

Highlights
-
•
Three classes of fusion glycoproteins have been hitherto identified.
-
•
Structures of their pre-fusion and post-fusion states have revealed a huge conformational change.
-
•
Intermediate structures during the structural transition have been recently characterized.
-
•
Published data are consistent with the existence of a ‘pre-hairpin’ intermediate.
Abstract
Entry of enveloped viruses into cells requires the fusion of viral and cellular membranes, driven by conformational changes in viral glycoproteins. Three different classes of viral fusion proteins have been hitherto identified based on common structural elements. Crystal structures have provided static pictures of pre-fusion and post-fusion conformations of these proteins and have revealed the dramatic reorganization of the molecules, but the transition pathway remains elusive. In this review, we will focus on recent data aiming to characterize intermediate structures during the conformational change. All these data support the existence of a pre-hairpin intermediate, but its oligomeric status is still a matter of debate.
Current Opinion in Virology 2013, 3:143–150
This review comes from a themed issue on Virus structure and function
Edited by Joseph Marcotrigiano and Felix Rey
For a complete overview see the Issue and the Editorial
Available online 3rd April 2013
1879-6257/$ – see front matter, © 2013 Elsevier B.V. All rights reserved.
Introduction
Entry of enveloped viruses into host cells requires fusion of the viral envelope with a cellular membrane. Fusion is mediated by viral fusogenic glycoproteins. Activation of their fusion capacity involves large structural rearrangements of these glycoproteins upon interaction with specific triggers (e.g. low pH, cellular receptors, etc.). These conformational changes result in the exposure of hydrophobic motifs (the so-called fusion peptides or fusion loops), which then interact with one or both of the participating membranes resulting in their destabilization and their merger [1]. Triggering of the conformational change in the absence of a target membrane leads to inactivation of the fusion properties of the fusogenic glycoprotein.
Structural studies have hitherto defined three classes of viral fusion glycoproteins [2]. Although the structure of these glycoproteins is quite different from one class to another, the membrane fusion pathway seems to be very similar for all the enveloped viruses studied so far [3, 4, 5]. It is generally assumed that fusion proceeds via the formation of an initial stalk that is a local lipidic connection between the outer leaflets of the fusing membranes. Radial expansion of the stalk induces the formation of a transient hemifusion diaphragm (i.e. a local bilayer made by the two initial inner leaflets). The next step is the formation of a pore in the fusion diaphragm. The pore's enlargement, which is the most energetically expensive step, leads to complete fusion [6].
Crystal structures of fusion glycoprotein ectodomains have hitherto provided static pictures of pre-fusion and post-fusion conformations, but the transition pathway still remains elusive. In this review, we will focus on recently published data concerning the structure of intermediate conformations during the structural transition.
Three structural classes of fusion glycoproteins
Three different classes of viral fusion proteins have been identified to date based on their common structural elements [2] (Figure 1 ). Class I fusion proteins are characterized by their post-fusion structure, which could be described as a trimer of hairpins [7]. The best-characterized members of this class are the influenza virus hemagglutinin (HA) [8, 9] and the fusion protein (F) of paramyxoviruses [10, 11], but this class also includes fusion proteins from retroviruses, coronaviruses and filoviruses [7]. Both the pre-fusion and post-fusion structures of class I fusion glycoproteins are trimeric. In most of the cases, the subunits constituting the trimer result from the proteolytic cleavage of a precursor into two fragments. The resulting C-terminal fragment, which is anchored in the viral membrane by a hydrophobic transmembrane (TM) domain, bears at or near its amino-terminal end a hydrophobic fusion peptide, buried at a protein–protein interface in the ectodomain of the pre-fusion state. Immediately downstream the fusion peptide, there is a first heptad repeat region (called HRA or HR1). A second one (called HRB or HR2) is located upstream the TM domain. In the post-fusion conformation, the HRA region forms a central trimeric coiled coil against which are packed, in an antiparallel manner, the HRB segments abutting the carboxy-terminal TM domains. Thus, in the post-fusion conformation, the protomer shape is an elongated hairpin-like structure with the fusion peptide and the TM domain located at the same end, as expected at the end of the fusion process. Except for this common central structural motif, the structures of class I fusion glycoproteins are very different from each other. Whether there is a common origin to all these proteins or if they represent convergent solutions to developing a membrane fusion machinery is still an open question.
Figure 1.
