

Graphical abstract
Keywords: Arterivirus, Equine arteritis virus, Porcine reproductive and respiratory syndrome virus, Signal peptide, Glycosylation, Membrane topology
Abbreviations: BHK, baby hamster kidney cells; BiP, binding immunoglobulin protein; BM2, M2 protein of influenza B; CD, cluster of differentiation; CHO, Chinese hamster ovary cells; DTT, dithiothreitol; EAV, equine arteritis virus; Endo H, endoglycosidase H; ER, endoplasmic reticulum; ERGIC, ER–Golgi intermediate compartment; ESCRT, endosomal sorting complexes required for transport; GFP, green fluorescent protein; Gp, glycoprotein; GPI, glycosylphosphatidylinositol; HIV, human immunodeficiency virus; HR, hydrophobic region; LC–MS/MS, liquid chromatography–tandem mass spectrometry; LDV, murine lactate dehydrogenase-elevating virus; MHC, major histocompatibility complex; ORF, open reading frame; OST, oligosaccharyltransferase; PAMs, porcine alveolar macrophages; PFU, plaque-forming unit; PI-PLC, phosphatidylinositol-specific phospholipase C; PNGase F, peptide:N-glycosidase F; PRRSV, porcine reproductive and respiratory syndrome virus; SARS, severe acute respiratory syndrome; SHFV, simian haemorrhagic fever virus; SPase, signal peptidase; SP, signal peptide; SRP, signal recognition particle; TMR, transmembrane region; VLP, virus-like particle; YFP, yellow fluorescent protein
Highlights
-
•
Arteriviruses are important pathogens in veterinary medicine.
-
•
We review the structure and processing of their membrane proteins.
-
•
Some features are unique from a cell biological point of view.
-
•
New data on this topic are also presented.
-
•
We speculate on the role of the membrane proteins during virus entry and budding.
Abstract
Arteriviruses, such as equine arteritis virus (EAV) and porcine reproductive and respiratory syndrome virus (PRRSV), are important pathogens in veterinary medicine. Despite their limited genome size, arterivirus particles contain a multitude of membrane proteins, the Gp5/M and the Gp2/3/4 complex, the small and hydrophobic E protein and the ORF5a protein. Their function during virus entry and budding is understood only incompletely.
We summarize current knowledge of their primary structure, membrane topology, (co-translational) processing and intracellular targeting to membranes of the exocytic pathway, which are the budding site. We profoundly describe experimental data that led to widely believed conceptions about the function of these proteins and also report new results about processing steps for each glycoprotein. Further, we depict the location and characteristics of epitopes in the membrane proteins since the late appearance of neutralizing antibodies may lead to persistence, a characteristic hallmark of arterivirus infection. Some molecular features of the arteriviral proteins are rare or even unique from a cell biological point of view, particularly the prevention of signal peptide cleavage by co-translational glycosylation, discovered in EAV-Gp3, and the efficient use of overlapping sequons for glycosylation. This article reviews the molecular mechanisms of these cellular processes. Based on this, we present hypotheses on the structure and variability of arteriviral membrane proteins and their role during virus entry and budding.
1. Introduction to arteriviruses
Arteriviridae is a family of enveloped, positive-stranded RNA viruses. Despite their importance in veterinary medicine, the arteriviruses are only poorly characterized in molecular terms. The prototype arterivirus is equine arteritis virus (EAV), which can cause substantial disease in horses; further arteriviruses are porcine reproductive and respiratory syndrome virus (PRRSV), the most important pathogen in the pig industry worldwide, the murine lactate dehydrogenase-elevating virus (LDV) and simian haemorrhagic fever virus (SHFV). To date, no arterivirus affecting humans has been encountered (for recent review, see Balasuriya et al., 2013, Meulenberg, 2000, Snijder et al., 2013).
Arterivirus infection may be subclinical (especially in the case of LDV), but can lead to severe symptoms, most prominently lesions of arteries (arteritis, hence the name of the virus family), oedema, respiratory symptoms/pneumonia as well as abortion in pregnant animals, with devastating implications in animal breeding (for review, see Cho and Dee, 2006, Nodelijk, 2002, Rossow, 1998). The primary target cells for arteriviruses are macrophages, but highly pathogenic PRRSV isolates may have an expanded tropism to include epithelial cells (Zhou and Yang, 2010). Transmission is mainly via the respiratory route; sexual transmission occurs as well (Cho and Dee, 2006, Nodelijk, 2002, Rossow, 1998).
One common, relevant trait of arterivirus infection is persistence. After acute infection, the virus is often not eliminated entirely, but continues to replicate at low levels in lymphoid tissues (PRRSV) or in the reproductive tract (EAV). It is generally assumed that the host's immune system is incapable of setting up a robust immune response against the virus. This is evidenced by the finding that neutralizing antibodies against PRRSV and LDV are generated only late after infection, and that their titres stay low. Several hypotheses about the mechanistic basis for persistence have been put forward (for review, see Chand et al., 2012, Darwich et al., 2010, Kimman et al., 2009, Mateu and Diaz, 2008, Murtaugh and Genzow, 2011).
This article will focus on the biochemical processing of the structural proteins of arteriviruses, with emphasis on EAV and PRRSV. We believe that understanding the biochemical properties of the viral proteins is highly relevant, not least in order to develop vaccines and treatments for the arteriviral diseases in a knowledge-based manner. In addition, some molecular features of the arteriviral proteins are very interesting from a cell biological point of view since they have been found to be unusual or even unique. For background information on the cellular biology of synthesis and processing of membrane proteins and the molecular features of the enzymes and proteins involved, the reader is referred to Addendum. Upon their first appearance in the main text, terms related to these topics are highlighted in italics and bold to refer the reader to Addendum.
1.1. The structural proteins of arteriviruses: overview
Due to similarities in genome organization and replication strategy, Arteriviridae are grouped in the order Nidovirales together with Coronaviridae, Roniviridae and Mesoniviridae. The biochemical properties of the structural proteins are, however, markedly different in Arteriviridae; yet, all Nidovirales share the property of possessing a nested genome, where the structural proteins are expressed from a set of 3′ co-terminal subgenomic mRNAs (Fang and Snijder, 2010, Gorbalenya et al., 2006). The open reading frames encoding the structural proteins are partially overlapping. Accordingly, one needs to keep in mind that a given amino acid context in a structural protein may actually be a by-product of evolutionary pressure on a different protein, encoded in part by the same RNA sequence. Also, this feature restricts the possibility to address virus protein mutations in recombinant viruses without affecting the overlapping ORFs.
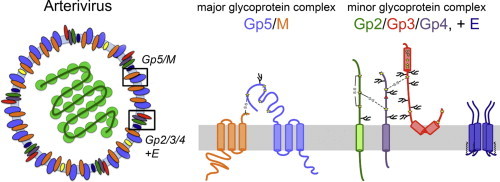
Arteriviruses contain at least seven structural proteins (Fig. 1 ; Music and Gagnon, 2010). The nucleocapsid protein N builds the scaffold for the genomic RNA, and several membrane proteins are incorporated into the viral envelope: the glycoproteins Gp2, Gp3, Gp4 and Gp5 as well as the non-glycosylated membrane proteins M and E. Gp2/3/4 form a disulphide-linked heterotrimeric complex in virus particles, and Gp5 is disulphide-linked to M. N, M and Gp5 are major virion components, whereas E, Gp2, Gp3 and Gp4 are referred to as minor structural proteins (de Vries et al., 1992). A novel membrane-anchored protein encoded by an alternative open reading frame of the subgenomic mRNA encoding Gp5 was discovered recently. This ORF5a protein is incorporated into virus particles, but probably as a very minor component (Firth et al., 2011, Johnson et al., 2011). Yet, it is relevant for replication: PRRSV does not replicate in the absence of the ORF5a protein, whereas recombinant EAV lacking the ORF5a protein could be generated, but grew to low titres and displayed a tiny plaque size (Firth et al., 2011, Sun et al., 2013). Very recently, the non-structural protein nsp2 was shown to be incorporated into virions of a variety of PRRSV strains (Kappes et al., 2013). This protein is predicted to have several transmembrane-spanning regions and assists in the induction of double membrane vesicles, ER-derived membrane scaffolds forming the viral replication and transcription complex (Snijder et al., 2001). The role of nsp2 in virions is not yet understood.
Fig. 1.
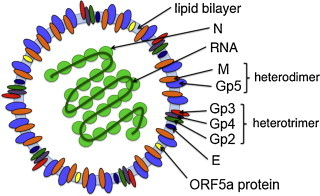
Scheme of an arterivirus particle. The membrane contains the major Gp5/M complex (blue and orange), the minor glycoprotein complex Gp2/3/4 (green, red and purple, respectively), the small hydrophobic E protein (dark blue) and the ORF5a protein (yellow).
From reverse genetics experiments with EAV and to some extent PRRSV, it is known that all structural proteins of either virus are essential for virus replication. If either Gp5 or M is deleted from the viral genome, no virus particles are released from infected cells. Thus, Gp5 and M are required for virus budding (which does not exclude additional functions, e.g. during virus entry). If expression of either Gp2 or Gp3 or Gp4 is abrogated, virus-like particles bud from cells, but the particles are not infectious, indicating that cell entry is disturbed in the absence of the minor glycoprotein complex (Wieringa et al., 2004, Wissink et al., 2005). Furthermore, the Gp2/3/4 complex governs cell tropism: When the ectodomains of EAV and PRRSV are swapped, the cell tropism of the resulting recombinant virus is altered, but not by exchanging the ectodomains of Gp5 and M (Dobbe et al., 2001, Lee and Yoo, 2006, Tian et al., 2012, Verheije et al., 2002). Interestingly, the Gp2/3/4 complex interacts either physically or functionally with E. Removal of E from the viral genome completely prevents incorporation of the Gp2/3/4 complex into virus particles, whereas in the absence of either Gp2, Gp3 or Gp4, the amount of E in virions is greatly reduced (Wieringa et al., 2004). Another peculiarity of the Gp2/3/4 complex is that Gp3 appears to be linked to the pre-formed Gp2/4 dimer only after virus budding (Wieringa et al., 2003b).
1.2. Virus particles, budding and cell entry of arteriviruses
Arterivirus particles are pleomorphic, but typically spherical or oval with a diameter of approx. 60 nm and a relatively smooth outer surface (for review, see Dokland, 2010, Snijder and Meulenberg, 1998). The nucleocapsid – comprising the viral RNA wrapped by the N protein – is arranged as a double-layered, spherical hollow core, separated from the lipid envelope by a gap of 2–3 nm as revealed by cryo-electron tomography (Spilman et al., 2009). The arterivirus particles are produced intracellularly; no budding is observed at the plasma membrane (Costers et al., 2006, Mardassi et al., 1994, Wada et al., 1995). According to various electron microscopy studies, virus particle generation proceeds by budding of pre-formed nucleocapsids into the lumen of the smooth endoplasmic reticulum (ER) and/or the Golgi apparatus (Pol et al., 1997, Snijder and Meulenberg, 1998, Wada et al., 1995, Wood et al., 1970). In analogy to the distantly related coronaviruses (Klumperman et al., 1994, Stertz et al., 2007), the ERGIC – the ER–Golgi intermediate compartment – could be the budding compartment, which has, however, not been directly shown for arteriviruses. After budding of the virions, they are transported along the secretory route and ultimately released by exocytosis.
PRRSV is the only arterivirus for which the entry mediators have been characterized to some extent. Initial contacts between the cell and the virus are mediated by binding of M to heparan sulphate proteoglycans, ubiquitous cellular membrane proteins. Sialic acid residues on Gp5 are then recognized by sialoadhesin (CD169), a lectin present only on macrophages, the primary target cells for arteriviruses. Sialoadhesin is supposed to be required for clathrin-dependent endocytosis of virus particles and their vesicular transport to early endosomes (for review, see Van Breedam et al., 2010). In an elegant biochemical study, it was shown that upon incubation with virus particles, purified sialoadhesin binds to Gp5, but not to Gp3 and Gp4, which, however, are also sialylated. The difference in the specificity was attributed to differences in the type of linkage between sialic acid and the following sugar residue and to a multivalency effect (binding of sialoadhesin to the glycoprotein that is more abundant on the virion surface, i.e. Gp5) (Van Breedam et al., 2010). However, the essential role of sialoadhesin for virus entry is controversial, since cells and even transgenic pigs lacking sialoadhesin are efficiently infected and support virus replication (Prather et al., 2013). Thus, sialoadhesin may be a non-essential attachment factor for PRRSV to bind and concentrate virions on the macrophage surface, hence allowing a more efficient infection.
In the endosome, the Gp2/3/4 complex binds to CD163, a scavenger receptor for haemoglobin clearance. CD163 appears to be the most specific receptor for PRRSV entry (for review, see Van Gorp et al., 2010, Welch and Calvert, 2010). The essential role of CD163 has been demonstrated most convincingly by rendering non-permissive cells susceptible to PRRSV infection upon transfection with CD163 (Calvert et al., 2007, Lee et al., 2010, Van Gorp et al., 2008). Interestingly, the host range specificity of PRRSV is not encoded in the cellular receptor, since CD163 and CD169 homologues from various mammalian species, including humans, can functionally replace the porcine receptors (Calvert et al., 2007, Van Breedam et al., 2013).
After receptor-mediated endocytosis of virus particles, a drop in endosomal pH, probably accompanied by the proteolytic activity of cathepsin E (Misinzo et al., 2008), activates the membrane fusion machinery of PRRSV (Van Breedam et al., 2010). However, it has not been identified so far for any of the arteriviruses what the target of cathepsin E is and which of the viral spikes mediates membrane fusion.
1.3. Virus replication in infected animals and immune response
In pigs, PRRSV replicates in porcine alveolar macrophages (PAMs) and other cells of the monocyte/macrophage lineage, which disseminate the virus to the blood. Highly pathogenic PRRSV isolates may possess an expanded tropism to include epithelial cells (Frydas et al., 2013, Zhou and Yang, 2010). Clinical symptoms, appearing early after infection, are mainly respiratory illness in growing pigs, whereas in pregnant sows, infection often leads to reproductive failure (Rossow, 1998, Zimmerman et al., 1997). Viraemia is sustained for up to 4 weeks after infection. Beyond that phase, however, the virus is typically not cleared from the body, but persists due to continuous low levels of replication, predominantly in lymphoid tissues, and continues to be shed (Chand et al., 2012). The molecular cause of persistence, one main obstacle to eliminate PRRSV from pig farms, is unclear. It is generally assumed that the host's immune system is incapable of setting up a robust immune response against the virus, leading to this persistence phenomenon (Mateu and Diaz, 2008). There is a strong antibody response directed against N and Gp5 few days after infection, yet these antibodies do not neutralize the virus. Neutralizing antibodies, however, appear only late, after more than 4 weeks after infection (Yoon et al., 1994). Their appearance coincides with clearing of virus from blood. The importance of neutralizing antibodies is reflected by the findings that serum from convalescent pigs (Lopez et al., 2007) and passive transfer of neutralizing antibodies (Osorio et al., 2002) both provide protection to homologous challenge and clear virus from blood. However, it is not universally accepted that neutralizing antibodies are crucial for immunity. Viraemia is often resolved before neutralizing antibodies are detected (Murtaugh and Genzow, 2011).
Several hypotheses about the mechanistic basis for persistence have been put forward (for review, see Kimman et al., 2009, Mateu and Diaz, 2008, Murtaugh and Genzow, 2011). One such hypothesis, important with regard to the viral membrane proteins, is the “decoy epitope” hypothesis (Lopez and Osorio, 2004), which predicts that a non-neutralizing, immunodominant epitope is present in Gp5, which decreases the induction or reactivity of antibodies against a nearby neutralizing epitope as described in more detail below.
Glycoproteins of PRRSV, especially Gp5 and Gp3, are the most variable proteins of any known viruses (Meng, 2000, Shi et al., 2010, Stadejek et al., 2013). Besides persistence of PRRSV, the variability of its antigens poses the biggest challenge for the successful control and elimination of the disease. Distinct PRRSV genotypes were identified in Europe (type 1, prototype: Lelystad virus) and North America (type 2, reference strain: VR-2332) in the early 1990s (Benfield et al., 1992, Wensvoort et al., 1991), but, meanwhile, the viruses have spread worldwide. Furthermore, new viruses continuously emerge, such as the highly virulent, type 2-related PRRSV in China (Zhou and Yang, 2010) and the virulent Lena strain in Eastern Europe (Karniychuk et al., 2010). The very low amino acid identity between glycoproteins may explain why the currently available vaccines protect against infections with homologous, but not heterologous strains.
Despite many efforts, it is still controversial against which surface protein(s) of PRRSV neutralizing antibodies are directed. Gp5 was favoured as the most relevant antigen (Lopez and Osorio, 2004), but a recent study described that a neutralizing pig serum that was affinity-purified with Gp5 did not neutralize infectivity of the virus (Li and Murtaugh, 2012). Neutralizing epitopes were also described and mapped in other PRRSV proteins, notably M (Yang et al., 2000), Gp3 (Vu et al., 2011) and – at least for the European subtype – Gp4 (Vanhee et al., 2010). However, epitopes were mostly determined by Pepscan analysis and phage display, peptide-based approaches that are error-prone and do not detect conformation-dependent antigens, which may arise by glycosylation, folding and oligomerization of proteins.
2. Membrane proteins of arteriviruses
2.1. The Gp5/M complex
M (∼160–170 amino acids, predicted MW ∼18 kDa) is the most conserved membrane protein of arteriviruses. It consists of a short ectodomain (15–17 amino acids), three putative transmembrane regions (20 amino acids each), which are connected by two short loops (∼6–9 amino acids), and a long, hydrophilic cytoplasmic tail (∼70–80 amino acids) (de Vries et al., 1992). The short N-terminal ectodomain does not contain a signal peptide or a consensus sequence for the attachment of N-linked glycans (Table 1 , Fig. 2 ). However, a motif for cotranslational attachment of myristic acid to the N-terminal glycine residue (consensus sequence GXXXS/T) is present in M of EAV (MGAIDSFCGD…). This site is predicted with high confidence to be myristoylated by a usually reliable bioinformatics tool (http://web.expasy.org/myristoylator/). Nevertheless, the acylation site is apparently not used as demonstrated by labelling experiments with [3H]-myristate (de Vries et al., 1995a and own unpublished results). In accordance, the N-termini of M from PRRSV of both the European (Lelystad: MGGLDDFCND…) and the American genotype (VR-2332: MGSSLDDFCH…) are not predicted to be myristoylated. Both retain the glycine residues, but the essential serine at position 6 is replaced by a large amino acid (D), which is not recognized by the N-myristoyltransferase (Bhatnagar et al., 1999).
Table 1.
N-terminal amino acid sequence of M from EAV and PRRSV.

Sequence alignment of M from PRRSV type 2 (VR-2332 strain, accession number AAD12130.1), PRRSV type 1 (strain Lelystad, Q04565) and EAV (strain Bucyrus, P28991). The unused myristoylation motif GXXXS/T present in M of EAV is in bold and underlined, the only cysteine linking M to Gp5 is in bold and charged amino acids delimiting the first and the beginning of the second transmembrane region (TMR) are shaded in grey.
Fig. 2.
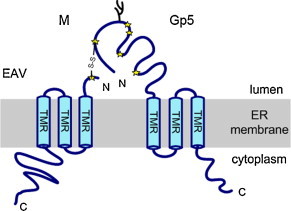
Scheme of the Gp5/M dimer of EAV. The N- and C-terminus of each protein is indicated. S—S indicates the disulphide bond between cysteine 8 of M and cysteine 27 in Gp5. Cysteine residues within the ectodomains are indicated as stars and glycans as branches. For the location of glycosylation sites in type 1 and 2 PRSSV strains, see Table 3, Table 4. TMR: putative transmembrane regions. For a different membrane topology of Gp5, see Table 2 and Fig. 3.
