

Abstract
Resistance to oseltamivir in pandemic (H1N1) 2009 influenza A virus is linked to an amino acid change from histidine (H) to tyrosine (Y) at position 275 in the neuraminidase protein (NA). A real-time one step RT-PCR assay using single nucleotide polymorphism (SNP) probes was developed to detect this mutation in respiratory specimens. The limit of detection was 47.6 copies/reaction for wild-type H275 RNA and 52.9 copies/reaction for the mutant H275Y RNA. The assay did not cross-react with other respiratory pathogens. The clinical sensitivity and specificity of the assay was compared to the gold standard Sanger sequencing method using 25 sensitive, 15 resistant and 20 negative samples. The sensitivity and specificity was 88.0% and 100% respectively with the SOIV_Osel_SEN probe designed to detect the H275 allele and 100% for the SOIV_Osel_RES probe detecting the 275Y allele. The sensitivity of the assay using nine admixtures of sensitive and resistant alleles was 88.9% and 77.8% with the SOIV_Osel_SEN probe and SOIV_Osel_RES probe respectively. The presence of mixed sensitive and resistant alleles in patient samples and mixtures of in vitro RNA were detected reproducibly. This assay can be used for screening of original samples for oseltamivir resistance without the need for culture and phenotypic testing.
Keywords: Pandemic (H1N1) 2009, Oseltamivir, Resistance, H275, H275Y, Single nucleotide polymorphism (SNP), Real-time RT-PCR
1. Introduction
The influenza pandemic of 2009 saw the emergence of a new influenza A (H1N1) 2009 strain that showed initial resistance to adamantanes and susceptibility to oseltamivir (Dawood et al., 2009). However, by the summer of 2009, there were reports of isolates with an amino acid change from histidine (H) to tyrosine (Y) at position 275 in the neuraminidase (NA) glycoprotein that was linked to oseltamivir resistance (Chen et al., 2009, Harvala et al., 2010, Janies et al., 2010, Thabet et al., 2010, WHO, 2009a). Some of these cases were identified in Alberta, Canada starting in July 2009. Although the prevalence of oseltamivir resistance in the 2009–2010 respiratory season has been low, there have been instances in the recent past of dramatic increases in oseltamivir and adamantane resistance in seasonal influenza A (IFVA) over a relatively short time frame in the absence of selective pressure by anti-viral usage (Deyde et al., 2007, Dharan et al., 2009, Hauge et al., 2009, Hurt et al., 2009, Janies et al., 2010, Nelson et al., 2009, Pabbaraju et al., 2010, Sheu et al., 2008). Resistance to oseltamivir has been reported to result from point mutations in several regions of the NA protein of the virus (e.g., D79G, S247G or S247N, and H275Y) (Sheu et al., 2008). Phylogenetic studies have revealed that the oseltamivir-resistant seasonal IFVA (H1N1) strain evolved from a susceptible strain (clade 2B) which circulated in the 2007–2008 season by acquiring the H275Y change in the NA gene (Zaraket et al., 2010).
Currently, the gold standard method for detection of the C823T nucleotide polymorphism that leads to the H275Y amino acid change is dideoxy Sanger sequencing of the NA gene. Neuraminidase inhibition assays including commercial kits such as the NA-Star® Influenza Neuraminidase Inhibitor Resistance assay (Applied Biosystems, Foster city, CA, USA) have been described for phenotypic resistance testing (Hurt et al., 2009). These phenotypic assays require culturing of the virus, which would not be possible in the case of novel sub-type variants such as the pandemic (H1N1) 2009 prior to characterization of the virus. Both sequencing and phenotypic testing are labor intensive, time-consuming, and expensive. The use of pyrosequencing for monitoring nucleotide changes has also be described, however this requires specialized instrumentation (Deyde et al., 2009, Deyde et al., 2010). A reverse-transcriptase PCR coupled with a restriction fragment length polymorphism assay (RTPCR-RFLP) has been described for seasonal H1N1 viruses but it is a lengthy process and not practical for high volume testing (Guo et al., 2009). A multiplex RT-PCR for subtyping and determination of oseltamivir resistance for seasonal and pandemic H1N1 influenza A viruses has been reported recently using liquid microarrays on the Luminex platform (Mahony et al., 2010). Real-time reverse-transcriptase (RT)-PCR assays using single nucleotide polymorphism (SNP) probes for allelic discrimination have been described for the detection of H275Y mutation in seasonal H1N1 viruses (Bolotin et al., 2009, Carr et al., 2008, Operario et al., 2010) and pandemic (H1N1) 2009 viruses (van der Vries et al., 2010). Rapid methods for the detection of mutations leading to oseltamivir resistance allow diagnostic laboratories to monitor the development and transmission of resistance especially in high risk populations such as immuno-compromised patients and patients requiring antiviral treatment.
This study describes the validation of a high throughput, sensitive and specific RT-PCR for detection of the C823T nucleotide polymorphism leading to the H275Y amino acid change in the NA protein of pandemic (H1N1) 2009 virus. This assay could be used in surveillance of tamiflu resistance, detection of a resistant virus and monitoring response to therapy.
