

Abstract
Purpose
Electronic cigarettes (e-cigs) are relatively new devices that allow the user to inhale a heated and aerosolized solution. At present, little is known about their health effects in the human lung, particularly in the small airways (<2 mm in diameter), a key site of airway obstruction and destruction in chronic obstructive pulmonary disease and other acute and chronic lung conditions. The aim of this study was to investigate the effect of e-cigarettes on human distal airway inflammation and remodeling.
Methods
We isolated primary small airway epithelial cells from donor lungs without known lung disease. Small airway epithelial cells were cultured at air–liquid interface and exposed to 15 puffs vapor obtained by heating a commercially available e-cigarette solution (e-vapor) with or without nicotine. After 24 hrs of e-vapor exposure, basolateral and apical media as well as cell lysates were collected to measure the pleiotropic cytokine interleukin 6 (IL6) and MUC5AC, one of the major components in mucus.
Results
Unlike the nicotine-containing e-vapor, nicotine-free e-vapor significantly increased the amount of IL6, which was coupled with increased levels of intracellular MUC5AC protein. Importantly, a neutralizing IL6 antibody (vs an IgG isotype control) significantly inhibited the production of MUC5AC induced by nicotine-free e-vapor.
Conclusion
Our results suggest that human small airway epithelial cells exposed to nicotine-free e-vapor increase the inflammatory response and mucin production, which may contribute to distal lung airflow limitation and airway obstruction.
Keywords: electronic cigarette, small airway epithelial cells, inflammation, interleukin 6, mucus
Plain Language Summary
The use of electronic cigarettes (e-cigs) is rapidly rising, but its health effect in human lungs particularly the distal lung remains unclear. In this study, we sought to determine the role of e-cigs in human distal airway inflammation and mucus production, two key components in the pathogenesis of chronic obstructive pulmonary disease. We collected human distal airway epithelial cells from donors without lung disease, and performed cell cultures to create a model mimicking in vivo small airway epithelium. The cultured cells were then exposed to e-cig vapor with or without nicotine to measure its damaging effects on cell inflammation, injury and mucus production. Our results demonstrated that nicotine-free e-cigs increased human distal airway epithelial production of pro-inflammatory cytokine IL6 and mucus. Our findings suggest that even a single exposure to e-cigs is detrimental to human lung health as excessive inflammation and mucus production could cause airway obstruction, a serious health condition that needs immediate medical care.
Introduction
Electronic cigarettes (e-cigs) are handheld battery-operated devices that recreate the feeling of traditional tobacco smoking without actual combustion occurring.1,2 They heat and aerosolize a solution of propylene glycol, vegetable glycerin, nicotine, flavorings and other components, which is then inhaled by the user (“vaping”). E-cigs are increasingly popular as an alternative to conventional smoking products due to a perception that they are safer than combustible products and may be useful in smoking cessation. While there are numerous studies demonstrating the harmful effects of tobacco smoking, given the relatively recent use of e-cigs, much less is known about their effects on human lungs.
Ex vivo studies into the human health effects of e-cigs have primarily involved the use of epithelial cells isolated from large airways, as they are more accessible for collection via bronchoscopy or nasal brushings. Small airways, typically defined as <2 mm diameter, are known to be a major site of airflow obstruction in diseases such as asthma and chronic obstructive pulmonary disease (COPD),3–5 yet they are understudied due to their poor accessibility. In epithelial cells isolated from large airways such as bronchi or trachea, both conventional cigarettes and e-cigs have been shown to alter the inflammatory response; in particular, it is widely accepted that conventional cigarettes promote inflammation and impair host defenses.6–8 Previous studies have used a wide variety of e-cig products and exposure methods, including diluted e-juice,9 e-vapor extract,10 and direct e-vapor exposure,11 and a plethora of flavorings and nicotine strengths, meaning care must be taken when comparing studies and drawing conclusions. Nevertheless, several studies appear to support the association between e-cigs and inflammation, including pro-inflammatory cytokines IL6 and IL8 in large airway epithelial cells.9–11 Moreover, e-cig exposure has been shown to increase airway mucin production in murine studies and clinical investigations.12,13 Increased mucus production, particularly mucin MUC5AC, and goblet cell hyperplasia are major contributing factors in the obstruction of small airways in COPD.14–17 Most investigations of the distal lung involve the use of the A549 human adenocarcinoma alveolar cell line, or inferred effects from bronchoalveolar lavage fluid, sputum, or brushings from 10–12th airway generation.13,18,19 At present, the health effect of e-cigs in human distal lung remains unknown. Nonetheless, limited numbers of studies20,21 suggest that the measured e-cig aerosol particle size range is alarming as some particles are small enough to be able to reach the alveolar compartment of the lung. To our knowledge, no report has studied the direct effect of e-cigarettes on cultured primary human small airway epithelial cells (SAEC). Our goal was to investigate the effects of e-cigarettes on primary SAEC cultured at the air-liquid interface, including inflammatory effects (eg, IL6) and tissue remodeling (eg, mucin production).
Materials and Methods
Small Airway Epithelial Cell Isolation and Expansion by Culture
Human lungs from de-identified organ donors whose lungs were not suitable for transplantation were donated for medical research through the National Disease Research Interchange (Philadelphia, PA), the International Institute for the Advancement of Medicine (Edison, NJ) or the Donor Alliance of Colorado. The Institutional Review Board (IRB) at National Jewish Health, Denver, Colorado deemed this research as nonhuman subject research. We selected donors with no history of clinical lung disease, absence of lung injury indicated by a PaO2/FiO2 ratio of >225, a chest radiograph that indicated no infection, and a time on the ventilator of <5 days. The sex, age, and race were variable and were not selection criteria; basic demographic information is detailed in Table 1. Small airway epithelial cells from Subject # 1, a smoker, was included in our study in order to increase the sample size as well as to demonstrate if cells from smokers and non-smokers may respond similarly to nicotine-free e-vapor.
Table 1.
Demographic Information of Lung Donors
| Experiments for e-Vapor Effect on IL6 and Mucin | IL6 Neutralization Experiments | ||||||
|---|---|---|---|---|---|---|---|
| Subject ID | Sex (M/F) |
Age (Years) | Smoker (Yes/No) |
Subject ID | Sex (M/F) |
Age (Years) | Smoker (Yes/No) |
| 1 | M | 27 | Yes | 6 | M | 35 | No |
| 7 | F | 35 | No | 12 | M | 48 | No |
| 9 | F | 77 | No | 13 | F | 18 | No |
| 10 | M | 39 | No | 21 | M | 87 | No |
| 12 | M | 48 | No | 23 | F | 61 | No |
| 20 | M | 46 | No | 24 | M | 38 | No |
| Mean±SEM | 45.3±7.1 | 47.8±9.8 | |||||
Abbreviation: IL6, interleukin 6.
