

Abstract
Sauropods were among the most diverse lineages of dinosaurs, with an ample geographic distribution throughout the Mesozoic. This evolutionary success is largely attributed to neck elongation and its impact on feeding efficiency. However, how neck elongation influenced exactly on feeding strategies is subject of debate. The process of mounting a nearly complete virtual skeleton of Spinophorosaurus nigerensis, from the Middle (?) Jurassic of Niger, has revealed several previously unknown osteological adaptations in this taxon. Wedged sacral and posterior dorsal vertebrae cause the presacral column to deflect antero-dorsally. This, together with elongated scapulae and humeri make the anterior region of the skeleton vertically lengthened. Also, elongated prezygapophyseal facets on the cervical vertebrae and a specialized first dorsal vertebra greatly increase the vertical range of motion of the neck. These characters support this early eusauropod as a more capable high browser than more basally branching sauropods. While limb proportions and zygapophyseal facets vary among Eusauropoda, the sacrum retained more than 10° of wedging in all Eusauropoda. This implied a functional constraint for sauropod species which evolved lower browsing feeding strategies: the antero-dorsal sloping caused by the sacrum had to be counteracted with further skeletal modifications, e.g. a ventrally curved mid to anterior presacral spine to hinder the dorsal slope of the whole presacral series caused by the wedged sacrum. This suggests that at least the last common ancestor of Eusauropoda developed high browsing capabilities, partially due to the modified wedged sacrum, likely a potential synapomorphy of the clade and key in the evolutionary history of the group.
Subject terms: Palaeontology, Biomechanics
Introduction
Sauropods were the earliest large phytophagous dinosaurs, with an unparalleled disparity in body size, since their Late Triassic origin until their demise at the end of the Cretaceous1. Their quadrupedal, long-necked, long-tailed body plan remained fixed during their evolution, although both relatively shorter necks2,3 and extremely long necks4–7 appeared on several different clades of sauropods. This characteristic body plan had a direct impact on the feeding efficiency of these animals1,8, with some changes on this body plan being likely feeding adaptations, key in their evolutionary history8. However, whether these adaptations allowed or not high browsing capabilities has been the subject of a lively debate9–15. Previous studies on sauropod feeding capabilities based on their postcrania have focused mostly on neck posture and range of motion6,9,11,12,15–17. Evidence from neutral articulation of the bones and computerized analyses suggested straighter, less flexible necks9,14,16, with some authors suggesting most sauropods could barely raise the neck above shoulder height9,16. However, evidence stemming from comparisons with extant relatives and analyses on inter-vertebral stress suggested elevated, more flexible and curved necks10–13,18. More recent studies have revealed that the relationship between sauropod neck posture and feeding habits is more complex than previously thought14,19: forelimb/hindlimb proportions15 or scapula orientation and position probably had a strong role in browsing capabilities as well14,20. Unfortunately, the fragmentary nature of the known sauropod fossil record has been a large caveat in understanding how the axial skeleton, girdles and limbs vary within sauropod dinosaurs, making the study of their functional morphology complex.
In 2007, the holotypic specimen of Spinophorosaurus nigerensis was unearthed from the Middle (?) Jurassic of Niger21,22, being one of the most complete single specimens retrieved among basally branching eusauropods. The exceptional preservation and completeness of this specimen enabled to generate a virtual skeletal mount with less uncertainty than previous virtual sauropod mounts. A high-resolution virtual skeleton was used to test (i) what the body plan and feeding capabilities of Spinophorosaurus were, (ii) previous claims on sauropod neck functional morphology and (iii) whether previous reconstructions were or not accurate.
Results
Spinophorosaurus body plan
Mounting the digital skeleton has revealed Spinophorosaurus had tall shoulders and an elevated neck, well above shoulder level, in osteologically induced curvature (OIC; Fig. 1, see Terminology in SI appendix, Section 1). The OIC of Spinophorosaurus is the result of articulating a skeleton in osteologically neutral pose (ONP, maximum zygapophyseal overlap, see Material and Methods below) with only two vertebrae visible at once. This makes the mount more a product of bone geometry alone, with preconceived notions minimized14,23. The resulting body plan, analogous to that of the more extremely verticalized brachiosaurid sauropods, is due to two main skeletal characters: (i) an elongated forelimb, the humerus being longer relative to the scapula and femur than those of most other sauropods, and the scapula being slightly longer than the femur (Table 1); and (ii) an acute wedged sacrum and posteriormost dorsal vertebrae, which deflect the presacral vertebrae dorsally in neutral articulation (Fig. 1).
Figure 1.
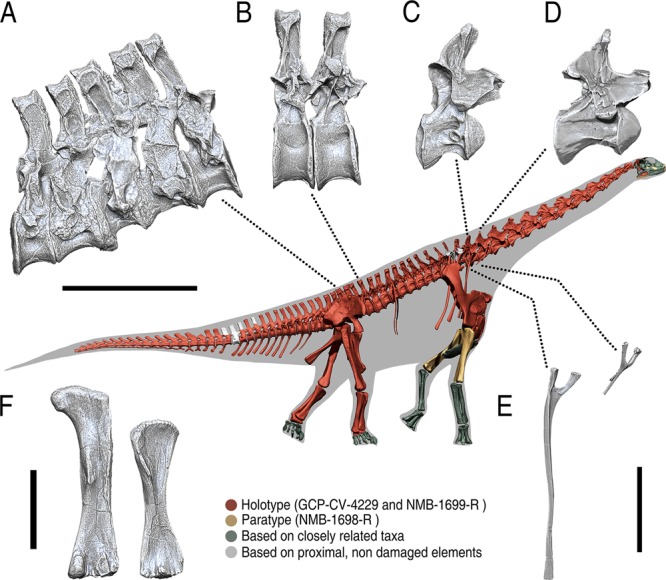
High browsing adaptations in Spinophorosaurus nigerensis. Skeletal reconstruction in osteologically neural pose with bones color-coded according to their provenance, with the holotype in red, paratype in yellow, and bones inferred from close relatives in green. White indicates reconstructed bone. (A) Sacrum, showing the 20° angle wedging. (B) 12th and 13th dorsal vertebrae, showing a slight acute wedging. (C) Partially cervicalized 1st dorsal vertebra. (D) 12th Cervical Vertebra. (E) First dorsal rib and last cervical rib. (F) Humerus and femur, to scale. Scales A-F = 500 mm.
Table 1.
