

Abstract
Viral diseases of shrimp have caused negative effects on the economy in several countries in Asia, South America and America, where they have numerous shrimp culture industries. The studies on the immunity of shrimp and other crustaceans have mainly focused on general aspects of immunity and as a consequence little is known about the antiviral responses in crustaceans. The aim of this review is to update recent knowledge of innate immunity against viral infections in crustaceans. Several antiviral molecules have been isolated and characterized recently from decapods. Characterization and identification of these molecules might provide a promising strategy for protection and treatment of these viral diseases. In addition dsRNA-induced antiviral immunity is also included.
Keywords: Antiviral immunity, Crustacean, WSSV, YHV, TSV
1. Crustacean viruses
Viruses are the most common pathogens in the sea and they are present up to ten billion per litre and several of them can infect many organisms [1]. Hence, viral infections are common diseases in crustaceans such as penaeid shrimp, which can be infected by more than twenty different viruses [2]. In crustaceans, research on viruses has increased considerably since the first report on the occurrence of a virus in the crab Macropipus depurator [3]. Crustacean viruses belong to or are related to various viral families like the Baculoviridae, Bunyaviridae, Herpesviridae, Picornaviridae, Parvoviridae, Reoviridae, Rhabdoviridae, Togaviridae, Iridoviridae or a new virus family, the Nimaviridae [4]. During the past decades, both extensive and intensive shrimp cultures have been established and as a result more detailed knowledge of viral-host interactions has been gained. Among these viruses, the most intensively studied viruses have been characterized from cultured penaeids such as the white spot syndrome virus (WSSV), yellow head virus (YHV), and Taura syndrome virus (TSV). Much less details are available for other viruses present in wild crustaceans [5]. Infection with these viruses has caused detrimental diseases in shrimp culturing, resulting in serious economic losses. WSSV, YHV, and TSV have been regarded as the most serious shrimp viruses (Fig. 1 ) [6].
Fig. 1.

Here is an intensive shrimp culture farm in Thailand. The main viral diseases in shrimp culture are caused by white spot syndrome virus (WSSV), yellow head virus (YHV) and Taura syndrome virus (TSV). Of these white spot disease is one of the most serious disease. It reduces shrimp production and caused serious economic losses worldwide. Mortality is very high and can reach 100% within 3–10 days from the onset of diseases.
WSSV is an enveloped large circular double stranded DNA virus containing about 300 Kbp [7], [8]. The enveloped virions (120–150 nm × 270–290 nm) are symmetrical and ellipsoid to bacilliform in shape similar to baculovirus [9]. Phylogenetic analysis suggests that WSSV is a new virus family named as the Nimaviridae, genus Whispovirus. WSSV has been widely studied and here we mainly focus on the interaction between this virus and its decapod hosts. The WSSV has a wide host range including all species of shrimps, some other crustaceans (e.g. crayfish, crab, and spiny lobster), and has even been detected in insects (Fig. 2 ) [10] and can be transmitted from broodstock to the offspring [11]. After infection, the shrimp exhibits very high mortality rates and the cumulative mortality usually reaches 100% within 3–10 days after the onset of the first visible gross signs. Histopathological studies have shown that WSSV targets many different tissues in the freshwater crayfish, Pacifastacus leniusculus (Fig. 3 ) [12].
Fig. 2.
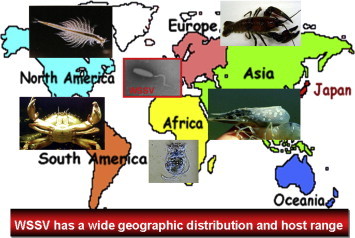
WSSV spreads to all countries which have shrimp farming from Asia to North and South America. This virus not only infects shrimp but also other crustaceans such as crayfish, crab, artemia and recently it was reported to infect rotifer in a shrimp pond.
Fig. 3.
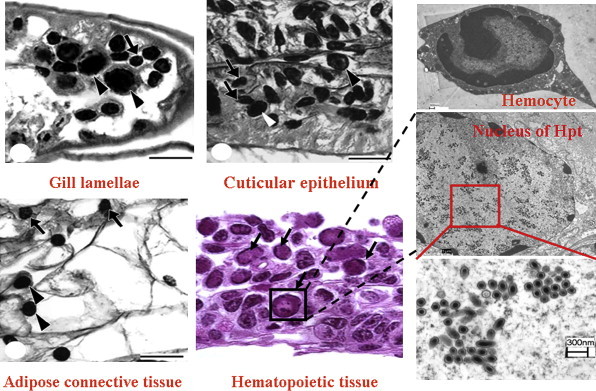
Target tissues of WSSV infection. Histopathological changes were found in various tissues of infected crayfish. For example gill, cuticular epidermis under shell and around the stomach, adipose connective tissue, hematopoietic tissue even hemocytes. The nuclei of infected cells are hypertrophied and the chromatin is marginated along the nuclear border (arrows). The arrow heads in the gill lamellae show infected hemocytes. By TEM observation inside these enlarged nuclei contains a massive of virus particles.
YHV is an enveloped viral particle (40–60 nm × 150–200 nm) with rounded ends containing a sense single-stranded RNA (approximately 26 Kbp). This virus contains three major structural proteins (gp116, gp64 and p20) and belongs to the genus Okavirus, Roniviridae, Nidovirales [13], [14], [15], [16]. The shrimps usually die within a few days after infection with a widespread necrosis in the lymphoid organ (LO), gills, connective tissues, hemocytes, and hematopoietic organs [17]. The LO is probably the primary target of YHV based on the histopathological studies and a YHV receptor study [18], [19].
TSV was originally reported in 1992 [20] and the virus was first isolated and characterized by Bonami et al. [21]. The viral particle is non-enveloped, icosahedral in shape with a diameter of 31–32 nm, and replicates within the cytoplasm of host cells. It contains a linear positive-sense ssRNA genome of about 10, 205 nucleotides, and the capsid possesses three major (55, 40, and 24 kDa) and one minor (58 kDa) polypeptides. This virus has been classified as a member of Picornaviridae [22], [23]. TSV is the first characterized picorna-like virus infecting an invertebrate other than insects. The cuticular epithelium of the foregut, gills, appendages, hindgut, and general body cuticle has been described as the predominant targeted cell type by TSV [6].
Unlike vertebrate immunity which is composed of both innate and adaptive responses, a general opinion is still that invertebrates rely on multiple innate defence reactions to combat infections. These reactions include the protection by physical barriers together with local (e.g. epithelial immunity) and systemic immune responses. Two main components, the humoral and cellular systems, are involved in the systemic innate immune response in invertebrates, both of which are activated upon immune challenge. Several reviews have described the histopathology, diagnostic techniques, epidemiology, and genome organization of these important viruses [6], [24], [25], [26]. This review is mainly focused on recent antiviral immunity described on these three major viruses in crustaceans.
2. Virus entry
Knowledge of virus entry into permissive cells is mainly from vertebrates and both cellular and viral factors are recruited when virus enters into cells. Most cells are capable of endocytosis through several distinct mechanisms including clathrin-coated vesicles, phagocytosis, macropinocytosis and caveolae [27]. Several animal viruses take advantage of the cell endocytic machinery for a successful infection. In this process, viruses are taken up into the cytoplasm, where they produce dsRNA and initiate replication. The incoming virus particles are entering endosomal structures, lysozymes, the endoplasmic reticulum (ER), and occasionally the Golgi complex. The mildly acidic pH in the endosome will trigger penetration and uncoating. During this process, cytoskeleton and cellular proteins are often recruited by the invading viruses.
How invertebrate viruses enter host cells remains largely unknown. Coronaviruses have been suggested to use envelope proteins as recognition molecules for viral-host binding and cell entry [28]. This is also supported by an in vitro study showing that a polyclonal antibody against gp116 (YHV glycoprotein 116) inhibited YHV entry into LO cells, but not an antibody to gp64 (YHV glycoprotein 116) [29]. YRP65 has been identified as a YHV receptor in Penaeus monodon. Both antibodies against YRP65 and silencing of the YRP65 are able to specifically inhibit the entry of the YHV into the Oka or lymphoid organ cells [18]. PmRab7, a shrimp small GTPase protein binding directly to VP28 of WSSV, was suggested to be involved in WSSV infection [30]. Silencing of PmRab7 dramatically inhibited WSSV-VP28 RNA and protein expression. Future work is still necessary to elucidate if this PmRab7 localizes on the surface of the host cells and how this protein is involved in virus transport into the cells. Further, the silencing of PmRab7 also inhibited YHV replication in the YHV-infected shrimp. These data indicate that PmRab7 is a common cellular factor required for WSSV or YHV replication in shrimp [31]. However, in a latter study, some shrimp injected with dsRNA did not show any antiviral protection and some of the RT-PCR results of host gene/viral gene are not in agreement with each other in the quantitation of their transcripts. Thus, the data presented in this study may need more detailed investigations for a fully understanding of the mechanism by which PmRab7 interacts with WSSV. Besides, another Rab GTPase from Penaeus japonicus was reported to regulate shrimp hemocytic phagocytosis through a proposed protein complex consisting of the PjRab, beta-actin, tropomyosin, and an envelope protein VP466 of WSSV, indicating a possible role of phagocytosis involved in virus evasion in shrimp [32]. A VP28 of WSSV was suggested to bind to shrimp cells as an attachment protein and in this way could help the virus to enter into the cytoplasm [33], but this speculation needs to be documented by more experimental data.
WSSV exhibits higher tropism to semigranular cells (SGCs) than to granular cells (GCs) in both crayfish and shrimp [12], [34]. Both the percentage of degranulated cells and cell spreading in WSSV-infected crayfish were significantly lower than that of sham infected crayfish when hemocyte lysate supernatant (HLS) or phorbol 12-myristate 13-acetate (PMA) was used [35]. Peroxinectin, a cell adhesive and opsonic peroxidase, has been shown to mediate attachment and spreading of the hemocytes in crayfish. This molecule can also trigger degranulation by a regulated exocytosis [36], [37]. Binding of peroxinectin to the cell surface via an extracellular superoxide dismutase (SOD) may produce a signal into the cell through an integrin for further cellular responses [38]. The cellular responses have been suggested to be triggered via a pathway that includes protein kinase C (PKC) activation and elevated protein tyrosine phosphorylation of a cellular protein of ∼80 kDa [39]. Hemocyte lysate supernatant (HLS) containing peroxinectin treated GCs and SGCs from WSSV-infected crayfish resulted in three different cell-reactions: non-spread, spread, and degranulated cells. The non-spread cell groups from both GCs and SGCs exhibited more WSSV positive cells than the degranulated cells. Taken these data together, it is likely that the PKC pathway might be somehow affected by WSSV during its replication within the cells [35].
3. Viral infection and hematopoiesis in crustaceans
In crustaceans, the number of freely circulating hemocytes varies in response to environmental stress, endocrine activity during the moulting cycle, and infection. Exposure to non-self molecules can cause a dramatic drop in total hemocyte count and the animals may die of an infection that they otherwise usually resist [37], [40]. WSSV affects the hemocytes in crayfish P. leniusculus. For instance, infection of WSSV has a significant effect on the proportion of different hemocyte types in the animals. The number of GCs was significantly higher in WSSV-injected crayfish compared to control animals [12]. Studies from crayfish and shrimp suggest that SGCs are more susceptible to WSSV and the virus replicates more rapidly in SGCs than GCs resulting in the gradually decreased SGCs from blood circulation [34], [35]. Other studies showed that the total numbers of circulating hemocytes in shrimp were dramatically decreased after infection by WSSV [41], [42], [43]. On the other hand, immune-relevant molecules from crustacean hemocytes are essential in defence against invasive pathogens [37], [44]. The data from recent studies also demonstrate that silencing of immune-relevant genes like anti-lipopolysaccharide factor (ALF) from crayfish hemocytes led to rapid infection of WSSV [45]. Therefore, immediate regeneration of hemocytes is critical for the survival of the animals against pathogenic intruders and hence the hematopoiesis, a process in which mature hemocytes are produced and subsequently released from the hematopoietic tissue, is therefore an important process during an infection with a microorganism.
