

Abstract
At the molecular and cellular levels, viruses behave quite differently from bacteria and protozoa but to some extent, this distinction disappears when viruses are considered at the levels of the whole animal and the population. Viruses, like other infectious agents, enter the animal body through one of its surfaces. They then spread either locally on one of the body surfaces or through lymphatic and blood vessels to produce systemic infection. To infect its host, a virus first attaches to and infects cells of one of the body surfaces: the skin, the mucosa of the digestive, respiratory, or urogenital tract, or the conjunctiva. Parenteral injection, either by needle or by the bite of an arthropod or vertebrate, bypasses the body surfaces. A lesion may be produced at the site of entry, although invasion may occur without the development of any local evidence of infection. The infection may remain localized in the skin, respiratory tract, or digestive tract or it may spread through the animal via the lymphatics, blood vessels, or nerves.
At the molecular and cellular levels, viruses behave quite differently from bacteria and protozoa, but to some extent this distinction disappears when viruses are considered at the levels of the whole animal and the population. Viruses, like other infectious agents, must enter the animal body through one of its surfaces (or enter the fetus across the placenta or through the egg). They must then spread, either locally on one of the body surfaces, or through lymphatic and blood vessels to produce systemic infection. Finally, if the infection is to be maintained in nature, infectious virus must be shed into the environment, or taken up by an arthropod vector or a needle, or passed congenitally.
ROUTES OF ENTRY
In order to infect its host, a virus first attaches to and infects cells of one of the body surfaces: the skin, or the mucosa of the digestive, respiratory, or urogenital tract or the conjunctiva (Fig. 7-1 ). Parenteral injection, either by needle or by the bite of an arthropod or vertebrate, bypasses the body surfaces. A lesion may be produced at the site of entry, although invasion may occur without the development of any local evidence of infection. The infection may remain localized in the skin, respiratory tract, or digestive tract, or it may spread through the animal via the lymphatics, blood vessels, or nerves.
FIG. 7-1.
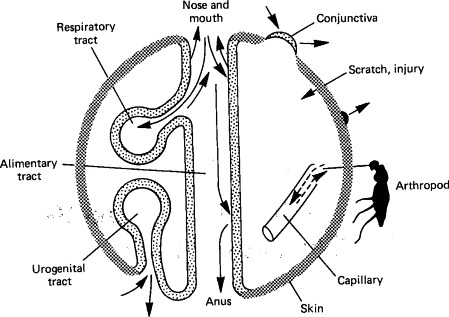
The surfaces of the body in relation to the entry and shedding of viruses.
(Modified from C. A. Mims and D. O. White, “Viral Pathogenesis and Immunology.” Blackwell Scientific Publications, Oxford, 1984.)
© 1987
Infection via the Skin
The largest organ in the body, the skin, provides a tough and impermeable barrier to the entry of viruses. However, some viruses replicate in the skin to produce local lesions, after entry through minor abrasions or by artificial puncture (Table 7-1 ). Several poxviruses—e.g., cowpox, orf, and fowlpox, and the papillomaviruses—cause local skin lesions. The most efficient way by which viruses are introduced through the skin is by the bite of an arthropod vector, such as a mosquito, tick, sandfly, or Culicoides. Such insects may be mechanical vectors (e.g., for myxomatosis and fowlpox), but most viruses introduced in this way replicate in the vector. Viruses that are transmitted by and replicate in arthropod vectors are called arboviruses, a term that carries no taxonomic implications. Infection can be acquired through the bite of an animal, as in rabies. Finally, introduction of a virus by skin penetration may be iatrogenic—the result of human intervention, e.g., transmission, of equine infectious anemia by a contaminated needle. Generalized infection of the skin (exanthema), such as is found in lumpyskin disease, sheeppox, and swine vesicular disease, for example, is due to viral spread via the bloodstream.
TABLE 7-1.