Length measurements and ratios of different sauropod taxa, with specimen number indicated.
| Taxa | Sc Length | H Length | F Length | F/H Ratio | Sacrum Angle |
|---|---|---|---|---|---|
| Spinophorosaurus nigerensis GCP-CV-4229 | 1243 | 1014* | 1215 | 1.20 | 20.1 |
| Shunosaurus lii ZDM T5402 | 902 | 670 | 1200 | 1.79 | 14.7 |
| Mamenchisaurus youngi ZDM0083 | 1190 | 825 | 1160 | 1.40 | 15.8 |
| Omeisaurus tianfuensis ZDM T5704 | 1330 | 1040 | 1280 | 1.23 | 18.5 |
| Jobaria tiguidensis MNN TIG4 | ? | 1040* | 1800 | 1.73 | 15.0 |
| Dicraeosaurus hansemanni MB.R.4886 | ? | 620 * | 1220 | 1.96* | 11.0 |
| Diplodocus carnegii. CM94 | 1240 | ? | 1542 | ? | 16.0 |
| Apatosaurus louisae CM3018 | 1640 | 1150 | 1785 | 1.55 | 15.5 |
| Brontosaurus excelsus YPM 1980 | ? | 1114 | 1910 | 1,72 | 15.0 |
| Tehuelchesaurus benitezii MPEF-PV-1125 | 1801 | 1183 | 1550 | 1.31 | 20? |
| Camarasaurus grandis YPM 1901 | 1.155 | 856 | 1172 | 1.37 | 17.5 |
| Brachiosaurus altithorax FMNH P25107 | ? | 2042 | 2025 | 0.99 | 28.8 |
| Giraffatitan brancai MB.R 2181 (formerly SII) | 1930 | 2130 | 1990* | 0.93 | 25.0 |
| Dreadnoughtus schrani MPM PV 1156 | 1772 | 1600 | 1910 | 1,19 | 21.1 |
| Opisthocoelicaudia skarzynski ZPAL MgD1/48 | 1180 | 1000 | 1395 | 1,39 | 14.1 |
| Kotasaurus yamanpalliensis | ? | *770 | 1130 | 1,46* | 3.7 |
| Melanorosaurus readi NM QR1551 | 489 | 450 | 638 | 1,41 | 1.0 |
| Lufengosaurus huenei IVPP 15 | ? | 335 | 560 | 1.67 | 5.4 |
| Jingshanosaurus xinwaensis LFGT-ZLJ0113 | ? | 470 | 850 | 1.80 | 1.5 |
| Yunnanosaurus huangi IVPP V20 | 305 | 231 | 435 | 1,88 | 2.1 |
Asterisk (*) indicates estimated measurements scaled from other specimens of the same species (see Supplementary Material). The sacrum angle and F/H ratio are plotted in the XY graphic on Fig. 2B. All lengths in mm. Angles in degrees. Sc = scapula, H = humerus, F = femur.
Given the position of the pelvis in Sauropoda, with an antero-posteriorly projected ilium and a mesopubic and opisthoischiatic condition24 (which allows the femur to be upright and graviportal), the coalesced sacrum is situated so that the posterior face of the last sacral centrum is sub-vertical, making the tail sub-horizontal and the presacral series to slope antero-dorsally in ONP. This way, when the last dorsal vertebra articulates with the sacrum, it deflects 20° from the centrum of the first caudal vertebra in lateral view (Fig. S4). In addition, the slightly wedged centra of the two last dorsal vertebrae (Fig. 1B) deflect the dorsal series further, making the first dorsal vertebra deflect 5° from the last dorsal centrum (overall 25° from the first caudal). The cervical series is almost straight, but with a slight sigmoidal dorsal deflection, with the axis deflecting 5° from the first dorsal vertebra (overall 30° from the first caudal), a little different from the straight and slightly ventrally deflected posture previously described for other sauropods9,15. When using the lateral semicircular canals (LSCs) of the inner ear25 to reconstruct the position of the skull, it forms an angle of 10° with the neck, compatible with the non-deflected, osteologically induced curvature of the presacral vertebrae (Fig. 1).
When all scapular girdle bones remain in articulation and are placed in the ribcage following the results of independent osteological, myological and phylogenetic bracketing (see Methods and SI appendix, Section 5), the scapular girdle is mounted in the following way: sub-vertical scapulae, coracoids antero-ventral to ribcage, scapular heads anterior to the ribcage and glenoids a bit ventral to the distal tips of anterior dorsal ribs. This allows for an upright articulation of the humerus, as is known to be the condition for sauropods. The distal humerus to floor distance is 1.11 m.
The forearm and hand are not yet known in any Spinophorosaurus specimen. However, the aforementioned distance implies a forearm (0.748 m ulna) and hand (0.278 m metacarpal III + 0.06 m carpals) with similar proportions relative to the humerus to those of individuals of closely related non-neosauropod eusauropods (SI appendix, Section 4; Tables S1, S2). These reconstructed forearm and hand are, therefore, more parsimonious than assuming a proportionately longer or shorter forearm and/or hand.
Summing up, the osteologically induced curvature of the presacral column of Spinophorosaurus makes its skull deflect 30° dorsally from the first caudal vertebra in osteologically neutral posture, with the snout situated at about 5 m above the ground, more than twice as high as the shoulder and acetabulum, at 2.15 m (see Fig. 2A).
Figure 2.

The verticalization of sauropod feeding envelopes. (A) Increased neck range of motion in Spinophorosaurus in the dorso-ventral plane, with the first dorsal vertebra as the vertex and 0° marking the ground. Poses shown: (1) maximum dorsiflexion; (2) highest vertical reach of the head (7.16 m from the ground), with the neck 90° deflected; (3) alert pose sensu Taylor Wedel and Naish13; (4) osteological neutral pose sensu Stevens14; (5) lowest vertical reach of the head (0.72 m from the ground at 0°), with the head as close to the ground without flexing the appendicular elements; (6) maximum ventriflexion. Blue indicates the arc described between maximum and minimum head heights. Grey indicates the arc described between maximum dorsiflexion and ventriflexion. (B) Bivariant plot comparing femur/humerus proportion with sacrum angle. The proportion of humerus and femur are compared as a ratio of femur maximum length/humerus maximum length. Sacrum angle measures the angle the presacral vertebral series are deflected from the caudal series by sacrum geometry in osteologically neutral pose. Measurements and taxa on Table 1. Scale = 1000 mm.