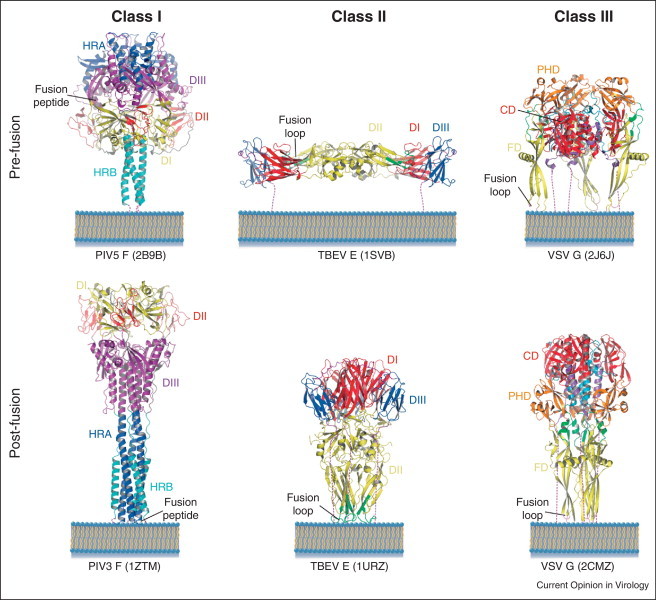
Ribbon illustrations of representative members of the three classes of viral fusion glycoproteins. Domains for the paramyxovirus (class I) and flavivirus (class II) proteins are colored and named according to [10, 11] and [12], respectively. Domains for the rhabdovirus (class III) protein are colored and named according to [21] with the exception that the central domain (CD) that was initially ascribed to two domains (DI and DII) is now considered as a single domain and corresponds to the previously described rigid block that remains invariant during the transition [21]. The dash lines represent the segments at the C-termini of the ectodomains that connect them to the TM domain (not represented). Respective pdb codes of each protein are showed in parentheses.
Among class II fusion proteins are the E protein of flaviviruses [12, 13] and E1 of alphaviruses [14, 15]. These proteins have a common origin and display a molecular architecture completely different from that of class I proteins [16]. They are synthesized and folded as a complex with a second viral envelope protein that acts as a chaperone. Proteolytic cleavage of the chaperone primes the fusion protein to trigger membrane merger at low pH. In their native conformation, they form homodimers (flaviviruses) or heterodimers (alphaviruses) that are organized in an icosahedral assembly. Their ectodomain is made of three domains made mostly of β-sheets (DI: central domain (CD); DII: fusion domain (FD); DIII: lateral, C-terminal domain) and lie flat or nearly flat at the viral surface. The fusion loop, located internally between two β-strands of domain II, is buried at a dimer interface. Considered in a single protomer, the conformational change is much less impressive than that observed for HA or for paramyxovirus F protein, since there is essentially no change in the secondary structure. Nevertheless, the post-fusion conformation is trimeric as for class I proteins and the shape of the pre-fusion trimer and interactions between its protomers are completely different than in the pre-fusion shell. Importantly, interactions between domains in a single protomer are also markedly different between the pre-fusion and post-fusion conformations. In particular, domain III, which is connected to the C-terminal part of the molecule, moves by about 35 Å (and also rotates by 65° for the flavivirus E), moving toward the fusion loop in the post-fusion trimer. The movement of domain III redirects the polypeptide chain, so that in the post-fusion trimeric form the TM segment is located near the fusion loop region. Therefore, here again, each protomer constituting the trimer has a hairpin structure.
The third class of fusion proteins was identified when the structures of the ectodomains of the fusogenic glycoproteins G [17] of vesicular stomatitis virus (VSV) and B (gB) of herpes simplex virus 1 (HSV-1) [18] were determined. Their comparison revealed an unanticipated homology between the two proteins, for which no sequence similarity had previously been detected. Epstein–Barr Virus (EBV) gB [19] and baculovirus gp64 [20], the structures of which were subsequently determined, have also been demonstrated to belong to this class.
VSV G is the only class III fusion protein for which the X-ray structures of both the pre-fusion [21] and post-fusion [17] states have been determined. Both states are trimeric and the polypeptide chain of G ectodomain folds into three distinct domains: the FD is inserted in a loop of a pleckstrin homology domain (PHD) that is itself inserted in a CD whose long helix is involved in trimerization of the molecule in both states. The CD also comprises a β-sheet rich region connecting to the C-terminal segment of the ectodomain. This segment itself connects to the TM domain in the intact glycoprotein. It is shorter in the VSV G post-fusion ectodomain crystal structure than in the ectodomains of gB [18] and gp64 [20], where the extra region forms an α helical structure that inserts between the other two protomers within the trimer. The organization of the class III FD is very similar to that of class II fusion proteins, although the class II and class III proteins are not homologous and the FDs’ topologies are inconsistent. Detailed points of convergence include an extended β-sheet structure terminating in a three-stranded sheet at the tip of which is located the membrane-interacting motif of the ectodomain. In VSV G both of the two loops at the tip of FD are fusion loops. In the pre-fusion structure, in contrast to class I and class II fusion proteins, the fusion loops are not buried at an oligomeric interface but point toward the viral membrane.