The only known modification of the ectodomain of M is the formation of a disulphide linkage with Gp5, as reported for EAV (de Vries et al., 1995a). This occurs through the single cysteine residue at position 8 of the ectodomain of M (Snijder et al., 2003), which is also present in M of PRRSV strains. Heterodimerization of M with Gp5 is required for transport of Gp5 and M from the ER to the Golgi apparatus, indicating that only properly assembled Gp5/M complexes can pass the quality control system of the ER. As a consequence, only Gp5/M heterodimers are incorporated into virus particles, and mutagenesis of cysteine 8 in M completely abolishes the release of infectious virus particles (Snijder et al., 2003). Folding and oligomerization of the Gp5/M complex seems to be a complicated and regulated process. Gp5 monomers rapidly form a disulphide linkage with M and virtually all Gp5 monomers end up in heterodimers, the association of M with Gp5 however proceeds at a slower rate, and a fraction of the M protein persists in cells as monomers. The M protein also forms covalently linked homodimers, which are either dead-end products or a mandatory step in the build-up of a functional Gp5/M spike (de Vries et al., 1995a).
Gp5 (previously termed GL, L for large) encompasses ∼200 amino acids in PRRSV, but 255 in EAV. Sequence alignment shows that the inserted amino acids are likely to be located in the ectodomain of Gp5. Gp5 comprises an N-terminal cleavable signal peptide directing protein synthesis to the rough ER, followed by an ectodomain of roughly 30 amino acids in PRRSV and 90 amino acids in EAV, containing at least one (EAV) or several (PRRSV) N-glycosylation sites. The region between residues 65 and 127 (PRRSV) or between 115 and 180 (EAV) is hydrophobic and assumed to span the membrane three times. The C-terminal part (128–200 in PRRSV, 181–255 in EAV) is hydrophilic and most probably located in the cytosol, ending up in the virus interior (Fig. 2). In PRRSV, Gp5 is the most variable structural protein with only ∼50% amino acid homology between North American and European isolates (Meng, 2000, Music and Gagnon, 2010).
2.1.1. Membrane topology of Gp5
Although Gp5 is usually depicted to span the membrane three times, the membrane topology of Gp5 has never been investigated experimentally, and bioinformatic tools do not always predict three TMRs for the Gp5 proteins of the various arteriviruses (Dokland, 2010).
Only the last part of the hydrophobic region of PRRSV-Gp5 complies with the properties of typical transmembrane regions, where a hydrophobic sequence of ∼20 amino acids is flanked on each side by charged residues (here, arginine, sequence RYVLSSIYAVCALAALTCFVIR, residues 105–126 in PRRSV VR-2332, Table 2 ). The preceding part of the hydrophobic region is ∼40 amino acids long (SFVIFPVLTHIVSYGALTTSHFLDTVALVTVSTAGFVHGR, residues 66–105 in PRRSV VR-2332) and thus could in principle span the membrane yet another two times. However, it is conceivable that this hydrophobic stretch rather forms a hairpin loop that does not fully transverse the membrane. In accordance with this, there is a conserved proline residue, which may induce a kink in the TMR's α-helix that will cause it to run back from the interior of the membrane to the lumen of the ER (Fig. 3 ). Furthermore, there is a charged residue (D or E in PRRSV or EAV, respectively) 15 residues from the end of this hydrophobic region, which is untypical for a transmembrane stretch. This charged residue may instead be exposed to the hydrophilic phase. In that case, the following stretch (DTVALVTVSTAGFVHGR, residues 89–105 in PRRSV VR-2332) would be exposed to the ER lumen (hence, the virus exterior in the mature virion).
Table 2.
Hydrophobic region of Gp5 from PRRSV, EAV and LDV.

Charged residues are shaded grey, the conserved proline is in bold and underlined. PRRSV 2: PRRSV strain VR-2332 (residues 65–126, accession number AAD12129.1); PRRSV 1: PRRSV strain Lelystad (67–128, Q04569); EAV: EAV strain Bucyrus (122–182, P28995); LDV: lactate dehydrogenase-elevating virus (65–126, NP_042577.1). See blue cylinders (labelled HR and TMR) in Fig. 3 for location of amino acids within the structure of Gp5.
Fig. 3.
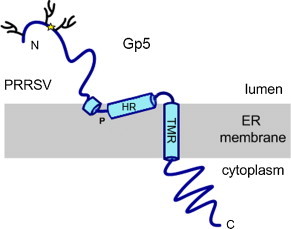
Speculation on an alternative membrane topology of Gp5. The first and second hydrophobic region may not cross the membrane completely (as depicted in Fig. 2), but form a hairpin structure. The kink might be induced by a conserved proline residue (Table 2), which is not compatible with the typical α-helical structure of transmembrane regions.
2.1.2. Antibody epitopes in Gp5
Generally, Gp5 is assumed to be the most relevant antigen for neutralizing antibodies in PRRSV (Lopez and Osorio, 2004), EAV (Balasuriya et al., 1997) and LDV (Plagemann, 2001).
Two epitopes have been mapped for Gp5 of type 2 PRRSV (reference strain: VR-2332), only one of which is neutralizing. Epitope A, encompassing residues 27–31, elicits an early and strong, but non-neutralizing immune response, while Epitope B (residues 37–44) is neutralizing, but less immunogenic. Antibodies directed against Epitope B appear only late in infection. Based on these findings, the “decoy hypothesis” was put forward to explain persistence of the virus (Lopez and Osorio). According to this hypothesis, the immunodominant Epitope A serves as a “decoy” to delay recognition of Epitope B. The region of Epitope A also harbours the predicted signal peptide cleavage sites and a variable number of potential N-glycosylation sites (Table 3 ). At the time of publication of the “decoy hypothesis”, it was not known whether and where the signal peptide is cleaved and whether the “decoy epitope” is present in virus particles, thus whether the molecular requirements for the hypothesis are fulfilled.
Table 3.
N-terminal amino acid sequence of Gp5 from various type 2 PRRSV strains.

The signal peptide is shaded in grey, either dark grey (cleavage of signal peptide at site 1), or light grey (cleavage at site 2). Epitope A (immunodominant, but non-neutralizing) and epitope B (neutralizing) are underlined and separated by a hypervariable region. Glycosylation sites are in bold. Accession numbers: VR-2332: ABB18278, Ingelvac (modified live vaccine): ABO87446, IAF Klop (strain from Quebec): AAC41206, HUB (isolate from China): ABW88233. See Fig. 3 for location of amino acids within the structure of Gp5.
We addressed this question and could show that the signal peptide of Gp5 from a selection of type 2 strains is cleaved, both during in vitro translation in the presence of microsomes and in transfected cells, and independently of neighbouring glycosylation sites. The exact signal peptide cleavage site in Gp5 of the reference strain VR-2332 was elucidated by mass spectrometry of recombinant and virus-derived protein. The results revealed that the signal peptide of Gp5 can be cleaved at two sites, between alanine 26 and valine 27 (site 1), or between alanine 31 and serine 32 (site 2). As a result, a mixture of Gp5 proteins exists in virus particles, some of which (cleaved at site 1) still contain the “decoy epitope” sequence. The mass spectrometry data are not quantitative, but SDS-PAGE migration patterns suggest that the majority of Gp5 molecules in transfected cells is cleaved at site 2 (Thaa et al., 2013). In accordance, sequencing the N-terminus of Gp5 from purified PRSSV particles revealed that the first residue of mature Gp5 is serine 32 (i.e. signal peptide cleavage at site 2) (Robinson et al., 2013a). However, sequencing is less sensitive than mass spectrometry, and therefore less abundant N-termini of Gp5 would not have been detected by this method. In summary, the “decoy epitope”-containing fraction of Gp5 in virions is probably small, but it needs to be tested whether it plays a role for immune evasion of the virus. The cellular fate of the cleaved signal peptides of Gp5 – most of which arise from cleavage at site 2 and thus contain the “decoy epitope” sequence – has however not been determined. They might be processed to be presented on either MHC class I or class II to fool the immune system of the host, similar to what has been shown for signal peptides from other virus families (Hegde and Bernstein, 2006, Martoglio and Dobberstein, 1998).
Intriguingly, the position of the neutralizing epitope in Gp5 appears not to be conserved between the PRRSV genotypes. For Gp5 of PRRSV genotype 1 only one neutralizing epitope was described for the strain Intervet-10 (I-10) using monoclonal antibodies from immunized mice (Weiland et al., 1999). Surprisingly, the antibodies did not neutralize the infectivity of the closely related Lelystad strain. This is due to a single amino acid difference (P→C) located in the signal peptide of Gp5 (Table 4 ) – yet, the epitope does not encompass this residue, but is located further downstream, right at the border between the predicted signal peptide and the ectodomain. It was speculated, but not shown, that the amino acid exchange affects the site of signal peptide cleavage, such that the epitope is present in the mature form of Gp5 from I-10, but not from Lelystad (Wissink et al., 2003). It is also an open question whether such monoclonal antibodies are also generated during a natural infection of pigs.
Table 4.
N-terminal amino acid sequence of Gp5 from type 1 PRRSV strains.

The (predicted) signal peptide sequence is shaded in grey. Glycosylation sites are in bold. The neutralizing epitope is underlined and bold. + and − indicate whether the respective strain is neutralized by a monoclonal antibody. The only amino acid difference in Gp5 between Lelystad and I-10 is in italic and underlined. Sequences are from Wissink et al. (2003).
2.1.3. N-glycosylation of Gp5
Gp5 (as well as the other arterivirus glycoproteins) contains only N-linked glycans; O-glycosylation at serine or threonine residues has not been described. The exact structure of the carbohydrate side chains (i.e. which specific sugars are attached to each glycosylation site and by which type of linkage) has not been determined for Gp5 or any other arterivirus glycoprotein. From digestion experiments with glycosidases it is known that (at least some) side chains are processed to an Endo-H resistant, complex form that contains terminal sialic acid (de Vries et al., 1995a, Delputte and Nauwynck, 2004). In addition, Gp5 (at least from EAV and LDV) contains repeating lactosamine units within the N-linked side chain, i.e. N-acetyl-glucosamine linked by a β1–4 linkage to galactose (de Vries et al., 1992, Li et al., 1998), a rather unusual modification of a viral glycoprotein. Attachment of lactosamine units usually occurs in the trans-Golgi cisternae and/or the trans-Golgi network, their number is variable (causing Gp5 to run as a smear in SDS-PAGE) and probably cell type-dependent. The role of polylactosamine glycans for the biology of arteriviruses is unknown, but potentially important, since polylactosamine on (at least cellular) glycoproteins influences basal levels of lymphocyte and macrophage activation (Togayachi et al., 2007).
The ectodomain of Gp5 from PRRSV contains a variable number of N-glycosylation sites. Two of these (N46 and N53 in type 1, N44 and N51 in type 2) are highly conserved. The relevance of the glycosylation sites for replication of PRRSV strains in cell culture has been addressed in several studies. In the Lelystad strain, removal of the site N53 had no effect on virus replication, whereas exchange of N46 strongly reduced the release of virus particles, which also had a lower specific infectivity (Wissink et al., 2004). A very similar effect regarding the significance of individual conserved glycosylation sites was reported for the American strain FL-12. Mutations involving N44 did not allow the production of infectious virus particles, whereas viruses with removal of N51 in Gp5 grew to lower titres (Ansari et al., 2006). In contrast, a recent study with the type 2 strain JXM 100 demonstrated that removal of the conserved glycosylation sites did not affect virus replication in cell culture. Only if two sites were exchanged simultaneously was the virus yield reduced (Wei et al., 2012a). In EAV the single conserved glycosylation site in Gp5 is essential for virus replication; yet, the virus tolerates removal of a large fragment of Gp5's ectodomain (Snijder et al., 2003).
Besides the two conserved glycosylation sites, Gp5 of PRRSV contains a number of variable potential glycosylation sites that are located in a hypervariable region at the border between the signal peptide and the ectodomain. This region is rich in serine and asparagine codons (AGC and AAC, respectively), and single nucleotide exchanges can easily lead to the addition or loss of N-glycosylation sites. Typically, 0–3 sites are present.
Addition or loss of glycosylation sites occurs also during infection of pigs. Serial passaging of VR-2332 virus between pigs resulted in numerous mutations in Gp5, including the changes N33S and D34N, removing and adding potential glycosylation sites, respectively (Kwon et al., 2008). The mutation D34N was also observed during the course of infection when the viral sequences in individual pigs were followed after experimental infection with PRRSV VR-2332 for 132 days (Chang et al., 2002). In addition, a PRRSV virus isolate with deleted glycosylation site N51 in Gp5 (and also one glycosylation site in Gp3) regained both sites after infection of piglets (Vu et al., 2011).
Using expression of Gp5 from PRRSV VR-2332, it was found that N30 does not contain carbohydrates in the mature protein, whereas N33 and also a site introduced at position N34 (leading to the overlapping sequon NNSS) are both glycosylated (Thaa et al., 2013). This is surprising since overlapping sequons are generally not modified at both asparagines (Karamyshev et al., 2005). When Gp5 present in virus particles was analysed by mass spectrometry, microheterogeneity was observed regarding the use of the glycosylation sites in the hypervariable region, not all the potential sites were used in every molecule (Thaa et al., 2013). Regarding the functional role of the variable glycosylation sites, it was reported that the carbohydrate attached to position N34 was required for efficient virus replication of one type 2 strain (Ansari et al., 2006), but that the adjacent site N35 had no effect on virus replication of another type 2 strain (Wei et al., 2012a). In the latter case the amino acid introduced for N34 also influenced the growth properties of recombinant virus particles.
In summary, the effects of N-glycosylation of Gp5 can hardly be generalized; there seems to be a certain flexibility in the number and position of glycans. Although not specifically addressed experimentally in any of the studies, it is likely that the co-translational attachment of carbohydrates affects folding of the polypeptide chain. Improperly folded proteins may not associate with M and/or will not pass the quality control system of the ER and will thus be degraded and cannot contribute to formation of virus particles. This is particularly relevant for the conserved glycosylation sites. Conversely, a minor fraction of Gp5 molecules may fold into the “right” conformation even in the absence of a given glycosylation site, as protein folding is a stochastic (probability-driven) process.
Besides its effect on protein folding, glycosylation is also supposed to affect the antigenicity of Gp5. This is generally referred to as “glycan shielding” as the presence of glycans appears to reduce the capacity of antibodies to neutralize PRRSV. Indeed, recombinant viral mutants with deleted glycosylation sites at N34, N51 and N34 + 51 exhibited enhanced sensitivity to neutralization by antibodies that had been raised against wild type PRRSV. In addition, these viruses also elicited a stronger neutralizing antibody response in infected piglets compared to wild type virus (Wei et al., 2012a). Likewise, a type 2 PRRSV field isolate lacking the conserved glycosylation site N51 in Gp5 (and also one glycosylation site in Gp3) raised a stronger antibody response than wild type (Vu et al., 2011). In another study, antibodies to the neutralization epitope were generated more rapidly when pigs were infected with a strain lacking the variable glycosylation sites. Conversely, there was no enhanced neutralization upon infection with a strain that lacks the glycosylation site N44, which is situated at the edge of the neutralization epitope (Faaberg et al., 2006).
It is easy to imagine that a bulky carbohydrate influences the accessibility of an adjacent epitope and thus prevents binding of a specific antibody by steric hindrance. However, to reduce the amount of antibodies elicited in infected pigs, some of the earlier steps in the humoral immune response (i.e. the activation of B cells) must be hindered by glycosylation of Gp5. According to current text book knowledge, this is assumed to occur by binding of the B cell receptor (membrane-bound IgM) to Gp5 present in virus particles or on cellular debris, followed by endocytosis of the complex, proteolysis of Gp5 and subsequent presentation of Gp5-derived peptides by MHC class II molecules to T helper cells. Since generation of peptides occurs in lysosomes, which also contain glycosidases, MHC-presentation of antigen seems unlikely to be affected by glycosylation of Gp5. However, the initial step, recognition of Gp5 by IgM present on the surface of B cells, resembles an antibody–antigen reaction and may thus be affected by glycan shielding.
2.1.4. Disulphide bond formation between Gp5 and M
As discussed above, Gp5 of EAV forms a disulphide linkage with cysteine 8 in the M protein, an essential modification required for viral infectivity (Snijder et al., 2003). In order to identify the counterpart in Gp5, the five cysteine residues present in its ectodomain were exchanged in a full-length EAV clone, but infectious virus could not be rescued. However, the isolation of a pseudorevertant containing a large deletion and only a single cysteine residue in Gp5 allowed for the identification of cysteine 34 as the binding partner of M. The deleterious effect of replacing the other cysteine residues (at positions 57, 63, 66, and 80) may be explained by prevention of essential intramolecular disulphide bridges between these residues (Snijder et al., 2003). It is also noteworthy that Gp5 of EAV tolerates a deletion of 19 amino acids including the single glycosylation site in its ectodomain; the resultant virus showed only reduced growth rates. A disulphide linkage between Gp5 and M is also essential for the infectivity of lactate dehydrogenase virus (Faaberg et al., 1995), but whether it exists in the respective glycoproteins of PRRSV has not been investigated so far.
In order to verify disulphide bond formation between Gp5 and M of PRSSV, we coexpressed Gp5 (fused at its C-terminus to a His-tag) and M (equipped with a C-terminal HA-tag) with a baculovirus system in insect cells. Western blotting after reducing SDS-PAGE revealed a single band for both Gp5-His and M-HA with the predicted molecular weight of ∼27 kDa and ∼20 kDa, respectively. After non-reducing SDS-PAGE an additional, hence disulphide-linked band of higher molecular weight (∼40 kDa) appeared (Fig. 4 A). To confirm that residue 48, the only cysteine present in the ectodomain of Gp5 after cleavage of the signal peptide, forms the disulphide bond with the M protein, it was replaced by serine. When the Gp5 C48S mutant was co-expressed with M, no band at around 40 kDa was observed under non-reducing conditions, regardless of whether the blot was probed with anti-HA or anti-M antibodies (Fig. 4B).
Fig. 4.
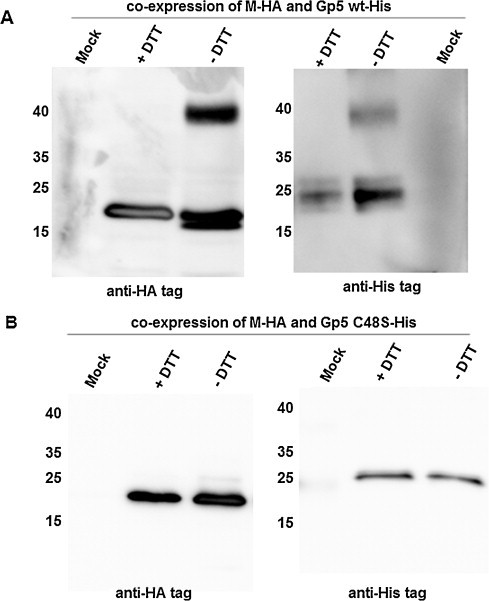
Cysteine 48 of PRRSV-Gp5 forms a disulphide bond with the M protein. Full-length genes of Gp5 from PRRSV VR-2332 (fused via the linker GS to a 7×His-tag) and M (fused via the same linker to the HA-tag) were co-expressed with baculovirus in insect cells as described (Thaa et al., 2013). Cells were lysed with reducing (+DTT) or non-reducing (−DTT) SDS-PAGE buffer, followed by SDS-PAGE (equal amounts were applied) and Western blot with anti-HA and anti-His antibodies, respectively. The numbers left to each blot indicate the mobility of molecular weight markers in kDa. Mock indicates that cells were infected with non-recombinant Baculovirus. (A) Western blot of cells expressing Gp5-His wt and M-HA. (B) Western blot of cells expressing Gp5-His C48S with M-HA.