Spinophorosaurus neck range of motion analysis
The neck of Spinophorosaurus has 12 moderately elongated cervical vertebrae (largest average Elongation Index, aEI = 3.63, see Supplementary Information Section 1 for details) with two features increasing range of motion in the dorsoventral plane. The prezygapophyseal facets on the cervical vertebrae are particularly antero-posteriorly elongated next to those of other sauropod cervical vertebrae (Fig. S5A). These elongated articulation facets provide a greater range of motion per joint than in other sauropods in comparable joint positions (Fig. S5C, D). The 12th cervical vertebra has all traits expected of a cervical, including a short and small cervical rib. The first dorsal vertebra of Spinophorosaurus shows a tendency to cervicalization, but it already bears a dorsal rib (Fig. 1E). This vertebra is more elongated than any of the other dorsal vertebrae and its centrum has a trapezoidal shape in lateral view (its anterior condyle is more dorsally located than the cotyle). Also, the prezygapophyseal rami are anteriorly projected, the prezygapophyseal facets have the same elongation present in the preceding cervical vertebra and the ventral keel is very pronounced. Most of these characters are missing in the second dorsal vertebra (Table S3). This partial cervicalization makes the first dorsal vertebra a functional cervicodorsal vertebra, sharing many convergences with the cervicalized first thoracic vertebra of giraffes: longer than wide prezygapophyseal facets, a ventral keel in the centrum or a centrum length intermediate between that of the preceding cervical and the following dorsal vertebra (Fig. S6, Table S3). Despite having a dorsal rib, the cervicothoracic vertebra of giraffes has a greater range of motion than any of the other thoracic vertebrae, but more reduced than the other cervical vertebrae26. This situation is also present in Spinophorosaurus, in which the first dorsal vertebra has an amount of osteological mobility greater than that of the following dorsal vertebrae but smaller than that of the preceding cervical vertebra (Table 2). This is particularly noticeable on its first dorsal vertebra and, to a lesser degree, on the second and third because their prezygapophyseal facets and centra are shorter than on the first dorsal.
Table 2.
Dorso-ventral range of motion values at the cervical-dorsal boundary for Spinophorosaurus nigerensis, Plateosaurus engledhardti, Okapia johnstoni and Giraffa camelopardalis, measured as degrees between maximum dorsiflexion and ventriflexion in the referred vertebra.
| DV 3 | DV 2 | DV 1 | LCV | PCV | |
|---|---|---|---|---|---|
| Okapia | 3 | 4 | 3 | 9 | 17.5 |
| Giraffa | 5 | 4 | 14 | 25–27 | 25–28 |
| Plateosaurus | 4 | 4 | 8 | 10 | 10 |
| Spinophorosaurus | *7 | *9 | 17 | 27 | 28 |
DV = Dorsal Vertebra; LCV = Last Cervical Vertebra; PCV = Penultimate Cervical Vertebra. *Asterisk indicates estimated measurements based on field pose (dorsiflexion) and osteological stop of ribs (ventriflexion). Data from Okapia and Giraffa from Gunji & Endo26. Data from Plateosaurus from Mallison52.
The range of motion of the neck alone (without deflecting the first dorsal from ONP) allows placing it vertical, with the skull antero-posterior axis perpendicular to the ground without disarticulation (Fig. 2A). The inner ear semicircular canals are relatively very large and slender25. This has been related with the perception of angular accelerations of the head of animals, predicting a highly flexible neck for Spinophorosaurus25, corroborated at least in the dorso-ventral plane by our range of motion analysis (Fig. 2A).
Discussion
Body plan and feeding capabilities of Spinophorosaurus
Although there has been a thorough debate on the feeding capabilities of different sauropod taxa, most analyses have focused on skull, teeth and neck functional morphology. Previous studies showed that pectoral girdle and forelimb position created a dorsal sloping of the presacral vertebrae14,20, suggesting that studying only cranial and cervical anatomy might be insufficient to understand sauropod body plan and feeding capabilities14. Mounting the virtual skeleton of Spinophorosaurus has revealed that the morphology of sacrum and posterior dorsal vertebrae also have a crucial role on the overall body plan by deflecting the presacral series from the caudal series in osteologically neutral pose (Fig. 1, Fig. 4S). Given the general position of the pelvis in Sauropoda, with an antero-posteriorly projected ilium and a mesopubic and opisthopubic condition24, the coalesced sacrum is situated so that the posterior face of the last sacral centrum is sub-vertical. This makes the presacral series to slope dorsally and the tail to be subhorizontal (Figs. 1 and 4S). Since a subhorizontal tail has been known to be present in the majority of known sauropods27–29, the OIC of the tail of Spinophorosaurus is therefore compatible with this condition.
Figure 4.
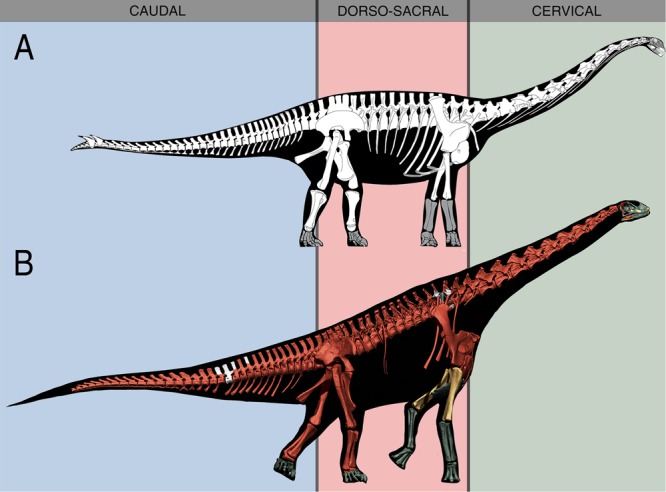
Skeletal reconstructions of Spinophorosaurus nigerensis. (A) Original skeletal reconstruction of Spinophorosaurus first published in 2009, modified from the original publication21. (B) Virtual skeletal reconstruction of Spinophorosaurus. All the skeletal reconstructions have been scaled to the same dorso-sacral sector length (between the last sacral vertebra and the first dorsal vertebra). Previous reconstruction lacked information on precise sacrum morphology, hence the radically different osteologically induced curvatures of the presacral vertebrae. Other bones which differ between the 2009 reconstruction and actual fossils (as further preparation has been carried out) are the ilium, the first three dorsal ribs, the cervicodorsal transition and the anterior caudal vertebrae.