Viruses of Animals That Initiate Infection via the Skin, Oral Mucosa, Genital Tract, or Eye
| Route | Family or genus | Viruses |
|---|---|---|
| Minor abrasion | Poxviridae | Cowpox, swinepox, orf, bovine papular stomatitis, pseudocowpox, fowlpox viruses |
| Herpesviridae | All viruses | |
| Papillomavirus | All viruses | |
| Picornaviridae | Swine vesicular disease virus | |
| Arthropod bite Mechanical | Poxviridae | Fowlpox, swinepox, myxoma viruses |
| Herpesviridae | Marek's disease virus | |
| Retroviridae | Equine infectious anemia virus | |
| Biological | Alphavirus | All viruses |
| Flaviviridae | All viruses | |
| Bunyaviridae | Rift Valley fever, Nairobi sheep disease viruses | |
| Rhabdoviridae | Bovine ephemeral fever, vesicular stomatitis viruses | |
| Orbivirus | Bluetongue, African horse sickness viruses | |
| Unclassified | African swine fever virus | |
| Bite of vertebrate | Rhabdoviridae | Rabies virus |
| Contaminated needle or equipment | Papillomavirus | All viruses |
| Togaviridae | Hog cholera, bovine virus diarrhea viruses | |
| Retroviridae | Equine infectious anemia, bovine leukemia viruses | |
| Genital tract | Herpesviridae | Many viruses |
| Papillomavirus | Bovine papilloma virus | |
| Togaviridae | Equine arteritis virus | |
| Conjunctiva | Herpesviridae | Infectious bovine rhinotracheitis virus, equine herpesvirus 1 |
Infection via the Respiratory Tract
Though lined by cells that are susceptible to infection by many viruses, the respiratory tract is ordinarily protected by effective defense mechanisms. A mucus blanket and ciliary escalator line the nasal cavity and most of the lower respiratory tract. Thus, inhaled foreign particles, including viral particles, deposited on this surface are trapped in mucus and carried by ciliary action from the nasal cavity and airways to the pharynx and then swallowed. Particles greater than 10 μm in diameter are usually deposited on the nasal mucosa over the turbinate bones, which project into the nasal cavity and act as baffle plates. Particles 5–10 μm in diameter may be carried to the trachea and bronchioles, where they are usually trapped in the mucociliary blanket. Smaller particles (≤5μm) are usually inhaled directly into the lungs and some may reach the alveoli, where virions may be destroyed by alveolar macrophages, or may infect such cells, or cells of the adjacent alveolar epithelial lining.
Despite these protective mechanisms, the respiratory tract is, overall, the most important entry site of viruses into the body (Table 7-2 ). Not only is the respiratory tract susceptible to infection via aerosols or particles that are inhaled, but infection may occur via fomites (i.e., contaminated objects), a mode of transfer well exemplified by the role of fingers in transferring rhinoviruses between humans but doubtless operative in other animals also.
TABLE 7-2.
Some Viruses of Animals That Initiate Infection via the Respiratory Tract
| Family | Viruses |
|---|---|
| Producing respiratory disease | |
| Herpesviridae | Most viruses |
| Adenoviridae | Most viruses |
| Parvoviridae | Feline panleukopenia virus, canine parvovirus |
| Picornaviridae | Rhinoviruses, aphthoviruses |
| Caliciviridae | Feline calicivirus |
| Orthomyxoviridae | Influenza viruses |
| Paramyxoviridae | Parainfluenza, respiratory syncytial viruses |
| Producing systemic disease, usually without initial respiratory signs | |
| Herpesviridae | Pseudorabies, bovine malignant catarrhal fever, Marek's disease |
| viruses | |
| Togaviridae | Hog cholera virus |
| Paramyxoviridae | Canine distemper, rinderpest, Newcastle disease viruses |
| Arenaviridae | Lymphocytic choriomeningitis virus |
Many viruses remain localized in the respiratory tract, but some viruses that enter via the respiratory tract produce their principal effects following systemic spread (e.g., foot-and-mouth disease, canine distemper, rinderpest, hog cholera, and Newcastle disease viruses).
Infection via the Digestive Tract
Many viruses, belonging to several families, are acquired by ingestion. They may either be swallowed or infect cells in the pharynx. The esophagus is rarely infected initially, probably because of its tough stratified squamous epithelium and the rapid passage of swallowed material over its surface. The intestinal tract is partially protected by mucus, which may contain secretory IgA antibodies, but the constant movement of the contents provides opportunities for virions to attach to specific receptors. Virions may also be taken up by specialized cells that overlie Peyer's patches in the ileum, from which they are passed to adjacent mononuclear cells in which they may be able to replicate.
There are other protective mechanisms in the intestinal tract, because substances involved in the digestion of food may also destroy viruses; from the stomach caudally, these are acid, bile, and proteolytic enzymes. In general, viruses that cause intestinal infection, such as enteroviruses, rotaviruses, caliciviruses, and parvoviruses (Table 7-3 ), are resistant to acid and bile. However, there are examples of acid- and bile-labile viruses that cause important intestinal infections. Bovine, porcine, and murine coronaviruses are protected by the buffering capacity of milk during passage through the stomach of young animals, and avian influenza viruses are assisted by the buffering capacity of food to survive passage through the upper digestive tract of birds. Because of the destructive effects of acid, bile, and proteolytic enzymes on viruses, dosage is important in determining whether infection will occur.
TABLE 7-3.