2. Materials and methods
2.1. Clinical specimens and virus strains
Respiratory specimens submitted to the Provincial Laboratory for Public Health (ProvLab), Alberta, Canada, between July 6th 2009 and January 4th 2010 for respiratory virus testing, were included in this analysis. The current diagnostic algorithm for testing of influenza A in our laboratory includes primary screening using a real-time RT-PCR assay (InfA) targeting the matrix gene of all human influenza A viruses (Dawood et al., 2009). Samples that test positive for IFVA are tested subsequently for H1, H3 and pandemic (H1N1) 2009 sub-types (Dawood et al., 2009, Pabbaraju et al., 2009). Specimens from patients with a significant travel history, epidemiological links to pandemic (H1N1) 2009 virus, severe respiratory infection, immuno-compromised/bone marrow transplant cases and intensive care unit patients were identified based on the requisitions submitted by the physician. Additionally, patients from the community with none of the above predisposing factors for development of resistance were also included in the analysis. Selected pandemic (H1N1) 2009 virus confirmed samples were tested for oseltamivir resistance using the gold standard Sanger sequencing method (n = 306). Sixty-nine specimens were selected for the assay validation, of which 45 were upper respiratory specimens including nasopharyngeal and throat swabs, 24 were lower respiratory specimens including endotracheal aspirate, bronchoalveolar lavage and auger suction.
2.2. Viral nucleic acid extraction
Extraction of total nucleic acid from all clinical specimens was performed on the easyMAG® automated extraction system (bioMérieux Canada Inc., Québec, Canada) following the manufacturer's instructions and as described previously (Pabbaraju et al., 2009).
2.3. Assay design and optimization
Primers and two minor-groove-binding (MGB) hydrolysis probes were designed based on the available NA gene sequences from GenBank (as of September 21, 2009) using the Primer Express™ software [version 3.0.0, Applied Biosystems (ABI), Foster City, CA, USA]. The two MGB probes were designed for discrimination of sensitive and resistant alleles. The probe specific for detection of the H275 allele that has been reported to lead to oseltamivir sensitivity (SOIV_Osel_SEN) was labelled with 6-carboxyfluorescein (FAM); and a probe specific for the H275Y allele that has been reported to lead to oseltamivir resistance (SOIV_Osel_RES) was labelled with VIC at the 5′ end; both probes had a non-fluorescent black hole quencher (BHQ) at the 3′ end. Nucleotide sequences of the primers and probes are indicated in Table 1 . Optimal primer concentrations were determined by checkerboard titrations using concentrations ranging from 400 nM to 800 nM. The optimal probe concentration was determined by testing concentrations ranging from 100 nM to 200 nM. Primers were synthesized at the Core DNA services, University of Calgary (Alberta, Canada) and the probes were purchased from ABI.
Table 1.
Primer and probe sequences designed for detection of the C823T mutation. The SNP position is highlighted in bold and underlined.
| Assay | Oligonucleotide name | Oligonucleotide sequence (5′–3′) | Amplicon (bp) | Locationa | Source |
|---|---|---|---|---|---|
| Sequencing (Partial NA) | NA675F_nested_sw | AACACAAGAGTCTGAATGTGC | 415 | 675–695 | In-house |
| NA1089R_nested_sw | ACCGTTTCTTGAACTAATGCTT | 1089–1110 | |||
| Real-time RT-PCR | SOIV_NA_781F | GGAAAGATAGTCAAATCAGTCGAAATG | 781–807 | In-house | |
| SOIV_NA_878R | CTGCACACACATGTGATTTCACTAG | 98 | 854–878 | ||
| SOIV_Osel_SEN | FAM – CCTCATAGTGATAATTA – MGB | 816–832 | |||
| SOIV_Osel_RES | VIC – CCTCATAGTAATAATTAG – MGB | 815–832 | |||
Nucleotide numbering is based on the neuraminidase (NA) gene of influenza A virus (A/California/04/2009(H1N1), GenBank accession no. GQ117044).
2.4. In vitro transcribed RNA
Full length amplification of the NA gene with the H275 and H275Y alleles from pandemic (H1N1) 2009 virus was performed using primers described previously (Mak et al., 2006). These amplified fragments were cloned into the plasmid vector pCRII-TOPO® and transformed into One Shot® TOP-10 chemically competent Escherichia coli cells (Invitrogen, Carlsbad, CA, USA). The recombinant plasmids with cloned NA fragments were purified using the QIAprep® Kit (Qiagen), linearized using the restriction enzyme Hind III and used as templates for transcription. RNA was transcribed using the T7 RiboMAX™ Express kit (Promega, Madison, WI, USA) and treated with TURBO® DNase (Applied Biosystems Canada, Streetsville, Ontario, Canada) for degradation of template DNA. The purified RNA was quantified using the NanoDrop™ spectrophotometer (Thermo Fisher Scientific, Wilmington, Delaware, USA) and the absorbance value was used to calculate the copy number of transcribed RNA.
2.5. One step RT-PCR amplification using SNP probes
Amplification was carried out using SuperScript™ III Platinum® One-step Quantitative Kit (Invitrogen) in a final volume of 25 μL with 5 μL of extracted RNA as template, 800 nM each of the forward and reverse primers, and 200 nM of each probe. Amplification and detection was performed on the 7500 Sequence Detection System (7500SDS, Applied Biosystem™ (ABI), Foster City, CA, USA). The amplification conditions were: reverse transcription at 48 °C for 30 min, Taq activation for 10 min at 95 °C (there was no difference in CT values with activation for 2 min), followed by 45 cycles of amplification comprising of denaturation for 15 s at 95 °C, annealing and primer extension for 1 min at 60 °C. A threshold of 0.3 was used for analysis of amplification curves and the baseline was calculated using default settings on the software (SDS version 1.4).