The dorsal sloping of presacral vertebrae in sauropod dinosaurs was noticed by Gilmore when assembling Diplodocus sp. USNM 10865, in which the posteriormost dorsal vertebrae would dorsally deflect from the sacrum when in articulation27. Gilmore also noticed that the posteriormost dorsal vertebrae of USNM 10865 had an anteriorly pointed neural spine which was perpendicular to the ground when vertebrae were in articulation with one-another and with the sacrum27. The neural spines of posterior dorsal vertebrae DV12 to DV9 are also anteriorly directed in the holotype of Spinophorosaurus (Fig. 1). However, Gilmore remarked that around mid-thoracic region, the dorsal vertebrae series of USNM 10865 reversed its curvature, deflecting ventrally, resulting in an arched torso27. This arching of the torso is not present in Spinophorosaurus in osteologically neutral pose, nor there is any wedging in middle or anterior dorsal vertebrae to create a ventral deflection (Figs. 1, S4). Instead, the presacral column of Spinophorosaurus is very straight, with only a slight dorsal deflection of 10° from the last dorsal vertebra to the axis. This configuration of a dorsally sloping presacral column in Spinophorosaurus is compatible with the sub-vertical, more antero-ventrally placed scapulocoracoid proposed by independent osteological20,30, myological31, biomechanical32 and phylogenetical bracketing20 studies (see Methods and SI appendix, Section 5), as well as with an elongated scapula and humerus.
Forelimb length and shoulder height are important factors for estimating the feeding capabilities of sauropod dinosaurs. Those with longer forelimbs relative to their hindlimbs are interpreted as having high browsing capabilities14,15. Camarasaurus is a genus typically interpreted as a capable medium to high browser based on its shoulder height14,33, with a humerus to scapula ratio around 0.74 and a femur to humerus ratio around 1.37. Both the humerus to scapula ratio (0.816) and the estimated femur to humerus ratio (1.21) of Spinophorosaurus indicate a relatively larger humerus than that of Camarasaurus. The scapula of Spinophorosaurus is also slightly longer than its femur, whereas that of Camarasaurus is slightly shorter than its femur (Table 1). Therefore the preserved forelimb and pectoral girdle elements of Spinophorosaurus are relatively longer than those of Camarasaurus. All in all, Spinophorosaurus had a humerus and scapula relatively longer, making its shoulders relatively taller than those of most known sauropods, with the exception of at least Atlasaurus34 brachiosaurids10,33,35 and some titanosaurs (Table 1, Fig. 2B).
Regarding the missing forearm and hand bones in Spinophorosaurus, if they were as long relative to the humerus as in other non-neosauropod eusauropods (Table S1), they would be compatible with those in the virtual mount. Shorter or taller shoulder height would require, respectively, shorter or longer hypothetical forearm and hand relative to the humerus. This implies that any hypothesis regarding much shorter or much taller shoulders for Spinophorosaurus than those proposed in this reconstruction requires additional evolutionary steps (to acquire relatively shorter or longer missing elements), and are therefore less parsimonious.
This shows that the overall body plan of this sauropod has a clear tendency toward vertical lengthening due to its sacral wedging as well as its humerus to femur and scapula to femur proportions, especially when compared with its earlier branching relatives (Table 1). This tendency to verticalization, coupled with the increased dorso-ventral neck flexibility granted by the elongated prezygapophyseal facets, and coordinated by its relatively large and slender inner ear25, reveals a feeding envelope with a large vertical component. This feeding envelope is greater than those calculated for Diplodocus carnegii9, Apatosaurus louisae9 or Mamenchisaurus youngi6, and possibly greater than those of Camarasaurus or Haplocanthosaurus (SI appendix, Section 8; Fig. S4). Summing up, Spinophorosaurus can be interpreted as adept at high browsing, like modern giraffes are36: its skeleton conferred Spinophorosaurus the capability to browse on vegetation at nearly three times the height of its shoulders, and hence it might have been part of its feeding strategy.
Sauropod sacra as an evolutionary innovation
Given the high disparity of hindlimb/forelimb length proportions among Sauropoda37,38, the evidence of different feeding capabilities among different sauropods3,11,14 and the role of the wedged sacrum on the high browsing capabilities in Spinophorosaurus (due its high impact on vertebral column OIC), a hypothesis can be formulated: only sauropod dinosaurs with moderately to extremely elongated forelimbs would have strongly acute wedged sacra and dorsal vertebrae.
Interestingly, a comparison of sacral wedging with the relative lengths of humeri and femora reveals a correlation: lower femur maximum length/humerus maximum length ratios are associated with a higher wedging angle of the sacrum and vice versa (Fig. 2B). However, all eusauropod sauropods have wedged sacra, even those with shorter humeri (Figs. 2B, 3). Sauropods with extremely short humeri (dicraeosaurids and some titanosaurs) have sacrum wedging angles of around 10° or a little higher, whereas extremely tall-shouldered sauropods with humeri longer than femora (Brachiosauridae) have sacrum angles of up to 30° (Table 1). This sacrum wedging to humerus relative length correlation, however, is only found in Eusauropoda. There are few known non-eusauropod sauropods preserving sacral and limb material, but those which preserve complete sacra have very little to no wedging (i.e. Kotasaurus39 or Barapasaurus40) despite having columnar fore and hindlimbs (Fig. 3) and being obligatory quadrupeds. The sacrum in non-sauropod sauropodomorphs is also hardly wedged even in large, quadrupedal species (i.e. Melanorosaurus41). Some of these quadrupedal non-sauropod sauropodomorphs have relatively larger humeri than some eusauropods, that also were obligatory quadrupeds but had wedged sacra (i.e. Shunosaurus42). Therefore, a rectangular or barely wedged sacrum would be the basal condition in Sauropodomorpha, which eventually derived to a more strongly wedged one in Eusauropoda, 10° or more (Fig. 3). From this point on, the relative length of the humerus correlates with the amount of wedging in the sacrum, which does not happen in non-eusauropod sauropods (Fig. 2B).
Figure 3.

Time calibrated sauropodomorph phylogenetic relationships with emphasis on different body proportioned taxa. Different body proportions for sauropodomorph dinosaurs include (1) facultative quadrupedalism and medium height browsing in non-sauropod sauropodomorphs (e.g. Melanorosaurus) with non-wedged sacrum; (2) obligatory quadrupedalism and medium height browsing in basally branching sauropods with longer forelimbs but no wedged sacrum (e.g. Kotasaurus); (3) medium-high browsing in non-neosauropod eusauropods, with longer necks and forelimbs and an acute wedged sacrum (e.g. Spinophorosaurus); (4) medium-ground level browsing in dicraeosaurid and rebbachisaurid diplodocoid sauropods, with short necks, shorter forelimbs than non-neosauropod sauropodomorphs but with acute wedged sacrum and obtuse wedged dorsal vertebrae (e.g. Dicraeosaurus); (5) medium-height browsing in basally branching macronarian sauropods, with a wedge shaped sacrum and retroverted pelvis (e.g. Camarasaurus); (6) extreme high browsing in brachiosaurid sauropods, with extremely elongated necks, humeri longer than femora and extremely wedged sacra (e.g. Brachiosaurus); (7) extreme high browsing in euhelopodid titanosauriforms, with extremely long necks and extremely wedged sacra (e.g. Euhelopus); and (8) medium-low browsing in some lithostrotian titanosaurs, with shorter forelimbs than other titanosaurs and titanosauriforms but still retaining a wedged sacrum and with obtuse wedged dorsal vertebrae (e.g. Saltasaurini). A wedged sacrum is only found in Eusauropoda, and albeit the degree of wedging varies among sauropods, it never returns to the basal condition. Schematic bones obtained directly from digitized 3D models, except in Melanorosaurus53, Kotasaurus39, Dicraeosaurus54,55 and Euhelopus56, where photographs were used as reference. Femora and humeri are proportioned to each other following Table 1. Schematic bone color coding: rectangular sacra (blue); acute wedged sacra (red); obtuse wedged mid-dorsal vertebrae (yellow and green). Time callibration of nodes after Xu et al.47.
