have been identified in different plant species [13, 18–22].
JA is one of the essential phytohormones which plays important functions in the protection of plants from various biotic and abiotic stresses [23, 24]. JA signaling is negatively regulated by JAZs, which inhibit the expression of JA responsive genes by binding to the transcription factors such as MYC2 [25, 26]. In response to environmental stimulus, JA promotes the interaction between JAZs and the SCFCOI1 ubiquitin ligase to trigger JAZ degradation via the ubiquitin/26S proteasome pathway [27]. The rapid activation of JA biosynthesis is stimulated by wound, a common response in plant development amid biotic and abiotic stresses such as insect attack, pathogen infection mechanical damage [23, 24]. In Arabidopsis, anthocyanin biosynthesis is induced by wound [28], and anthocyanin accumulation was regulated by JA signaling via the degradation of JAZ proteins to release bHLH and MYB TFs in the MBW complex [29]. In cotton, F3H and its downstream genes in proanthocyanidins (PAs) biosynthesis were significantly induced by V. dahliae infection and wound [30]. In apple tree, MdWRKY40 functions as a key modulator in the wounding-induced anthocyanin biosynthesis [31].
To date, many anthocyanin associated R2R3-MYB TFs have been isolated from different plants [16, 32, 33]. However, their regulatory mechanisms in wound induced anthocyanin biosynthesis are still largely unknown. Previously, we demonstrated that PdMYB118 regulated the biosynthesis of anthocyanin in poplar [34]. In this work, we report that PdMYB118 functioned in anthocyanin biosynthesis by interacting with PdTT8 to activate the expression of ABGs in poplar. Wound treatment induced JA accumulation, triggered the degradation of PtrJAZ1, and released its repression on the transcriptional activation activity of PdMYB/PdTT8 complex by directly interacting with PdTT8, and then activated the expression of ABGs for anthocyanin biosynthesis.
Results
PdMYB118 is involved in wound induced anthocyanin biosynthesis in poplar
To explore the possible functions of PdMYB118 in anthocyanin biosynthesis in poplar, we previously generated transgenic Shanxin Yang plants [34]. Overexpression of PdMYB118 led to red leafed phenotype in adult transgenic plants grown in both green house and field, whereas young tissue culture plantlets showed green leaves as did the wide type (WT) (Additional file 1: Fig. S1a). When young plantlets were sub-cultured onto new MS medium, their leaves turned red and produced more anthocyanins after being cut off from their mother plants (Additional file 1: Fig. S1b-d). Therefore, wound may have prompted the biosynthesis of anthocyanin in transgenic plants. To confirm this speculation, wound induction was applied to both young leaves of tissue cultured transgenic plantlets and mature leaves of greenhouse grown transgenic plants. As we have hypothesized, both WT and transgenic leaves from tissue cultured plantlets were green, but transgenic leaf discs formed more red speckles 2 days after wound induction (Fig. 1a). Similar results were also observed in the mature leaf discs of greenhouse grown transgenic plants (Fig. 1b). Transgenic leaf discs produced more anthocyanins than did the WT control (Fig. 1c, d). These results indicate that PdMYB118 is involved in wound induced anthocyanin biosynthesis in poplar.
Fig. 1.

Wound induced accumulation of anthocyanins in the leaves of transgenic plants overexpressing PdMYB118. a Wound prompts anthocyanin biosynthesis in the leaves of transgenic plants. The leaves of WT and transgenic plantlets cultured on MS medium were cut into pieces and transferred onto MS medium for 2 days. Images in the third row were the higher magnification of the framed area of the images in the secondary row. Scale bars for the images in the first and secondary rows stand for 1 cm and in the third row stand for 1 mm. b Accumulated anthocyanin biosynthesis in the wounded leaves of transgenic plants grown in greenhouse. Matured leaves were cut into leaf discs and transferred onto MS medium for 2 days. The black arrows indicate the red speckles. Scale bar = 0.5 cm. c Anthocyanin extracted from the wounded leaf discs in (b). Scale bar = 0.5 cm. d Relative content of anthocyanins extracted from the leaves in (b). Control, before wound induction; Wound, after wound induction on MS medium for 2 days; WT, leaves of wild type Shanxin Yang plants; L2 and L7, leaves of transgenic plants overexpressing PdMYB118; WT-W, wounded leaves of wild type plant; L2-W and L7-W, wounded leaves of transgenic plants (Lines L2 and L7). Values are means and standard deviations of three biological replicates (n = 3). ** and ***, significant differences in comparison to WT, L2 and L7 at P < 0.01 and P < 0.001, respectively (Student’s t-test)
Wound induces the expression of ABGs
Based on the observation that wound promoted anthocyanin accumulation in PdMYB118 overexpressing transgenic plants, we postulated that wound signaling may also induce the expression of ABGs in wild type poplar. We found that PdMYB118 transcripts were gradually increased in the leaf discs during wound induction, and reached to the highest transcription level at 24 h (Fig. 2). Some ABGs, including PtrF3’H, PtrDFR2 and PtrANS1, showed similar expression pattern as PdMYB118. A responsive expression of PtrCHS1 to wound induction was also observed: it increased at 1 h, then gradually reduced within 12 h and restored to the high expression level at 24 h. PdTT8 was slight induced in wounded leaves within 3 h, and obviously up-regulated at 24 h. PtrCHI1 and PtrF3H were down-regulated within 3 to 12 h after wound induction and then reached to the normal expression level. The transcripts of PtrF3’5’H2 reached a peak level within 1 h after wound induction and rapidly decreased to lower levels at later time points. From these data, we identified four wound-inducible ABGs: PtrCHS1, PtrF3’H, PtrDFR2 and PtrANS1.
Fig. 2.

qRT-PCR analyses of PdMYB118 and ABGs in the wounded leaves of wild type poplar plants. The mature leaves of poplar plants grown in greenhouse were treated using leaf-disc method for various time intervals for RNA extraction. Gene expression in the untreated leaves (0 h) was set to 1. PtrMYB118 and PtrTT8 are respectively the homology of PdMYB118 and PdTT8, and PtrCHS1, PtrCHI1, PtrF3H, PtrF3’H, PtrF3’5’H, PtrDFR2 and PtrANS1 are the ABG genes respectively encoding chalcone synthase, chalcone isomerase, flavanone 3-hydroxylase, flavanone 3′-hydroxylase, flavonoid 3′5’-hydroxylase, dihydroflavonol 4-reductase and anthocyanidin synthase in Populus. Three independent replicates of measurements were performed for each time point, and the values are means and standard deviations (n = 3). ** and ***, significant differences at P < 0.01 and P < 0.001 (Student’s t-test)
The transcriptional activation activity of PdMYB118 is enhanced by a bHLH TF PdTT8
To clarify the exact function of PdMYB118 in wound induced anthocyanin biosynthesis, we first transiently expressed it in the leaf protoplasts of wild type Shanxin Yang plants. We found that transient expression of PdMYB118 enhanced the transcription of most ABGs, including the wound-/JA-inducibe ABGs: PtrCHS1, PtrF3’H, PtrDFR2 and PtrANS1 (Fig. 3a). In plants, anthocyanin-related MYB factors interact closely with bHLH TFs to control anthocyanin biosynthesis [15]. We cloned a poplar bHLH TF PdTT8, the homolog of MdbHLH3 in apple and TT8 in Arabidopsis, which interacted with different MYB TFs, and investigated its transcription activity. PdTT8 alone did not regulate the expressions of ABGs, but could significantly enhance the transcriptional activation activity of PdMYB118. When PdTT8 was transiently co-expressed with PdMYB118 in the protoplasts, the transcription levels of all ABGs were remarkably higher than those expressing PdMYB118 alone (Fig. 3a). These results imply that PdTT8 could be an efficient enhancer of PdMYB118 to regulate the expression of ABGs.