Small airway (<2 mm diameter) epithelial cells (SAEC) were collected from cut pieces of the distal lung using a 2 mm bronchoscopy brush (Conmed Greenwood Village, CO). The flexible bristles of the brush allowed penetration of airways <2 mm (measured with a ruler) which were penetrated no deeper than the length of the brush, after which the brush was placed in sterile PBS. The cells were isolated by centrifugation, resuspended in PBS, counted, and plated onto an irradiated NIH 3T3 fibroblast feeder layer in F-media. Once large visible colonies had formed (7–10 days), they were removed with 0.25% trypsin (Corning 25-053-CI) and plated onto double collagen-coated 12-well transwells (Advanced BioMatrix 5005 and Corning Costar 3460) for air–liquid interface (ALI) culture. Cells were maintained in F6 medium, a 50/50 mixture of DMEM and Airway Epithelial Growth Medium (PromoCell) supplemented with 0.5 µg/mL bovine serum albumin (BSA), 50 mM ethanolamine, 30 mM MgCl2, 4.0 mM MgSO4, 1.0 mM CaCl2, 30 ng/mL retinoic acid, 10 ng/mL human epidermal growth factor, 5 µg/mL insulin, 10 µg/mL transferrin, 0.5 µg/mL hydrocortisone, 0.5 µg/mL epinephrine, 0.004 mL/mL bovine pituitary extract, 10 µg/mL gentamicin and 0.25 µg/mL amphotericin B. Cells on the transwells were under the submerged culture for ~7 days, and then shifted to ALI for 14 days when cells clearly demonstrated mucociliary differentiation (Figure 1). Moreover, we performed RNA sequencing on these cells, and found abundant mRNA expression of SCGB1A1, a specific marker for club cells found in the small airways, and surfactant proteins (eg, SP-A). Furthermore, we were able to detect secreted surfactant protein A in apical surface of cultured small airway epithelial cells. Our data are in line with a recent publication that characterized cultured human small airway epithelial cells.22 Specifically, when epithelial cells in the transwells were expanded for ~7 days under submerged condition, 250 µL medium was added to the apical surface (the upper chamber). To switch from submerged to ALI culture, medium in the upper chamber was reduced to 50 µL to keep the cells hydrated and at the same time promote cell mucociliary differentiation as we previously reported.23
Figure 1.

Small airway epithelial cells from a non-smoker donor demonstrated mucociliary differentiation after 14 days of air-liquid interface culture. Green and black arrows indicate cilia and mucous goblet cells, respectively. Hematoxylin and eosin (H&E) staining, x400 magnification.
E-Cigarette Vapor (e-Vapor) Exposure of Cells
White Cloud Electronic Cigarettes (www.whitecloudelectroniccigarettes.com) regular (tobacco) flavor at 0% and 2.4% nicotine strengths were used for this study. According to the manufacturer, the e-liquids consist of 84.6 – 92% propylene glycol and glycerol in an approximately 80:20 ratio, and 8 – 10% proprietary flavorings. E-cigarette vapor (e-vapor) was generated by a Cirrus 2 battery (White Cloud Electronic Cigarettes, Tarpon Springs, FL, no specifications on the atomizer were provided to us) attached via tubing to a MasterFlex L/S Economy Variable Speed Drive.24,25 After at least 14 days of ALI culture, apical surfaces of SAEC were exposed directly to 15 puffs of e-vapor, each puff defined as 35 mL over 4 s every minute (modified from ISO standard 3308–2000 for conventional cigarettes) in a British American Tobacco (London, UK) smoke exposure chamber containing initially warm (37°C) DMEM circulated by a Fisher Scientific variable flow mini pump. A monoclonal anti-IL6 antibody (40 ng/mL, clone MQ2-13A5, ThermoFisher, Waltham, MA) or IgG isotype control (Santa Cruz sc-2026) was added to both apical and basolateral chambers of the transwell plate at least 30 min prior to air or e-vapor exposure, and for the entire 24 hrs post-exposure. Specifically, the IL6 antibody or IgG control in the 100 µL of media was added to the apical surface of small airway epithelial cells in the transwells. This volume is enough to cover the surface and maintain mucociliary differentiation of the cells over the 24 hr period of the experiment. In a pilot study, we did a time course study by evaluating the effects of e-vapor on small airway epithelial IL6 production at both 24 and 48 hrs. Our pilot study demonstrated that IL6 levels were not significantly altered by nicotine-free e-vapor at 48 hr. Thus, we focused on the 24 hr time point.
Nicotine and Cotinine Measurements in the Media of e-Vapor-Exposed SAEC
Nicotine and cotinine concentrations in the apical media of e-vapor-exposed SAEC were detected and measured by GC/MS based on a previously described method.26 The instruments used were a Focus GC coupled to a DSQ II mass spectrometer and an AS 3000 autosampler. A 15 m RTX-5MS column (0.25 mm i.d., 0.25 µm film thickness; Restek, Bellefonte, PA) was used with ultra-high purity helium as the carrier gas at a constant flow rate of 1.0mL/min. Each sample (1 µL) was injected into the 250ºC inlet using the splitless mode with a split flow of 10mL/min and a splitless time of 1.2 min. The initial oven temperature was 62ºC then ramped to 190ºC at 20ºC/min, followed by an increase in temperature to 290ºC at 50ºC/min which was held for 1 min. The MS transfer line was kept at 290ºC and the quadrupole at 200ºC. Ions were detected by electron impact SIM mode for nicotine at m/z 84 and 162, d3-nicotine at m/z 87 and 165, cotinine at m/z 98 and 176; and d3-cotinine at m/z 101 and 179. Under these conditions, nicotine-d3 exhibited a retention time of 4.16 min, followed by nicotine at 4.17 min, cotinine-d3 at 6.18 min, and cotinine at 6.19 min. Nicotine and nicotine-d3 standards were purchased from Cayman Chemical (16,535 and 18,256, Ann Arbor, MI) while cotinine and cotinine-d3 standards were purchased from Cerilliant (C-016 and C-017, Round Rock, TX). While cotinine was not detectable, nicotine at the apical surface of cultured SAEC was detected in the 0.5–3 µg/mL range 24 hrs after exposure to e-vapor (Figure 2). This concentration is relevant to levels of exposure of humans in vivo, where serum nicotine concentrations (typically up to 16.3 ng/mL27) were determined to be ~1000 times lower than those in the airway epithelial lining fluid.28
Figure 2.