2.6. Sensitivity, dynamic range, specificity and reproducibility of the RT-PCR
Analytical sensitivity of the real-time RT-PCR was determined by testing serial dilutions of quantified in vitro transcribed RNA for both the H275 and H275Y alleles in eight replicates. Serial dilutions of RNA spanning eight logs of template concentration were tested in triplicate to determine the dynamic range of the assay. The primers and probes were designed to be specific for the pandemic (H1N1) 2009 virus sequence, however, they were evaluated experimentally for cross-reactivity with the following targets using extracted DNA/RNA from high titre viral and bacterial cultures or positive respiratory samples as template: IFVA (seasonal H1N1, n = 13; seasonal H3N2, n = 11; H1N2, n = 1; recombinant H3N2, n = 1), influenza B (IFVB) (n = 2), parainfluenza (PIV) (types 1, 2, 3, 4B), respiratory syncytial virus (RSV) (A and B), human coronaviruses (hCoV) (229E, OC43, NL63 and HKU1), human metapneumovirus (hMPV) (lineages 1 and 2), human rhinovirus 1B, coxsackievirus A16, echovirus 2, human adenovirus type 3, human bocavirus, Chlamydophila pneumoniae, Legionella pneumophila and Mycoplasma pneumoniae.
The inter-assay and intra-assay variability was assessed using three clinical samples each with the H275 and H275Y alleles at three levels of viral load as determined by the cycle threshold (Ct) values. The samples were tested in triplicate on three independent runs.
2.7. Accuracy testing using clinical specimens and in vitro RNA
Sixty-nine samples that had been confirmed previously as positive for pandemic (H1N1) 2009 virus by real-time RT-PCR and sequenced for the NA gene were used for testing assay accuracy. These included 25 sensitive (C823) and 15 resistant (823T) samples. Nine samples containing admixtures of C823 and 823T alleles as determined by close observation of the electropherogram were also included. Twenty samples which had tested negative for IFVA viruses were included as negative controls.
In vitro transcribed RNA with the C823 and 823T markers were selected at 2 different concentrations to represent high and low viral titre samples. The copy numbers for high titre RNA were 4.76 × 104 and 5.29 × 104 per reaction for C823 and 823T, respectively. The copy numbers for low titre RNA were 4.76 × 102 and 5.29 × 102 per reaction for C823 and 823T, respectively. At each concentration, a mixture of RNA was prepared ranging from 5% to 95% of each allele. The RNA mixtures were tested in triplicate for the detection of mixed clones and estimation of sensitivity for detection when both alleles are present.
2.8. Sanger sequencing of NA gene
Conventional Sanger sequencing of the region of interest in the NA gene was used as the gold-standard method for this study. Three hundred and six clinical samples that tested positive for pandemic (H1N1) 2009 virus from immuno-compromised, intensive care unit cases, patients who had received oseltamivir treatment, had a travel history and from the community were selected for sequencing of the partial NA gene including the H275Y (C823T) region. A 415 bp fragment of the NA gene was amplified using primers (NA675F_nested_sw and NA1089R_nested_sw) (Table 1), and the amplified product was purified using the EZ-10 PCR purification kit (Bio Basic Inc., Markham, Ontario, Canada). Sequencing was performed on both strands using the BigDye® Terminator v3.1 Cycle Sequencing Kit (ABI) in the ABI PRISM® 3130-Avant Genetic Analyzer. Careful manual examination of the electropherogram at base pair 823 was also performed for the presence of mixed bases.
3. Results
3.1. Dynamic range, sensitivity, specificity and reproducibility of the assay
This RT-PCR assay showed a broad dynamic range using in vitro transcribed RNA. Linear amplification of template was observed between 4.76 × 101 to 4.76 × 108 and 5.29 × 102 to 5.29 × 109 copies per reaction for H275 and H275Y alleles, respectively (Fig. 1 a and b). The assay efficiency was 98.8% and 98.0% for the SOIV_Osel_SEN probe and SOIV_Osel_RES probe respectively, and the R 2 values comparing template concentrations and Ct values were 1.0 for both probes.
Fig. 1.
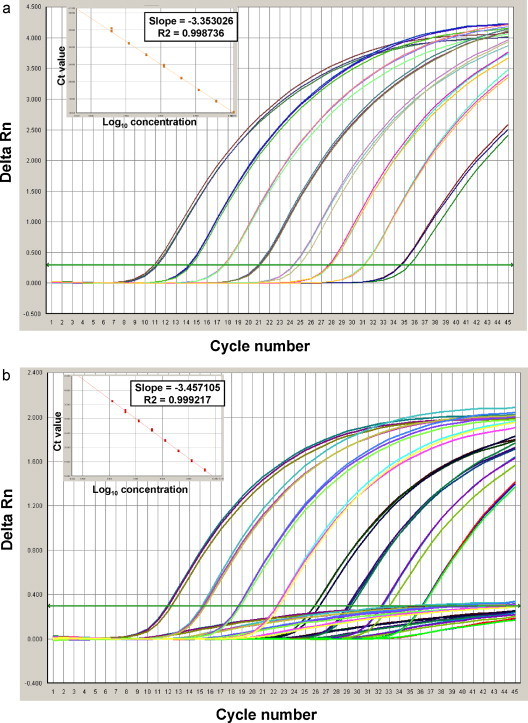
(a) Dynamic range of the developed RT-PCR assay. Amplification curves (Delta Rn vs. Cycle number) and standard curve (Ct vs. log10 concentration) for serial dilutions of the plasmid with C823 nucleotide. Assay dynamic range was linear between template concentrations ranging from 4.76 × 101 copies/reaction to 4.76 × 108 copies/reactions. (b) Dynamic range of the developed RT-PCR assay. Amplification curves (Delta Rn vs. Cycle number) and standard curve (Ct vs. log10 concentration) for serial dilutions of the plasmid with C823T nucleotide. Assay dynamic range was linear between template concentrations ranging from 5.29 × 102 copies/reaction to 5.29 × 109 copies/reactions.