Fig. 3.
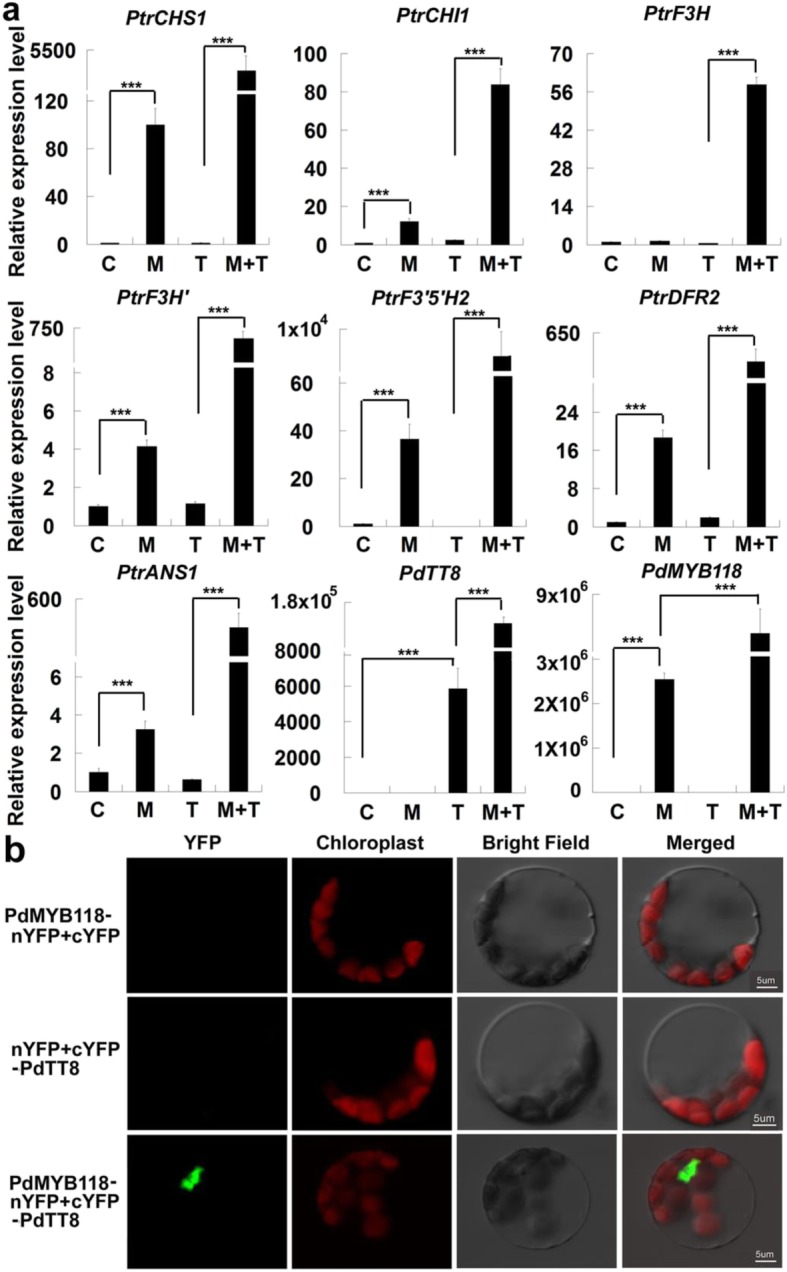
PdTT8 enhances the transcriptional activation activity of PdMYB118. a Transient expression of PdMYB118 or PdTT8 in poplar mesophyll protoplasts. The constructs of pGreenII62-SK-PdMYB118, pGreenII62-SK-PdTT8 or pGreenII62-SK-PdMYB118 + pGreenII62-SK-PdTT8 were transfected into the poplar leaf protoplasts, respectively. The empty vector was used as a negative control. RNA was extracted from the transfected protoplasts for qRT-PCR analyses of ABGs, PdTT8 and PdMYB118. PtrCHS1, PtrCHI1, PtrF3H, PtrF3’H, PtrF3’5’H, PtrDFR2 and PtrANS1 are the ABG genes respectively encoding chalcone synthase, chalcone isomerase, flavanone 3-hydroxylase, flavanone 3′-hydroxylase, flavonoid 3′5’-hydroxylase, dihydroflavonol 4-reductase and anthocyanidin synthase in Populus. C, protoplasts transfected with pGreenII62-SK (control); M, protoplasts transfected with pGreenII62-SK-PdMYB118; T, protoplasts transfected with pGreenII62-SK-PdTT8; M + T; protoplasts transfected with pGreenII62-SK-PdMYB118 and pGreenII62-SK-PdTT8. b BiFC assays to detect the interaction of PdTT8 and PdMYB118. PdMYB118 or PdTT8 was respectively fused with N-terminal and C-terminal fragments of YFP. Construct pairs indicated on the left were co-expressed in the leaf protoplasts of WT poplar plants. Gene expression level in the control sample was set to 1. Values are means and standard deviations of three biological replicates (n = 3). ***, significant difference in comparison to C and T at P < 0.001, respectively (Student’s t-test)
The enhanced transcriptional activation function of PdMYB118 by PdTT8 suggested a possible interaction of these two proteins. Therefore, we performed BiFC assays to confirm this possibility. The N- and C-terminal fragment of yellow fluorescent protein (nYFP and cYFP) was fused with PdMYB118 and PdTT8, respectively. As expected, when PdMYB118-nYFP and cYFP-PdTT8 were co-transfected into poplar leaf mesophyll protoplasts, the nuclei of protoplasts showed strong YFP fluorescence; whereas no signal was detected in the control (Fig. 3b). These results demonstrate that PdTT8 could directly interact with PdMYB118 to enhance its transcriptional activation activity.
Wound prompts JA biosynthesis and JAZ1 degradation
JA biosynthesis is catalyzed by a series of biosynthetic enzymes step by step [26]. It is widely considered that JA is rapidly synthesized in plant leaves suffering wound stimulus [23, 24]. To detect the JA changes in response to wound, we examined the expressions of JA biosynthesis genes PtrAOC2–3, PtrOPR3–1, PtrACX1–2 and PtrJAR1–1. All these genes were rapidly expressed to their peak levels within 1 h after wound induction and then reduced gradually as the treatment elongated (Fig. 4a). The responsive expression of JA biosynthesis genes may result in JA accumulation. We then measured JA and JA-Ile contents at two time points: 1 h (short time) and 24 h (long time) after mechanical damage. JA and JA-Ile contents were about 10 ng g− 1 and 0.58 ng g− 1 fresh weight tissues (FW) in undamaged leaves, respectively. However, after 1 h of wound induction, the contents of JA and JA-Ile increased by about 16 and 200 folds (Fig. 4b). After 24 h, JA and JA-Ile contents reduced to about 1.9 and 0.53 ng g− 1 FW. The changes in JA and JA-Ile contents in wounded poplar leaves were tightly correlated with the expression changes of JA biosynthesis genes.
Fig. 4.