Some Viruses of Animals That Initiate Infection via the Intestinal Tract
| Family | Viruses |
|---|---|
| Producing scours | |
| Coronaviridae | Some coronaviruses |
| Reoviridae | Rotaviruses |
| Unclassified | Astroviruses, toroviruses |
| Producing systemic disease, usually without local signs | |
| Adenoviridae | Some adenoviruses |
| Caliciviridae | Vesicular exanthema of swine virus |
| Picornaviridae | Enteroviruses |
Coronaviruses, rotaviruses, parvoviruses, and caliciviruses are now recognized as the major causes of viral diarrhea. The great majority of intestinal infections by enteroviruses and adenoviruses are asymptomatic. Some of the enteroviruses (e.g., porcine, avian, and murine encephalomyelitis viruses) are important causes of generalized infection but do not produce signs referable to the intestinal tract. Parvoviruses cause diarrhea after reaching cells of the intestinal tract via viremic spread.
Infection by Other Routes
The genital tract (in coitus) is the route of entry of several important pathogens, e.g., bovine herpesvirus 1, equine herpesvirus 3, and porcine papillomavirus. The conjunctiva, though much less resistant to viral invasion than the skin, is constantly cleansed by the flow of secretion (tears) and is wiped by the eyelids. The conjunctiva is unproven as a portal of entry of viruses in animals, although experimentally infection with a wide range of viruses can be achieved via this route.
MECHANISMS OF SPREAD IN THE BODY
Viruses may remain localized to the body surface through which they entered (i.e., skin, respiratory tract, intestine, genital tract, or conjunctiva), or they may cause generalized infections, which are usually associated with viremia and subsequent localization in particular organs.
Local Spread in Epithelia
Many viruses, e.g., poxviruses and papillomaviruses, replicate in epithelial cells at the site of entry, produce a localized or spreading infection in the epithelium, and are then shed directly into the environment. Infection spreads by sequential infection of neighboring cells. Papillomaviruses initiate infection in the basal layer of the epidermis, but maturation, with the production of virions, occurs only when the cells become keratinized as they move toward the skin surface. Since this is a slow process, taking several weeks, warts have a long incubation period. Many poxviruses produce infection via the skin. In contrast to the papillomaviruses, however, there is also subepithelial and lymphatic spread. In infection with vaccinia virus, which may become as common for domestic animals as it used to be in humans if vaccinia is used as a vector for veterinary vaccines (see Chapter 14), a few epidermal cells are infected by scarification and virus spreads locally from cell to cell, primarily in the epidermis, before spreading to the local lymph nodes. Some poxviruses, e.g., orf, rabbit fibroma, and Yaba poxvirus, remain localized in the skin and produce a proliferative response and a localized tumor.
In contrast to the skin, viruses that enter the body via the mouth or respiratory tract can spread rapidly over the moist epithelial surfaces. After infections of the respiratory tract by paramyxoviruses and influenza virus, or the intestinal tract by rotaviruses or coronaviruses, there is little or no invasion beyond the epithelium. Although these viruses usually enter the lymphatics and thus have the potential to spread, they do not appear to replicate in the deeper tissues, possibly because the appropriate virus receptors are restricted to epithelial cells. However, restriction to an epithelial surface cannot be equated with lack of severity of clinical disease. Large areas of intestinal epithelium may be damaged by rotaviruses or coronaviruses, for example, causing severe diarrhea. The severity of localized infections of the respiratory tract depends on their location: infections of the upper respiratory tract may produce severe rhinitis but few other signs; infection of the bronchioles or alveoli produces more severe respiratory distress and may predispose to secondary bacterial invasion.
Subepithelial Invasion and Lymphatic Spread
After traversing the epithelium to reach the subepithelial tissues, virions are immediately exposed to tissue macrophages and can enter lymphatics (Fig. 7-2 ). Macrophages play an important role in controlling viral infections in many sites in the body and are discussed below; suffice it to say here that they phagocytose and destroy most viruses.
FIG. 7-2.
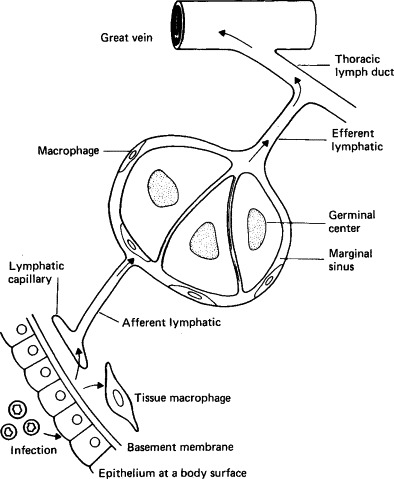
Subepithelial invasion and lymphatic spread of viruses.
(From C. A. Mims and D. O. White, “Viral Pathogenesis and Immunology.” Blackwell Scientific Publications, Oxford, 1984.)
© 1987
A network of lymphatics lies beneath the skin and all mucosal epithelia. Virions that enter lymphatics are carried to local lymph nodes. As they enter, they are exposed to macrophages lining marginal sinuses and may be engulfed. Virions may be inactivated and processed and their component antigens presented to adjacent lymphocytes in such a way that an immune response is initiated (see Chapter 9). Some viruses, however, replicate in macrophages (e.g., canine distemper virus, some adenoviruses, and some herpesviruses). Some virions may pass straight through lymph nodes to enter the bloodstream.