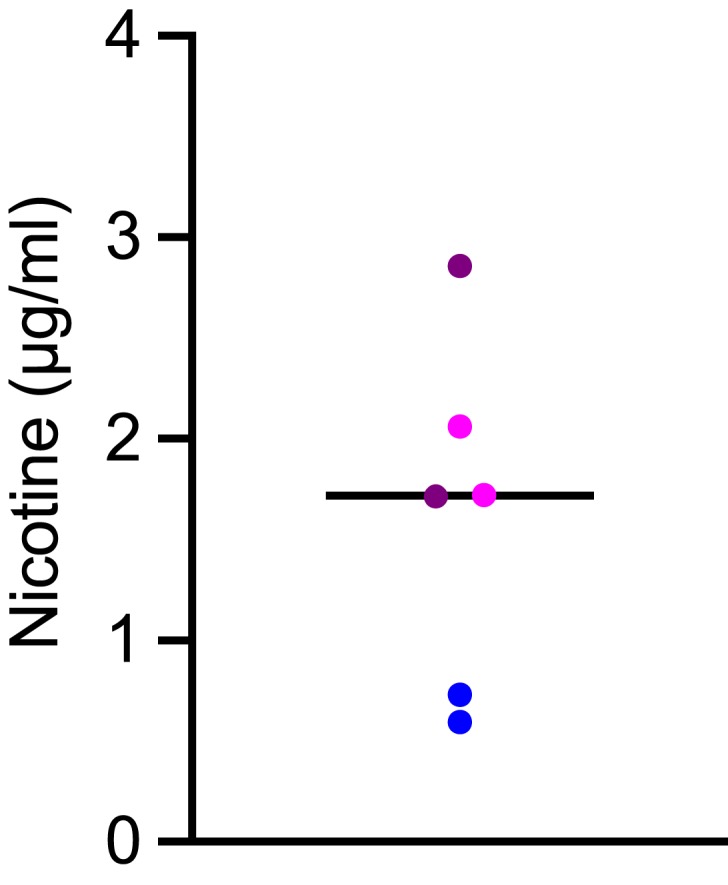
Nicotine concentrations in apical media of human small airway epithelial cells (SAEC, n = 3 donors; experimental replicates from the same donor shown in the same color) exposed to nicotine-containing e-vapor (tobacco flavor) for 24 hrs. The horizontal bar shows the mean value.
MUC5AC Dot Blot
Mucins including MUC5AC are highly glycosylated proteins with molecular weight >250 kD, which is very difficult for Western blot analysis. Given the high specificity of the MUC5AC antibody used in the current study and previous ones,23,29-32 dot blot serves as a specific and reliable method to measure mucin protein levels in supernatants and cell lysates. MUC5AC protein detected by dot blot as we previously described.23 Apical supernatant was first treated with Sputolysin (Calbiochem, San Diego, CA) according to the manufacturer’s directions in order to reduce disulfide bonds. Equal amounts of protein from heat-denatured cell lysates (500 ng in PBS), or equal volumes of apical supernatant (100 µL of 1/25 dilution in PBS), were loaded onto a 0.45 µm pore nitrocellulose membrane sandwiched in the dot blot apparatus (BioRad, Hercules, California). After vacuum filtration of the sample, the wells of the dot blot were washed twice with 200 µL PBS. The membrane was blocked for 1 hr in 5% skim milk powder in tris-buffered saline containing 0.05% tween-20 (TBST), then probed for mucins using a MUC5AC antibody (clone 45M1, NeoMarkers MS-145-P1) in 5% BSA in TBST. The membranes were washed with TBST then probed with a goat-anti-mouse-HRP-conjugated antibody (1858413, ThermoFisher Scientific, Waltham, MA), washed again, and visualized by chemiluminescent reaction of ECL substrate (RPN2236, GE Healthcare, Waukesha, WI). The membrane was stained for total protein with Amido Black (A8181, Sigma, St. Louis, MO). The intensity of the MUC5AC protein signal was measured by densitometry using the NIH ImageJ software. MUC5AC data in cell lysates were expressed as the ratio of MUC5AC/total protein, while the MUC5AC data in the apical supernatants were expressed as arbitrary units. To validate MUC5AC dot blot assay, we performed a dot blot by using five series of twofold dilutions of lysates (total protein levels at 2000, 1000, 500, 250 and 125 ng, in duplicate) of SAEC exposed to nicotine-free e-vapor. A linear relationship was observed between the MUC5AC protein intensity and the total protein levels (r2=0.83, p=0.03), suggesting that the MUC5AC dot blot assay was appropriate for qualifying MUC5AC protein expression levels.
Quantitative PCR (qPCR) for MUC5B mRNA
Since, using our methodology, MUC5B protein levels were undetectable in SAEC, we performed quantitative real-time PCR to measure MUC5B mRNA expression using forward and reverse primers and a FAM-labeled probe as reported in a previous publication.33 Total RNA was extracted using the GenCatch Total RNA Miniprep Kit following manufacturer’s instructions. qPCR was performed using a Taqman probe-based method (ThermoFisher Scientific). GAPDH was used as a reference gene (4352934E, ThermoFisher Scientific).
Lactate Dehydrogenase (LDH) Assay
To quantitate the cytotoxic effects of e-vapor, LDH levels were measured in the basolateral supernatants of SAEC using a cytotoxicity detection kit (Roche Diagnostics, Indianapolis, IN) according to the manufacturer’s instruction. OD value at 450 nm was measured using a microplate reader. Data were expressed as the percentage cytotoxicity of cells with various treatments as compared to medium control alone.9
Measurement of Trans-Epithelial Electrical Resistance (TEER)
TEER was measured using an epithelial volt/ohm (TEER) meter (World Precision Instruments, Sarasota, FL) in ALI cultures of SAEC as previously reported.34 Data were presented as ohms/cm2.