JA biosynthesis gene expression, JA content and JAZ1 degradation analyses. The mature leaves of poplar plants grown in greenhouse were treated using leaf-disc method for various time intervals and then used for RNA extraction. a qRT-PCR analyses of JA biosynthesis genes in the wounded leaves. PtrAOC2–3, a putative allene oxide cyclase gene in Populus; PtrOPR3–1, a putative OPDA reductase 3 gene in Populus; PtrACX1–2, a putative acyl-CoA oxidase gene in Populus; PtrJAR1–1, a putative JASMONATE RESISTANT gene (a JA-Ile biosynthesis gene) in Populus. b JA and JA-Ile content in the wounded leaves of poplar plants grown in greenhouse. The leaves treated for various time intervals were used to be tested. c qRT-PCR analyses of PtrJAZ1 expression in the wounded leaves. d Western blotting analyses of JAZ1 protein in the wounded leaves. Total proteins were extracted from the leaf-discs treated for various time intervals. An amount of 30 μg proteins were separated by 10% SDS-PAGE and hybridized with Arabidopsis JAZ1 antibodies (JAZ1) or plant actin antibodies (Actin), respectively. Each gene expression in the untreated leaves (0 h) was set to 1. Three independent replicates of measurements were performed for each time point, and the values are means and standard deviations (n = 3). * and ***, significant differences at P < 0.05 and P < 0.001 (Student’s t-test). The original uncropped blot image was shown in additional file 6: Fig. S6
It has been reported that JAZ genes are obviously induced by mechanical wounding [23]. We analyzed the expression of a poplar JAZ gene PtrJAZ1 during wound induction, and found that PtrJAZ1 transcripts were quickly up-regulated within 1 h after wound induction, rapidly decreased within 3 h, and dropped to the lowest level after 24 h (Fig. 4c). The change in PtrJAZ1 protein content in response to wound induction was then tested by Western blotting. Consistent with its gene expression variation in wounded leaves, PtrJAZ1 protein accumulation reached to a higher level within 3 h, began to decline after 8 h, and reduced to a lower level after 12 h (Fig. 4d). These results indicate that wound induced anthocyanin biosynthesis in poplar may be mediated by JA signaling.
JA regulates the expressions of ABGs in poplar
We further examined whether JA can regulate the expression of ABGs. Poplar leaves were sprayed with MeJA solution for qRT-PCR analyses. As expected, the expression levels of wound inducible ABGs (PtrCHS1, PtrF3’H, PtrDFR2 and PtrANS1) were up-regulated more than 3 to 20 folds by MeJA treatment (Fig. 5). JA signaling is regulated by JAZ proteins, which inhibit the expression of JA response genes by binding to other transcription factor [35]. We transiently expressed PtrJAZ1 in the leaf protoplasts of PdMYB118 transgenic plants. The up-regulated expression of most ABGs by PdMYB118 was inhibited by the expression of PtrJAZ1 (Fig. 6a, b). These results indicate that JA induced expression of ABGs in poplar is negatively regulated by PtrJAZ1.
Fig. 5.

qRT-PCR analyses of ABGs in MeJA treated leaves. PtrCHS1, PtrCHI1, PtrF3H, PtrF3’H, PtrF3’5’H, PtrDFR2 and PtrANS1 are the ABG genes respectively encoding chalcone synthase, chalcone isomerase, flavanone 3-hydroxylase, flavanone 3′-hydroxylase, flavonoid 3′5’-hydroxylase, dihydroflavonol 4-reductase and anthocyanidin synthase in Populus. Gene expression in the leaves sprayed with water (control) was set to 1. Values are means and standard deviations of three biological replicates (n = 3). ** and ***, significant differences at P < 0.01 and P < 0.001 (Student’s t-test)
Fig. 6.

Expression levels of ABGs in transgenic protoplasts transiently expressing PtrJAZ1. Protoplasts isolated from the leaves of WT and transgenic plants overexpressing PdMYB118 were used for the analyses. a Expression level of PtrJAZ1. b Expression levels of the ABG genes PtrCHS1, PtrCHI1, PtrF3H, PtrF3’H, PtrF3’5’H, PtrDFR2 and PtrANS1 respectively encoding chalcone synthase, chalcone isomerase, flavanone 3-hydroxylase, flavanone 3′-hydroxylase, flavonoid 3′5’-hydroxylase, dihydroflavonol 4-reductase and anthocyanidin synthase in Populus. Gene expression level in the WT was set to 1. WT, wild type protoplasts transfected with pGreenII62-SK; M, transgenic protoplasts transfected with pGreenII62-SK; M + J, transgenic protoplasts transfected with pGreenII62-SK-PtrJAZ1. Values (n = 3) are mean ± SD and different letters on the histograms indicate that the values differ significantly (one-way ANOVA; P < 0.01)
PtrJAZ1 represses the transcriptional activation activity of PdMYB118/PdTT8 complex by binding to PdTT8
The suppressed transcriptional activation activity of PdMYB118 by PtrJAZ1 could be regulated by PdTT8. Therefore, we transiently expressed PdTT8 and PtrJAZ1 alone or together in the leaf protoplasts of PdMYB118 transgenic plants (Additional file 2: Fig. S2a, b). We observed that transient expression of PdTT8 in the leaf protoplasts of PdMYB118 transgenic plants further increased the expression levels of ABGs (Fig. 7a). However, when PtrJAZ1 and PdTT8 were co-expressed in the leaf protoplasts of PdMYB118 transgenic plants, the expression levels of ABGs were repressed to the levels of control samples (Fig. 7a). These results suggest that PtrJAZ1 suppressed the transcription activity of PdMYB118/PdTT8 complex. Then, we performed BiFC assays to examine whether PtrJAZ1 interacts with PdMYB118 or PdTT8. PtrJAZ1-nYFP was transiently co-expressed with cYFP-PdTT8 or cYFP-PdMYB118 in the leaf mesophyll protoplasts of wide type poplar. Co-expression of PtrJAZ1-nYFP with cYFP-PdTT8 produced strong YFP fluorescence in the nuclei of protoplasts, whereas no fluorescence signal was detected in the protoplasts co-expressing PtrJAZ1-nYFP and cYFP-PdMYB118 (Fig. 7b; Additional file 3: Fig. S3). This result is consistent with the finding in apple that MdJAZs interact with MdbHLH3 (the analogue of TT8) but not with the PA biosynthesis associated MdMYB9/11 [36].
Fig. 7.