Limit-of-detection limit for the assay was determined by testing 10-fold serial dilutions of quantified RNA in triplicate, resulting in 47.6 copies and 52.9 copies for the H275 and the H275Y alleles per reaction, respectively.
The primers and probes did not cross-react with high titre respiratory viruses and bacteria listed in the materials and methods section resulting in 100% specificity.
The intra-assay coefficient of variation in Ct values for three clinical samples representing a range of viral loads was tested in triplicate on three independent runs. This parameter ranged from 0.08% to 6.06% with the H275 allele and 0.02–2.95% for H275Y samples. The inter-assay variability ranged from 0.56% to 2.13% and 0.27–1.01% with the H275 and H275Y alleles, respectively.
3.2. Accuracy of the one step RT-PCR assay
Accuracy of the RT-PCR assay was determined using 25 samples with C823, 15 with 823T and nine with a mixture of C and T nucleotides at position 823 by the gold standard Sanger sequencing method. Additionally, 20 samples that were negative for IFVA were also tested. The 24 specimens containing the H275Y allele and H275/H275Y admixtures populations were from 10 patients belong to immuno-compromised and intensive care unit cases identified in the described screening process previously.
The 20 IFVA negative samples were not detected by the assay indicating good primer and probe specificity. Of the 25 samples with the H275 allele, the SOIV_Osel_SEN probe resulted 22 as positive and three samples tested negative (Table 2a ). These samples had been tested previously by a screening assay for influenza targeting the matrix gene (InfA) (Dawood et al., 2009). The Ct values by this screening assay for the three discordant samples were 26.44, 35.48, and 36.79. A negative result suggests either a low viral load or sequence variation in the primer or probe binding region. The sequence of the assay detection region was determined for these samples and analyzed for the presence of mutations. The sample with a Ct value of 35.48 had no change in the probe binding region indicating that the sample possibly tested negative as a result of low viral load. The samples with Ct values of 26.44 and 36.79 were found to have a base pair mutation at different positions in the probe binding region (Fig. 2 ). Researchers using this region for the design of probes to detect oseltamivir resistance should be aware of the presence of these mutations in some isolates. These 25 samples were not detected by the SOIV_Osel_RES probe as expected by the probe design. Fig. 2 shows an alignment of the two SNP probes and mutations detected in this region.
Table 2a.
Performance of RT-PCR assay using SOIV_Osel_SEN probe in comparison to the gold standard Sanger sequencing method.
| Sequencing result for pandemic (H1N1) 2009 virus | Samples (N) | RT-PCR assay |
Sensitivity (percentage) | Specificity (percentage) | |
|---|---|---|---|---|---|
| + | − | ||||
| Sensitive (C823) | 25 | 22 | 3a | 88.0 | 100.0 |
| Resistant (823T) | 15b | 0 | 15 | 0.0 | 100.0 |
| Resistant admixtures (C823/C823T) | 9c | 8 | 1d | 88.9 | 100.0 |
| IFVA negative | 20 | 0 | 0 | 0.0 | 100.0 |
Three samples were not detected by the SOIV_Osel_SEN probe, one sample had a Ct of 35.48 by the InfA assay indicating low viral load; the other two samples had Ct values of 26.44 and 36.79, and were found to have a base pair mutation in the probe region.
Fifteen samples contained 100% T at base pair 823.
Nine samples had a mixed population of resistant and sensitive alleles (mixture of C and T at nucleotide position 823).
One admixture was not detected by the SOIV_Osel_SEN probe and showed T as the major nucleotide and C as the minor nucleotide present. It had a Ct value of 35.48 by the InfA assay indicating a low viral load.
Fig. 2.
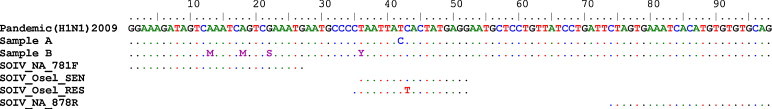
Alignment of primers and probes with oseltamivir-sensitive pandemic (H1N1) 2009 samples (samples A and B) that were not detected by the SOIV_Osel_SEN probe. Sample A had a T to C mutation and sample B had a mixture of C and T nucleotides at the variant position in the probe region.
Samples with the H275Y allele were present either as 823T (n = 15) or admixtures of C823 and 823T (n = 9). All of the 15 samples with only the 823T were detected by the SOIV_Osel_RES probe and not by the SOIV_Osel_SEN probe. Of the 9 admixtures, the SOIV_Osel_SEN probe detected eight as positive and the SOIV_Osel_RES probe detected seven as positive. One admixture that was not detected by the SOIV_Osel_SEN probe (but detected by the SOIV_Osel_RES probe) showed a major 823T population and a minor C823 population based on peak height in the electropherogram and also had a Ct value of 35.36 for detection of IFVA, indicating a low viral load (Table 2a). Two samples with admixtures that had minor populations of the 823T nucleotide present as determined by sequencing were not detected by the SOIV_Osel_RES probe (Table 2b ).
Table 2b.
Performance of RT-PCR assay using SOIV_Osel_RES probe in comparison to the gold standard Sanger sequencing method.
| Sequencing result for pandemic (H1N1) 2009 virus | Samples (N) | RT-PCR assay |
Sensitivity (percentage) | Specificity (percentage) | |
|---|---|---|---|---|---|
| + | − | ||||
| Sensitive (C823) | 25 | 0 | 25 | 0.0 | 100.0 |
| Resistant (C823T) | 15a | 15 | 0 | 100.0 | 100.0 |
| Resistant admixtures (C823/C823T) | 9 | 7 | 2b | 77.8 | 100.0 |
| Negative for IFVA | 20 | 0 | 0 | 0.0 | 100.0 |
Fifteen samples contained 100% T at nucleotide position 823.