So far, little is known about hematopoiesis under a viral infection in crustaceans. A previous study showed that the hematopoietic tissue (Hpt) of crayfish is the organ in which proliferation of hemocytes occurs. Injection of a β-1,3-glucan caused a severe loss of hemocytes followed by a rapid recovery due to release from the Hpt organ [46]. This study also indicated that the hemocytes were synthesized and partly differentiated in the Hpt, but the final differentiation into functional hemocytes was not completed until they were released into the circulation. Hematopoiesis is tightly regulated by the lineage restricted activity of various factors that promote cell diversification [47], [48], [49]. The hematopoietic tissue has been studied in several crustaceans, but the mechanisms for the release of blood cells into circulation in crustaceans are still unclear. The identification of three different proteins/transcripts can be used as markers/indicators for hematopoietic cell proliferation and specific differentiation into SGCs or GCs. Thus, the study of hemocyte lineage marker proteins would be useful for future investigations of hematopoiesis under normal or microbial challenge conditions in crustaceans [50].
4. Antiviral immune responses
Viral components like genomic DNA and RNA or dsRNA generated in virally infected cells can be sensed by host pattern recognition receptors (PRRs). After recognition, PRRs trigger effective and appropriate antiviral responses, including production of various cytokines and induction of inflammatory and adaptive immune reactions [51]. The molecular mechanisms that underlie the majority of crustacean antiviral immune responses are still unknown and are only starting to be addressed. So far, few host cellular genes/proteins involved in TSV and YHV pathogenesis have been studied. An increasing number of immune responsive genes/proteins involved in WSSV pathogenesis have been described in shrimp and crayfish. Here we summarize the major progresses made in anti-WSSV response in shrimp, crayfish, and other crustaceans.
4.1. Antiviral-related proteins/genes in crustaceans
To find antiviral substances, the substances have been isolated from tissue extracts of shrimp, blue crab, and crayfish. These extracts can bind to various DNA and RNA viruses such as Sindbis virus, vaccinia virus, vesicular stomatitis virus, mengovirus, Banzi virus, and poliomyelitis virus. The mechanism of this inhibitory activity remains unclear [52]. Recently, many studies using different techniques have been carried out on host-WSSV interactions in crustaceans [45], [53], [54], [55], [56]. It has been reported that virus-inhibiting proteins could be produced and some genes were up-regulated upon viral infection in crustaceans [52], [57], [58], [59], [60], [61]. Genes induced by viral infections and genes whose expression are associated with the ability of shrimp to survive from viral infections have been reported but their importance to produce antiviral substances is little known (Table 1 ).
Table 1.
Proteins/genes involved in anti-WSSV responses or interacting with WSSV in crustaceans.
| Proteins/genes involved in anti-WSSV or interacting with WSSV | Species |
|---|---|
| Actin | Litopenaeus vannamei[75] |
| ALF | Pacifastacus leniusculus[45] |
| Beta-integrin | Marsupenaeus japonicus[69] |
| Calreticulin | Fenneropenaeus chinensis[80] |
| Caspase-3 like gene | Penaeus monodon[79] |
| C-type lectin | Litopenaeus vannamei[76] |
| C-type lectin (LvCTL1) | Litopenaeus vannamei[68] |
| FcLectin | Fenneropenaeus chinensis[83] |
| Fortilin | Penaeus monodon[70] |
| Hemocyanin | Marsupenaeus japonicus[73] |
| Penaeus monodon[72] | |
| LGBP | Penaeus stylirostris[58] |
| Manganese superoxide dismutase | Fenneropenaeus chinensis[82] |
| PmAV | Penaeus monodon[67] |
| PmCBP | Penaeus monodon[74] |
| PmRab7 | Penaeus monodon[30] |
| Rab GTPase | Marsupenaeus japonicus[81] |
| Ran protein | Marsupenaeus japonicus[78] |
| Syntenin | Penaeus monodon[71] |
| Syntenin-like protein gene | Penaeus monodon[84] |
A well-known cationic protein, the anti-lipopolysaccharide factor (ALF) originally isolated from the horseshoe crab Limulus polyphemus [62], [63] has been well studied in crustaceans for its antibacterial activity [64], [65]. Interestingly, the crayfish ALF was up-regulated by a WSSV challenge and was shown to be involved in antiviral response against WSSV. Silencing of ALF specially resulted in higher rates of WSSV propagation both in the animals and in a cell culture of Hpt from crayfish. In contrast, enhanced expression of ALF in the animals by the administration of UV-treated WSSV led to lower viral replication and a partial protection against a subsequent challenge with the active virus [45]. ALF from shrimp was also up-regulated in Litopenaeus vannamei upon WSSV challenge [66]. Silencing of LvALF1 resulted in a significant increase of mortality in L. vannamei challenged by Vibrio penaeicida and Fusarium oxysporum indicating that LvALF1 has a role in protecting shrimp from both bacterial and fungal infections, but not to a WSSV infection even if it was up-regulated by WSSV infection [65]. This is different from crayfish ALF which showed antiviral property both in vivo and in vitro [45]. The mechanism for the antiviral activity of crayfish ALF is still unknown. Further studies have shown that recombinant ALFPm3, from shrimp P. monodon affected the viral infection by inhibiting the attachment of virus to the crayfish Hpt cells suggesting that ALF may act during the initial stages of virus entry to host cells (unpublished data). The mechanism of this inhibition still needs further investigations. A gene named as PmAV was found to be up-regulated in virus resistant shrimp and the PmAV protein has a C-type lectin-like domain (CTLD). Recombinant PmAV protein displayed a strong antiviral activity in inhibiting virus-induced cytopathic effect in fish cells in vitro. Further experiments showed that PmAV did not bind to the WSSV implying that the antiviral mechanism of this protein was not due to inhibition of the attachment of virus to target host cell [67]. A novel C-type lectin (LvCTL1) showed strong affinity to WSSV and interacted with several envelope proteins of WSSV. Binding of recombinant LvCTL1 to WSSV could protect the shrimp from viral infection and could delay the survival of shrimps against WSSV infection [68]. Besides, a beta-integrin was found to interact with a WSSV envelope protein VP187 containing the RGD motif. Soluble integrin, integrin-specific antibody and an RGD containing peptide could block the WSSV infection in vivo and in vitro. Silencing of beta-integrin efficiently inhibited the virus infection. These data suggest that this beta-integrin may function as a cellular receptor for WSSV infection [69]. Injection of recombinant shrimp fortilin followed by WSSV challenge resulted in decreased viral infection by an unknown mechanism in P. monodon [70]. A syntenin and its protein partner alpha-2-macroglobulin co-precipitated with each other and both of them were up-regulated in the acute-phase of a WSSV infection [71]. Hemocyanin, the respiratory protein of arthropods and molluscs, was found to exhibit non-specific antiviral properties [72] and to delay the infection of WSSV in vivo in P. japonicus [73]. Other host proteins like a chitin-binding protein [74] and actin microfilaments have been shown to interact with WSSV component proteins [75]. Some other genes have been found to be up-regulated as a response to WSSV infection in the animals, but no mechanistic studies have been performed. These genes include a lipopolysaccharide and beta-1,3-glucan binding protein gene [58], a C-type lectin [76], a catalase gene [77], a Ras-related nuclear protein (Ran protein) gene [78], a caspase-3 like gene [79], calreticulin [80], a Rab GTPase gene [81], manganese superoxide dismutase [82], Fclectin [83], and a syntenin-like protein gene (Table 1) [84]. The functional characterization of these genes/proteins will bring interesting insights into the antiviral defence in crustaceans.
4.2. Antiviral activity induced by antibody or immune stimulants
WSSV proteins have been studied as possible immune therapeutic agents to prevent WSSV infections. For instance, a number of WSSV envelope proteins, such as VP28, have been proposed to be involved in viral infectivity based on the ability of specific antibodies to attenuate WSSV-induced mortality in vivo. When injected intramuscularly or administered orally with VP28, the shrimps obtained a higher and prolonged survival rates after WSSV challenge [85], [86]. Different protein/DNA vaccinations against WSSV infection were reported for the protection of shrimp/crayfish [87], [88], [89], [90], [91], [92], [93], [94]. Other antibodies against WSSV envelope proteins, such as VP68, VP281 and VP466 were also shown to reduce and delay the mortality of shrimp challenged with WSSV [95]. However, strong inactivation of WSSV by some normal rabbit sera was observed in a manner independent of anti-VP28 antibodies questioning the potential of anti-VP28 preimmune antibodies to specifically neutralize WSSV [96]. Thus, further investigations are necessary for the availability of these antibodies against different viral proteins. Preincubation of WSSV with mature synthetic mytilin (a synthetic antibacterial peptide from Mytilus galloprovincialis) significantly reduced shrimp mortality. This production was suggested to be caused by the interaction between the peptide and virus, but no mechanism was present [97].
Some other immune stimulants have also been described such as: glucans derived from yeast [98], [99], LPS from bacteria [100], inactivated viruses [101], [102], and dsRNA [103], [104], [105], [106]. Among these immune stimulants, viral proteins and dsRNA might be of particular interest for further investigations, because as virus-associated molecules they are likely to be the targets of immune recognition in the context of natural viral infections [107]. Recently, grossly normal shrimp has been found sometimes to be infected with one or more viruses [108], [109]. A viral accommodation concept has been introduced to the shrimp–virus interaction which provides more clues for a better understanding of the crustacean–virus interaction [110].
4.3. Cytokine activation mediated antiviral response
In vertebrates, most cell types respond to invading viral infection by rapidly releasing antiviral cytokines such as interferon alpha/beta (IFN-α/β). IFN-α/β is able to trigger activation of multiple noncytolytic intracellular antiviral pathways that can interfere with many steps in the life cycles of virus, thereby limiting the amplification and speed of the virus and attenuating the infection [111]. After the cells are activated by IFNs, signal transduction through the Janus kinase/signal transducer and activator of transcription (JAK/STAT) system will induce the expression of hundreds of genes [112], [113]. The widely studied type I IFN-induced genes are the dsRNA-activated serine/threonine protein kinase (PKR), the myxovirus-resistance (Mx) proteins, oligoadenylate synthetase, RNaseL, RNA-specific adenosine deaminase ADAR and IFNs themselves [114]. These self-amplifying systems can be triggered not only by IFN, but also directly by viral components. Cytokine activation through JAK/STAT pathway of a number of genes has been suggested in countering viral infection in Drosophila [115]. Flies deficient in the JAK kinase Hopscotch show increased susceptibility to Drosophila C virus and contain a higher viral load. These data indicate that flies produce antiviral molecules in a JAK-STAT-dependent way [116]. The JAK-STAT signaling pathways are also considered as a part of the antiviral response in mosquitoes [117].