PtrJAZ1 binds to PdTT8 to inhibit the transcriptional activation activity of the PdMYB118/PdTT8 complex. PtrCHS1, PtrCHI1, PtrF3H, PtrF3’H, PtrF3’5’H, PtrDFR2 and PtrANS1 are the ABG genes respectively encoding chalcone synthase, chalcone isomerase, flavanone 3-hydroxylase, flavanone 3′-hydroxylase, flavonoid 3′5’-hydroxylase, dihydroflavonol 4-reductase and anthocyanidin synthase in Populus. a PtrJAZ1 represses the transcriptional activation activity of the PdMYB118/PdTT8 complex. Protoplasts isolated from the leaves of transgenic plants overexpressing PdMYB118 were transformed with pGreenII62-SK-PdTT8 alone, or co-transformed with pGreenII62-SK-PdTT8 and pGreenII62-SK-PtrJAZ1. qRT-PCR analysis of ABGs in the transfected protoplasts was performed. The empty vector was used as a negative control and the gene expression level in the control was set to 1. Values are means and standard deviations of three biological replicates (n = 3). ** and ***, significant differences at P < 0.01 and P < 0.001 (Student’s t-test). M, transgenic protoplasts transfected with pGreenII62-SK; T, transgenic protoplasts transfected with pGreenII62-SK-PdTT8; T + J, transgenic protoplasts co-transfected with pGreenII62-SK-PdTT8 and pGreenII62-SK-PtrJAZ1. b BiFC assays to test the interaction of PtrJAZ1 with PdTT8. PtrJAZ1 was fused with the N-terminal fragment of YFP, and PdTT8 was linked to the C-terminal fragment of YFP, respectively. Construct pairs indicated on the left were co-expressed in poplar leaf protoplasts. Scale bar = 10 μm. c-e Transient transcription dual-luciferase assays to show that PtrJAZ1 inhibits the transcriptional activation activity of the MYB118/TT8 complex to the promoters of PtrCHS1 (c), PtrDFR2 (d) and PtrANS1 (e). The effectors (pGreenII62-SK-PdMYB118, pGreenII62-SK-PdTT8, pGreenII62-SK-PdMYB118 + pGreenII62-SK-PdTT8 or pGreenII62-SK-PdMYB118 + pGreenII62-SK-PdTT8 + pGreenII62-SK-PtrJAZ1) and reporters (35S::REN-ProPtrCHS1::LUC, 35S::REN-ProPtrDFR2::LUC or 35S::REN-ProPtrANS1::LUC) were co-expressed in poplar protoplasts. The expression level of REN was used as an internal control. The LUC/REN ratio represents relative activity of the PtrCHS1, PtrDFR2 and PtrANS1 promoters. Error bars represent the SDs from three biological replicates. C, protoplasts transfected with pGreenII62-SK (control); M, protoplasts transfected with pGreenII62-SK-PdMYB118; T, protoplasts transfected with pGreenII62-SK-PdTT8; M + T, protoplasts transfected with pGreenII62-SK-PdMYB118 and pGreenII62-SK-PdTT8; M + T + J, protoplasts transfected with pGreenII62-SK-PdMYB118, pGreenII62-SK-PdTT8 and pGreenII62-SK-PtrJAZ1. Values are means and standard deviations of three biological replicates (n = 3). ***, significant differences at P < 0.001 (Student’s t-test)
We further confirmed the inhibition of PtrJAZ1 on the transcriptional activation activity of PdMYB118/PdTT8 complex by dual-luciferase assays. Three wound inducible ABGs (PtrCHS1, PtrDFR2 and PtrANS1) were selected. The transcriptional activation activity of PdMYB118 on the promoters of PtrCHS1, PtrDFR2 and PtrANS1 was higher than that of PdTT8, showing a higher ratio of LUC/REN (Fig. 7c-e). The highest ratio values were observed in the protoplasts co-expressing PdMYB118 and PdTT8. And the transcription activity of PdMYB118/PdTT8 complex decreased obviously when PtrJAZ1 was co-transformed with PdMYB118 and PdTT8 (Fig. 7c-e). These results suggest that by binding to PdTT8, PtrJAZ1 could restrain the transcriptional activation activity of PdMYB118/PdTT8 complex.
Degradation of JAZ1 proteins promotes anthocyanin biosynthesis in the wounded leaves of transgenic plants
Based on the observations that expression of ABGs was inhibited by PtrJAZ1 and JAZ1 proteins were degraded upon wound induction, we speculated that the increased anthocyanin biosynthesis in the wounded transgenic leaves may be a result of JAZ1 degradation. By Western blotting analysis using Arabidopsis JAZ1 antibodies, we found that JAZ1 accumulation in the wounded leaves of transgenic plants were distinctly lower than that in the no-treated control leaves (Fig. 8a). Therefore, the decreased JAZ1 protein level may have released the transcriptional activation activity of the MYB118/TT8 complex and increased the expression of wound-inducible ABGs. We further analyzed the expression of ABGs in the leaves of transgenic plants overexpressing PdMYB118. In the untreated leaves of transgenic plants (lines L2 and L7), wound−/JA-inducible ABGs (PtrCHS1, PtrDFR2, PtrF3H and PtrANS1) were up-regulated in transgenic plants. Upon wound induction, their expression levels increased in the leaves of both WT and transgenic plants, with a more significant increase in the wounded leaves of transgenic plants (Fig. 8b). Similarly, the expression level of PdMYB118 increased in the wounded leaves of both WT and transgenic plants, but the increase was significantly higher in transgenic plants (Additional file 4: Fig. S4a). However, the expression level of PdTT8 was only slightly changed in the leaves of both WT and transgenic leaves (Additional file 4: Fig. S4b). These results indicate that the increased anthocyanin accumulation in the wounded leaves of transgenic plants could be due to the enhanced expression of ABGs resulted from the wound-induced degradation of JAZ1 proteins.
Fig. 8.

JAZ1 protein content and ABG expressions in the wounded leaves of transgenic poplar plants. PtrCHS1, PtrCHI1, PtrF3H, PtrF3’H, PtrF3’5’H, PtrDFR2 and PtrANS1 are the ABG genes respectively encoding chalcone synthase, chalcone isomerase, flavanone 3-hydroxylase, flavanone 3′-hydroxylase, flavonoid 3′5’-hydroxylase, dihydroflavonol 4-reductase and anthocyanidin synthase in Populus. a Western blotting analysis of JAZ1 contents. Total proteins were extracted from the untreated or treated leaves of PdMYB118 overexpressing plants. JAZ1 antibodies (JAZ1) and plant actin antibodies (Actin) were used for the hybridization. The original uncropped blot image was shown in additional file 7: Fig. S7. b qRT-PCR analyses of ABGs in the untreated or treated leaves of wild type and PdMYB118 overexpressing plants. The leaves were mechanically wounded by leaf-disc method for 2 days. Then RNA was extracted for qRT-PCR analyses. Gene expression level in wide type leaves was set to 1. Values are means and standard deviations of three biological replicates (n = 3). ** and ***, significant differences at P < 0.01 and P < 0.001 (Student’s t-test). WT, leaves of wide type poplar plant; L2 and L7, leaves of transgenic plants overexpressing PdMYB118; WT-W, wounded leaves of wide type poplar; L2-W and L7-W, wounded leaves of transgenic plants
Discussion
Although wound induced anthocyanin accumulation has been widely observed in plants, the molecular mechanism that how wound regulates anthocyanin biosynthesis in poplar still remains unknown [28, 37]. Recently, we reported that PdMYB118 regulated the biosynthesis of anthocyanin in poplar [34]. Overexpression of PdMYB118 obviously activated the expression of anthocyanin biosynthesis genes but not the two proanthocyanin synthesis genes, leading to red leafed phenotype in transgenic plants. Interestingly, during the process of shoot propagation, we observed that the leaves of tissue cultured plantlets or green house grown young transgenic plants remained in green or less red color, but once the shoots were cut off, the leaves of new cut shoots turn into red color (Additional file 1: Fig. S1a-d). The stability of this phenomenon (wound-induced anthocyanin biosynthesis) was further confirmed using green leaves of both tissue cultured plantlets and greenhouse grown plants (Fig. 1a-d).
Previous studies have showed that JA regulates WD-repeat/bHLH/MYB (WBM) complex-mediated anthocyanin accumulation via the interaction of JAZ proteins with bHLH and MYB transcription factors [29]. PA-specific R2R3-MYB factors like MYB134 in poplar and MdMYB9/11 in apple dramatically responded to wound treatment, whereas the anthocyanin biosynthesis R2R3-MYB factor MdMYB1 in apple did not [8, 36]. MYB/bHLH complex, such as PAP1/TT8 in Arabidopsis, AN4/AN1 in Petunia hybrid, VvMYBA1/VvMYC1 in grape, and MdMYB10/MdbHLH3 in apple, are also involved in anthocyanin synthesis [13, 38–40]. We found that expression of most ABGs was responsive to wound treatment (Fig. 2). The poplar bHLH TF PdTT8 physically interacted with PdMYB118 to form the PdMYB118/PdTT8 complex, and more efficiently activated the expressions of ABGs (Fig. 3a, b). Therefore, PdTT8 may function as an enhancer to promote the transcriptional activation activity of PdMYB118.