As for the other classes, the conformational change of VSV G from pre-fusion to post-fusion state involves a huge reorganization of the glycoprotein [21]. During this low pH-induced structural transition, both relocation of domains and major secondary structure modifications occur. The three domains retain their tertiary structure but undergo large rearrangements in their relative orientation. This is driven by refolding of large hinge regions between domains, the change in the two segments connecting CD to PHD being particularly dramatic and leading to lengthening of the CD central helix. Similarly, the C-terminal segment of the ectodomain both completely relocates and refolds from a series of short β-strands, helices and loops buried in part in the trimeric interface to a single α-helix on the surface of the trimeric core. This conformational change exhibits striking overall similarities to that of class I proteins such as paramyxovirus fusion protein F and influenza HA [22]. As for these proteins, the pre-fusion and post-fusion states of the G protomer are related by flipping both the FD and the C-terminal segment relative to CD. During this change, both the fusion loops and the TM domain move from one end of the molecule to the other. In the trimeric post-fusion state, a central core is made by the three CD central helices, in the grooves of which the three C-terminal helices position themselves in an antiparallel manner to form a six-helix bundle. The resulting overall organization of the post-fusion state is reminiscent of that of class I fusion glycoproteins.
Thus, even though the structures of fusion proteins from the three different classes are unrelated, the mechanisms for refolding share two key common features. First, the fusion peptides/loops are exposed and projected toward the end of the glycoprotein distal from the viral membrane. Second, the folding-back of the C-terminal region onto a trimeric N-terminal region leads to the formation of a post-fusion protein structure with the outer regions zipped up against the inner trimeric core.
Structural intermediates during the conformational changes
Using hydrophobic photolabeling, for both rhabdoviruses and influenza virus, it has been demonstrated that one of the earliest stages of the membrane fusion process is the interaction of the fusion peptide (for influenza HA) [23, 24] or the fusion loops (for rhabdovirus G) [25] with the target membrane. This indicates that an intermediate state during the structural transition exposes the fusion peptides that become accessible and are orientated toward the target membrane.
For class I proteins, it is assumed that the intermediate state interacting with the target membrane is in an extended conformation, sometimes called pre-hairpin, in which the central trimeric coiled coil (made of HR1/HRA segments) is already formed but the relocation of HR2/HRB segments against this central core has not occurred. Several lines of evidence support this model. First, for both paramyxoviruses [26] and retroviruses [27, 28], peptides corresponding to the HR2 segments can inhibit the fusion process (Figure 2 ). Second, it has been shown that when paramyxoviruses are trapped in the process of fusing with solid-supported bilayers, the distance between the viral membrane and the supported bilayer is precisely that predicted from a pre-hairpin model [29•]. Third, cryo-electron tomography has been recently performed on avian sarcoma/leucosis virus (ASLV, a retrovirus) incubated with liposomes, in the presence of a soluble form of the receptor in order to trigger the conformational change of the spikes. Subtomogram averaging revealed that an oblate globular domain is connected to both the target and the viral membrane by 2.5-nm-long stalks consistent with the existence of an elongated pre-hairpin conformation [30•].
Figure 2.