2.1.5. Function of Gp5/M
The prime function of the Gp5/M complex for the viral life cycle is that it is required for budding of virus particles. Circumstantial evidence for this stems from the finding that no virus particles are released when the expression of either Gp5 or M from a full-length genome of PRRSV or EAV is prevented (Wieringa et al., 2004, Wissink et al., 2005). In addition, it was reported that upon co-expression of Gp5 and M with a baculovirus expression system, virus-like particles (VLPs, vesicles with the same density and appearance as complete virus particles) containing Gp5 and M are released from cells (Nam et al., 2013). This suggests that the Gp5/M complex is the driving force for the generation of membrane curvature in order to create a bud.
The molecular mechanism of arterivirus budding is largely unexplored. The assembly and budding process of an enveloped virus can be generally divided into several steps or tasks, each of which must be executed by a viral (or cellular) protein (Thaa et al., 2010). As the first step, the membrane from which the virus is supposed to bud must be defined, and the components of the prospective virus particle must be transported specifically to the budding site. Since Gp5 and M appear to be the driving force for budding, it is likely that they also define the budding site. This is probably achieved by unidentified retention or retrieval signals that retain the protein in intracellular membranes, the ERGIC and/or cis-Golgi.
Further, it needs to be ensured that all the essential components of the virus are incorporated into the virion, in particular the viral genome. Interestingly, the structure of the C-terminal domain of the N protein reveals an MHC-like fold, suggesting that it might bind short peptide motifs (Doan and Dokland, 2003). One might speculate that the long cytoplasmic tails of M and/or Gp5 bind to the MHC-like domain of N (complexed to the viral RNA), thereby recruiting the viral genome to the viral budding site. Such an interaction may be transient. The Gp5/M complex may not be sufficient for budding; it is conceivable that the nucleocapsid is required at least for initiation of budding. Such a mechanism would ensure that the viral genome is contained in the virion. In infected cells, the N protein has been observed to form protein tubules, intertwined with the replication complex membranes (Knoops et al., 2012, Wada et al., 1995); these tubules might extend towards the virus assembly/budding site. There may even be a connection between the membrane rearrangements that induce the formation of double-membrane vesicles (replication complexes) and virion assembly, involving for instance the N protein or nsp2, recently discovered to be contained in mature virions (Kappes et al., 2013).
Although the Gp2/3/4 complex and the E protein are apparently not required for budding of VLPs, these proteins must be also integrated into particles in order to get an infectious virus. Targeting to the budding site might be achieved by intrinsic, but unidentified sorting signals present in Gp2, Gp3 and/or Gp4, but how the viral spike is enriched in viral particles is completely unknown.
After assembly of all components, the budding process itself needs a protein which physically drives membrane curvature. In principle, membrane bending can be achieved by variety of different, but not mutually exclusive mechanisms: by (1) a scaffolding mechanism, using either a rigid protein with intrinsic curvature (“banana-shaped”) or a coat protein that polymerizes at the surface of or within the membrane, by (2) wedge-like insertion of an amphiphilic helix into one bilayer of the membrane in order to create membrane curvature, or (3) by a crowding mechanism exerted by a “mushroom-shaped” protein, for example a viral spike with a bulky ectodomain and a short cytoplasmic tail (Derganc et al., 2013, Zimmerberg and Kozlov, 2006). The latter mechanism seems unlikely to work for arteriviruses since the ectodomains of the viral glycoproteins are only small. Furthermore, arteriviruses do not possess a cytosolic matrix protein that polymerizes at the assembly site to induce bud formation, as has been described for several negative-strand RNA viruses (Garoff et al., 1998). However, the Gp5/M dimer might form higher-order oligomers, possibly induced by interactions between the membrane-embedded domains of both proteins. This is reminiscent of a mechanism proposed for the budding of caveolae (Bauer and Pelkmans, 2006) and the formation of the highly curved edges of ER tubules (Hu et al., 2011). The proteins involved (caveolin and reticulons/DP1, respectively) have two membrane segments with a hairpin-like topology, similar to what was considered above for parts of the hydrophobic region of Gp5 (see Fig. 3). The hairpin can be regarded as a wedge-like structure that displaces the lipids preferentially in one leaflet of the lipid bilayer, thus causing local curvature.
Formation of a bud is often not sufficient for release of virus particles. Budding may also require cellular factors, such as the ESCRT complex (endosomal sorting complexes required for transport). In the cellular context, this reversibly assembled multisubunit particle is involved in sorting of proteins into vesicles that bud into the lumen of endosomes, thereby forming multivesicular bodies. Some viral proteins recruit parts of the ESCRT complex by so-called “late domains” (short peptide motifs such as PTAP or YPDL) and target them to the viral budding site to cause scission of assembled particles (for review, see Votteler and Sundquist, 2013). However, the ESCRT complex is not hijacked by every virus for efficient budding. Influenza virus, for instance, instead employs the viral protein M2 for virus particle scission. M2 is located at the edge of the viral budding site, and an amphiphilic helix inserts into the membrane to catalyze scission of virus particles (Rossman et al., 2010). For arteriviruses, it has not been investigated whether an amphiphilic helix in one of its proteins mediates scission of virus particles, whether budding depends on ESCRT or whether any other cellular factor is required for budding.
In addition to the role of the Gp5/M complex for virus assembly and budding, the complex is believed to be involved in virus entry. Gp5/M appears to mediate the initial interactions of PRRSV with two cell entry mediators, heparane sulphate glycosaminoglycans and sialoadhesin/CD169 (Van Breedam et al., 2010; see Section 1). Further research is needed to address the issue whether the Gp5/M complex is essential for cell entry of PRRSV. As neutralizing antibodies usually inhibit virus entry, it is important to identify the essential cell receptors and especially their ligand on the viral surface.
2.2. ORF5a protein
The amino acid length of the ORF5a protein varies from 43 to 51 in PRRSV and from 47 to 64 in the other arteriviruses. The predicted molecular mass of the ORF5a protein is 5–6 kDa, but it migrates in SDS-PAGE at approximately 10 kDa. The protein is predicted to have a type III membrane topology, i.e. a central hydrophobic transmembrane region that may function as signal peptide, a short N-terminal ER-luminal ectodomain (5–12 aa) without any glycosylation sequon and a long (14–31 aa) C-terminal endodomain (Firth et al., 2011, Johnson et al., 2011) (see Fig. 5 ). The length of the endodomain could be extended to 63 amino acids by inactivation of putative stop-codons (Sun et al., 2013). There is very little amino acid conservation between ORF5a proteins from arteriviruses except for the sequence MF at the N-terminus and an arginine-/glutamine-rich sequence in the endodomain, penultimate to the predicted transmembrane region. Interestingly, the conserved RQ-motif of the ORF5a protein, which is weakly pronounced in EAV, is encoded by the same RNA sequence that encodes the hypervariable glycosylation-rich ectodomain region of Gp5 (Firth et al., 2011, Johnson et al., 2011). Thus, the genome of PRRSV has to maintain the (probably essential) RQ-motif in ORF5a while at the same time allowing flexibility in the hypervariable region of Gp5 (Robinson et al., 2013a).
Fig. 5.
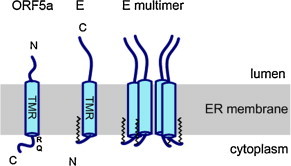
Scheme of the ORF5a and E protein. RQ denotes the presence of an arginine-/glutamine-rich sequence in ORF5a. The zigzag line denotes the attachment of myristate to the N-terminus of E.
The ORF5a protein is essential for replication of type 1 and 2 PRRSV strains, whereas recombinant EAV lacking the ORF5a protein could be generated, but were greatly compromised in growth (Firth et al., 2011, Sun et al., 2013). The function of the ORF5a protein is unknown, but it was suggested to be involved in RNA-binding, similar to cellular proteins containing an RQ-motif (Johnson et al., 2011). The ORF5a protein, both during a natural infection and after immunization with ORF5a protein, elicits a specific antibody response. However, the antibodies produced are neither neutralizing nor do they protect against infection in challenge experiments (Robinson et al., 2013b).
2.3. E protein
2.3.1. Structure, topology, processing and intracellular transport
The E protein, encoded by ORF2a in the genome of EAV and by ORF2b in PRRSV, was identified as a component of arterivirus particles (Snijder et al., 1999, Wu et al., 2005). It is a small protein (8 kDa, 69–73 amino acids) composed of three domains (see Fig. 5): an N-terminal domain (∼25 amino acids) predicted not to be a signal peptide, a hydrophobic transmembrane region (∼20 amino acids), and a hydrophilic, polybasic C-terminal domain (∼25 amino acids) (Snijder et al., 1999). Fluorescence protease protection assays indicate that the hydrophobic domain spans the membrane once; the C-terminus of the E protein is oriented towards the lumen of the ER (and is hence present on the surface of virus particles), whereas the N-terminus is cytosolic (and hence inside virus particles) (Matczuk et al., 2013, Yu et al., 2010). Cross-linking experiments have shown that E forms homo-oligomers, which are not covalently linked by disulphide bonds (Lee and Yoo, 2006). In accordance, conserved cysteine residues present in the C-terminal (luminal) domain of E are not essential for viral replication, indicating that they play no structural role (Lee and Yoo, 2005). However, the exact stoichiometry of the homo-oligomers (i.e. whether E forms dimers, trimers, tetramers or higher-order oligomers) needs to be defined.
Immunoelectron microscopy demonstrated staining of ER and Golgi membranes with E-specific antibodies, and E co-localizes with fluorescent markers of the ER and of various regions of the Golgi (mainly with the ERGIC and with cis-Golgi stacks) in confocal microscopy, both in transfected and in infected cells (Lee and Yoo, 2006, Snijder et al., 1999, Thaa et al., 2009). Transport of E to the plasma membrane has never been observed, indicating that E is retained inside cells, but none of the known retention or retrieval signals are present in the amino acid sequence of E. Using a variety of truncated E constructs fused to GFP, it was concluded that amino acids 1–15, 23–50 and 50–73 may be ER localization sequences (Yu et al., 2010). However, it remains to be shown whether these artificial constructs are representative for the authentic E protein.
The only known covalent modification of E is the co-translational attachment of the fatty acid myristate (C14:0) to the N-terminal glycine residue (Du et al., 2010, Thaa et al., 2009). The myristoylation signal (MGXXXS/T, the initiator methionine is cleaved prior to myristoylation) is strictly conserved in all arteriviruses (Snijder et al., 1999, Thaa et al., 2009). The hydrophobic modification is not essential for initial targeting and anchoring of E to intracellular membranes, which implies that both functions are likely performed by the central hydrophobic domain of the molecule (Thaa et al., 2009). The observation that the total release of particles is not affected in viral mutants with non-myristoylated E suggests that E myristoylation is not required for budding of virus particles and more likely has a function during virus entry (Du et al., 2010, Thaa et al., 2009).
Myristoylation of E is not essential for virus replication, but contributes significantly to virus infectivity. The presence of a myristoylation inhibitor during virus replication completely blocked E acylation and reduced virus titres by 1–2 log10. Similarly, mutant EAV and PRRSV with non-myristoylatable E grew to a titre that was 10–100 fold lower than that of the wild type virus and exhibited a reduced plaque size (Du et al., 2010, Thaa et al., 2009).
2.3.2. Function of E
Reverse genetic experiments revealed that cells release virus-like particles (consisting of RNA and the Gp5, M and N proteins) when the expression of the E protein is prevented in the context of the full-length genome of EAV (Wieringa et al., 2004). Thus, E is not essential for assembly and budding of VLPs. Yet, E is essential for the production of infectious viruses as the inactivation of E expression prevents incorporation of the Gp2/3/4 complex into VLPs. In the absence of the Gp2/3/4 complex, the amount of E present in VLPs is greatly (60–80%) reduced (Wieringa et al., 2004, Wissink et al., 2005). Thus, the E protein and the Gp2/3/4 complex appear to interact structurally (forming a physical complex) or at least functionally during virus assembly.
The E protein of PRRSV was proposed to exert an ion channel-like activity, possibly allowing uncoating of the viral genome in the acidic pH of the endosome (Lee and Yoo, 2006). Since cell entry of both PRRSV and EAV requires clathrin-dependent virus uptake and exposure to a low-pH compartment (Kreutz and Ackermann, 1996, Nauwynck et al., 1999, Nitschke et al., 2008), such a function of E is conceivable. In addition, the structure of E (small, hydrophobic and oligomeric transmembrane protein) resembles the structure of better characterized viroporins, such as M2 of influenza virus (Pinto and Lamb, 2006).
The claim which speaks in favour of an ion-channel function of E is based on two lines of evidence: Replication of PRRSV is blocked by amantadine, a well-known inhibitor of the influenza virus M2 channel, and Escherichia coli cells expressing the E protein become susceptible to a (membrane-impermeable) inhibitor of protein synthesis, suggesting that E has an effect on the membrane permeability (Lee and Yoo, 2006). However, amantadine is a very selective drug; it only inhibits the M2 proton channel of just some influenza A strains (Pinto and Lamb, 2006). Furthermore, the proton conductance of M2 depends on the presence of the strictly conserved sequence motif HXXXW, which is the only sequence element shared between M2 from influenza A virus and BM2 of influenza B virus (Pielak and Chou, 2011). However, this motif is not present in the transmembrane region of E from EAV or PRRSV.
An alternative explanation for the observed effect of amantadine might be that the compound, which is a weak base, raises the pH in the endosome, thereby preventing low-pH triggered membrane fusion and hence virus entry, similar to the other drugs used in the same study (ammonium chloride, chloroquine; Lee and Yoo, 2006). The effect of E on the membrane permeability of E. coli might be due to a general toxic effect exerted by overexpression of a hydrophobic protein. Furthermore, it is not known whether E integrates into the membrane of E. coli cells in a similar orientation and oligomerization status as in virus particles; this is rather unlikely considering the vast difference in membrane-targeting signals between bacteria and higher cells.
Thus, more sophisticated experiments are required to prove that E is indeed a proton channel. For the M2 protein of influenza virus, the proton channel activity was convincingly demonstrated by expression of the protein at the plasma membrane of Xenopus oocytes and subsequent electrophysiological recordings (Pinto et al., 1992). Our attempts to pursue a similar approach for E, fused at its C-terminus to YFP, were unsuccessful as the protein could not be detected at the cell surface of transfected oocytes, which precluded any measurements (M.V. and Thomas Friedrich, Technical University Berlin, unpublished). Thus, new approaches, such as reconstitution of purified E into artificial membranes, are required to test the assumption that E functions as an ion channel.
2.4. The Gp2/3/4 complex
The minor glycoprotein complex of arteriviruses (Gp2/3/4) is believed to be formed by late association of Gp3 with a preformed Gp2/4 dimer (Wieringa et al., 2003b). Therefore, we will first describe the processing of Gp2/4 and then describe and discuss the unique processing scheme of Gp3.
2.4.1. Gp2/4
2.4.1.1. Membrane topology of Gp2 and Gp4
Gp2 is also termed Gp2b in EAV, but Gp2a in PRRSV due to the organization of the genes on the respective genomes; in early publications, Gp2 is referred to as GS (S for small). The protein is composed of 227 amino acids in EAV and 249 in PRRSV. It contains an N-terminal signal peptide, a large ectodomain (∼150 amino acids in EAV, ∼170 amino acids in PRRSV), one hydrophobic transmembrane region (∼25 amino acids in PRRSV, 30 amino acids in EAV, but containing a negatively charged glutamic acid) and a short cytoplasmic tail (∼20 residues in PRRSV, ∼30 in EAV) (de Vries et al., 1995b) (see Fig. 6 ).
Fig. 6.
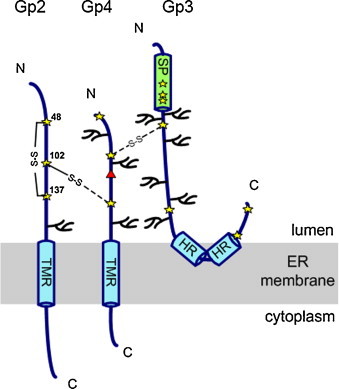
Scheme of the Gp2/3/4 complex of EAV. Cysteine residues within the ectodomains are indicated as stars and used glycosylation sites with branches. The red triangle indicates the probably unused glycosylation site NPT in Gp4. S—S indicates a disulphide bond between cysteine 102 of Gp2 and an unidentified cysteine in Gp4. Gp2 contains also an intramolecular disulphide bond between cysteines 48 and 137. One of the seven cysteines in Gp3 is linked via a disulphide bond with Gp4. TMR: transmembrane regions; HR: hydrophobic region.
Gp4 is composed of 152 amino acids in EAV and 178 in PRRSV. It contains an N-terminal signal peptide, an ectodomain (∼100 amino acids in EAV, ∼130 amino acids in PRRSV), one short hydrophobic transmembrane region (only 15 amino acids in EAV and 17 amino acids in PRRSV if positively charged arginines are excluded) and no (PRRSV) or a very short (EAV, 11 amino acids) cytoplasmic tail. One report claims that the short transmembrane region of PRRSV-Gp4 may serve as a signal sequence for replacement by a glycosylphosphatidylinositol (GPI) anchor, which would result in peripheral membrane attachment of Gp4 rather than transmembrane anchorage. In that study the TMR of a glycolipid-anchored protein was replaced by the TMR of Gp4 and the resulting construct could be released from transfected cells by phosphatidylinositol-specific phospholipase C (PI-PLC), a characteristic hallmark for a GPI-anchored protein (Du et al., 2012). However, it was neither demonstrated that native Gp4 can be released by PI-PLC nor that it contains a GPI-anchor, and thus this notion is not widely accepted.
2.4.1.2. Signal peptide cleavage from Gp2 and Gp4
Using in vitro translation in the presence of microsomes, it was demonstrated that the signal peptide is cleaved from both Gp2 and Gp4 of EAV (de Vries et al., 1992, Wieringa et al., 2002). In order to identify the signal peptide cleavage sites by mass spectrometry, we co-expressed the ectodomains of both proteins, C-terminally fused to either a HA-tag (Gp2-HA) or a Strep-tag (Gp4-strep) with a baculovirus expression system in insect cells. Gp2-HA was co-purified from the supernatant of infected insect cells with Gp4-strep using a Strep-tactin column (Fig. 7A). From this result, we conclude that both proteins were secreted as a disulphide-linked complex. Following digestion of glycans with PNGase F and SDS-PAGE, protein bands corresponding in size to deglycosylated Gp2 and Gp4 were excised from the gel and digested with either elastase or chymotrypsin. The resulting peptides were eluted from the gel slices and subjected to liquid chromatography–tandem mass spectrometry (LC–MS/MS). Processed Gp2 and Gp4 were identified with high confidence in all samples as these proteins were the first hits in the MASCOT search, where mass spectrometric fragments are matched with database entries to determine protein identity. No peptides corresponding in mass to the N-terminal (signal peptide) region were detected for both proteins. This implies that the signal peptide was completely cleaved from both Gp2 and Gp4. In the Gp4 samples, digested with elastase, various peptides starting with threonine 22 were identified (Fig. 7C). As this is not a cleavage site for elastase, the result indicates that Gp4 was cleaved after cysteine 21 by signal peptidase (LPCCWC↓TFYPCH). The most N-terminal peptides detected in chymotrypsin-digested Gp2 samples started with arginine 32, which is preceded by two tryptophan residues in the amino acid sequence (Fig. 7B). Since tryptophan is a chymotrypsin cleavage site, the two residues were most probably removed from the mature Gp4 protein during sample preparation. This suggests that signal peptide cleavage occurred after alanine 29 (GSLLPSAAA↓WWRGVHEVRV). Results from digestion of Gp2 with elastase and Gp4 with chymotrypsin (not shown) are consistent with the conclusion that Gp4 is cleaved after alanine 29 and Gp2 after cysteine 21. No evidence was found for alternative signal peptide cleavage sites such as L24 (GSLL↓PSAA…), predicted in an older report (de Vries et al., 1995a). Thus, the signal peptide is cleaved from both Gp2 and Gp4 at a single site and, since the proteins are secreted from cells after removal of the C-terminal hydrophobic region, both Gp2 and Gp4 are typical type I transmembrane proteins, which corroborates earlier investigations (de Vries et al., 1992, Wieringa et al., 2002).