Wound can rapidly activate the biosynthesis of JA, which functions as an important plant defense signal in response to various stresses [23, 24]. In plants, the biosynthetic pathway of JA is initiated from the tri-unsaturated fatty acids α-linolenic acid (18:3) [26]. After two oxidation steps, α-linolenic acid is converted to an unstable allene oxide, which is subsequently catalyzed to 12-oxophytodienoic acid (OPDA) by an allene oxide cyclase (AOC) in chloroplasts. Then the second half of JA biosynthesis is finished in peroxisomes, where OPDA is reduced to OPC-8 by OPDA reductase 3 (OPR3) [41]. After β-oxidation, JA is transported to cytosol, where it is conjugated to amino acids by JA-amino acid synthetase (JAR1) to form jasmonoyl-isoleucine (JA-Ile), a major biologically active jasmonate among a growing number of jasmonate derivatives. We found that wound treatment rapidly up-regulated the expression of JA biosynthesis genes within 1 h (Fig. 4a), leading to the rapid output of JA and JA-Ile in the wounded leaves (Fig. 4b). These results indicate that JA biosynthesis responds rapidly to wound induction in poplar. Increased JA biosynthesis by wound treatment could activate the JA signaling. Indeed, similar to the expression pattern of JA biosynthesis gene, PtrJAZ1 was also up-regulated within 1 h after the initiation of wound induction, and then gradually decreased (Fig. 4c). This result is consistent with previous reports that most JAZ genes were strongly induced within 1 h after wound induction and declined at later time points [23, 24]. JA signaling also promotes the degradation of JAZ transcriptional repressors through the activity of the E3 ubiquitin-ligase SCFCOI1 JAZs [27]. We found that the content of poplar JAZ proteins was also obviously decreased after 8 h of wound treatment (Fig. 4d). Therefore, in poplar, wound induction could rapidly induce JA biosynthesis by activating its biosynthesis genes and promote the degradation of JAZ protein.
JA regulates several anthocyanin biosynthetic genes involved in anthocyanin accumulation [42, 43]. In poplar, MeJA treatment could induce the expression of some ABGs such as PtrCHS1, PtrF3H’, PtrDFR2 and PtrANS1 (Fig. 5). Interestingly, expressions of these ABGs were also induced by wound, indicating that wound-induced anthocyanin biosynthesis may be mediated by JA signaling. It has been reported that JA regulates WD-repeat/bHLH/MYB complex-mediated anthocyanin accumulation via the interaction of JAZ proteins with bHLH and MYB factors [29]. We found that PtrJAZ1 specifically bound to PdTT8 and attenuated the transcriptional activation activity of PdMYB118/PdTT8 complex on the promoter of JA/wound-inducible ABGs (Fig. 7a-e). JAZ regulated expression of ABGs in poplar is similar to that in Arabidopsis [29], but different from that in apple, where MdJAZ2 inhibits the recruitment of MdbHLH3 to the promoters of MdMYB9 and MdMYB11 which regulate the biosynthesis of anthocyanin and proanthocyanin [24]. Therefore, JA induced JAZ protein degradation could abolish the interaction of JAZ proteins with bHLH and MYB factors, and then activate the biosynthesis of anthocyanin [29]. In agreement with this speculation, when PtJAZ1 protein was degraded in the wounded leaves of transgenic plants, JA−/wound-inducible ABGs were highly induced to promote anthocyanin biosynthesis (Fig. 8a, b).
Conclusions
Taken together, our study illustrates the biological function of PdMYB118 in wound-induced anthocyanin synthesis in poplar. Although more detailed studies are still needed to completely understand the mechanism of PdMYB118 in anthocyanin synthesis, our data presented here imply a possible model of wound-induced anthocyanin synthesis in poplar: upon wound induction, JA biosynthetic genes are rapidly activated for JA biosynthesis, subsequently triggered the degradation of PtrJAZ1; Then, the transcriptional activation activity of PdMYB118/PdTT8 complex is restored to promote the expression of anthocyanin biosynthesis genes; Finally, anthocyanins were biosynthesized, leading to red leafed phenotype in transgenic plants (Additional file 5: Fig. S5a, b).
Methods
Plant materials and growth conditions
Wild type Shanxin Yang (WT) and transgenic Shanxin Yang (P. davidiana × P. bolleana) overexpressing PdMYB118 used in our previous report were propagated on MS medium, transferred to soil, and grown in greenhouse [34, 44]. Shanxin Yang was provided by Prof. Gui-Feng Liu (Northeast Forestry University, China). Populus deltoids and its red leaf mutant were purchased from Qingyuan HiTech Ltd. (Yantai, China). Plants were grown in greenhouse at 25 °C (day)/18 °C (night) in a 12 h light/12 h dark photoperiod.
Wound treatments
For wound induction, the leaves of wild type and transgenic poplar grown on MS medium or in green house were cut into 1.5 cm dices and transferred onto MS medium. At various time points after wound induction, leaf discs were harvested, frozen in liquid nitrogen, and stored at − 80 °C for RNA, protein, JA and anthocyanin extraction.
MeJA treatment
To analyze the induction of PdMYB118 and ABGs by JA, 100 μM MeJA (Sigma, Shanghai, China) solution was sprayed onto the mature leaves of WT poplar plants grown in greenhouse. After 3 h, leaves were collected and used for RNA extraction. Leaves sprayed with water were used as a negative control.
Quantitative real-time RT-PCR
Total RNA was extracted from leaves and leaf protoplasts with RNAiso Reagent (Takara, Shanghai, China) and qPCR was performed as described previously [45]. The relative expression of each target gene was normalized using PtrEF1β. Gene specific primers used in this study were listed in additional file 8: Table S1. Three independent replicates of measurements were performed for each sample.
Anatomical observations
For histological observation of anthocyanin speckles, leaf dices of wild type and transgenic plants were examined with a light microscope after wound treatment for 48 h. Images were captured under the SMZ800 microscope.
Anthocyanin content determination
After wound induction for 48 h, anthocyanin content in the leaf discs of WT and transgenic plants grown in greenhouse was measured as described previously [46]. To detect the quantity of anthocyanin, A530 per gram fresh weight (FW) was used. Three replicates were carried out for each measurement, and the variability was indicated with the standard deviation (SD).
Western blotting
Western blotting analyses were performed as described previously [47]. Briefly, a total amount of 30 μg proteins were extracted from the wounded leaf discs at various time points and separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Then total protein was electrotransferred onto polyvinylidene difluoride membranes. Immunoblotting was performed using JAZ1 antibodies against the Arabidopsis JAZ1 protein (Agrisera, http://www.agrisera.com/), then incubated with the secondary antibody goat anti-rabbit IgG-horseradish peroxidase (HRP) (Abmart, China). To detect the Actin, the primary antibody (mouse monoclonal Actin antibody) and the secondary antibody (goat anti-mouse IgG-HRP) were used to perform the immune reaction (Abmart, Shanghai, China). After incubation in the chemiluminescence detection solution LumiGLO (KPL, USA), membranes were imaged with a chemiluminescence image system Tanon 5500 (Tanon, Shanghai, China). Proteins were quantified with a BCA Protein Assay kit (Thermo, Shanghai, China).
JA content assays
For JA and JA-Ile content assays, the wounded leaves of wild type poplar plants at 0 h, 1 h and 24 h time points were ground into fine powder in liquid nitrogen, respectively. JA extraction was performed as described previously [48]. Three replicates were carried out for each assay, and the variability was indicated with the standard deviation (SD).