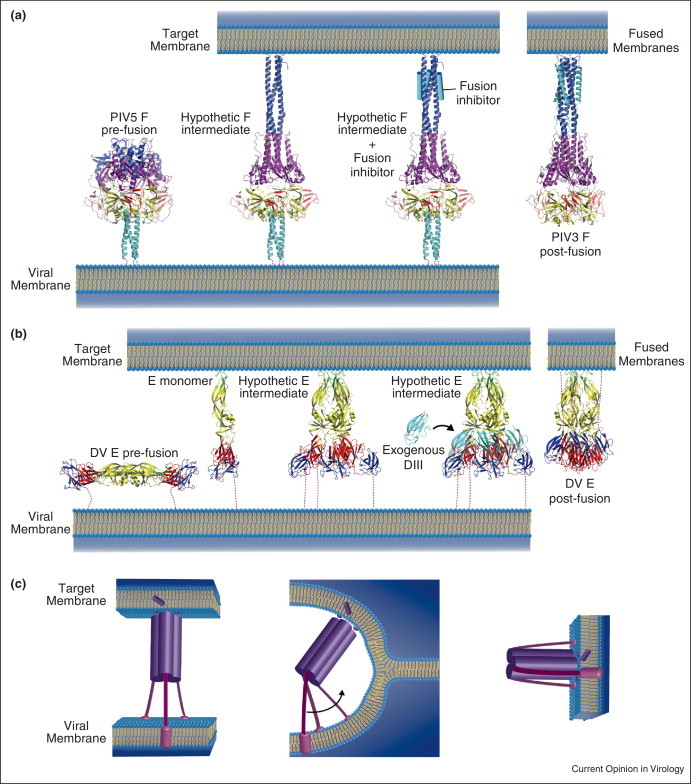
Models of viral fusion protein intermediates for class I (a) and class II (b) glycoproteins according to [29•, 34]. (a) In the case of paramyxovirus F glycoprotein, a trimeric pre-hairpin intermediate has been suggested to interact with the target membrane. This intermediate could be trapped by using a peptide derived from HRB (cyan cylinder), which inhibits fusion. It is believed that the interaction between the inhibitor and HRA (dark blue domain) mimics the six-helix bundle observed in the post-fusion state. (b) For class II fusion glycoproteins (here represented by E from Dengue virus), upon lowering the pH, the dimeric prefusion forms dissociates. Monomers interact with the target membrane and have been suggested to form trimeric pre-hairpin intermediates. In these intermediates, DIII have not folded back yet toward DI as in the E post-fusion form. The interaction of exogenous DIII with the trimeric intermediates prevents DIII foldback and blocks fusion. (c) Cartoon showing the complexity of the transition from the trimeric pre-hairpin intermediate to the trimeric post-fusion form. This transition requires a break of the threefold symmetry of the trimer (through bending of the molecule) together with a rotation of at least one of the C-terminal segments.
Class II fusion proteins are known to transit from a (homo- or hetero-) pre-fusion dimer to a post-fusion trimer through a monomeric intermediate [31, 32]. The questions for this class regard the oligomeric status of the glycoprotein when it interacts with the target membrane and the stage of the refolding process at which trimerization occurs. For tick-borne encephalitis virus, it has been suggested that the initial interaction with the target membrane involves the insertion of multiple copies of E monomers and that trimerization requires coupling to domain III folding-back [33]. However, it has been demonstrated that exogenous domain III can function as a dominant-negative inhibitor of alphavirus and flavivirus membrane fusion [34], which is consistent with the existence of a relatively long-lived core trimer intermediate with which domain III interacts. Since, for Semliki forest virus, DI/II trimers are stable in the absence of DIII, it has been proposed that DI/II trimer formation is relatively rapid, while the folding-back of domain III is a late and relatively slow step in the structural transition pathway [35, 36].
A complex final rearrangement
For class I and class II fusion glycoproteins, the transition from the trimeric extended intermediate (Figure 2) to the final trimeric post-fusion state cannot maintain strict threefold symmetry. This symmetry is disrupted by the refolding-back of the C-terminal portion of the molecule.
Furthermore, in a putative trimeric pre-hairpin intermediate conformation, the FDs are located outside the pyramidal volume defined by the viral membrane and the three connecting segments, whereas in the post-fusion state, they are located inside (Figure 2c). It is worth noting that if the central trimeric core does not dissociate during the structural transition from this putative intermediate to the final post-fusion state, then it is absolutely required that the TM segments are at least temporarily separated because they need to turn on themselves to achieve the transition.
VSV G structural transition proceeds through monomerization
Global refolding of VSV G from the pre-fusion to the post-fusion state exhibits striking similarities to that of class I fusion glycoproteins [22]. Recently, the low-pH-induced structural transition of a soluble form of VSV G ectodomain (the fragment that was previously crystallized) has been characterized by using an array of biophysical techniques [37•]. This has provided important insights into the conformation of intermediates during the transition. While the post-fusion trimer was the major species detected at low pH, the pre-fusion trimer was never observed in solution. Instead, at a slightly basic or neutral pH, the soluble ectodomain appeared to be a flexible monomer that populates a manifold of conformations in a pH-dependent way. The presence of monomers at the viral surface was also detected by electron microscopy. In fact, at pH 7.5, only a few pre-fusion trimers can be detected and most of the glycoproteins are likely to be in a monomeric state. At pH 6.7, rod-shaped elongated structures were observed, the dimensions of which were consistent with a monomeric state. Taken together, these data suggested that the conformational change proceeds through monomeric states of G [37•] (Figure 3 ). Consistent with this view are the large differences between the trimeric interfaces of the pre-fusion and post-fusion structure [21] as well as the fact that the VSV G trimers are unstable after detergent solubilization [38].