This correlation between the sacrum and forelimbs appears to have worked as a functional module during sauropod evolution, with evolutionary changes in limb proportions and sacra happening in a reciprocal way (Fig. 2B): changes in limb proportions resulted in modifications on the angulation of the sacrum wedging. In those sauropods with extremely short forelimbs and low/ground level browsing capabilities, the sacral wedging diminishes, making the presacral series deflect dorsally with a lesser angle. The sacrum, however, never returned to the basal condition of a more rectangular sacrum (Fig. 2B, Table 1). The acute sacrum wedging in sauropods with extremely short forelimbs, such as Dicraeosaurus, is also counteracted by an obtuse wedging in the dorsal vertebrae, which makes the presacral vertebral series deflect ventrally (Fig. 3, Figure 6.5C in Stevens and Parrish15). Some of these sauropods with shorter humeri had proportionately shorter necks than other sauropods43, and medium-height browsing44,45 or ground level browsing3,45 capabilities have been proposed for them. Some titanosaurs with relatively shorter necks and forelimbs might also have been medium to ground level browsers, and they also have wedged sacra and obtuse wedging in the dorsal vertebrae (Fig. 3, Saltasaurini), although more functional analyses are necessary. Sauropods with shorter forelimbs arose separately in at least two different clades according to phylogenetic analyses38,46–48 (Fig. 3, Diplodocoidea and Titanosauria) and all of them had close relatives with longer forelimbs and more wedged sacra. This implies that the acute wedged sacrum became irreversible for Eusauropoda, thus fixed in their body plan, likely a constraint for sauropods that evolved lower browsing feeding strategies.
The evolution of an acute wedged sacrum in sauropods appears to have been abrupt (Fig. 3), turning the non-wedged sacrum of basally branching Early to Middle Jurassic sauropods such as Barapasaurus and Kotasaurus, into a quite acute wedged one in Middle Jurassic sauropods such as Shunosaurus42 or Patagosaurus49. This derived sacrum evolved well after sauropods had already become obligate quadrupeds, earlier in their evolution46, implying it was not linked to the evolution of quadrupedality. Neck elongation has typically been regarded as one of the most important key innovations in sauropod evolution48, directly affecting the size of the feeding envelope1. Feeding envelope size, and with it, feeding efficiency, increased primarily from neck elongation8. However, the appearance of strongly wedged sacral and dorsal vertebrae and changes in limb proportions might also have been important factors on feeding envelope size. As evidenced for the first time in the almost complete skeleton of Spinophorosaurus (Figs. 1, 2B), further increase in the vertical component of feeding envelopes could have been achieved by relative forelimb elongation, longer prezygapophyseal facets in cervical vertebrae, cervicalization of anterior dorsals, and an acute wedged sacrum, the latter of which occurred in sauropods more deeply nested than Kotasaurus and Barapasaurus (Fig. 3).
The fact that an acute wedged sacrum of more than 10° remained present in all eusauropods, supports the trait as a synapomorphy rather than a convergent character (Fig. 3). Moreover, it may represent a new factor in the evolutionary cascade proposed for sauropod gigantism1,8, directly affecting the energy-efficient feeding selective advantage proposed in that evolutionary cascade.
Conclusions
The virtual skeleton of Spinophorosaurus nigerensis, the first digital mount using a single specimen from a sauropod dinosaur so complete, reveals a body plan very different from previous reconstructions of this animal, with the presacral column antero-dorsally deflected and relatively tall shoulders (Fig. 4). A 20° wedged sacrum in lateral view (Fig. 1) causes most of this dorsal sloping of the presacral column. Elongated pre and postzygapophyses and a partially cervicalized first dorsal vertebra enabled a greater dorso-ventral range of motion in Spinophorosaurus than in all previously studied sauropods. This enlarged vertical component of the feeding envelope, together with the dorsal sloping, show Spinophorosaurus had high browsing capabilities. Thus, skeletal adaptations to high browsing in sauropods were present as early as the Early-Middle Jurassic (Fig. 3).
All eusauropods shared a sacrum wedged more than 10° inherited from a common ancestor, which evolved in correlation with humerus/femur proportions in what could be part of a functional module: the greater the sacral wedging, the longer the humerus relative to the femur. This skeletal adaptation was likely a synapomorphy and a key factor on the evolutionary cascade proposed for sauropod gigantism and for the evolution of their body plan, as it remained present in all known eusauropods until their extinction at the K-Pg boundary.
Material and Methods
Spinophorosaurusdigitization
The holotype (GCP-CV-4229 and NMB-1699-R) and paratype (NMB-1698-R), found next to the holotype21 (SI appendix, Section 2), were both scanned by digital photogrammetry, using the protocol described by Mallison and Wings50. As many bones were collected in multiple good-fitting fragments but were not put back together after preparation when the scans were performed, the fragments were photographed separately and put together in Agisoft Photoscan 1.3 using a virtual alignment technique (SI appendix, Section 3). Although the skeleton is exquisitely preserved with minor symmetrical latero-medial compression, two middle caudal vertebrae and dorsal vertebrae DV2 and DV3 were affected by a fault, and therefore had slight shear distortion and some breakages. They were digitally restored under the protocol proposed by Vidal and Díez Díaz51 (SI appendix, detailed in Sections 3 and 4).