Transient expression of TFs in poplar mesophyll protoplasts
To analyze the transcription function of PdMYB118, PdTT8 and PdMYB118/PdTT8 complex, CaMV 35S promoter driven transcriptional factor effectors were produced by inserting PdMYB118 or PdTT8 into pGreenII62-SK [49]. Leaf protoplasts were isolated from the leaves of Shanxin Yang as described previously [34]. The resultant constructs pGreenII62-SK-PdMYB118, pGreenII62-SK-PdTT8 and pGreenII62-SK-PdMYB118 + pGreenII62-SK-PdTT8 were transferred into protoplasts, respectively. The empty vector was used as a negative control. After kept in dark for 16 h, the transfected protoplasts were collected for RNA extraction and qRT-PCR analyses of ABGs as described above.
To detect the effects of PdTT8 or PtrJAZ1 on ABGs expression in the leaves of transgenic plants overexpressing PdMYB118, effectors pGreenII62-SK-PdTT8, pGreenII62-SK-PtrJAZ1 and pGreenII62-SK-PdTT8 + pGreenII62-SK-PtrJAZ1 were transformed into the mesophyll protoplasts of transgenic plants, respectively. The transfected protoplasts were used for qRT-PCR analyses of ABGs. Three replicates were carried out for each assay, and the variability was indicated with the standard deviation (SD).
BiFC assays
Full-length coding sequences of PtrJAZ1, PdMYB118 and PdTT8 were individually cloned and subsequently recombined into YFP BiFC vectors so that they were fused with the N- or C-terminal of YFP (nYFP or cYFP) to generate pSAT4-nYFP-PdMYB118/PtrJAZ1 and pSAT4-cYFP-PdTT8/PdMYB118 plasmids. Primers used for gene clone are given in Supporting Information Table S1. To detect the interaction of PtrJAZ1 with PdTT8 or PdMYB118, and the interaction of PdMYB118 with PdTT8, the relative constructs were co-transfected into the mesophyll protoplasts of wild type plants. After incubated at 23 °C for 16 h, the protoplasts were analyzed using a confocal microscope at 514 nm wavelength (Zeiss LSM 510 META). Three replicates were carried out for each assay, and the variability was indicated with the standard deviation (SD).
Transient transcription dual-luciferase assays
For dual-luciferase assays, the LUC reporter constructs were generated by cloning the promoter of PtrCHS1, PtrDRF2 or PtrANS1 into pGreenII0800-LUC [47]. The resultant pGreenII62-SK-PdMYB118, pGreenII62-SK-PdTT8 or pGreenII62-SK-PtrJAZ1 was used as effector construct as described above. To detect the inhibition of PtrJAZ1 to the transcription activity of the PdMYB118/PdTT8 complex, effectors PdMYB118, PdTT8, PdMYB118 + PdTT8 or PdMYB118 + PdTT8 + PtrJAZ1 were repetitively co-expressed with each reporter construct in poplar leaf protoplasts [45]. The LUC/REN ratio was used to represent the relative activity of the transcriptional factors. Three replicates were carried out for each assay, and the variability was indicated with the standard deviation (SD).
Statistical analysis
All data were obtained from three biological replicates each. For statistical analyses, Student’s t-test (two-tailed) or ANOVA (one-way) was used to generate every P value. The variability was indicated with the standard deviation (SD). *, ** and *** indicate p-values < 0.05, <0.01 and < 0.001, respectively.
Supplementary information
Additional file 1: Figure S1. Wound induced anthocyanin biosynthesis in the leaf of transgenic plants.
Additional file 2: Figure S2. BiFC assays to test the interaction of PtrJAZ1 with PdMYB118.
Additional file 3: Figure S3. Expression of PtrJAZ1 and PdTT8 in the protoplasts isolated from the leaves of transgenic plants overexpressing PdMYB118.
Additional file 4: Figure S4. qRT-PCR analyses of PdMYB118 and PdTT8 in the wounded leaves of WT and transgenic plants overexpressing PdMYB118.
Additional file 5: Figure S5. A proposed model of wound induced anthocyanin biosynthesis in poplar.
Additional file 6: Figure S6. Western blotting analyses of JAZ1 protein in the wounded leaves.
Additional file 7: Figure S7. Western blotting analyses of JAZ1 protein in the wounded leaves of transgenic poplar plants.
Additional file 8: Table S1. Primer sequences used in this study.
Acknowledgements
We thank Prof. Gui-Feng Liu (Northeast Forestry University, China) for kindly providing us the Shanxin yang (P. davidiana × P. bolleana) clone.
Abbreviations
- TF
Transcription factors
- ABG
Anthocyanin biosynthesis gene
- MeJA
Methyl jasmonate
- JAZ
JASMONATE ZIM-domain
- CHS
Chalcone synthase
- CHI
Chalcone isomerase
- F3H
Flavanone 3-hydroxylase
- F3’H
Flavanone 3′-hydroxylase
- F3′5’H
Flavonoid 3’5’-hydroxylase
- DFR
Dihydroflavonol 4-reductase
- ANS
Anthocyanidin synthase
- bHLH
Basic helix-loop-helix
- WT
Wild type
Authors’ contributions
HW, XW, CY, CW and YL performed the experiments and analyzed data. HW and HZ wrote the manuscript and arranged all data. All authors contributed revision of the manuscript. All authors read and approved the manuscript.
Funding
This work was supported by the following grants: the National Key R & D Program of China (2016YFD0600106, 2019YFD1000500); the National Key Program on Transgenic Research (2018ZX08020002–003-004); the National Natural Science Foundation of China (31700524, 31870576, 31871625); the Modern Agricultural Industry Technology System Innovation Team of Shandong Province of China (SDAIT-02-05); the Agricultural Variety Improvement Project of Shandong Province (2019LZGC009). The funders played no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript .
Availability of data and materials
All materials and data analyzed in this study are available from the corresponding author
Ethics approval and consent to participate
This study does not contain any research requiring ethical consent or approval.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Haihai Wang and Xiaoqing Wang contributed equally to this work.
Supplementary information
Supplementary information accompanies this paper at 10.1186/s12870-020-02389-1.