Figure 3.
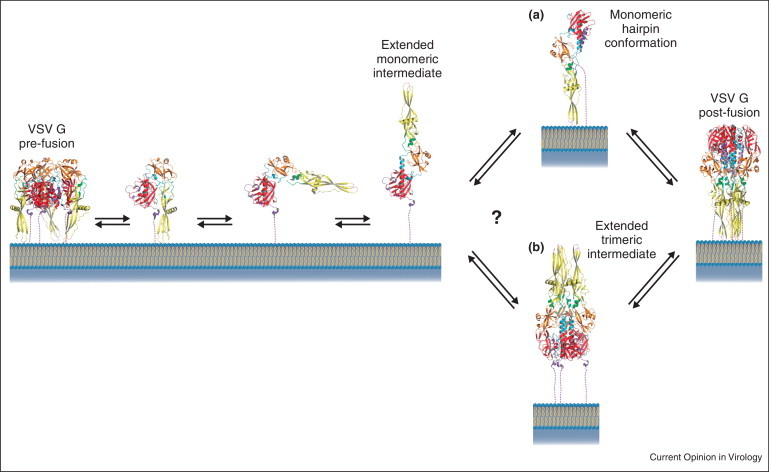
Plausible structural transition pathways of G at the viral surface. At pH 7.5, pre-fusion trimers and flexible monomers are in equilibrium at the viral surface. At pH 6.7, elongated monomeric structures are observed. Whether trimerization occurs after (pathway A) of before (pathway B) hairpin formation is not known for VSV G.
The stage at which the monomeric intermediates re-associate to form the final post-fusion form is not yet known (Figure 3). Therefore, whether an extended trimeric conformation exists for VSV G is still an open question.
However, monomerization poses a problem. How is it compatible with the cooperativity between glycoproteins, which seems to be required for membrane merger? First, several monomers could simultaneously bind to the target membrane through their fusion loops to account for the cooperativity required at the early stage of the fusion process (i.e. stalk and initial pore formation). Second, for VSV, post-fusion trimers located outside the contact zone between virion and cellular membranes can reorganize into regular arrays, the formation of which has been suggested to induce pore enlargement [39•]. This latter hypothesis is consistent with the model of Kozlov and Chernomordik [40], according to which fusion proteins outside the contact zone generate the driving force for pore enlargement by forming a coat around the fusion site. It is also in agreement with experimental data from the same group obtained on influenza, showing that HAs located outside the fusion site are involved in late stages of the fusion reaction [41].
Conclusions and perspectives
The structures of intermediate conformations of fusion proteins, how they cooperate and how they interact with and deform the fusing membranes are still very elusive. Electron microscopy and tomography have given some clues [29•, 30•, 37•, 42] but the resolution of the reconstructions is too low to unambiguously conclude on the nature of these intermediates. The only crystal structure of an intermediate has been obtained on an HA mutant with a mutation in HA2 FD that stabilizes the HA trimer in a prefusion-like state at and below fusogenic pH [43]. Only very subtle differences could be detected between this structure and HA prefusion structure indicating that, in the crystal, HA was trapped in a very early intermediate conformation. Gaining high-resolution structural information on intermediate conformations of fusion proteins (ideally complete with their TM domain and in association with bilayers) is now the new frontier for scientists working in the field to reach.
Conflict of interest
The authors declare no conflict of interest.
References and recommended reading
Papers of particular interest, published within the period of review, have been highlighted as:
• of special interest
Acknowledgements
This project was supported by CNRS and grants from ANR including post-doctoral fellowship to AA (ANR-08-BLAN-0256) and from the European Union (7th framework program, ITN ‘Virus entry’) including pre-doctoral fellowship to EB.