Two admixtures had a low proportion of T at position 823 and were not detected by the SOIV_Osel_RES probe.
Assay sensitivity for the detection of samples with C823 by the SOIV_Osel_SEN probe was 88% and detection of 823T by the SOIV_Osel_RES probes was 100%. Sensitivity for the detection of samples with mixtures of C823 and 823T by SOIV_Osel_SEN probe and SOIV_Osel_RES probe was 88.9% and 77.8%, respectively. There was no non-specific detection of the H275 or H275Y alleles, thus both probes were 100% specific (Table 2a, Table 2b). Representative amplification curves for the detection of C823, 823T and a mixture of both nucleotides are shown in Fig. 3 . The sensitive and resistant samples showed a single amplification curve; whereas the admixtures had two detectable amplification curves as indicated in Fig. 3.
Fig. 3.
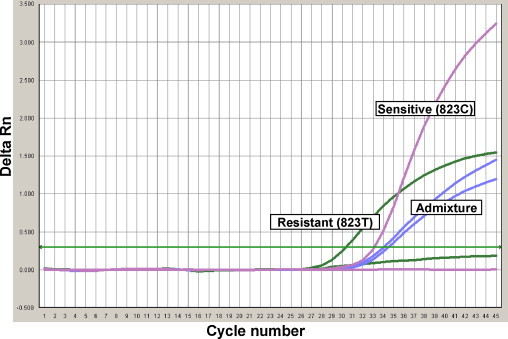
Amplification curves for templates containing sensitive (C823), resistant (C823T) and mixed alleles (C823 and C823T).
3.3. Detection of mixed alleles using in vitro transcribed RNA
To test the ability of the assay for detection of both sequence variants when they are present in various ratios and at different viral loads, a mixture of known template concentration of resistant and sensitive in vitro RNA was tested. Two mixtures were prepared, one at a low copy number with 4.76 × 102 and 5.29 × 102 copies/reaction and the other mixture with 4.76 × 105 and 5.29 × 105 copies/reaction for the sensitive and resistant RNA, respectively. In vitro RNA with the two sequence variants was mixed in ratios from 5% to 95% at both viral loads.
In the mixture prepared using higher copies of in vitro RNA, the SOIV_Osel_RES probe detected the presence of 823T when its proportion ranged from 20% to 100% of the total template concentration; this probe did not give a positive result when 823T was present at concentrations less than 20%. In the same mixtures, the presence of C823 was detected by SOIV_Osel_SEN probe when present at a concentrations ranging from 5% to 100% of total template as indicated in Table 3a .
Table 3a.
Detection of mixed templates prepared using quantitated in vitro transcribed RNA at a high copy number.
| % of template 823C RNA | % of template 823T RNA | Copies/reaction of total RNAa | Results for resistant probe | Results for sensitive probe |
|---|---|---|---|---|
| 100 | 0 | 4.76E+04 | Not detected | Detected |
| 95 | 5 | 4.78E+04 | Not detected | Detected |
| 80 | 20 | 4.87E+04 | Detected | Detected |
| 60 | 40 | 4.98E+04 | Detected | Detected |
| 50 | 50 | 5.03E+04 | Detected | Detected |
| 40 | 60 | 5.07E+04 | Detected | Detected |
| 20 | 80 | 5.18E+04 | Detected | Detected |
| 5 | 95 | 5.27E+04 | Detected | Detected |
| 0 | 100 | 5.29E+04 | Detected | Not detected |
Total copy number for both variants of in vitro RNA present in the reaction. Copy number for individuals alleles of in vitro RNA used to make the mixtures is indicated in Section 2.
For mixtures with low copies of in vitro RNA, the SOIV_Osel_RES probe detected the presence of 823T when its proportion ranged from 20% to 100% of the total template concentration; this probe did not give a positive result when 823T was present at concentrations less than 20%. In the same mixtures, the presence of C823 was detected by SOIV_Osel_SEN probe when present at a concentrations ranging from 20% to 100% of total template and presence of C823 at lower than 20% was not detected (Table 3b ).
Table 3b.
Detection of mixed templates prepared using quantitated in vitro transcribed RNA at a low copy number.
| % of template 823C RNA | % of template 823T RNA | Copies/reaction of total RNAa | Results for resistant probe | Results for sensitive probe |
|---|---|---|---|---|
| 100 | 0 | 4.76E+02 | Not detected | Detected |
| 95 | 5 | 4.78E+02 | Not detected | Detected |
| 80 | 20 | 4.87E+02 | Detected | Detected |
| 60 | 40 | 4.98E+02 | Detected | Detected |
| 50 | 50 | 5.03E+02 | Detected | Detected |
| 40 | 60 | 5.07E+02 | Detected | Detected |
| 20 | 80 | 5.18E+02 | Detected | Detected |
| 5 | 95 | 5.27E+02 | Detected | Not detected |
| 0 | 100 | 5.29E+02 | Detected | Not detected |
Total copy number for both variants of in vitro RNA present in the reaction. Copy number for individuals alleles of in vitro RNA used to make the mixtures is indicated in Section 2.