The WSSV immediate early gene (ie1) was shown to employ a shrimp STAT as a transcription factor to enhance its expression and this resulted in high promoter activity in the host cells (Fig. 4 ) [118]. A further study showed that shrimp STAT was activated in response to WSSV infection and the WSSV does not disrupt JAK-STAT pathway but benefits from STAT activation in the shrimp [119]. WSSV ie1 protein also exhibits transactivation, dimerization, and DNA binding activity. This might provide more insights into the mechanism of how the replication of WSSV is manipulated in the host cells [120]. In addition, some components of the Toll pathway (Toll and Dif) have also been shown to be of importance for the resistance against Drosophila X virus [121]. But although a few Toll receptors have been found in shrimp [122], [123], their importance in any innate immune reactions in crustacean is totally unknown.
Fig. 4.
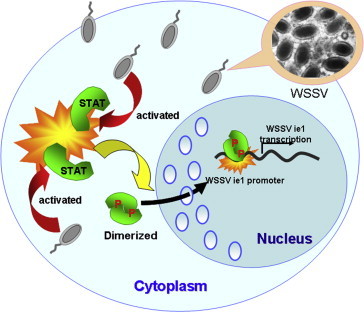
STAT–WSSV interaction based on Lo's work [118]. This model is modified from Arbouzova et al. [182]. This model suggests that the host STAT can be activated after WSSV invasion. Activated STAT will dimerize and be phosphorylated, which then enters the nucleus and activates the promoter of WSSV ie1 gene for viral transcription.
4.4. Apoptosis
Apoptosis is a critical cellular process for removing unnecessary or potentially harmful cells and for development [124]. Apoptosis is also regulated as an innate cellular response to limit virus production and also decrease or eliminate spread of progeny virus in the host [125]. Many viruses have evolved genes encoding proteins to effectively suppress or delay apoptosis for producing sufficient quantities of progeny. For instance, Baculovirus contains p35 and inhibitor of apoptosis proteins [126] which can inhibit multiple caspases [127], [128], [129]. Besides, an increasing number of viruses are shown to induce apoptosis actively at late stages of infection. This process may function as a final and key step in the spread of progeny virus to neighbouring cells and also to evade from host immune inflammatory responses and protect progeny virus from enzymes and antibodies [130]. Increasing numbers of cells showed DNA fragmentation as the animals approach death following WSSV or YHV infections, suggesting that apoptosis occurs after infections by WSSV [43], [131], [132] or YHV [133] in shrimp. Caspases are central effectors in apoptosis [134] and if in a shrimp, Marsupenaeus japonicus, the Pjcaspase gene was silenced, the WSSV-induced apoptosis was significantly inhibited and this resulted in an increase of viral copies, indicating that apoptosis may play a role in antiviral processes of shrimp [135]. This proposal however needs to be ascertained by more experiments.
The percentage of apoptotic hemocytes in WSSV-infected crayfish was very low, but it was significantly higher than that in the sham-injected crayfish on day 3 or 5 post-infection [35]. Apoptosis has been considered to constitute a defence responsible for eliminating virus in naturally TSV infected shrimp [136]. However, other studies [132], [137] suggest that apoptosis cannot be taken as an effective protection against WSSV in shrimp. For example by knocking down caspase-3 by RNAi this reduces mortality in Pacific white shrimp challenged with a low dose of WSSV but not if a high-dose of WSSV is used. This suggests that apoptosis may increase rather than decrease mortality in WSSV-challenged shrimp [138]. Another study indicates that shrimp with gross signs of WSSV infection from shrimp farms exhibited up to 40% apoptotic cells, and it was suggested that apoptosis might be implicated in shrimp death [132]. Similarly, the widespread and progressive occurrence of apoptosis in P. monodon infected with YHV is a major cause of dysfunction and death of the host. YHV-infected cells show the signs of viral-triggered apoptosis like nuclear pyknosis and karyorrhexis [133]. The expression of ribophorin I was up-regulated and remained high until the moribund stage in YHV-infected shrimp [139]. Ribophorin I belongs to the oligosaccharyltransferase (OST) complex involved in apoptosis [140]. Whereas, the transcriptional level of defender against apoptotic death 1 (DAD1), a negative regulator of apoptosis [141], decreased dramatically after YHV challenge in P. monodon, suggesting that DAD1 plays a role in mortality caused by YHV [142]. A shrimp translationally controlled tumor protein (TCTP) is an anti-apoptotic protein which has been implied to play a key role in shrimp defence responses or in control over viral-triggered apoptosis [143]. Whereas a hepatopancreatic nuclease is shown unlikely to be involved in viral-triggered apoptosis in shrimp [144]. The DNA fragmentation for WSSV-infected shrimp is the possible result of WSSV nuclease activity rather than shrimp nuclease activity. WSSV nuclease was identified and clearly shown to be expressed in the hepatopancreas of WSSV-infected shrimp [145]. However, signs of endogenous apoptosis have been shown in the shrimp lymphoid organ without known viral infections [133], [136]. Thus, still quite a lot of controversy remains for the role of apoptosis in antiviral responses in crustaceans and this may be due to different species used for the assays, the big variations of animals, different experimental manipulations, etc. Besides, apoptosis is a complex process involving the interaction of many proteins. Few of these proteins have been characterized from crustaceans. Therefore, studies are still wanted of the process and the role of apoptosis in the antiviral response in crustaceans.
4.5. Antiviral activity induced by RNA interference or injection of dsRNA
RNA interference (RNAi) is triggered by dsRNA processed into shorter 21–25 bp small interfering RNAs (siRNA) by the type III endonuclease Dicer [146]. The siRNA are then incorporated into the RNA-induced silencing complex (RISC) [147], [148], which facilitates the binding of the siRNA to the homologous mRNA upon which the targeted mRNA will be then degraded. RNAi has been proven to be a natural antiviral mechanism in plants [149], fruit flies [150], [151], [152], [153], mosquitoes [154], nematodes [155], [156], and mammalian cells [157], [158], [159]. The RNAi technique is now explored as an alternative and more specific approach to counteract virus infections in shrimps. Injection of dsRNA/siRNA specific to viral genes can block viral disease progression. This effect has been confirmed with different unrelated viruses. For instance, viral replication was efficiently suppressed with injection of WSSV-specific dsRNA/siRNA targeting VP19, VP28, VP281, or WSSV protein kinase in penaeid shrimp [103], [160], [161]. Lower YHV replication was observed in shrimp primary cell cultures by transfecting the cells with dsRNA targeted to the viral nonstructural genes [162]. Inhibition of YHV replication by cognate dsRNA significantly resulted in lower mortality in the black tiger shrimp [105], [163]. All these data strongly imply the important role of RNAi in antiviral protection, which might also offer applied practice in shrimp culture in the near future.
On the other hand, dsRNA is a potent inducer of the IFN response in vertebrates. It is a molecule that often forms during viral infection as a result of viral genomic replication and viral RNAs with extensive secondary structure [164]. In mammals, dsRNA is recognized by TLR3, which activates myeloid differentiation factor 88 (Myd88)-dependent and an independent signal transduction cascade, resulting in expression of IFN-β [165], [166]. DsRNA can also induce antiviral responses intracellularly by directly activating PKR, which results in inhibition of cellular and viral protein synthesis via phosphorylation of eukaryotic translation initiation factor 2 α (eIF2α) [167]. Further, some of the components of the pro-apoptotic pathways (e.g. PKR, RNaseL, IFN regulatory factor 3, and c-Jun N-terminal kinase) can be activated by dsRNA [168]. Hence, dsRNA is involved in both signaling viral infection and inducing apoptosis. It has been generally agreed that these dsRNA-induced IFNs are lacking in invertebrates [169], [170], [171]. Recent studies reveal the existence of both innate (non-sequence specific) and RNAi related (sequence specific) antiviral phenomena in a crustacean model [103], [104], [107], [172]. But the protection induced by dsRNA could be overwhelmed by a higher dose (8-fold) of infectious virus, which means that sequence independent dsRNA induces quite a low induction of resistance to virus infection. The mechanism of this difference between higher and lower amount of infectious virus remains unknown. The protective efficiency for WSSV infection by specific dsRNAs is also varied between different viral genes targeted and no explanations have been addressed for these differences in this study. Other studies have shown that viral RNAs exhibit different sensitivities to RNAi attack [173]. This may results from protective proteins that bind to the viral RNA, or protective RNA structure because the viral RNA presents in a virus particle or a subcellular compartment that is not accessible to the RNAi machinery [173]. It remains unclear whether the antiviral protection by virus-specific long dsRNA is the result of RNAi mechanism alone or the combination of innate immune activation and RNAi in shrimp. Further, the readout from these studies was mainly based on the cumulative mortalities and no mechanism so far is available for these antiviral phenomena. Importantly, these studies suggest a possible evolutionary link (recognition of dsRNA) between innate antiviral immunity in invertebrates and vertebrates (Fig. 5 ). DsRNAs have been im-plied as important regulators for gene expression [174], [175]. The distinction between self and non-self dsRNA is suggested by different structural features including 5′ triphosphate, 3′ protruding end or other specific motifs. Following the identification of RNA helicases, retinoic acid inducible gene-I, melanoma differentiation associated gene-5, and TLRs 7/8, more details will be elucidated on how different dsRNAs are selectively recognized by different sensors of the innate immune responses [168]. Based on the antiviral activity induced by dsRNA in shrimp, it would be also interesting to elucidate the possible role, potency and application of dsRNA in antiviral immunity in crustaceans.
Fig. 5.
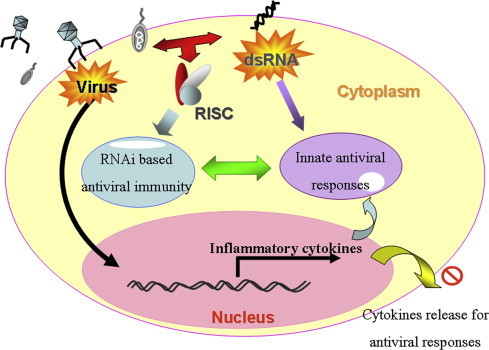
A hypothetical scheme for dsRNA-induced antiviral immunity in crustacean (modified from Robalino et al. 2007) [107]. Viral dsRNA or extracellular dsRNA enters the RNAi pathway, and/or activates innate antiviral responses directly or indirectly. Signal transduction triggered by dsRNA recognition may regulate transcriptomic expression leading to antiviral reactions. These pathways may function independently as well as interact together to facilitate the antiviral responses.
5. Concluding remarks
Crustaceans (e.g. crayfish, crabs, lobster and shrimp) mount strong innate immune responses against microbes such as bacteria, fungi, and viruses. Studies on the mechanism of these responses have been hampered by absence of genome, tools for genetic manipulation and mutants, and stable long-term cell lines for in vitro studies. We have succeeded in developing an Hpt cell culture from crayfish, which can be a useful tool for gene functional studies in crustaceans [46]. This Hpt cultures can also be used to replicate WSSV and to study host–virus interactions (Fig. 4) [176]. A novel dsRNA mediated RNAi technology has been developed for crustacean cells [177], which is useful for antiviral studies in crustaceans.