References
- 1.Albertini A., Bressanelli S., Lepault J., Gaudin Y. Structure and working of viral fusion machinery. In: Benos D.J., Balaban R., Simon S., editors. vol. 68. Elsevier Academic Press Inc.; San Diego, CA, USA: 2011. pp. 49–80. (Current Topics in Membranes). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Harrison S.C. Viral membrane fusion. Nat Struct Mol Biol. 2008;15:690–698. doi: 10.1038/nsmb.1456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chernomordik L.V., Frolov V.A., Leikina E., Bronk P., Zimmerberg J. The pathway of membrane fusion catalyzed by influenza hemagglutinin: restriction of lipids, hemifusion, and lipidic fusion pore formation. J Cell Biol. 1998;140:1369–1382. doi: 10.1083/jcb.140.6.1369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gaudin Y. Rabies virus-induced membrane fusion pathway. J Cell Biol. 2000;150:601–612. doi: 10.1083/jcb.150.3.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zaitseva E., Mittal A., Griffin D.E., Chernomordik L.V. Class II fusion protein of alphaviruses drives membrane fusion through the same pathway as class I proteins. J Cell Biol. 2005;169:167–177. doi: 10.1083/jcb.200412059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chernomordik L.V., Kozlov M.M. Mechanics of membrane fusion. Nat Struct Mol Biol. 2008;15:675–683. doi: 10.1038/nsmb.1455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lamb R.A., Jardetzky T.S. Structural basis of viral invasion: lessons from paramyxovirus F. Curr Opin Struct Biol. 2007;17:427–436. doi: 10.1016/j.sbi.2007.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wilson I.A., Skehel J.J., Wiley D.C. Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3 A resolution. Nature. 1981;289:366–373. doi: 10.1038/289366a0. [DOI] [PubMed] [Google Scholar]
- 9.Bullough P.A., Hughson F.M., Skehel J.J., Wiley D.C. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature. 1994;371:37–43. doi: 10.1038/371037a0. [DOI] [PubMed] [Google Scholar]
- 10.Yin H.S., Paterson R.G., Wen X., Lamb R.A., Jardetzky T.S. Structure of the uncleaved ectodomain of the paramyxovirus (hPIV3) fusion protein. Proc Natl Acad Sci U S A. 2005;102:9288–9293. doi: 10.1073/pnas.0503989102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yin H.S., Wen X., Paterson R.G., Lamb R.A., Jardetzky T.S. Structure of the parainfluenza virus 5 F protein in its metastable, prefusion conformation. Nature. 2006;439:38–44. doi: 10.1038/nature04322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rey F.A., Heinz F.X., Mandl C., Kunz C., Harrison S.C. The envelope glycoprotein from tick-borne encephalitis virus at 2 A resolution. Nature. 1995;375:291–298. doi: 10.1038/375291a0. [DOI] [PubMed] [Google Scholar]
- 13.Modis Y., Ogata S., Clements D., Harrison S.C. Structure of the dengue virus envelope protein after membrane fusion. Nature. 2004;427:313–319. doi: 10.1038/nature02165. [DOI] [PubMed] [Google Scholar]
- 14.Lescar J., Roussel A., Wien M.W., Navaza J., Fuller S.D., Wengler G., Wengler G., Rey F.A. The Fusion glycoprotein shell of Semliki Forest virus: an icosahedral assembly primed for fusogenic activation at endosomal pH. Cell. 2001;105:137–148. doi: 10.1016/s0092-8674(01)00303-8. [DOI] [PubMed] [Google Scholar]
- 15.Gibbons D.L., Vaney M.C., Roussel A., Vigouroux A., Reilly B., Lepault J., Kielian M., Rey F.A. Conformational change and protein–protein interactions of the fusion protein of Semliki Forest virus. Nature. 2004;427:320–325. doi: 10.1038/nature02239. [DOI] [PubMed] [Google Scholar]
- 16.Vaney M.C., Rey F.A. Class II enveloped viruses. Cell Microbiol. 2011;13:1451–1459. doi: 10.1111/j.1462-5822.2011.01653.x. [DOI] [PubMed] [Google Scholar]
- 17.Roche S., Bressanelli S., Rey F.A., Gaudin Y. Crystal structure of the low-pH form of the vesicular stomatitis virus glycoprotein G. Science. 2006;313:187–191. doi: 10.1126/science.1127683. [DOI] [PubMed] [Google Scholar]