Fig. 7.
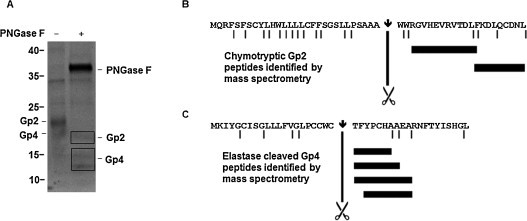
Identification of the signal peptide cleavage sites of recombinant Gp2 and Gp4 by mass spectrometry. (A) The ectodomains of Gp2 from EAV Bucyrus strain (amino acids 1–177 fused to a His-tag) and Gp4 (amino acids 1–120 fused to a Strep-tag) were co-expressed in Sf9 insect cells by infection with recombinant baculovirus prepared as described (Thaa et al., 2013). Gp2-Gp4 complexes were enriched from the cell culture supernatants using a Strep-tactin superflow column (Qiagen). The eluted proteins were digested with PNGase F and subjected to reducing SDS-PAGE and Coomassie staining. One band with the size of deglycosylated Gp2-HA and a double band for Gp4-strep (black boxes) were excised from the gel, digested with chymotrypsin or elastase and analysed by LC–MS/MS as described (Thaa et al., 2013). The left lane shows the purified proteins prior to PNGase F digestion. In both cases peptides corresponding to Gp2 and Gp4, respectively, were the predominant peaks in the MS spectra. (B) Illustrative result from mass spectrometry of Gp2. The first 51 residues of Gp2-HA are shown with the predicted chymotrypsin cleavage sites marked with black lines and the signal peptide cleavage site indicated by an arrow and scissors. Chymotryptic peptides that were identified are denoted as black bars. (C) Illustrative result from mass spectrometry of Gp4. The first 41 residues of Gp4 are shown with the predicted elastase cleavage sites marked with black lines and the signal peptide cleavage site indicated by an arrow and scissors. Identified peptides resulting from digestion with elastase are denoted as black bars.
2.4.1.3. Glycosylation of Gp2 and Gp4
Gp2 proteins of many (but not all) PRRSV strains have two glycosylation sites, N178 and N184, which are located ∼25 amino acids away from the transmembrane region. Gp2 of EAV has just one glycosylation site, located in the same region of the protein. Gp4 proteins of PRRSV have four conserved glycosylation sites, which are distributed over the whole length of the sequence (N37, N84, N120, N130 in VR-2332). All the described glycosylation sites are occupied with carbohydrates, which are usually Endo-H sensitive in infected cells, but are processed to an Endo-H resistant form in virus particles (Das et al., 2011, de Vries et al., 1995b).
Gp4 of EAV also contains four glycosylation sequons (the potential glycosylation sites being N33, N55, N65 and N90). To analyse whether all of them are used, we expressed Gp4 fused at its C-terminus to yellow fluorescent protein (YFP) in CHO cells and digested the [35S]-methionine-labelled and immunoprecipitated protein with Endo H prior to SDS-PAGE (Fig. 8 ). Stepwise reduction of the amount of the glycosidase yielded a ladder-like band appearance that allowed counting of four bands. Since the lower band is completely deglycosylated, one can conclude that Gp4 contains three carbohydrates. Thus, one potential glycosylation site is not used in Gp4. This is most likely N65, which is part of the sequon NPT. Such a sequon is very rarely recognized by the glycosyltransferase (Kaplan et al., 1987) as the proline typically prevents glycosylation (Gavel and von Heijne, 1990).
Fig. 8.
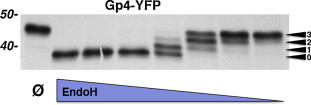
Number of carbohydrates attached to Gp4 of EAV. Gp4-YFP, labelled for 2 h with [35S]-methionine/cysteine in transfected CHO cells, was immunoprecipitated with anti-GFP antibodies from cell lysates and digested with serial twofold dilutions of Endo H prior to SDS-PAGE and fluorography as described (Matczuk et al., 2013). The arrows indicate the number of carbohydrates that have remained on the protein bands.
Various studies have analysed the role of individual glycosylation sites for replication of PRRSV and antigenicity of Gp2 and Gp4. For Gp2, it was reported that none of the two sites affected efficient replication of recombinant viruses (Wei et al., 2012b, Wissink et al., 2004); in another study, N184 was critical for recovery of virus, whereas N178 had no effect (Das et al., 2011). For Gp4 it was found that mutation of individual sequons does not affect virus replication, but double, triple or quadruple mutants of the glycosylation sites are lethal for virus production. None of the viable Gp2 or Gp4 mutants was more sensitive to antibody neutralization or elicited higher titres of neutralizing antibodies after infection of piglets (Das et al., 2011, Wei et al., 2012b, Wissink et al., 2004). Thus, a glycan shielding effect of antibody epitopes, as observed for Gp5, appears not to occur in Gp2 and Gp4.
2.4.1.4. Disulphide bond formation in Gp2 and Gp4
Disulphide bonds play an essential role during processing of the Gp2/4 complex as demonstrated in detailed studies with EAV. Gp2 contains three cysteine residues in its ectodomain, located at positions 48, 102 and 137. Non-reducing SDS-PAGE showed that Gp2 runs as several monomeric bands which are likely to be conformational variants (de Vries et al., 1995b). Mutation of individual residues revealed that cysteine 48 forms an intramolecular bond with cysteine 137, whereas cysteine 102 links Gp2 with Gp4. Incorporation of Gp2 into virus particles is reduced to 10–20% upon mutation of one of the cysteine residues. This suggests that transport of the Gp2/4 complex to the viral budding site is rendered inefficient or retarded by mutating the cysteines, probably due to a folding defect. The viruses that are produced are greatly impaired in replication (C48S, C137S) or are completely non-infectious (C102S). Thus, the disulphide linkage between Gp2 and Gp4 is essential for infectivity of EAV, whereas the intramolecular bond in Gp2 is required for efficient replication (Wieringa et al., 2003a).
After cleavage of the signal peptide between C21 and T22 (see Section 2.4.1.2), three cysteine residues remain in the ectodomain of EAV-Gp4, located at positions 26, 50 and 77. One of these cysteines must link Gp4 to cysteine 102 in Gp2; one or potentially two other cysteines form a disulphide bond with Gp3 in virus particles (see Section 2.4.3). In order to assess the relevance of each cysteine in Gp4 for virus replication, we exchanged them individually to serine in an infectious clone of the Bucyrus strain of EAV. Only in the case of the mutation C50S could infectious virus be rescued, indicating that C26 and C77 are essential for virus infectivity. The virus with the Gp4 mutation C50S produced only tiny plaques (Fig. 9 ) and grew to very low titres (5 × 102 PFU/ml), which precluded further analysis by biochemical means. Thus, similarly to Gp2, the cysteines in Gp4 are either essential for infectivity (C26, C77) or required for efficient virus replication (C50). All of them are thus likely to play a crucial structural role in the Gp2/3/4 complex.
Fig. 9.
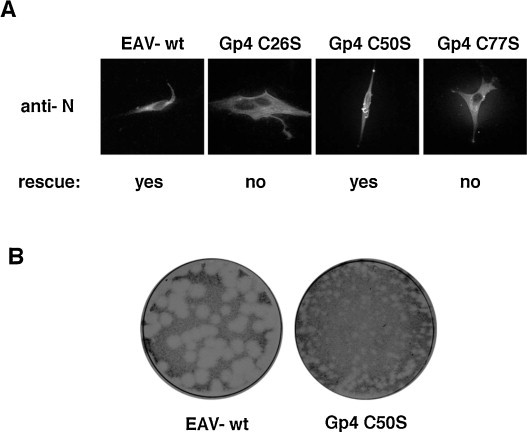
Gp4 cysteine mutants, virus rescue and plaque morphology. (A) BHK cells were nucleoporated with RNA, in vitro synthesized from pEAV211wt and the respective Gp4 mutants as described (Thaa et al., 2009). Immunofluorescence with anti-N antibody performed 16 h after nucleoporation showed that cells express newly synthesized N protein, i.e. they were successfully transfected. (B) The supernatants of transfected cells were collected 3 days after nucleoporation and analysed with a plaque test. No infectious virus could be detected for virus with the Gp4 mutations C26S and C77S, and plaques obtained for virus with the Gp4 mutation C50S were much smaller compared to wild type virus.
2.4.1.5. Neutralizing antibody epitopes in Gp4
The ectodomain of Gp4 may be a relevant antibody target. In Gp4 of the PRRSV strain Lelystad, a linear epitope was identified using a panel of monoclonal antibodies generated in mice. This epitope comprises amino acids 59–67, which are located in a highly variable region of Gp4. In accordance, the monoclonal antibodies neutralize the infectivity of the Lelystad strain, but do not react with Gp4 from the American genotype strain VR-2332 (Meulenberg et al., 1997). A later study revealed that naïve pigs produce antibodies against this epitope 2–3 weeks after experimental infection, even if virus strains with large amino acid differences in this region are used for infection. These antibodies reduce replication of homologous, but not heterologous virus strains in PAMs (Vanhee et al., 2010). Finally, antibody-escape mutants isolated from pigs contained single amino acid substitutions in this epitope and acquired resistance against neutralization by a Gp4-specific monoclonal antibody. However, other antibody-escape mutant variants did not contain amino acid substitutions in the epitope of Gp4, which implies that further neutralizing epitopes might exist in membrane proteins of PRRSV (Costers et al., 2010). Since the neutralizing epitope of Gp4 is highly variable, this part of the molecule is unlikely to be crucial for its function during cell entry. One can speculate that it forms a loop on the surface of the viral spike, similar to the antibody binding sites in influenza virus haemagglutinin (Wiley et al., 1981).
2.4.2. Gp3
The Gp3 gene encodes 163 amino acids in EAV, but the protein is much larger (265 amino acids) in PRRSV. Gp3 consists of an N-terminal signal peptide, an ectodomain containing six (EAV) or seven (PRRSV) potential N-glycosylation sites, a hydrophobic region and a very short (EAV) or longer (PRRSV) hydrophilic domain.
2.4.2.1. Signal peptide cleavage and membrane topology
In vitro translation experiments in the presence of microsomes showed that the N-terminal signal peptide is not cleaved from Gp3, indicating that it might function as a signal anchor (Faaberg and Plagemann, 1997, Hedges et al., 1999, Wieringa et al., 2002). Based on these results, two different topologies were proposed for Gp3, either a type II topology with an N-terminal membrane anchor and a C-terminal ectodomain or, if the hydrophobic C-terminus also spans the membrane, a hairpin structure with two transmembrane regions.
Surprisingly, the bioinformatics tool SignalP 4.0 predicts signal peptide cleavage from Gp3 with high probability. However, SignalP does not take into account possible protein modifications that may affect cleavage. There is an overlapping sequon NNTT adjacent to the signal peptide in Gp3 of EAV, which we have shown to be efficiently N-glycosylated at both asparagines (Matczuk et al., 2013). This is a unique observation in glycobiology, since only one asparagine per molecule is usually used in overlapping sequons (Karamyshev et al., 2005). The signal peptide is retained in the mature protein. However, the signal peptide of Gp3 is cleaved upon deletion of both (but not one) glycosylation sites of the overlapping sequon or when glycosylation is inhibited pharmacologically. This indicates that co-translational attachment of carbohydrates prevents access of the signal peptidase to the cleavage site of Gp3 (Matczuk et al., 2013). An identical result was obtained when the overlapping sequon was exchanged in the context of the viral genome (Matczuk and Veit, 2014).
Since glycosylation usually does not occur close to membrane-spanning segments (Nilsson and von Heijne, 1993), we analysed the topology of Gp3. Membrane fractionation and secretion experiments revealed that the signal peptide does not act as a membrane anchor, suggesting that it is completely translocated into the lumen of the ER. Membrane attachment is caused by the hydrophobic C-terminus of Gp3, which however does not span the membrane, but rather attaches the protein peripherally to the membrane (Fig. 6; Matczuk et al., 2013).
This conclusion is supported by investigations with recombinant viruses with disabled glycosylation sites. When the signal peptide of Gp3 is cleaved (upon mutation of the overlapping sequon), the infectivity of the recombinant EAV is not impaired, and Gp3 with cleaved signal peptide associates with Gp2/4 in virus particles. In contrast, viruses containing Gp3 with deleted hydrophobic C-terminus rapidly reverted back to wild type (Matczuk and Veit, 2014).
Shielding of the signal peptide cleavage site by glycosylation may be a feature common to all arteriviruses: Gp3 proteins from other arteriviruses contain at least one glycosylation site near the signal peptide (Table 5 ), which is also predicted to be cleaved by bioinformatics analyses.
Table 5.
Gp3 proteins from arteriviruses contain at least one glycosylation site near the signal peptide, which is predicted to be cleaved by bioinformatic analyses.

N-terminal Gp3 sequences from various arteriviruses. Glycosylation sequons are in bold, the signal peptide is underlined and a gap in the sequence indicates the most probable cleavage site as predicted with SignalP (http://www.cbs.dtu.dk/services/SignalP/). Shaded in grey is the predicted, but unused, N-terminal myristoylation site (GXXXS) present in Gp3 of EAV (see text). Accession numbers: EAV (Bucyrus strain): P28993, VR-2332: AAD12127, Lelystad: Q04567, SHFV: AAB63392, LDV: NP_042575.
In PRRSV, there is a remarkable difference in the biophysical properties of the C-terminus of Gp3: it is strongly hydrophilic in Lelystad virus, the prototype type 1 strain, but rather hydrophobic in type 2 strains and also the emerging virulent type 1-related strain Lena (Fig. 10 ). Variation in the C-terminus might affect the mode of membrane attachment of Gp3. It is conceivable that PRRSV-Gp3 is only weakly membrane-anchored in some strains. These differences might explain why incorporation of Gp3 into virus particles has been questioned and why a significant fraction of Gp3 molecules is secreted from transfected cells (Mardassi et al., 1998). It is tempting to speculate that a secreted Gp3 protein acts as a “decoy” that “distracts” antibodies from virus particles. Indeed, antibodies against various regions of the Gp3 protein were elicited in the majority of experimentally infected piglets, and most of them had no or only little neutralizing activity (de Lima et al., 2006, Oleksiewicz et al., 2001, Oleksiewicz et al., 2002, Vanhee et al., 2011).
Fig. 10.
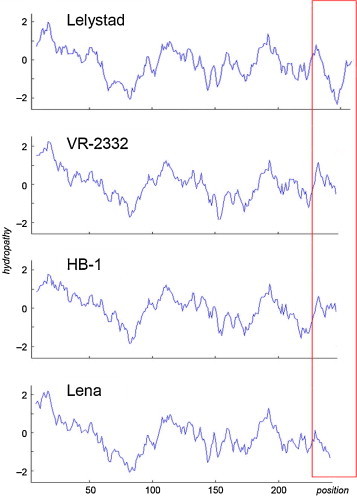
Kyte–Doolittle hydropathy plots for Gp3 from the indicated PRRSV strains (Lelystad/Type 1, accession number Q04567.1; VR-2332/type 2 reference strain, accession number AAD12127.1; HB-1/type 2-related Chinese variant, accession number ABY66112.1; Lena/virulent type 1-related strain, accession number ADZ45529.1). Hydrophobicity (y-axis, windows size 15 amino acids) is plotted against the amino acid number (x-axis). Note that the overall pattern looks similar for all four Gp3 variants except for the C-terminus (red box): The C-terminus is strongly hydrophilic for Gp3 from Lelystad, but rather hydrophobic in the other variants.
The N-terminus of Gp3, but only from EAV, contains a potential myristoylation site (consensus sequence MGXXXS/T, Table 5). Since the N-terminal signal peptide is retained in the mature protein, we asked whether Gp3 is myristoylated. We expressed Gp3 (fused at its C-terminus to YFP) in CHO cells and labelled the protein with [35S]-methionine and with [3H]-myristate, respectively, followed by immunoprecipitation, SDS-PAGE and fluorography. Whereas the E-protein of EAV, expressed and labelled in parallel, was clearly [3H]-myristoylated, no myristoylated band with the size of Gp3 was detected (Fig. 11 ). Gp3 was even not myristoylated when arginine at position 3 was changed to alanine, a mutation that greatly increases the confidence level with which myristoylation of Gp3 is predicted by a bioinformatics tool (http://web.expasy.org/myristoylator/). Thus, the N-terminus of Gp3 is probably shielded from the cytosolic N-myristoyltransferase by binding to the signal recognition particle (SRP), which associates with the signal peptide as soon as it emerges from the ribosome. In the case where myristoylation of an N-terminal signal peptide for a viral glycoprotein was reported (glycoprotein C of arena virus; Schrempf et al., 2007), the signal peptide is much longer (58 amino acids), giving the N-myristoyltransferase more time for interaction until the site for recognition by SRP emerges from the ribosome.
Fig. 11.
Gp3 from EAV is not myristoylated. (A) N-terminal amino acid sequence of E and Gp3 from EAV. “Prediction” indicates the confidence level calculated by the “myristoylator”-tool. The score of the predictor is based on the average responses of 25 artificial neural networks. Values below 0.5 indicate that the majority of networks predict that the protein is not myristoylated. (B) CHO cells, expressing the indicated proteins, were labelled for 2 h with [35S]-methionine/cysteine and [3H]-myristate, respectively. The proteins were immunoprecipitated from cellular extracts with antibodies against GFP and subjected to SDS-PAGE and fluorography as described (Thaa et al., 2009). Note that there are unspecific [35S]-methionine/cysteine labelled bands marked with an asterisk present in each lane running just below E-YFP.
2.4.2.2. Glycosylation of Gp3
Gp3 is a highly glycosylated protein. In EAV, Gp3 contains six potential N-glycosylation sites (including the overlapping sequon), which are all used (Matczuk et al., 2013). Gp3 from PRRRSV contains seven potential N-glycosylation sites. Compared with EAV-Gp3, there is only one site adjacent to the signal peptide, but two additional sites in the C-terminal region. Except the last one (N195), which might by shielded by the adjacent hydrophobic region, all the sites contain carbohydrates (Das et al., 2011).
Various studies have analysed the role of individual glycosylation sites for virus replication and antigenicity of PRRSV-Gp3. In one study, N42, N50 and N131 proved to be required for infectious particle formation in the infectious clone FL-12, whereas N29, N152 and N160 had little (if any) effect, virus growth was impaired only if these sites were exchanged simultaneously (Das et al., 2011). In contrast, recombinant viruses could be generated with all Gp3 glycosylation mutants in another study, where only replacement of N42 and N50 resulted in reduced growth kinetics (Wei et al., 2012b). No effect regarding reactivity with antisera or induction of neutralizing antibodies was seen for any of the viruses with hypoglycosylated Gp3 (Das et al., 2011, Wei et al., 2012b). On the other hand, a field isolate lacking the glycosylation site N131 in Gp3 provoked a stronger neutralizing antibody response (Vu et al., 2011).