References
- 1.Winkel-Shirley B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001;126:485–493. doi: 10.1104/pp.126.2.485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Christie PJ, Alfenito MR, Walbot V. Impact of low-temperature stress on general phenylpropanoid and anthocyanin pathways: enhancement of transcript abundance and anthocyanin pigmentation in maize seedlings. Planta. 1994;194:541–549. doi: 10.1007/BF00714468. [DOI] [Google Scholar]
- 3.Sarma AD, Sharma R. Anthocyanin-dna copigmentation complex: mutual protection against oxidative damage. Phytochemistry. 1999;52:1313–1318. doi: 10.1016/S0031-9422(99)00427-6. [DOI] [Google Scholar]
- 4.Steyn W, Wand S, Holcroft D, Jacobs G. Anthocyanins in vegetative tissues: a proposed unified function in photoprotection. New Physiol. 2002;155:349–361. doi: 10.1046/j.1469-8137.2002.00482.x. [DOI] [PubMed] [Google Scholar]
- 5.Lorenc-Kukuła K, Jafra S, Oszmiański J, Szopa J. Ectopic expression of anthocyanin 5-o-glucosyltransferase in potato tuber causes increased resistance to bacteria. J Agr Food Chem. 2005;53:272–281. doi: 10.1021/jf048449p. [DOI] [PubMed] [Google Scholar]
- 6.Nakabayashi R, Yonekura-Sakakibara K, Urano K, Suzuki M, Yamada Y, Nishizawa T, Matsuda F, Kojima M, Sakakibara H, Shinozaki K, Michael AJ, Tohge T, Yamazaki M, Saito K. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014;77:367–379. doi: 10.1111/tpj.12388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tsai CJ, Harding SA, Tschaplinski TJ, Lindroth RL, Yuan YN. Genome-wide analysis of the structural genes regulating defense phenylpropanoid metabolism in Populus. New Physiol. 2006;172:47–62. doi: 10.1111/j.1469-8137.2006.01798.x. [DOI] [PubMed] [Google Scholar]
- 8.Mellway RD, Tran LT, Prouse MB, Campbell MM, Constabel CP. The wound-, pathogen-, and ultraviolet B-responsive MYB134 gene encodes an R2R3 MYB transcription factor that regulates proanthocyanidin synthesis in poplar. Plant Physiol. 2009;150:924–941. doi: 10.1104/pp.109.139071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.James AM, Ma D, Mellway R, Gesell A, Yoshida K, Walker V, Tran L, Stewart D, Reichelt M, Suvanto J, Salminen JP, Gershenzon J, Séguin A, Constabel CP. Poplar MYB115 and MYB134 transcription actors regulate proanthocyanidin synthesis and structure. Plant Physiol. 2017;174:154–171. doi: 10.1104/pp.16.01962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wang L, Ran L, Hou Y, Tian Q, Li C, Liu R, Fan D, Luo K. The transcription factor MYB115 contributes to the regulation of proanthocyanidin biosynthesis and enhances fungal resistance in poplar. New Phytologist. 2017;215:351–67. [DOI] [PubMed]
- 11.Ullah C, Tsai CJ, Unsicker SB, Xue LJ, Reichelt M, Gershenzon J. Hammerbacher A. Salicylic acid activates poplar defense against the biotrophic rust fungus Melampsora larici-populina via increased biosynthesis of catechin and proanthocyanidins. New Physiol 2019a;221:960–975. [DOI] [PMC free article] [PubMed]
- 12.Ullah C, Unsicker SB, Reichelt M, Gershenzon J. Hammerbacher A. Accumulation of catechin and proanthocyanidins in black poplar stems after infection by Plectosphaerella populi: hormonal regulation, biosynthesis and antifungal activity. Front Plant Sci 2019b;10:1–10. [DOI] [PMC free article] [PubMed]
- 13.Espley RV, Hellens RP, Putterill J, Stevenson DE, Kutty-Amma S, Allan AC. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007;49:414–427. doi: 10.1111/j.1365-313X.2006.02964.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hancock KR, Collette V, Fraser K, Greig M, Xue H, Richardson K, Jones C, Rasmussen S. Expression of the R2R3-MYB transcription factor TaMYB14 from Trifolium arvense activates proanthocyanidin biosynthesis in the legumes Trifolium repens and Medicago sativa. Plant Physiol. 2012;159:1204–1220. doi: 10.1104/pp.112.195420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hichri I, Barrieu F, Bogs J, Kappel C, Delrot S, Lauvergeat V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J Exp Bot. 2011;62:2465–2483. doi: 10.1093/jxb/erq442. [DOI] [PubMed] [Google Scholar]
- 16.Petroni K, Tonelli C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 2001;181:219–229. doi: 10.1016/j.plantsci.2011.05.009. [DOI] [PubMed] [Google Scholar]
- 17.Xu W, Dubos C, Lepiniec L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends in Plant Sci. 2015;20:176–185. doi: 10.1016/j.tplants.2014.12.001. [DOI] [PubMed] [Google Scholar]
- 18.Grotewold E, Sainz MB, Tagliani L, Hernandez JM, Bowen B, Chandler VL. Identification of the residues in the Myb domain of maize C1 that specify the interaction with the bHLH cofactor R. P Natl Acad Sci USA. 2000;97:13579–13584. doi: 10.1073/pnas.250379897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Elomaa P, Uimari A, Mehto M, Albert VA, Laitinen RA, Teeri TH. Activation of anthocyanin biosynthesis in Gerbera hybrida (Asteraceae) suggests conserved protein–protein and protein–promoter interactions between the anciently diverged monocots and eudicots. Plant Physiol. 2003;133:1831–1842. doi: 10.1104/pp.103.026039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sweeney MT, Thomson MJ, Pfeil BE, McCouch S. Caught red-handed: Rc encodes a basic helix–loop–helix protein conditioning red pericarp in rice. Plant Cell. 2006;18:283–294. doi: 10.1105/tpc.105.038430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Nakatsuka T, Haruta KS, Pitaksutheepong C, Abe Y, Kakizaki Y, Yamamoto K, Shimada N, Yamamura S, Nishihara M. Identification and characterization of R2R3-MYB and bHLH transcription factors regulating anthocyanin biosynthesis in gentian flowers. Plant Cell Physiol. 2008;49:1818–1829. doi: 10.1093/pcp/pcn163. [DOI] [PubMed] [Google Scholar]
- 22.Matus JT, Poupin MJ, Canon P, Bordeu E, Alcalde JA, Arce-Johnson P. Isolation of WDR and bHLH genes related to flavonoid synthesis in grapevine (Vitis vinifera L.) Plant Mol Biol. 2010;72:607–620. doi: 10.1007/s11103-010-9597-4. [DOI] [PubMed] [Google Scholar]
- 23.Chung HS, Koo AJK, Gao X, Jayanty S, Thines B, Jones AD, Howe GA. Regulation and function of Arabidopsis JASMONATE ZIM-domain genes in response to wounding and herbivory. Plant Physiol. 2008;146:952–964. doi: 10.1104/pp.107.115691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Koo AJ, Gao X, Jones AD, Howe GA. A rapid wound signal activates the systemic synthesis of bioactive jasmonates in Arabidopsis. Plant J. 2009;59:974–986. doi: 10.1111/j.1365-313X.2009.03924.x. [DOI] [PubMed] [Google Scholar]
- 25.Staswick PE. JAZing up jasmonate signaling. Trends in Plant Sci. 2008;13:66–71. doi: 10.1016/j.tplants.2007.11.011. [DOI] [PubMed] [Google Scholar]
- 26.Gfeller A, Dubugnon L, Liechti R, Farmer EE. Jasmonate biochemical pathway. Sci Signal. 2010;3:332. doi: 10.1126/scisignal.3109cm3. [DOI] [PubMed] [Google Scholar]
- 27.Thines B, Katsir L, Melotto M, Niu Y, Mandaokar A, Liu G, Nomura K, He SY, Howe GA, Browse J. JAZ repressor proteins are targets of the SCF (COI1) complex during jasmonate signalling. Nature. 2007;448:661–665. doi: 10.1038/nature05960. [DOI] [PubMed] [Google Scholar]
- 28.Gan Y, Li H, Xie Y, Wu W, Li M, Wang X, Huang J. THF1 mutations lead to increased basal and wound-induced levels of oxylipins that stimulate anthocyanin biosynthesis via COI1 signaling in Arabidopsis. J Integr Plant Biol. 2014;56:916–927. doi: 10.1111/jipb.12177. [DOI] [PubMed] [Google Scholar]