RNAi is a powerful tool for antiviral responses [173]. Compared to other pharmacological interventions, RNAi is an attractive antiviral therapeutic because it allows interference with the target gene in a highly sequence-specific manner, and therefore essential viral genes can be targeted by design with little or no risk of unexpected off-target effects. Recent studies have demonstrated that the viral disease progression can be blocked by injecting shrimp with dsRNA/siRNA specific to viral genes. This strategy is effective against three unrelated viruses: WSSV, TSV and YHV [104], [105]. The mechanism for this phenomenon is still not clear. In these studies, positive strand RNA virus (e.g. TSV and YHV) and DNA virus (e.g. WSSV) often induce the formation of dsRNAs during their infectious cycles such as genomic replicative intermediates, intramolecular interactions within viral transcripts, and bi-directional transcription [178]. And these dsRNAs might engage the shrimp RNAi pathway, resulting in effective antiviral responses. Thus it will be of interest to investigate whether viral dsRNA accumulates in crustacean cells infected with WSSV, TSV, or YHV. Two components of RNA silencing, Argonaute and Dicer-1, have been characterized from P. monodon [179], [180], [181]. Based on the studies in crustaceans above, plants [149], fruit flies [150], [151], [152], mosquitoes [154], and nematodes [155], [156], it is likely that RNAi indeed exists as a natural antiviral mechanism in several organisms including crustaceans. These studies and further investigations of genes involved in RNAi in crustaceans should provide the possibility by reverse-genetic approaches to test this hypothesis directly.
Acknowledgement
This work has been supported by Swedish Research Council, Formas (to KS) and The Swedish Research Council to K.S. and P.J. A special thank to Mr. Apiruck Watthanasurorot for drawing the figures.
References
- 1.Fuhrman J.A. Marine viruses and their biogeochemical and ecological effects. Nature. 1999;399:541–548. doi: 10.1038/21119. [DOI] [PubMed] [Google Scholar]
- 2.Loh P.C., Tapay L.M., Lu Y., Nadala E.C., Jr. Viral pathogens of the penaeid shrimp. Adv Virus Res. 1997;48:263–312. doi: 10.1016/S0065-3527(08)60290-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Vago C. A virus disease in Crustacea. Nature. 1966;209:1290. [Google Scholar]
- 4.Jiravanichpaisal P. Uppsala University; 2005. White spot syndrome virus interaction with a freshwater crayfish. Digital comprehensive summaries of Uppsala dissertations. [Google Scholar]
- 5.Bonami J.R., Lightner D.V. Unclassified viruses of crustacean. In: Adams J.R., Bonami J.R., editors. CRC Press; Boca Raton: 1991. pp. 597–622. [Google Scholar]
- 6.Dhar A.K., Cowley J.A., Hasson K.W., Walker P.J. Genomic organization, biology, and diagnosis of Taura syndrome virus and yellowhead virus of penaeid shrimp. Adv Virus Res. 2004;63:353–421. doi: 10.1016/S0065-3527(04)63006-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.van Hulten M.C.W., Witteveldt J., Peters S., Kloosterboer N., Tarchini R., Fiers M. The white spot syndrome virus DNA genome sequence. Virology. 2001;286:7–22. doi: 10.1006/viro.2001.1002. [DOI] [PubMed] [Google Scholar]
- 8.Yang F., He J., Lin X., Li Q., Pan D., Zhang X. Complete genome sequence of the shrimp white spot bacilliform virus. J Virol. 2001;75:11811–11820. doi: 10.1128/JVI.75.23.11811-11820.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Durand S., Lightner D.V., Redman R.M., Bonami J.R. Ultrastructure and morphogenesis of white spot syndrome baculovirus (WSSV) Dis Aquat Org. 1997;29:205–211. [Google Scholar]
- 10.Escobedo-Bonilla C.M., Alday-Sanz V., Wille M., Sorgeloos P., Pensaert M.B., Nauwynck H.J. A review on the morphology, molecular characterization, morphogenesis and pathogenesis of white spot syndrome virus. J Fish Dis. 2008;31:1–18. doi: 10.1111/j.1365-2761.2007.00877.x. [DOI] [PubMed] [Google Scholar]
- 11.Lo C.F., Ho C.H., Chert C.H., Liu K.F., Chiu Y.L., Yeh P.Y. Detection and tissue tropism of white spot syndrome baculovirus (WSBV) in captured brooders of Penaeus monodon with a special emphasis in reproductive organs. Dis Aquat Org. 1997;30:53–72. [Google Scholar]
- 12.Jiravanichpaisal P., Bangyeekhun E., Söderhäll K., Söderhäll I. Experimental infection of white spot syndrome virus in freshwater crayfish Pacifastacus leniusculus. Dis Aquat Org. 2001;47:151–157. doi: 10.3354/dao047151. [DOI] [PubMed] [Google Scholar]
- 13.Wongteerasupaya C., Sriurairatana S., Vickers J.E., Anutara A., Boonsaeng V., Panyim S. Yellowhead virus of Penaeus monodon is an RNA virus. Dis Aquat Org. 1995;22:45–50. [Google Scholar]
- 14.Wang Y.C., Chang P.S. Yellowhead virus infection in the giant tiger prawn Penaeus monodon cultured in Taiwan. Fish Pathol. 2000;35:1–10. [Google Scholar]
- 15.Boonyaratpalin S., Supamataya K., Kasornchnadra J., Direkbusarakom S., Aekpanithanpong U., Chantanachookin C. Non-occluded baculo-like virus, the causative agent of yellow-head disease in the black tiger shrimp (Penaeus monodon) Fish Pathol. 1993;28:103–109. [Google Scholar]
- 16.Chantanachookin C.S., Boonyaratpalin J., Kasornchnadra S., Direkbusarakom U., Ekpanithanpong K., Supamataya S. Histology and ultrastructure reveal a new granulosis-like virus in Penaeus monodon affected by yellowhead disease. Dis Aqua Org. 1993;17:145–157. [Google Scholar]
- 17.Wang C.S., Tang K.J., Kuo G.H., Chen S.N. Yellowhead disease-like virus infection in the kuruma shrimp Penaeus japonicus cultured in Taiwan. Fish Pathol. 1996;31:177–182. [Google Scholar]
- 18.Assavalapsakul W., Smith D.R., Panyim S. Identification and characterization of a Penaeus monodon lymphoid cell-expressed receptor for the yellow head virus. J Virol. 2006;80:262–269. doi: 10.1128/JVI.80.1.262-269.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lu Y., Tapay L.M., Loh P.C., Brock J.A., Gose R.B. Distribution of yellow head virus in selected tissue and organs of penaeid shrimp Penaeus vannamei. Dis Aqua Org. 1995;23:67–70. [Google Scholar]
- 20.Jimenez R. Sindrome de taura (Resumen) Acua Ecuador. 1992;1:1–16. [Google Scholar]
- 21.Bonami J.R., Hasson K.W., Mari J., Poulos B.T., Lightner D.V. Taura syndrome of marine penaeid shrimp: characterization of the viral agent. J Gen Virol. 1997;78(Pt 2):313–319. doi: 10.1099/0022-1317-78-2-313. [DOI] [PubMed] [Google Scholar]
- 22.Mari J., Poulos B.T., Lightner D.V., Bonami J.R. Shrimp taura syndrome virus: genomic characterization and similarity with members of the genus Cricket paralysis-like viruses. J Gen Virol. 2002;83:915–926. doi: 10.1099/0022-1317-83-4-915. [DOI] [PubMed] [Google Scholar]
- 23.van Regenmortel M.H.V., Fauquet C.M., Bishop D.H.L., Carstens E.B., Estes M.K., Lemon S.M. The seventh report of the international committee on taxonomy of viruses. Academic Press; San Diego, CA: 2000. Virus taxonomy: the classification and nomenclature of viruses”. [Google Scholar]
- 24.Flegel T.W. Special topic review: major viral diseases of the black tiger prawn (Penaeus monodon) in Thailand. World J Microbiol Biotechnol. 1997;13:433–442. [Google Scholar]
- 25.Lightner D.V. Epizootiology, distribution, and the impact on international trade of two penaeid shrimp viruses in the Americas. Rev Sci Off Int Epiz. 1996;5:579–601. doi: 10.20506/rst.15.2.944. [DOI] [PubMed] [Google Scholar]
- 26.Lightner D.V., Redman R.M. Shrimp diseases and current diagnostic methods. Aquaculture. 1998;164:201–220. [Google Scholar]
- 27.Klasse P.J., Bron R., Marsh M. Mechanisms of enveloped virus entry into animal cells. Adv Drug Deliv Rev. 1998;34:65–91. doi: 10.1016/S0169-409X(98)00002-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Spaan W., Cavanagh D., Horzinek M.C. Coronaviruses: structure and genome expression. J Gen Virol. 1988;69(Pt 12):2939–2952. doi: 10.1099/0022-1317-69-12-2939. [DOI] [PubMed] [Google Scholar]
- 29.Assavalapsakul W., Tirasophon W., Panyim S. Antiserum to the gp116 glycoprotein of yellow head virus neutralizes infectivity in primary lymphoid organ cells of Penaeus monodon. Dis Aqua Org. 2005;63:85–88. doi: 10.3354/dao063085. [DOI] [PubMed] [Google Scholar]
- 30.Sritunyalucksana K., Wannapapho W., Lo C.F., Flegel T.W. PmRab7 is a VP28-binding protein involved in white spot syndrome virus infection in shrimp. J Virol. 2006;80:10734–10742. doi: 10.1128/JVI.00349-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ongvarrasopone C., Chanasakulniyom M., Sritunyalucksana K., Panyim S. Suppression of PmRab7 by dsRNA Inhibits WSSV or YHV Infection in Shrimp. Mar Biotechnol (NY) 2008;10:374–381. doi: 10.1007/s10126-007-9073-6. [DOI] [PubMed] [Google Scholar]
- 32.Wu W., Zong R., Xu J., Zhang X. Antiviral phagocytosis is regulated by a novel Rab-dependent complex in shrimp Penaeus japonicus. J Proteome Res. 2008;7:424–431. doi: 10.1021/pr700639t. [DOI] [PubMed] [Google Scholar]
- 33.Yi G., Wang Z., Qi Y., Yao L., Qian J., Hu L. Vp28 of shrimp white spot syndrome virus is involved in the attachment and penetration into shrimp cells. J Biochem Mol Biol. 2004;37:726–734. doi: 10.5483/bmbrep.2004.37.6.726. [DOI] [PubMed] [Google Scholar]
- 34.Wang Y.T., Liu W., Seah J.N., Lam C.S., Xiang J.H., Korzh V. White spot syndrome virus (WSSV) infects specific hemocytes of the shrimp Penaeus merguiensis. Dis Aquat Org. 2002;52:249–259. doi: 10.3354/dao052249. [DOI] [PubMed] [Google Scholar]