Enzyme-Linked Immunosorbent Assay (ELISA)
Human IL6 levels were measured using a Duoset ELISA kit from R&D systems (Minneapolis, MN) by diluting epithelial supernatants (cell-free) by 2-fold in ELISA buffer according to the manufacturer.
Statistics
All statistics were performed using a JMP program (SAS Institute, Cary, USA). A paired t-test was used for two group comparisons. Analysis of variance (ANOVA) was used for multiple group comparisons. Correlations were performed by using Spearman’s rank correlation coefficients. P≤0.05 was considered statistically significant.
Results
Nicotine-Free e-Cigarettes Increased IL6 Production in SAECs
To investigate if single exposure to e-vapor is sufficient to trigger an inflammatory response, primary SAEC were exposed once to 15 puffs of e-vapor obtained from heating e-cig solutions of the same tobacco flavor and brand that were nicotine-containing (Nic) and nicotine-free (NF). Interestingly, 24 hrs after e-vapor exposure, a consistent and statistically significant increase in IL6 was observed in SAEC exposed to NF e-vapor compared to those exposed to ambient air (Figure 3A, Table 2). However, exposure to Nic e-vapor did not significantly increase IL6 as SAEC from 2 subjects decreased IL6 following Nic e-vapor (Figure 3B, Table 2).
Figure 3.

IL6 levels in basolateral supernatants of air-liquid interface cultured human small airway epithelial cells (SAEC, n=6 donors) measured 24 hrs after exposure to ambient air, nicotine-free (NF) or nicotine-containing (Nic) e-vapor (tobacco flavor). (A) Change in interleukin 6 (IL6) levels in each individual donor following NF e-vapor exposure as compared to air exposure. Each color represents an individual donor. The square shape represents the smoker. (B) Change in IL6 levels in each individual donor following Nic e-vapor exposure as compared to air exposure. As the IL6 data varied among the subjects, the log transformation of the data was used for the paired t-test.
Table 2.
IL6 Levels (pg/ml) in Basolateral Supernatants of Air-Liquid Interface Cultured Human Small Airway Epithelial Cells Exposed to Air, Nicotine-Free (NF) or Nicotine-Containing (Nic) e-Vapor
| Subject # | Air | NF e-Vapor | Nic e-Vapor |
|---|---|---|---|
| 1 | 149 | 314 | 189 |
| 7 | 81 | 98 | 61 |
| 9 | 131 | 139 | 93 |
| 10 | 172 | 360 | 261 |
| 12 | 74 | 117 | 81 |
| 20 | 159 | 173 | 199 |
Abbreviation: IL6, interleukin 6.
To determine the potential mechanisms by which Nic e-vapor did not significantly increase IL6, we measured epithelial integrity as indicated by trans-epithelial electrical resistance (TEER), and cytotoxicity as indicated by the levels of lactate dehydrogenase (LDH). TEER levels were similar among air-exposed cells from different subjects. Moreover, both NF e-vapor and Nic e-vapor did not significantly affect TEER values (Figure 4A). Likewise, LDH levels were not significantly altered by NF e-vapor or Nic e-vapor (Figure 4B).
Figure 4.

(A) Trans-epithelial electrical resistance (TEER) was measured using an epithelial volt/ohm meter in air-liquid interface cultured human small airway epithelial cells (n = 4 subjects) exposed to ambient air, nicotine-free (NF) or nicotine-containing (Nic) e-vapor (tobacco flavor). (B) Lactate dehydrogenase (LDH) levels were quantified in basolateral supernatants of air-liquid interface cultured human small airway epithelial cells (n = 6 subjects) exposed to ambient air, nicotine-free (NF) or nicotine-containing (Nic) e-vapor (tobacco flavor). Each color represents an individual donor. The square shape represents the smoker.
Nicotine-Free e-Cigarettes Increased Intracellular MUC5AC Production
Airway mucins are secreted from airway epithelial goblet cells and submucosal glands and play critical roles in maintaining health, including hydration, trapping of foreign particles, immunomodulation, and regulation of cell proliferation.35 Conventional cigarette smokers and e-cig users have been shown to have increased MUC5AC and MUC5B mucin secretion in large airway epithelial cells and induced sputum samples.13,14 However, smokers with severe airway obstructive disease and impaired mucociliary clearance exhibit decreased MUC5B in large airway epithelial cells.36,37 To determine if mucins were altered by e-vapor in our model, we assessed mucin protein production and secretion by dot blot. MUC5AC was increased in cell lysates from SAEC exposed to NF e-vapor (Figure 5A, Table 3), similar to the observation for IL6. Similarly, NF e-vapor (vs air) increased MUC5AC mRNA expression in SAEC from 5 out of 6 subjects, but this increase was not statistically significant (p>0.05). In contrast, MUC5AC secretion by SAEC in the apical media was decreased by NF e-vapor (Figure 5B, Table 3). There was no change in MUC5B mRNA expression as assessed by RT-qPCR (Figure 5C). MUC5B protein could not be detected. These results suggest that a single exposure NF e-vapor was sufficient to increase the intracellular accumulation of mucin MUC5AC in SAEC, while inhibiting its secretion.
Figure 5.

Human small airway epithelial cells (SAEC) mucin expression after 24 hrs of nicotine-free (NF) e-vapor (tobacco flavor) exposure. (A) MUC5AC protein in lysates of SAEC, detected by dot blot; representative dot blot from one subject shown below. (B) MUC5AC protein in apical supernatants of SAEC, detected by dot blot; representative dot blot from one subject shown below. (C) MUC5B mRNA in SAEC lysates. Each color represents a different subject. The square shape represents the smoker.
Table 3.