2.4.3. Disulphide linkage of Gp3 with Gp2/Gp4
The formation of the disulphide linkage between Gp3 and the Gp2/4 complex of EAV is a unique process: Unlike what has been described for any other viral or cellular protein, Gp3 is not linked to Gp2/4 in the lumen of the ER, where that process could be assisted by PDI and folding factors (Braakman and Bulleid, 2011, Oka and Bulleid, 2013), but only after release of virus particles from infected cells. Disulphide bond formation between Gp3 and Gp2/4 is assumed to be a spontaneous process, accelerated by physiological temperature, oxidizing conditions and slightly basic pH (Wieringa et al., 2003b). Since two cysteine residues must be in very close vicinity to form a covalent bond, Gp3 is likely to be non-covalently associated with the Gp2/4 dimer already in budding virus particles (Wieringa et al., 2003b). Accordingly, the minor glycoproteins can associate with each other independently of glycosylation and disulphide bonds, as demonstrated by refolding experiments with the ectodomains of these proteins purified from E. coli (Kabatek and Veit, 2012).
The cysteine residue(s) linking Gp3 to one (or two) of the available cysteines of Gp4 have not been identified. Gp3 of EAV contains nine cysteines, two of which are located in the (partly hydrophobic) C-terminus, three in the signal peptide and three within (NCS) or very close (NASCN and NTTIC) to used glycosylation sites. Glycosylation may shield the cysteines, but could also enhance their accessibility (by exposing the cysteine-containing amino acid stretch at the surface of the molecule).
For PRRSV, it has not been analysed whether Gp3 associates with the Gp2/4 dimer via a disulphide linkage. It was originally supposed that Gp3 of PRRSV and LDV is a non-structural protein (Faaberg and Plagemann, 1997, Gonin et al., 1998), and that Gp3 is secreted from infected cells. Yet, it has now been demonstrated for both genotypes 1 and 2 of PRRRSV that Gp3 is indeed a structural component of virus particles that may be covalently linked to Gp2/4 (de Lima et al., 2009, Meulenberg et al., 1995).
2.4.4. Building the Gp2/3/4 spike
In summary, the Gp2/3/4 spike is generated by a unique and highly complex process. First, intramolecular disulphide bonds are formed in Gp2 (and probably also in Gp4), which are rearranged until a properly folded product is achieved. This is followed by association of both proteins and stabilization of the resulting dimer by an intermolecular disulphide bond (Wieringa et al., 2003a). Since Gp3 is capable to associate with Gp2/4 in virus particles and also with Gp2 and Gp4 in vitro (Kabatek and Veit, 2012), the question arises what prevents formation of the Gp2/3/4 trimer already in the ER and what triggers its build-up later on. One might speculate that a conformational rearrangement occurs in Gp2/4 and Gp3 after transport from the ER to the Golgi, which might be triggered by the different milieu present in the Golgi lumen. This may now result in closer interaction of Gp3 with Gp2/4 to bring the relevant cysteine(s) into close proximity. Since the lumen of the Golgi lacks PDI and is slightly acidic (Paroutis et al., 2004), disulphide bond formation does not occur here, but only after release of virus particles.
The E protein is likely to play a role in processing of Gp2/3/4 since it greatly facilitates incorporation of Gp2/3/4 into virus particles (Wieringa et al., 2004). This might be due to a direct physical interaction between E and Gp2/3/4, which has, however, not been demonstrated so far. Also, E might conduct protons (or other ions) to adjust the milieu of the lumen of ER or Golgi, which may regulate interactions between Gp2/4 and Gp3. A similar intracellular function has been reported for the proton channel M2 of influenza virus, which neutralizes the pH of the Golgi lumen to prevent an unwanted proton-driven conformational change of the viral spike haemagglutinin (Ohuchi et al., 1994).
Recently, expression of each glycoprotein of PRRSV, either individually or in combination, combined with co-immunoprecipitation assays revealed a strong interaction of Gp5 with Gp4, which is required for the formation of a multiprotein complex containing also Gp2 and Gp3. Thus, Gp5 might help Gp2, Gp3 and Gp4 to fold in a suitable conformation. However, Gp5 was expressed in the absence of its natural binding partner, the M protein, and it is therefore not known whether such complexes would form in virus-infected cells (Das et al., 2010).
Gp2, 3 and 4 need to be transported to the budding site for incorporation into virions. It is only poorly investigated which oligomerization status this requires. When Gp2, Gp3 and Gp4 of EAV are expressed together, all proteins remain Endo-H sensitive, indicating that they are not transported to and beyond the medial-Golgi (Wieringa et al., 2003b). However, it has not been analysed whether oligomerization of the minor glycoproteins occurs under these conditions and whether a certain oligomer (e.g. the Gp2/4 dimer) is transported to the cis-Golgi and can thus pass the quality control system of the ER (Ellgaard and Helenius, 2003). For Gp2/3/4 of PRRSV it has been reported that only co-expression of the three proteins with E allows processing of the proteins into an Endo-H resistant form and partial transport to the plasma membrane. Whether E is actually required for transport of Gp2/3/4 was not determined since Gp2 and E are encoded by overlapping genes and were simultaneously expressed from the same vector in that study (Wissink et al., 2005). Nevertheless, the results suggest that oligomer formation is the critical step for exit of Gp2/3/4 from the ER and further transport. It is totally unclear whether there is a motif in any of the minor glycoproteins that is relevant for subcellular targeting. Note that none of the known retention signals is present in any of the minor glycoproteins. A potential exception could be the transmembrane domain of Gp4, which is rather short, and the cytosolic C-terminus of EAV-Gp2 of EAV that ends with the sequence RRRKIL, reminiscent of the canonical ER retention signal KKXX. However, PRRSV-Gp2 does not contain an identical or similar sequence, and this region is not even conserved between PRRSV strains.
2.4.5. Creating protein diversity by competition between co-translational modifications?
As soon as the nascent peptide chain emerges from the translocon into the lumen of the ER, it can be accessed by the oligosaccharyltransferase (OST) and signal peptidase, which both (but probably not simultaneously) associate with the translocon. Thus, these co-translational processes may affect each other if they compete for overlapping or neighbouring sites in a protein. It is striking that both Gp3 and Gp5 contain N-glycosylation sites in close proximity to the (predicted) signal peptide cleavage sites. This is a rather rare location for a sequon; from the ∼3000 mouse or human protein sequences from the GenBank database that are annotated to contain a signal peptide, only 1.7% possess an N-glycosylation site within five amino acids behind the signal peptide cleavage site (Matczuk et al., 2013).
In the case of Gp3 and Gp5, both modifications affect each other in a different manner. Attachment of a carbohydrate two residues downstream of the signal peptide cleavage site of Gp5 does not inhibit signal peptide processing. Even if an additional site is introduced, generating a double-glycosylated overlapping sequon, signal peptide cleavage is not affected (Thaa et al., 2013). This is in accordance with results on cellular model proteins that also demonstrated that the signal peptide needs to be cleaved before a sequon four residues downstream of the cleavage site can be glycosylated efficiently (Chen et al., 2001). It was concluded that signal peptidase and OST act in a sequential and dependent manner with their substrate, i.e. signal peptide cleavage occurs first (Chen et al., 2001). In contrast, attachment of glycans to sequons two residues downstream of the predicted signal peptide cleavage site prevents cleavage in EAV-Gp3 (Matczuk et al., 2013). Thus, one can conclude for Gp3 that glycosylation must occur first to prevent signal peptide cleavage.
We speculate that it must be a subject of regulation whether signal peptidase or OST has privileged access to the growing polypeptide chain if they compete for neighbouring sites (Fig. 12 ). In the absence of regulation, i.e. if OST and signal peptidase had random access to neighbouring sites, a mixed protein population would be produced: a fraction of proteins would and the other would not contain the signal peptide. However, such heterogeneous protein processing has never been observed.
Fig. 12.
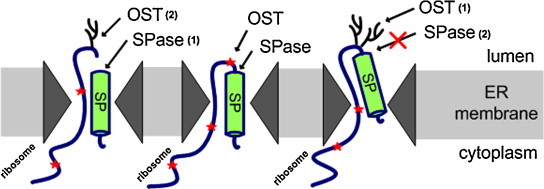
Competition of signal peptidase (SPase) and oligosaccharyltransferase (OST) for neighbouring sites. The middle part of the figure shows the signal peptide (SP) within the translocon presenting both the signal peptide cleavage site and the N-glycosylation sites (red asterisk) to signal peptidase (SPase) and oligosaccharyltransferase (OST). The left part shows the experimental result obtained for Gp5 of PRRSV, where the signal peptide is first cleaved and N-glycosylation occurs after that (Thaa et al., 2013). The right part shows the data for Gp3 of EAV; N-glycosylation occurs first and inhibits signal peptide cleavage (Matczuk et al., 2013).
In addition, another co-translational modification (possibly) competing with signal peptide cleavage and N-glycosylation is disulphide bond formation. For a few cellular proteins it has been shown that disulphide bond formation generally reduces glycosylation at adjacent sites, probably because a folded and fixed protein conformation cannot be processed by OST (Allen et al., 1995, Tanaka et al., 2002). Inspection of the N-terminal amino acid sequences of Gp5 and Gp3 (Table 6 ) shows that they contain multiple cysteine residues, either within the signal peptide or penultimate to the signal peptide cleavage site. Some of these cysteines may form (possibly transient) disulphide bonds to prevent signal peptide cleavage or N-glycosylation.
Table 6.
N-terminal amino acid sequence of Gp3 of EAV and Gp5 of PRRSV.

“No” and “yes” indicate whether the signal peptide is cleaved from the respective wild type protein or indicated mutant (Matczuk et al., 2013, Thaa et al., 2013). The predicted signal peptide is underlined, used glycosylation sites are in bold and underlined and cysteine residues are shaded grey in the wild type protein.
Finally, signal peptide cleavage at more than one site and heterogeneity in the usage of N-terminal glycosylation sites, as demonstrated for Gp5 (Thaa et al., 2013), might create further variability within the N-terminal region of a protein.
It is rarely investigated experimentally whether (and how efficiently) a glycosylation site is used and especially whether (and at which site) a signal peptide is cleaved. Most investigators rely on bioinformatics predictions, which are usually trustworthy, but do not take into account the other co-translational modifications, which are, however, relevant for processing.
How the sequence of co-translational, possibly competitive modifications is regulated is unknown, but the signal peptide itself may play a role. It has emerged that different signal peptides interact with different binding sites within the translocon and that these differences can substantially affect protein biogenesis (Hegde and Bernstein, 2006). One can imagine that a signal peptide recruits either OST or signal peptidase to the translocon to allow privileged access to sites that emerge from the translocon. This in turn might influence whether a cysteine is able to form a disulphide bond and whether co-translational formation of a disulphide bond affects glycosylation or the site of signal peptide cleavage. Thus, mutations in the N-terminal region of Gp3 or Gp5 (and even in the signal peptide) might affect all co-translational modifications in an unpredictable manner. In principle, it needs to be determined experimentally for each protein (and variant thereof) how competition for neighbouring processing sites is resolved.
The biological relevance (if any) of this competition for co-translational processing in Gp3 and Gp5 is not known. Regulation of signal peptide cleavage by glycosylation may allow variability of processing. One may speculate that during virus infection of animals the signal peptide is cleaved from Gp3 in certain cell types only. Furthermore, mutations in the N-terminal region of Gp3 occurring during virus propagation in animals might have a similar effect. We have shown that EAV containing Gp3 with cleaved signal peptide is as infectious as wild type EAV, at least in cell culture (Matczuk and Veit, 2014). EAV isolated from the semen of a persistently infected stallion has lost the glycosylation site in the N-terminal region of Gp3, which is otherwise highly conserved (Balasuriya et al., 1999). Whether or not the co-translational modifications are accomplished has a strong impact on a protein's structure, its interaction with other viral proteins and hence the antigenicity of a viral spike. Changes in the co-translational modifications might cause exposure or masking of antigenic epitopes and thus may contribute to viral persistence, a characteristic hallmark of arterivirus infections.
2.4.6. Function of Gp2/3/4 during virus entry
Using an infectious clone of EAV or PRRSV it was demonstrated that abrogation of expression of either Gp2 or Gp3 or Gp4 does not disturb budding of (non-infectious) virus-like particles (containing Gp5, M and the viral genome), indicating that the minor glycoprotein complex (together with E) is essential for at least one step in cell entry of viruses, either binding to the receptor and/or execution of membrane fusion (Wieringa et al., 2004, Wissink et al., 2005). In line with a role in receptor binding, Gp2 and Gp4, but not Gp3 and Gp5 of PRRSV, could be co-immunoprecipitated with the CD163 receptor if these proteins were co-expressed (Das et al., 2010). Although it is plausible that Gp2 and Gp4 mediate receptor binding, the set-up of the experiments was not appropriate to analyse cell entry of PRRSV since the viral proteins were expressed individually and thus could not form a complex with their binding partner. In the same study a strong interaction between Gp4 and Gp5 was observed, which may be required either for build-up of the Gp2/3/4 spike (as discussed in Section 2.4.4) or for cell entry of PRRSV, suggesting that a network of protein interactions governs virus entry.
A similar conclusion was drawn from a comprehensive study where the amino acid changes between the virulent Bucyrus strain of EAV and attenuated modified live vaccines with a restricted cell tropism were identified. Using chimeric viruses it was concluded that both the Gp5/M and the Gp2/3/4 complex, but neither E nor the non-structural proteins, determine cell tropism. The observed amino acid changes are located mainly in the ectodomains of Gp2, Gp3, Gp4 and Gp5, suggesting that interactions with cellular receptor(s) need to be modified to alter cell tropism. Interestingly, the changes in Gp3 (including a five-amino-acid extension at its C-terminus) were observed only in a vaccine strain with a restricted tropism for CD14+ monocytes, whereas a virus with blocked infectivity for CD3+ T-lymphocytes had no mutations in Gp3. Thus, amino acid changes in Gp3 might be required only for adaption of EAV to some cell types, implying that it recognizes a different receptor than Gp2/4 (Go et al., 2010).
It has not been determined for any arterivirus which of the viral spikes mediates fusion between the viral envelope and cellular membranes. One characteristic hallmark of viral fusion proteins is the presence of a fusion peptide, a hydrophobic region within the protein sequence that – after a conformational change of the fusion protein – inserts into the membrane of the target cell to initiate fusion (Earp et al., 2005, Kielian and Rey, 2006). A Kyte–Doolittle plot reveals that Gp2, Gp3 and Gp4 (but not Gp5 and M) contain an additional region with similar hydrophobicity in the middle of each protein besides the N-terminal signal peptide and the hydrophobic C-terminus (Kabatek and Veit, 2012). These proteins thus resemble classical viral fusion proteins in this respect. Another characteristic feature of viral (and cellular) fusion proteins is the presence of so-called heptad repeats, amino acid sequences with a high propensity to form coiled coil domains. Such heptad repeats are not be predicted for any of the arteriviral membrane proteins (http://toolkit.tuebingen.mpg.de/pcoils).
Thus, it remains unclear whether the Gp2/3/4 complex mediates membrane fusion. To resolve this question experimentally, the Gp2/3/4 complex has to be purified in a native form and tested in established membrane fusion assays, such as haemolysis of erythrocytes (a feature of EAV particles; Veit et al., 2008) or fusion with fluorescently labelled liposomes. Also, it would be instructive to identify the target of cathepsin E (Misinzo et al., 2008), which might cleave the viral glycoprotein that subsequently executes membrane fusion, similar to what has been proposed for SARS and Ebola viruses (Heald-Sargent and Gallagher, 2012, Hunt et al., 2012). Other proteolytic steps that are typically needed for activation of viral membrane fusion activity, such as cleavage at mono- or polybasic sites (Klenk and Garten, 1994), have not been described for any glycoprotein of arteriviruses.
3. Concluding remarks, open questions
Arteriviruses are not as well-studied as important human pathogens such as influenza virus and HIV – despite the increasing relevance especially of PRRSV in the pig industry. Accordingly, our knowledge of arteriviral features is still sparse. Yet, research in the last years has increased our understanding of these viruses, also in terms of their molecular protein biochemistry, which we have reviewed here. Of particular note, this research has revealed peculiarities that appear to be unique in cell biology, for example the inhibition of signal peptide cleavage by adjacent carbohydrates in Gp3 and the late formation of disulphide bonds between Gp3 and Gp2/4 in virus particles. However, such research is not purely academic since it may contribute to a knowledge-based design of vaccines. Arterivirus infections are likely to remain significant in veterinary medicine and might even have zoonotic potential.
Even though our understanding of the molecular and cell biological features of arteriviruses has advanced, crucial questions have remained unresolved. As outlined above, these comprise the role of Gp5/M for virus budding, the identity and function of the viral membrane fusion machinery (which is probably dependent on Gp2/3/4) and the question whether the E protein is an ion channel and how it interacts with the Gp2/3/4 complex.
Acknowledgements
Funding in the authors’ lab was provided by the German Research Foundation (DFG) through project Ve 141/11-1 and Ve 141/13-1, by the EU FP7 through the Marie Curie Initial Training Network “Molecular mechanism of cell entry of enveloped viruses” and by Boehringer Ingelheim Vetmedica GmbH through a PRRSV Research Award. We thank Ineke Braakman and Peter Rottier (Utrecht) for valuable suggestions on processing of Gp3 and Udeni Balasuriya (Kentucky), Kay Faaberg (Iowa), Hans Nauwynck (Ghent), Eric Snijder (Leiden) and Peter Rottier (Utrecht) for clones and antibodies.
Addendum.
Background on processing of membrane proteins
Synthesis of membrane proteins starts at cytoplasmic ribosomes, but is directed to the endoplasmic reticulum (ER) by an N-terminal signal peptide, 15–30 (sometimes up to 50) amino acids that comprise a basic n-region, a hydrophobic h-region and a polar c-region without further sequence consensus (Martoglio and Dobberstein, 1998). The signal peptide is bound by the signal recognition particle (SRP), a cytosolic multisubunit complex (Halic and Beckmann, 2005). SRP then binds to its receptor at the ER membrane, which targets the complex of ribosome and nascent protein chain to the ER. Proteins without an N-terminal signal peptide may be targeted to the ER by other hydrophobic domains (internal signal peptides).
Protein synthesis then continues by insertion of the nascent peptide chain into the translocon (Sec61 complex), a protein-conducting channel in the ER membrane. Hydrophilic peptide stretches are typically passed through the translocon into the lumen of the ER, but hydrophobic stretches are released laterally into the surrounding membrane (Park and Rapoport, 2012, von Heijne, 2006). Hydrophobicity/hydropathy (i.e. little propensity to be water-soluble) is the prime determinant for the formation of membrane-spanning stretches. Hydropathy values of amino acids/peptides can be displayed in hydropathy plots (Kyte–Doolittle plots, such as in Fig. 10); they are also the basis of algorithms to predict transmembrane regions of proteins (Rath and Deber, 2012, von Heijne, 2006).
To form a transmembrane region (TMR), a peptide should have an uninterrupted sequence of at least 20 hydrophobic amino acids (Ala, Ile, Val, Leu, Gly, Phe, also compatible are Ser, Cys and Thr) in order to span the membrane bilayer (thickness: ∼8 nm) completely. Rarely, one (or very few) charged residues (Asp, Glu, Arg, Lys) might be present inside the transmembrane span, especially if they are required for the function of the protein (e.g. in ion channels), but usually such residues and other hydrophilic amino acids (Asn, Gln) flank both sides of the transmembrane segment. The aromatic amino acids (Trp, Tyr) and also histidine favour a position at the interface region between the hydrophobic membrane core and the water-exposed surface (Rath and Deber, 2012, von Heijne, 2006). Shorter hydrophobic spans can form membrane-inserted hairpins. Amphipathic helices have a hydrophilic (hence water-exposed) and a hydrophobic side, the latter is integrated into a membrane.