4. Discussion
Resistance to oseltamivir in seasonal and pandemic influenza A (H1N1) has been linked primarily to an H275Y polymorphism in NA glycoprotein (Okomo-Adhiambo et al., 2010, Sheu et al., 2008). The H275Y amino acid change was first reported in pandemic (H1N1) virus in June 2009 in an immuno-compromised patient treated with oseltamivir (WHO 2009 briefly note 18), and more cases have since been reported sporadically worldwide (Baz et al., 2009, Chen et al., 2009, Le et al., 2010). As there are still some questions as to whether or not this H275Y polymorphism impairs viral fitness and transmission (Seibert et al., 2010), we must be prepared for the possibility that oseltamivir-resistant isolates of the pandemic (H1N1) 2009 virus could expand in a manner similar to that seen in the previous seasonal H1N1 viruses (Baranovich et al., 2010, Besselaar et al., 2008, Hurt et al., 2009, Lackenby et al., 2008, Meijer et al., 2009, Sheu et al., 2008).
Screening tools should take into account the low prevalence of oseltamivir-resistance in the pandemic (H1N1) 2009 strain and ideally detection should be approached with cost effective and simple screening methods (Dawood et al., 2009, Harvala et al., 2010, Pabbaraju et al., 2009, Thabet et al., 2010). The current gold standard methods for detection of resistance include phenotypic detection using the NA-Star® assay from Applied Biosystems or genetic detection of relevant mutations using Sanger sequencing. Both of these methodologies are labor intensive, time-consuming and expensive. Additionally, phenotypic methods require viral culture which may not be feasible for novel sub-type influenza viruses that are restricted to high-level biosafety environments. Other methods such as reverse-transcriptase PCR coupled with restriction fragment length polymorphism assay (RTPCR-RFLP), pyrosequencing, microarray and multiplex RT-PCR with Luminex detection technologies have also been reported for detection of resistance but require specialized equipment and training (Bolotin et al., 2009, Carr et al., 2008, Deyde et al., 2009, Deyde et al., 2010, Guo et al., 2009, Hindiyeh et al., 2010, Mahony et al., 2010, Operario et al., 2010, van der Vries et al., 2010).
Real-time RT-PCR using probe technology is now commonplace in molecular diagnostic laboratories; these assays are high-throughput and amenable to large scale testing of patient samples with a quick turn-around time for results. Many laboratories are currently equipped to utilize SNP hydrolysis probes for the detection of oseltamivir resistance in pandemic (H1N1) 2009 virus. This approach has been reported previously for the detection of the C823T mutation in seasonal H1N1 (Bolotin et al., 2009, Carr et al., 2008, Operario et al., 2010) and pandemic (H1N1) 2009 viruses (Hindiyeh et al., 2010, van der Vries et al., 2010).
The possible caveats of using SNP probes to detect H275 and H275Y include the presence of mutations in the probe region adjacent to the SNP positions. This may result in false negative results; the presence of such mutations was detected in this study and has been published previously (Hindiyeh et al., 2010). To address this issue degenerate probes have been utilized by other investigators, however, the issue of cross reactivity and sensitivity was not investigated thoroughly (Hindiyeh et al., 2010). SNP assays may also be unable to detect minor populations in heterogenous mixtures of H275 and H275Y alleles within some specimens. Given the concerns about how H275Y SNP assays perform in the context of mixed viral populations and mutations in the probe-binding region, the authors feel that the results of a SNP assay should be interpreted with caution and ambiguous results should be confirmed by sequencing.
In conclusion, this manuscript describes the development and validation of a SNP assay for the detection of H275Y mutations associated with oseltamivir resistance in pandemic (H1N1) 2009 virus. The strength of this assay is its ability to identify H275Y mutations rapidly without specialized sequencing equipment. This tool is still relevant in the post-pandemic period as the pandemic strain is currently the circulating seasonal H1N1 influenza A strain of the 2010–2011 respiratory season.
Acknowledgements
We would like to thank the staff in Molecular Diagnostics and Virology departments at Provincial Laboratory for Public Health (ProvLab) for their technical assistance in undertaking routine respiratory virus testing. In particular, we thank Ms. Kara Gill and Ms Danielle Zarra for their excellent technical support for sequencing of samples.