There is often a local inflammatory response, the extent of which depends on the extent of tissue damage. Local blood vessels are dilated and rendered more permeable, so that macrophages and lymphocytes, immunoglobulins, and complement components can be delivered directly to the site of infection, especially after the immune response has been initiated.
Spread by the Bloodstream
Once a virus has reached the bloodstream, usually via the lymphatic system (Fig. 7-2), it can localize in any part of the body within minutes. The presence of virus in the blood is called viremia. In some infections it is possible to distinguish a primary from a secondary viremia. A primary viremia follows directly after entry of virions via the lymphatics (Fig. 7-3 ); usually it is recognized only because of the invasion of distant organs. Further replication in these sites leads to the sustained liberation of much higher concentrations of virus, producing a secondary viremia, which can in turn lead to the establishment of infection in yet other parts of the body.
FIG. 7-3.
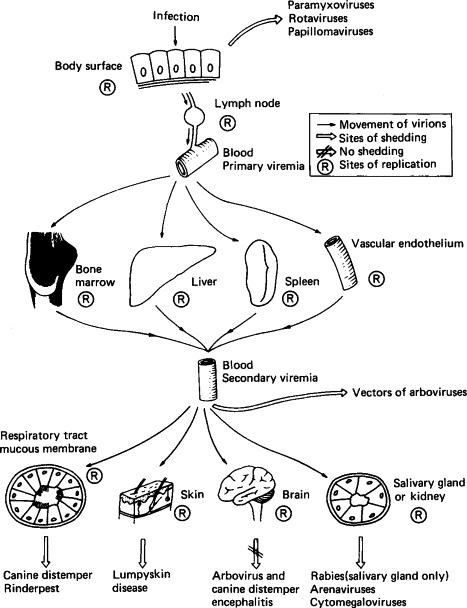
Spread of virions through the body, indicating sites of replication and important routes of shedding of various viruses.
In the blood, virions may be free in the plasma or may be associated with particular types of leukocytes, with platelets, or with erythrocytes. Leukocyte-associated viremia is a feature of several types of infection, including canine distemper, bluetongue, and many herpesvirus infections, for example. As already mentioned, several viruses replicate in macrophages; others, e.g., Marek's virus, replicate in lymphocytes. Rarely, as in African swine fever and Colorado tick fever, virions may be associated with erythrocytes. All the togaviruses and flaviviruses, and the enteroviruses that cause viremia, circulate free in the plasma.
Virions circulating in the bloodstream encounter many kinds of cells, but two kinds play a special role in determining their subsequent fate and thus merit special consideration; macrophages and vascular endothelial cells.
Role of Macrophages
The term “reticuloendothelial system” is used to designate macrophages, fixed and circulating, throughout the body. These very efficient phagocytes are present in all compartments of the body, in alveoli, subepithelial tissues, sinusoids of the lymph nodes, free in plasma, and above all in the sinusoids of the liver, spleen, and bone marrow. The kinds of interactions that may occur between macrophages and virions can be described in relation to those found in the sinusoids of the liver, the Kupffer cells (Fig. 7-4 ; the numbers in the list refer to the circled numbers in this diagram).
-
1.
Virions may resist phagocytosis by macrophages. This occurs in Venezuelan equine encephalitis virus infection, and is a factor that favors prolonged viremia.
-
2.
Virions may be phagocytosed and destroyed. This is the usual fate of picornaviruses that circulate in the plasma. Since the macrophage system is so efficient, viremia with such viruses can be maintained only if virions enter the blood as fast as they are removed.
-
3.
Virions may be phagocytosed and then passively transferred to adjacent cells (hepatocytes in the liver). If the virus cannot replicate in hepatocytes the situation resembles (2) above; if, like Rift Valley fever virus, the virus replicates in these cells it can cause clinical hepatitis and the virus produced in the liver can sustain a high level of viremia.
-
4.
Virions may be phagocytosed by macrophages and then replicate in them. With some viruses, such as lactic dehydrogenase virus in mice, only macrophages are infected [Fig. 7-4 (4A)] and progeny virions enhance the viremia, which reaches an extremely high level. More commonly [Fig. 7-4 (4B)], as in infectious canine hepatitis, virus replicates in both macrophages and hepatic cells, producing severe hepatitis.
FIG. 7-4.
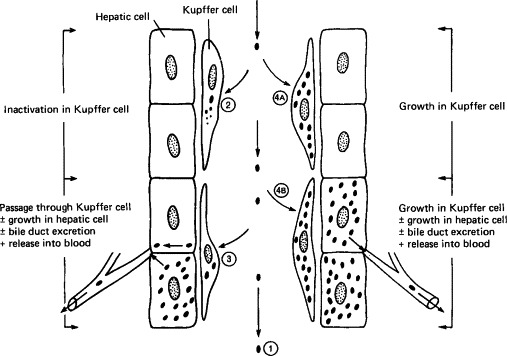
Types of interaction between viruses and macrophages, exemplified by the Kupffer cells lining a sinusoid in the liver.