MUC5AC Protein Levels in Lysates and Apical Surface (Secreted) of Air-Liquid Interface Cultured Human Small Airway Epithelial Cells Exposed to Air, Nicotine-Free (NF) e-Vapor
| MUC5AC/Total Protein (Cell Lysates) | Secreted MUC5AC (Arbitrary Units) | |||
|---|---|---|---|---|
| Subject no. | Air | NF e-Vapor | Air | NF e-Vapor |
| 1 | 0.31 | 0.46 | 29,801 | 26,268 |
| 7 | 0.24 | 0.46 | 28,840 | 21,259 |
| 9 | 0.13 | 0.60 | 21,757 | 20,179 |
| 10 | 0.32 | 0.46 | 29,150 | 22,290 |
| 12 | 0.32 | 0.45 | 25,100 | 21,464 |
| 20 | 0.02 | 0.62 | 22,954 | 20,128 |
IL6 Neutralization Inhibited Mucin Production
Several cytokines, such as IL6 and IL13, can lead to increased mucin production in large airway epithelial cells.38,39 To investigate the IL6 signaling dependence of mucin production in our model, SAEC were exposed to NF e-vapor in the presence of an anti-IL6 neutralizing antibody or an isotype control (IgG) antibody. As expected, the anti-IL6 antibody effectively neutralized IL6 as it markedly inhibited IL6 detection by ELISA (Figure 6A). Neutralization of IL6 in NF-exposed SAEC consistently and significantly decreased intracellular MUC5AC protein levels in NF e-cig vapor-exposed cells, when compared to the control antibody (Figure 6B). This indicated that increased IL6 contributed to the increased MUC5AC in NF e-vapor-exposed SAEC.
Figure 6.

Effect of an anti-interleukin 6 (IL6) neutralizing antibody compared to immunoglobulin G (IgG) isotype on (A) secreted IL6, as detected by enzyme-linked immunosorbent assay (ELISA), and (B) MUC5AC protein levels, detected by dot blot (representative blot shown below) in human small airway epithelial cells (SAEC) exposed to nicotine-free (NF) e-vapor (tobacco flavor) for 24 hrs. Each color represents a different subject.
Discussion
Cigarette smoke is a major cause of acute and chronic inflammation leading to substantial airway remodeling and tissue damage, particularly in the small airways where the major pathologic features occur.4 In contrast to the widely known detrimental effects of conventional cigarettes, the effect of e-cigs on airway inflammation and remodeling remains unclear. This study demonstrated an increase of IL6 and intracellular mucin by nicotine-free e-vapor, but not nicotine-containing e-vapor. Our small airway epithelial cell culture data have extended the previous studies that demonstrated an increase in IL6 upon exposure to nicotine-free e-cig juice in cultured human tracheobronchial cells and Calu-3 epithelial cells, and alveolar macrophages.9–11,40 Our data showing no increase of IL6 by nicotine-containing e-vapor are consistent with those recently reported by Czekala L et al.41 We realize that the data presented here showed a large variation of IL6 responses to e-vapor in SAEC from different donors although cells from all the six donors demonstrated increased IL6. The smoking status, age and sex did not appear to play a role in such variation, but given the relatively small sample size (n=6), further investigation into the effects of age and sex on the IL-6 response to e-vapor is warranted. Together, studies from us and others suggest that an increase in IL6 following e-vapor does not appear to be dependent on nicotine nor on brand of e-cig used. The dampening effect of nicotine on the production of IL6 observed in our current study may be due to known immune suppressive effects of nicotine,42–44 or to the short duration of exposure in our model.
The clinical implication of IL6 induction by acute exposure to nicotine-free e-vapor deserves further research, especially in terms of long-term effects of repeated exposures in those who chose nicotine-free e-cig products. Conversely, the lack of IL6 production by acute exposures to Nic-containing e-cig vapor may also have clinical implications, in situations when IL6 production is necessary for host defense. Further, once tobacco smoke reaches the small airways, nicotine can be quickly absorbed into the bloodstream and metabolized to many metabolites including cotinine.45 It is, therefore, possible that in vivo, following the inhalation of nicotine-containing e-vapor, nicotine is quickly absorbed and removed from the small airways, leaving the viscous nicotine-free e-cig components including propylene glycol on the small airway mucosa, which may then contribute to a low-grade and persistent pro-inflammatory response. Indeed, studies have demonstrated wheezing and chest tightness in subjects with acute inhalation of propylene glycol or vegetable glycerin,46–48 indicating that these e-cig constituents are detrimental to lung health. Further, the very recent surge in cases of acute lung injury induced by e-cig vaping suggests that certain e-cig formulation may produce exuberant inflammatory responses in the small airway epithelium and lung parenchyma. This effect in vivo is likely to be more pronounced than in our SAEC culture model, due to simultaneous injurious effects of e-cig vaping on other cell types not studied here, such as inflammatory cells, fibroblasts, and lung endothelial cells as we previously reported.49
Airway mucins are large polymeric O-linked glycoproteins that form about 30% of the solid components of airway mucus.17 Mucins are produced and then secreted by airway goblet cells and by submucosal glands in the large conducting airways. Cigarette smoking is a major risk factor for the development of goblet cell metaplasia and mucus hypersecretion; indeed, small airway mucus obstruction is a characteristic feature of COPD.15–17 While there are only a few studies, a link is emerging between e-cigs, mucus production and mucociliary clearance. Elevated MUC5AC has been observed in the sputum of e-cigarette users compared to non-users,13 while decreased ciliary beat frequency associated with patches of secretory material on ciliated cells have been detected in cultured nasal epithelial cells exposed to e-vapor.50 Furthermore, animal models have demonstrated an increase in mucus production in mice exposed to e-cigarettes and an impairment of mucociliary clearance in e-cig-exposed sheep.51,52 Several cytokines, including IL6, have been shown to induce the production of MUC5AC in epithelial cells.38,39,53 Our study demonstrates the first connection between MUC5AC production and IL6 in SAEC due to exposure to e-vapor. Our data demonstrated that an acute exposure of SAEC to NF e-vapor but not Nic e-vapor increased IL6 which directly contributed to an increase in airway MUC5AC production as an anti-IL6 antibody significantly inhibited the mucin induction by NF e-vapor. These results are based on the study of six separate human donor lungs from both males and females, free of acute or chronic lung disease, and overwhelmingly nonsmokers. Thus, it is unlikely that our study on the acute effects of NF e-cig vaping on SAEC is limited by replicates from a single individual, and may, therefore, be generalizable to all healthy smoking-naïve SAEC.
One intriguing finding in the current study is that increased intracellular mucin production by NF e-vapor was not accompanied by increased mucin secretion into the apical surface of SAEC cultured at air-liquid interface. This may be because MUC5AC production and secretion are regulated by distinct processes15 that are differentially affected by e-vapor. In uninflamed airways, the rate of mucin production and the rate of secretion is roughly equivalent, such that there is only minimal mucin stored in secretory granules. Stimulation of mucin production, for example, by IL6, IL13 or IL17, leads to an increase of stored intracellular mucin but not necessarily to a large increase in secreted mucin. To dramatically increase the rate of secretion, goblet cells must be stimulated by high levels of extracellular agonists such as ATP, acetylcholine, or histamine.16,54 Since no extracellular agonist was applied in our SAEC culture model, it may not be surprising that secreted MUC5AC levels did not increase. Accumulation of intracellular MUC5AC due to repeated exposures of SAEC to e-cigs over several days may lead to a small increase in secreted MUC5AC and should be investigated. Changes in the amount of secreted mucins may also occur due to goblet cell metaplasia, but for this to occur, repeated exposures of cells to e-cigs over a time period much longer than 24 hrs used here would be necessary.14,55 This may explain why secreted mucin was elevated in the sputum of e-cig users,13 but not in ex vitro acute exposure models.