- 29.Qi T, Song S, Ren Q, Wu D, Huang H, Chen Y, Fan M, Peng W, Ren C, Xie D. The Jasmonate-ZIM-domain proteins interact with the WD-repeat/bHLH/MYB complexes to regulate jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell. 2011;23:1795–1814. doi: 10.1105/tpc.111.083261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Xie C, Wang D, Kou F, Kang D, Yang X. The pathogen and wound induces expression of genes related to proanthocyanidins (PAs) synthesis in cotton leaves. Am J Plant Sci. 2012;3:228–234. doi: 10.4236/ajps.2012.32027. [DOI] [Google Scholar]
- 31.An J, Zhang X, You C, Bi S, Wang X, Hao Y. MdWRKY40 promotes wounding-induced anthocyanin biosynthesis in association with MdMYB1 and undergoes MdBT2-mediated degradation. New Phytol. 2019;224:380–395. doi: 10.1111/nph.16008. [DOI] [PubMed] [Google Scholar]
- 32.Wilkins O, Nahal H, Foong J, Provart NJ, Campbell MM. Expansion and diversification of the Populus R2R3-MYB family of transcription factors. Plant Physiol. 2009;149:981–993. doi: 10.1104/pp.108.132795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cho JS, Nguyen VP, Jeon HW, Kim MH, Eom SH, Lim YJ, Kim WC, Park EJ, Choi YI, Ko JH. Overexpression of PtrMYB119, a R2R3-MYB transcription factor from Populus trichocarpa, promotes anthocyanin production in hybrid poplar. Tree Physiol. 2016;36:1162–1176. doi: 10.1093/treephys/tpw046. [DOI] [PubMed] [Google Scholar]
- 34.Wang HH, Wang XQ, Song WM, Bao Y, Jin YL, Jiang CM, Wang CT, Li B, Zhang HX. PdMYB118, isolated from a red leaf mutant of Populus deltoids, is a new transcription factor regulating anthocyanin biosynthesis in poplar. Plant Cell Rep. 2019;38:927–936. doi: 10.1007/s00299-019-02413-1. [DOI] [PubMed] [Google Scholar]
- 35.Sheard LB, Tan X, Mao H, Withers J, Ben-Nissan G, TR RH, Kobayashi Y, Hsu FF, Sharon M, Browse J, He SY, Rizo J, Howe GA, Zheng N. Jasmonate perception by inositol-phosphate-potentiated COI1–JAZ co-receptor. Nature. 2010;468:400–405. doi: 10.1038/nature09430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.An XH, Tian Y, Chen KQ, Liu XJ, Liu DD, Xie XB, Cheng CG, Cong PH, Hao YJ. MdMYB9 and MdMYB11 are involved in the regulation of the JA-induced biosynthesis of anthocyanin and proanthocyanidin in apples. Plant Cell Physiol. 2015;56:650–662. doi: 10.1093/pcp/pcu205. [DOI] [PubMed] [Google Scholar]
- 37.Tamari G, Borochov A, Atzorn R, Weiss D. Methyl jasmonate induces pigmentation and flavonoid gene expression in petunia corollas: A possible role in wound response. Physiol Plantarum. 1995;94:45–50. doi: 10.1111/j.1399-3054.1995.tb00782.x. [DOI] [Google Scholar]
- 38.Nesi N, Jond C, Debeaujon I, Caboche M, Lepiniec L. The Arabidopsis TT2 gene encodes an R2R3 MYB domain protein that acts as a key determinant for proanthocyanidin accumulation in developing seed. Plant Cell. 2001;13:2099–2114. doi: 10.1105/TPC.010098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Quattrocchio F, Verweij W, Kroon A, Spelt C, Mol J, Koes R. PH4 of petunia is an R2R3 MYB protein that activates vacuolar acidification through interactions with basic-helix-loop-helix transcription factors of the anthocyanin pathway. Plant Cell. 2006;18:1274–1291. doi: 10.1105/tpc.105.034041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hichri I, Heppel SC, Pillet J, Léon C, Czemmel S, Delrot S, Lauvergeat V, Bogs J. The basic helix-loop-helix transcription factor MYC1 is involved in the regulation of the flavonoid biosynthesis pathway in grapevine. Mol Plant. 2010;3:509–523. doi: 10.1093/mp/ssp118. [DOI] [PubMed] [Google Scholar]
- 41.Wasternack C, Song S. Jasmonates: biosynthesis, metabolism, and signaling by proteins activating and repressing transcription. J Exp Bot. 2017;68:1303–1321. doi: 10.1093/jxb/erw443. [DOI] [PubMed] [Google Scholar]
- 42.Loreti E, Povero G, Novi G, Solfanelli C, Alpi A, Perata P. Gibberellins, jasmonate and abscisic acid modulate the sucrose-induced expression of anthocyanin biosynthetic genes in Arabidopsis. New Phytol. 2008;179:1004–1016. doi: 10.1111/j.1469-8137.2008.02511.x. [DOI] [PubMed] [Google Scholar]
- 43.Shan X, Zhang Y, Peng W, Wang Z, Xie D. Molecular mechanism for jasmonate-induction of anthocyanin accumulation in Arabidopsis. J Exp Bot. 2009;60:3849–3860. doi: 10.1093/jxb/erp223. [DOI] [PubMed] [Google Scholar]
- 44.Wang HH, Wang CT, Liu H, Tang RJ, Zhang HX. An efficient agrobacterium-mediated transformation and regeneration system for leaf explants of two elite aspen hybrid clones Populus alba × P. berolinensis and Populus davidiana × P. bolleana. Plant Cell Rep. 2011;30:2037–2044. doi: 10.1007/s00299-011-1111-1. [DOI] [PubMed] [Google Scholar]
- 45.Wang HH, Tang RJ, Liu H, Chen HY, Liu JY, Jiang XN, Zhang HX. Chimeric repressor of PtSND2 severely affects wood formation in transgenic Populus. Tree Physiol. 2013;33:878–886. doi: 10.1093/treephys/tpt058. [DOI] [PubMed] [Google Scholar]
- 46.Yoshida K, Ma D, Constabel CP. The MYB182 protein down-regulates proanthocyanidin and anthocyanin biosynthesis in poplar by repressing both structural and regulatory flavonoid genes. Plant Physiol. 2015;167:693–710. doi: 10.1104/pp.114.253674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang HH, Jiang CM, Wang CT, Yang Y, Gao XY, Zhang HX. Antisense expression of the fasciclin-like arabinogalactan protein PtFLA6 gene in Populus inhibits expression of its homologous genes and alters stem biomechanics and cell wall composition in transgenic trees. J Exp Bot. 2015;66:1291–1302. doi: 10.1093/jxb/eru479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chen ML, Fu XM, Liu JQ, Ye TT, Hou SY, Huang YQ, Yuan BF, Wu Y, Feng YQ. Highly sensitive and quantitative profiling of acidic phytohormones using derivatization approach coupled with nano-LC-ESI-Q-TOF-MS analysis. J Chromatogr B. 2012;905:67–74. doi: 10.1016/j.jchromb.2012.08.005. [DOI] [PubMed] [Google Scholar]
- 49.Hellens RP, Allan AC, Friel EN, Bolitho K, Grafton K, Templeton MD, Karunairetnam S, Gleave AP, Laing WA. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods. 2005;1:13. doi: 10.1186/1746-4811-1-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Figure S1. Wound induced anthocyanin biosynthesis in the leaf of transgenic plants.
Additional file 2: Figure S2. BiFC assays to test the interaction of PtrJAZ1 with PdMYB118.
Additional file 3: Figure S3. Expression of PtrJAZ1 and PdTT8 in the protoplasts isolated from the leaves of transgenic plants overexpressing PdMYB118.
Additional file 4: Figure S4. qRT-PCR analyses of PdMYB118 and PdTT8 in the wounded leaves of WT and transgenic plants overexpressing PdMYB118.
Additional file 5: Figure S5. A proposed model of wound induced anthocyanin biosynthesis in poplar.
Additional file 6: Figure S6. Western blotting analyses of JAZ1 protein in the wounded leaves.
Additional file 7: Figure S7. Western blotting analyses of JAZ1 protein in the wounded leaves of transgenic poplar plants.
Additional file 8: Table S1. Primer sequences used in this study.
Data Availability Statement
All materials and data analyzed in this study are available from the corresponding author