References
- Baranovich T., Saito R., Suzuki Y., Zaraket H., Dapat C., Caperig-Dapat I., Oguma T., Shabana I.I., Saito T., Suzuki H. Emergence of H274Y oseltamivir-resistant A(H1N1) influenza viruses in Japan during the 2008–2009 season. J. Clin. Virol. 2010;47:23–28. doi: 10.1016/j.jcv.2009.11.003. [DOI] [PubMed] [Google Scholar]
- Baz M., Abed Y., Papenburg J., Bouhy X., Hamelin M.E., Boivin G. Emergence of oseltamivir-resistant pandemic H1N1 virus during prophylaxis. N. Engl. J. Med. 2009;361:2296–2297. doi: 10.1056/NEJMc0910060. [DOI] [PubMed] [Google Scholar]
- Besselaar T.G., Naidoo D., Buys A., Gregory V., McAnerney J., Manamela J.M., Blumberg L., Schoub B.D. Widespread oseltamivir resistance in influenza A viruses (H1N1), South Africa. Emerg. Infect. Dis. 2008;14:1809–1810. doi: 10.3201/eid1411.080958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolotin S., Robertson A.V., Eshaghi A., De L.C., Lombos E., Chong-King E., Burton L., Mazzulli T., Drews S.J. Development of a novel real-time reverse-transcriptase PCR method for the detection of H275Y positive influenza A H1N1 isolates. J. Virol. Methods. 2009;158:190–194. doi: 10.1016/j.jviromet.2009.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr M.J., Sayre N., Duffy M., Connell J., Hall W.W. Rapid molecular detection of the H275Y oseltamivir resistance gene mutation in circulating influenza A (H1N1) viruses. J. Virol. Methods. 2008;153:257–262. doi: 10.1016/j.jviromet.2008.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen H., Cheung C.L., Tai H., Zhao P., Chan J.F., Cheng V.C., Chan K.H., Yuen K.Y. Oseltamivir-resistant influenza A pandemic (H1N1) 2009 virus, Hong Kong, China. Emerg. Infect. Dis. 2009;15:1970–1972. doi: 10.3201/eid1512.091057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dawood F.S., Jain S., Finelli L., Shaw M.W., Lindstrom S., Garten R.J., Gubareva L.V., Xu X., Bridges C.B., Uyeki T.M. Emergence of a novel swine-origin influenza A (H1N1) virus in humans. N. Engl. J. Med. 2009;360:2605–2615. doi: 10.1056/NEJMoa0903810. [DOI] [PubMed] [Google Scholar]
- Deyde V.M., Xu X., Bright R.A., Shaw M., Smith C.B., Zhang Y., Shu Y., Gubareva L.V., Cox N.J., Klimov A.I. Surveillance of resistance to adamantanes among influenza A(H3N2) and A(H1N1) viruses isolated worldwide. J. Infect. Dis. 2007;196:249–257. doi: 10.1086/518936. [DOI] [PubMed] [Google Scholar]
- Deyde V.M., Okomo-Adhiambo M., Sheu T.G., Wallis T.R., Fry A., Dharan N., Klimov A.I., Gubareva L.V. Pyrosequencing as a tool to detect molecular markers of resistance to neuraminidase inhibitors in seasonal influenza A viruses. Antiviral Res. 2009;81:16–24. doi: 10.1016/j.antiviral.2008.08.008. [DOI] [PubMed] [Google Scholar]
- Deyde V.M., Sheu T.G., Trujillo A.A., Okomo-Adhiambo M., Garten R., Klimov A.I., Gubareva L.V. Detection of molecular markers of drug resistance in 2009 pandemic influenza A (H1N1) viruses by pyrosequencing. Antimicrob. Agents Chemother. 2010;54:1102–1110. doi: 10.1128/AAC.01417-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dharan N.J., Gubareva L.V., Meyer J.J., Okomo-Adhiambo M., McClinton R.C., Marshall S.A., St G.K., Epperson S., Brammer L., Klimov A.I., Bresee J.S., Fry A.M. Infections with oseltamivir-resistant influenza A(H1N1) virus in the United States. JAMA. 2009;301:1034–1041. doi: 10.1001/jama.2009.294. [DOI] [PubMed] [Google Scholar]
- Guo L., Garten R.J., Foust A.S., Sessions W.M., Okomo-Adhiambo M., Gubareva L.V., Klimov A.I., Xu X. Rapid identification of oseltamivir-resistant influenza A(H1N1) viruses with H274Y mutation by RT-PCR/restriction fragment length polymorphism assay. Antiviral Res. 2009;82:29–33. doi: 10.1016/j.antiviral.2009.01.004. [DOI] [PubMed] [Google Scholar]
- Harvala H., Gunson R., Simmonds P., Hardie A., Bennett S., Scott F., Roddie H., McKnight J., Walsh T., Rowney D., Clark A., Bremner J., Aitken C., Templeton K. The emergence of oseltamivir-resistant pandemic influenza A(H1N1) 2009 virus amongst hospitalised immunocompromised patients in Scotland, November–December, 2009. Euro. Surveill. 2010;15 [PubMed] [Google Scholar]
- Hauge S.H., Dudman S., Borgen K., Lackenby A., Hungnes O. Oseltamivir-resistant influenza viruses A (H1N1), Norway, 2007–2008. Emerg. Infect. Dis. 2009;15:155–162. doi: 10.3201/eid1502.081031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hindiyeh M., Ram D., Mandelboim M., Meningher T., Hirsh S., Robinov J., Levy V., Orzitzer S., Azar R., Grossman Z., Mendelson E. Rapid detection of influenza A pandemic (H1N1) 2009 virus neuraminidase resistance mutation H275Y by real-time reverse transcriptase PCR. J. Clin. Microbiol. 2010;48:1884–1887. doi: 10.1128/JCM.02540-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurt A.C., Ernest J., Deng Y.M., Iannello P., Besselaar T.G., Birch C., Buchy P., Chittaganpitch M., Chiu S.C., Dwyer D., Guigon A., Harrower B., Kei I.P., Kok T., Lin C., McPhie K., Mohd A., Olveda R., Panayotou T., Rawlinson W., Scott L., Smith D., D'Souza H., Komadina N., Shaw R., Kelso A., Barr I.G. Emergence and spread of oseltamivir-resistant A(H1N1) influenza viruses in Oceania, South East Asia and South Africa. Antiviral Res. 2009;83:90–93. doi: 10.1016/j.antiviral.2009.03.003. [DOI] [PubMed] [Google Scholar]