Mounting the virtualSpinophorosaurusskeleton
The skeleton was assembled in ZBrush 4R6 in osteologically neutral pose (ONP, see Terminology in SI appendix, Section 1), that is, maximum overlap of the pre- and postzygapophyseal facets. To reconstruct missing bones from the holotype specimen, the approach was a combination of scaling known elements from the paratype and phylogenetic interpolation for elements unknown in both specimens. Broken dorsal vertebrae DV2 and DV3 and caudal vertebra CdV15 were reconstructed using field photographs as reference, as well as the immediately anterior or posterior elements with good preservation (SI appendix, detailed in Section 4). The humerus to scapula maximum length ratio was measured in the paratype specimen of Spinophorosaurus as 0.816. The missing humerus of the holotype was therefore estimated to be 1014 mm long (0.816 times the 1243 mm scapula). The missing forelimb elements (ulna, radius and hand) were estimated based on known forelimb proportions of closely related non-neosauropod eusauropods (Table S3). The final model has a 748 mm ulna and a 278 mm longest metacarpal, both closer to the shorter end of the spectrum of the estimated lengths (Table S2) and compatible with the skeletal reconstruction proposed (SI appendix, detailed in Section 4 and Tables S2, S3). Axial elements were assembled following the protocol of Mallison52 (SI appendix, Section 5) in which vertebrae were articulated in pairs (only two elements visible at once, one remained static while the other was articulated in ONP) in order to minimize preconceived notions on axial skeleton “neutral” posture, maximizing an axial skeleton curvature based on bone geometry. It was done anterior to posterior and vice versa. If both skeletal assemblages had the same osteologically induced curvature (OIC), the mount was approved for further work.
The pelvic girdle was posed with an antero-posteriorly oriented ilium and a mesopubic condition, the widespread condition for all sauropods24. The pectoral girdle was mounted with sub-vertical scapulae with the coracoids antero-ventral to the ribcage, the scapular heads anterior to the ribcage and the glenoids a bit lower than the distalmost tips of the anterior dorsal ribs (Fig. 1). This position and orientation of the pectoral girdle is the only possible position which allows (i) to keep the scapulocoracoids articulated with the clavicles and interclavicle30 and fit within the ribcage, (ii) to not have the ribcage become an osteological stop for humerus retraction, (iii) to have functional cingulo-axial and shoulder musculature lines, particularly for M. subcoracoscapularis pars scapularis, which has its origin on the acromial region, on the medial side31,32 and which would otherwise be obstructed by the ribs, (iv) to place the costo-coracoideal articulation subparallel to the distal ribs axis as is the case of all extant non-mammalian tetrapods20, (v) to leave room dorsal to the distal expansion for the cartilaginous suprascapula, which would be the insertion point for M. rhomboideus31,32 (SI appendix, detailed in Section 5). This configuration of the scapulocoracoid has been also been independently reconstructed on previous works20,31,32. The appendicular skeleton was articulated in ONP, then posed in a fast walking gait for the figures.
Range of motion analysis
While there are standards for comparing skeletal neutral postures, there are no standards defined yet for range of motion analyses: different authors follow different criteria for assessing maximum articular excursions (SI appendix, Section 6). Here, the protocol of Mallison52 was used: vertebrae were deflected until only a minimum overlap of the facets was retained, that is, before they completely disarticulated (Fig. S5). That way, if accounting for a larger facet in vivo (as happens in extant archosaurs52), the range of motion is underestimated rather than overestimated. The center of rotation was the anteriormost part of the condyle of the posterior vertebra when the articulation was opisthocoelus, and at midheight of the posterior centrum face in platycoelus articulations. This would prevent the misalignment of the neural canals (SI appendix, detailed in Section 6). When discussing browsing heights (low/medium/high browsing) we are not referring to behavior/paleobiology, but to the physical capabilities the osteological range of motion enabled (See Terminology in SI).
Supplementary information
Acknowledgements
We thank I. Fierro and J.M. Marín from the PALDES project for their help while researching Spinophorosaurus and Diego Cobo for his help in illustrating Fig. 3. Thanks to L. Alcalá and R. Royo-Torres (Fundación Conjunto-Paleontológico de Teruel Dinópolis), D. Schwarz and V. Díez Díaz (Museum für Naturkunde, Berlin), I. Cerda (Museo de Cinco Saltos), D. Pol, J. Sterli and E. Ruigomez (Museo Paleontológico Egidio Feruglio), P. Ortiz and R. González (Instituto Miguel Lillo), M. Lamanna and A. Heinrici (Carnegie Museum of Natural History), D. Brickman and M. Fox (Yale Peabody Museum), M. Norell and C. Mehling (American Museum of Natural History), P. Makovicky and W. Simpson (Field Museum of Natural History) and P. Sereno, T. Keillor and E. Fitzgerald (University of Chicago) for their help when consulting specimens under their care. Thanks to K.A. Stevens and the AMNH for providing access to the virtual skeleton of a giraffe. Thanks to two anonymous reviewers whose comments, remarks and suggestions have helped us to improve key areas of this manuscript. Research was supported by the project CGL2015-68363-P of the Spanish Ministerio de Economía y Competitividad (MINECO). DV’s research is funded by a predoctoral FPI UNED Grant (Ref. 0531174813 Y0SC001170). PM’s research is funded by FCT/MCTES for one CEEC Individual contract (CEECIND/00726/2017).
Author contributions
D.V., P.M., F.O. and J.L.S. conceived the study. D.V. digitized the specimens and assembled the virtual skeleton. A.A. curated the holotype specimen. D.V., P.M., A.A. and F.O. analyzed the data. D.V., P.M., F.O., and J.L.S. wrote the paper.
Data availability
Most fossil material for the holotype specimen of Spinophorosaurus nigerensis (GCP-CV-4229) is temporally deposited at the Museo Paleontológico de Elche (Elche, Spain). The paratype specimen (NMB-1699-R) and part of the holotype (NMB-1698-R) of Spinophorosaurus nigerensis are temporally deposited at the Staatliches Naturhistorisches Museum (Braunchsweig, Germany). Both specimens will eventually be returned to the Musée National Boubou Hama (Niamey, Niger). The digital fossils used to build the virtual skeleton are deposited and accessioned at the Museo Paleontológico de Elche.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
is available for this paper at 10.1038/s41598-020-63439-0.