(Modified from C. A. Mims and D. O. White, “Viral Pathogenesis and Immunology.” Blackwell Scientific Publications, Oxford, 1984.)
© 1987
The situation in vivo is much more complex than this simple scheme suggests, since macrophages comprise several subpopulations that differ in their properties according to their site in the body and depending on whether they have been activated.
Vascular Endothelial Cells
The vascular endothelium with its basement membrane and tight cell junctions constitutes the blood-tissue interface—for particles such as virions, often a barrier. Since most tissues and organs lack sinusoids, parenchymal invasion by circulating virions depends on localization in the endothelial cells of capillaries and venules, where blood flow is slowest and the barrier thinnest. Virions may move passively between or through endothelial cells and basement membrane, or they may infect endothelial cells and “grow” through this barrier. This subject has been most intensively studied in relation to viral invasion of the central nervous system (see below), but it also applies to secondary invasion of the skin, pulmonary epithelium, salivary gland epithelium, intestinal epithelium, kidney, and placenta.
Maintenance of Viremia
Since virions circulating in the blood are continuously removed by cells of the reticuloendothelial system, viremia can be maintained only (1) if there is a continuing introduction of virus into the blood from infected tissues, or (2) if there is impairment of the reticuloendothelial system. Circulating leukocytes can themselves constitute a site for viral replication; indeed, blood leukocytes maintained in culture support replication of many viruses. However, viremia is usually maintained by infection of the parenchymal cells of organs with extensive sinusoids, like the liver, spleen, lymph nodes, and bone marrow. In some infections (e.g., hog cholera) the viremia is partly maintained by infection of endothelial cells. Striated and smooth muscle cells may be an important site of replication of some enteroviruses, togaviruses, and rhabdoviruses; virions are transferred to the blood via the lymph.
Invasion of the Skin
As well as being a site of initial infection, the skin may be invaded via the bloodstream, producing a rash. Rashes are more easily seen in humans than other animals, where fur or feathers usually restrict observation. The individual lesions in generalized rashes are described as macules, papules, vesicles, or pustules. A lasting local dilation of subpapillary dermal blood vessels produces a macule, which becomes a papule if there is also edema and infiltration of cells into the area. Primary involvement of the epidermis or separation of epidermis from dermis by fluid pressure results in vesiculation. Erosion or sloughing of the epithelium results in ulceration and scabbing, but prior to ulceration a vesicle may be converted to a pustule by cellular infiltration. More severe involvement of the dermal vessels may lead to petechial or hemorrhagic rashes, although coagulation defects and thrombocytopenia may also be important in the genesis of such lesions.
Invasion of the Central Nervous System
Because of its critical physiological importance and its vulnerability to damage by any process that damages neurons directly or via increased intracranial pressure, viral invasion of the central nervous system is always a serious matter. Viruses can spread from the blood to the brain by two routes: via the bloodstream (Fig. 7-5 ) or via peripheral nerve fibers (Fig. 7-6 ). When invasion occurs from the bloodstream, the early stages in pathogenesis (upper part of Fig. 7-5) are the same as in all generalized infections (see Fig. 7-3). Viruses can pass from the blood into the brain or cerebrospinal fluid at several anatomical sites and by several different mechanisms. Although the cerebral capillaries represent a morphological blood-brain barrier, most viruses that invade the central nervous system cross these vessels. Some viruses infect the vascular endothelial cells prior to infection of the cells of the brain parenchyma; others appear to be transported across the capillary walls without endothelial cell infection. Rarely, virus may be carried across capillary walls into the brain parenchyma via infected leukocytes.
FIG. 7-5.
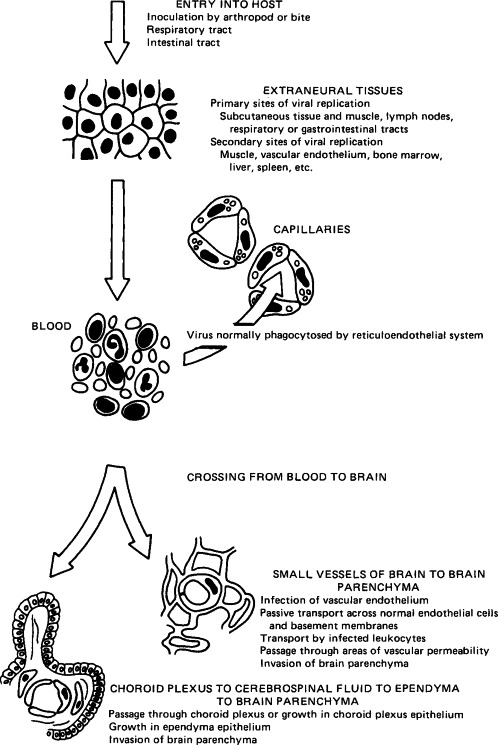
Steps in the hematogenous spread of virus into the central nervous system.