- 35.Jiravanichpaisal P., Sricharoen S., Söderhäll I., Söderhäll K. White spot syndrome virus (WSSV) interaction with crayfish haemocytes. Fish Shellfish Immunol. 2006;20:718–727. doi: 10.1016/j.fsi.2005.09.002. [DOI] [PubMed] [Google Scholar]
- 36.Johansson M.W. Cell adhesion molecules in invertebrate immunity. Dev Comp Immunol. 1999;23:303–315. doi: 10.1016/s0145-305x(99)00013-0. [DOI] [PubMed] [Google Scholar]
- 37.Johansson M.W., Keyser P., Sritunyaluksana K., Söderhäll K. Crustacean hemocytes and haematopoiesis. Aquaculture. 2000;191:45–52. [Google Scholar]
- 38.Johansson M.W., Holmblad T., Thörnqvist P.O., Cammarata M., Parrinello N., Söderhäll K. A cell-surface superoxide dismutase is a binding protein for peroxinectin, a cell-adhesive peroxidase in crayfish. J Cell Sci. 1999;112(Pt 6):917–925. doi: 10.1242/jcs.112.6.917. [DOI] [PubMed] [Google Scholar]
- 39.Johansson M.W., Söderhäll K. Intracellular signaling in arthropod blood cells: involvement of protein kinase C and protein tyrosine phosphorylation in the response to the 76-kDa protein or the beta-1,3-glucan-binding protein in crayfish. Dev Comp Immunol. 1993;17:495–500. doi: 10.1016/s0145-305x(05)80005-9. [DOI] [PubMed] [Google Scholar]
- 40.Jiravanichpaisal P., Lee B.L., Söderhäll K. Cell-mediated immunity in arthropods: hematopoiesis, coagulation, melanization and opsonization. Immunobiology. 2006;211:213–236. doi: 10.1016/j.imbio.2005.10.015. [DOI] [PubMed] [Google Scholar]
- 41.van de Braak C.B., Botterblom M.H., Huisman E.A., Rombout J.H., van der Knaap W.P. Preliminary study on haemocyte response to white spot syndrome virus infection in black tiger shrimp Penaeus monodon. Dis Aquat Org. 2002;51:149–155. doi: 10.3354/dao051149. [DOI] [PubMed] [Google Scholar]
- 42.Guan Y., Yu Z., Li C. The effects of temperature on white spot syndrome infections in Marsupenaeus japonicus. J Invertebr Pathol. 2003;83:257–260. doi: 10.1016/s0022-2011(03)00068-5. [DOI] [PubMed] [Google Scholar]
- 43.Wongprasert K., Khanobdee K., Glunukarn S.S., Meeratana P., Withyachumnarnkul B. Time-course and levels of apoptosis in various tissues of black tiger shrimp Penaeus monodon infected with white-spot syndrome virus. Dis Aquat Org. 2003;55:3–10. doi: 10.3354/dao055003. [DOI] [PubMed] [Google Scholar]
- 44.Cerenius L., Lee B.L., Söderhäll K. The proPO-system: pros and cons for its role in invertebrate immunity. Trends Immunol. 2008;29:263–271. doi: 10.1016/j.it.2008.02.009. [DOI] [PubMed] [Google Scholar]
- 45.Liu H., Jiravanichpaisal P., Söderhäll I., Cerenius L., Söderhäll K. Antilipopolysaccharide Factor interferes with white spot syndrome virus replication in vitro and in vivo in the crayfish Pacifastacus leniusculus. J Virol. 2006;80:10365–10371. doi: 10.1128/JVI.01101-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Söderhäll I., Bangyeekhun E., Mayo S., Söderhäll K. Hemocyte production and maturation in an invertebrate animal; proliferation and gene expression in hematopoietic stem cells of Pacifastacus leniusculus. Dev Comp Immunol. 2003;27:661–672. doi: 10.1016/s0145-305x(03)00039-9. [DOI] [PubMed] [Google Scholar]
- 47.Orkin S.H. Embryonic stem cells and transgenic mice in the study of hematopoiesis. Int J Dev Biol. 1998;42:927–934. [PubMed] [Google Scholar]
- 48.Orkin S.H. Diversification of haematopoietic stem cells to specific lineages. Nat Rev Genet. 2000;1:57–64. doi: 10.1038/35049577. [DOI] [PubMed] [Google Scholar]
- 49.Sieweke M.H., Graf T. A transcription factor party during blood cell differentiation. Curr Opin Genet Dev. 1998;8:545–551. doi: 10.1016/s0959-437x(98)80009-9. [DOI] [PubMed] [Google Scholar]
- 50.Wu C., Söderhäll I., Kim Y., Liu H., Söderhäll K. Hemocyte lineage marker proteins in a crustacean, the freshwater crayfish, Pacifastacus leniusculus. Proteomics. 2008;8:4226–4235. doi: 10.1002/pmic.200800177. [DOI] [PubMed] [Google Scholar]
- 51.Kawai T., Akira S. Innate immune recognition of viral infection. Nat Immunol. 2006;7:131–137. doi: 10.1038/ni1303. [DOI] [PubMed] [Google Scholar]
- 52.Pan J., Kurosky A., Xu B., Chopra A.K., Coppenhaver D.H., Singh I.P. Broad antiviral activity in tissues of crustaceans. Antiviral Res. 2000;48:39–47. doi: 10.1016/s0166-3542(00)00117-0. [DOI] [PubMed] [Google Scholar]
- 53.Lan Y., Xu X., Yang F., Zhang X. Transcriptional profile of shrimp white spot syndrome virus (WSSV) genes with DNA microarray. Arch Virol. 2006;151:1723–1733. doi: 10.1007/s00705-006-0751-x. [DOI] [PubMed] [Google Scholar]
- 54.Wang B., Li F., Dong B., Zhang X., Zhang C., Xiang J. Discovery of the genes in response to white spot syndrome virus (WSSV) infection in Fenneropenaeus chinensis through cDNA microarray. Mar Biotechnol (NY) 2006;8:491–500. doi: 10.1007/s10126-005-6136-4. [DOI] [PubMed] [Google Scholar]
- 55.Wang H.C., Wang H.C., Leu J.H., Kou G.H., Wang A.H., Lo C.F. Protein expression profiling of the shrimp cellular response to white spot syndrome virus infection. Dev Comp Immunol. 2007;31:672–686. doi: 10.1016/j.dci.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 56.Zhao Z.Y., Yin Z.X., Weng S.P., Guan H.J., Li S.D., Xing K. Profiling of differentially expressed genes in hepatopancreas of white spot syndrome virus-resistant shrimp (Litopenaeus vannamei) by suppression subtractive hybridisation. Fish Shellfish Immunol. 2007;22:520–534. doi: 10.1016/j.fsi.2006.07.003. [DOI] [PubMed] [Google Scholar]
- 57.Rojtinnakorn J., Hirono I., Itami T., Takahashi Y., Aoki T. Gene expression in haemocytes of kuruma prawn, Penaeus japonicus, in response to infection with WSSV by EST approach. Fish Shellfish Immunol. 2002;13:69–83. doi: 10.1006/fsim.2001.0382. [DOI] [PubMed] [Google Scholar]
- 58.Roux M.M., Pain A., Klimpel K.R., Dhar A.K. The lipopolysaccharide and beta-1,3-glucan binding protein gene is upregulated in white spot virus-infected shrimp (Penaeus stylirostris) J Virol. 2002;76:7140–7149. doi: 10.1128/JVI.76.14.7140-7149.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Pan D., He N., Yang Z., Liu H., Xu X. Differential gene expression profile in hepatopancreas of WSSV-resistant shrimp (Penaeus japonicus) by suppression subtractive hybridization. Dev Comp Immunol. 2005;29:103–112. doi: 10.1016/j.dci.2004.07.001. [DOI] [PubMed] [Google Scholar]
- 60.He N., Qin Q., Xu X. Differential profile of genes expressed in hemocytes of White Spot Syndrome Virus-resistant shrimp (Penaeus japonicus) by combining suppression subtractive hybridization and differential hybridization. Antiviral Res. 2005;66:39–45. doi: 10.1016/j.antiviral.2004.12.010. [DOI] [PubMed] [Google Scholar]
- 61.Dhar A.K., Dettori A., Roux M.M., Klimpel K.R., Read B. Identification of differentially expressed genes in shrimp (Penaeus stylirostris) infected with White spot syndrome virus by cDNA microarrays. Arch Virol. 2003;148:2381–2396. doi: 10.1007/s00705-003-0172-z. [DOI] [PubMed] [Google Scholar]
- 62.Muta T., Miyata T., Tokunaga F., Nakamura T., Iwanaga S. Primary structure of anti-lipopolysaccharide factor from American horseshoe crab, Limulus polyphemus. J Biochem. 1987;101:1321–1330. doi: 10.1093/oxfordjournals.jbchem.a121999. [DOI] [PubMed] [Google Scholar]
- 63.Aketagawa J., Miyata T., Ohtsubo S., Nakamura T., Morita T., Hayashida H. Primary structure of limulus anticoagulant anti-lipopolysaccharide factor. J Biol Chem. 1986;261:7357–7365. [PubMed] [Google Scholar]
- 64.Pan C.Y., Chao T.T., Chen J.C., Chen J.Y., Liu W.C., Lin C.H. Shrimp (Penaeus monodon) anti-lipopolysaccharide factor reduces the lethality of Pseudomonas aeruginosa sepsis in mice. Int Immunopharmacol. 2007;7:687–700. doi: 10.1016/j.intimp.2007.01.006. [DOI] [PubMed] [Google Scholar]
- 65.de la Vega E., O'Leary N.A., Shockey J.E., Robalino J., Payne C., Browdy C.L. Anti-lipopolysaccharide factor in Litopenaeus vannamei (LvALF): a broad spectrum antimicrobial peptide essential for shrimp immunity against bacterial and fungal infection. Mol Immunol. 2008;45:1916–1925. doi: 10.1016/j.molimm.2007.10.039. [DOI] [PubMed] [Google Scholar]
- 66.Robalino J., Almeida J.S., McKillen D., Colglazier J., Trent H.F., III, Chen Y.A. Insights into the immune transcriptome of the shrimp Litopenaeus vannamei: tissue-specific expression profiles and transcriptomic responses to immune challenge. Physiol Genomics. 2007;29:44–56. doi: 10.1152/physiolgenomics.00165.2006. [DOI] [PubMed] [Google Scholar]
- 67.Luo T., Zhang X., Shao Z., Xu X. PmAV, a novel gene involved in virus resistance of shrimp Penaeus monodon. FEBS Lett. 2003;551:53–57. doi: 10.1016/s0014-5793(03)00891-3. [DOI] [PubMed] [Google Scholar]
- 68.Zhao Z.Y., Yin Z.X., Xu X.P., Weng S.P., Rao X.Y., Dai Z.X. A novel C-type lectin from the shrimp Litopenaeus vannamei possesses anti-WSSV activity. J Virol. 2008;83:347–356. doi: 10.1128/JVI.00707-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Li D.F., Zhang M.C., Yang H.J., Zhu Y.B., Xu X. Beta-integrin mediates WSSV infection. Virology. 2007;368:122–132. doi: 10.1016/j.virol.2007.06.027. [DOI] [PubMed] [Google Scholar]
- 70.Tonganunt M., Nupan B., Saengsakda M., Suklour S., Wanna W., Senapin S. The role of Pm-fortilin in protecting shrimp from white spot syndrome virus (WSSV) infection. Fish Shellfish Immunol. 2008 doi: 10.1016/j.fsi.2008.08.006. [DOI] [PubMed] [Google Scholar]
- 71.Tonganunt M., Phongdara A., Chotigeat W., Fujise K. Identification and characterization of syntenin binding protein in the black tiger shrimp Penaeus monodon. J Biotechnol. 2005;120:135–145. doi: 10.1016/j.jbiotec.2005.06.006. [DOI] [PubMed] [Google Scholar]