The topology of a membrane protein (its orientation with respect to the membrane) is usually determined during translation: When a TMD leaves the translocon (through a lateral gate), the following segment of the nascent protein chain is located in the cytosol until a further hydrophobic region inserts into the translocon. In multipass membrane proteins, the amino acids at the cytosolic end of a TMD usually have more basic amino acids compared to the luminal part (“positive inside rule”; White and von Heijne, 2004).
A protein may adopt multiple topologies, for example caused by an inefficient signal peptide (a fraction of the protein then remains cytosolic) or by inefficient insertion of a hydrophobic region into the membrane. Dynamic topologies have also been described; a transmembrane helix can be reorientated or inserted into the membrane post-translationally. If this occurs in just a fraction of all molecules, multiple stable topologies (and thus functions) can arise from the same amino acid chain (von Heijne, 2006).
For single-pass transmembrane proteins, three types are distinguished: Type I membrane proteins have an N-terminal signal peptide, which is cleaved (see below), followed by an ER-luminal stretch, a single TMD, and the C-terminal region facing the cytosol. In type II membrane proteins, the signal peptide is not cleaved and forms a membrane anchor in the mature protein; the topology is opposite (N-terminus towards the cytosol, C-terminus towards the ER lumen). Type III membrane proteins do not have a canonical N-terminal signal peptide; here the N-terminus is exposed to the ER-lumen, followed by a transmembrane stretch (that targets the protein to the ER membrane) and a C-terminal cytosolic domain. In viral membrane proteins, the domain facing the lumen of the ER is referred to as the ectodomain (as it is pointing towards the exterior in the mature virus particle), the cytoplasmic region is the endodomain.
Co-translational modifications of membrane proteins are generated while the protein is being made and sequestered through the translocon. They are important for protein folding, oligomerization, function and antigenicity.
Signal peptide cleavage is performed co-translationally by the signal peptidase, a membrane-bound multimeric protein complex oriented with its catalytic domain towards the ER lumen. During translocation, the c-region of a signal peptide becomes exposed to the lumen of the ER and hence to the signal peptidase. For cleavage to occur, small and uncharged amino acids (Ala, Gly, Ser, Cys) need to be present at positions −1 and −3 upstream of the cleavage site (e.g. …ANA↓SND…). Signal peptides that are not cleaved and contain an h-region long enough to span a lipid bilayer permanently attach the protein to membranes and are hence termed signal-anchor domains. Proteins with a cleaved signal peptide and no further hydrophobic regions often get secreted. There are reliable bioinformatics tools such as SignalP (http://www.cbs.dtu.dk/services/SignalP/) to predict whether the N-terminus of a protein functions as a signal peptide and whether or not it is cleaved.
It was originally thought that signal peptides are interchangeable targeting domains without any further function, but recent studies have revealed a remarkable complexity. Some signal peptides are cleaved very slowly, which affects subsequent folding and N-glycosylation of the respective protein (Land et al., 2003). It is also possible that a signal peptide interacts only weakly with the translocon such that a fraction of the protein remains cytosolic. After cleavage from the protein, the signal peptide can have post-cleavage functions, especially in the life cycle of viruses: Some cleaved signal peptides are relevant for virus particle egress (e.g. in the retrovirus Foamy virus (Lindemann et al., 2001) or as cofactors for protein maturation (of the Lassa virus glycoprotein; Eichler et al., 2003)). Typically, the signal peptide is further processed by the intramembraneous signal peptide peptidase; the resulting peptide fragments can be loaded onto MHC class I and presented on the cell surface to inhibit natural killer cells (Hegde and Bernstein, 2006).
N-glycosylation is performed by oligosaccharyltransferase (OST), a multimeric membrane protein complex with the active site facing the ER lumen. A preassembled core oligosaccharide (glycan) is transferred from the membrane-embedded lipid dolichol to asparagine residues in the target proteins. The asparagine has to be in the context of the sequence motif (sequon) N-X-S/T, where X can be every amino acid except proline. Importantly, only about 2/3 of the potential sites are used as substrate by OST. The residues in and around the sequon affect the efficiency of glycosylation, e.g. NXT is preferred over NXS (Aebi, 2013, Kelleher and Gilmore, 2006, Yan and Lennarz, 2005). Further, folding of a protein can prevent accessibility of a potential glycosylation site. In addition, steric hindrance generally prevents glycosylation of sequons that are too close to a transmembrane span (<13 amino acids; Nilsson and von Heijne, 1993) or too close to another sequon (a distance of at least two residues is required to achieve partial glycosylation of both sites). Overlapping sequons (e.g. NNTT) are either not used or glycosylated at only one site per molecule (Karamyshev et al., 2005, Kelleher and Gilmore, 2006, Reddy et al., 1999). The only known exception are the glycoproteins Gp3 and Gp5 of arteriviruses, which often contain overlapping sequons that are used quite efficiently at both sites (see main text, Section 2.4.2.1). Along the secretory pathway, the core glycosylation (as attached by OST) gets sequentially processed by a multitude of enzymes. In consequence, the glycans usually become resistant to digestion with the enzyme Endoglycosidase H (Endo H) upon passage of the protein through the medial-Golgi, whereas all N-glycosylations can be removed from proteins with Peptide:N-glycosidase F (PNGase F).
Disulphide bonds are formed by oxidation of sulfhydryl (—SH) groups of two cysteine residues, resulting in a covalent bond (—S—S—) between the two cysteines. These bonds support structural stability within a polypeptide chain (intramolecular disulphide bonds) and promote or stabilize the assembly of multi-protein complexes (intermolecular disulphide bonds). Potentially any two cysteine residues in close spatial proximity can form a disulphide bond spontaneously, especially in the oxidizing milieu of the ER lumen. In that case non-native disulphides may arise, which can prevent correct folding. In the ER, such disulphide bonds can be reduced and reformed by protein disulphide isomerases (PDI; Oka and Bulleid, 2013).
Myristoylation is the covalent attachment of the saturated 14-carbon fatty acid myristate to the N-terminal glycine residue in the consensus sequence (M-)G-X-X-X-S/T, catalyzed by N-myristoyltransferase. The start methionine is cleaved off, and myristate is attached by an amide bond to the amino group of the glycine. This typically happens co-translationally in the cytoplasm, but post-translational myristoylation is also possible (after proteolytic cleavage of a protein and generation of an N-terminal glycine). Myristoylation is relevant for membrane targeting of proteins (Bhatnagar et al., 1999, Farazi et al., 2001).
Protein folding and oligomerization. The folding of a membrane protein starts during translocation into the lumen of the ER and continues after release of the completed protein from the translocon. In principle, a protein can fold spontaneously into its native conformation, but the process is assisted by chaperones such as heat shock proteins, BiP, calnexin and calreticulin. These folding factors do not catalyze protein folding, but rather shield newly synthesized proteins – especially hydrophobic regions exposed at their surface – from aggregation (Braakman and Bulleid, 2011, Ellgaard and Helenius, 2003). Correct folding is also governed by glycosylation as well as formation of disulphide bonds and their isomerization by PDI.
Viral membrane proteins form oligomeric spike proteins, either composed of multiple copies of the same protein (homooligomers) or (in the case of Nidovirales) of different viral proteins (heterooligomers). Folding and oligomerization are intimately linked: A glycoprotein with a folding defect does not oligomerize and vice versa. Thus, if a single protein of an heterooligomeric spike is expressed alone (as it often has been reported for membrane proteins of arteriviruses), it is rather unlikely that the product has the same structure and antigenicity as the native counterpart (Doms et al., 1993).
Only properly folded proteins can exit the ER. Unfolded, misfolded and aggregated proteins are disposed of via the ER-associated degradation system (Ellgaard and Helenius, 2003). ER-resident proteins, not supposed to leave the ER, have retention or retrieval signals. The most common of these are the KDEL motif, the di-lysine motif and arginine-based motifs. KDEL is found at the C-terminal end of soluble proteins, which are retrieved from other compartments back to the ER by action of a membrane-bound KDEL receptor (Pelham, 1990). The di-lysine motif (usually KKXX, but also RKXX, KXKXX, KXHXX) is located at the cytoplasmic tail of transmembrane proteins and bound by the cytosolic coat of COPI transport vesicles for recycling of “escaped” proteins back to the ER (Cosson and Letourneur, 1994). Arginine-based motifs occur in various positions in the cytoplasmic tail of heterooligomeric protein complexes and maintain unfolded proteins in the ER until the signal is masked by protein assembly (Michelsen et al., 2005).
The length of the transmembrane region also affects protein targeting. In comparison to the membranes of ER and Golgi, the plasma membrane is thicker and thus harbours proteins with longer transmembrane spans (typically 24 amino acids; Sharpe et al., 2010).
References
- Aebi M. N-linked protein glycosylation in the ER. Biochim. Biophys. Acta. 2013;1833(11):2430–2437. doi: 10.1016/j.bbamcr.2013.04.001. [DOI] [PubMed] [Google Scholar]
- Allen S., Naim H.Y., Bulleid N.J. Intracellular folding of tissue-type plasminogen activator. Effects of disulfide bond formation on N-linked glycosylation and secretion. J. Biol. Chem. 1995;270(9):4797–4804. doi: 10.1074/jbc.270.9.4797. [DOI] [PubMed] [Google Scholar]
- Ansari I.H., Kwon B., Osorio F.A., Pattnaik A.K. Influence of N-linked glycosylation of porcine reproductive and respiratory syndrome virus GP5 on virus infectivity, antigenicity, and ability to induce neutralizing antibodies. J. Virol. 2006;80(8):3994–4004. doi: 10.1128/JVI.80.8.3994-4004.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balasuriya U.B., Go Y.Y., MacLachlan N.J. Equine arteritis virus. Vet. Microbiol. 2013;167(1–2):93–122. doi: 10.1016/j.vetmic.2013.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balasuriya U.B., Hedges J.F., Nadler S.A., McCollum W.H., Timoney P.J., MacLachlan N.J. Genetic stability of equine arteritis virus during horizontal and vertical transmission in an outbreak of equine viral arteritis. J. Gen. Virol. 1999;80(Pt 8):1949–1958. doi: 10.1099/0022-1317-80-8-1949. [DOI] [PubMed] [Google Scholar]
- Balasuriya U.B., Patton J.F., Rossitto P.V., Timoney P.J., McCollum W.H., MacLachlan N.J. Neutralization determinants of laboratory strains and field isolates of equine arteritis virus: identification of four neutralization sites in the amino-terminal ectodomain of the G(L) envelope glycoprotein. Virology. 1997;232(1):114–128. doi: 10.1006/viro.1997.8551. [DOI] [PubMed] [Google Scholar]
- Bauer M., Pelkmans L. A new paradigm for membrane-organizing and -shaping scaffolds. FEBS Lett. 2006;580(23):5559–5564. doi: 10.1016/j.febslet.2006.08.077. [DOI] [PubMed] [Google Scholar]
- Benfield D.A., Nelson E., Collins J.E., Harris L., Goyal S.M., Robison D., Christianson W.T., Morrison R.B., Gorcyca D., Chladek D. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332) J. Vet. Diagn. Invest. 1992;4(2):127–133. doi: 10.1177/104063879200400202. [DOI] [PubMed] [Google Scholar]
- Bhatnagar R.S., Futterer K., Waksman G., Gordon J.I. The structure of myristoyl-CoA:protein N-myristoyltransferase. Biochim. Biophys. Acta. 1999;1441(2–3):162–172. doi: 10.1016/s1388-1981(99)00155-9. [DOI] [PubMed] [Google Scholar]
- Braakman I., Bulleid N.J. Protein folding and modification in the mammalian endoplasmic reticulum. Annu. Rev. Biochem. 2011;80:71–99. doi: 10.1146/annurev-biochem-062209-093836. [DOI] [PubMed] [Google Scholar]
- Calvert J.G., Slade D.E., Shields S.L., Jolie R., Mannan R.M., Ankenbauer R.G., Welch S.K. CD163 expression confers susceptibility to porcine reproductive and respiratory syndrome viruses. J. Virol. 2007;81(14):7371–7379. doi: 10.1128/JVI.00513-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chand R.J., Trible B.R., Rowland R.R. Pathogenesis of porcine reproductive and respiratory syndrome virus. Curr. Opin. Virol. 2012;2(3):256–263. doi: 10.1016/j.coviro.2012.02.002. [DOI] [PubMed] [Google Scholar]
- Chang C.C., Yoon K.J., Zimmerman J.J., Harmon K.M., Dixon P.M., Dvorak C.M., Murtaugh M.P. Evolution of porcine reproductive and respiratory syndrome virus during sequential passages in pigs. J. Virol. 2002;76(10):4750–4763. doi: 10.1128/JVI.76.10.4750-4763.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X., VanValkenburgh C., Liang H., Fang H., Green N. Signal peptidase and oligosaccharyltransferase interact in a sequential and dependent manner within the endoplasmic reticulum. J. Biol. Chem. 2001;276(4):2411–2416. doi: 10.1074/jbc.M007723200. [DOI] [PubMed] [Google Scholar]
- Cho J.G., Dee S.A. Porcine reproductive and respiratory syndrome virus. Theriogenology. 2006;66(3):655–662. doi: 10.1016/j.theriogenology.2006.04.024. [DOI] [PubMed] [Google Scholar]
- Cosson P., Letourneur F. Coatomer interaction with di-lysine endoplasmic reticulum retention motifs. Science. 1994;263(5153):1629–1631. doi: 10.1126/science.8128252. [DOI] [PubMed] [Google Scholar]
- Costers S., Delputte P.L., Nauwynck H.J. Porcine reproductive and respiratory syndrome virus-infected alveolar macrophages contain no detectable levels of viral proteins in their plasma membrane and are protected against antibody-dependent, complement-mediated cell lysis. J. Gen. Virol. 2006;87(Pt 8):2341–2351. doi: 10.1099/vir.0.81808-0. [DOI] [PubMed] [Google Scholar]
- Costers S., Vanhee M., Van Breedam W., Van Doorsselaere J., Geldhof M., Nauwynck H.J. GP4-specific neutralizing antibodies might be a driving force in PRRSV evolution. Virus Res. 2010;154(1–2):104–113. doi: 10.1016/j.virusres.2010.08.026. [DOI] [PubMed] [Google Scholar]
- Darwich L., Diaz I., Mateu E. Certainties, doubts and hypotheses in porcine reproductive and respiratory syndrome virus immunobiology. Virus Res. 2010;154(1–2):123–132. doi: 10.1016/j.virusres.2010.07.017. [DOI] [PubMed] [Google Scholar]
- Das P.B., Dinh P.X., Ansari I.H., de Lima M., Osorio F.A., Pattnaik A.K. The minor envelope glycoproteins GP2a and GP4 of porcine reproductive and respiratory syndrome virus interact with the receptor CD163. J. Virol. 2010;84(4):1731–1740. doi: 10.1128/JVI.01774-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das P.B., Vu H.L., Dinh P.X., Cooney J.L., Kwon B., Osorio F.A., Pattnaik A.K. Glycosylation of minor envelope glycoproteins of porcine reproductive and respiratory syndrome virus in infectious virus recovery, receptor interaction, and immune response. Virology. 2011;410(2):385–394. doi: 10.1016/j.virol.2010.12.002. [DOI] [PubMed] [Google Scholar]
- de Lima M., Ansari I.H., Das P.B., Ku B.J., Martinez-Lobo F.J., Pattnaik A.K., Osorio F.A. GP3 is a structural component of the PRRSV type II (US) virion. Virology. 2009;390(1):31–36. doi: 10.1016/j.virol.2009.04.017. [DOI] [PubMed] [Google Scholar]
- de Lima M., Pattnaik A.K., Flores E.F., Osorio F.A. Serologic marker candidates identified among B-cell linear epitopes of Nsp2 and structural proteins of a North American strain of porcine reproductive and respiratory syndrome virus. Virology. 2006;353(2):410–421. doi: 10.1016/j.virol.2006.05.036. [DOI] [PubMed] [Google Scholar]
- de Vries A.A., Chirnside E.D., Horzinek M.C., Rottier P.J. Structural proteins of equine arteritis virus. J. Virol. 1992;66(11):6294–6303. doi: 10.1128/jvi.66.11.6294-6303.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Vries A.A., Post S.M., Raamsman M.J., Horzinek M.C., Rottier P.J. The two major envelope proteins of equine arteritis virus associate into disulfide-linked heterodimers. J. Virol. 1995;69(8):4668–4674. doi: 10.1128/jvi.69.8.4668-4674.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Vries A.A., Raamsman M.J., van Dijk H.A., Horzinek M.C., Rottier P.J. The small envelope glycoprotein (GS) of equine arteritis virus folds into three distinct monomers and a disulfide-linked dimer. J. Virol. 1995;69(6):3441–3448. doi: 10.1128/jvi.69.6.3441-3448.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delputte P.L., Nauwynck H.J. Porcine arterivirus infection of alveolar macrophages is mediated by sialic acid on the virus. J. Virol. 2004;78(15):8094–8101. doi: 10.1128/JVI.78.15.8094-8101.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derganc J., Antonny B., Copic A. Membrane bending: the power of protein imbalance. Trends Biochem. Sci. 2013;38(11):576–584. doi: 10.1016/j.tibs.2013.08.006. [DOI] [PubMed] [Google Scholar]
- Doan D.N., Dokland T. Structure of the nucleocapsid protein of porcine reproductive and respiratory syndrome virus. Structure. 2003;11(11):1445–1451. doi: 10.1016/j.str.2003.09.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dobbe J.C., van der Meer Y., Spaan W.J., Snijder E.J. Construction of chimeric arteriviruses reveals that the ectodomain of the major glycoprotein is not the main determinant of equine arteritis virus tropism in cell culture. Virology. 2001;288(2):283–294. doi: 10.1006/viro.2001.1074. [DOI] [PubMed] [Google Scholar]
- Dokland T. The structural biology of PRRSV. Virus Res. 2010;154(1–2):86–97. doi: 10.1016/j.virusres.2010.07.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doms R.W., Lamb R.A., Rose J.K., Helenius A. Folding and assembly of viral membrane proteins. Virology. 1993;193(2):545–562. doi: 10.1006/viro.1993.1164. [DOI] [PubMed] [Google Scholar]