(Modified from R. T. Johnson, “Viral Infections of the Nervous System.” Raven Press, New York, 1982.)
© 1987
FIG. 7-6.
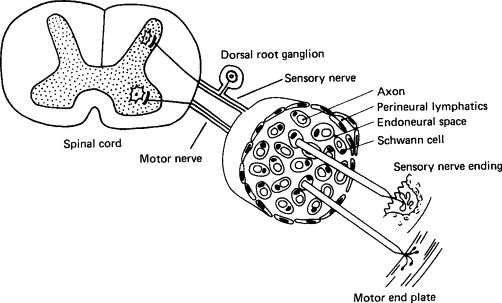
Neural pathways of central nervous system infection. Virions can be taken up at sensory, motor, or autonomic nerve endings and moved centripetally within axons, endoneural space, or perineural lymphatics, or by infection of Schwann cells. If transport is via axons, viruses taken up at sensory nerve endings will be delivered selectively to dorsal root ganglia neurons and then to the spinal cord; they may then move via the nerve tracts to the brain stem. Viruses taken up at motor nerve endings will be delivered to ventral nerve roots in the spinal cord and then to the brain stem.
(Modified from R. T. Johnson, “Viral Infections of the Nervous System.” Raven Press, New York, 1982.)
© 1987
Viruses that cause meningitis rather than encephalitis may traverse the blood–cerebrospinal fluid junction in the meninges or choroid plexus or may grow in the epithelium of the choroid plexus, thus causing an alteration in the delicately balanced osmotic pressure of the interstitial spaces of the brain. Interstitial edema in the brain may, in turn, open the way to more extensive viral spread.
The other important route of infection of the central nervous system is via the peripheral nerves, as seen, for example, in rabies, herpes B virus encephalitis, and pseudorabies in secondary hosts (Fig. 7-6). Herpesvirus particles travel to the central nervous system in axon cytoplasm, but while doing so also sequentially infect the Schwann cells of the nerve sheath. Rabies virus also travels to the central nervous system in axon cytoplasm, but usually does not infect the nerve sheath. Sensory, motor, and autonomic nerves may be involved in the neural spread of these viruses. Rabies and some togaviruses are able to use the olfactory nerve for movement to the central nervous system; they can infect olfactory neuroepithelial cells, which are exposed in the epithelium of the nares and have their axonal endings in the olfactory bulb of the brain.
Lytic infections of neurons, whether due to togaviruses, herpesviruses, or other viruses, lead to the three histological hallmarks of encephalitis: neuronal necrosis, phagocytosis of neurons by phagocytic cells (neuronophagia), and perivascular infiltration of inflammatory cells (perivascular cuffing)—the latter a manifestation of cell-mediated immunity (see Chapter 9). The cause of clinical neurological signs in other central nervous system infections is more obscure. Rabies virus infection is noncytocidal; it evokes little of the inflammatory reaction or cell necrosis found in other encephalitides, yet it is highly lethal for most mammalian species. With certain other viruses, infection of neurons causes no clinical signs; for example, the extensive central nervous system infection of mice congenitally infected with lymphocytic choriomeningitis virus, which is readily demonstrable by fluorescent-antibody staining, has no recognizable deleterious effect. Still other pathological changes are produced by some of the viruses that cause slowly progressive diseases of the central nervous system (see Chapter 11). In scrapie of sheep, for example, there is slow neuronal degeneration and vacuolization; in visna (a chronic retrovirus infection of sheep), changes in glial cell membranes lead to demyelination.
Invasion of Other Organs
Almost any organ may be infected via the bloodstream with one or another kind of virus, but most viruses have well-defined organ and tissue tropisms. The clinical importance of infection of various organs and tissues depends in part on their role in the economy of the body. Thus invasion of the liver, causing severe hepatitis, as in Rift Valley fever and infectious canine hepatitis, is a life-threatening situation. The critical importance of such organs as the brain, heart, and lungs is self-evident.
Infection of the testis or accessory sexual organs may lead to excretion of virus in the semen and the risk of transmission during coitus (see Table 7-5 ). The lungs may be infected via the bloodstream as well as the airways. Sometimes viremic infections reach the respiratory epithelium, causing bronchitis and pneumonia, with shedding of virus in the respiratory secretions; such infections, like airways-derived infections, may cause an interstitial pneumonia.
TABLE 7-5.