Several limitations are present in our current study. Our results using a model of acute exposure of SAEC to e-vapor cannot be directly extrapolated to chronic or repeated e-vapor exposures, which will need to be addressed in future studies. It is unclear which components in NF-e-vapor may be responsible for the increased IL6 and MUC5AC production. Much of the non-nicotine components of e-cigs reflect a proprietary mixture of chemicals that may have wide-ranging effects on cytotoxicity and cytokine production.2 A more global exploration of the upstream signaling pathways involved in IL6 production following acute NF e-vapor exposures will provide further insight into how these chemicals engage host cell responses. The effects of e-cig on IL6 production vary among different studies, likely due to the differences in the cell types used, e-cig exposure methods and brands (flavored vs non-flavored). Some studies have shown that IL-6 release was reduced when cells were exposed to flavored e-cig.56,57 Given the recent report of death related to vaping associated lung injury (EVALI),58,59 studies using the human lung tissue such as precision-cut lung slices are warranted to better understand how e-vapor with various additives such as vitamin E acetate causes lung injury.
Conclusion
By leveraging our access to human small airway epithelial cells of multiple human donors, this study has provided the first evidence that a single exposure to e-vapor is sufficient to increase distal airway mucin production in an IL6-dependent manner.
Funding Statement
This work was supported by the National Institutes of Health under grant R01 HL144396.
Disclosure
Ms Taylor Crue reports grants from National Institutes of Health, during the conduct of the study. Dr Brian Day reports grants from NIH, during the conduct of the study; personal fees from Aldeyra Therapeutics, outside the submitted work. Dr Mari Numata reports grants from NIH, grants from Flight Attendant Medical Research Institute, Inc. (FAMRI) Clinical Innovator Award Program, during the conduct of the study; In addition, Dr Mari Numata has a patent Surfactant Lipids, Composition Thereof and Uses Thereof. US Patent 8367643 issued, a patent Surfactant Lipids, Composition Thereof and Uses Thereof. US Patent 8796243 issued, a patent Surfactant Lipids, Composition Thereof and Uses Thereof. US Patent 9861649 issued. Dr Dennis R Voelker reports grants from NIH-NIGMS, grants from NIH-NHLBI, grants from FAMRI, during the conduct of the study. Dr Irina Petrache reports personal fees from Allinaire Therapeutics, outside the submitted work. The authors report no other conflicts of interest in this work.
References
- 1.Bhalerao A, Sivandzade F, Archie SR, Cucullo L. Public health policies on E-cigarettes. Curr Cardiol Rep. 2019;21(10):111. doi: 10.1007/s11886-019-1204-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Clapp PW, Jaspers I. electronic cigarettes: their constituents and potential links to asthma. Curr Allergy Asthma Rep. 2017;17(11):79. doi: 10.1007/s11882-017-0747-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hogg JC, Macklem PT, Thurlbeck WM. Site and nature of airway obstruction in chronic obstructive lung disease. N Engl J Med. 1968;278(25):1355–1360. doi: 10.1056/NEJM196806202782501 [DOI] [PubMed] [Google Scholar]
- 4.van den Berge M, Ten Hacken NHT, Cohen J, Douma WR, Postma DS. Small airway disease in asthma and COPD: clinical implications. Chest. 2011;139(2):412–423. doi: 10.1378/chest.10-1210 [DOI] [PubMed] [Google Scholar]
- 5.Stockley JA, Cooper BG, Stockley RA, Sapey E. Small airways disease: time for a revisit? Int J Chron Obstruct Pulmon Dis. 2017;12:2343–2353. doi: 10.2147/COPD.S138540 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Eddleston J, Lee RU, Doerner AM, Herschbach J, Zuraw BL. Cigarette smoke decreases innate responses of epithelial cells to rhinovirus infection. Am J Respir Cell Mol Biol. 2011;44(1):118–126. doi: 10.1165/rcmb.2009-0266OC [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Miyashita L, Suri R, Dearing E, et al. E-cigarette vapour enhances pneumococcal adherence to airway epithelial cells. Eur Respir J. 2018;51(2):1701592. doi: 10.1183/13993003.01592-2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gaurav R. Vaping Away Epithelial Integrity. Am J Respir Cell Mol Biol. 2019;61(2):127–129. doi: 10.1165/rcmb.2019-0016ED [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wu Q, Jiang D, Minor M, Chu HW. Electronic cigarette liquid increases inflammation and virus infection in primary human airway epithelial cells. PLoS One. 2014;9(9):e108342. doi: 10.1371/journal.pone.0108342 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Higham A, Bostock D, Booth G, Dungwa JV, Singh D. The effect of electronic cigarette and tobacco smoke exposure on COPD bronchial epithelial cell inflammatory responses. Int J Chron Obstruct Pulmon Dis. 2018;13:989–1000. doi: 10.2147/COPD.S157728 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Shen Y, Wolkowicz MJ, Kotova T, Fan L, Timko MP. Transcriptome sequencing reveals e-cigarette vapor and mainstream-smoke from tobacco cigarettes activate different gene expression profiles in human bronchial epithelial cells. Sci Rep. 2016;6(1):23984. doi: 10.1038/srep23984 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Glynos C, Bibli SI, Katsaounou P, et al. Comparison of the effects of e-cigarette vapor with cigarette smoke on lung function and inflammation in mice. Am J Physiol Lung Cell Mol Physiol. 2018;315(5):L662–L672. doi: 10.1152/ajplung.00389 [DOI] [PubMed] [Google Scholar]