- 18.Heldwein E.E., Lou H., Bender F.C., Cohen G.H., Eisenberg R.J., Harrison S.C. Crystal structure of glycoprotein B from herpes simplex virus 1. Science. 2006;313:217–220. doi: 10.1126/science.1126548. [DOI] [PubMed] [Google Scholar]
- 19.Backovic M., Longnecker R., Jardetzky T.S. Structure of a trimeric variant of the Epstein–Barr virus glycoprotein B. Proc Natl Acad Sci U S A. 2009;106:2880–2885. doi: 10.1073/pnas.0810530106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kadlec J., Loureiro S., Abrescia N.G., Stuart D.I., Jones I.M. The postfusion structure of baculovirus gp64 supports a unified view of viral fusion machines. Nat Struct Mol Biol. 2008;15:1024–1030. doi: 10.1038/nsmb.1484. [DOI] [PubMed] [Google Scholar]
- 21.Roche S., Rey F.A., Gaudin Y., Bressanelli S. Structure of the prefusion form of the vesicular stomatitis virus glycoprotein g. Science. 2007;315:843–848. doi: 10.1126/science.1135710. [DOI] [PubMed] [Google Scholar]
- 22.Roche S., Albertini A.A., Lepault J., Bressanelli S., Gaudin Y. Structures of vesicular stomatitis virus glycoprotein: membrane fusion revisited. Cell Mol Life Sci. 2008;65:1716–1728. doi: 10.1007/s00018-008-7534-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Stegmann T., Delfino J.M., Richards F.M., Helenius A. The HA2 subunit of influenza hemagglutinin inserts into the target membrane prior to fusion. J Biol Chem. 1991;266:18404–18410. [PubMed] [Google Scholar]
- 24.Tsurudome M., Gluck R., Graf R., Falchetto R., Schaller U., Brunner J. Lipid interactions of the hemagglutinin HA2 NH2-terminal segment during influenza virus-induced membrane fusion. J Biol Chem. 1992;267:20225–20232. [PubMed] [Google Scholar]
- 25.Durrer P., Gaudin Y., Ruigrok R.W., Graf R., Brunner J. Photolabeling identifies a putative fusion domain in the envelope glycoprotein of rabies and vesicular stomatitis viruses. J Biol Chem. 1995;270:17575–17581. doi: 10.1074/jbc.270.29.17575. [DOI] [PubMed] [Google Scholar]
- 26.Russell C.J., Jardetzky T.S., Lamb R.A. Membrane fusion machines of paramyxoviruses: capture of intermediates of fusion. EMBO J. 2001;20:4024–4034. doi: 10.1093/emboj/20.15.4024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wild C.T., Shugars D.C., Greenwell T.K., McDanal C.B., Matthews T.J. Peptides corresponding to a predictive alpha-helical domain of human immunodeficiency virus type 1 gp41 are potent inhibitors of virus infection. Proc Natl Acad Sci U S A. 1994;91:9770–9774. doi: 10.1073/pnas.91.21.9770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Furuta R.A., Wild C.T., Weng Y., Weiss C.D. Capture of an early fusion-active conformation of HIV-1 gp41. Nat Struct Biol. 1998;5:276–279. doi: 10.1038/nsb0498-276. [DOI] [PubMed] [Google Scholar]
- 29•.Kim Y.H., Donald J.E., Grigoryan G., Leser G.P., Fadeev A.Y., Lamb R.A., DeGrado W.F. Capture and imaging of a prehairpin fusion intermediate of the paramyxovirus PIV5. Proc Natl Acad Sci U S A. 2011;108:20992–20997. doi: 10.1073/pnas.1116034108. [DOI] [PMC free article] [PubMed] [Google Scholar]; The authors have measured the interbilayer spacing of a paramyxovirus trapped in the process of fusing with solid-supported bilayers. The interbilayer spacing is precisely that predicted from a computational model of the pre-hairpin intermediate, providing strong evidence for its structure and functional role.
- 30•.Cardone G., Brecher M., Fontana J., Winkler D.C., Butan C., White J.M., Steven A.C. Visualization of the two-step fusion process of the retrovirus avian sarcoma/leukosis virus by cryo-electron tomography. J Virol. 2012;86:12129–12137. doi: 10.1128/JVI.01880-12. [DOI] [PMC free article] [PubMed] [Google Scholar]; Cryo-electron tomography was used to study virion/receptor/liposome complexes that simulate the interactions of avian sarcoma/leukosis virus with cells. Subtomogram averaging revealed oblate globular domains (presumably the receptor-binding domain) connected to both the target and the viral membrane by 2.5-nm-long stalks. The authors propose that this conformation of the viral glycoprotein corresponds to the prehairpin intermediate.