Some Viruses Excreted in the Semen of Domestic Animals
| Family | Virus | Animal | Comments |
|---|---|---|---|
| Herpesviridae | Infectious bovine rhinotracheitis virus | Cattle | Virus persists in ganglia, reinfecting genital tract with excretion in semen intermittently |
| Retroviridae | Mammary tumor virus | Mouse | Virus transmitted via sperm in some strains of mice |
| Bovine leukemia virus | Cattle | Virus persists in lymphoid tissues and leukocytes, with excretion in semen continuously | |
| Reoviridae | Bluetongue viruses | Cattle | Virus infects leukocytes and is excreted in semen continuously or intermittently as long as bull is viremic |
| Picornaviridae | Foot-and-mouth disease virus | Cattle | Virus infects genital tract with excretion in semen |
Delivery of circulating virus to the salivary glands or mammary glands may lead to lesions in those organs and excretion in the saliva or milk. Infection of muscle cells occurs with several togaviruses and cox-sackieviruses, while infection of the synovial cells of goats by caprine arthritis–encephalomyelitis virus produces arthritis.
Infection of the Fetus
Many viral infections of the dam have no harmful effect on the fetus, but some viruses regularly cross the placenta to reach the fetal circulation, sometimes after establishing foci of infection in the placenta. Severe cytolytic infections of the fetus cause fetal death and resorption or abortion, outcomes which are common in, for example, pseudorabies and parvovirus infections in swine. Also important are the teratogenic effects of less lethal viruses like bovine virus diarrhea virus and Akabane virus infections in cattle (Table 7-4 ).
TABLE 7-4.
Some Congenital Viral Infections in Domestic Animals
| Animal | Viral family | Virus | Syndrome |
|---|---|---|---|
| Cattle | Herpesviridae | Infectious bovine rhinotracheitis virus | Fetal death, abortion |
| Togaviridae | Bovine virus diarrhea virus | Fetal death, abortion, congenital defects, inapparent infection with lifelong carrier state and shedding | |
| Bunyaviridae | Akabane virus | Fetal death, abortion, stillbirth, congenital defects | |
| Retroviridae | Bovine leukemia virus | Inapparent infection, leukemia | |
| Reoviridae | Bluetongue viruses | Fetal death, abortion, congenital defects | |
| Horse | Herpesviridae | Equine herpesvirus 1 | Fetal death, abortion, neonatal disease |
| Togaviridae | Equine arteritis virus | Fetal death, abortion | |
| Swine | Herpesviridae | Pseudorabies virus | Fetal death, abortion |
| Parvoviridae | Parvovirus | Fetal death, abortion, mummification, stillbirth, infertility | |
| Flaviviridae | Japanese encephalitis virus | Fetal death, abortion | |
| Togaviridae | Hog cholera virus | Fetal death, abortion, congenital defects, inapparent infection with lifelong carrier state | |
| Sheep | Togaviridae | Border disease virus | Congenital defects |
| Bunyaviridae | Rift Valley fever virus | Fetal death, abortion | |
| Nairobi sheep disease virus | Fetal death, abortion | ||
| Reoviridae | Bluetongue virus | Fetal death, abortion, congenital defects | |
| Dog | Herpesviridae | Canine herpesvirus 1 | Perinatal death |
| Cat | Parvoviridae | Feline panleukopenia virus | Cerebellar hypoplasia |
| Retroviridae | Feline leukemia virus | Inapparent, leukemia, fetal death | |
| Mouse | Parvoviridae | Rat virus | Fetal death |
| Arenaviridae | Lymphocytic choriomeningitis virus | Inapparent, with lifelong carrier state | |
| Chicken | Picornaviridae | Avian encephalomyelitis virus | Congenital defects, fetal death |
| Retroviridae | Avian leukosis–sarcoma viruses | Inapparent, leukemia, other disease |
Generally, infection in the first and second trimesters of gestation is most damaging. Little is known of the pathogenesis of most fetal infections, but experimental studies of bluetongue virus and Akabane virus infections in bovine fetuses, and parvovirus infections in swine fetuses have led to some general understanding. The source of the virus is most often from maternal infection with viremia and transplacental passage.
In the second half of pregnancy, the course of most fetal infections is influenced by the developing fetal immune response. For example, infections with porcine parvovirus after days 65–70 of gestation result not in fetal death but in fetal antibody production and recovery. When bovine virus diarrhea virus infects the fetus in the first half of gestation, immune tolerance with an inapparent, lifelong carrier state may develop; infection late in gestation usually leads to an effective immune response. When viral replication in the fetus is rapid, as in alphaherpesvirus infections of horse, cow, pig, and dog, fetal death and abortion can occur even during the last trimester, because the immune response is inadequate.
Germ-line transmission of the endogenous retroviruses, as integrated proviral DNA, occurs in many species of animals.