- 13.Reidel B, Radicioni G, Clapp PW, et al. E-cigarette use causes a unique innate immune response in the lung, involving increased neutrophilic activation and altered mucin secretion. Am J Respir Crit Care Med. 2018;197(4):492–501. doi: 10.1164/rccm.201708-1590OC [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cao X, Wang Y, Xiong R, et al. Cigarette whole smoke solutions disturb mucin homeostasis in a human in vitro airway tissue model. Toxicology. 2018;409:119–128. doi: 10.1016/j.tox.2018.07.015 [DOI] [PubMed] [Google Scholar]
- 15.Dijkstra AE, de Jong K, Boezen HM, et al. Risk factors for chronic mucus hypersecretion in individuals with and without COPD: influence of smoking and job exposure on CMH. Occup Environ Med. 2014;71(5):346–352. doi: 10.1136/oemed-2013-101654 [DOI] [PubMed] [Google Scholar]
- 16.Jaramillo AM, Azzegagh Z, Tuvim MJ, Dickey BF. Airway mucin secretion. Ann Am Thorac Soc. 2018;15(Suppl3):S164–S170. doi: 10.1513/AnnalsATS.201806-371AW [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fahy JV, Dickey BF. Airway mucus function and dysfunction. N Engl J Med. 2010;363(23):2233–2247. doi: 10.1056/NEJMra0910061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chun LF, Moazed F, Calfee CS, Matthay MA, Gotts JE. Pulmonary toxicity of e-cigarettes. Am J Physiol Lung Cell Mol Physiol. 2017;313(2):L193–L206. doi: 10.1152/ajplung.00071.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Staudt MR, Salit J, Kaner RJ, Hollmann C, Crystal RG. Altered lung biology of healthy never smokers following acute inhalation of E-cigarettes. Respir Res. 2018;19(1):78. doi: 10.1186/s12931-018-0778-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sosnowski TR, Odziomek M. Particle size dynamics: toward a better understanding of electronic cigarette aerosol interactions with the respiratory system. Front Physiol. 2018;9:853. doi: 10.3389/fphys.2018.00853 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pichelstorfer L, Hofmann W, Renate W-H, Yurteri CÜ, McAughey J. Simulation of aerosol dynamics and deposition of combustible and electronic cigarette aerosols in the human respiratory tract. J Aerosol Sci. 2016;99:8. doi: 10.1016/j.jaerosci.2016.01.017 [DOI] [Google Scholar]
- 22.Wang G, Lou HH, Salit J, et al. Characterization of an immortalized human small airway basal stem/progenitor cell line with airway region-specific differentiation capacity. Respir Res. 2019;20(1):196. doi: 10.1186/s12931-019-1140-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chu HW, Balzar S, Seedorf GJ, et al. Transforming growth factor-beta2 induces bronchial epithelial mucin expression in asthma. Am J Pathol. 2004;165:1097–1106. doi: 10.1016/s0002-9440(10)63371-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhang W, Case S, Bowler RP, Martin RJ, Jiang D, Chu HW. Cigarette smoke modulates PGE(2) and host defence against Moraxella catarrhalis infection in human airway epithelial cells. Respirology. 2011;16(3):508–516. doi: 10.1111/j.1440-1843.2010.01920.x [DOI] [PubMed] [Google Scholar]
- 25.Thorne D, Adamson J. A review of in vitro cigarette smoke exposure systems. Exp Toxicol Pathol. 2013;65(7–8):1183–1193. doi: 10.1016/j.etp.2013.06.001 [DOI] [PubMed] [Google Scholar]
- 26.Chiadmi F, Schlatter J. Simultaneous determination of cotinine and trans-3-hydroxycotinine in urine by automated solid-phase extraction using gas chromatography-mass spectrometry. Biomed Chromatogr. 2014;28(4):453–458. doi: 10.1002/bmc.3159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Marsot A, Simon N. Nicotine and cotinine levels with electronic cigarette: a review. Int J Toxicol. 2016;35(2):179–185. doi: 10.1177/1091581815618935 [DOI] [PubMed] [Google Scholar]
- 28.Baqir M, Chen CZ, Martin RJ, et al. Cigarette smoke decreases MARCO expression in macrophages: implication in Mycoplasma pneumoniae infection. Respir Med. 2008;102(11):1604–1610. doi: 10.1016/j.rmed.2008.05.002 [DOI] [PubMed] [Google Scholar]
- 29.Wu Q, Martin RJ, Rino JG, Jeyaseelan S, Breed R, Chu HW. A deficient TLR2 signaling promotes airway mucin production in Mycoplasma pneumoniae-infected allergic mice. Am J Physiol Lung Cell Mol Physiol. 2007;292(5):L1064–1072. doi: 10.1152/ajplung.00301.2006 [DOI] [PubMed] [Google Scholar]
- 30.Bartlett JA, Albertolle ME, Wohlford-Lenane C, et al. Protein composition of bronchoalveolar lavage fluid and airway surface liquid from newborn pigs. Am J Physiol Lung Cell Mol Physiol. 2013;305(3):L256–66. doi: 10.1152/ajplung.00056.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Song KS, Choi YH, Kim JM, Lee H, Lee TJ, Yoon JH. Suppression of prostaglandin E2-induced MUC5AC overproduction by RGS4 in the airway. Am J Physiol Lung Cell Mol Physiol. 2009;296(4):L684–692. doi: 10.1152/ajplung.90396.2008 [DOI] [PubMed] [Google Scholar]
- 32.Zheng T, Liu W, Oh SY, et al. IL-13 receptor alpha2 selectively inhibits IL-13-induced responses in the murine lung. J Immunol. 2008;180(1):522–529. doi: 10.4049/jimmunol.180.1.522 [DOI] [PubMed] [Google Scholar]
- 33.Ordonez CL, Khashayar R, Wong HH, et al. Mild and moderate asthma is associated with airway goblet cell hyperplasia and abnormalities in mucin gene expression. Am J Respir Crit Care Med. 2001;163(2):517–523. doi: 10.1164/ajrccm.163.2.2004039 [DOI] [PubMed] [Google Scholar]
- 34.Wang H, He L, Liu B, et al. Establishment and comparison of air-liquid interface culture systems for primary and immortalized swine tracheal epithelial cells. BMC Cell Biol. 2018;19(1):10. doi: 10.1186/s12860-018-0162-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zanin M, Baviskar P, Webster R, Webby R. The interaction between respiratory pathogens and mucus. Cell Host Microbe. 2016;19(2):159–168. doi: 10.1016/j.chom.2016.01.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Innes AL, Woodruff PG, Ferrando RE, et al. Epithelial mucin stores are increased in the large airways of smokers with airflow obstruction. Chest. 2006;130(4):1102–1108. doi: 10.1378/chest.130.4.1102 [DOI] [PubMed] [Google Scholar]