- 31.Wahlberg J.M., Garoff H. Membrane fusion process of Semliki Forest virus. I: Low pH-induced rearrangement in spike protein quaternary structure precedes virus penetration into cells. J Cell Biol. 1992;116:339–348. doi: 10.1083/jcb.116.2.339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Stiasny K., Allison S.L., Marchler-Bauer A., Kunz C., Heinz F.X. Structural requirements for low-pH-induced rearrangements in the envelope glycoprotein of tick-borne encephalitis virus. J Virol. 1996;70:8142–8147. doi: 10.1128/jvi.70.11.8142-8147.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Stiasny K., Kossl C., Lepault J., Rey F.A., Heinz F.X. Characterization of a structural intermediate of flavivirus membrane fusion. PLoS Pathog. 2007;3:e20. doi: 10.1371/journal.ppat.0030020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liao M., Kielian M. Domain III from class II fusion proteins functions as a dominant-negative inhibitor of virus membrane fusion. J Cell Biol. 2005;171:111–120. doi: 10.1083/jcb.200507075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sanchez-San Martin C., Sosa H., Kielian M. A stable prefusion intermediate of the alphavirus fusion protein reveals critical features of class II membrane fusion. Cell Host Microbe. 2008;4:600–608. doi: 10.1016/j.chom.2008.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Roman-Sosa G., Kielian M. The interaction of alphavirus E1 protein with exogenous domain III defines stages in virus–membrane fusion. J Virol. 2011;85:12271–12279. doi: 10.1128/JVI.05902-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37•.Albertini A.A., Merigoux C., Libersou S., Madiona K., Bressanelli S., Roche S., Lepault J., Melki R., Vachette P., Gaudin Y. Characterization of monomeric intermediates during VSV glycoprotein structural transition. PLoS Pathog. 2012;8:e1002556. doi: 10.1371/journal.ppat.1002556. [DOI] [PMC free article] [PubMed] [Google Scholar]; By several biophysical techniques, it is shown that a soluble form of VSV G ectodomain is a flexible monomer that explores a large conformational space. The monomeric population exhibits a marked pH-dependence and adopts more elongated conformations when pH decreases. Solution studies are complemented by electron micrographs of negatively stained viral particles in which monomeric ectodomains of G are observed at the viral surface at both pH 7.5 and pH 6.7. The authors propose that the monomers are intermediates during the conformational change and thus that VSV G trimers dissociate at the viral surface during the structural transition.
- 38.Doms R.W., Keller D.S., Helenius A., Balch W.E. Role for adenosine triphosphate in regulating the assembly and transport of vesicular stomatitis virus G protein trimers. J Cell Biol. 1987;105:1957–1969. doi: 10.1083/jcb.105.5.1957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39•.Libersou S., Albertini A.A., Ouldali M., Maury V., Maheu C., Raux H., de Haas F., Roche S., Gaudin Y., Lepault J. Distinct structural rearrangements of the VSV glycoprotein drive membrane fusion. J Cell Biol. 2010;191:199–210. doi: 10.1083/jcb.201006116. [DOI] [PMC free article] [PubMed] [Google Scholar]; Individual fusion events between VSV and liposomes were visualized by electron microscopy. Fusion appears to be driven by two successive structural rearrangements of G at different sites on the virion. Rhabdoviruses are bullet-shaped and fusion is initiated at the flat base of the particle. Glycoproteins located outside the contact zone between virions and liposomes then reorganize into regular arrays. The authors suggest that the formation of these arrays, which is demonstrated to be an intrinsic property of the G ectodomain, induces membrane constraints, completing the fusion reaction.
- 40.Kozlov M.M., Chernomordik L.V. The protein coat in membrane fusion: lessons from fission. Traffic. 2002;3:256–267. doi: 10.1034/j.1600-0854.2002.030403.x. [DOI] [PubMed] [Google Scholar]
- 41.Leikina E., Mittal A., Cho M.S., Melikov K., Kozlov M.M., Chernomordik L.V. Influenza hemagglutinins outside of the contact zone are necessary for fusion pore expansion. J Biol Chem. 2004;279:26526–26532. doi: 10.1074/jbc.M401883200. [DOI] [PubMed] [Google Scholar]
- 42.Fontana J., Cardone G., Heymann J.B., Winkler D.C., Steven A.C. Structural changes in Influenza virus at low pH characterized by cryo-electron tomography. J Virol. 2012;86:2919–2929. doi: 10.1128/JVI.06698-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Xu R., Wilson I.A. Structural characterization of an early fusion intermediate of influenza virus hemagglutinin. J Virol. 2011;85:5172–5182. doi: 10.1128/JVI.02430-10. [DOI] [PMC free article] [PubMed] [Google Scholar]