- 72.Zhang X., Huang C., Qin Q. Antiviral properties of hemocyanin isolated from shrimp Penaeus monodon. Antiviral Res. 2004;61:93–99. doi: 10.1016/j.antiviral.2003.08.019. [DOI] [PubMed] [Google Scholar]
- 73.Lei K., Li F., Zhang M., Yang H., Luo T., Xu X. Difference between hemocyanin subunits from shrimp Penaeus japonicus in anti-WSSV defense. Dev Comp Immunol. 2008;32:808–813. doi: 10.1016/j.dci.2007.11.010. [DOI] [PubMed] [Google Scholar]
- 74.Chen L.L., Lu L.C., Wu W.J., Lo C.F., Huang W.P. White spot syndrome virus envelope protein VP53A interacts with Penaeus monodon chitin-binding protein (PmCBP) Dis Aquat Org. 2007;74:171–178. doi: 10.3354/dao074171. [DOI] [PubMed] [Google Scholar]
- 75.Xie X., Yang F. Interaction of white spot syndrome virus VP26 protein with actin. Virology. 2005;336:93–99. doi: 10.1016/j.virol.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 76.Ma T.H., Tiu S.H., He J.G., Chan S.M. Molecular cloning of a C-type lectin (LvLT) from the shrimp Litopenaeus vannamei: early gene down-regulation after WSSV infection. Fish Shellfish Immunol. 2007;23:430–437. doi: 10.1016/j.fsi.2006.12.005. [DOI] [PubMed] [Google Scholar]
- 77.Zhang Q., Li F., Zhang X., Dong B., Zhang J., Xie Y. cDNA cloning, characterization and expression analysis of the antioxidant enzyme gene, catalase, of Chinese shrimp Fenneropenaeus chinensis. Fish Shellfish Immunol. 2008;24:584–591. doi: 10.1016/j.fsi.2008.01.008. [DOI] [PubMed] [Google Scholar]
- 78.Han F., Zhang X. Characterization of a ras-related nuclear protein (Ran protein) up-regulated in shrimp antiviral immunity. Fish Shellfish Immunol. 2007;23:937–944. doi: 10.1016/j.fsi.2007.01.022. [DOI] [PubMed] [Google Scholar]
- 79.Wongprasert K., Sangsuriya P., Phongdara A., Senapin S. Cloning and characterization of a caspase gene from black tiger shrimp (Penaeus monodon)-infected with white spot syndrome virus (WSSV) J Biotechnol. 2007;131:9–19. doi: 10.1016/j.jbiotec.2007.05.032. [DOI] [PubMed] [Google Scholar]
- 80.Luana W., Li F., Wang B., Zhang X., Liu Y., Xiang J. Molecular characteristics and expression analysis of calreticulin in Chinese shrimp Fenneropenaeus chinensis. Comp Biochem Physiol B Biochem Mol Biol. 2007;147:482–491. doi: 10.1016/j.cbpb.2007.03.001. [DOI] [PubMed] [Google Scholar]
- 81.Wu W., Zhang X. Characterization of a Rab GTPase up-regulated in the shrimp Peneaus japonicus by virus infection. Fish Shellfish Immunol. 2007;23:438–445. doi: 10.1016/j.fsi.2007.01.001. [DOI] [PubMed] [Google Scholar]
- 82.Zhang Q., Li F., Wang B., Zhang J., Liu Y., Zhou Q. The mitochondrial manganese superoxide dismutase gene in Chinese shrimp Fenneropenaeus chinensis: cloning, distribution and expression. Dev Comp Immunol. 2007;31:429–440. doi: 10.1016/j.dci.2006.08.005. [DOI] [PubMed] [Google Scholar]
- 83.Liu Y.C., Li F.H., Dong B., Wang B., Luan W., Zhang X.J. Molecular cloning, characterization and expression analysis of a putative C-type lectin (Fclectin) gene in Chinese shrimp Fenneropenaeus chinensis. Mol Immunol. 2007;44:598–607. doi: 10.1016/j.molimm.2006.01.015. [DOI] [PubMed] [Google Scholar]
- 84.Bangrak P., Graidist P., Chotigeat W., Supamattaya K., Phongdara A. A syntenin-like protein with postsynaptic density protein (PDZ) domains produced by black tiger shrimp Penaeus monodon in response to white spot syndrome virus infection. Dis Aquat Org. 2002;49:19–25. doi: 10.3354/dao049019. [DOI] [PubMed] [Google Scholar]
- 85.Witteveldt J., Cifuentes C.C., Vlak J.M., van Hulten M.C. Protection of Penaeus monodon against white spot syndrome virus by oral vaccination. J Virol. 2004;78:2057–2061. doi: 10.1128/JVI.78.4.2057-2061.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Witteveldt J., Vlak J.M., van Hulten M.C. Protection of Penaeus monodon against white spot syndrome virus using a WSSV subunit vaccine. Fish Shellfish Immunol. 2004;16:571–579. doi: 10.1016/j.fsi.2003.09.006. [DOI] [PubMed] [Google Scholar]
- 87.Kumar S.R., Ahamed V.P.I., Sarathi M., Basha A.N., Hameed A.S.S. Immunological responses of Penaeus monodon to DNA vaccine and its efficacy to protect shrimp against white spot syndrome virus (WSSV) Fish Shellfish Immunol. 2008;24:467–478. doi: 10.1016/j.fsi.2008.01.004. [DOI] [PubMed] [Google Scholar]
- 88.Jha R.K., Xu Z.R., Shen J., Bai S.J., Sun J.Y., Li W.F. The efficacy of recombinant vaccines against white spot syndrome virus in Procambarus clarkii. Immunol Lett. 2006;105:68–76. doi: 10.1016/j.imlet.2006.01.004. [DOI] [PubMed] [Google Scholar]
- 89.Li L.J., Yuan J.F., Cai C.A., Gu W.G., Shi Z.L. Multiple envelope proteins are involved in white spot syndrome virus (WSSV) infection in crayfish. Arch Virol. 2006;151:1309–1317. doi: 10.1007/s00705-005-0719-2. [DOI] [PubMed] [Google Scholar]
- 90.Rout N., Kumar S., Jaganmohan S., Murugan V. DNA vaccines encoding viral envelope proteins confer protective immunity against WSSV in black tiger shrimp. Vaccine. 2007;25:2778–2786. doi: 10.1016/j.vaccine.2006.12.056. [DOI] [PubMed] [Google Scholar]
- 91.Vaseeharan B., Prem Anand T., Murugan T., Chen J.C. Shrimp vaccination trials with the VP292 protein of white spot syndrome virus. Lett Appl Microbiol. 2006;43:137–142. doi: 10.1111/j.1472-765X.2006.01941.x. [DOI] [PubMed] [Google Scholar]
- 92.Witteveldt J., Vlak J.M., van Hulten M.C. Increased tolerance of Litopenaeus vannamei to white spot syndrome virus (WSSV) infection after oral application of the viral envelope protein VP28. Dis Aquat Org. 2006;70:167–170. doi: 10.3354/dao070167. [DOI] [PubMed] [Google Scholar]
- 93.Ha Y.M., Soo-Jung G., Thi-Hoai N., Ra C.H., Kim K.H., Nam Y.K. Vaccination of shrimp (Penaeus chinensis) against white spot syndrome virus (WSSV) J Microbiol Biotechnol. 2008;18:964–967. [PubMed] [Google Scholar]
- 94.Fu L.L., Li W.F., Du H.H., Dai W., Xu Z.R. Oral vaccination with envelope protein VP28 against white spot syndrome virus in Procambarus clarkii using Bacillus subtilis as delivery vehicles. Lett Appl Microbiol. 2008;46:581–586. doi: 10.1111/j.1472-765X.2008.02355.x. [DOI] [PubMed] [Google Scholar]
- 95.Wu W., Wang L., Zhang X. Identification of white spot syndrome virus (WSSV) envelope proteins involved in shrimp infection. Virology. 2005;332:578–583. doi: 10.1016/j.virol.2004.12.011. [DOI] [PubMed] [Google Scholar]
- 96.Robalino J., Payne C., Parnell P., Shepard E., Grimes A.C., Metz A. Inactivation of White Spot Syndrome Virus (WSSV) by normal rabbit serum: implications for the role of the envelope protein VP28 in WSSV infection of shrimp. Virus Res. 2006;118:55–61. doi: 10.1016/j.virusres.2005.11.011. [DOI] [PubMed] [Google Scholar]
- 97.Dupuy J.W., Bonami J.R., Roch P. A synthetic antibacterial peptide from Mytilus galloprovincialis reduces mortality due to white spot syndrome virus in palaemonid shrimp. J Fish Dis. 2004;27:57–64. doi: 10.1046/j.1365-2761.2003.00516.x. [DOI] [PubMed] [Google Scholar]
- 98.Chang C.F., Su M.S., Chen H.Y., Liao I.C. Dietary beta-1,3-glucan effectively improves immunity and survival of Penaeus monodon challenged with white spot syndrome virus. Fish Shellfish Immunol. 2003;15:297–310. doi: 10.1016/s1050-4648(02)00167-5. [DOI] [PubMed] [Google Scholar]
- 99.Huang C.C., Song Y.L. Maternal transmission of immunity to white spot syndrome associated virus (WSSV) in shrimp (Penaeus monodon) Dev Comp Immunol. 1999;23:545–552. doi: 10.1016/s0145-305x(99)00038-5. [DOI] [PubMed] [Google Scholar]
- 100.Takahashi Y., Kondo M., Itami T., Honda T., Inagawa H., Nishizawa T. Enhancement of disease resistance against penaeid acute viraemia and induction of virus-inactivating activity in haemolymph of kuruma shrimp, Penaeus japonicus, by oral administration of Pantoea agglomerans lipopolysaccharide (LPS) Fish Shellfish Immunol. 2000;10:555–558. doi: 10.1006/fsim.2000.0268. [DOI] [PubMed] [Google Scholar]
- 101.Melena J., Bayot B., Betancourt I., Amano Y., Panchana F., Alday V. Pre-exposure to infectious hypodermal and haematopoietic necrosis virus or to inactivated white spot syndrome virus (WSSV) confers protection against WSSV in Penaeus vannamei (Boone) post-larvae. J Fish Dis. 2006;29:589–600. doi: 10.1111/j.1365-2761.2006.00739.x. [DOI] [PubMed] [Google Scholar]
- 102.Bright Singh I.S., Manjusha M., Pai S.S., Philip R. Fenneropenaeus indicus is protected from white spot disease by oral administration of inactivated white spot syndrome virus. Dis Aquat Org. 2005;66:265–270. doi: 10.3354/dao066265. [DOI] [PubMed] [Google Scholar]
- 103.Robalino J., Bartlett T., Shepard E., Prior S., Jaramillo G., Scura E. Double-stranded RNA induces sequence-specific antiviral silencing in addition to nonspecific immunity in a marine shrimp: convergence of RNA interference and innate immunity in the invertebrate antiviral response? J Virol. 2005;79:13561–13571. doi: 10.1128/JVI.79.21.13561-13571.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Robalino J., Browdy C.L., Prior S., Metz A., Parnell P., Gross P. Induction of antiviral immunity by double-stranded RNA in a marine invertebrate. J Virol. 2004;78:10442–10448. doi: 10.1128/JVI.78.19.10442-10448.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Yodmuang S., Tirasophon W., Roshorm Y., Chinnirunvong W., Panyim S. YHV-protease dsRNA inhibits YHV replication in Penaeus monodon and prevents mortality. Biochem Biophys Res Commun. 2006;341:351–356. doi: 10.1016/j.bbrc.2005.12.186. [DOI] [PubMed] [Google Scholar]