- 37.Roy MG, Livraghi-Butrico A, Fletcher AA, et al. Muc5b is required for airway defence. Nature. 2014;505(7483):412–416. doi: 10.1038/nature12807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chen Y, Thai P, Zhao YH, Ho YS, DeSouza MM, Wu R. Stimulation of airway mucin gene expression by interleukin (IL)-17 through IL-6 paracrine/autocrine loop. J Biol Chem. 2003;278(19):17036–17043. doi: 10.1074/jbc.M210429200 [DOI] [PubMed] [Google Scholar]
- 39.Neveu WA, Allard JB, Dienz O, et al. IL-6 is required for airway mucus production induced by inhaled fungal allergens. J Immunol. 2009;183(3):1732–1738. doi: 10.4049/jimmunol.0802923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Scott A, Lugg ST, Aldridge K, et al. Pro-inflammatory effects of e-cigarette vapour condensate on human alveolar macrophages. Thorax. 2018;73(12):1161–1169. doi: 10.1136/thoraxjnl-2018-211663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Czekala L, Simms L, Stevenson M, Tschierske N, Maione AG, Walele T. Toxicological comparison of cigarette smoke and e-cigarette aerosol using a 3D in vitro human respiratory model. Regul Toxicol Pharmacol. 2019;103:314–324. doi: 10.1016/j.yrtph.2019.01.036 [DOI] [PubMed] [Google Scholar]
- 42.Kalra R, Singh SP, Pena-Philippides JC, Langley RJ, Razani-Boroujerdi S, Sopori ML. Immunosuppressive and anti-inflammatory effects of nicotine administered by patch in an animal model. Clin Diagn Lab Immunol. 2004;11(3):563–568. doi: 10.1128/CDLI.11.3.563-568.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sopori M. Effects of cigarette smoke on the immune system. Nat Rev Immunol. 2002;2(5):372–377. doi: 10.1038/nri803 [DOI] [PubMed] [Google Scholar]
- 44.Gomes JP, Watad A, Shoenfeld Y. Nicotine and autoimmunity: the lotus’ flower in tobacco. Pharmacol Res. 2018;128:101–109. doi: 10.1016/j.phrs.2017.10.005 [DOI] [PubMed] [Google Scholar]
- 45.Benowitz NL, Hukkanen J, Jacob P 3rd. Nicotine chemistry, metabolism, kinetics and biomarkers. Handb Exp Pharmacol. 2009;192:29–60. doi: 10.1007/978-3-540-69248-5_2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Phillips B, Titz B, Kogel U, et al. Toxicity of the main electronic cigarette components, propylene glycol, glycerin, and nicotine, in Sprague-Dawley rats in a 90-day OECD inhalation study complemented by molecular endpoints. Food Chem Toxicol. 2017;109:315–332. doi: 10.1016/j.fct.2017.09.001 [DOI] [PubMed] [Google Scholar]
- 47.Boulay ME, Henry C, Bosse Y, Boulet LP, Morissette MC. Acute effects of nicotine-free and flavour-free electronic cigarette use on lung functions in healthy and asthmatic individuals. Respir Res. 2017;18(1):33. doi: 10.1186/s12931-017-0518-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Varughese S, Teschke K, Brauer M, Chow Y, van Netten C, Kennedy SM. Effects of theatrical smokes and fogs on respiratory health in the entertainment industry. Am J Ind Med. 2005;47(5):411–418. doi: 10.1002/ajim.20151 [DOI] [PubMed] [Google Scholar]
- 49.Schweitzer KS, Chen SX, Law S, et al. Endothelial disruptive proinflammatory effects of nicotine and e-cigarette vapor exposures. Am J Physiol Lung Cell Mol Physiol. 2005;47:411–418. doi: 10.1002/ajim.20151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Carson JL, Zhou L, Brighton L M, et al. Temporal structure/function variation in cultured differentiated human nasal epithelium associated with acute single exposure to tobacco smoke or E-cigarette vapor. Inhal Toxicol. 2017;29(3):137–144. doi: 10.1080/08958378.2017.1318985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Garcia-Arcos I, Geraghty P, Baumlin N, et al. Chronic electronic cigarette exposure in mice induces features of COPD in a nicotine-dependent manner. Thorax. 2016;71(12):1119–1129. doi: 10.1136/thoraxjnl-2015-208039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Chung S, Baumlin N, Dennis JS, et al. Electronic cigarette vapor with nicotine causes airway mucociliary dysfunction preferentially via TRPA1 receptors. Am J Respir Crit Care Med. 2019;200(9):1134–1145. doi: 10.1164/rccm.201811-2087OC [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Smirnova MG, Kiselev SL, Birchall JP, Pearson JP. Up-regulation of mucin secretion in HT29-MTX cells by the pro-inflammatory cytokines tumor necrosis factor-alpha and interleukin-6. Eur Cytokine Netw. 2001;12(1):119–125. [PubMed] [Google Scholar]
- 54.Shumilov D, Popov A, Fudala R, et al. Real-time imaging of exocytotic mucin release and swelling in Calu-3 cells using acridine orange. Methods. 2014;66(2):312–324. doi: 10.1016/j.ymeth.2013.09.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Schamberger AC, Staab-Weijnitz CA, Mise-Racek N, Eickelberg O. Cigarette smoke alters primary human bronchial epithelial cell differentiation at the air-liquid interface. Sci Rep. 2015;5(1):8163. doi: 10.1038/srep08163 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Cervellati F, Muresan XM, Sticozzi C, et al. Comparative effects between electronic and cigarette smoke in human keratinocytes and epithelial lung cells. Toxicol in Vitro. 2014;28(5):999–1005. doi: 10.1016/j.tiv.2014.04.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ween MP, Hamon R, Macowan MG, Thredgold L, Reynolds PR, Hodge SJ. Effects of E-cigarette E-liquid components on bronchial epithelial cells: demonstration of dysfunctional efferocytosis. Respirology. 2019. doi: 10.1111/resp.13696 [DOI] [PubMed] [Google Scholar]
- 58.Prevention CfDCa. Outbreak of lung injury associated with the use of E-cigarette, or vaping, products. 2019.
- 59.NIH. Public Health Consequences of E-Cigarettes. In:2019.