VIRUS SHEDDING
The shedding of infectious virions maintains infection in populations of animals (see Chapter 15, on epidemiology), and usually occurs via one of the body openings or surfaces involved in the entry of viruses. With localized infections the same openings are involved in both entry and exit (see Fig. 7-1); in generalized infections a greater variety of modes of shedding is recognized (see Fig. 7-3).
Skin
Although skin lesions are produced in several generalized diseases, only a few viruses are shed from skin lesions in a way that leads to transmission. Virions are not excreted from the skin lesions associated with most picornavirus, togavirus, or flavivirus infections. Foot-and-mouth disease, vesicular stomatitis, poxvirus, and some herpesvirus infections, on the other hand, produce vesicular or pustular lesions from which virus is readily shed. Even here, however, virus shed in saliva and aerosols is often more important, as far as transmission is concerned, than that shed via the skin lesions.
The skin is an important source of virus in diseases in which transmission is by direct contact (rubbing) via small abrasions, e.g., papillomatosis. Several poxviruses may be spread from animal to human, and sometimes from human to animal, by contact with skin lesions, e.g., the viruses of cowpox, vaccinia, orf, and pseudocowpox. Localization of virus in the feather follicles is important in providing a mechanism for the shedding of Marek's disease virus from infected chickens.
Respiratory Tract
Many different viruses that cause either localized disease of the respiratory tract or generalized infections are shed in fluid expelled from the respiratory tract. Large droplets fall rapidly to the ground or may contaminate various fomites. Smaller droplets, with their relatively large surface-to-volume ratio, evaporate rapidly and are reduced to small droplet nuclei which remain airborne. Respiratory viruses, like infectious bovine rhinotracheitis virus, paramyxoviruses, some coronaviruses, and bovine respiratory syncytial virus, are excreted in both nasal and oral secretions. Shedding may continue to occur during convalescence or recurrently after that time, especially with herpesviruses.
Digestive Tract
Enteric viruses are shed in the feces and are in general more resistant to inactivation by environmental conditions than are the respiratory viruses, especially when suspended in water, e.g., in dams and troughs contaminated with feces. Thus, unlike the respiratory viruses, which usually spread directly from infected to susceptible animals, enteric viruses can persist for some time outside the body, especially as some, like rotaviruses and enteroviruses, are relatively heat stable.
Genital Tract
There may be transfer of virus from male to female and vice versa during coitus, and several viruses that cause important diseases of cattle and sheep are excreted in the semen (Table 7-5). Artificial insemination and embryo transfer are being promoted to avoid the introduction of sexually transmitted viruses into herds, as well as to avoid the risk of introduction of exotic viruses into disease-free countries. Knowledge of the transmission characteristics of viruses in semen and ova is extremely important in this regard.
Other Routes of Shedding
Urinary Tract
Urine, like feces, tends to contaminate food supplies and housing. A number of viruses, e.g., rinderpest, infectious canine hepatitis, and foot-and-mouth disease viruses, replicate in tubular epithelial cells in the kidney and are shed in the urine. Viruria is lifelong in arenavirus infections of rodents and constitutes the principal mode of contamination of the environment by these viruses.
Milk
Several kinds of viruses are excreted in milk, which may serve as a route of transmission of caprine arthritis-encephalitis virus, mouse mammary tumor virus, and some of the tick-borne flaviviruses. Foot-and-mouth disease virus can be spread by feeding calves with infected milk.
Blood and Internal Organs
Blood is important as the source from which arthropods acquire arboviruses, and blood may also be the route of transfer of viruses to the ovum or fetus. Equine infectious anemia virus and bovine leukemia virus are often transmitted by needles and other equipment contaminated with blood. Carnivores and omnivores may be infected by consuming virus-containing meat; e.g., hog cholera, African swine fever, and vesicular exanthema of swine viruses are often transmitted to swine that eat garbage containing contaminated pork scraps.
FURTHER READING
- Johnson R.T. “Viral Infections of the Nervous System.”. Raven Press; New York: 1982. [Google Scholar]
- Mims C.A. Vertical transmission of viruses. Microbiol. Rev. 1981;45(267) doi: 10.1128/mr.45.2.267-286.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mims C.A., White D.O. “Viral Pathogenesis and Immunology.”. Blackwell Scientific Publications; Oxford: 1984. [Google Scholar]
- Notkins A.L., Oldstone M.B.A., editors. Springer-Verlag; New York: 1984. (“Concepts in Viral Pathogenesis.”). [Google Scholar]
- Smith H., Sweet C. The pathogenicity of viruses. In: Brown F., Wilson G., editors. Vol. 4. Williams and Wilkins; Baltimore: 1985. p. 94. (In “Topley and Wilson's Principles of Bacteriology, Virology and Immunology”). 7th edition. [Google Scholar]
- Tyrrell D.A.J. How do viruses invade mucous surfaces? Phil. Trans. R. Soc. Lond. 1983;B303(75) [Google Scholar]