References
- 1.Sander PM, et al. Biology of the sauropod dinosaurs: The evolution of gigantism. Biol. Rev. 2011;86:117–155. doi: 10.1111/j.1469-185X.2010.00137.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rauhut OWM, Remes K, Fechner R, Cladera G, Puerta P. Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia. Nature. 2005;435:670–672. doi: 10.1038/nature03623. [DOI] [PubMed] [Google Scholar]
- 3.Sereno PC, et al. Structural extremes in a cretaceous dinosaur. PLoS One. 2007;2(11):e1230. doi: 10.1371/journal.pone.0001230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Jensen JA. Three new sauropod dinosaurs from the Upper Jurassic of Colorado. Gt. Basin Nat. 1985;45:697–709. [Google Scholar]
- 5.Wedel MJ, Cifelli RL, Sanders RK. Sauroposeidon proteles, a new sauropod from the Early Cretaceous of Oklahoma. J. Vertebr. Paleontol. 2000;20:109–114. doi: 10.1671/0272-4634(2000)020[0109:SPANSF]2.0.CO;2. [DOI] [Google Scholar]
- 6.Christian A, et al. Biomechanical reconstructions and selective advantages of neck poses and feeding strategies of Sauropods with the example of Mamenchisaurus youngi. PLoS One. 2013;8(10):e71172. doi: 10.1371/journal.pone.0071172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Taylor MP, Wedel MJ. The neck of Barosaurus: longer, wider and weirder than those of Diplodocus and other diplodocines. PeerJ Prepr. 2016;4:e67v2. [Google Scholar]
- 8.Sander PM. An evolutionary cascade model for sauropod dinosaur gigantism–overview, update and tests. PloS one. 2013;8(10):e78573. doi: 10.1371/journal.pone.0078573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Stevens KA, Parrish MJ. Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs. Science. 1999;284:798–800. doi: 10.1126/science.284.5415.798. [DOI] [PubMed] [Google Scholar]
- 10.Christian A. Neck posture and overall body design in sauropods. Foss. Rec. 2002;5:271–281. doi: 10.1002/mmng.20020050116. [DOI] [Google Scholar]
- 11.Christian A, Dzemski G. Reconstruction of the cervical skeleton posture of Brachiosaurus brancai Janensch, 1914 by an analysis of the intervertebral stress along the neck and a comparison with the results of different approaches. Foss. Rec. 2007;10:38–49. doi: 10.1002/mmng.200600017. [DOI] [Google Scholar]
- 12.Christian A. Some sauropods raised their necks—evidence for high browsing in Euhelopus zdanskyi. Biol. Lett. 2010;6:823–825. doi: 10.1098/rsbl.2010.0359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Taylor MP, Wedel MJ, Naish D. Head and Neck Posture in Sauropod Dinosaurs Inferred from Extant Animals. Acta Palaeontol. Pol. 2009;54:213–220. doi: 10.4202/app.2009.0007. [DOI] [Google Scholar]
- 14.Stevens KA. The articulation of sauropod necks: methodology and mythology. PLoS One. 2013;8(10):e78572. doi: 10.1371/journal.pone.0078572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Stevens, K. A. & Parrish, M. J. Neck posture, dentition, and feeding strategies in Jurassic sauropod dinosaurs. In Thunder-lizards: TheSauropodomorphDinosaurs (eds. Tidwell, V. & Carpenter, K.) 212–232 (Indiana University Press, 2005).
- 16.Martin, J. Mobility and feeding of Cetiosaurus (Saurischia, Sauropoda): why the long neck? In 4th Symp. MesozoicTerrestrialEcosystems (eds. Currie, P. J. & Koster, E. H.) 154–159 (1987).
- 17.Martin J, Martin-Rolland V, Frey E. Not cranes or masts, but beams: the biomechanics of sauropod necks. Oryctos. 1998;1:113–120. [Google Scholar]
- 18.Dzemski G, Christian A. Flexibility along the neck of the ostrich (Struthio camelus) and consequences for the reconstruction of dinosaurs with extreme neck length. J. Morphol. 2007;268:701–714. doi: 10.1002/jmor.10542. [DOI] [PubMed] [Google Scholar]
- 19.Taylor MP. Almost all known sauropod necks are incomplete and distorted. PeerJ PrePrints. 2015;3:e1418v1. doi: 10.7287/peerj.preprints.1418v1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schwarz D, Frey E, Meyer CA. Novel reconstruction of the orientation of the pectoral girdle in sauropods. Anat. Rec. 2007;290:32–47. doi: 10.1002/ar.20405. [DOI] [PubMed] [Google Scholar]
- 21.Remes K, et al. A new basal sauropod dinosaur from the middle Jurassic of Niger and the early evolution of sauropoda. PLoS One. 2009;4:e6924. doi: 10.1371/journal.pone.0006924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mocho, P., Ortega, F., Aberasturi, A. & Escaso, F. Spinophorosaurus (Sauropoda), a new look inside eusauropod evolution. In VI Jornadas Internacionales sobre Paleontologia de dinosaurios Abstract Book 89–90 (2013).
- 23.Mallison H. The Digital Plateosaurus I: Body Mass, Mass Distribution and Posture Assessed Using Cad and Cae on a Digitally Mounted Complete Skeleton. Palaeontol. Electron. 2010;2:1–26. [Google Scholar]
- 24.Rasskin-Gutman, D. & Buscalioni, A. D. Theoretical morphology of the Archosaur (Reptilia: Diapsida) pelvic girdle. Paleobiology27, 59–78, http://dx.doi.org/10.1666/0094-8373(2001)027<0059:TMOTAR>2.0.CO;2 (2001).
- 25.Knoll F, Witmer LM, Ortega F, Ridgely RC, Schwarz-Wings D. The braincase of the basal sauropod dinosaur Spinophorosaurus and 3D reconstructions of the cranial endocast and inner ear. PLoS One. 2012;7:e30060. doi: 10.1371/journal.pone.0030060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gunji M, Endo H. Functional cervicothoracic boundary modified by anatomical shifts in the neck of giraffes. R. Soc. Open Sci. 2016;3:150604. doi: 10.1098/rsos.150604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gilmore CW. On a newly mounted skeleton of Diplodocus in the United States National Museum. Proc. United States Natl. Museum. 1932;81:1–21. doi: 10.5479/si.00963801.81-2941.1. [DOI] [Google Scholar]
- 28.Coombs WP. Sauropod habits and habitats. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1975;17:1–33. doi: 10.1016/0031-0182(75)90027-9. [DOI] [Google Scholar]
- 29.Bakker RT. The Superiority of Dinosaurs. Discovery. 1968;3:11–22. [Google Scholar]
- 30.Tschopp E, Mateus O. Clavicles, interclavicles, gastralia, and sternal ribs in sauropod dinosaurs: New reports from Diplodocidae and their morphological, functional and evolutionary implications. J. Anat. 2013;222:321–340. doi: 10.1111/joa.12012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Remes, K. Evolution of the Pectoral girdle and Forelimb in Sauropodomorpha (Dinosauria, Saurischia): Osteology, Myology and Fuction. Evolution (Ludwig-Maximilians-Universität München, 2007).