- Du Y., Pattnaik A.K., Song C., Yoo D., Li G. Glycosyl-phosphatidylinositol (GPI)-anchored membrane association of the porcine reproductive and respiratory syndrome virus GP4 glycoprotein and its co-localization with CD163 in lipid rafts. Virology. 2012;424(1):18–32. doi: 10.1016/j.virol.2011.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du Y., Zuckermann F.A., Yoo D. Myristoylation of the small envelope protein of porcine reproductive and respiratory syndrome virus is non-essential for virus infectivity but promotes its growth. Virus Res. 2010;147(2):294–299. doi: 10.1016/j.virusres.2009.11.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Earp L.J., Delos S.E., Park H.E., White J.M. The many mechanisms of viral membrane fusion proteins. Curr. Top. Microbiol. Immunol. 2005;285:25–66. doi: 10.1007/3-540-26764-6_2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eichler R., Lenz O., Strecker T., Eickmann M., Klenk H.D., Garten W. Identification of Lassa virus glycoprotein signal peptide as a trans-acting maturation factor. EMBO Rep. 2003;4(11):1084–1088. doi: 10.1038/sj.embor.7400002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellgaard L., Helenius A. Quality control in the endoplasmic reticulum. Nat. Rev. Mol. Cell Biol. 2003;4(3):181–191. doi: 10.1038/nrm1052. [DOI] [PubMed] [Google Scholar]
- Faaberg K.S., Even C., Palmer G.A., Plagemann P.G. Disulfide bonds between two envelope proteins of lactate dehydrogenase-elevating virus are essential for viral infectivity. J. Virol. 1995;69(1):613–617. doi: 10.1128/jvi.69.1.613-617.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faaberg K.S., Hocker J.D., Erdman M.M., Harris D.L., Nelson E.A., Torremorell M., Plagemann P.G. Neutralizing antibody responses of pigs infected with natural GP5 N-glycan mutants of porcine reproductive and respiratory syndrome virus. Viral Immunol. 2006;19(2):294–304. doi: 10.1089/vim.2006.19.294. [DOI] [PubMed] [Google Scholar]
- Faaberg K.S., Plagemann P.G. ORF 3 of lactate dehydrogenase-elevating virus encodes a soluble, nonstructural, highly glycosylated, and antigenic protein. Virology. 1997;227(1):245–251. doi: 10.1006/viro.1996.8310. [DOI] [PubMed] [Google Scholar]
- Fang Y., Snijder E.J. The PRRSV replicase: exploring the multifunctionality of an intriguing set of nonstructural proteins. Virus Res. 2010;154(1–2):61–76. doi: 10.1016/j.virusres.2010.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farazi T.A., Waksman G., Gordon J.I. The biology and enzymology of protein N-myristoylation. J. Biol. Chem. 2001;276(43):39501–39504. doi: 10.1074/jbc.R100042200. [DOI] [PubMed] [Google Scholar]
- Firth A.E., Zevenhoven-Dobbe J.C., Wills N.M., Go Y.Y., Balasuriya U.B., Atkins J.F., Snijder E.J., Posthuma C.C. Discovery of a small arterivirus gene that overlaps the GP5 coding sequence and is important for virus production. J. Gen. Virol. 2011;92(Pt 5):1097–1106. doi: 10.1099/vir.0.029264-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frydas I.S., Verbeeck M., Cao J., Nauwynck H.J. Replication characteristics of porcine reproductive and respiratory syndrome virus (PRRSV) European subtype 1 (Lelystad) and subtype 3 (Lena) strains in nasal mucosa and cells of the monocytic lineage: indications for the use of new receptors of PRRSV (Lena) Vet. Res. 2013;44:73. doi: 10.1186/1297-9716-44-73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garoff H., Hewson R., Opstelten D.J. Virus maturation by budding. Microbiol. Mol. Biol. Rev. 1998;62(4):1171–1190. doi: 10.1128/mmbr.62.4.1171-1190.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gavel Y., von Heijne G. Sequence differences between glycosylated and non-glycosylated Asn-X-Thr/Ser acceptor sites: implications for protein engineering. Protein Eng. 1990;3(5):433–442. doi: 10.1093/protein/3.5.433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Go Y.Y., Zhang J., Timoney P.J., Cook R.F., Horohov D.W., Balasuriya U.B. Complex interactions between the major and minor envelope proteins of equine arteritis virus determine its tropism for equine CD3+ T lymphocytes and CD14+ monocytes. J. Virol. 2010;84(10):4898–4911. doi: 10.1128/JVI.02743-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonin P., Mardassi H., Gagnon C.A., Massie B., Dea S. A nonstructural and antigenic glycoprotein is encoded by ORF3 of the IAF-Klop strain of porcine reproductive and respiratory syndrome virus. Arch. Virol. 1998;143(10):1927–1940. doi: 10.1007/s007050050430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorbalenya A.E., Enjuanes L., Ziebuhr J., Snijder E.J. Nidovirales: evolving the largest RNA virus genome. Virus Res. 2006;117(1):17–37. doi: 10.1016/j.virusres.2006.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halic M., Beckmann R. The signal recognition particle and its interactions during protein targeting. Curr. Opin. Struct. Biol. 2005;15(1):116–125. doi: 10.1016/j.sbi.2005.01.013. [DOI] [PubMed] [Google Scholar]
- Heald-Sargent T., Gallagher T. Ready, set, fuse! The coronavirus spike protein and acquisition of fusion competence. Viruses. 2012;4(4):557–580. doi: 10.3390/v4040557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedges J.F., Balasuriya U.B., MacLachlan N.J. The open reading frame 3 of equine arteritis virus encodes an immunogenic glycosylated, integral membrane protein. Virology. 1999;264(1):92–98. doi: 10.1006/viro.1999.9982. [DOI] [PubMed] [Google Scholar]
- Hegde R.S., Bernstein H.D. The surprising complexity of signal sequences. Trends Biochem. Sci. 2006;31(10):563–571. doi: 10.1016/j.tibs.2006.08.004. [DOI] [PubMed] [Google Scholar]
- Hu J., Prinz W.A., Rapoport T.A. Weaving the web of ER tubules. Cell. 2011;147(6):1226–1231. doi: 10.1016/j.cell.2011.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt C.L., Lennemann N.J., Maury W. Filovirus entry: a novelty in the viral fusion world. Viruses. 2012;4(2):258–275. doi: 10.3390/v4020258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson C.R., Griggs T.F., Gnanandarajah J., Murtaugh M.P. Novel structural protein in porcine reproductive and respiratory syndrome virus encoded by an alternative ORF5 present in all arteriviruses. J. Gen. Virol. 2011;92(Pt 5):1107–1116. doi: 10.1099/vir.0.030213-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kabatek A., Veit M. Folding and oligomerization of the gp2b/gp3/gp4 spike proteins of equine arteritis virus in vitro. Viruses. 2012;4(3):414–423. doi: 10.3390/v4030414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan H.A., Welply J.K., Lennarz W.J. Oligosaccharyl transferase: the central enzyme in the pathway of glycoprotein assembly. Biochim. Biophys. Acta. 1987;906(2):161–173. doi: 10.1016/0304-4157(87)90010-4. [DOI] [PubMed] [Google Scholar]
- Kappes M.A., Miller C.L., Faaberg K.S. Highly divergent strains of porcine reproductive and respiratory syndrome virus incorporate multiple isoforms of nonstructural protein 2 into virions. J. Virol. 2013;87(24):13456–13465. doi: 10.1128/JVI.02435-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karamyshev A.L., Kelleher D.J., Gilmore R., Johnson A.E., von Heijne G., Nilsson I. Mapping the interaction of the STT3 subunit of the oligosaccharyl transferase complex with nascent polypeptide chains. J. Biol. Chem. 2005;280(49):40489–40493. doi: 10.1074/jbc.M509168200. [DOI] [PubMed] [Google Scholar]
- Karniychuk U.U., Geldhof M., Vanhee M., Van Doorsselaere J., Saveleva T.A., Nauwynck H.J. Pathogenesis and antigenic characterization of a new East European subtype 3 porcine reproductive and respiratory syndrome virus isolate. BMC Vet. Res. 2010;6:30. doi: 10.1186/1746-6148-6-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelleher D.J., Gilmore R. An evolving view of the eukaryotic oligosaccharyltransferase. Glycobiology. 2006;16(4):47R–62R. doi: 10.1093/glycob/cwj066. [DOI] [PubMed] [Google Scholar]
- Kielian M., Rey F.A. Virus membrane-fusion proteins: more than one way to make a hairpin. Nat. Rev. Microbiol. 2006;4(1):67–76. doi: 10.1038/nrmicro1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimman T.G., Cornelissen L.A., Moormann R.J., Rebel J.M., Stockhofe-Zurwieden N. Challenges for porcine reproductive and respiratory syndrome virus (PRRSV) vaccinology. Vaccine. 2009;27(28):3704–3718. doi: 10.1016/j.vaccine.2009.04.022. [DOI] [PubMed] [Google Scholar]
- Klenk H.D., Garten W. Host cell proteases controlling virus pathogenicity. Trends Microbiol. 1994;2(2):39–43. doi: 10.1016/0966-842x(94)90123-6. [DOI] [PubMed] [Google Scholar]
- Klumperman J., Locker J.K., Meijer A., Horzinek M.C., Geuze H.J., Rottier P.J. Coronavirus M proteins accumulate in the Golgi complex beyond the site of virion budding. J. Virol. 1994;68(10):6523–6534. doi: 10.1128/jvi.68.10.6523-6534.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knoops K., Barcena M., Limpens R.W., Koster A.J., Mommaas A.M., Snijder E.J. Ultrastructural characterization of arterivirus replication structures: reshaping the endoplasmic reticulum to accommodate viral RNA synthesis. J. Virol. 2012;86(5):2474–2487. doi: 10.1128/JVI.06677-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreutz L.C., Ackermann M.R. Porcine reproductive and respiratory syndrome virus enters cells through a low pH-dependent endocytic pathway. Virus Res. 1996;42(1–2):137–147. doi: 10.1016/0168-1702(96)01313-5. [DOI] [PubMed] [Google Scholar]
- Kwon B., Ansari I.H., Pattnaik A.K., Osorio F.A. Identification of virulence determinants of porcine reproductive and respiratory syndrome virus through construction of chimeric clones. Virology. 2008;380(2):371–378. doi: 10.1016/j.virol.2008.07.030. [DOI] [PubMed] [Google Scholar]
- Land A., Zonneveld D., Braakman I. Folding of HIV-1 envelope glycoprotein involves extensive isomerization of disulfide bonds and conformation-dependent leader peptide cleavage. FASEB J. 2003;17(9):1058–1067. doi: 10.1096/fj.02-0811com. [DOI] [PubMed] [Google Scholar]
- Lee C., Yoo D. Cysteine residues of the porcine reproductive and respiratory syndrome virus small envelope protein are non-essential for virus infectivity. J. Gen. Virol. 2005;86(Pt 11):3091–3096. doi: 10.1099/vir.0.81160-0. [DOI] [PubMed] [Google Scholar]
- Lee C., Yoo D. The small envelope protein of porcine reproductive and respiratory syndrome virus possesses ion channel protein-like properties. Virology. 2006;355(1):30–43. doi: 10.1016/j.virol.2006.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee Y.J., Park C.K., Nam E., Kim S.H., Lee O.S., Lee du S., Lee C. Generation of a porcine alveolar macrophage cell line for the growth of porcine reproductive and respiratory syndrome virus. J. Virol. Methods. 2010;163(2):410–415. doi: 10.1016/j.jviromet.2009.11.003. [DOI] [PubMed] [Google Scholar]
- Li J., Murtaugh M.P. Dissociation of porcine reproductive and respiratory syndrome virus neutralization from antibodies specific to major envelope protein surface epitopes. Virology. 2012;433(2):367–376. doi: 10.1016/j.virol.2012.08.026. [DOI] [PubMed] [Google Scholar]
- Li K., Chen Z., Plagemann P. The neutralization epitope of lactate dehydrogenase-elevating virus is located on the short ectodomain of the primary envelope glycoprotein. Virology. 1998;242(2):239–245. doi: 10.1006/viro.1997.9014. [DOI] [PubMed] [Google Scholar]
- Lindemann D., Pietschmann T., Picard-Maureau M., Berg A., Heinkelein M., Thurow J., Knaus P., Zentgraf H., Rethwilm A. A particle-associated glycoprotein signal peptide essential for virus maturation and infectivity. J. Virol. 2001;75(13):5762–5771. doi: 10.1128/JVI.75.13.5762-5771.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez O.J., Oliveira M.F., Garcia E.A., Kwon B.J., Doster A., Osorio F.A. Protection against porcine reproductive and respiratory syndrome virus (PRRSV) infection through passive transfer of PRRSV-neutralizing antibodies is dose dependent. Clin. Vaccine Immunol. 2007;14(3):269–275. doi: 10.1128/CVI.00304-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez O.J., Osorio F.A. Role of neutralizing antibodies in PRRSV protective immunity. Vet. Immunol. Immunopathol. 2004;102(3):155–163. doi: 10.1016/j.vetimm.2004.09.005. [DOI] [PubMed] [Google Scholar]
- Mardassi H., Athanassious R., Mounir S., Dea S. Porcine reproductive and respiratory syndrome virus: morphological, biochemical and serological characteristics of Quebec isolates associated with acute and chronic outbreaks of porcine reproductive and respiratory syndrome. Can. J. Vet. Res. 1994;58(1):55–64. [PMC free article] [PubMed] [Google Scholar]
- Mardassi H., Gonin P., Gagnon C.A., Massie B., Dea S. A subset of porcine reproductive and respiratory syndrome virus GP3 glycoprotein is released into the culture medium of cells as a non-virion-associated and membrane-free (soluble) form. J. Virol. 1998;72(8):6298–6306. doi: 10.1128/jvi.72.8.6298-6306.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martoglio B., Dobberstein B. Signal sequences: more than just greasy peptides. Trends Cell Biol. 1998;8(10):410–415. doi: 10.1016/s0962-8924(98)01360-9. [DOI] [PubMed] [Google Scholar]
- Matczuk A.K., Kunec D., Veit M. Co-translational processing of glycoprotein 3 from equine arteritis virus: N-glycosylation adjacent to the signal peptide prevents cleavage. J. Biol. Chem. 2013;288(49):35396–35405. doi: 10.1074/jbc.M113.505420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matczuk A.K., Veit M. Signal peptide cleavage from GP3 enabled by removal of adjacent glycosylation sites does not impair replication of equine arteritis virus in cell culture, but the hydrophobic C-terminus is essential. Virus Res. 2014;183:107–111. doi: 10.1016/j.virusres.2014.02.005. [DOI] [PubMed] [Google Scholar]
- Mateu E., Diaz I. The challenge of PRRS immunology. Vet. J. 2008;177(3):345–351. doi: 10.1016/j.tvjl.2007.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng X.J. Heterogeneity of porcine reproductive and respiratory syndrome virus: implications for current vaccine efficacy and future vaccine development. Vet. Microbiol. 2000;74(4):309–329. doi: 10.1016/S0378-1135(00)00196-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meulenberg J.J. PRRSV, the virus. Vet. Res. 2000;31(1):11–21. doi: 10.1051/vetres:2000103. [DOI] [PubMed] [Google Scholar]
- Meulenberg J.J., Petersen-den Besten A., De Kluyver E.P., Moormann R.J., Schaaper W.M., Wensvoort G. Characterization of proteins encoded by ORFs 2 to 7 of Lelystad virus. Virology. 1995;206(1):155–163. doi: 10.1016/S0042-6822(95)80030-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meulenberg J.J., van Nieuwstadt A.P., van Essen-Zandbergen A., Langeveld J.P. Posttranslational processing and identification of a neutralization domain of the GP4 protein encoded by ORF4 of Lelystad virus. J. Virol. 1997;71(8):6061–6067. doi: 10.1128/jvi.71.8.6061-6067.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michelsen K., Yuan H., Schwappach B. Hide and run, arginine-based endoplasmic-reticulum-sorting motifs in the assembly of heteromultimeric membrane proteins. EMBO Rep. 2005;6(8):717–722. doi: 10.1038/sj.embor.7400480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misinzo G.M., Delputte P.L., Nauwynck H.J. Involvement of proteases in porcine reproductive and respiratory syndrome virus uncoating upon internalization in primary macrophages. Vet. Res. 2008;39(6):55. doi: 10.1051/vetres:2008031. [DOI] [PubMed] [Google Scholar]
- Murtaugh M.P., Genzow M. Immunological solutions for treatment and prevention of porcine reproductive and respiratory syndrome (PRRS) Vaccine. 2011;29(46):8192–8204. doi: 10.1016/j.vaccine.2011.09.013. [DOI] [PubMed] [Google Scholar]
- Music N., Gagnon C.A. The role of porcine reproductive and respiratory syndrome (PRRS) virus structural and non-structural proteins in virus pathogenesis. Anim. Health Res. Rev. 2010;11(2):135–163. doi: 10.1017/S1466252310000034. [DOI] [PubMed] [Google Scholar]
- Nam H.M., Chae K.S., Song Y.J., Lee N.H., Lee J.B., Park S.Y., Song C.S., Seo K.H., Kang S.M., Kim M.C., Choi I.S. Immune responses in mice vaccinated with virus-like particles composed of the GP5 and M proteins of porcine reproductive and respiratory syndrome virus. Arch. Virol. 2013;158(6):1275–1285. doi: 10.1007/s00705-013-1612-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nauwynck H.J., Duan X., Favoreel H.W., Van Oostveldt P., Pensaert M.B. Entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages via receptor-mediated endocytosis. J. Gen. Virol. 1999;80(Pt 2):297–305. doi: 10.1099/0022-1317-80-2-297. [DOI] [PubMed] [Google Scholar]
- Nilsson I.M., von Heijne G. Determination of the distance between the oligosaccharyltransferase active site and the endoplasmic reticulum membrane. J. Biol. Chem. 1993;268(8):5798–5801. [PubMed] [Google Scholar]
- Nitschke M., Korte T., Tielesch C., Ter-Avetisyan G., Tunnemann G., Cardoso M.C., Veit M., Herrmann A. Equine arteritis virus is delivered to an acidic compartment of host cells via clathrin-dependent endocytosis. Virology. 2008;377(2):248–254. doi: 10.1016/j.virol.2008.04.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nodelijk G. Porcine reproductive and respiratory syndrome (PRRS) with special reference to clinical aspects and diagnosis. A review. Vet. Q. 2002;24(2):95–100. doi: 10.1080/01652176.2002.9695128. [DOI] [PubMed] [Google Scholar]
- Ohuchi M., Cramer A., Vey M., Ohuchi R., Garten W., Klenk H.D. Rescue of vector-expressed fowl plague virus hemagglutinin in biologically active form by acidotropic agents and coexpressed M2 protein. J. Virol. 1994;68(2):920–926. doi: 10.1128/jvi.68.2.920-926.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oka O.B., Bulleid N.J. Forming disulfides in the endoplasmic reticulum. Biochim. Biophys. Acta. 2013;1833(11):2425–2429. doi: 10.1016/j.bbamcr.2013.02.007. [DOI] [PubMed] [Google Scholar]
- Oleksiewicz M.B., Botner A., Normann P. Porcine B-cells recognize epitopes that are conserved between the structural proteins of American- and European-type porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2002;83(Pt 6):1407–1418. doi: 10.1099/0022-1317-83-6-1407. [DOI] [PubMed] [Google Scholar]
- Oleksiewicz M.B., Botner A., Toft P., Normann P., Storgaard T. Epitope mapping porcine reproductive and respiratory syndrome virus by phage display: the nsp2 fragment of the replicase polyprotein contains a cluster of B-cell epitopes. J. Virol. 2001;75(7):3277–3290. doi: 10.1128/JVI.75.7.3277-3290.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osorio F.A., Galeota J.A., Nelson E., Brodersen B., Doster A., Wills R., Zuckermann F., Laegreid W.W. Passive transfer of virus-specific antibodies confers protection against reproductive failure induced by a virulent strain of porcine reproductive and respiratory syndrome virus and establishes sterilizing immunity. Virology. 2002;302(1):9–20. doi: 10.1006/viro.2002.1612. [DOI] [PubMed] [Google Scholar]