- Janies D.A., Voronkin I.O., Studer J., Hardman J., Alexandrov B.B., Treseder T.W., Valson C. Selection for resistance to oseltamivir in seasonal and pandemic H1N1 influenza and widespread co-circulation of the lineages. Int. J. Health Geogr. 2010;9:13. doi: 10.1186/1476-072X-9-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lackenby A., Hungnes O., Dudman S.G., Meijer A., Paget W.J., Hay A.J., Zambon M.C. Emergence of resistance to oseltamivir among influenza A(H1N1) viruses in Europe. Euro. Surveill. 2008;13 doi: 10.2807/ese.13.05.08026-en. [DOI] [PubMed] [Google Scholar]
- Le Q.M., Wertheim H.F., Tran N.D., van Doorn H.R., Nguyen T.H., Horby P. A community cluster of oseltamivir-resistant cases of 2009 H1N1 influenza. N. Engl. J. Med. 2010;362:86–87. doi: 10.1056/NEJMc0910448. [DOI] [PubMed] [Google Scholar]
- Mahony J.B., Chong S., Luinstra K., Petrich A., Smieja M. Development of a novel bead-based multiplex PCR assay for combined subtyping and oseltamivir resistance genotyping (H275Y) of seasonal and pandemic H1N1 influenza A viruses. J. Clin. Virol. 2010;49:277–282. doi: 10.1016/j.jcv.2010.08.006. [DOI] [PubMed] [Google Scholar]
- Mak A., Rahmanian R., Lei V., Lawrence D., Krajden M., Brunham R.C., Skowronski D., Li Y., Booth T., Goh S.H., Petric M. Longitudinal analysis of genotype distribution of influenza A virus from 2003 to 2005. J. Clin. Microbiol. 2006;44:3583–3588. doi: 10.1128/JCM.02601-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meijer A., Lackenby A., Hungnes O., Lina B., van-der-Werf S., Schweiger B., Opp M., Paget J., van-de-Kassteele J., Hay A., Zambon M. Oseltamivir-resistant influenza virus A (H1N1), Europe, 2007–2008 season. Emerg. Infect. Dis. 2009;15:552–560. doi: 10.3201/eid1504.081280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson M.I., Simonsen L., Viboud C., Miller M.A., Holmes E.C. The origin and global emergence of adamantane resistant A/H3N2 influenza viruses. Virology. 2009;388:270–278. doi: 10.1016/j.virol.2009.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okomo-Adhiambo M., Sleeman K., Ballenger K., Nguyen H.T., Mishin V.P., Sheu T.G., Smagala J., Li Y., Klimov A.I., Gubareva L.V. Neuraminidase inhibitor susceptibility testing in human influenza viruses: a laboratory surveillance perspective. Viruses. 2010;2:2269–2289. doi: 10.3390/v2102269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Operario D.J., Moser M.J., St. George K. Highly sensitive and quantitative detection of the H274Y oseltamivir resistance mutation in seasonal A/H1N1 influenza virus. J. Clin. Microbiol. 2010;48:3517–3524. doi: 10.1128/JCM.01031-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pabbaraju K., Wong S., Wong A.A., Appleyard G.D., Chui L., Pang X.L., Yanow S.K., Fonseca K., Lee B.E., Fox J.D., Preiksaitis J.K. Design and validation of real-time reverse transcription-PCR assays for detection of pandemic (H1N1) 2009 virus. J. Clin. Microbiol. 2009;47:3454–3460. doi: 10.1128/JCM.01103-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pabbaraju K., Wong S., Kits D.K., Fox J.D. Adamantane resistance in seasonal human influenza A viruses from Calgary, Alberta (January 2007–August 2008) Can. J. Infect. Dis. Med. Microbiol. 2010;21(2):e87–e91. doi: 10.1155/2010/710149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seibert C.W., Kaminski M., Philipp J., Rubbenstroth D., Albrecht R.A., Schwalm F., Stertz S., Medina R.A., Kochs G., Garcia-Sastre A., Staeheli P., Palese P. Oseltamivir-resistant variants of the 2009 pandemic H1N1 influenza A virus are not attenuated in the guinea pig and ferret transmission models. J. Virol. 2010;84:11219–11226. doi: 10.1128/JVI.01424-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheu T.G., Deyde V.M., Okomo-Adhiambo M., Garten R.J., Xu X., Bright R.A., Butler E.N., Wallis T.R., Klimov A.I., Gubareva L.V. Surveillance for neuraminidase inhibitor resistance among human influenza A and B viruses circulating worldwide from 2004 to 2008. Antimicrob. Agents Chemother. 2008;52:3284–3292. doi: 10.1128/AAC.00555-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thabet A.A., Al-Bahlooli S.H., Al-Kohlani A., Shoja’a A. Oseltamivir-resistant pandemic (H1N1)2009 in Yemen – case report. Virol. J. 2010;7:88. doi: 10.1186/1743-422X-7-88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Vries E., Jonges M., Herfst S., Maaskant J., Van der Linden A., Guldemeester J., Aron G.I., Bestebroer T.M., Koopmans M., Meijer A., Fouchier R.A., Osterhaus A.D., Boucher C.A., Schutten M. Evaluation of a rapid molecular algorithm for detection of pandemic influenza A (H1N1) 2009 virus and screening for a key oseltamivir resistance (H275Y) substitution in neuraminidase. J. Clin. Virol. 2010;47:34–37. doi: 10.1016/j.jcv.2009.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- World Health Organization First oseltamivir-resistant pandemic (H1N1) 2009 influenza from EMR. Wkly Epidemiol. Monit. 2009;2(44) [Google Scholar]
- World Health Organization, 2009b. Oseltamivir resistance in immunocompromised hospital patients pandemic (H1N1) 2009 breifing note 18. http://www.who.int/csr/disease/swineflu/notes/briefing_20091202/en/.
- Zaraket H., Saito R., Suzuki Y., Baranovich T., Dapat C., Caperig-Dapat I., Suzuki H. Genetic makeup of amantadine-resistant and oseltamivir-resistant human influenza A/H1N1 viruses. J. Clin. Microbiol. 2010;48:1085–1092. doi: 10.1128/JCM.01532-09. [DOI] [PMC free article] [PubMed] [Google Scholar]