- 32.Hohn-Schulte, B. Form and Function of the Shoulder Girdle in Sauropod Dinosaurs: A Biomechanical Investigation with the Aid of Finite Elements. (Ruhr- Universität Bochum, 2010).
- 33.Paul GS. Restoring Maximum Vertical Browsing Reach in Sauropod Dinosaurs. Anat. Rec. 2017;300:1802–1825. doi: 10.1002/ar.23617. [DOI] [PubMed] [Google Scholar]
- 34.Monbaron M, Russell DA, Taquet P. Atlasaurus imelakei n.g., n.sp., a brachiosaurid-like sauropod from the Middle Jurassic of Morocco. Comptes Rendus de l’Académie des Sciences - Series IIA - Earth and Planetary Science. 1999;329:519–526. [Google Scholar]
- 35.Janensch, W. Die Skelettrekonstruktion von Brachiosaurus brancai. Palaeontographica Supplement, 95–103 (1950).
- 36.Young TP, Isbell LA. Sex Differences in Giraffe Feeding Ecology: Energetic and Social Constraints. Ethology. 1991;87:79–89. doi: 10.1111/j.1439-0310.1991.tb01190.x. [DOI] [Google Scholar]
- 37.McIntosh, J. S. Sauropoda. In The Dinosauria (eds. Weishampel, D. B., Dodson, P. & Osmólska, H.) 345–401 (University of California Press, 1990).
- 38.Upchurch, P., Barrett, P. M. & Dodson, P. Sauropoda. In The Dinosauria, 2nd edition (eds. Weishampel, D. B., Dodson, P. & Osmólska, H.) 259–322 (University of California Press, 2004).
- 39.Yadagiri, P. The osteology of Kotasaurus yamanpalliensis, a sauropod dinosaur from the Early Jurassic Kota Formation of India. J. Vertebr. Paleontol. 21, 242–252, https://doi.org/10.1671/0272-4634(2001)021[0242:TOOKYA]2.0.CO;2 (2001).
- 40.Jain, S. L., Kutty, T. S., RoyChowdhury, T. K. R. & Chatterjee, S. Some characteristics of Barapasaurus tagorei, a sauropod dinosaur from the Lower Jurassic of Deccan, India. In Forth International Gondwana Symposium: Culcutta, India, 1977 - papers (Vol 1) 204–216 (1979).
- 41.Bonnan MF, Yates AM. A new description of the forelimb of the basal sauropodomorph Melanorosaurus: Implications for the evolution of pronation, manus shape and quadrupedalism in sauropod dinosaurs. Evol. Palaeobiology. 2007;77:157–168. [Google Scholar]
- 42.Zhang, Y. The Middle Jurassic dinosaur fauna from Dashanpu, Zigong, Sichuan Vol I. Sauropod dinosaurs (1). Shunosaurus. Sichuan Publ. House Sci. Technol. 1–89 (1988).
- 43.Taylor MP, Wedel MJ. Why sauropods had long necks; and why giraffes have short necks. PeerJ. 2013;1:e36. doi: 10.7717/peerj.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Carabajal AP, Carballido JL, Currie PJ. Braincase, neuroanatomy, and neck posture of Amargasaurus cazaui (Sauropoda, Dicraeosauridae) and its implications for understanding head posture in sauropods. J. Vertebr. Paleontol. 2014;34(4):870–882. doi: 10.1080/02724634.2014.838174. [DOI] [Google Scholar]
- 45.Whitlock JA. Inferences of diplodocoid (Sauropoda: Dinosauria) feeding behavior from snout shape and microwear analyses. PLoS One. 2011;6(4):e18304. doi: 10.1371/journal.pone.0018304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wilson JA. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zool. J. Linn. Soc. 2002;136:215–275. doi: 10.1046/j.1096-3642.2002.00029.x. [DOI] [Google Scholar]
- 47.Xu X, et al. A new Middle Jurassic diplodocoid suggests an earlier dispersal and diversification of sauropod dinosaurs. Nat. Commun. 2018;9(2700):1. doi: 10.1038/s41467-018-05128-. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bates KT, et al. Temporal and phylogenetic evolution of the sauropod dinosaur body plan. R. Soc. Open Sci. 2016;3:1–17. doi: 10.1098/rsos.150636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bonaparte JF. Les dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétiosauridés) du Jurassique moyen de Cerro. Condor (Chubut, Argentine). Ann. Paléontologie. 1986;72:325–386. [Google Scholar]
- 50.Mallison H, Wings O. Photogrammetry in paleontology - A practical guide. J. Paleontol. Tech. 2014;12:1–31. [Google Scholar]
- 51.Vidal D, Díez Díaz V. Reconstructing hypothetical sauropod tails by means of 3D digitization: Lirainosaurus astibiae as case study. J. Iber. Geol. 2017;43:293–305. doi: 10.1007/s41513-017-0022-6. [DOI] [Google Scholar]
- 52.Mallison H. The Digital Plateosaurus II: An Assessment of the Range of Motion of the Limbs and Vertebral Column and of Previous Reconstructions using a Digital Skeletal Mount. Acta Palaeontol. Pol. 2010;55:433–458. doi: 10.4202/app.2009.0075. [DOI] [Google Scholar]
- 53.Galton, P. M., Van Heerden, J. & Yates, A. M. Postcranial Anatomy of Referred Specimens of Melanorosaurus. In Thunder-lizards: The SauropodomorphDinosaurs (eds. Carpenter, K. & Tidwell, V.) 1–37 (Indiana University Press, 2005).
- 54.Janensch WDieWirbelsäule. der Gattung Dicraeosaurus hausemanni. Palaeontographica. 1929;3:39–133. [Google Scholar]
- 55.Janensch W. Die Gliedmaszen und Gliedmaszengürtel der Sauropoden der Tendaguru- Schichten. Palaeontographica. 1961;3:177–235. [Google Scholar]
- 56.Wiman C. Die Kreide-Dinosaurier aus Shantung. Palaeontol. Sin. Ser. C. 1929;6:1–67. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Most fossil material for the holotype specimen of Spinophorosaurus nigerensis (GCP-CV-4229) is temporally deposited at the Museo Paleontológico de Elche (Elche, Spain). The paratype specimen (NMB-1699-R) and part of the holotype (NMB-1698-R) of Spinophorosaurus nigerensis are temporally deposited at the Staatliches Naturhistorisches Museum (Braunchsweig, Germany). Both specimens will eventually be returned to the Musée National Boubou Hama (Niamey, Niger). The digital fossils used to build the virtual skeleton are deposited and accessioned at the Museo Paleontológico de Elche.