- 106.Sarathi M., Simon M.C., Venkatesan C., Hameed A.S. Oral administration of bacterially expressed VP28dsRNA to protect Penaeus monodon from white spot syndrome virus. Mar Biotechnol (NY) 2008;10:242–249. doi: 10.1007/s10126-007-9057-6. [DOI] [PubMed] [Google Scholar]
- 107.Robalino J., Bartlett T.C., Chapman R.W., Gross P.S., Browdy C.L., Warr G.W. Double-stranded RNA and antiviral immunity in marine shrimp: inducible host mechanisms and evidence for the evolution of viral counter-responses. Dev Comp Immunol. 2007;31:539–547. doi: 10.1016/j.dci.2006.08.011. [DOI] [PubMed] [Google Scholar]
- 108.Flegel T.W., Nielsen L., Thamavit V., kongtim S., Pasharawipas T. Presence of multiple viruses in non-diseased, cultivated shrimp at harvest. Aquaculture. 2004;240:55–68. [Google Scholar]
- 109.Chayaburakul K., Nash G., Pratanpipat P., Sriurairatana S., Withyachumnarnkul B. Multiple pathogens found in growth retarded black tiger shrimp Penaeus monodon cultivated in Thailand. Dis Aquat Organ. 2004;60:89–96. doi: 10.3354/dao060089. [DOI] [PubMed] [Google Scholar]
- 110.Flegel T.W. Update on viral accommodation, a model for host-viral interaction in shrimp and other arthropods. Dev Comp Immunol. 2007;31:217–231. doi: 10.1016/j.dci.2006.06.009. [DOI] [PubMed] [Google Scholar]
- 111.Guidotti L.G., Chisari F.V. Noncytolytic control of viral infections by the innate and adaptive immune response. Annu Rev Immunol. 2001;19:65–91. doi: 10.1146/annurev.immunol.19.1.65. [DOI] [PubMed] [Google Scholar]
- 112.de Veer M.J., Holko M., Frevel M., Walker E., Der S., Paranjape J.M. Functional classification of interferon-stimulated genes identified using microarrays. J Leukoc Biol. 2001;69:912–920. [PubMed] [Google Scholar]
- 113.Ehrt S., Schnappinger D., Bekiranov S., Drenkow J., Shi S., Gingeras T.R. Reprogramming of the macrophage transcriptome in response to interferon-gamma and Mycobacterium tuberculosis: signaling roles of nitric oxide synthase-2 and phagocyte oxidase. J Exp Med. 2001;194:1123–1140. doi: 10.1084/jem.194.8.1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Katze M.G., He Y., Gale M., Jr. Viruses and interferon: a fight for supremacy. Nat Rev Immunol. 2002;2:675–687. doi: 10.1038/nri888. [DOI] [PubMed] [Google Scholar]
- 115.Lemaitre B., Hoffmann J. The host defense of Drosophila melanogaster. Annu Rev Immunol. 2007;25:697–743. doi: 10.1146/annurev.immunol.25.022106.141615. [DOI] [PubMed] [Google Scholar]
- 116.Dostert C., Jouanguy E., Irving P., Troxler L., Galiana-Arnoux D., Hetru C. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of Drosophila. Nat Immunol. 2005;6:946–953. doi: 10.1038/ni1237. [DOI] [PubMed] [Google Scholar]
- 117.Lin C.C., Chou C.M., Hsu Y.L., Lien J.C., Wang Y.M., Chen S.T. Characterization of two mosquito STATs, AaSTAT and CtSTAT. Differential regulation of tyrosine phosphorylation and DNA binding activity by lipopolysaccharide treatment and by Japanese encephalitis virus infection. J Biol Chem. 2004;279:3308–3317. doi: 10.1074/jbc.M309749200. [DOI] [PubMed] [Google Scholar]
- 118.Liu W.J., Chang Y.S., Wang A.H., Kou G.H., Lo C.F. White spot syndrome virus annexes a shrimp STAT to enhance expression of the immediate-early gene ie1. J Virol. 2007;81:1461–1471. doi: 10.1128/JVI.01880-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Chen W.Y., Ho K.C., Leu J.H., Liu K.F., Wang H.C., Kou G.H. WSSV infection activates STAT in shrimp. Dev Comp Immunol. 2008;32:1142–1150. doi: 10.1016/j.dci.2008.03.003. [DOI] [PubMed] [Google Scholar]
- 120.Liu W.J., Chang Y.S., Wang H.C., Leu J.H., Kou G.H., Lo C.F. Transactivation, dimerization, and DNA-binding activity of white spot syndrome virus immediate early protein IE1. J Virol. 2008 doi: 10.1128/JVI.01244-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Zambon R.A., Nandakumar M., Vakharia V.N., Wu L.P. The Toll pathway is important for an antiviral response in Drosophila. Proc Natl Acad Sci U S A. 2005;102:7257–7262. doi: 10.1073/pnas.0409181102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Arts J.A., Cornelissen F.H., Cijsouw T., Hermsen T., Savelkoul H.F., Stet R.J. Molecular cloning and expression of a Toll receptor in the giant tiger shrimp, Penaeus monodon. Fish Shellfish Immunol. 2007;23:504–513. doi: 10.1016/j.fsi.2006.08.018. [DOI] [PubMed] [Google Scholar]
- 123.Yang C., Zhang J., Li F., Ma H., Zhang Q., Jose Priya T.A. A Toll receptor from Chinese shrimp Fenneropenaeus chinensis is responsive to Vibrio anguillarum infection. Fish Shellfish Immunol. 2008;24:564–574. doi: 10.1016/j.fsi.2007.12.012. [DOI] [PubMed] [Google Scholar]
- 124.Bortner C.D., Oldenburg N.B., Cidlowski J.A. The role of DNA fragmentation in apoptosis. Trends Cell Biol. 1995;5:21–26. doi: 10.1016/s0962-8924(00)88932-1. [DOI] [PubMed] [Google Scholar]
- 125.Aubert M., Jerome K.R. Apoptosis prevention as a mechanism of immune evasion. Int Rev Immunol. 2003;22:361–371. doi: 10.1080/08830180305213. [DOI] [PubMed] [Google Scholar]
- 126.Sugumaran M., Nellaiappan K. Characterization of a new phenoloxidase inhibitor from the cuticle of Manduca sexta. Biochem Biophys Res Commun. 2000;268:379–383. doi: 10.1006/bbrc.2000.2138. [DOI] [PubMed] [Google Scholar]
- 127.Everett H., McFadden G. Apoptosis: an innate immune response to virus infection. Trends Microbiol. 1999;7:160–165. doi: 10.1016/s0966-842x(99)01487-0. [DOI] [PubMed] [Google Scholar]
- 128.Kalvakolanu D.V. Virus interception of cytokine-regulated pathways. Trends Microbiol. 1999;7:166–171. doi: 10.1016/s0966-842x(99)01476-6. [DOI] [PubMed] [Google Scholar]
- 129.Tortorella D., Gewurz B.E., Furman M.H., Schust D.J., Ploegh H.L. Viral subversion of the immune system. Annu Rev Immunol. 2000;18:861–926. doi: 10.1146/annurev.immunol.18.1.861. [DOI] [PubMed] [Google Scholar]
- 130.Teodoro J.G., Branton P.E. Regulation of apoptosis by viral gene products. J Virol. 1997;71:1739–1746. doi: 10.1128/jvi.71.3.1739-1746.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Granja C.B., Aranguren L.F., Vidal O.M., Aragón L., Salazar M. Does hyperthermia increase apoptosis in white spot syndrome virus (WSSV)-infected Litopenaeus vannamei? Dis Aquat Org. 2003;54:73–78. doi: 10.3354/dao054073. [DOI] [PubMed] [Google Scholar]
- 132.Sahtout A.H., Hassan M.D., Shariff M. DNA fragmentation, an indicator of apoptosis, in cultured black tiger shrimp Penaeus monodon infected with white spot syndrome virus (WSSV) Dis Aquat Org. 2001;44:155–159. doi: 10.3354/dao044155. [DOI] [PubMed] [Google Scholar]
- 133.Khanobdee K., Soowannayan C., Flegel T.W., Ubol S., Withyachumnarnkul B. Evidence for apoptosis correlated with mortality in the giant black tiger shrimp Penaeus monodon infected with yellow head virus. Dis Aquat Org. 2002;48:79–90. doi: 10.3354/dao048079. [DOI] [PubMed] [Google Scholar]
- 134.Thornberry N.A., Lazebnik Y. Caspases: enemies within. Science. 1998;281:1312–1316. doi: 10.1126/science.281.5381.1312. [DOI] [PubMed] [Google Scholar]
- 135.Wang L., Zhi B., Wu W., Zhang X. Requirement for shrimp caspase in apoptosis against virus infection. Dev Comp Immunol. 2008;32:706–715. doi: 10.1016/j.dci.2007.10.010. [DOI] [PubMed] [Google Scholar]
- 136.Anggraeni M.S., Owens L. The haemocytic origin of lymphoid organ spheroid cells in the penaeid prawn Penaeus monodon. Dis Aquat Org. 2000;40:85–92. doi: 10.3354/dao040085. [DOI] [PubMed] [Google Scholar]
- 137.Wu J.L., Muroga K. Apoptosis does not play an important role in the resistance of ‘immune’ Penaeus japonicus against white spot syndrome virus. J Fish Dis. 2004;27:15–21. doi: 10.1046/j.1365-2761.2003.00491.x. [DOI] [PubMed] [Google Scholar]
- 138.Rijiravanich A., Browdy C.L., Withyachumnarnkul B. Knocking down caspase-3 by RNAi reduces mortality in Pacific white shrimp Penaeus (Litopenaeus) vannamei challenged with a low dose of white-spot syndrome virus. Fish Shellfish Immunol. 2008;24:308–313. doi: 10.1016/j.fsi.2007.11.017. [DOI] [PubMed] [Google Scholar]
- 139.Molthathong S., Buaklin A., Senapin S., Klinbunga S., Rojtinnakorn J., Flegel T.W. Up-regulation of ribophorin I after yellow head virus (YHV) challenge in black tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 2008;25:40–46. doi: 10.1016/j.fsi.2007.12.003. [DOI] [PubMed] [Google Scholar]
- 140.Lindholm P., Kuittinen T., Sorri O., Guo D., Merits A., Tormakangas K. Glycosylation of phytepsin and expression of dad1, dad2 and ost1 during onset of cell death in germinating barley scutella. Mech Dev. 2000;93:169–173. doi: 10.1016/s0925-4773(00)00254-9. [DOI] [PubMed] [Google Scholar]
- 141.Nakashima T., Sekiguchi T., Kuraoka A., Fukushima K., Shibata Y., Komiyama S. Molecular cloning of a human cDNA encoding a novel protein, DAD1, whose defect causes apoptotic cell death in hamster BHK21 cells. Mol Cell Biol. 1993;13:6367–6374. doi: 10.1128/mcb.13.10.6367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Molthathong S., Senapin S., Klinbunga S., Puanglarp N., Rojtinnakorn J., Flegel T.W. Down-regulation of defender against apoptotic death (DAD1) after yellow head virus (YHV) challenge in black tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 2008;24:173–179. doi: 10.1016/j.fsi.2007.10.013. [DOI] [PubMed] [Google Scholar]
- 143.Bangrak P., Graidist P., Chotigeat W., Phongdara A. Molecular cloning and expression of a mammalian homologue of a translationally controlled tumor protein (TCTP) gene from Penaeus monodon shrimp. J Biotechnol. 2004;108:219–226. doi: 10.1016/j.jbiotec.2003.12.007. [DOI] [PubMed] [Google Scholar]