- Park E., Rapoport T.A. Mechanisms of Sec61/SecY-mediated protein translocation across membranes. Annu. Rev. Biophys. 2012;41:21–40. doi: 10.1146/annurev-biophys-050511-102312. [DOI] [PubMed] [Google Scholar]
- Paroutis P., Touret N., Grinstein S. The pH of the secretory pathway: measurement, determinants, and regulation. Physiology (Bethesda) 2004;19:207–215. doi: 10.1152/physiol.00005.2004. [DOI] [PubMed] [Google Scholar]
- Pelham H.R. The retention signal for soluble proteins of the endoplasmic reticulum. Trends Biochem. Sci. 1990;15(12):483–486. doi: 10.1016/0968-0004(90)90303-s. [DOI] [PubMed] [Google Scholar]
- Pielak R.M., Chou J.J. Influenza M2 proton channels. Biochim. Biophys. Acta. 2011;1808(2):522–529. doi: 10.1016/j.bbamem.2010.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinto L.H., Holsinger L.J., Lamb R.A. Influenza virus M2 protein has ion channel activity. Cell. 1992;69(3):517–528. doi: 10.1016/0092-8674(92)90452-i. [DOI] [PubMed] [Google Scholar]
- Pinto L.H., Lamb R.A. The M2 proton channels of influenza A and B viruses. J. Biol. Chem. 2006;281(14):8997–9000. doi: 10.1074/jbc.R500020200. [DOI] [PubMed] [Google Scholar]
- Plagemann P.G. Complexity of the single linear neutralization epitope of the mouse arterivirus lactate dehydrogenase-elevating virus. Virology. 2001;290(1):11–20. doi: 10.1006/viro.2001.1139. [DOI] [PubMed] [Google Scholar]
- Pol J.M., Wagenaar F., Reus J.E. Comparative morphogenesis of three PRRS virus strains. Vet. Microbiol. 1997;55(1–4):203–208. doi: 10.1016/s0378-1135(96)01329-6. [DOI] [PubMed] [Google Scholar]
- Prather R.S., Rowland R.R., Ewen C., Trible B., Kerrigan M., Bawa B., Teson J.M., Mao J., Lee K., Samuel M.S., Whitworth K.M., Murphy C.N., Egen T., Green J.A. An intact sialoadhesin (Sn/SIGLEC1/CD169) is not required for attachment/internalization of the porcine reproductive and respiratory syndrome virus. J. Virol. 2013;87(17):9538–9546. doi: 10.1128/JVI.00177-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rath A., Deber C.M. Protein structure in membrane domains. Annu. Rev. Biophys. 2012;41:135–155. doi: 10.1146/annurev-biophys-050511-102310. [DOI] [PubMed] [Google Scholar]
- Reddy A., Gibbs B.S., Liu Y.L., Coward J.K., Changchien L.M., Maley F. Glycosylation of the overlapping sequons in yeast external invertase: effect of amino acid variation on site selectivity in vivo and in vitro. Glycobiology. 1999;9(6):547–555. doi: 10.1093/glycob/9.6.547. [DOI] [PubMed] [Google Scholar]
- Robinson S.R., Abrahante J.E., Johnson C.R., Murtaugh M.P. Purifying selection in porcine reproductive and respiratory syndrome virus ORF5a protein influences variation in envelope glycoprotein 5 glycosylation. Infect. Genet. Evol. 2013;20:362–368. doi: 10.1016/j.meegid.2013.09.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson S.R., Figueiredo M.C., Abrahante J.E., Murtaugh M.P. Immune response to ORF5a protein immunization is not protective against porcine reproductive and respiratory syndrome virus infection. Vet. Microbiol. 2013;164(3–4):281–285. doi: 10.1016/j.vetmic.2013.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossman J.S., Jing X., Leser G.P., Lamb R.A. Influenza virus M2 protein mediates ESCRT-independent membrane scission. Cell. 2010;142(6):902–913. doi: 10.1016/j.cell.2010.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossow K.D. Porcine reproductive and respiratory syndrome. Vet. Pathol. 1998;35(1):1–20. doi: 10.1177/030098589803500101. [DOI] [PubMed] [Google Scholar]
- Schrempf S., Froeschke M., Giroglou T., von Laer D., Dobberstein B. Signal peptide requirements for lymphocytic choriomeningitis virus glycoprotein C maturation and virus infectivity. J. Virol. 2007;81(22):12515–12524. doi: 10.1128/JVI.01481-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharpe H.J., Stevens T.J., Munro S. A comprehensive comparison of transmembrane domains reveals organelle-specific properties. Cell. 2010;142(1):158–169. doi: 10.1016/j.cell.2010.05.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi M., Lam T.T., Hon C.C., Hui R.K., Faaberg K.S., Wennblom T., Murtaugh M.P., Stadejek T., Leung F.C. Molecular epidemiology of PRRSV: a phylogenetic perspective. Virus Res. 2010;154(1–2):7–17. doi: 10.1016/j.virusres.2010.08.014. [DOI] [PubMed] [Google Scholar]
- Snijder E.J., Dobbe J.C., Spaan W.J. Heterodimerization of the two major envelope proteins is essential for arterivirus infectivity. J. Virol. 2003;77(1):97–104. doi: 10.1128/JVI.77.1.97-104.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snijder E.J., Kikkert M., Fang Y. Arterivirus molecular biology and pathogenesis. J. Gen. Virol. 2013;94(Pt 10):2141–2163. doi: 10.1099/vir.0.056341-0. [DOI] [PubMed] [Google Scholar]
- Snijder E.J., Meulenberg J.J. The molecular biology of arteriviruses. J. Gen. Virol. 1998;79(Pt 5):961–979. doi: 10.1099/0022-1317-79-5-961. [DOI] [PubMed] [Google Scholar]
- Snijder E.J., van Tol H., Pedersen K.W., Raamsman M.J., de Vries A.A. Identification of a novel structural protein of arteriviruses. J. Virol. 1999;73(8):6335–6345. doi: 10.1128/jvi.73.8.6335-6345.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snijder E.J., van Tol H., Roos N., Pedersen K.W. Non-structural proteins 2 and 3 interact to modify host cell membranes during the formation of the arterivirus replication complex. J. Gen. Virol. 2001;82(Pt 5):985–994. doi: 10.1099/0022-1317-82-5-985. [DOI] [PubMed] [Google Scholar]
- Spilman M.S., Welbon C., Nelson E., Dokland T. Cryo-electron tomography of porcine reproductive and respiratory syndrome virus: organization of the nucleocapsid. J. Gen. Virol. 2009;90(Pt 3):527–535. doi: 10.1099/vir.0.007674-0. [DOI] [PubMed] [Google Scholar]
- Stadejek T., Stankevicius A., Murtaugh M.P., Oleksiewicz M.B. Molecular evolution of PRRSV in Europe: current state of play. Vet. Microbiol. 2013;165(1–2):21–28. doi: 10.1016/j.vetmic.2013.02.029. [DOI] [PubMed] [Google Scholar]
- Stertz S., Reichelt M., Spiegel M., Kuri T., Martinez-Sobrido L., Garcia-Sastre A., Weber F., Kochs G. The intracellular sites of early replication and budding of SARS-coronavirus. Virology. 2007;361(2):304–315. doi: 10.1016/j.virol.2006.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun L., Li Y., Liu R., Wang X., Gao F., Lin T., Huang T., Yao H., Tong G., Fan H., Wei Z., Yuan S. Porcine reproductive and respiratory syndrome virus ORF5a protein is essential for virus viability. Virus Res. 2013;171(1):178–185. doi: 10.1016/j.virusres.2012.11.005. [DOI] [PubMed] [Google Scholar]
- Tanaka K., Kitagawa Y., Kadowaki T. Drosophila segment polarity gene product porcupine stimulates the posttranslational N-glycosylation of wingless in the endoplasmic reticulum. J. Biol. Chem. 2002;277(15):12816–12823. doi: 10.1074/jbc.M200187200. [DOI] [PubMed] [Google Scholar]
- Thaa B., Hofmann K.P., Veit M. Viruses as vesicular carriers of the viral genome: a functional module perspective. Biochim. Biophys. Acta. 2010;1803(4):507–519. doi: 10.1016/j.bbamcr.2010.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thaa B., Kabatek A., Zevenhoven-Dobbe J.C., Snijder E.J., Herrmann A., Veit M. Myristoylation of the arterivirus E protein: the fatty acid modification is not essential for membrane association but contributes significantly to virus infectivity. J. Gen. Virol. 2009;90(Pt 11):2704–2712. doi: 10.1099/vir.0.011957-0. [DOI] [PubMed] [Google Scholar]
- Thaa B., Sinhadri B.C., Tielesch C., Krause E., Veit M. Signal peptide cleavage from GP5 of PRRSV: a minor fraction of molecules retains the decoy epitope, a presumed molecular cause for viral persistence. PLoS ONE. 2013;8(6):e65548. doi: 10.1371/journal.pone.0065548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian D., Wei Z., Zevenhoven-Dobbe J.C., Liu R., Tong G., Snijder E.J., Yuan S. Arterivirus minor envelope proteins are a major determinant of viral tropism in cell culture. J. Virol. 2012;86(7):3701–3712. doi: 10.1128/JVI.06836-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Togayachi A., Kozono Y., Ishida H., Abe S., Suzuki N., Tsunoda Y., Hagiwara K., Kuno A., Ohkura T., Sato N., Sato T., Hirabayashi J., Ikehara Y., Tachibana K., Narimatsu H. Polylactosamine on glycoproteins influences basal levels of lymphocyte and macrophage activation. Proc. Natl. Acad. Sci. U. S. A. 2007;104(40):15829–15834. doi: 10.1073/pnas.0707426104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Breedam W., Delputte P.L., Van Gorp H., Misinzo G., Vanderheijden N., Duan X., Nauwynck H.J. Porcine reproductive and respiratory syndrome virus entry into the porcine macrophage. J. Gen. Virol. 2010;91(Pt 7):1659–1667. doi: 10.1099/vir.0.020503-0. [DOI] [PubMed] [Google Scholar]
- Van Breedam W., Verbeeck M., Christiaens I., Van Gorp H., Nauwynck H.J. Porcine, murine and human sialoadhesin (Sn/Siglec-1/CD169): portals for porcine reproductive and respiratory syndrome virus entry into target cells. J. Gen. Virol. 2013;94(Pt 9):1955–1960. doi: 10.1099/vir.0.053082-0. [DOI] [PubMed] [Google Scholar]
- Van Gorp H., Delputte P.L., Nauwynck H.J. Scavenger receptor CD163, a Jack-of-all-trades and potential target for cell-directed therapy. Mol. Immunol. 2010;47(7–8):1650–1660. doi: 10.1016/j.molimm.2010.02.008. [DOI] [PubMed] [Google Scholar]
- Van Gorp H., Van Breedam W., Delputte P.L., Nauwynck H.J. Sialoadhesin and CD163 join forces during entry of the porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2008;89(Pt 12):2943–2953. doi: 10.1099/vir.0.2008/005009-0. [DOI] [PubMed] [Google Scholar]
- Vanhee M., Costers S., Van Breedam W., Geldhof M.F., Van Doorsselaere J., Nauwynck H.J. A variable region in GP4 of European-type porcine reproductive and respiratory syndrome virus induces neutralizing antibodies against homologous but not heterologous virus strains. Viral Immunol. 2010;23(4):403–413. doi: 10.1089/vim.2010.0025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanhee M., Van Breedam W., Costers S., Geldhof M., Noppe Y., Nauwynck H. Characterization of antigenic regions in the porcine reproductive and respiratory syndrome virus by the use of peptide-specific serum antibodies. Vaccine. 2011;29(29–30):4794–4804. doi: 10.1016/j.vaccine.2011.04.071. [DOI] [PubMed] [Google Scholar]
- Veit M., Kabatek A., Tielesch C., Hermann A. Characterization of equine arteritis virus particles and demonstration of their hemolytic activity. Arch. Virol. 2008;153(2):351–356. doi: 10.1007/s00705-007-1094-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verheije M.H., Welting T.J., Jansen H.T., Rottier P.J., Meulenberg J.J. Chimeric arteriviruses generated by swapping of the M protein ectodomain rule out a role of this domain in viral targeting. Virology. 2002;303(2):364–373. doi: 10.1006/viro.2002.1711. [DOI] [PubMed] [Google Scholar]
- von Heijne G. Membrane-protein topology. Nat. Rev. Mol. Cell Biol. 2006;7(12):909–918. doi: 10.1038/nrm2063. [DOI] [PubMed] [Google Scholar]
- Votteler J., Sundquist W.I. Virus budding and the ESCRT pathway. Cell Host Microbe. 2013;14(3):232–241. doi: 10.1016/j.chom.2013.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vu H.L., Kwon B., Yoon K.J., Laegreid W.W., Pattnaik A.K., Osorio F.A. Immune evasion of porcine reproductive and respiratory syndrome virus through glycan shielding involves both glycoprotein 5 as well as glycoprotein 3. J. Virol. 2011;85(11):5555–5564. doi: 10.1128/JVI.00189-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wada R., Fukunaga Y., Kondo T., Kanemaru T. Ultrastructure and immuno-cytochemistry of BHK-21 cells infected with a modified Bucyrus strain of equine arteritis virus. Arch. Virol. 1995;140(7):1173–1180. doi: 10.1007/BF01322744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei Z., Lin T., Sun L., Li Y., Wang X., Gao F., Liu R., Chen C., Tong G., Yuan S. N-linked glycosylation of GP5 of porcine reproductive and respiratory syndrome virus is critically important for virus replication in vivo. J. Virol. 2012;86(18):9941–9951. doi: 10.1128/JVI.07067-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei Z., Tian D., Sun L., Lin T., Gao F., Liu R., Tong G., Yuan S. Influence of N-linked glycosylation of minor proteins of porcine reproductive and respiratory syndrome virus on infectious virus recovery and receptor interaction. Virology. 2012;429(1):1–11. doi: 10.1016/j.virol.2012.03.010. [DOI] [PubMed] [Google Scholar]
- Weiland E., Wieczorek-Krohmer M., Kohl D., Conzelmann K.K., Weiland F. Monoclonal antibodies to the GP5 of porcine reproductive and respiratory syndrome virus are more effective in virus neutralization than monoclonal antibodies to the GP4. Vet. Microbiol. 1999;66(3):171–186. doi: 10.1016/s0378-1135(99)00006-1. [DOI] [PubMed] [Google Scholar]
- Welch S.K., Calvert J.G. A brief review of CD163 and its role in PRRSV infection. Virus Res. 2010;154(1–2):98–103. doi: 10.1016/j.virusres.2010.07.018. [DOI] [PubMed] [Google Scholar]
- Wensvoort G., Terpstra C., Pol J.M., ter Laak E.A., Bloemraad M., de Kluyver E.P., Kragten C., van Buiten L., den Besten A., Wagenaar F. Mystery swine disease in The Netherlands: the isolation of Lelystad virus. Vet. Q. 1991;13(3):121–130. doi: 10.1080/01652176.1991.9694296. [DOI] [PubMed] [Google Scholar]
- White S.H., von Heijne G. The machinery of membrane protein assembly. Curr. Opin. Struct. Biol. 2004;14(4):397–404. doi: 10.1016/j.sbi.2004.07.003. [DOI] [PubMed] [Google Scholar]
- Wieringa R., De Vries A.A., Post S.M., Rottier P.J. Intra- and intermolecular disulfide bonds of the GP2b glycoprotein of equine arteritis virus: relevance for virus assembly and infectivity. J. Virol. 2003;77(24):12996–13004. doi: 10.1128/JVI.77.24.12996-13004.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wieringa R., de Vries A.A., Raamsman M.J., Rottier P.J. Characterization of two new structural glycoproteins, GP(3) and GP(4), of equine arteritis virus. J. Virol. 2002;76(21):10829–10840. doi: 10.1128/JVI.76.21.10829-10840.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wieringa R., de Vries A.A., Rottier P.J. Formation of disulfide-linked complexes between the three minor envelope glycoproteins (GP2b, GP3, and GP4) of equine arteritis virus. J. Virol. 2003;77(11):6216–6226. doi: 10.1128/JVI.77.11.6216-6226.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wieringa R., de Vries A.A., van der Meulen J., Godeke G.J., Onderwater J.J., van Tol H., Koerten H.K., Mommaas A.M., Snijder E.J., Rottier P.J. Structural protein requirements in equine arteritis virus assembly. J. Virol. 2004;78(23):13019–13027. doi: 10.1128/JVI.78.23.13019-13027.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiley D.C., Wilson I.A., Skehel J.J. Structural identification of the antibody-binding sites of Hong Kong influenza haemagglutinin and their involvement in antigenic variation. Nature. 1981;289(5796):373–378. doi: 10.1038/289373a0. [DOI] [PubMed] [Google Scholar]
- Wissink E.H., Kroese M.V., Maneschijn-Bonsing J.G., Meulenberg J.J., van Rijn P.A., Rijsewijk F.A., Rottier P.J. Significance of the oligosaccharides of the porcine reproductive and respiratory syndrome virus glycoproteins GP2a and GP5 for infectious virus production. J. Gen. Virol. 2004;85(Pt 12):3715–3723. doi: 10.1099/vir.0.80402-0. [DOI] [PubMed] [Google Scholar]
- Wissink E.H., Kroese M.V., van Wijk H.A., Rijsewijk F.A., Meulenberg J.J., Rottier P.J. Envelope protein requirements for the assembly of infectious virions of porcine reproductive and respiratory syndrome virus. J. Virol. 2005;79(19):12495–12506. doi: 10.1128/JVI.79.19.12495-12506.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wissink E.H., van Wijk H.A., Kroese M.V., Weiland E., Meulenberg J.J., Rottier P.J., van Rijn P.A. The major envelope protein GP5, of a European porcine reproductive and respiratory syndrome virus contains a neutralization epitope in its N-terminal ectodomain. J. Gen. Virol. 2003;84(Pt 6):1535–1543. doi: 10.1099/vir.0.18957-0. [DOI] [PubMed] [Google Scholar]
- Wood O., Tauraso N., Liebhaber H. Electron microscopic study of tissue cultures infected with simian haemorrhagic fever virus. J. Gen. Virol. 1970;7(2):129–136. doi: 10.1099/0022-1317-7-2-129. [DOI] [PubMed] [Google Scholar]
- Wu W.H., Fang Y., Rowland R.R., Lawson S.R., Christopher-Hennings J., Yoon K.J., Nelson E.A. The 2b protein as a minor structural component of PRRSV. Virus Res. 2005;114(1–2):177–181. doi: 10.1016/j.virusres.2005.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan A., Lennarz W.J. Unraveling the mechanism of protein N-glycosylation. J. Biol. Chem. 2005;280(5):3121–3124. doi: 10.1074/jbc.R400036200. [DOI] [PubMed] [Google Scholar]
- Yang L., Frey M.L., Yoon K.J., Zimmerman J.J., Platt K.B. Categorization of North American porcine reproductive and respiratory syndrome viruses: epitopic profiles of the N, M, GP5 and GP3 proteins and susceptibility to neutralization. Arch. Virol. 2000;145(8):1599–1619. doi: 10.1007/s007050070079. [DOI] [PubMed] [Google Scholar]
- Yoon I.J., Joo H.S., Goyal S.M., Molitor T.W. A modified serum neutralization test for the detection of antibody to porcine reproductive and respiratory syndrome virus in swine sera. J. Vet. Diagn. Invest. 1994;6(3):289–292. doi: 10.1177/104063879400600326. [DOI] [PubMed] [Google Scholar]
- Yu M., Liu X., Sun L., Chen C., Ma G., Kitamura Y., Gao G.F., Liu W. Subcellular localization and topology of porcine reproductive and respiratory syndrome virus E protein. Virus Res. 2010;152(1–2):104–114. doi: 10.1016/j.virusres.2010.06.012. [DOI] [PubMed] [Google Scholar]
- Zhou L., Yang H. Porcine reproductive and respiratory syndrome in China. Virus Res. 2010;154(1–2):31–37. doi: 10.1016/j.virusres.2010.07.016. [DOI] [PubMed] [Google Scholar]
- Zimmerberg J., Kozlov M.M. How proteins produce cellular membrane curvature. Nat. Rev. Mol. Cell Biol. 2006;7(1):9–19. doi: 10.1038/nrm1784. [DOI] [PubMed] [Google Scholar]
- Zimmerman J.J., Yoon K.J., Wills R.W., Swenson S.L. General overview of PRRSV: a perspective from the United States. Vet. Microbiol. 1997;55(1–4):187–196. doi: 10.1016/s0378-1135(96)01330-2. [DOI] [PubMed] [Google Scholar]