- 144.Molthathong S., Rojtinnakorn J., Senapin S., Flegel T.W. Hepatopancreatic nuclease of black tiger shrimp Penaeus monodon unlikely to be involved in viral triggered apoptosis. Fish Shellfish Immunol. 2007;22:617–627. doi: 10.1016/j.fsi.2006.08.008. [DOI] [PubMed] [Google Scholar]
- 145.Li L., Lin S., Yanga F. Functional identification of the non-specific nuclease from white spot syndrome virus. Virology. 2005;337:399–406. doi: 10.1016/j.virol.2005.04.017. [DOI] [PubMed] [Google Scholar]
- 146.Hammond S.M., Bernstein E., Beach D., Hannon G.J. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature. 2000;404:293–296. doi: 10.1038/35005107. [DOI] [PubMed] [Google Scholar]
- 147.Hammond S.M., Boettcher S., Caudy A.A., Kobayashi R., Hannon G.J. Argonaute2, a link between genetic and biochemical analyses of RNAi. Science. 2001;293:1146–1150. doi: 10.1126/science.1064023. [DOI] [PubMed] [Google Scholar]
- 148.Martinez J., Patkaniowska A., Urlaub H., Luhrmann R., Tuschl T. Single-stranded antisense siRNAs guide target RNA cleavage in RNAi. Cell. 2002;110:563–574. doi: 10.1016/s0092-8674(02)00908-x. [DOI] [PubMed] [Google Scholar]
- 149.Lecellier C.H., Voinnet O. RNA silencing: no mercy for viruses? Immunol Rev. 2004;198:285–303. doi: 10.1111/j.0105-2896.2004.00128.x. [DOI] [PubMed] [Google Scholar]
- 150.Galiana-Arnoux D., Dostert C., Schneemann A., Hoffmann J.A., Imler J.L. Essential function in vivo for Dicer-2 in host defense against RNA viruses in Drosophila. Nat Immunol. 2006;7:590–597. doi: 10.1038/ni1335. [DOI] [PubMed] [Google Scholar]
- 151.Wang X.H., Aliyari R., Li W.X., Li H.W., Kim K., Carthew R. RNA interference directs innate immunity against viruses in adult Drosophila. Science. 2006;312:452–454. doi: 10.1126/science.1125694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Zambon R.A., Vakharia V.N., Wu L.P. RNAi is an antiviral immune response against a dsRNA virus in Drosophila melanogaster. Cell Microbiol. 2006;8:880–889. doi: 10.1111/j.1462-5822.2006.00688.x. [DOI] [PubMed] [Google Scholar]
- 153.Li H., Li W.X., Ding S.W. Induction and suppression of RNA silencing by an animal virus. Science. 2002;296:1319–1321. doi: 10.1126/science.1070948. [DOI] [PubMed] [Google Scholar]
- 154.Keene K.M., Foy B.D., Sanchez-Vargas I., Beaty B.J., Blair C.D., Olson K.E. RNA interference acts as a natural antiviral response to O'nyong-nyong virus (Alphavirus; Togaviridae) infection of Anopheles gambiae. Proc Natl Acad Sci U S A. 2004;101:17240–17245. doi: 10.1073/pnas.0406983101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Lu R., Maduro M., Li F., Li H.W., Broitman-Maduro G., Li W.X. Animal virus replication and RNAi-mediated antiviral silencing in Caenorhabditis elegans. Nature. 2005;436:1040–1043. doi: 10.1038/nature03870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Wilkins C., Dishongh R., Moore S.C., Whitt M.A., Chow M., Machaca K. RNA interference is an antiviral defence mechanism in Caenorhabditis elegans. Nature. 2005;436:1044–1047. doi: 10.1038/nature03957. [DOI] [PubMed] [Google Scholar]
- 157.Bennasser Y., Le S.Y., Benkirane M., Jeang K.T. Evidence that HIV-1 encodes an siRNA and a suppressor of RNA silencing. Immunity. 2005;22:607–619. doi: 10.1016/j.immuni.2005.03.010. [DOI] [PubMed] [Google Scholar]
- 158.Cullen B.R. Is RNA interference involved in intrinsic antiviral immunity in mammals? Nat Immunol. 2006;7:563–567. doi: 10.1038/ni1352. [DOI] [PubMed] [Google Scholar]
- 159.Berkhout B., Jeang K.T. RISCy business: MicroRNAs, pathogenesis, and viruses. J Biol Chem. 2007;282:26641–26645. doi: 10.1074/jbc.R700023200. [DOI] [PubMed] [Google Scholar]
- 160.Kim C.S., Kosuke Z., Nam Y.K., Kim S.K., Kim K.H. Protection of shrimp (Penaeus chinensis) against white spot syndrome virus (WSSV) challenge by double-stranded RNA. Fish Shellfish Immunol. 2007;23:242–246. doi: 10.1016/j.fsi.2006.10.012. [DOI] [PubMed] [Google Scholar]
- 161.Xu J., Han F., Zhang X. Silencing shrimp white spot syndrome virus (WSSV) genes by siRNA. Antiviral Res. 2007;73:126–131. doi: 10.1016/j.antiviral.2006.08.007. [DOI] [PubMed] [Google Scholar]
- 162.Tirasophon W., Roshorm Y., Panyim S. Silencing of yellow head virus replication in penaeid shrimp cells by dsRNA. Biochem Biophys Res Commun. 2005;334:102–107. doi: 10.1016/j.bbrc.2005.06.063. [DOI] [PubMed] [Google Scholar]
- 163.Tirasophon W., Yodmuang S., Chinnirunvong W., Plongthongkum N., Panyim S. Therapeutic inhibition of yellow head virus multiplication in infected shrimps by YHV-protease dsRNA. Antiviral Res. 2007;74:150–155. doi: 10.1016/j.antiviral.2006.11.002. [DOI] [PubMed] [Google Scholar]
- 164.Jacobs B.L., Langland J.O. When two strands are better than one: the mediators and modulators of the cellular responses to double-stranded RNA. Virology. 1996;219:339–349. doi: 10.1006/viro.1996.0259. [DOI] [PubMed] [Google Scholar]
- 165.Alexopoulou L., Holt A.C., Medzhitov R., Flavell R.A. Recognition of double-stranded RNA and activation of NF-kappaB by toll-like receptor 3. Nature. 2001;413:732–738. doi: 10.1038/35099560. [DOI] [PubMed] [Google Scholar]
- 166.Oshiumi H., Matsumoto M., Funami K., Akazawa T., Seya T. TICAM-1, an adaptor molecule that participates in toll-like receptor 3-mediated interferon-beta induction. Nat Immunol. 2003;4:161–167. doi: 10.1038/ni886. [DOI] [PubMed] [Google Scholar]
- 167.Meurs E., Chong K., Galabru J., Thomas N.S., Kerr I.M., Williams B.R. Molecular cloning and characterization of the human double-stranded RNA-activated protein kinase induced by interferon. Cell. 1990;62:379–390. doi: 10.1016/0092-8674(90)90374-n. [DOI] [PubMed] [Google Scholar]
- 168.Gantier M.P., Williams B.R. The response of mammalian cells to double-stranded RNA. Cytokine Growth Factor Rev. 2007;18:363–371. doi: 10.1016/j.cytogfr.2007.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Adams M.D., Celniker S.E., Holt R.A., Evans C.A., Gocayne J.D., Amanatides P.G. The genome sequence of Drosophila melanogaster. Science. 2000;287:2185–2195. doi: 10.1126/science.287.5461.2185. [DOI] [PubMed] [Google Scholar]
- 170.Dehal P., Satou Y., Campbell R.K., Chapman J., Degnan B., De Tomaso A. The draft genome of Ciona intestinalis: insights into chordate and vertebrate origins. Science. 2002;298:2157–2167. doi: 10.1126/science.1080049. [DOI] [PubMed] [Google Scholar]
- 171.Holt R.A., Subramanian G.M., Halpern A., Sutton G.G., Charlab R., Nusskern D.R. The genome sequence of the malaria mosquito Anopheles gambiae. Science. 2002;298:129–149. doi: 10.1126/science.1076181. [DOI] [PubMed] [Google Scholar]
- 172.Westenberg M., Heinhuis B., Zuidema D., Vlak J.M. siRNA injection induces sequence-independent protection in Penaeus monodon against white spot syndrome virus. Virus Res. 2005;114:133–139. doi: 10.1016/j.virusres.2005.06.006. [DOI] [PubMed] [Google Scholar]
- 173.Haasnoot J., Westerhout E.M., Berkhout B. RNA interference against viruses: strike and counterstrike. Nat Biotechnol. 2007;25:1435–1443. doi: 10.1038/nbt1369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Matzke M.A., Birchler J.A. RNAi-mediated pathways in the nucleus. Nat Rev Genet. 2005;6:24–35. doi: 10.1038/nrg1500. [DOI] [PubMed] [Google Scholar]
- 175.Kim D.H., Villeneuve L.M., Morris K.V., Rossi J.J. Argonaute-1 directs siRNA-mediated transcriptional gene silencing in human cells. Nat Struct Mol Biol. 2006;13:793–797. doi: 10.1038/nsmb1142. [DOI] [PubMed] [Google Scholar]
- 176.Jiravanichpaisal P., Söderhäll K., Söderhäll I. Characterization of white spot syndrome virus replication in in vitro-cultured haematopoietic stem cells of freshwater crayfish, Pacifastacus leniusculus. J Gen Virol. 2006;87:847–854. doi: 10.1099/vir.0.81758-0. [DOI] [PubMed] [Google Scholar]
- 177.Liu H., Söderhäll I. Histone H2A as a transfection agent in crayfish hematopoietic tissue cells. Dev Comp Immunol. 2007;31:340–346. doi: 10.1016/j.dci.2006.07.007. [DOI] [PubMed] [Google Scholar]
- 178.Weber F., Wagner V., Rasmussen S.B., Hartmann R., Paludan S.R. Double-stranded RNA is produced by positive-strand RNA viruses and DNA viruses but not in detectable amounts by negative-strand RNA viruses. J Virol. 2006;80:5059–5064. doi: 10.1128/JVI.80.10.5059-5064.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Dechklar M., Udomkit A., Panyim S. Characterization of argonaute cDNA from Penaeus monodon and implication of its role in RNA interference. Biochem Biophys Res Commun. 2008;367:768–774. doi: 10.1016/j.bbrc.2008.01.031. [DOI] [PubMed] [Google Scholar]
- 180.Unajak S., Boonsaeng V., Jitrapakdee S. Isolation and characterization of cDNA encoding Argonaute, a component of RNA silencing in shrimp (Penaeus monodon) Comp Biochem Physiol B Biochem Mol Biol. 2006;145:179–187. doi: 10.1016/j.cbpb.2006.07.002. [DOI] [PubMed] [Google Scholar]
- 181.Su J., Oanh D.T., Lyons R.E., Leeton L., van Hulten M.C., Tan S.H. A key gene of the RNA interference pathway in the black tiger shrimp, Penaeus monodon: identification and functional characterisation of Dicer-1. Fish Shellfish Immunol. 2008;24:223–233. doi: 10.1016/j.fsi.2007.11.006. [DOI] [PubMed] [Google Scholar]
- 182.Arbouzova N.I., Zeidler M.P. JAK/STAT signalling in Drosophila: insights into conserved regulatory and cellular functions. Development. 2006;133:2605–2616. doi: 10.1242/dev.02411. [DOI] [PubMed] [Google Scholar]